
Expression and Function of IL-12 and IL-18 Receptors on Human Tonsillar B Cells
10
of June 26, 2015. This information is current as Receptors on Human Tonsillar B Cells Expression and Function of IL-12 and IL-18 Trinchieri and Vito Pistoia Corcione, Paola Facchetti, Roberta Guglielmino, Giorgio Irma Airoldi, Giorgia Gri, Jason D. Marshall, Anna http://www.jimmunol.org/content/165/12/6880 doi: 10.4049/jimmunol.165.12.6880 2000; 165:6880-6888; ; J Immunol References http://www.jimmunol.org/content/165/12/6880.full#ref-list-1 , 30 of which you can access for free at: cites 60 articles This article Subscriptions http://jimmunol.org/subscriptions is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/ji/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/cgi/alerts/etoc Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2000 by The American Association of 9650 Rockville Pike, Bethesda, MD 20814-3994. The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on June 26, 2015 http://www.jimmunol.org/ Downloaded from by guest on June 26, 2015 http://www.jimmunol.org/ Downloaded from
Transcript of Expression and Function of IL-12 and IL-18 Receptors on Human Tonsillar B Cells

of June 26, 2015.This information is current as
Receptors on Human Tonsillar B CellsExpression and Function of IL-12 and IL-18
Trinchieri and Vito PistoiaCorcione, Paola Facchetti, Roberta Guglielmino, Giorgio Irma Airoldi, Giorgia Gri, Jason D. Marshall, Anna
http://www.jimmunol.org/content/165/12/6880doi: 10.4049/jimmunol.165.12.6880
2000; 165:6880-6888; ;J Immunol
Referenceshttp://www.jimmunol.org/content/165/12/6880.full#ref-list-1
, 30 of which you can access for free at: cites 60 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2000 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on June 26, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on June 26, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

Expression and Function of IL-12 and IL-18 Receptors onHuman Tonsillar B Cells1
Irma Airoldi, 2*† Giorgia Gri, †‡ Jason D. Marshall,†§ Anna Corcione,* Paola Facchetti,*Roberta Guglielmino,* Giorgio Trinchieri, † and Vito Pistoia*
IL-12 activates murine and human B cells, but little information is available as to the expression and function of IL-12R on humanB lymphocytes. Here we show that the latter cells, freshly isolated from human tonsils, expressed the transcripts of bothb1 andb2 chains of IL-12R and that b2 chain mRNA was selectively increased (4- to 5-fold) by incubation withStaphylococcus aureusCowan I bacteria or IL-12. B cell stimulation with IL-12 induced de novo expression of the transcripts of the two chains of IL-18R,i.e., IL-1 receptor-related protein and accessory protein-like. Functional studies showed that both IL-12 and IL-18 signaled to Bcells through the NF-kB pathway. In the case of IL-12, no involvement of STAT transcription factors, and in particular of STAT-4,was detected. c-reland p50 were identified as the members of NF-kB family involved in IL-12-mediated signal transduction to Bcells. IL-12 and IL-18 synergized in the induction of IFN-g production by tonsillar B cells, but not in the stimulation of B celldifferentiation, although either cytokine promoted IgM secretion in culture supernatants. Finally, naive but not germinal centeror memory, tonsillar B cells were identified as the exclusive IL-12 targets in terms of induction of NF-kB activation and of IFN-gproduction. The Journal of Immunology,2000, 165: 6880–6888.
I nterleukin-12 is an immunomodulatory cytokine that repre-sents a functional bridge between innate resistance and Ag-specific adaptive immunity (1). IL-12 is produced by various
cell types, such as phagocytic cells, dendritic cells, and B lym-phocytes, and induces production of cytokines, especially IFN-g,from T and NK cells after binding to its own receptor (2–9).
The IL-12R is composed of two subunits designatedb1 andb2(10, 11). The tissue distribution of theb2 subunit is more restrictedthan that of theb1, and regulation ofb2 expression is likely torepresent a central mechanism whereby IL-12 responsiveness iscontrolled (12–17). In T and NK cells, IL-12 induces activation ofSTAT family members, i.e., STAT3 and STAT4, which bind to thepromoter regions of different genes, including the IFN-g gene pro-moter (18–23). However, STAT-4 activation is restricted to IL-12and IFN-a, whereas STAT-3 is activated by different cytokines(18–23).
Recently, murine dendritic cells have been found to transduceIL-12 signaling through NF-kB activation, leading to functionalresponses (24). IL-12 mediates activation of human and murine Bcells (25–30). It is well known that human B lymphocytes expressconstitutively the IL-12Rb1 chain, but no information is availableas to the expression of theb2 chain in these cells (31, 32). Fur-
thermore, the signal transduction pathways initiated in B lympho-cytes by IL-12 triggering are unknown.
Another IFN-g-inducing factor sharing functional similaritieswith IL-12, i.e., IL-18, has been recently cloned (33). IL-18 issynthesized by various cell types (34–36) as an inactive precursormolecule that becomes functional following cleavage by caspase-1(37, 38). IL-18 synergizes with IL-12 in the enhancement of NKcell cytotoxicity and of T cell proliferation, as well as in the in-duction of IFN-g production by T cells (39, 40). The IL-18R iscomposed of two chains, named IL-1 receptor-related protein (IL-1Rrp)3 and accessory protein-like (AcPL), respectively, both ofwhich are required for IL-18-driven induction of NF-kB activation(41–44). IL-12 induces the expression of both IL-1Rrp and AcPLin different cell types (39, 45).
Here we have investigated IL-12R expression and function infreshly isolated or cultured human tonsillar B lymphocytes and intheir subsets. Because IL-12 was found to induce de novo expres-sion of IL-18R chains, the interactions of IL-12 and IL-18 withrespect to different B cell functions were also investigated.
Materials and MethodsCell separation and culture
Mononuclear cells (MNC) were isolated from surgically removed tonsilson Ficoll-Hypaque density gradients and depleted of T cells by rosettingwith neuraminidase-treated sheep erythrocytes. Non-T cells were subse-quently deprived of macrophages and NK cells by incubation with CD68and CD56 mAbs, respectively, followed by immune rosetting (46). Theresulting cell preparations contained on average 99% B cells, as assessedby staining with CD19 or CD20 mAbs (Becton Dickinson, San Jose, CA).CD31 T cells, CD561 NK cells, and CD681 macrophages were virtuallyabsent (,1%) from these cell suspensions, as assessed by flow cytometry.CD3, CD56, and CD68 mAbs were all obtained from Becton Dickinson.Controls were cells stained with isotype-matched mAbs of unrelated spec-ificities; at least 10,000 events were acquired for each staining.
*Laboratory of Oncology, G. Gaslini Institute, Genoa, Italy;†Wistar Institute of Anat-omy and Biology, Philadelphia, PA 19104;‡Division of Experimental Oncology,Istituto per lo Studio e la Cura dei tumori, Milan, Italy; and§Dynavax Technologies,Berkeley, CA 94705
Received for publication February 1, 2000. Accepted for publication September26, 2000.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisementin accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by grants from the Associazione Italiana per la Ricerca sulCancro, Italy and Ministero della Sanita, Progetti Finalizzati, Italy (to V.P.). I.A. wassupported by an Italian Foundation for Cancer Research (FIRC) fellowship.2 Address correspondence and reprint requests to Dr. Irma Airoldi, Laboratory ofOncology, G. Gaslini Institute, Largo G. Gaslini, 5 16148 Genova, Italy. E-mailaddress: [email protected]
3 Abbreviations used in this paper: IL-1Rrp, IL-1 receptor-related protein; SAC,Staphylococcus aureusCowan I bacteria; AcPL, accessory protein-like; MNC, mono-nuclear cells; GC, germinal center; HPRT, hypoxanthine-guanine phosphoribosyl-transferase; RPA, RNase protection assay.
Copyright © 2000 by The American Association of Immunologists 0022-1767/00/$02.00
by guest on June 26, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

The purity of B cell fractions was further checked by extracting RNAand searching for the expression of the CD3g (T cell-specific), CD56 (NKcell-specific), and CD19 (B cell-specific) genes by RT-PCR (see below forthe sequences of the primers and the details of these procedures).
As shown in Fig. 1A, the CD3g and CD56 transcripts were not detectedin B cell-enriched suspensions, whereas they were consistently found inneuraminidase-treated sheep erythrocytes-rosette forming tonsillar cellfractions. The opposite pattern of expression was observed for the B cell-specific CD19 gene. These experiments demonstrate that there were no Tor NK cell contaminants in the tonsillar B suspensions used throughout thisstudy.
Fractionation of tonsillar B lymphocytes into germinal center (GC), naive,and memory cell subsets was performed as follows. Naive B lymphocyteswere isolated as IgD1 cells from total B lymphocyte suspensions by immunerosetting (46). The IgD2 B cell fractions were further separated into CD381
(GC) cells and CD382 (memory) cells by the same technique (46).All of the separation procedures were performed at 4°C to prevent spon-
taneous apoptosis of GC B cells. Purified B cells were used immediatelyafter isolation or were cultured for the indicated times at the concentrationof 1 3 106/ml in RPMI 1640 medium (Seromed-Biochrom KG, Berlin,Germany) supplemented with 10% FCS (Seromed) in the presence of dif-ferent stimuli (see below).
