
Evaluation of the Binding of Radiolabeled Rituximab to CD20Positive Lymphoma Cells: An In Vitro...
11
CANCER BIOTHERAPY & RADIOPHARMACEUTICALS Volume 22, Number 4, 2007 © Mary Ann Liebert, Inc. DOI: 10.1089/cbr.2007.371 Evaluation of the Binding of Radiolabeled Rituximab to CD20-Positive Lymphoma Cells: An In Vitro Feasibility Study Concerning Low-Dose-Rate Radioimmunotherapy with the -Emitter 227 Th Katrine B. Melhus, Roy H. Larsen, Trond Stokke, Olav Kaalhus, Pål K. Selbo, and Jostein Dahle Department of Radiation Biology, Institute for Cancer Research, The Norwegian Radium Hospital, Montebello, Oslo, Norway ABSTRACT Radioimmunotherapy (RIT) with the alpha-emitter 227 Th is currently under evaluation. 227 Th is conjugated to the chimeric anti-CD20 monoclonal antibody rituximab, using the chelator p-isothiocyanato-benzyl-DOTA. In this study, the binding of 227 Th-DOTA-p-benzyl-rituximab to three different CD-20-positive lymphoma cell lines, Raji, Rael, and Daudi, were evaluated. Equilibrium and kinetic binding experiments were used to de- termine binding parameters, including the association and dissociation rate constants, the equilibrium disso- ciation constants, and the total number of antigens for Raji, Rael, and Daudi cells. There were significant dif- ferences between the cell lines with respect to both K d and the total number of antigens. Rael cells had more than three times as many antigens as the other two cell lines, and the functional K d found for Rael cells was significantly higher than that found for Raji and Daudi cells. These results were confirmed using flow cy- tometry. Rituximab was found to be localized in patches on the cell membrane. The findings indicated that 227 Th-labeled rituximab has relevant antigen-targeting properties for radioimmunotherapy. Key words: -particle-radioimmunotherapy, antibody-binding parameters, antibody affinity, 227 Th, rituximab 469 INTRODUCTION Monoclonal antibodies (MAbs) labeled with al- pha-emitting radionuclides are a new potential treatment modality of disseminated and metasta- tic cancer. 1–3 Alpha particles have a short track length of typically less than 100 m, high rela- tive biological effectiveness, and a low oxygen enhancement ratio, which may give therapeutic advantages over radiation qualities of lower lin- ear energy transfer (LET). 4 However, the alpha- emitting radionuclides tested so far are challeng- ing to apply in clinical settings because of their short half-lives, low production capability, and/or limited supply of source material. 2,5 Cationic ra- dium-223 (t 1/2 11.4 d) has shown considerable promise in clinical trials in patients with skeletal metastases. 6,7 Although 223 Ra can be stably re- tained in liposomes, it cannot currently be stably conjugated to mAbs. 8,9 We are now testing Address reprint requests to: Jostein Dahle; Department of Radiation Biology, Institute for Cancer Research, The Nor- wegian Radium Hospital; Building J, Montebello, 0310 Oslo, Norway; Tel.: 47 22 78 12 03; Fax: 47 22 78 12 07 E-mail: [email protected]
-
Upload
rr-research -
Category
Documents
-
view
0 -
download
0
Transcript of Evaluation of the Binding of Radiolabeled Rituximab to CD20Positive Lymphoma Cells: An In Vitro...

CANCER BIOTHERAPY & RADIOPHARMACEUTICALSVolume 22, Number 4, 2007© Mary Ann Liebert, Inc.DOI: 10.1089/cbr.2007.371
Evaluation of the Binding of Radiolabeled Rituximabto CD20-Positive Lymphoma Cells: An In VitroFeasibility Study Concerning Low-Dose-RateRadioimmunotherapy with the �-Emitter 227Th
Katrine B. Melhus, Roy H. Larsen, Trond Stokke, Olav Kaalhus, Pål K. Selbo, and Jostein DahleDepartment of Radiation Biology, Institute for Cancer Research, The Norwegian Radium Hospital,Montebello, Oslo, Norway
ABSTRACT
Radioimmunotherapy (RIT) with the alpha-emitter 227Th is currently under evaluation. 227Th is conjugated tothe chimeric anti-CD20 monoclonal antibody rituximab, using the chelator p-isothiocyanato-benzyl-DOTA.In this study, the binding of 227Th-DOTA-p-benzyl-rituximab to three different CD-20-positive lymphoma celllines, Raji, Rael, and Daudi, were evaluated. Equilibrium and kinetic binding experiments were used to de-termine binding parameters, including the association and dissociation rate constants, the equilibrium disso-ciation constants, and the total number of antigens for Raji, Rael, and Daudi cells. There were significant dif-ferences between the cell lines with respect to both Kd and the total number of antigens. Rael cells had morethan three times as many antigens as the other two cell lines, and the functional Kd found for Rael cells wassignificantly higher than that found for Raji and Daudi cells. These results were confirmed using flow cy-tometry. Rituximab was found to be localized in patches on the cell membrane. The findings indicated that227Th-labeled rituximab has relevant antigen-targeting properties for radioimmunotherapy.
