
Response of digestive cysteine proteinases from the colorado potato beetle (Leptinotarsa...
14
Archives of Insect Biochemistry and Physiology 31 :451-464 (1 996) Response of Digestive Cysteine Proteinases From the Colorado Potato Beetle (Leptinotarsa decemlineata) and the Black Vine Weevil (Ofiorynchus sukafus) to a Recombinant Form of Human Stefin A Dominique Michaud, Binh Nguyen-Quoc,Thierry C. Vrain, Dunne Fong, and Serge Yelle Centre de Recherche en Horticulture, Dkpartement de phytologie, Facultt? des Sciences de I'Agricultu~e et de I'Alimentation, Uniziersitt? Laval, Qukbec (D.M., B.N.-Q., S.Y.) and Pacific Agriculture Research Centre, Agriculture and Agri-Food Canada, Vancouver, B.C. (T.C.V.), Canada; Department of Biological Sciences, Rutgers, The State University of New Jersey, Piscataway, New Jersey (1J.F.) The effects of the cystatins, human stefin A (HSA) and oryzacystatin I (OCI) on digestive cysteine proteinases of the Colorado potato beetle (CPB), Leptinotarsa decemlineata, and the black vine weevil (BVW), Otiorynchus sulcatus, were assessed using complementary inhibition assays, cystatin-affinity chromatogra- phy, and recombinant forms of the two inhibitors. For both insects, either HSA and OCI used in excess (10 or 20 LM) caused partial and stable inhibition of total proteolytic (azocaseinase) activity, but unlike for OCI the HSA-mediated inhibitions were significantly increased when the inhibitor was used in large excess (1 00 pM). As demonstrated by complementary inhibition assays, this two-step inhibition of the insect proteases by HSA was due to the differential inactivation of two distinct cysteine proteinase populations in either insect ex- tracts, the rapidly (strongly) inhibited population corresponding to the OCI- sensitive fraction. After removing the cystatin-sensitive proteinases from CPB and BVW midgut extracts using OCI- (or HSA-) affinity chromatography, the effects of the insect "non-target" proteases on the structural integrity of the two cystatins were assessed. While OCI remained essentially stable, HSA was sub- jected to hydrolysis without the accumulation of detectable stable intermedi- ates, suggesting the presence of multiple exposed cleavage sites sensitive to the action of the insect proteases on this cystatin. This apparent susceptibility of HSA to proteolytic cleavage may partially explain its low efficiency to inactivate Acknowledgments: This work was supported by grants from the Natural Science and Engineer- ing Research Council (NSERC) of Canada and from the Conseil des Recherches en Peche et Agro-Alimentaire du Quebec to S.Y. D.M. was the recipient of an NSERC fellowship. Received May 8,1995; accepted November 27, 1995. Address reprint requests to Dominique Michaud, Centre de Recherche en Horticulture, Pavillon de I'Envirotron, Universite Laval, Quebec, Canada G1 K 7P4. 0 1996 Wiley-Liss, Inc.
Transcript of Response of digestive cysteine proteinases from the colorado potato beetle (Leptinotarsa...

Archives of Insect Biochemistry and Physiology 31 :451-464 (1 996)
Response of Digestive Cysteine Proteinases From the Colorado Potato Beetle (Leptinotarsa decemlineata) and the Black Vine Weevil (Ofiorynchus sukafus) to a Recombinant Form of Human Stefin A Dominique Michaud, Binh Nguyen-Quoc, Thierry C. Vrain, Dunne Fong, and Serge Yelle Centre de Recherche en Horticulture, Dkpartement de phytologie, Facultt? des Sciences de I'Agricultu~e et de I'Alimentation, Uniziersitt? Laval, Qukbec (D.M., B.N.-Q., S.Y.) and Pacific Agriculture Research Centre, Agriculture and Agri-Food Canada, Vancouver, B.C. (T.C.V.), Canada; Department of Biological Sciences, Rutgers, The State University of New Jersey, Piscataway, New Jersey (1J.F.)