RNA preparation, PCR, and RNase protectionassay (RPA)
RNA was extracted from cultured cells using Ultraspec (Biotecx Labora-tories, Houston, TX). The stimuli tested were:Staphylococcus aureusCowan I bacteria (SAC; 1:10,000) (Calbiochem, La Jolla, CA); rIL-12 (20ng/ml) (Genetics Institute, Cambridge, MA); and anti-k (1 mg/106 cells)(Southern Biotechnology Associates, Birmingham, AL) in combinationwith anti-l (1 mg/106 cells) Ig light chain mAbs (Southern BiotechnologyAssociates); CD40 mAb (1mg/ml) (Immunotech, Marseille, France), aloneor in combination with rIL-4 (10 ng/ml) (Genzyme, Cambridge, MA);rIL-4 alone (10 ng/ml); and IFN-g (1000 U/ml) (Boehringer Mannheim,Mannheim, Germany). Primer sequences and profiles of amplification wereas follows: hypoxanthine-guanine phosphoribosyltransferase (HPRT)sense CCTGCTGGATTACATCAAAGCACTG, antisense TCCAACACTTCGTGGGGTCCT, 94°C for 1 min, 61°C for 1 min, 72°C for 1 min,30 cycles; IL-12Rb1 sense TCTTCCTCTTCCTGCTGTCC, antisenseCAGCCGCCTCCTCCCATCC, 94°C for 1 min, 55°C for 1 min, 72°C for1 min, 30 cycles; IL-12Rb2 sense ACCGGAAATTGGGCTGTGGCTG,antisense AATGCTTGGTGCCACAAACGCC 94°C for 1 min, 55°C for 1min, 72°C for 1 min, 30 cycles; IL-1Rrp sense ATTACCCTTGACCCTTTGGG, antisense TCAAACTCGGCGTTCTTCTT, 94°C for 1 min, 55°Cfor 1 min, 58°C for 1 min, 27 cycles; AcPL sense CCTGCCCTTCATGGGTAGTA, antisense ATCCACTACGATTCGGTTGC, 94°C for 1 min,55°C for 1 min, 60°C for 1 min, 26 cycles; GAPDH sense ACATCGCTCAGAACACCTATGG, antisense GGGTCTACATGGCAACTGTGAG,94°C for 1 min, 60°C for 1 min, 72°C for 2 min, 30 cycles; CD3g senseGGTTCGGTACTTCTGACT, antisense TGGTTTTGACTTGTTCTG,94°C for 1 min, 48°C for 1 min, 72°C for 1 min, 32 cycles; CD19 senseACCTCCTCGCCTCCTCTTC, antisense TCCCCTTCCTCTTCTTCTG,94°C for 1 min, 57°C for 1 min, 72°C for 1 min, 32 cycles. Ten microlitersof each sample were electrophoresed through a 1.5% agarose gel withethidium bromide and scanned via FluorImager (Molecular Dynamics,Sunnyvale, CA). The specificity of the PCR products was checked both byconfirming the known base pair sequence length and by Southern blot ofthe amplified bands using internal probes specific for each gene investi-gated. Bands were assigned densitometric values by ImageQuaNT program(Molecular Dynamics), and these values were normalized to HPRT values.
RPA was performed using the PharMingen probe kit hCR-3 (San Diego,CA) with 5 mg/lane total RNA, according to the manufacturer’s protocol.Products were resolved on 6% denaturing polyacrylamide gels, and theprotected fragments were visualized and quantitated using a PhosphorIm-ager 445SI (Molecular Dynamics). Relative radioactivity values for IL-12Rb1 and IL-12Rb2 transcripts were determined by normalizing to thevalues obtained for the L32 housekeeping gene, which was used as internalcontrol for equal RNA loading.
Assay for Ig secretion
B cells were cultured in the presence of IL-12 (20 ng/ml) or SAC (1:10,000) for 48 h, washed, and incubated with IL-12 (20 ng/ml) or IL-18 (50ng/ml) (Harlan Breeders, Indianapolis, IN) alone or in combination, for 7days. Culture supernatants and human Ig standards (Behring, Marburg,Germany) were diluted in PBS containing 2% BSA (Sigma, St. Louis,MO). IgG, IgA, and IgM concentration was measured by ELISA. Samples(100 ml) were added to 96-well polystyrene ELISA/RIA plates (Dynatech
Laboratories, Chantilly, VA) previously coated for 16 h at 4°C with rabbitanti-human IgG, IgA, or IgM antisera (Dako, Glostrup, Denmark). Fol-lowing a 2-h incubation at 37°C and three washings with PBS containing0.001% Tween 20, HRP-conjugated goat anti-human IgG, IgA, or IgM(Dako) antiserum was added. After addition ofO-phenylenediamine (Sig-ma), substrate color development was stopped by 5 M sulfuric acid, andabsorbance was measured at 492 nm using a Gralis Ad Reader (Bouty,Milan, Italy). The concentration of Ig in the culture supernatants was cal-culated from the standard curve.
Detection of IFN-g in B cell culture supernatants
Purified tonsillar B cells pretreated for 48 h with IL-12 (20 ng/ml) or SAC(1:10,000) were washed and subsequently cultured for 24 or 48 h withIL-12 (20 ng/ml) and/or IL-18 (50 ng/ml). IgD1 (naive) and IgD2 (GC andmemory) B cell fractions were cultured for 24 h in the presence of IL-12(20 ng/ml). Supernatants were then collected and tested in triplicate forIFN-g by ELISA (BioSource International, Camarillo, CA).
IFN-g intracellular staining
Purified tonsillar B cells or MNC were cultured for 48 h with IL-12 andIL-18, washed, and incubated for an additional 5 h with 20 ng/ml PMA,250 ng/ml calcium ionophore, and 5mg/ml brefeldin A (all reagents pur-chased from Sigma). After washing, cells were single- or double-stainedfor a pan-B surface marker (CD20) and/or intracytoplasmic IFN-g. Fordouble staining, B cells were first incubated with PE-conjugated CD20mAb (Becton Dickinson) or with the appropriate control (see below) for 30min at 4°C, then fixed with 4% paraformaldehyde (Sigma) for 20 min at4°C and permeabilized with 0.1% saponin (Sigma) in PBS. In some ex-periments, a PE-conjugated CD3 mAb (Becton Dickinson) was used in theplace of CD20 mAb following the same protocol. Cells were subsequentlyincubated with an FITC-conjugated anti-IFN-g mAb (PharMingen) or withthe appropriate controls (see below) for 30 min at 4°C, washed, and enu-merated by flow cytometry. Control for CD20 surface staining were cellsincubated with a PE-conjugated, isotype-matched mAb of irrelevant spec-ificity. Controls for intracytoplasmic IFN-g staining were cells incubatedwith an FITC-conjugated, isotype-matched mAb of irrelevant specificity orwith the anti-IFN-g mAb preincubated with rIFN-g before staining. Singlestaining of B cells by CD20 or anti-IFN-g mAbs was conducted using theabove procedures separately.
Nuclear extracts and EMSA
Purified B lymphocytes were used directly or following stimulation withSAC (1:10,000) or IL-12 (20 ng/ml) for 48 h, washed with serum-freeRPMI 1640, and starved for 3 h at 37°C in RPMI 1640 supplemented with1% albumin. Cells were then resuspended at a concentration of 23 107/mlin medium with or without IL-12 (20 ng/ml) or IL-18 (50 ng/ml), eitheralone or in combination, and incubated at 37°C for 30 min. Nuclear ex-tracts were subsequently prepared as previously described (47). Freshlyisolated IgD1 and IgD2 B cells starved as described above were incubatedfor 30 min with IL-12 before preparing nuclear extract.
End-labeled DNA probes (50,000 cpm/sample) were mixed with 4mgof crude nuclear extract and incubated at room temperature for 25 min inthe presence of 1mg of poly (dI-dC) (Boehringer Mannheim) in a volumeof 10 ml of buffer C (20 mM HEPES, pH 7.9; 0.4 M NaCl; 0.1 mM EGTA;1 mM DTT; 1 mM PMSF; 10mg/ml aprotinin; 10mg/ml leupeptin; 1mg/ml pepstatin A; and 1 mM sodium orthovanadate). The mix was thenfractionated through a 6% polyacrylamide gel in 0.53Tris borate EDTAfor 1.5 h at 150 V. The gel was dried, exposed in a PhosphorImager storagescreen, and scanned. Supershift experiments were conducted by preincu-bating the nuclear extract with 1–2mg polyclonal anti-p50, anti-p65, anti-c-Rel, anti-Rel-B, or anti-ets2 rabbit antisera (all purchased from SantaCruz Biotechnology, Santa Cruz, CA) or normal rabbit serum for 30 minat room temperature after the addition of radiolabeled probe. The followingdouble-strand oligonucleotides were used in EMSA as labeled or compet-itor probes: IFN-g activation site (GAS), 59-GTATTTCCCAGAAAAG-GAAC 39, NF-kB 59-ATGTGAGGGGACTTTCCCAGG-39.
Total extracts and Western blot analysis
Purified tonsillar B cells were stimulated with SAC (1:10,000) or mediumfor 48 h, washed with serum-free RPMI 1640, and incubated for an addi-tional 30 min with IL-12 (20 ng/ml) or medium alone. Cells were subse-quently incubated for 30 min on ice with lysis buffer containing 20 mMHEPES, 150 mM NaCl, 10% glycerol, 0.5% Nonidet P-40, 1 mM EDTA,2.5 mM DTT, 10mg/ml aprotinin, 10mg/ml leupeptin, 1mg/ml pepstatinA, 1 mM PMSF, and 1 mM Na3VO4. During this time interval, cells weresubjected to vortex mixing every 5 min. Thereafter, lysates were centrifuged at
6881The Journal of Immunology
by guest on June 26, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
12,000 rpm for 5 min at 4°C, and supernatants were quantitated by the BCAkit assay (Pierce, Rockford, IL). Equal amounts of protein (10mg) were loadedon 8% SDS-polyacrylamide gel and boiled 3 min before application. Gel wasblotted onto Protean nitrocellulose membrane (Schleicher & Schuell, Dassel,Germany) for 1 h at 100 V. Blots were incubated in a blocking buffer con-taining BSA and 0.5% Tween 20 in TBS for 1 h, followed by incubation withrabbit anti-human STAT4 antiserum (Santa Cruz Biotechnology). After threewashings in TBS-Tween 20, blots were incubated for 1 h with goat anti-rabbitIg conjugated with HRP (Santa Cruz Biotechnology) at the final concentrationof 50 ng/ml in TBS-Tween 20 containing 1% BSA. Detection was performedby enhanced chemiluminescence (ECL, Pierce).