Key words: �-particle-radioimmunotherapy, antibody-binding parameters, antibody affinity, 227Th,rituximab
469
INTRODUCTION
Monoclonal antibodies (MAbs) labeled with al-pha-emitting radionuclides are a new potentialtreatment modality of disseminated and metasta-tic cancer.1–3 Alpha particles have a short tracklength of typically less than 100 �m, high rela-
tive biological effectiveness, and a low oxygenenhancement ratio, which may give therapeuticadvantages over radiation qualities of lower lin-ear energy transfer (LET).4 However, the alpha-emitting radionuclides tested so far are challeng-ing to apply in clinical settings because of theirshort half-lives, low production capability, and/orlimited supply of source material.2,5 Cationic ra-dium-223 (t1/2 � 11.4 d) has shown considerablepromise in clinical trials in patients with skeletalmetastases.6,7 Although 223Ra can be stably re-tained in liposomes, it cannot currently be stablyconjugated to mAbs.8,9 We are now testing
Address reprint requests to: Jostein Dahle; Department ofRadiation Biology, Institute for Cancer Research, The Nor-wegian Radium Hospital; Building J, Montebello, 0310 Oslo,Norway; Tel.: �47 22 78 12 03; Fax: �47 22 78 12 07E-mail: [email protected]

whether 227Th, the parent of 223Ra, is suitable forradioimmunotherapy (RIT). Thorium-227 has ahalf-life of 18.7 days and can be conjugated torituximab by using the bifunctional chelator p-isothiocyanato-benzyl-DOTA. Long-term oper-ating generators for 227Th can be prepared. 227Th-DOTA-p-benzyl-rituximab is stable both in vitroand in vivo.10 Low initial activity levels of 227Th-DOTA-p-benzyl-rituximab inhibit growth ofCD20 positive cells in vitro.10 The long half-lifeof 227Th may permit the administration and tar-geting of a 227Th-labeled radioimmunoconjugate(RIC) to occur before a significant amount of223Ra is generated. The relatively long half-lifeof 223Ra, in turn, assures that this nuclide wouldbe largely excreted or trapped in skeletal hy-droxyapatite before decay occurs.6 Although aconsiderable amount of 223Ra is taken up inbone,10 this will probably not cause bone marrowtoxicity because of the short range of the alpha-particles.11 Thus, a therapeutic window may ex-ist, which permits therapy with 227Th.
To assess the potential of 227Th in �-RIT, thebinding to antigen-positive cells is important toevaluate. Because of the relatively long half-lifeof 227Th, the dose rate would probably have to belower than for other �-emitters considered for ther-apy. It is, therefore, necessary to have a high affin-ity RIC and a specific activity that ensures thatenough 227Th-atoms decay at the cells. This paperrefers to CD20-positive lymphoma cells targetedwith rituximab as the experimental system.
MATERIALS AND METHODS
Cell Lines
Three B-cell lymphoma lines, Raji, Rael, andDaudi, and one B-cell leukemia line, Reh, wereused. The B-cell lymphoma cell lines express theCD20 antigen, whereas the Reh cells are CD20negative and were used as control cells. Singlecell suspensions were grown in RPMI 1640medium (PAA; Linz, Austria), supplementedwith 13% heat-inactivated fetal calf serum (FCS)(PAA), 1% L-glutamine (PAA), and 1% peni-cillin-streptomycin (PAA) in a humid atmospherewith 95% air/5% CO2.
Radiolabeling of Rituximab
The mAb rituximab (MabThera®; Roche, BaselSwitzerland) targeting CD20 was labeled with227Th. The preparation of 227Th-DOTA complexes
and the conjugation of 227Th-p-isothiocyanato-ben-zyl-DOTA to rituximab has been described be-fore.10 Briefly, 227Ac was produced through thethermal neutron irradiation of 226Ra followed by ��
decay of 227Ra (t1/2 � 42.2 m) to 227Ac.3 227Th wasselectively retained from a 227Ac decay mixture in7 M of HNO3 solution by anion exchange chro-matography.8 After 227Ac, 223Ra and its daughtershad eluted from the column, and 227Th was ex-tracted from the column with 12 M of HCl. The227Th, prepared this way, was more than 99.93%pure. The 227Th-DOTA complex was prepared byadding 20 �L of a 10-mg/mL solution of p-SCN-benzyl-DOTA (Macrocyclics Inc.; Dallas, TX) to asolution containing 20 �L (150 mg/mL) of L-ascor-bic acid (Sigma-Aldrich Co. Ltd; Gillingham, UK)and 20–150 �L of 100–300 MBq/mL of 227Th inHCl in a 2-mL glass vial. Then, 1–2 mg of ritux-imab in 100–200 �L was added to the 227Th andp-SCN-benzyl-DOTA reaction. Thereafter, the re-action mixture was incubated for 40 minutes at ap-proximately 55°C by using a Thermomixer Com-fort (Eppendorf AG; Hamburg, Germany). Thesolution was cooled down to 37°C, and the pH wasadjusted to 8–9 by adding 1 M of NaCO3/NaHCO3buffer. Then, the rituximab was added. After 45minutes of reaction, 20 �L of saturated diethylenetriamine pentaacetic acid (DTPA, Fluka Chemie,Buchs, Switzerland) was added and the mixture fur-ther incubated for 5 minutes. Thereafter, the reac-tion mixture was purified by gel filtration (Econo-Pac10 DG; Bio-Rad, Hercules, CA) and eluted with1% bovine serum albumin (BSA; Sigma ChemicalCo., St Louis, MO) in phosphate-buffered salinePBS; pH, 7.4. Finally, the purified product was ster-ile filtered (Millex GV-13; Millipore Co., Bedford,MA) into a sterile 10-mL glass vial (Wheaton; Mill-ville, NJ), which was subsequently capped with asterile rubber cap (Sigma; St. Louis, MO). The spe-cific activity of the RIC was typically 650–5300Bq/�g.
Quantification of 227Th
Quantification of 227Th can be problematic in thepresence of 223Ra. Therefore, the RIC was alwayspurified by gel filtration, as described above, be-fore the binding experiments. The activity wascounted in a gamma counter (Cobra Auto-Gamma; Packard; Downers Grove, IL).
Measurement of Immunoreactive Fraction
The quality of the RIC was measured using lym-phoma cells and cell concentrations up to 108 Raji
470

cells/ml to compensate for the modest specificactivity and counting efficacy of the RIC.10,13
The immunoreactive fraction (IRF) of 227Th-p-isothiocyanato-benzyl-DOTA-rituximab wasbetween 50 and 70%.
