
Ethyl propionate: Synergistic kairomone for african palm weevil, Rhynchophorus phoenicis L....
9
Jpurnal of Chemical Ecology, Vol. 20, No. 4, 1994 .ETHYL PROPIONATE: SYNERGISTIC KAIROMONE FOR AFRICAN PALM WEEVIL, Rhynchophorus phoenicis L. (COLEOPTERA: CURCULIONIDAE) GERHARD GRIES,I,* REGINE GRIES,I ALICE L. PEREZ,2 LILLIANA M. GONZALES,2 HAROLD D. PIERCE, JR.,2 A. CAMERON OEHLSCHLAGER,2 MARC RHAINDS,I MESMER ZEBEYOU, 3 and BROU KOUAME 3 I Centre for Pest Management, Department of Biological Sciences 2Department of Chemistry Simon Fraser University Burnaby, Britisñ Columbia, Canada V5A 1S6 3/nstituts des Foréts (/DEFOR) Departement des Plantes Oléagineuses (DPO) Station Principale de La Me 13 B.P. 989 Abidjan 13, Cote d'Ivoire (Received August 17, 1993; accepted December 6, 1993) Abstract-Small trunk pieces of a freshly felled lO-year-old oil palm, Elaeis quineensis (Jacq.), were placed in a modified Nalgene desiccator, and volatiles captured for six days on Porapak Q. Gas chromatographic (GC) analysis of Porapak-Q-trapped volatiles with both f1ame ionization (FID) and electroan- tennographic detection (EAD) using male or female R. phoenicis antennae revealed several EAD-active compounds. They were identified as: ethyl ace- tate, ethyl propionate, isobutyl propionate, ethyl butyrate, and ethyl isobu- tyrate. In field experiments in the La Me Research Station, Cote d'Ivoire, ethyl propionate (50 mg/24 hr) but not all esters combined (50 mg/24 hr each) significantly increased capture of R. phoenicis in pheromone-baited (3 mg/24 hr) traps. One kilogram of 1- to 3-day-old palm tissue was significantly more effective than ethyl propionate in enhancing pheromone attraction. Superior attraction of palm tissue may be attributed to additional as yet unknown semi- ochemicals. Altematively, release rates and/or ratios of synthetic volatiles differed from those of palm tissue at peak attraction. Key Words-Coleoptera, Rhynchophorus phoenicis, Elaeis quineensis, kai- romone, synergism, oil palm, palm weevil, palm volatiles, host selection, *To whom correspondence should be addressed. 889 0098-0331/94/0400-0889$07.00/0 © 1994 Plenum Publishing Corporation
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Ethyl propionate: Synergistic kairomone for african palm weevil, Rhynchophorus phoenicis L....

Jpurnal of Chemical Ecology, Vol. 20, No. 4, 1994
.ETHYL PROPIONATE: SYNERGISTIC KAIROMONE FORAFRICAN PALM WEEVIL, Rhynchophorus phoenicis L.
(COLEOPTERA: CURCULIONIDAE)
GERHARD GRIES,I,* REGINE GRIES,I ALICE L. PEREZ,2LILLIANA M. GONZALES,2 HAROLD D. PIERCE, JR.,2A. CAMERON OEHLSCHLAGER,2 MARC RHAINDS,I
MESMER ZEBEYOU, 3 and BROU KOUAME3
ICentre for Pest Management, Department of Biological Sciences2Department of ChemistrySimon Fraser University
Burnaby, Britisñ Columbia, Canada V5A 1S63/nstituts des Foréts (/DEFOR)
Departement des Plantes Oléagineuses (DPO)Station Principale de La Me
13 B.P. 989 Abidjan 13, Cote d'Ivoire
(Received August 17, 1993; accepted December 6, 1993)
Abstract-Small trunk pieces of a freshly felled lO-year-old oil palm, Elaeisquineensis (Jacq.), were placed in a modified Nalgene desiccator, and volatilescaptured for six days on Porapak Q. Gas chromatographic (GC) analysis ofPorapak-Q-trapped volatiles with both f1ame ionization (FID) and electroan-tennographic detection (EAD) using male or female R. phoenicis antennaerevealed several EAD-active compounds. They were identified as: ethyl ace-tate, ethyl propionate, isobutyl propionate, ethyl butyrate, and ethyl isobu-tyrate. In field experiments in the La Me Research Station, Cote d'Ivoire,ethyl propionate (50 mg/24 hr) but not all esters combined (50 mg/24 hr each)significantly increased capture of R. phoenicis in pheromone-baited (3 mg/24hr) traps. One kilogram of 1- to 3-day-old palm tissue was significantly moreeffective than ethyl propionate in enhancing pheromone attraction. Superiorattraction of palm tissue may be attributed to additional as yet unknown semi-ochemicals. Altematively, release rates and/or ratios of synthetic volatilesdiffered from those of palm tissue at peak attraction.