ResultsIL-12R and IL-18R expression in human tonsillar B cells
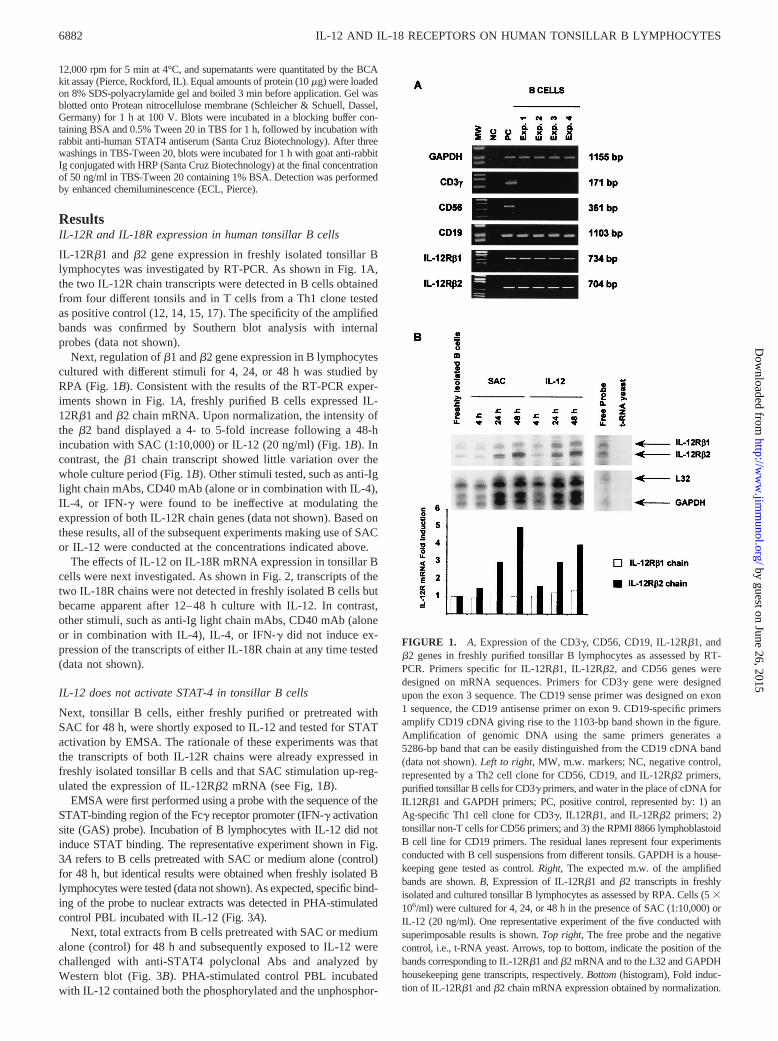
IL-12Rb1 andb2 gene expression in freshly isolated tonsillar Blymphocytes was investigated by RT-PCR. As shown in Fig. 1A,the two IL-12R chain transcripts were detected in B cells obtainedfrom four different tonsils and in T cells from a Th1 clone testedas positive control (12, 14, 15, 17). The specificity of the amplifiedbands was confirmed by Southern blot analysis with internalprobes (data not shown).
Next, regulation ofb1 andb2 gene expression in B lymphocytescultured with different stimuli for 4, 24, or 48 h was studied byRPA (Fig. 1B). Consistent with the results of the RT-PCR exper-iments shown in Fig. 1A, freshly purified B cells expressed IL-12Rb1 andb2 chain mRNA. Upon normalization, the intensity ofthe b2 band displayed a 4- to 5-fold increase following a 48-hincubation with SAC (1:10,000) or IL-12 (20 ng/ml) (Fig. 1B). Incontrast, theb1 chain transcript showed little variation over thewhole culture period (Fig. 1B). Other stimuli tested, such as anti-Iglight chain mAbs, CD40 mAb (alone or in combination with IL-4),IL-4, or IFN-g were found to be ineffective at modulating theexpression of both IL-12R chain genes (data not shown). Based onthese results, all of the subsequent experiments making use of SACor IL-12 were conducted at the concentrations indicated above.
The effects of IL-12 on IL-18R mRNA expression in tonsillar Bcells were next investigated. As shown in Fig. 2, transcripts of thetwo IL-18R chains were not detected in freshly isolated B cells butbecame apparent after 12–48 h culture with IL-12. In contrast,other stimuli, such as anti-Ig light chain mAbs, CD40 mAb (aloneor in combination with IL-4), IL-4, or IFN-g did not induce ex-pression of the transcripts of either IL-18R chain at any time tested(data not shown).
IL-12 does not activate STAT-4 in tonsillar B cells
Next, tonsillar B cells, either freshly purified or pretreated withSAC for 48 h, were shortly exposed to IL-12 and tested for STATactivation by EMSA. The rationale of these experiments was thatthe transcripts of both IL-12R chains were already expressed infreshly isolated tonsillar B cells and that SAC stimulation up-reg-ulated the expression of IL-12Rb2 mRNA (see Fig, 1B).
EMSA were first performed using a probe with the sequence of theSTAT-binding region of the Fcg receptor promoter (IFN-g activationsite (GAS) probe). Incubation of B lymphocytes with IL-12 did notinduce STAT binding. The representative experiment shown in Fig.3A refers to B cells pretreated with SAC or medium alone (control)for 48 h, but identical results were obtained when freshly isolated Blymphocytes were tested (data not shown). As expected, specific bind-ing of the probe to nuclear extracts was detected in PHA-stimulatedcontrol PBL incubated with IL-12 (Fig. 3A).
Next, total extracts from B cells pretreated with SAC or mediumalone (control) for 48 h and subsequently exposed to IL-12 werechallenged with anti-STAT4 polyclonal Abs and analyzed byWestern blot (Fig. 3B). PHA-stimulated control PBL incubatedwith IL-12 contained both the phosphorylated and the unphosphor-
FIGURE 1. A, Expression of the CD3g, CD56, CD19, IL-12Rb1, andb2 genes in freshly purified tonsillar B lymphocytes as assessed by RT-PCR. Primers specific for IL-12Rb1, IL-12Rb2, and CD56 genes weredesigned on mRNA sequences. Primers for CD3g gene were designedupon the exon 3 sequence. The CD19 sense primer was designed on exon1 sequence, the CD19 antisense primer on exon 9. CD19-specific primersamplify CD19 cDNA giving rise to the 1103-bp band shown in the figure.Amplification of genomic DNA using the same primers generates a5286-bp band that can be easily distinguished from the CD19 cDNA band(data not shown).Left to right, MW, m.w. markers; NC, negative control,represented by a Th2 cell clone for CD56, CD19, and IL-12Rb2 primers,purified tonsillar B cells for CD3g primers, and water in the place of cDNA forIL12Rb1 and GAPDH primers; PC, positive control, represented by: 1) anAg-specific Th1 cell clone for CD3g, IL12Rb1, and IL-12Rb2 primers; 2)tonsillar non-T cells for CD56 primers; and 3) the RPMI 8866 lymphoblastoidB cell line for CD19 primers. The residual lanes represent four experimentsconducted with B cell suspensions from different tonsils. GAPDH is a house-keeping gene tested as control.Right, The expected m.w. of the amplifiedbands are shown.B, Expression of IL-12Rb1 andb2 transcripts in freshlyisolated and cultured tonsillar B lymphocytes as assessed by RPA. Cells (53106/ml) were cultured for 4, 24, or 48 h in the presence of SAC (1:10,000) orIL-12 (20 ng/ml). One representative experiment of the five conducted withsuperimposable results is shown.Top right, The free probe and the negativecontrol, i.e., t-RNA yeast. Arrows, top to bottom, indicate the position of thebands corresponding to IL-12Rb1 andb2 mRNA and to the L32 and GAPDHhousekeeping gene transcripts, respectively.Bottom(histogram), Fold induc-tion of IL-12Rb1 andb2 chain mRNA expression obtained by normalization.
6882 IL-12 AND IL-18 RECEPTORS ON HUMAN TONSILLAR B LYMPHOCYTES
by guest on June 26, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
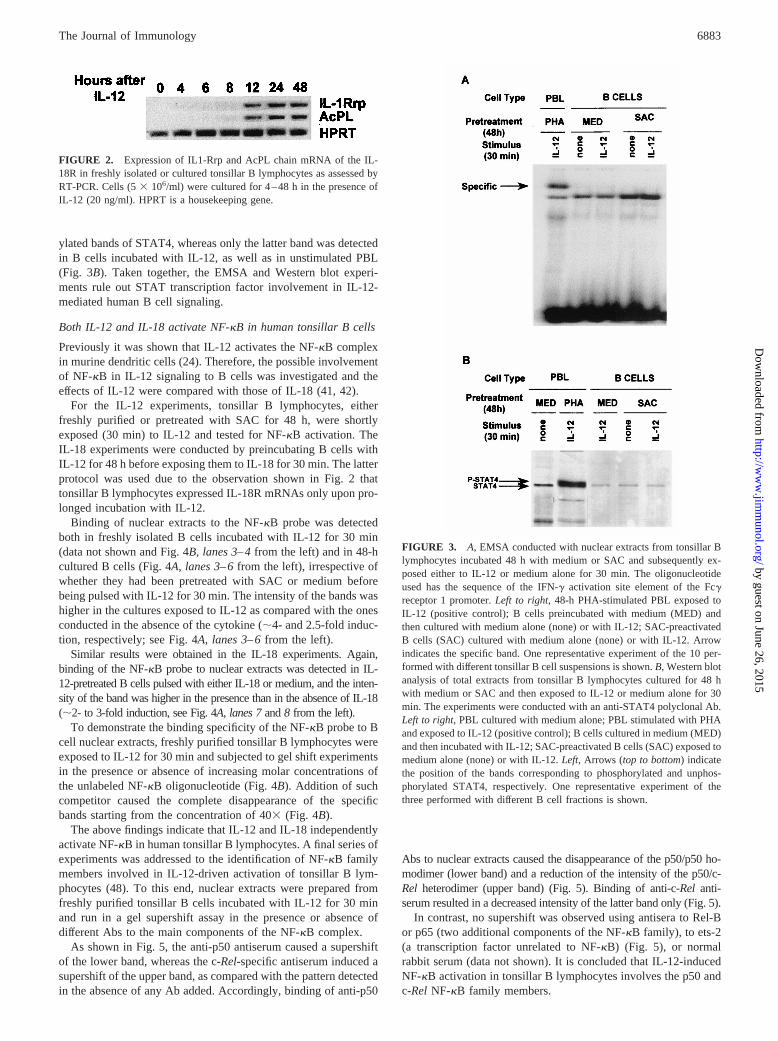
ylated bands of STAT4, whereas only the latter band was detectedin B cells incubated with IL-12, as well as in unstimulated PBL(Fig. 3B). Taken together, the EMSA and Western blot experi-ments rule out STAT transcription factor involvement in IL-12-mediated human B cell signaling.