Equilibrium Binding Experiments
Specific binding was measured at seven differ-ent concentrations of RIC to determine the to-tal number of antigens, Bmax, and the equilib-rium dissociation constant, Kd, by Scatchardanalysis.14,15 Two (2) million cells in 0.4 mLof PBS were incubated with 10–50,000 ng/mLof RIC for 2 hours in a shaker at 4°C. Then, thecells were washed twice with PBS containing1% FCS, and the activity was counted in agamma counter (Packard). Both cells and su-pernatant were counted. Nonspecific bindingwas measured by blocking with cold rituximab.The analysis of equilibrium binding experi-ments by the Scatchard method has been ex-plained before.14,15 Briefly, specific binding, B,divided by the concentration of free antibody,[Ab], is plotted versus specific binding. Theslope of this plot equals �1/Kd and the inter-cept with the x-axis gives the total number ofantigens, Bmax, directly:
��1�Kd
� B � (1)
Binding of Cold Rituximab
Cells were incubated with unlabeled rituximab(Roche, Basel, Switzerland) or herceptin (Roche)(irrelevant mAb) for 1 hour in PBS at 4°C, andwashed twice with PBS. Then, the cells were in-cubated with fluorescein isothiocyanate (FITC)-labeled mouse-anti-human Ab (BD Biosciences;Erembodegen, Belgium) for 0.5 hours in PBS at4°C. Subsequently, the cells were washed twice,and the fluorescence was measured on viablecells in a FACSCalibur flowcytometer (BD Bio-sciences). Dead cells were excluded by addingpropidium iodide (PI; Calbiochem, La Jolla, CA)to the cells directly before flow cytometry andgating on the PI-negative cells.
Fluorescence Labeling of Rituximab
Rituximab was labeled with the fluorescing dye,Alexa Fluor 488 (hereby named Alexa-ritux-imab), according to the instructions of the man-ufacturer (Sigma).
Bmax�Kd
B�[Ab]
Measurements of Binding Using Alexa-Rituximab
Raji and Rael cells were incubated with 0.1�g/mL of Alexa-rituximab for 24 hours at 37°C.The cells were then washed twice with PBS andstudied by using 100� magnification and a flu-orescence microscope (AX70; Olympus, Ham-burg, Germany) equipped with a charge-coupleddevice (CCD) camera. PI was used to excludedead cells.
In another experiment, 5 million Rael cellswere stained with 2 �g/mL of Hoechst 33342 for0.5 hours at 37°C, then washed and mixed with5 million unstained Raji cells. Subsequently, thecell suspension was incubated with 0.01–100�g/mL of Alexa-rituximab for 1 hour at 4°C, thenwashed and analyzed by flow cytometry by gat-ing on the Hoechst 33342 fluorescence (Stokke,unpublished date). The experiment was per-formed three times and each time the cell linestained with Hoechst 33342 was switched.
Dissociation Rate Constant
The dissociation rate constant kd was found byallowing RIC to bind to cells for 1.5 hours at 4°C,and then further binding was blocked by addingunlabeled rituximab in excess. Five (5) millioncells per mL of PBS with 50–300 ng/mL of RICwere incubated for 1.5 hours and washed threetimes with PBS and then resuspended in freshmedium with 100 �g/mL of unlabeled rituximab.Activity was counted in a gamma counter(Packard) before the addition of unlabeled ritux-imab. Subsequently, three 0.4-mL aliquots weresubtracted at several time points, and cell-boundactivity was measured. Nonspecific binding wasmeasured, using the same cells blocked with 25�g/mL of rituximab. The dissociation rate con-stant was determined by plotting total bindingversus time and fitting the data to Equation 2 us-ing Sigma plot (version 7; SPSS; San Jose, CA).
Btot � Bunspecific � a � e�kd � t � b � e�kd2 � t (2)
Association Rate Constant
Two (2) million cells in 0.4 mL of PBS were in-cubated from 5 minutes to 3.5 hours at 4°C with50–300 ng/mL of RIC, washed twice with PBSas for the Scatchard analysis, and binding wasmeasured in a gamma counter (Cobra Auto-Gamma, Packard). Nonspecific binding was mea-sured by using the same cells blocked with 25�g/mL of rituximab. The observed rate of asso-
471

ciation, kobs, was determined by plotting specificbinding versus time and fitting the data to Equa-tion 3, using the Sigma plot (version 7, SPSS):
B � Ymax � (1 � e�kobs � t) (3)
Here, kobs is a measure of how long it takes toreach equilibrium between association and dis-sociation. Thus, it depends on the associationrate, ka, the dissociation rate, kd, and the start con-centrations of Ab and Ag, [Ab]0 and Bmax, re-spectively. That is, kobs � ka([Ab]0 � Bmax) �kd. Consequently, an expression for ka can befound:
ka � (4)
Equations 3 and 4 can also be found as approxi-mations of the solution of the differential equa-tion describing the net rate of formation of theantigen-antibody complex.16
RESULTS
Scatchard Analysis
The specific activity of a RIC affects the mea-surements of IRF and may also affect the mea-surements of affinity and number of binding sites.Affinity and number of binding sites were mea-sured using the Scatchard analysis and three dif-ferent CD20-positive cell lines: Raji, Rael, andDaudi. Figure 1 shows representative Scatchardexperiments, and Table 1 shows the resulting
kobs � kd��[Ab]0 � Bmax
mean number of antigens, Bmax, and the meanequilibrium dissociation constant, Kd, for thethree different cell lines.
The specific activity obtained for 227Th-DOTA-p-benzyl-rituximab was typically approx-imately 1 kBq/�g, which was low, comparedwith most radioimmunoconjugates.1–3 However,in the prescribing information for rituximab, theaffinity is 8 nM, which is similar to our data forRaji and Daudi cells, indicating that the low spe-cific activity and the labeling procedure did notalter the binding properties of the antibody. Fur-thermore, results from the Scatchard experimentswith 125I-rituximab did not differ considerablyfrom the results for 227Th-DOTA-p-benzyl-ritux-imab (not shown). Thus, the values found for227Th-DOTA-p-benzyl-rituximab seem to be ofthe right order of magnitude.
Significant differences were found between theRael cell line and the two other cell lines. Therewere 2.5 times more available antigens on theRael cells than on the Raji and Daudi cells, andthe Kd was 2.3 times higher for Rael cells thanfor Raji and Daudi cells.