Key Words-Coleoptera, Rhynchophorus phoenicis, Elaeis quineensis, kai-romone, synergism, oil palm, palm weevil, palm volatiles, host selection,
*To whom correspondence should be addressed.
889
0098-0331/94/0400-0889$07.00/0 © 1994 Plenum Publishing Corporation

890 GRIES ET AL.
primary attraction, aggregation pheromone, 3-methyl-octan-4-ol, ethyl ace-tate, ethyl propionate, isobutyl propionate, ethyl butyrate, ethyl isobutyrate.
INTRODUCTION
Rynchophorus palm weevils are destructive pests of commercial and ornamentalpalms in the tropics (Hill, 1983). Weevils are attracted to wounded or stressedpalms or fermenting palm sap (Kalshoven, 1950; Lever, 1969; Leong, 1987;Giblin-Davis and Howard, 1989; Sadakathulla, 1991). After oviposition on softpalm tissue, larvae tunnel into the terminal bud or trunk of the tree, leading toits death. In recent laboratory and field studies, methyl-branched, secondaryaleohols have been identified as aggregation pheromones in the American palmweevil, R. palmarum L. (Rochat et al., 1991; Oeh1schlager et al., 1992a); theAfrican palm weevil, R. phoenicis L. (Gries et al., 1993; Rochat et al., 1993);the Asian palm weevils, R. vulneratus (Panz.), R. ferrugineus (Oliv.) (Hallettet al., 1993a,b), R. bilineatus (Montr.) (Oehlschlager et al., 1993b); and thesabal palmetto weevil, R. cruentatus (F.) (Weissling et al., 1994). As in themaize weevil, Sitophilus zeamais Motschulsky (Walgenbach et al., 1987),attraction of Rhynchophorus weevils to aggregation pheromones invariablyrequired the presence of synergistic plant tissue (Oehlschlager et al., 1992a, b,1993a,b,c; Gries et al., 1993; Hallett et al., 1993a,b; Rochat et al., 1993;Weissling et al., 1994). We report the identification and field testing of oil palmconstituent ethyl propionate, which enhances attraction of the R. phoenicisaggregation pheromone "phoenicol" (3-methyl-octan-4-01).
METHODS AND MA TERIALS
Volatile Collection. In the La Me Research Station, Cote d'lvoire, lowertrunk sections of a freshly felled lO-year-old oil palm were cut into small cubesand placed in a modified Nalgene desiccator. An aspirator-driven, charcoal-filtered airstream was maintained through the desiccator for five days, collectingpalm volatiles on Porapak Q. Volatiles were eluted from Porapak Q with pentaneand concentrated by distillation.
Instrumental Methods. Porapak Q extracts were analyzed by gas chromato-graphic-electroantennographic detection (GC-EAD) (Arn et al., 1975), employ-ing a Hewlett Packard (HP) 5885B gas chromatograph equipped with aSP-lOOO-coated, fused silica column (30 m X 0.25 mm ID) (Supeleo, Inc.Bellefonte, Pennsylvania). A HP 5985B coupled GC-mass spectrometer (GC-MS) fitted with the same column was used for GC-MS analyses of plant volatilesin both electro n impact (El) and chemical ionization (CI) modes.