Both IL-12 and IL-18 activate NF-kB in human tonsillar B cells
Previously it was shown that IL-12 activates the NF-kB complexin murine dendritic cells (24). Therefore, the possible involvementof NF-kB in IL-12 signaling to B cells was investigated and theeffects of IL-12 were compared with those of IL-18 (41, 42).
For the IL-12 experiments, tonsillar B lymphocytes, eitherfreshly purified or pretreated with SAC for 48 h, were shortlyexposed (30 min) to IL-12 and tested for NF-kB activation. TheIL-18 experiments were conducted by preincubating B cells withIL-12 for 48 h before exposing them to IL-18 for 30 min. The latterprotocol was used due to the observation shown in Fig. 2 thattonsillar B lymphocytes expressed IL-18R mRNAs only upon pro-longed incubation with IL-12.
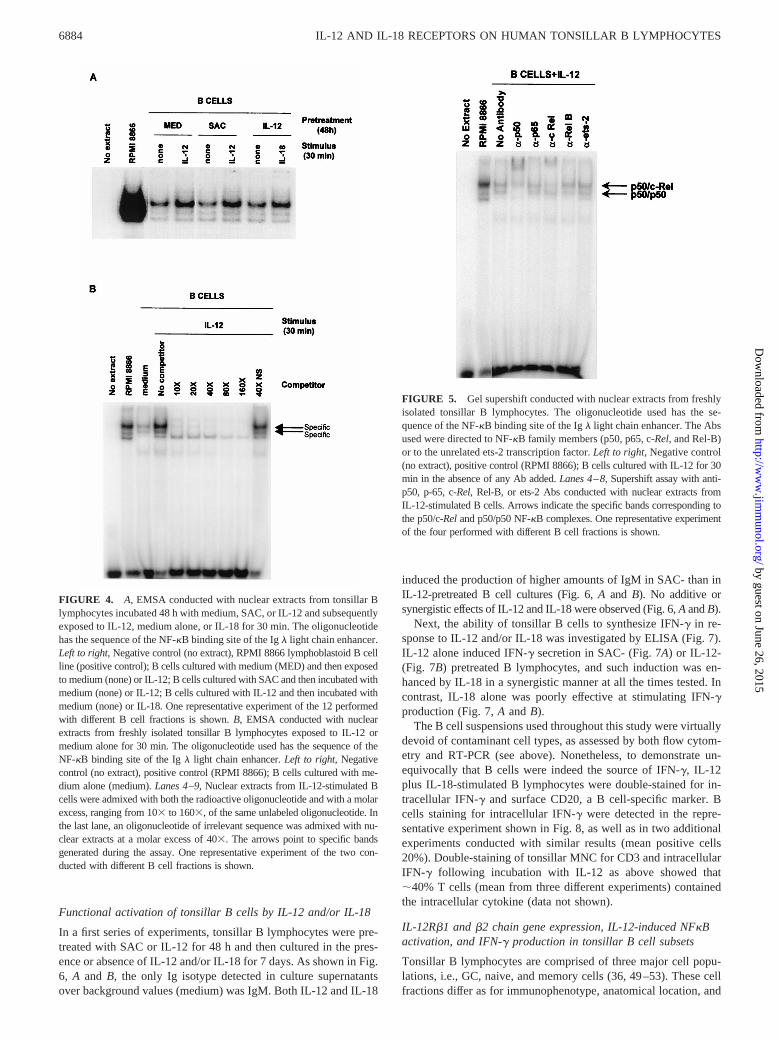
Binding of nuclear extracts to the NF-kB probe was detectedboth in freshly isolated B cells incubated with IL-12 for 30 min(data not shown and Fig. 4B,lanes 3–4from the left) and in 48-hcultured B cells (Fig. 4A,lanes 3–6from the left), irrespective ofwhether they had been pretreated with SAC or medium beforebeing pulsed with IL-12 for 30 min. The intensity of the bands washigher in the cultures exposed to IL-12 as compared with the onesconducted in the absence of the cytokine (;4- and 2.5-fold induc-tion, respectively; see Fig. 4A, lanes 3–6from the left).
Similar results were obtained in the IL-18 experiments. Again,binding of the NF-kB probe to nuclear extracts was detected in IL-12-pretreated B cells pulsed with either IL-18 or medium, and the inten-sity of the band was higher in the presence than in the absence of IL-18(;2- to 3-fold induction, see Fig. 4A, lanes 7and8 from the left).
To demonstrate the binding specificity of the NF-kB probe to Bcell nuclear extracts, freshly purified tonsillar B lymphocytes wereexposed to IL-12 for 30 min and subjected to gel shift experimentsin the presence or absence of increasing molar concentrations ofthe unlabeled NF-kB oligonucleotide (Fig. 4B). Addition of suchcompetitor caused the complete disappearance of the specificbands starting from the concentration of 403 (Fig. 4B).
The above findings indicate that IL-12 and IL-18 independentlyactivate NF-kB in human tonsillar B lymphocytes. A final series ofexperiments was addressed to the identification of NF-kB familymembers involved in IL-12-driven activation of tonsillar B lym-phocytes (48). To this end, nuclear extracts were prepared fromfreshly purified tonsillar B cells incubated with IL-12 for 30 minand run in a gel supershift assay in the presence or absence ofdifferent Abs to the main components of the NF-kB complex.
As shown in Fig. 5, the anti-p50 antiserum caused a supershiftof the lower band, whereas the c-Rel-specific antiserum induced asupershift of the upper band, as compared with the pattern detectedin the absence of any Ab added. Accordingly, binding of anti-p50
Abs to nuclear extracts caused the disappearance of the p50/p50 ho-modimer (lower band) and a reduction of the intensity of the p50/c-Rel heterodimer (upper band) (Fig. 5). Binding of anti-c-Rel anti-serum resulted in a decreased intensity of the latter band only (Fig. 5).
In contrast, no supershift was observed using antisera to Rel-Bor p65 (two additional components of the NF-kB family), to ets-2(a transcription factor unrelated to NF-kB) (Fig. 5), or normalrabbit serum (data not shown). It is concluded that IL-12-inducedNF-kB activation in tonsillar B lymphocytes involves the p50 andc-RelNF-kB family members.
FIGURE 2. Expression of IL1-Rrp and AcPL chain mRNA of the IL-18R in freshly isolated or cultured tonsillar B lymphocytes as assessed byRT-PCR. Cells (53 106/ml) were cultured for 4–48 h in the presence ofIL-12 (20 ng/ml). HPRT is a housekeeping gene.
FIGURE 3. A, EMSA conducted with nuclear extracts from tonsillar Blymphocytes incubated 48 h with medium or SAC and subsequently ex-posed either to IL-12 or medium alone for 30 min. The oligonucleotideused has the sequence of the IFN-g activation site element of the Fcgreceptor 1 promoter.Left to right, 48-h PHA-stimulated PBL exposed toIL-12 (positive control); B cells preincubated with medium (MED) andthen cultured with medium alone (none) or with IL-12; SAC-preactivatedB cells (SAC) cultured with medium alone (none) or with IL-12. Arrowindicates the specific band. One representative experiment of the 10 per-formed with different tonsillar B cell suspensions is shown.B, Western blotanalysis of total extracts from tonsillar B lymphocytes cultured for 48 hwith medium or SAC and then exposed to IL-12 or medium alone for 30min. The experiments were conducted with an anti-STAT4 polyclonal Ab.Left to right, PBL cultured with medium alone; PBL stimulated with PHAand exposed to IL-12 (positive control); B cells cultured in medium (MED)and then incubated with IL-12; SAC-preactivated B cells (SAC) exposed tomedium alone (none) or with IL-12.Left, Arrows (top to bottom) indicatethe position of the bands corresponding to phosphorylated and unphos-phorylated STAT4, respectively. One representative experiment of thethree performed with different B cell fractions is shown.