Binding of Unlabeled Rituximab and Alexa-Rituximab to CD20-PositiveLymphoma Cells
The binding of unlabeled rituximab and Alexa-Rituximab was assessed using flow cytometryand fluorescence microscopy. Figure 2 shows thebinding of 0.1 �g/mL of unlabeled rituximab toRaji and Rael lymphoma cells. There was no dif-ference in fluorescence signal from herceptin,rituximab, or secondary antibody-labeled CD20-
472
Figure 1. Scatchard analysis. Representative plots of specific binding/free antibody (Ab) concentration versus number of anti-gens per cell for (A) Raji, (B) Rael, and (C) Daudi cells labeled with 227Th-DOTA-p-benzyl-rituximab.
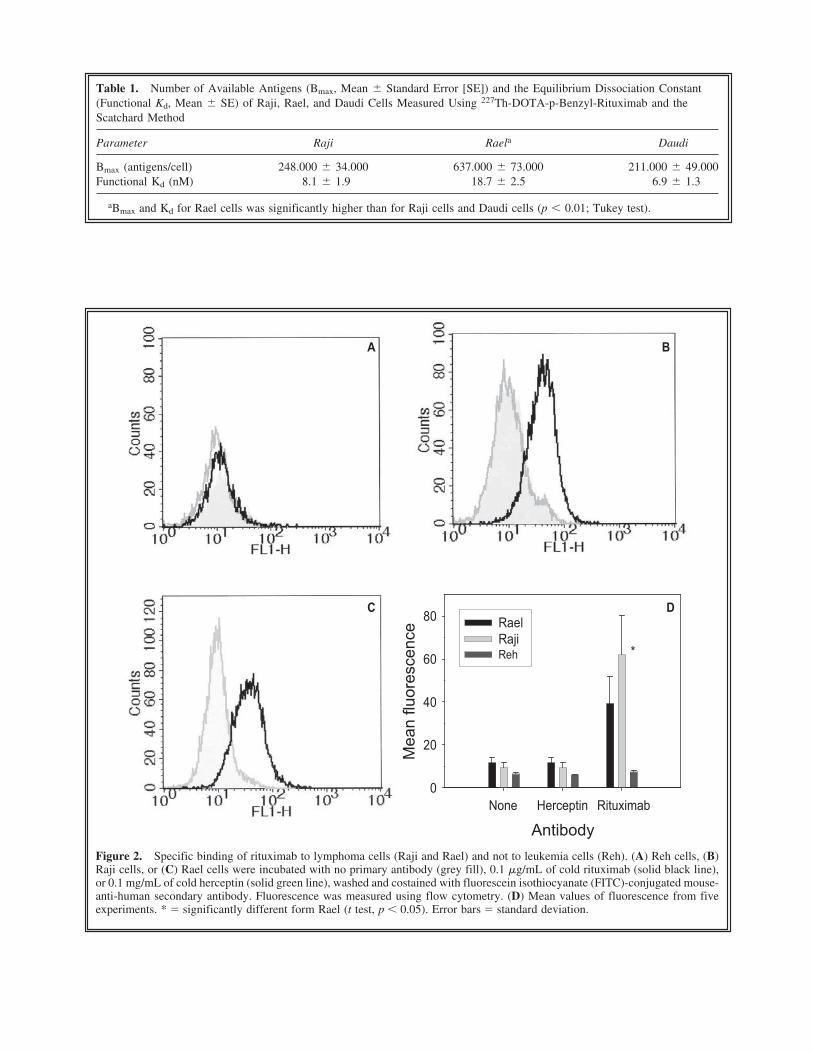
Figure 2. Specific binding of rituximab to lymphoma cells (Raji and Rael) and not to leukemia cells (Reh). (A) Reh cells, (B)Raji cells, or (C) Rael cells were incubated with no primary antibody (grey fill), 0.1 �g/mL of cold rituximab (solid black line),or 0.1 mg/mL of cold herceptin (solid green line), washed and costained with fluorescein isothiocyanate (FITC)-conjugated mouse-anti-human secondary antibody. Fluorescence was measured using flow cytometry. (D) Mean values of fluorescence from fiveexperiments. * � significantly different form Rael (t test, p � 0.05). Error bars � standard deviation.
Table 1. Number of Available Antigens (Bmax, Mean Standard Error [SE]) and the Equilibrium Dissociation Constant(Functional Kd, Mean SE) of Raji, Rael, and Daudi Cells Measured Using 227Th-DOTA-p-Benzyl-Rituximab and theScatchard Method
Parameter Raji Raela Daudi
Bmax (antigens/cell) 248.000 34.000 637.000 73.000 211.000 49.000Functional Kd (nM) 8.1 1.9 18.7 2.5 6.9 1.3
aBmax and Kd for Rael cells was significantly higher than for Raji cells and Daudi cells (p � 0.01; Tukey test).
negative Reh cells (Fig. 2A). Raji and Rael cellslabeled with Rituximab fluoresced significantlystronger than cells labeled with herceptin or thesecondary antibody only (Fig. 2B and 2C, t test,p � 0.05). The fluorescence signal from Rajicells was significantly stronger than from Raelcells. Similar results as for Raji cells were foundfor Daudi cells (not shown). The antigen expres-sion for all three cell lines resembled log normaldistributions and varied with at least a factor of50 between the cells with the lowest and the cellswith the highest antigen expression. The antigenexpression of rituximab-labeled cells overlappedwith the herceptin background distribution for thelower fluorescence intensities.