Field Bioassay. Weevil response to synthetic palm volatiles was tested in

PALM WEEVIL KAIROMONE 891
6- to lO-year-old oil palm stand s of the La Me Research Station. Fifteen-literbucket traps (Oehlschlager et al., 1992a) were attached at breast height to palmtrees in complete randomized blocks with traps at 27-m intervals and blocks27-100 m apart. Release devices suspended 5 cm below the trap lid dispensedtest chemicals. Synthetic, stereoisomeric phoenicol (98 % purity) (Gries et al.,
, 1993), was released at 3 mg/day (at 25°C), whereas each of the palm volatiles, was released at 50 mg/day (at 25°C). A S-cm-wide ring of petrolatum (Ana-
chemia, Rouses Pount, New York 12979) on the inner upper surface of eachtrap, and a wet yellow sponge on the trap bottom, treated with insecticidal(biodegradable) Evisect -" S" (0.3 % thiocyclam-hydrogenoxalate in water),retained captured weevils.
The first prescreening seven-treatment experiment tested phoenicol aloneand in combination with either one or all of the following compounds: ethylacetate (EA), ethyl propionate (EP), isobutyl propionate (IBP) , ethyl butyrate(EB), or ethyl isobutyrate (EIB). The second four-treatrnent experiment testedphoenicol alone and in combination with either EA, EP, or both. A final three-treatment experiment tested phoenicol alone and combined with either EP or 1kg of freshly cut oil palm tissue. All compounds were purchased (Aldrich Chem.Company, Inc., Milwaukee, Wisconsin 53233) and were >98% chemicallypureo
RESULTS
Four palm volatiles elicited good antennal responses in GC-EAD analyses(Figure 1). El and CI mas s spectra ofthese compounds indicated that they were,respectively, EA, EP, EIB, and EB (Figure 1). Palm volatiles and authenticstandards had comparable antennal activity and identical retention and mas sspectrometric characteristics. Additional esters, such as isobutyl propionate, wereidentified in palm volatiles but were only weakly EAD active. In the prescreen-ing field experiment, EA or EP but not all synthetic esters combined enhancedattraction to phoenicol. Ethyl propionate alone more than in combination withEA enhanced pheromone attraction (Experiment 2, Figure 2), but palm tissuewas two to three times more synergistic than EP (Experiment 3, Figure 3).
DISCUSSION
Selection of specific plants for feeding and reproduction has been welldocumented in the Coleoptera, particularly for scolytid and curculionid beetles(Wattanapongsiri, 1966; Moeck et al., 1981 and references therein; Ryker andOester, 1982; Gara et al., 1984; Miller et al., 1986; Moeck and Simmons,1991; Byers, 1992 and references therein; Tunset et al., 1993). Short-chain