6883The Journal of Immunology
by guest on June 26, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
Functional activation of tonsillar B cells by IL-12 and/or IL-18
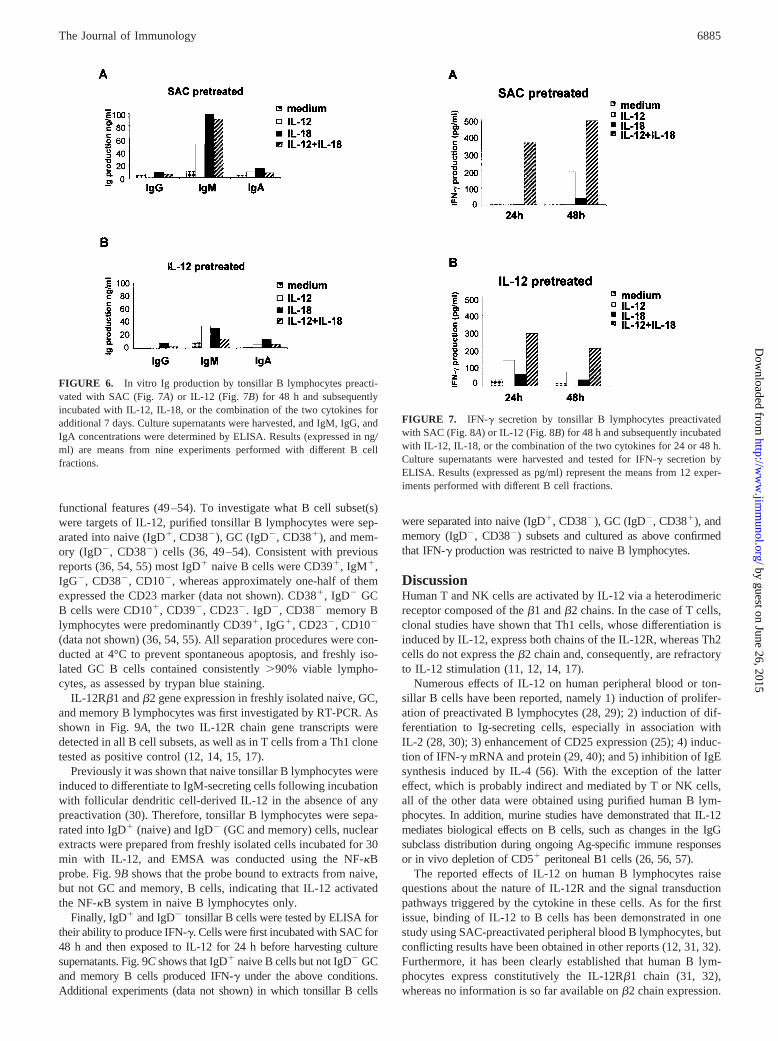
In a first series of experiments, tonsillar B lymphocytes were pre-treated with SAC or IL-12 for 48 h and then cultured in the pres-ence or absence of IL-12 and/or IL-18 for 7 days. As shown in Fig.6, A and B, the only Ig isotype detected in culture supernatantsover background values (medium) was IgM. Both IL-12 and IL-18
induced the production of higher amounts of IgM in SAC- than inIL-12-pretreated B cell cultures (Fig. 6,A and B). No additive orsynergistic effects of IL-12 and IL-18 were observed (Fig. 6,A andB).
Next, the ability of tonsillar B cells to synthesize IFN-g in re-sponse to IL-12 and/or IL-18 was investigated by ELISA (Fig. 7).IL-12 alone induced IFN-g secretion in SAC- (Fig. 7A) or IL-12-(Fig. 7B) pretreated B lymphocytes, and such induction was en-hanced by IL-18 in a synergistic manner at all the times tested. Incontrast, IL-18 alone was poorly effective at stimulating IFN-gproduction (Fig. 7,A andB).
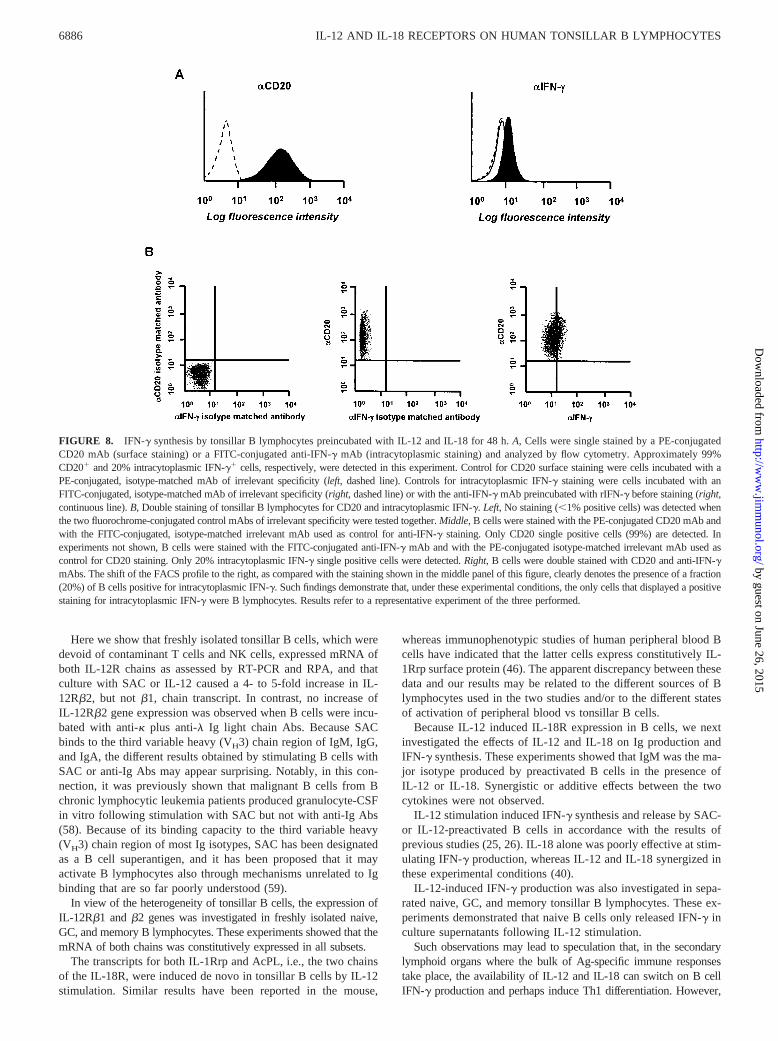
The B cell suspensions used throughout this study were virtuallydevoid of contaminant cell types, as assessed by both flow cytom-etry and RT-PCR (see above). Nonetheless, to demonstrate un-equivocally that B cells were indeed the source of IFN-g, IL-12plus IL-18-stimulated B lymphocytes were double-stained for in-tracellular IFN-g and surface CD20, a B cell-specific marker. Bcells staining for intracellular IFN-g were detected in the repre-sentative experiment shown in Fig. 8, as well as in two additionalexperiments conducted with similar results (mean positive cells20%). Double-staining of tonsillar MNC for CD3 and intracellularIFN-g following incubation with IL-12 as above showed that;40% T cells (mean from three different experiments) containedthe intracellular cytokine (data not shown).
IL-12Rb1 andb2 chain gene expression, IL-12-induced NFkBactivation, and IFN-g production in tonsillar B cell subsets
Tonsillar B lymphocytes are comprised of three major cell popu-lations, i.e., GC, naive, and memory cells (36, 49–53). These cellfractions differ as for immunophenotype, anatomical location, and
FIGURE 4. A, EMSA conducted with nuclear extracts from tonsillar Blymphocytes incubated 48 h with medium, SAC, or IL-12 and subsequentlyexposed to IL-12, medium alone, or IL-18 for 30 min. The oligonucleotidehas the sequence of the NF-kB binding site of the Igl light chain enhancer.Left to right, Negative control (no extract), RPMI 8866 lymphoblastoid B cellline (positive control); B cells cultured with medium (MED) and then exposedto medium (none) or IL-12; B cells cultured with SAC and then incubated withmedium (none) or IL-12; B cells cultured with IL-12 and then incubated withmedium (none) or IL-18. One representative experiment of the 12 performedwith different B cell fractions is shown.B, EMSA conducted with nuclearextracts from freshly isolated tonsillar B lymphocytes exposed to IL-12 ormedium alone for 30 min. The oligonucleotide used has the sequence of theNF-kB binding site of the Igl light chain enhancer.Left to right, Negativecontrol (no extract), positive control (RPMI 8866); B cells cultured with me-dium alone (medium).Lanes 4–9,Nuclear extracts from IL-12-stimulated Bcells were admixed with both the radioactive oligonucleotide and with a molarexcess, ranging from 103 to 1603, of the same unlabeled oligonucleotide. Inthe last lane, an oligonucleotide of irrelevant sequence was admixed with nu-clear extracts at a molar excess of 403. The arrows point to specific bandsgenerated during the assay. One representative experiment of the two con-ducted with different B cell fractions is shown.
FIGURE 5. Gel supershift conducted with nuclear extracts from freshlyisolated tonsillar B lymphocytes. The oligonucleotide used has the se-quence of the NF-kB binding site of the Igl light chain enhancer. The Absused were directed to NF-kB family members (p50, p65, c-Rel, and Rel-B)or to the unrelated ets-2 transcription factor.Left to right, Negative control(no extract), positive control (RPMI 8866); B cells cultured with IL-12 for 30min in the absence of any Ab added.Lanes 4–8, Supershift assay with anti-p50, p-65, c-Rel, Rel-B, or ets-2 Abs conducted with nuclear extracts fromIL-12-stimulated B cells. Arrows indicate the specific bands corresponding tothe p50/c-Reland p50/p50 NF-kB complexes. One representative experimentof the four performed with different B cell fractions is shown.
6884 IL-12 AND IL-18 RECEPTORS ON HUMAN TONSILLAR B LYMPHOCYTES
by guest on June 26, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
functional features (49–54). To investigate what B cell subset(s)were targets of IL-12, purified tonsillar B lymphocytes were sep-arated into naive (IgD1, CD382), GC (IgD2, CD381), and mem-ory (IgD2, CD382) cells (36, 49–54). Consistent with previousreports (36, 54, 55) most IgD1 naive B cells were CD391, IgM1,IgG2, CD382, CD102, whereas approximately one-half of themexpressed the CD23 marker (data not shown). CD381, IgD2 GCB cells were CD101, CD392, CD232. IgD2, CD382 memory Blymphocytes were predominantly CD391, IgG1, CD232, CD102
(data not shown) (36, 54, 55). All separation procedures were con-ducted at 4°C to prevent spontaneous apoptosis, and freshly iso-lated GC B cells contained consistently.90% viable lympho-cytes, as assessed by trypan blue staining.
IL-12Rb1 andb2 gene expression in freshly isolated naive, GC,and memory B lymphocytes was first investigated by RT-PCR. Asshown in Fig. 9A, the two IL-12R chain gene transcripts weredetected in all B cell subsets, as well as in T cells from a Th1 clonetested as positive control (12, 14, 15, 17).