The effects of the cystatins, human stefin A (HSA) and oryzacystatin I (OCI) on digestive cysteine proteinases of the Colorado potato beetle (CPB), Leptinotarsa decemlineata, and the black vine weevil (BVW), Otiorynchus sulcatus, were assessed using complementary inhibition assays, cystatin-affinity chromatogra- phy, and recombinant forms of the two inhibitors. For both insects, either HSA and OCI used in excess (10 or 20 LM) caused partial and stable inhibition of total proteolytic (azocaseinase) activity, but unlike for OCI the HSA-mediated inhibitions were significantly increased when the inhibitor was used in large excess (1 00 pM). As demonstrated by complementary inhibition assays, this two-step inhibition of the insect proteases by HSA was due to the differential inactivation of two distinct cysteine proteinase populations in either insect ex- tracts, the rapidly (strongly) inhibited population corresponding to the OCI- sensitive fraction. After removing the cystatin-sensitive proteinases from CPB and BVW midgut extracts using OCI- (or HSA-) affinity chromatography, the effects of the insect "non-target" proteases on the structural integrity of the two cystatins were assessed. While OCI remained essentially stable, HSA was sub- jected to hydrolysis without the accumulation of detectable stable intermedi- ates, suggesting the presence of multiple exposed cleavage sites sensitive to the action of the insect proteases on this cystatin. This apparent susceptibility of HSA to proteolytic cleavage may partially explain its low efficiency to inactivate
Acknowledgments: This work was supported by grants from the Natural Science and Engineer- ing Research Council (NSERC) of Canada and from the Conseil des Recherches en Peche et Agro-Alimentaire du Quebec to S.Y. D.M. was the recipient of an NSERC fellowship.
Received May 8,1995; accepted November 27, 1995.
Address reprint requests to Dominique Michaud, Centre de Recherche en Horticulture, Pavillon de I'Envirotron, Universite Laval, Quebec, Canada G1 K 7P4.
0 1996 Wiley-Liss, Inc.

452 Michaud et al.
the insect OCI-insensitive cysteine proteinases when not used in large excess. It could also have major implications when planning the use of cystatin-expressing transgenic plants for the control of coleopteran pests. o 1996 WiIey-Liss, Inc.
Key words: Coleoptera, digestive cysteine proteinases, human stefin A, oryzacystatin I
INTRODUCTION
Transformation of plant genomes with proteinaceous cysteine proteinase inhibitor (cystatin) cDNA clones of plant origin represents an attractive way to control several insect pests belonging to the order of Coleoptera. Recent studies have demonstrated the in vitro inhibition of midgut proteinases from several species of this group by either plant or animal cystatins (Liang et al., 1991; Hines et al., 1991; Gillikin et al., 1992; Chen et al., 1992; Michaud et al., 1993, 1995a; Orr et al., 1994) and deleterious effects of plant cystatins against the red flour beetle (Triboliurn castaneum) (Chen et al., 1992), the western corn rootworm (Diabvofica virgifern) (Orr et al., 19941, and the Colorado potato beetle (Leptinotarsa decemlineata) (Michaud et al., 1995b) have recently been reported. Based on these observations, cystatins were proposed as effective molecules to control insects that use cysteine proteinases for protein digestion, and oryzacystatin I (OC1")-expressing transgenic canola (Bonadk-Bottino, 1993), tobacco (Masoud et al., 1993), rice (Hosoyama et al., 1994), potato (Benchekroun et al., 1995), and poplar (Lepli. et al., 1995) plants were recently developed.
Despite these promising developments, the real potential of cystatins for the control of Coleoptera has not been clearly demonstrated. The significant effect of OCI on red flour beetle growth, notably, required incorporation of a very high level of inhibitor (10% w/w) into the diet (Chen et al., 19921, and few studies have yet been published on the repressive effect of cystatin-ex- pressing transgenic plants against insect growth and development (Leplk et al., 1995). As recently proposed, this apparent inefficiency of cystatins may be due to their instability in insect midguts (Orr et al., 1994; Michaud et al., 1995a). While in several cases coleopteran pests use cathepsin B- and cathep- sin H-like proteinases for protein digestion (Hilder et al., 1993), plant cystatins show no or little activity against both human and coleopteran cathepsins B (Kondo et al., 1990; Michaud et al., 1993). Cathepsin H-like proteinase from the major potato pest, Colorado potato beetle (CPB; L. decernlineata), for instance, is strongly inhibited by OCI and oryzacystatin I1 (OCII), but the cathepsin B-like activity, which constitutes an important part of the total endoproteolytic activity (Thie and Houseman, 1990), remains unchanged (Michaud et al., 1993). The presence of free (non-target) proteases in the in- sect midgut could lead, in this case, to a certain physiological complementa- tion of cathepsin H-related functions and/or to a significant degradation of the inhibitor, thereby affecting its effectiveness for the control of CPB.