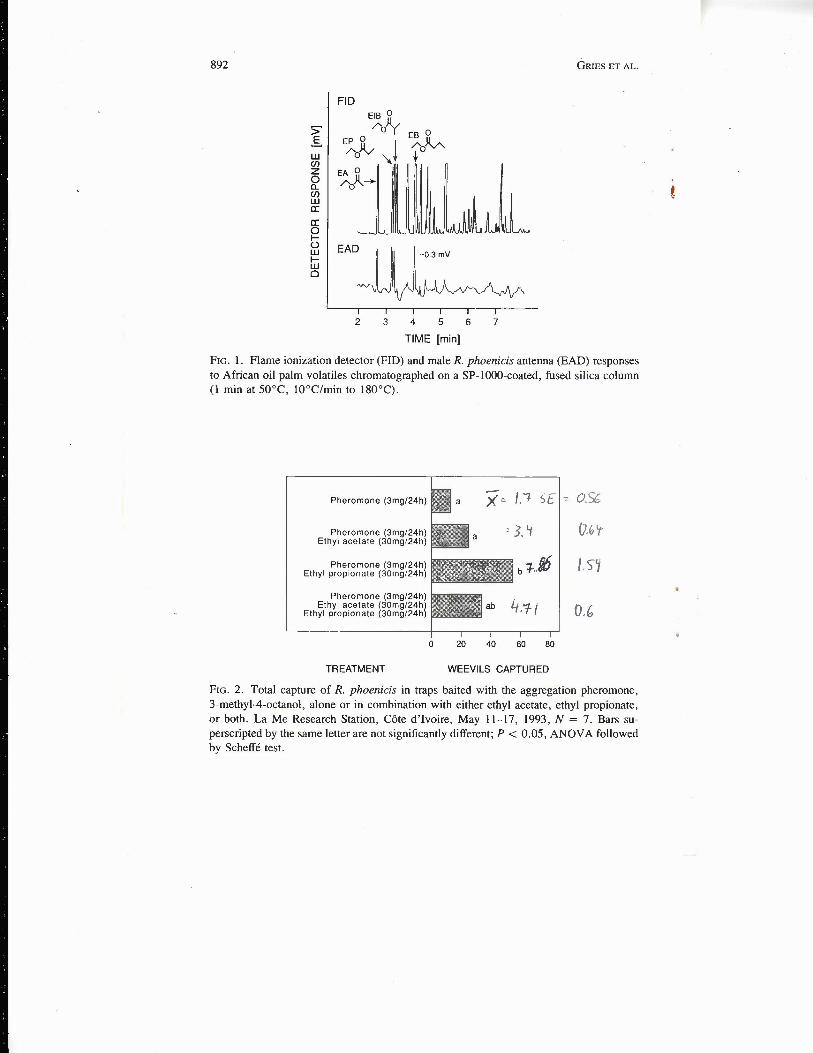
892 GRIES ET AL.
5'.sw(f)
zOa..(f)wa:a:~OW1-WO
FID
EAD -0.3 mV
2 3 4 5 6
TIME [rnin]7
FIG. 1. Flame ionization detector (FID) and male R. phoenicis antenna (EAD) responsesto African oil palm volatiles chromatographed on a SP-1OOO-coated, fused silica column(1 min at 50°C, lQ°C/min to 180°C).
Pheromone (3mg/24h) - 1.1- SE 0.56X> -e
'3. ~ (J.6'f'
b 1-..15 ISj
4.1f O.b
o 20 40 60 80
WEEVILS CAPTURED
Pheromone (3mg/24h)Ethyl acetate (30mg!24h)
Pheromone (3mg/24h)Ethyl propionate (30mg/24h)
Pheromone (3mg/24h)Ethyl acetate (30mg/24h)
Ethyl propionate (30mg/24h)
TREATMENT
FIG. 2. Total capture of R. phoenicis in traps baited with the aggregation pheromone,3-methyl-4-octanol, alone or in combination with either ethyl acetate, ethyl propionate,or both. La Me Research Station, Cote d'Ivoire, May 11-17, 1993, N = 7. Bars su-perscripted by the same letter are not significantly different; P < 0.05, ANOVA followedby Scheffé test.

PALM WEEVIL KAIROMONE
.,
Z ·1
0,3Pheromone (3mg/24h)
e
Pheromone (3mg/24h)Ethyl propionate (30mg/24h)
Pheromone (3mg/24h)Palm tissue (1 kg)
TREATMENTo 20 40 60 80
WEEVILS CAPTURED
FIG. 3. Total capture of R. phoenicis in traps baited with pheromone alone or in com-bination with either ethyl propionate or I kg of 1- to 3-day-old palm tissue. La MeResearch Station, Cote d'Ivoire, May 14-17, 1993, N = 10. Bars superscripted by thesame letter are not significantly different; P < 0.05, ANOVA followed by Scheffé test.
aleohols, oleoresin, monoterpenes, and sesquiterpenes alone or in combinationhave been associated with host selection behavior (Byers, 1992 and referencestherein).
Plant-derived volatiles and beetle-produced pheromones have ofien beenshown to mediate synergistically host selection behavior. Although aggregationpheromones of many beetles are known (Borden, 1985; Byers, 1989), few syn-ergistic host plant kairomones have been identified. Ethanol, myrcene, e-pinene,¡1-phellandrene, and camphene enhance response to aggregation pheromone invarious bark and ambrosia beetles (Bedard et al., 1969; Moeck, 1970; Vité andBakke, 1979; Kohnle, 1985; Paiva and Kiesel, 1985; Byers et al., 1988; Millerand Borden, 1990). o-Cubebene, typically released from moribund elm trees,enhances pheromone attraction in the elm bark beetle, Scolytus multistriatus(Marsham) Peacock et al., 1984). Propanoic and butanoic acids, methanol,2-propanol, l-heptanol, methyl butanoate, and propanal were most effective insynergizing pheromone attraction of the dried fruit beetle, Carpophilus hemi-pterus (L.) (Dowd and Bartelt, 1991). In mimicking whole-wheat bread doughodor, a blend of synthetic acetaldehyde, ethyl acetate, ethanol, l-propanol,2-methylpropanol, 2-methylbutanol, and 3-methylbutanol attracted the nitidulidsCarpophilus lugubris Murray, Glischrochilus quadrisignatus (Say), and G. fas-ciatus (Olivier) (Lin and Phelan, 1991a,b) and synergistically enhanced attrac-tion of C. lugubris pheromone (Lin et al., 1992). We report EP as a synergistichost plant kairomone for R. phoenicis (Figure 2 and 3).