Previously it was shown that naive tonsillar B lymphocytes wereinduced to differentiate to IgM-secreting cells following incubationwith follicular dendritic cell-derived IL-12 in the absence of anypreactivation (30). Therefore, tonsillar B lymphocytes were sepa-rated into IgD1 (naive) and IgD2 (GC and memory) cells, nuclearextracts were prepared from freshly isolated cells incubated for 30min with IL-12, and EMSA was conducted using the NF-kBprobe. Fig. 9Bshows that the probe bound to extracts from naive,but not GC and memory, B cells, indicating that IL-12 activatedthe NF-kB system in naive B lymphocytes only.
Finally, IgD1 and IgD2 tonsillar B cells were tested by ELISA fortheir ability to produce IFN-g. Cells were first incubated with SAC for48 h and then exposed to IL-12 for 24 h before harvesting culturesupernatants. Fig. 9Cshows that IgD1 naive B cells but not IgD2 GCand memory B cells produced IFN-g under the above conditions.Additional experiments (data not shown) in which tonsillar B cells
were separated into naive (IgD1, CD382), GC (IgD2, CD381), andmemory (IgD2, CD382) subsets and cultured as above confirmedthat IFN-g production was restricted to naive B lymphocytes.
DiscussionHuman T and NK cells are activated by IL-12 via a heterodimericreceptor composed of theb1 andb2 chains. In the case of T cells,clonal studies have shown that Th1 cells, whose differentiation isinduced by IL-12, express both chains of the IL-12R, whereas Th2cells do not express theb2 chain and, consequently, are refractoryto IL-12 stimulation (11, 12, 14, 17).
Numerous effects of IL-12 on human peripheral blood or ton-sillar B cells have been reported, namely 1) induction of prolifer-ation of preactivated B lymphocytes (28, 29); 2) induction of dif-ferentiation to Ig-secreting cells, especially in association withIL-2 (28, 30); 3) enhancement of CD25 expression (25); 4) induc-tion of IFN-g mRNA and protein (29, 40); and 5) inhibition of IgEsynthesis induced by IL-4 (56). With the exception of the lattereffect, which is probably indirect and mediated by T or NK cells,all of the other data were obtained using purified human B lym-phocytes. In addition, murine studies have demonstrated that IL-12mediates biological effects on B cells, such as changes in the IgGsubclass distribution during ongoing Ag-specific immune responsesor in vivo depletion of CD51 peritoneal B1 cells (26, 56, 57).
The reported effects of IL-12 on human B lymphocytes raisequestions about the nature of IL-12R and the signal transductionpathways triggered by the cytokine in these cells. As for the firstissue, binding of IL-12 to B cells has been demonstrated in onestudy using SAC-preactivated peripheral blood B lymphocytes, butconflicting results have been obtained in other reports (12, 31, 32).Furthermore, it has been clearly established that human B lym-phocytes express constitutively the IL-12Rb1 chain (31, 32),whereas no information is so far available onb2 chain expression.
FIGURE 6. In vitro Ig production by tonsillar B lymphocytes preacti-vated with SAC (Fig. 7A) or IL-12 (Fig. 7B) for 48 h and subsequentlyincubated with IL-12, IL-18, or the combination of the two cytokines foradditional 7 days. Culture supernatants were harvested, and IgM, IgG, andIgA concentrations were determined by ELISA. Results (expressed in ng/ml) are means from nine experiments performed with different B cellfractions.
FIGURE 7. IFN-g secretion by tonsillar B lymphocytes preactivatedwith SAC (Fig. 8A) or IL-12 (Fig. 8B) for 48 h and subsequently incubatedwith IL-12, IL-18, or the combination of the two cytokines for 24 or 48 h.Culture supernatants were harvested and tested for IFN-g secretion byELISA. Results (expressed as pg/ml) represent the means from 12 exper-iments performed with different B cell fractions.
6885The Journal of Immunology
by guest on June 26, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
Here we show that freshly isolated tonsillar B cells, which weredevoid of contaminant T cells and NK cells, expressed mRNA ofboth IL-12R chains as assessed by RT-PCR and RPA, and thatculture with SAC or IL-12 caused a 4- to 5-fold increase in IL-12Rb2, but notb1, chain transcript. In contrast, no increase ofIL-12Rb2 gene expression was observed when B cells were incu-bated with anti-k plus anti-l Ig light chain Abs. Because SACbinds to the third variable heavy (VH3) chain region of IgM, IgG,and IgA, the different results obtained by stimulating B cells withSAC or anti-Ig Abs may appear surprising. Notably, in this con-nection, it was previously shown that malignant B cells from Bchronic lymphocytic leukemia patients produced granulocyte-CSFin vitro following stimulation with SAC but not with anti-Ig Abs(58). Because of its binding capacity to the third variable heavy(VH3) chain region of most Ig isotypes, SAC has been designatedas a B cell superantigen, and it has been proposed that it mayactivate B lymphocytes also through mechanisms unrelated to Igbinding that are so far poorly understood (59).
In view of the heterogeneity of tonsillar B cells, the expression ofIL-12Rb1 andb2 genes was investigated in freshly isolated naive,GC, and memory B lymphocytes. These experiments showed that themRNA of both chains was constitutively expressed in all subsets.
The transcripts for both IL-1Rrp and AcPL, i.e., the two chainsof the IL-18R, were induced de novo in tonsillar B cells by IL-12stimulation. Similar results have been reported in the mouse,
whereas immunophenotypic studies of human peripheral blood Bcells have indicated that the latter cells express constitutively IL-1Rrp surface protein (46). The apparent discrepancy between thesedata and our results may be related to the different sources of Blymphocytes used in the two studies and/or to the different statesof activation of peripheral blood vs tonsillar B cells.
Because IL-12 induced IL-18R expression in B cells, we nextinvestigated the effects of IL-12 and IL-18 on Ig production andIFN-g synthesis. These experiments showed that IgM was the ma-jor isotype produced by preactivated B cells in the presence ofIL-12 or IL-18. Synergistic or additive effects between the twocytokines were not observed.
IL-12 stimulation induced IFN-g synthesis and release by SAC-or IL-12-preactivated B cells in accordance with the results ofprevious studies (25, 26). IL-18 alone was poorly effective at stim-ulating IFN-g production, whereas IL-12 and IL-18 synergized inthese experimental conditions (40).
IL-12-induced IFN-g production was also investigated in sepa-rated naive, GC, and memory tonsillar B lymphocytes. These ex-periments demonstrated that naive B cells only released IFN-g inculture supernatants following IL-12 stimulation.
Such observations may lead to speculation that, in the secondarylymphoid organs where the bulk of Ag-specific immune responsestake place, the availability of IL-12 and IL-18 can switch on B cellIFN-g production and perhaps induce Th1 differentiation. However,
FIGURE 8. IFN-g synthesis by tonsillar B lymphocytes preincubated with IL-12 and IL-18 for 48 h.A, Cells were single stained by a PE-conjugatedCD20 mAb (surface staining) or a FITC-conjugated anti-IFN-g mAb (intracytoplasmic staining) and analyzed by flow cytometry. Approximately 99%CD201 and 20% intracytoplasmic IFN-g1 cells, respectively, were detected in this experiment. Control for CD20 surface staining were cells incubated with aPE-conjugated, isotype-matched mAb of irrelevant specificity (left, dashed line). Controls for intracytoplasmic IFN-g staining were cells incubated with anFITC-conjugated, isotype-matched mAb of irrelevant specificity (right, dashed line) or with the anti-IFN-g mAb preincubated with rIFN-g before staining (right,continuous line).B, Double staining of tonsillar B lymphocytes for CD20 and intracytoplasmic IFN-g. Left, No staining (,1% positive cells) was detected whenthe two fluorochrome-conjugated control mAbs of irrelevant specificity were tested together.Middle, B cells were stained with the PE-conjugated CD20 mAb andwith the FITC-conjugated, isotype-matched irrelevant mAb used as control for anti-IFN-g staining. Only CD20 single positive cells (99%) are detected. Inexperiments not shown, B cells were stained with the FITC-conjugated anti-IFN-g mAb and with the PE-conjugated isotype-matched irrelevant mAb used ascontrol for CD20 staining. Only 20% intracytoplasmic IFN-g single positive cells were detected.Right, B cells were double stained with CD20 and anti-IFN-gmAbs. The shift of the FACS profile to the right, as compared with the staining shown in the middle panel of this figure, clearly denotes the presence of a fraction(20%) of B cells positive for intracytoplasmic IFN-g. Such findings demonstrate that, under these experimental conditions, the only cells that displayed a positivestaining for intracytoplasmic IFN-g were B lymphocytes. Results refer to a representative experiment of the three performed.
6886 IL-12 AND IL-18 RECEPTORS ON HUMAN TONSILLAR B LYMPHOCYTES
by guest on June 26, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

current models of B cell activation indicate that the latter cells are com-mitted to maintain the Th1 or Th2 phenotype of already differentiatedCD41 cells rather than to promote such differentiation (8, 60).
STAT3 and STAT4 are the major transcription factors involvedin IL-12-mediated signaling to T and NK cells (18–22). An alter-native pathway of IL-12-driven signal transduction has been re-
ported for murine dendritic cells, in which NF-kB is activated byIL-12 in the absence of STAT3 or STAT4 involvement (24).
In this study, EMSA and Western blot experiments excluded theactivation of any STAT family member in tonsillar B cells exposedto IL-12. However, in experiments not shown here, specific bind-ing of nuclear extracts from IFN-a-treated B cells to the IFN-gactivation site probe was observed, indicating that tonsillar B lym-phocytes activated STAT 4 under these conditions.
We speculated that NF-kB could be a candidate family of tran-scription factors involved in IL-12-mediated signaling to B cells.NF-kB was found to be constitutively activated in freshly sepa-rated or cultured B cells, but IL-12 or IL-18 stimulation caused aclear-cut increase of NF-kB binding to DNA, thus lending supportto our hypothesis.