*Abbreviations used: BVW, black vine weevil; E-64, trans-epoxysuccinyl-t-leucylamido-(4- guanidino) butane; GSH, reduced glutathione; GST, glutathione 5-transferase; HSA, human stefin A; IPTG, isopropyl-P-D-thiogalactoside; OCI, oryzacystatin 1; OCII, oryzacystatin II; PCR, poly- merase chain reaction; SDS, sodium dodecyl sulfate.

Inhibition of Coleoptera Cysteine Proteinases 453
In an attempt to find cystatins with a wider inhibitory spectrum against the proteinases of Coleoptera, the present study analyzed the response of CPB and black vine weevil (BVW; Otiovynchus sulcalus) digestive proteinases to a recombinant form of human stefin A (HSA), a cystatin well-known for its affinity to human cathepsin B (Barrett et al., 1986; Fong et al., 1989). After producing this inhibitor in Escherichia coli using the glutathione S-transferase (GST) gene fusion system (Smith and Johnson, 1988), its ability to comple- ment OCI-mediated inhibitions was assessed. The effects of the insect “non- target” proteases on the stability of both cystatins were also studied.
MATERIALS AND METHODS Reagents
Azocasein, BSA, tvans-epoxysuccinyl-L-leucylamido (4-guanidino) butane (E-641, isopropyl-P-o-thiogalactoside (IPTG), reduced glutathione (GSH), GSH- agarose beads, and papain (from papaya latex; E.C.3.4.22.2) were purchased from Sigma (St. Louis, MO). Cathepsin B (from human liver; E.C.3.4.22.1), cathepsin H (from human kidney; E.C.3.4.22.6), cathepsin L (from human liver; E.C.3.4.22.151, factor Xa (from human plasma; E.C.3.4.21.6), lysozyme (from hen egg white; E.C.3.2.1.17), and thrombin (from human plasma; E.C.3.4.21.5) were from Calbiochem (LaJolla, CA). Expression vectors pGEX-2T and pGEX- 3X were from I‘harmacia (Uppsala, Sweden). Electrophoretic reagents were from Bio-Rad (Richmond, CAI.
Construction of the Expression Vectors Large amounts of active cystatins were produced using the GST gene fu-
sion system (Smith and Johnson, 1988). For OCI (Abe et al., 1987), the plas- mid pGEX-3XOCI was constructed as described previously (Michaud et al., 1994). For HSA, the full encoding region from the inhibitor cDNA clone 242 (Kartasova et al., 1987) was amplified by polymerase chain reaction (PCR) using the following primers: 5’ AGGATCCAAAATGATACCTG 3’, compris- ing a BamHl cleavage sequence; and 3’ AGGTACCAAGGATCATGACTCAG 5’, comprising a KpnI cleavage sequence. After PCR, the HSA-encoding se- quence was subcloned into pUCl8 using a BamHI-KprzI digestion, verified by DNA sequencing, and transferred into the commercial vector, pGEX-2T, us- ing a BnrnHI-EcoRI digestion. This construct (pGEX-2THSA) encoded a 37- kDa fusion protein consisting of GST (26 kDa) and HSA (11 kDa). The two recombinant plasmids, pGEX-3XOCI and PGEX-~THSA, were transformed into E. coli JM109 for expression of the fusion proteins.
Expression, Purification, and Cleavage of the Fusion Proteins The expression procedure was adapted from Michaud et al. (1994). Briefly,
25 ml of an overnight culture of E . coli JM109 transformed with the appropri- ate expression vector was inoculated into 225 mi of LB medium containing 100 pg/ml ampicillin. The culture was incubated for 2 h at 37°C with vigor- ous agitation before adding IPTG to 0.1 mM final concentration. After incu- bation for various periods at 37”C, the cells were harvested by centrifugation at 8,0008 for 10 min, and suspended into 3 ml of 50 mM Tris, pH 8.0, contain-

454 Michaud et al.
ing 5% (w/v) sucrose, 50 mM EDTA, 5% (v/v) Triton X-100, and 0.025% (w/ v) lysozyme. After standing 5 min on ice, the bacterial lysate was centrifuged at 8,OOOg for 10 min at 4"C, and the supernatant containing soluble proteins was saved. Purification of the fusion proteins using GSH-agarose beads was performed as described previously (Michaud et al., 1994). Cleavage of the GST-OCI fusion with human factor Xa was done according to Nagai and Thogersen (1984). Cleavage of the GST-HSA fusion with human thrombin was done according to Guan and Dixon (1991). Purity of the protein prod- ucts was analyzed by 15% (w/v) SDS-PAGE (Laemmli, 1970).