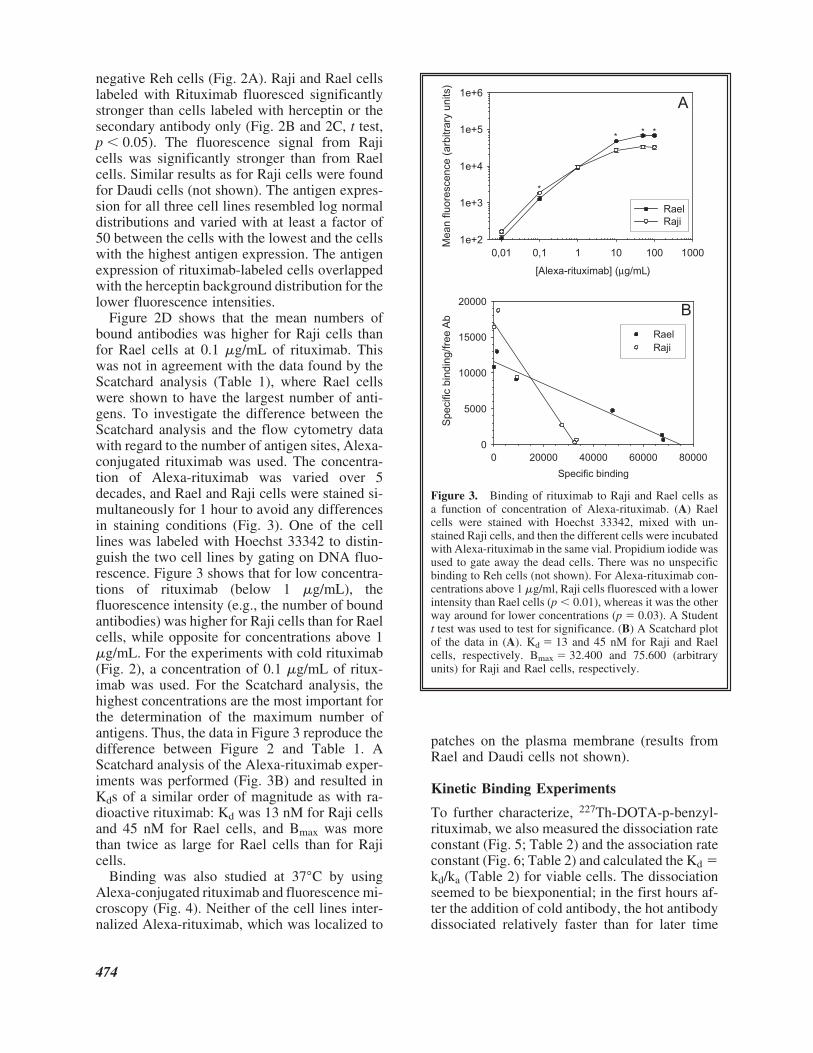
Figure 2D shows that the mean numbers ofbound antibodies was higher for Raji cells thanfor Rael cells at 0.1 �g/mL of rituximab. Thiswas not in agreement with the data found by theScatchard analysis (Table 1), where Rael cellswere shown to have the largest number of anti-gens. To investigate the difference between theScatchard analysis and the flow cytometry datawith regard to the number of antigen sites, Alexa-conjugated rituximab was used. The concentra-tion of Alexa-rituximab was varied over 5decades, and Rael and Raji cells were stained si-multaneously for 1 hour to avoid any differencesin staining conditions (Fig. 3). One of the celllines was labeled with Hoechst 33342 to distin-guish the two cell lines by gating on DNA fluo-rescence. Figure 3 shows that for low concentra-tions of rituximab (below 1 �g/mL), thefluorescence intensity (e.g., the number of boundantibodies) was higher for Raji cells than for Raelcells, while opposite for concentrations above 1�g/mL. For the experiments with cold rituximab(Fig. 2), a concentration of 0.1 �g/mL of ritux-imab was used. For the Scatchard analysis, thehighest concentrations are the most important forthe determination of the maximum number ofantigens. Thus, the data in Figure 3 reproduce thedifference between Figure 2 and Table 1. AScatchard analysis of the Alexa-rituximab exper-iments was performed (Fig. 3B) and resulted inKds of a similar order of magnitude as with ra-dioactive rituximab: Kd was 13 nM for Raji cellsand 45 nM for Rael cells, and Bmax was morethan twice as large for Rael cells than for Rajicells.
Binding was also studied at 37°C by usingAlexa-conjugated rituximab and fluorescence mi-croscopy (Fig. 4). Neither of the cell lines inter-nalized Alexa-rituximab, which was localized to
patches on the plasma membrane (results fromRael and Daudi cells not shown).
Kinetic Binding Experiments
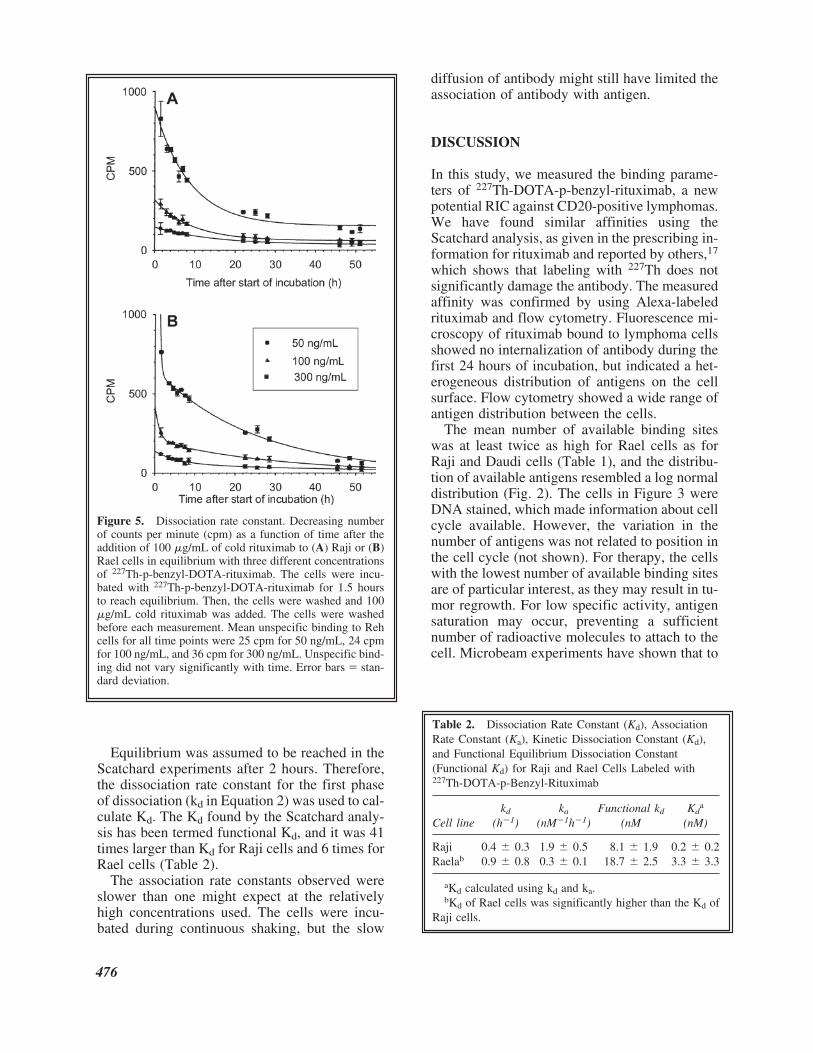
To further characterize, 227Th-DOTA-p-benzyl-rituximab, we also measured the dissociation rateconstant (Fig. 5; Table 2) and the association rateconstant (Fig. 6; Table 2) and calculated the Kd �kd/ka (Table 2) for viable cells. The dissociationseemed to be biexponential; in the first hours af-ter the addition of cold antibody, the hot antibodydissociated relatively faster than for later time
474
Figure 3. Binding of rituximab to Raji and Rael cells asa function of concentration of Alexa-rituximab. (A) Raelcells were stained with Hoechst 33342, mixed with un-stained Raji cells, and then the different cells were incubatedwith Alexa-rituximab in the same vial. Propidium iodide wasused to gate away the dead cells. There was no unspecificbinding to Reh cells (not shown). For Alexa-rituximab con-centrations above 1 �g/ml, Raji cells fluoresced with a lowerintensity than Rael cells (p � 0.01), whereas it was the otherway around for lower concentrations (p � 0.03). A Studentt test was used to test for significance. (B) A Scatchard plotof the data in (A). Kd � 13 and 45 nM for Raji and Raelcells, respectively. Bmax � 32.400 and 75.600 (arbitraryunits) for Raji and Rael cells, respectively.