Cell fractionation experiments showed that IL-12-inducedNF-kB activation occurred in naive, but not in GC or memory Bcells. The reason why signal transduction in response to IL-12 wasdetected only in naive B cells, whereas expression of IL-12Rchains was demonstrated also in GC and memory B cells, is pres-ently unknown. One possibility is that posttranscriptional modifi-cations of theb1 and/orb2 IL12R chain mRNA lead to the pro-duction of the functional protein(s) in naive, but not in GC ormemory B cells. Further studies are needed to clarify this issue.
IL-12 and IL-18 share the ability to activate NF-kB (24, 41, 42)and to induce IFN-g production by T and NK cells (2). The pos-sibility that, in our experimental conditions, part of the biologicalactivities attributed to IL-12 were mediated by endogenous IL-18possibly produced by the B cells themselves is made highly un-likely because: 1) IL-12-induced B cell signaling via NF-kB wasclearly demonstrated using freshly isolated tonsillar B lympho-cytes that did not express IL-18R; and 2) IL-18 was a poor stim-ulator of IFN-g production by B cells pretreated with either SACor IL-12.
The human NF-kB family is comprised of five members, i.e.,p50, p65, c-Rel, Rel-B, and p52 (48). To investigate what NF-kBcomponents were involved in IL-12-mediated B cell signaling, gelsupershift experiments were conducted with specific Abs. It wasfound that p50 and c-Relwere the members of the NF-kB familyresponsible for IL-12-delivered signals. Interestingly, p50 andRel-B were previously identified as the proteins involved in IL-12signaling to murine dendritic cells (24).
In conclusion, this study shows that IL-12 signals to humantonsillar B cells through the NF-kB pathway by interacting withthe classic heterodimeric IL-12R composed of theb1 and b2chains. IL-12R is constitutively expressed by freshly isolated ton-sillar B cells and, in particular, by the naive, GC, and memory cellsubsets, but exposure to IL-12 activates signal transduction andIFN-g production in naive B cells only.
References1. Trinchieri, G. 1995. Interleukin-12: a proinflammatory cytokine with immuno-
regulatory functions that bridges innate resistance and antigen-specific adaptiveimmunity. Annu. Rev. Immunol. 13:251.
2. Kobayashi, M., L. Fitz, M. Ryan, R. M. Hewick, S. C. Clark, S. Chan, R. Loudon,F. Sherman, B. Perussia, and G. Trinchieri. 1989. Identification and purificationof natural killer cell stimulatory factor (NKSF), a cytokine with multiple biologiceffects on human lymphocytes.J. Exp. Med. 170:827.
3. Stern, A. S., F. J. Podlaski, J. D. Hulmes, Y. C. Pan, P. M. Quinn, A. G. Wolitzky,P. C. Familletti, D. L. Stremlo, T. Truitt, R. Chizzonite, and M. K. Gately. 1990.Purification to homogeneity and partial characterization of cytotoxic lymphocytematuration factor from human B-lymphoblastoid cells.Proc. Natl. Acad. Sci.USA 87:6808.
4. Wolf, S. F., P. A. Temple, M. Kobayashi, D. Young, M. Dicig, L. Lowe,R. Dzialo, L. Fitz, C. Ferenz, R. M. Hewick, et al. 1991. Cloning of cDNA fornatural killer stimulatory factor, a heterodimeric cytokine with multiple biologiceffects on T and natural killer cells.J. Immunol. 146:3074.
5. Manetti, R., F. Gerosa, M. G. Giudizi, R. Biagiotti, P. Parronchi, M. P. Piccinni,S. Sampognaro, E. Maggi, S. Romagnani, and G. Trinchieri. 1994. Interleukin 12
FIGURE 9. Expression of IL-12Rb1 andb2 chain mRNA, IL-12-in-duced NF-kB activation, and IFN-g production in human tonsillar B cellsubsets.A, Expression of IL-12Rb1 andb2 transcripts in freshly isolatedB cell subsets as assessed by RT-PCR.Left to right, MW, m.w. markers;negative control NC, represented by water in the place of cRNA; an Ag-specific Th1 cell clone; total tonsillar B cells; GC (IgD2, CD381), naive(IgD1), and memory (IgD2, CD382) B cells. B, EMSA conducted withnuclear extracts from freshly isolated tonsillar B cell subsets exposed toIL-12 for 30 min. The oligonucleotide has the sequence of the NF-kBbinding site of the Igl light chain enhancer.Left to right, RPMI 8866lymphoblastoid B cell line (positive control); IgD2 (GC and memory) B cellsincubated with medium (none) or IL-12; IgD1 (naive) B cells incubated withmedium (none) or IL-12. One representative experiment of the four performedwith different B cell fractions is shown.C, IFN-g secretion by tonsillar B cellsubsets incubated with IL-12 for 24 h. The IgD2 B cell fraction contains GCand memory B lymphocytes, whereas the IgD1 B cell fraction contains naiveB cells. Culture supernatants were harvested and tested for IFN-g secretion byELISA. Results (expressed as pg/ml) represent the means from four experi-ments performed with different B cell suspensions.
6887The Journal of Immunology
by guest on June 26, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

induces stable priming for interferon gamma (IFN-g) production during differ-entiation of human T helper (Th) cells and transient IFN-g production in estab-lished Th2 clones.J. Exp. Med. 179:1273.
6. Magram, J., S. E. Connaughton, R. R. Warrier, D. M. Carvajal, C.-Y. Wu,J. Ferrante, C. Stewart, U. Sarmiento, D. A. Faherty, and M. K. Gately. 1996.IL-12-deficient mice are defective in IFN-g production and type 1 cytokine re-sponses.Immunity 4:471.
7. Macatonia, S. E., N. A. Hosken, M. Litton, P. Vieira, C. Hsieh, J. A. Culpepper,M. Wysocka, G. Trinchieri, K. M. Murphy, and A. O’Garra. 1995. Dendritic cellsproduce IL-12 and direct the development of Th1 cells from naive CD41 T cells.J. Immunol. 154:5071.
8. Shultze, J. L., S. Michalak, J. Lowne, A. Wong, M. H. Gilleece, J. G. Gribben,and L. M. Nadler. 1999. Human non-germinal center B cell interleukin (IL)-12production is primarily regulated by T cell signal CD40 ligand, interferong, andIL-10: role of B cells in the maintenance of T cell responses.J. Exp. Med. 189:1.
9. Peritt, D., S. Robertson, G. Gri, M. Aste-Amezaga, and G. Trinchieri. 1998.Differentiation of human NK cells into NK1 and NK2 subsets.J. Immunol. 161:5821.
10. Chua, A. O., R. Chizzonite, B. B. Desai, T. P. Truitt, P. Nunes, L. J. Minetti,R. R. Warrier, D. H. Presky, J. F. Levine, M. K. Gately, and U. Gubler. 1994.Expression cloning of a human IL-12 receptor component.J. Immunol. 153:128.
11. Presky, D. H., Y. Hong, L. J. Minetti, A. O. Chua, N. Nabavi, C. Y. Wu,M. K. Gately, and U. Gubler. 1996. A functional interleukin 12 receptor complexis composed of twob-type cytokine receptor subunits.Proc. Natl. Acad. Sci. USA93:14002.
12. Desai, B. B., P. M. Quinn, A. G. Wolitzky, P. K. A. Mongini, R. Chizzonite, andM. K. Gately. 1992. IL-12 receptor. II. Distribution and regulation of receptorexpression.J. Immunol. 148:3125.
13. Wu, C., R. R. Warrier, X. Wang, D. H. Presky, and M. K. Gately. 1997. Regu-lation of IL-12 receptorb1 chain expression and IL-12 binding by human pe-ripheral blood mononuclear cells.Eur. J. Immunol. 27:147.
14. Szabo, S., A. S. Dighe, U. Gubler, and K. M. Murphy. 1997. Regulation of theinterleukin (IL)-12Rb2 subunit expression.J. Exp. Med. 185:817.
15. Gollob, J. A., H. Kawasaki, and J. Ritz. 1997. Interferon-g and interleukin-4regulate T cell interleukin-12 responsiveness through the differential modulationof high-affinity interleukin-12 receptor expression.Eur. J. Immunol. 3:647.
16. Igarashi, O., H. Yamane, S. Imajoh-Ohmi, and H. Nariuchi. 1998. IL-12 receptor(IL-12R) expression and accumulation of IL-12Rb1 and IL-12Rb2 mRNAs inCD41 T cells by costimulation with B7-2 molecules.J. Immunol. 160:1638.
17. Rogge, L., A. Papi, D. H. Presky, M. Biffi, L. J. Minetti, D. Miotto, C. Agostini,G. Semenzato, L. M. Fabbri, and F. Sinigaglia. 1999. Antibodies to the IL-12receptorb2 chain mark human Th1 but not Th2 cells in vitro and in vivo.J. Im-munol. 162:3926.
18. Bacon, C. M., E. F. Petricoin, J. R. Ortaldo, R. C. Rees, A. C. Larner,J. A. Johnston, and J. J. O’Shea. 1995. IL-12 induces tyrosine phosphorylationand activation of STAT4 in human lymphocytes.Proc. Natl. Acad. Sci. USA92:7307.
19. Cho, S. S., C. M. Bacon, C. Sudarshan, R. C. Rees, D. Flimbloom, R. Pine, andJ. J. O’Shea. 1996. Activation of STAT4 by IL-12 and IFN-a: evidence for theinvolvement of ligand-induced tyrosine and serine phosphorylation.J. Immunol.157:4781.
20. Murphy, K. M., T. L. Murphy, S. J. Szabo, N. J. Jacobson, M. L. Guler,J. D. Gorham, and U. Gubler. 1997. Regulation of IL-12 receptor in early T-helper responses implies two phases of Th1 differentiation: capacitance and de-velopment.Chem. Immunol. 68:54.
21. Thierfelder, W. E., J. M. van Deursen, K. Yamamoto, R. A. Tripp, S. R. Sasawar,R. T. Carson, M. Y. Sangster, D. A. Vignali, P. C. Doherty, G. C. Grosveld, andJ. N. Ihle. 1996. Requirement for STAT-4 in interleukin-12-mediated responsesof natural killer and T cells.Nature 382:171.