EP significantly increased capture of R. phoenicis in phoenicol-baited traps(Figure 2) but was stillless effective thanfreshly cut palrn tissue (Figure 3). R.phoenicis preference for palm- over EP-baited traps (Figure 3) was unlikely dueto different microclimates within traps. The weevil-preferred humid environment
893 S' t
o 1),

894 GRIES ET AL.
(Weissling and Giblin-Davis, 1993; Oehlschlager et al., 1993c), as provided bypalm tissue, was prevailing in al! traps due to .wet sponges on trap bottoms.Superior synergism of palm tissue may therefore be attributed to additional, asyet unknown palm volatiles.
Increase, culmination, and decrease of palm tissue attraction within threeto five days after cutting (Weissling et al., 1992; Gries et al., 1993; Hallett etal., 1993a; Oehlschlager et al., 1993a) indicate proportional changes of semi- ~ochemicals that may be critical for optimal attraction of weevils. Potato leafvolatiles, e.g., cis-3-hexen-l-01, cis-3-hexenyl acetate, trans-2-hexenal, andtrans-2-hexen-l-01 only induce positive anemotaxis in the Colorado beetle, Lep-tinotarsa decemlineata (Say), when released in natural ratio s (Vis ser, 1979).Antennally active palm volatiles should therefore be field tested at ratios andrelease rates equivalent to palm tissue at peak attraction. Abundance and ratioof palm volatiles in volatile extracts may not accurately reflect natural releaserates of palm tissue and may in part depend on the volatile collection technique.Aside from Porapak Q, other absorbents, such as charcoal, Tenax, glasswool,or cryogenic traps (Golub and Weatherston, 1984) need to be evaluated, partic-ularly for trapping low-boiling-palm volatiles.
Ethyl propionate, EA, EB, and EIB are comrnon volatile constituents in(fermenting) African oil palm, coconut palm, Cocos nucifera L., and cabbagepalmetto, SabaLpaLmetto (Walter) (Gries et al., unpublished). As all these esterselicit antennal responses in R. phoenicis and congeneric R. paLmarum, R. cruen-tatus, R. ferrugineus, R. vuLneratus, and R. biLineatus (Gries et al., unpub-lished), EP or any other "palm ester" alone or in combination with other palmvolatiles may be a primary attractant for Rynchophorus palm weevils. In a recentempirical study, EA has been shown to be a semiochemical for R. cruentatus(Giblin-Davis et al., 1993) and for R. paLmarum (Jaffé et al., 1993). Syntheticpalm volatiles in pherornone-baited traps could replace currently used insecti-cide-treated sugar cane or palm tissue, thereby promoting the further develop-ment of comrnercial, semiochemical-based management of Rynchophorusweevils in oil, coconut, and date palm (Oehlschlager et al., 1992b; Chinchillaet al., 1993).
Acknowledgments- We thank G. Owen for mas s spectrometry. The research was supportedby an NSERC research grant to A.C.O.
REFERENCES
ARN, H., STADLER,E., and RAUSCHER,S. 1975. The e1ectroantennographic detector-a selectiveand sensitive tool in the gas chromatographic analysis of insect pheromones. Z. Naturforsch.30c:722-725.
BEDARD,W.D., TILDEN, P.E., WOOD, D.L., SILVERSTEIN,R.M., BROWNLEE,R.G., and RODlN,

PALM WEEVIL KAIROMONE 895
J.O. 1969. Westem pine beetle: Field response to its sex pheromone and a synergistic hostterpene, myrcene. Science 164:1284-1285.