Insect Proteinase Extracts Digestive proteinase extracts were prepared from adult insects as described
previously (Michaud et al., 1993), except that the pH of the extraction buffer was adjusted to 6.0 instead of 5.5. Protein concentration in the extracts was adjusted to 5 mg/ml with extraction buffer using the Bradford procedure (Bradford, 1976), with BSA as a protein standard.
Proteinase and Proteinase Inhibitor Assays
The proteinase assay procedure was adapted from a previously described method (Michaud et al., 1993). Unless otherwise indicated, the enzyme ex- tracts (5 p1 per assay) were incubated with 35 pl of assay buffer (100 mM citrate phosphate, pH 5.0 [BVWI or 6.0 [CPBI, 5 mM L-cysteine, 0.1% [v/vl Triton X-100) and 80 1.11 of 2% (w/v) azocasein dissolved in assay buffer, for 3 h at 37°C. After proteolysis, 300 pl of 10% (w/v) trichloroacetic acid was added to the mixture, and the residual azocasein was removed by centrifugation for 5 min at 13,000~. The supernatant (350 1.11) was added to 300 pl of 1N NaOH, and the absorbance ( A ) was measured at 440 nm using a Spectronic 1000 Plus spectrophotometer (Milton Roy, Rochester, NU). The A440 of blanks, which consisted of complete mixtures incubated for 0 h, was subtracted from each value. For inhibition assays and active-site titrations, proteinase inhibitors were diluted to the appropriate concentrations in the 35 p1 of assay buffer of the reaction mixture. The enzyme and inhibitor solutions were allowed to react for 15 min at 37°C before addition of the substrate. Ki values were esti- mated from Dixon plots (Dixon and Webb, 19791, as described previously (Michaud et al., 1995a).
Removal of OCI- and HSA-Sensitive Proteinases From Insect Crude Extracts OCI- and HSA-sensitive proteinases were removed from CPB and BVW
crude extracts using the GST fusion proteins pre-adsorbed onto GSH-agar- ose beads. Briefly, 1 ml of the GST/OCI (or GST/HSA) solution previously prepared from E. coli (2 mg/ml in 50 mM Tris, pH 8.0) was mixed with 1 ml of 50% (w/v) GSH-agarose beads suspended in 50 mM Tris, pH 8.0. After adsorption of the fusion protein on GSH for 15 min, the mixture was centrifuged for 10 min at 8,0OOg, and the supernatant was discarded. The CPB (or BVW) crude extract (1 ml) containing both OCI- (or HSA-) sensitive and OCI- (or HSA-) insensitive proteases was then mixed with the pellet composed of GSH-agarose beads with the adsorbed GST/OCI (or the GST/HSA) fusion. After the insect proteinases reacted with OCI

Inhibition of Coleoptera Cysteine Proteinases 455
(or HSA) for 15 min, the mixture was centrifuged for 10 min at 8,0008, and the supernatant corresponding to the CPB (or BVW) extract without OCI- (or HSA-) sensitive proteinases was collected for further analyses. All steps were carried out at room temperature, with gentle agitation dur- ing the two adsorption procedures.
Cleavage of OCI and HSA by the Insect "Non-Target" Proteases
The susceptibility of recombinant OCI and HSA to proteolytic cleavage by CPB and BVW proteases was assessed by incubating the purified inhibitors with the insect modified (sensitive proteinases-free) extracts. Briefly, 5 pl of the insect extracts (25 pg protein) diluted ten-fold in a 200 mM citrate phos- phate buffer, pH 6.0, were incubated for 60 min at 37°C with 10 pl of 400 mM sodium acetate, pH 5.0, and 5 pl (10 pg) of OCI (or HSA). After incubation, proteolysis was blocked by adding 20 pl SDS-PAGE sample buffer 2x (Laemmli, 1970) to the enzyme/inhibitor solutions, and the complete mix- tures were placed for 3 rnin in a boiling water bath. Analysis of the proteolysis products was done by 15% (w/v) SDS-PAGE (Laemmli, 1970). Semi-quantita- tive assay of cystatins in gels was done by densitometry, with a Microtek ScanMaker I1 scanner (Microtek Lab, Torrance, CA) and the image analysis software NIH Image 1.51 (National Institutes of Health, Bethesda, MD).