22. Kaplan, M. H., Y. L. Sun, T. Heey, and M. J. Grusby. 1996. Impaired IL-12responses and enhanced development of Th2 cells in STAT4 deficient mice.Nature 382:174.
23. Leonard, W. J., and J. J. O’Shea. 1998. JAKs and STATs: biological implica-tions.Annu. Rev. Immunol. 16:293.
24. Grohmann, U., M. L. Belladonna, R. Bianchi, C. Orabona, E. Ayroldi,M. C. Fioretti, and P. Puccetti. 1998. IL-12 acts directly on DC to promotenuclear localization of NF-kB and primes DC for IL-12 production.Immunity.9:315.
25. Metzger, D. W., L. A. Vogel, V. H. Van Cleave, T. L. Lester, andJ. M. Buchanan. 1995. The effects of IL12 on B cell subset function.Res. Im-munol. 147:499.
26. Vogel, L. A., T. L. Lester, V. H. Van Cleave, and D. W. Metzger. 1996. Inhi-bition of murine B1 lymphocytes by interleukin-12.Eur. J. Immunol. 26:219.
27. Jones, B. M. 1996. Effect of IL-12 neutralizing anti-cytokine antibodies on invitro activation of B-cells: interleukin-12 is required by B1a but not B2 cells.Scand. J. Immunol. 43:64.
28. Jelinek, D. F., and J. K. Braaten. 1995. Role of IL-12 in human B lymphocyteproliferation and differentiation.J. Immunol. 154:1606.
29. Li, L., D. Young, S. F. Wolf, and Y. S. Choi. 1996. Interleukin-12 stimulates Bcell growth by inducing IFN-g. Cell. Immunol. 168:133.
30. Dubois, B., C. Massacrier, B. Vanbervliet, J. Faiette, F. Briere, J. Banchereau,and C. Caux. 1998. Critical role of IL-12 in dendritic cell-induced differentiationof naive B lymphocytes.J. Immunol. 161:2223.
31. Vogel, L. A., L. C. Showe, T. L. Lester, R. M. McNutt, V. H. Van Cleave, andD. W. Metzger. 1996. Direct binding of IL-12 to human and murine B lympho-cytes.Int. Immunol. 8:1955.
32. Benjamin, B., V. Sharma, M. Kubin, J. L. Klein, A. Sartori, J. Holliday, andG. Trinchieri. 1996. IL-12 expression in AIDS-related lymphoma B cell lines.J. Immunol. 156:1626.
33. Okamura, H., T. Tsutsui, T. Komatsu, M. Yutsudo, A. Hakura, T. Tanimoto,K. Torigoe, T. Okura, Y. Nukada, K. Harttori, et al. 1995. Cloning of a newcytokine that induces IFN-g production by T cells.Nature 378:88.
34. Stoll, S., G. Muller, M. Kurimoto, J. Saloga, T. Tanimoto, H. Yamauchi,H. Okamura, J. Knop, and A. H. Enk. 1997. Production of IL-18 (IFN-g inducingfactor) messenger RNA and functional protein by murine keratinocytes.J. Im-munol. 159:198.
35. Conti, B., J. Jahng, C. Tinti, J. Son, and T. Joh. 1997. Induction of IFN-g in-ducing factor in the adrenal cortex.J. Biol. Chem. 264:2025.
36. Tsutsui, H., K. Matsui, N. Kawada, Y. Hyodo, N. Hayashi, H. Okamura,K. Higashino, and K. Nakanishi. 1997. IL-18 accounts for both TNF-a and FasL-mediated hepatotoxic pathways in endotoxin-induced liver injury.J. Immunol.159:3961.
37. Ghayur, T., S. Banerjee, M. Hugunin, D. Butler, L. Herzog, A. Carter, L. Quinta,L. Sekut, R. Talanian, M. Paskind, et al. 1997. Caspase-1 processes IFN-g in-ducing factor and regulates LPS-induced IFN-g production.Nature 386:619.
38. Gu, Y., K. Kuida, H. Tsutsui, G. Ku, K. Hsiao, M. Fleming, H. Hayashi,K. Higashino, H. Okamura, K. Nakanishi, et al. 1997. Activation of IFN-g in-ducing factor mediated by interleukin-1b converting enzyme.Science 275:206.
39. Yoshimoto, T., K. Takeda, T. Tanaka, K. Ohkusu, S. Kashiwamura, H. Okamura,S. Akira, and K. Nakanishi. 1998. IL-12 up-regulates IL-18 receptor expressionon T cells, Th1 cells, and B cells: synergism with IL-18 for IFN-g production.J. Immunol. 161:3400.
40. Barbulescu, K., C. Becker, J. F. Schlaak, E. Schmitt, K.-H. Meyer zum Buschen-felde, and M. F. Neurath. 1998. Cutting edge: IL-12 and IL-18 differentiallyregulate the transcriptional activity of the human IFN-g promoter in primaryCD41 T lymphocytes.J. Immunol. 160:3642.
41. Robinson, D., K. Shibuia, A. Mui, F. Zonin, E. Murphy, T. Sana, S. B. Hartley,S. Menon, R. Kalstelein, F. Bazan, and A. O’Garra. 1997. IGIF does not driveTh1 development but synergizes with IL-12 for interferon-g production and ac-tivates IRAK and NFkB. Immunity 7:571.
42. Sica, A., L. Dorman, V. Viggiano, M. Cippitelli, P. Ghosh, N. Rice, andH. A. Young. 1997. Interaction of NF-kB and NFAT with the interferon-g pro-moter.J. Biol. Chem. 373:30412.
43. Torigoe, K., S. Ushio, T. Okura, S. Kobayashi, M. Taniai, T. Kunikata,T. Murakami, O. Sanou, H. Kojima, M. Fujii, et al. 1997. Purification and char-acterization of the human interleukin-18 receptor.J. Biol. Chem. 272:25737.
44. Born, T. L., E. Thomassen, T. A. Bird, and J. E. Sims. 1998. Cloning of a novelreceptor subunit, AcPL, required for interleukin-18 signaling.J. Biol. Chem. 272:29445.
45. Kunikata, T., K. Torigoe, S. Ushio, T. Okura, C. Ushio, H. Yamauchi, M. Ikeda,H. Ikegami, and M. Kurimoto. 1998. Constitutive and induced IL-18 receptorexpression by various peripheral blood cell subsets as determined by anti-hIL-18R monoclonal antibody.Cell. Immunol. 189:135.
46. Lagresle, C., C. Bella, and T. Defrance. 1993. Phenotypic and functional heter-ogeneity of the IgD2 B cell compartment: identification of two major tonsillar Bcell subsets.Int. Immunol. 5:1259.
47. Schreiber, E., P. Matthias, M. M. Muller, and W. Schaffner. 1989. Rapid detec-tion of octamer binding proteins with “mini-extracts”, prepared from a smallnumber of cells.Nucleic Acids Res. 17:6419.
48. Ghosh, S., M. J. May, and E. B. Kopp. 1998. NFkB and Rel proteins: evolu-tionary conserved mediators of immune responses.Annu. Rev. Immunol. 16:225.
49. Liu, Y. J., D. E. Joshua, G. T. Williams, C. A. Smith, J. Gordon, andI. C. M. MacLennan. 1989. Mechanism of antigen-driven selection in germinalcenters.Nature 342:929.
50. Berek, C., A. Berger, and M. Apel. 1991. Maturation of immune responses ingerminal centers.Cell 67:1121.
51. Leanderson, T., E. Kallberg, and D. Gray. 1992. Expansion, selection and mu-tation of antigen-specific B cells in germinal centers.Immunol. Rev. 126:47.
52. MacLennan, I. C. M. 1994. Germinal centers.Annu. Rev. Immunol. 12:117.53. Pascual, V., Y. J. Liu, A. Magalski, O. de Bouteiller, J. Banchereau, and
J. D. Capra. 1994. Analysis of somatic mutation in five B cell subsets of humantonsil. J. Exp. Med. 180:329.
54. Szakal, A. K. H., M. Kosko, and J. G. Tew. 1989. Microanatomy of lymphoidtissues during humoral immune responses: structure function relationships.Annu.Rev. Immunol. 7:91.
55. Liu, Y. D., C. Barthelemy, O. de Bouteiller, C. Arpin, I. Durand, andJ. Banchereau. 1995. Memory B cells from human tonsils colonize mucosal ep-ithelium and directly present antigen to T cell by rapid up-regulation of B7-1 andB7-2. Immunity 2:239.
56. Kiniwa, M., M. Gately, U. Gubler, R. Chizzonite, C. Fargeas, and G. Delespesse.1992. Recombinant interleukin-12 suppresses the synthesis of immunoglobulin Eby interleukin-4 stimulated human lymphocytes.J. Clin. Invest. 90:262.
57. Morris, S. C., K. B. Madden, J. J. Adamovicz, W. C. Gause, B. R. Hubbard,M. K. Gately, and F. D. Finkelman. 1994. Effects of IL-12 on in vivo cytokinegene expression and Ig isotype selection.J. Immunol. 152:1047.
58. Corcione, A., M. V. Corrias, S. Daniele, S. Zupo, M. Spriano, and V. Pistoia.1996. Expression of granulocyte colony-stimulating factor and granulocyte col-ony-stimulating factor receptor genes in partially overlapping monoclonal B-cellpopulations from chronic lymphocytic leukemia patients.Blood 87:2861.
59. Silverman, G. J. 1997. B-cell superantigens.Immunol. Today 18:379.60. Skok, J., J. Poudrier, and D. Gray. 1999. Dendritic cell-derived IL-12 promotes
B cell induction of Th2 differentiation: a feedback regulation of Th1 develop-ment.J. Immunol. 163:4284.
6888 IL-12 AND IL-18 RECEPTORS ON HUMAN TONSILLAR B LYMPHOCYTES
by guest on June 26, 2015http://w
ww
.jimm
unol.org/D
ownloaded from




