BORDEN,J.H. 1985. Aggregation pheromones, pp. 257-285, in G.A. Kerkut and L.!. Gilbert(eds.).Comprehensive Insect Physiology, Biochemistry and Pharmacology. Pergamon Press, Oxford.
BYERS, J.A. 1989. Chemical ecology ofbark beetles. Experientia 45:271-283.BYERS, J.A. 1992. Attraction of bark beetles Tomicus piniperda, Hylurgops palliatus and Trypo-
dendron domesticum and other insects to short-chain alcohols and monoterpenes. J. Chem.Ecol. 18:2385-2402.
BYERS, J.A., BIRGERSSON,G., LÓFQVIST,J., and BERGSTRÓM,G. 1988. Synergistic pheromonesand monoterpenes enable aggregation and host recognition by a bark beetle. Naturwissenschaf-ten 75:153-155.
CHINCHILLA,C.M., OEHLSCHLAGER,A.C., and GONZALES,L.M. 1993. Management of red ringdisease in oil palm through pheromone-based trapping of Rhynchophorus palmarum (L.). Pro-ceedings of the Intemational Conference on Oil Palm (PORIM), Kuala Lumpur, Malaysia.
DOWD, P.F., and BARTELT,R.J. 1991. Host-derived volatiles as attractants and pheromone syner-gists for driedfruit beetle, Carpophilus hemipterus. J. Chem. Ecol. 17:285-308.
GARA, R.I., GEISZLER,D.R., and LITTKE, W.R. 1984. Primary attraction of the mountain pinebeetle to lodgepole pine in Oregon. Ann. Entorno!. Soco Am. 77:333-334.
GIBLIN-DAVIS,R., and HOWARD, F.W. 1989. Vulnerability of stressed palms to attack by Rhyn-chophorus cruentatus (Coleoptera: Curculionidae) and insecticial control of the pest. J. Econ.Entomol.82:1185-1190.
GIBLIN-DAVIS,R.M., WEISSLING,T.J., OEHLSCHLAGER,A.C., and GONZALES,L.M. 1993. Fieldresponse of Rhynchophorus cruentatus (F.) (Coleoptera: Curculionidae) to its aggregation pher-omone and fermenting plant volatiles. Fl. Entomol. In press.
GOLUB, M.A., and WEATHERSTON,I. 1984. Techniques for extracting and collecting sex phero-mones from live insects and from artificial sources, pp. 223-285, in H.E. Hummel and T.A.Miller (eds.). Techniques in Pheromone Research. Springer, New York.
GRIES,G., GRIES,R., PEREZ, A.L., OEHLSCHLAGER,A.C., GONZALES,L.M., PIERCE,H.D., KOUDA,M., ZEBEYOU,M., and NANOU, N. 1993. Aggregation pheromone of the African palm weevil,Rhynchophorus phoenicis F. Naturwissenschaften 80:90-91.
HALLETT, R,H., GRIES, G., GRIES, R., BORDEN, J.H., CZYZEWSKA,E., OEHLSCHLAGER,A.C.,PIERCE, H.D., JR., ANGERILLI,N.P.D., and RAUF, A. 1993a. Aggregation pheromone oftwoAsian palm weevils, Rhynchophorus ferrugineus (Oliv.) and R. vulneratus (Panz.). Naturwis-senschaften 80: 328-331.
HALLETT,R.H., OEHLSCHLAGER,A.C., GRIES, G., ANGERILLI,N.D.P., AL SHAREQUI,R.H., GAS-SOUMA,M.S., and BORDEN,J.H. 1993b. Field activity of aggregation pheromone of two Asianpalm weevils. Proceedings of the Intemational Conference on Oil Palm (PORIM), KualaLumpur, Malaysia.
HILL, D.S. 1983. Agricultural Insect Pests of the Tropics and Their Control. Cambridge UniversityPress, New York.
JAFFÉ, K., SÁNCHEZ,P., CERDA, H., HERNÁNDEZ,J.V., JAFFÉ, R., URDANETA,N., GUERRA,G.,MARTÍÑEZ, R., and MIRAS, B. 1993. Chemical ecology of the palm weevil Rhynchophoruspalmarum (L.) (Coleoptera: Curculionidae): Attraction to host plants and to a male-producedaggregation pheromone. J. Chem. Ecol. 19:1703-1720.