RESULTS High-Yield Bacterial Production of HSA
The time-course of accumulation of the GST/HSA fusion protein in E . coli gave a sigmoidal pattern roughly similar to the growth curve usually ob- served for this bacterium, with a maximal production of the fusion protein 16 h after induction by IPTG (Fig. 1A). The amount of fusion protein purified using GSH-agarose beads was mathematically predicted by the following equation:
GST/HSA (mg/l) = 9.14 t - 0.27 t2 - 7.64 (r2 = 0.93) (Eq. 1)
where t represents the time (h) after induction by IPTG, and r2 the correla- tion coefficient of the quadratic equation. The maximal yield of fusion prod- uct was estimated to -70 mg/l €. coli culture (Fig. lA), and the yield of the fusion cleavage with human thrombin was about 85% (not shown). Consis- tent with the vector construct, the molecular weights of GST/HSA and HSA purified after cleavage were estimated at -37 kDa (26-kDa GST + ll-kDa HSA) and -11 kDa, respectively (Fig. 1B). Both the fusion protein and the liberated recombinant HSA were biologically active, as shown by their Ki values against cysteine proteinases (Table 1). The affinities (Ki constants) of both GST/HSA and free recombinant HSA for papain and human cathepsins (Table 1) were similar to those reported for the natural HSA isolated from human skin (Barrett et al., 1986). Unlike recombinant OCI (Michaud et al., 1994), the recombinant human inhibitor showed measurable activity against human cathepsin B (Table 1).

456 Michaud et al.
0 10 20 Time after induction (h)
A Fig. I . Bacterial expression and purification of recombinant HSA using the CST gene fusion system. A: Time-course accumulation of the GSTIHSA fusion protein in E. coli JMI 09. Quantitation of the fusion was done according to Bradford ( I 976), after its purification on GSH-agarose beads. B: Cleavage of the CST/HSA fusion protein using human thrombin. Isolation and cleavage ot the fusion protein was monitored using 15% SDS-PAGE. Estimated molecular masses for GSTiHSA (lane 1) and free HSA (lane 2) were 37 and 11 kUa , respectively.
Differential Inhibition of CPB and BVW Cysteine Proteinases by OCI and HSA
Active-site titration of cysteine proteinases in CPB and BVW extracts was performed using the large-spectrum and highly specific cysteine proteinase inhibitor E-64 as an active-site titrant (Fig. 2). The cysteine proteinase content in the extracts of both insects was estimated to 7-8 pM (active-site equiva- lent) under the conditions used, and the inhibition by E-64 was partial, sug- gesting the occurrence in CPB and BVW midguts of proteases belonging to other mechanistic classes and/or of cysteine proteinases insensitive to E-64.
TABLE 1. Response of Papain and Human Cathepsins to Recombinant HSA
- 97.4 - 66.2 - 45.0
- 31.0
- 21.5
- 14.4
1 2 3
B
Ki value (nM) Protcinase GSTiHSA Free F1SA
Cathepsin B 4.3 3.0 Cathepsin H 0.79 0.5 Cathepsin L 3.0 2.1 Papain 0.08 0.04

Inhibition of Coleoptera Cysteine Proteinases 457
l:] 60 \ CPB
E 40 .z 20 x >
I . . . I . I x " . - ' - ' . I -
BVW
260 i! 'r \ ,.;, 4
4 40
20 - 0-1 . ' . a . ' . I ! ! - ' . *
0 2 4 6 8 50 100
E-64 (W) Fig. 2 . Active-site titration of CPB and BVW cysteine proteinases with E-64 resent the maximal inhibitions under the conditions used. Arrows indicate thr centration of E-64-sensitive cysteine proteinases in the extracts.