points (Fig. 5). The half-life for the first phase ofdissociation was 1.8 hours and 46 minutes,whereas it was 10.3 hours and 28.7 hours for thesecond phase, for Raji and Rael cells, respec-tively. Notably, Figure 5 shows that even 2 daysafter adding cold rituximab, in excess of10%–20% of the RIC was still binding to bothtypes of cells. This retention might have been theresult of the internalization of at least a portionof the antibody. Furthermore, the dissociationrate constants determined in this paper werefound by adding a large amount of unlabeled an-
tibody to inhibit further binding. Thus, the dis-sociation might be even slower, as an eventualfraction of monovalently bound hot antibodycould be competed out by the cold antibody. Ad-ditionally, a bivalently bound antibody is proba-bly continuously wobbling, such that one armbecomes transiently detached. Therefore, the dis-sociation rate constant might depend on theamount of unlabeled antibody added. To preventthis artifact, the dissociation rate constant mightinstead be measured after a large dilution of thereaction mixture.
475
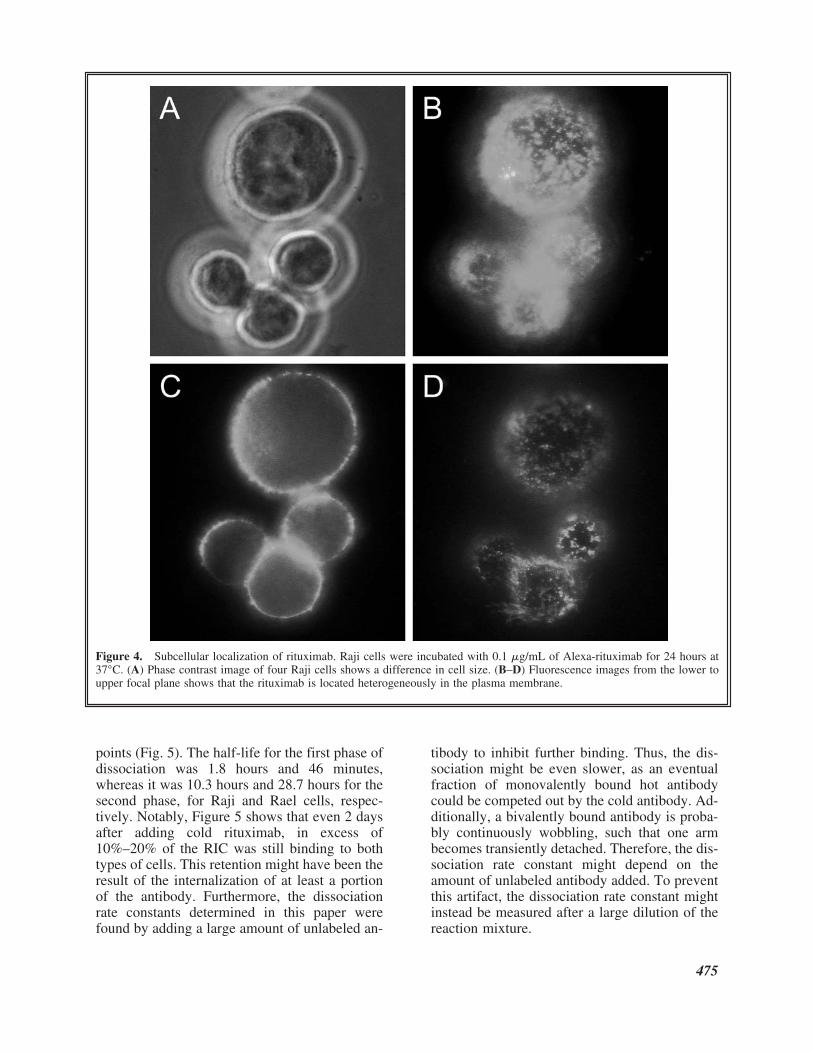
Figure 4. Subcellular localization of rituximab. Raji cells were incubated with 0.1 �g/mL of Alexa-rituximab for 24 hours at37°C. (A) Phase contrast image of four Raji cells shows a difference in cell size. (B–D) Fluorescence images from the lower toupper focal plane shows that the rituximab is located heterogeneously in the plasma membrane.
Equilibrium was assumed to be reached in theScatchard experiments after 2 hours. Therefore,the dissociation rate constant for the first phaseof dissociation (kd in Equation 2) was used to cal-culate Kd. The Kd found by the Scatchard analy-sis has been termed functional Kd, and it was 41times larger than Kd for Raji cells and 6 times forRael cells (Table 2).