KALSHOVEN,L.G.E. 1950. Pest of Crops in Indonesia. P.T. Ichtiar Baru-Van Hoeve, Jakarta.Revised and translated by P.A. Van der Laan (1981). 701 pp.
KOHNLE,U. 1985. Untersuchungen über die Pheromonsysteme sekundárer Borkenkafer (Col., Scol-ytidae). z. Angew. Entomol. 100: 197-218.
LEONG, C.T. 1987. Sago worm culture. Annual Report of the Research Branch, Department ofAgriculture for the year 1986. Ministry of Agriculture and Cornrnunity Development, Sarawak.

896 GRIES ET AL.
LEVER, R.J.A.W. 1969. Pests of the Coconut Palm. FAO Agricultura! Studies No. 77, Food andAgriculture Organization of the United Nations, Rome. 190 pp.
LIN, H., and PHELAN,P.L. 1991a. Identification offood volatiles attractive to the dusky sap beetle,Carpophilus lugubris (Coleoptera: Nitidulidae). J. Chem. Ecol. 17:1273-1286.
LIN, H., and PHELAN,P.L. 1991b. Characterization of host odor attractive to Glischrochilus quad-risignatus and Glischrochilus fasciatus. J. Chem. Ecol. 17:2469-2480.
UN, H., PHELAN, P.L., and BARTELT,RJ. 1992. Synergism between synthetic food odors and theaggregation pheromone for attracting Carpophilus lugubris in the field (Coleoptera: Nitiduli-dae). Environ. Entomol. 21:156-159.
MILLER, D.R., and BORDEN,J.H. 1990. f3-Phellandrene: kairomone for pine engraver, lps pini Say(Coleoptera: Scolytidae) J. Chem. Ecol. 16:2519-2532.
MILLER, D.R., MADDEN,J.L., and BORDEN,J.H. 1986. Primary attraction of Ips latidens (LeConte)and Hylastis gracilis LeConte (Coleoptera: Scolytidae) to high-girdled lodgepole pine, Pinuscontorta var. latifolia Engelmann. Can. Entomol. 118:85-85.
MOECK, H.A. 1970. Ethanol as the primary attractant for the ambrosia beetle, Trypodendron linea-tum (Coleoptera: Scolytidae). Can. Entomo!. 113:939-942.
MOECK, H.A., and SIMMONS,C.S. 1991. Primary attraction of mountain pine beetle, Dendroctonusponderosae Hopk. (Coleoptera: Scolytidae), to bolts of lodgepole pine. Can. Entomo!. 123:299-304.
MOECK, H.A., WOOD, D.L., and LINDAHL,K.Q., JR. 1981. Host selection behavior ofbark beetles(Coleoptera: Scolytidae) attacking Pinus ponderosa with special emphasis on the western pinebeetle, Dendroctonus brevicomis. J. Chem. Ecol. 7:49-83.
OEHLSCHLAGER,A.C., PIERCE, H.D., JR., MORGAN,B., WIMALARATNE,P.D.C., SLESSOR,K.N.,KING, G.G.S., ORlES, 0., GRIES, R., BORDE.N,J.H., JIRON, L.F., CHINCHILLA,C.M., andMEXZAN,R.G. 1992a. Chirality and field activity of rhynchophorol, the aggregation pheromoneof the American palm weevil. Naturwissenschaften 79: 134-135.
OEHLSCHLAGER,A.C., CHINCHILLA,C.M., and GONZALES,L.M. 1992b. Management of red ringdisease of oil palm through a pheromone-based trapping of Rhynchophorus palmarum. Pro-ceedings of the intemational seminar on coconut research, Kingston, Jamaica.
OEHLSCHLAGER,A.C., CHINCHILLA,C.M., GONZALES,L.M., JIRON, L.F., MEXZAN, R.O., andMORGAN, B. 1993a. Development of a pheromone-based trapping system for Rhynchophoruspalmarum (Coleoptera: Curculionidae). J. Econ. Entomol. 86: 1381-1392.
OEHLSCHLAGER,A.C., PEREZ, A.L., PIERCE, H.D., JR., OONZALES,L.M., PRIOR, B., GRIES, R.,and GRIES, O. 1993b. Aggregation pheromone of the Asian palm weevil, Rhynchophorusbilineatus. Naturwissenschaften Submitted.
OEHLSCHLAGER,A.C., CHINCHILLA,C.M., and GONZALES,L.M. 1993c. Optimization of a pher-omone-based trap for the American palm weevil. Proceedings of the international conferenceon oil pa1m (PORIM), Kuala Lumpur, Malaysia.
PAIVA, M.R., and KIESEL, K. 1985. Field response of Trypodendron spp. (Coleoptera: Scolytidae)to different concentrations of 1ineatin and o-pínene. Z. Angew. Entomo!. 99:442-447.
PEACOCK,J.W., WRIGHT, S.L., and FORO, R.D. 1984. Elm volatiles increase attraction of Scolytusmultistriatus to multilure. Environ. Entomol. 13:394-398.
RocHAT, D., MALOSSE,C., LETTERE,M., DUCROT,P.H., ZAGATTI,M., RENou, M., and DESCOINS,C. 1991. Male-produced aggregation pheromone ofthe American palm weevil, Rhynchophoruspalmarum (L.) (Coleoptera: Curculionidae): Collection, identification, electrophysiologicalactivity and laboratory bioassay. J. Chem. Ecol. 17:2127-2140.
RocHAT, D., DESCOIN,C., MALOSSE,C., NAGNAN,P., ZAGATTI,P., AKAMou, F., and MARIAU,D. 1993. Ecologie chimique des charancons des palmiers, Rhynchophorus spp. (Coleoptera).Oleagineux 48:225-236.
e

PALM WEEVIL KAIROMONE 897
:
RYKER, L.C., and OESTER, P.T. 1982. Pseudohylesinus nebulosus (LeConte) (Col., Scolytidae):Aggregation by primary attraction. Z. Angew. Entomol. 94:377-382.
SADAKATHULLA,S. 1991. Management of red palm weevil, Rhynchophorus ferrugineus F., incoconut plantations. Planter 67:415-419.
TUNSET, K., NILSSON, A.C., and ANDERSON,J. 1993. Primary attraction in host recognition ofconiferous bark beetles and bark weevils (Col., Scolytidae and Curculionidae). 1. Appl. Ento-mol. 115:155-169.
VISSER, J.H. 1979. Olfaction of the Colorado potato beetle at the onset of host selection. PhDthesis. Wageningen, The Netherlands.
VITÉ, J.P., and BAKKE,A. 1979. Synergism between chemical and physical stimuli in host colo-nization by an ambrosia beetle. Naturwissenschaften 66:528-529.
WALGENBACH,C.A., BURKHOLDER,W.E., CURTIS,M.J., and KHAN, Z.A. 1987. Laboratory trap-ping studies with Sitophilus zeamais (Coleoptera: Curculionidae). J. Econ. Entomol. 80:763-767.
WATTANAPONGSIRI,A. 1966. A revision of the genera Rhynchophorus and Dynamis (Coleoptera:Curculionidae). Dep. Agr. Sci. Bull. Bankok 1: 1-328.
WEISSLING,T.J., and GIBLIN-DAVIS,R.M. 1993. Water loss dynamics and humidity preference ofRhynchophorus cruentatus (Coleoptera; Curculionidae) adults. Enviran. Entomol. 22:93-98.
WEISSLING,T.J., GIBLIN-DAVIS,R.M., SCHEFFRAHN,R.H., and MENDOZA,N.M. 1992. Trap forcapturing and retaining Rhynchophorus cruentatus (Coleoptera: Curculionidae) adults usingSabal palmetto as bait. Fla. Entomol. 75:212-221.
WEISSLING,T.J., GIBLIN-DAVIS,R.M., GRIES, G., GRIES, R., PEREZ, A.L., PIERCE, H.D., JR.,and OEHLSCHLAGER,A.e. 1994. Aggregation pheromone of the palmetto weevil, Rhyncho-phorus cruentatus (F.) (Coleoptera: Curculionidae) J. Chem. Ecol. 20:505-515.




