The association rate constants observed wereslower than one might expect at the relativelyhigh concentrations used. The cells were incu-bated during continuous shaking, but the slow
diffusion of antibody might still have limited theassociation of antibody with antigen.
DISCUSSION
In this study, we measured the binding parame-ters of 227Th-DOTA-p-benzyl-rituximab, a newpotential RIC against CD20-positive lymphomas.We have found similar affinities using theScatchard analysis, as given in the prescribing in-formation for rituximab and reported by others,17
which shows that labeling with 227Th does notsignificantly damage the antibody. The measuredaffinity was confirmed by using Alexa-labeledrituximab and flow cytometry. Fluorescence mi-croscopy of rituximab bound to lymphoma cellsshowed no internalization of antibody during thefirst 24 hours of incubation, but indicated a het-erogeneous distribution of antigens on the cellsurface. Flow cytometry showed a wide range ofantigen distribution between the cells.
The mean number of available binding siteswas at least twice as high for Rael cells as forRaji and Daudi cells (Table 1), and the distribu-tion of available antigens resembled a log normaldistribution (Fig. 2). The cells in Figure 3 wereDNA stained, which made information about cellcycle available. However, the variation in thenumber of antigens was not related to position inthe cell cycle (not shown). For therapy, the cellswith the lowest number of available binding sitesare of particular interest, as they may result in tu-mor regrowth. For low specific activity, antigensaturation may occur, preventing a sufficientnumber of radioactive molecules to attach to thecell. Microbeam experiments have shown that to
476
Table 2. Dissociation Rate Constant (Kd), AssociationRate Constant (Ka), Kinetic Dissociation Constant (Kd),and Functional Equilibrium Dissociation Constant(Functional Kd) for Raji and Rael Cells Labeled with227Th-DOTA-p-Benzyl-Rituximab
kd ka Functional kd Kda
Cell line (h�1) (nM�1h�1) (nM (nM)
Raji 0.4 0.3 1.9 0.5 8.1 1.9 0.2 0.2Raelab 0.9 0.8 0.3 0.1 18.7 2.5 3.3 3.3
aKd calculated using kd and ka.bKd of Rael cells was significantly higher than the Kd of
Raji cells.
Figure 5. Dissociation rate constant. Decreasing numberof counts per minute (cpm) as a function of time after theaddition of 100 �g/mL of cold rituximab to (A) Raji or (B)Rael cells in equilibrium with three different concentrationsof 227Th-p-benzyl-DOTA-rituximab. The cells were incu-bated with 227Th-p-benzyl-DOTA-rituximab for 1.5 hoursto reach equilibrium. Then, the cells were washed and 100�g/mL cold rituximab was added. The cells were washedbefore each measurement. Mean unspecific binding to Rehcells for all time points were 25 cpm for 50 ng/mL, 24 cpmfor 100 ng/mL, and 36 cpm for 300 ng/mL. Unspecific bind-ing did not vary significantly with time. Error bars � stan-dard deviation.

obtain a 1% cell survival probability, 21 alphaparticles on average have to hit the nucleus,18
which fits well with microdosimetry calculationsof the alpha-RIT of lymphoma cells11 and withsimulations of tumor-control probability.19 Lym-phoma cells have a relatively large nucleus,which ensures that approximately one third of thealpha-particles emitted from the cell surface willhit the nucleus.20 Thus, at least 63 227Th-ritux-imab conjugates have to decay at the cell surfaceto get a cell kill probability of 99%. Hence, as-suming log normal distributions of available anti-gens, the number of antigens in the 0.14% (mean-3�) of cells with the lowest number of antigenswas calculated. For these cells, a specific activ-ity of 2375 Bq/�g for Raji cells, 1275 Bq/�g forRael cells, and 6000 Bq/�g for Daudi cells wasnecessary to obtain 63 227Th-rituximab conju-
gates on the cell surface (Table 3). We have pre-viously shown that the incubation of cells withonly 200 Bq/mL of 227Th-rituximab significantlyinhibited the growth of Raji cells.10 This corre-sponded to only 0.2 �g/mL of 227Th-rituximab(specific activity, 1000 Bq/�g), which meansthat, on average, approximately Bmax/7 antigenshad bound antibody (Fig. 3). For the 0.14% cellswith the lowest number of antigens, this meansapproximately four 227Th atoms per Raji cell. Inaddition, daughter cells get half of the 227Th-rit-uximab of the mother, so for the calculationsabove to be valid, we have to assume no tumorgrowth occurred during the decay of 227Th. How-ever, because 227Th-rituximab has a long biolog-ical half-life in vivo (5.3 days in mice10), tumorcells undergoing division may later be irradiatedowing to a continuous association of RIC fromthe pool of circulating compound.
For low concentrations of rituximab, a largeramount of the antigens on Raji cells versus Raelcells had bound rituximab (Fig. 2D) and viceversa at concentrations above 1 �g/mL (Fig. 3).This result may be explained by differences inthe association rate for the two cell lines. For lowconcentrations of antibody, only a small fractionof the available antigens is occupied by antibodyand it takes a longer time to reach equilibrium.Thus, if the association constant is higher on theRaji cells than on the Rael cells, the Raji cellswill bind antibody faster and thus appear to have
477
Figure 6. Association rate constant. Increasing number ofcounts per minute (cpm) as a function of time after the ad-dition of three different concentrations of 227Th-p-benzyl-DOTA-rituximab to (A) Raji or (B) Rael cells. The cellswere washed before measurement and unspecific binding toReh cells was subtracted before plotting. Error bars � stan-dard deviation.
Table 3. The 0.14% Limit of Number of Antigens andthe Specific Activity Necessary to Get 63227Th DecaysPer Cell
Raji Rael Daudi
Number of antigensa 47,150 87,950 18,780Specific activity (Bq/�g)b 2375 1275 6000
aAssuming a log normal distribution with a standard de-viation of � � FWHM/2(2ln2), the percentage of cells withthe lower number of available binding sites than
is 0.14%.
bSpecific activity � [#227Th] � �Tln
1
2
/2�
� �MN
W
A� ,
where T1/2 is the half-life of 227Th, Mw, is the molecularweight of rituximab, and NA is Avogadro’s number.
Bmax��e�3� � �
�
2
2��
Bmax��e�3� � �
�
2
2��

more antigen sites than the Rael cells. For higherconcentrations of antibody, it takes a shorter timeto reach equilibrium and then kinetics have lessimpact. We have found that the association ratewas seven times higher for Raji cells than for Raelcells (Table 2), so this explanation might be theanswer. Furthermore, it is tempting to speculatethat the reason for the lower ka and higher kd ofthe CD20 antigen on Rael cells than on Raji cellsmight be related to differences in organization ofthe antigens on the cell surface. Other explana-tions may be mutation in the CD20 gene or post-translational modifications of the CD20 proteinin either of the cell lines.
There was a large difference between the functional Kd and Kd determined by the kinetic experiments, which might indicate that the ra-dioimmunoconjugate bound irreversibly, andconsequently, had an almost infinite affinity orKd almost equal to zero. If kd2 (Equation 2) wasused, the values of Kd were approximately1 � 10�15 M for both Raji and Rael cells. Thus,the difference between functional Kd and Kdwere certainly larger than an order of magni-tude. The reason for this difference might bethe disagreement between the assumption aboutequilibrium that is the basis for the Scatchardanalysis and the two-phase dissociation of theantibody conjugate. These results imply that theScatchard analysis does not always give reli-able estimates of antibody affinity.
CONCLUSIONS
The findings of this study show that 227Th-DOTA-p-benzyl-rituximab has relevant antigen-binding properties for use in targeted �-im-munotherapy against CD20-positive lymphomasand that the association and dissociation proper-ties of the antigen-antibody complex is compati-ble with the relatively long half-life of 227Th.Thus, low-dose-rate �-RIT might be a promisingstrategy against CD20-positive tumors.
ACKNOWLEDGMENTS
We are grateful to Jørgen Borrebæk of AlgetaASA, Oslo, Norway, for the production of 227Th-DOTA-p-benzyl-rituximab. This study was fi-nanced by the MEDKAP program of The Nor-wegian Research Council.
REFERENCES
1. Jurcic JG, Larson SM, Sgouros G, et al. Targeted al-pha-particle immunotherapy for myeloid leukemia.Blood 2002;100:1233.
2. McDevitt MR, Ma D, Lai LT, et al. Tumor therapy withtargeted atomic nanogenerators. Science 2001;294:1537.
3. Zalutsky MR, Zhao XG, Alston KL et al. High-levelproduction of alpha-particle-emitting 211At and prepa-ration of 211At-labeled antibodies for clinical use. J NuclMed 2001;42:1508.
4. Hall EJ, Astor M, Bedford J, et al. Basic radiobiology.Am J Clin Oncol 1988;11:220.
5. Miederer M, McDevitt MR, Borchardt P, et al. Treat-ment of neuroblastoma meningeal carcinomatosis withintrathecal application of alpha-emitting atomic nano-generators targeting disialo-ganglioside GD2. Clin Can-cer Res 2004;10:6985.
6. Nilsson S, Larsen RH, Fossa SD, et al. First clinical ex-perience with alpha-emitting radium-223 in the treat-ment of skeletal metastases. Clin Cancer Res 2005;11:4451.
7. Bruland O, Nilsson S, Fisher DR, et al. High-linear en-ergy transfer irradiation targeted to skeletal metastasesby the a-emitter 223Ra: Adjuvant or Alternative to Con-ventional Modalities? Clin Cancer Res 2006;12(Suppl.20):S1.
8. Henriksen G, Hoff P, Larsen RH. Evaluation of poten-tial chelating agents for radium. Appl Radiat Isot2002;56:667.
9. Jonasdottir TJ, Fisher DR, Borrebaek J, et al. First invivo evaluation of liposome-encapsulated 223Ra as a po-tential alpha-particle-emitting cancer therapeutic agent.Anticancer Res 2006;26:2841.
10. Dahle J, Borrebaek J, Melhus KB, et al. Initial evalua-tion of 227Th-p-benzyl-DOTA-rituximab for low-doserate alpha-particle radioimmunotherapy. Nucl Med Biol2006;33:271.
11. Aurlien E, Kvinnsland Y, Larsen RH, et al. Radiationdoses to non-Hodgkin’s lymphoma cells and normalbone marrow exposed in vitro: Comparison of an alpha-emitting radioimmunoconjugate and external gamma-irradiation. Int J Radiat Biol 2002;78:133.
12. Press OW. Radioimmunotherapy for non-Hodgkin’slymphomas: A historical perspective. Semin Oncol2003;30:10.
13. Lindmo T, Bunn PA, Jr. Determination of the true im-munoreactive fraction of monoclonal antibodies afterradiolabeling. Meth Enzymol 1986;121:678.
14. Rosenthal HE. A graphic method for the determinationand presentation of binding parameters in a complexsystem. Anal Biochem 1967;20:525.
15. Limbird LE. Cell Surface Receptors: A Short Courseon Theory and Methods. Kluwer Academic, Norwell,MA, 1996.
16. Dahle J, Krogh C, Melhus KB, et al. Alternative methodfor determination of maximum number of bound anti-
478

bodies, affinity, and association constant for antibodybinding. Nucl Med Commun in press.
17. Prescribing information for rituximab, 2006. Onlinedocument at: www.rituxan.com/ra/prescribing-info.jspAccessed July 16, 2007.
18. Sawant SG, Randers-Pehrson G, Metting NF, et al.Adaptive response and the bystander effect induced byradiation in C3H 10T(1/2) cells in culture. Radiat Res2001;156:177.
19. Roeske JC, Stinchcomb TG. The average number of al-pha-particle hits to the cell nucleus required to eradi-cate a tumour cell population. Phys Med Biol 2006;51:N179.
20. Kvinnsland Y, Stokke T, Aurlien E. Radioimmuno-therapy with alpha-particle emitters: Microdosimetry ofcells with a heterogeneous antigen expression and withvarious diameters of cells and nuclei. Radiat Res 2001;155:288.
479
All in-text references underlined in blue are linked to publications on ResearchGate, letting you access and read them immediately.




