- - -.-.- 4
x > .z 20 - 0 .CI
BVW
260 ,.;, 4
4 40
20 - 0-1 . ' . a . ' . I ! ! - ' . *
0 2 4 6 8 50 100
E-64 (W) Fig. 2 . Active-site titration of CPB and BVW cysteine proteinases with E-64 resent the maximal inhibitions under the conditions used. Arrows indicate thr centration of E-64-sensitive cysteine proteinases in the extracts.
. Dashed lincs rep- ? approximate con-
When used in excess recombinant OCI and HSA had a partial effect, but un- like OCI the degree of inhibition by the human inhibitor increased with its concentration (Table 2). While the rates of enzyme activity determined in the presence of HSA were similar to those measured for OCI when the inhibitors were used in "limited" excess (e.g., 10 or 20 pM), the HSA-mediated inhibi- tions were significantly increased when larger concentrations were used (e.g., 100 yM; Table 2). Consistent with the wide inhibition spectrum of HSA for
TABLE 2. Inhibition of CPB and BVW Proteinases by E-64, OCI, and HSA
Residual activitv (%)a
Inhibitor Working concentration (pM) CPB BVW
E-64 10 12.9 a 37.7 a 20 13.0 a 38.1 a
100 12.7 a 37.4 a OCI 10 57.5 b 62.1 b
20 56.4 b 64.2 b 100 56.9 b 64.0 b
HSA 10 55.7 b 63.2 b 20 57.2 b 61.1 b
100 24.3 c 40.8 a
'Data are expressed as the remaining activity as comparcd to a control to which no inhibitor was added. In each column, means followed by the same letter are not significantly different, P=0.05 (Duncan's multiple range test). Each datum represents the mean of three detemnations.

458 Michaud et al.
human cathepsins (Table l), these observations suggest the occurrence in both CPB and BVW extracts of two cathepsin-like proteinase populations differ- entially inactivated by the human inhibitor. This putative two-step inhibition of the insect cysteine proteinases by HSA was confirmed by complementary inhibition studies, which did not show any complementary inhibition between HSA and OCI used at 10 pM concentrations (Fig. 3) . Despite a larger inhibi- tion spectrum for cysteine proteinases (Tables 1 and 2), recombinant HSA used at low concentrations thus seemed to cause partial inhibition of CPB and BVW cysteine proteinases similar to those observed with OCI (Fig. 3 ) . The partial inhibition of CPB E-64-sensitive proteinases probably resulted from the selective inactivation of cathepsin H-like proteinases, since OCI was pre- viously shown to selectively inhibit this fraction of CPB digestive protein- ases without altering the cathepsin B-like activity (Michaud et al., 1993). Globally, these results provide evidence that HSA inhibits CPB and BVW cys- teine proteinases in a two-step manner, the rapidly (strongly) inhibited pro- teinases corresponding to the fraction also recognized by OCI.
Removal of OCI- and HSA-Sensitive Proteinases From CPB and BVW Extracts OCI- and HSA-sensitive proteinases were removed from the insect extracts
by incubating the crude midgut preparations with the corresponding GST fusion proteins pre-adsorbed onto GSH-agarose beads (Fig. 4). The maximal inhibitions of CPB and BVW proteases by OCI, estimated at -45% and -35%, respectively, for the crude extracts (Table 21, dramatically decreased follow- ing removal of the sensitive enzymes during the affinity step (Fig. 4). Con- versely, the removal of HSA-sensitive proteinases using the GST/HSA fusion protein was partial, leaving an important fraction of HSA-sensitive cysteine proteinases in both extracts (Fig. 4). For either CPB and BVW, cysteine pro- teinases not removed from the crude extracts probably correspond to those "weakly" sensitive to HSA and insensitive to OCI (Table 2; Fig. 3) , as demon- strated by the negligible effect of OCI on the protease activity measured in the extracts modified using the GST/HSA-GSH-agarose beads (Fig. 4).
Effects of CPB and BVW Non-Target Proteases on the Stability of OCI and HSA The comparative effects of CPB and BVW OCI-insensitive proteases on the
stability of OCI and HSA were analyzed by incubating the partially purified
CPB BVW 100 50 0 Inhibition (%) 0 50 100
I ' I I I I L I I I I I I I I I I I I I I I
10 pM OCI + 10 pM HSA 100 pM OCI + 10 pM HSA 10 pM OCI + 100 pM HSA
1 0 0 pM OCI + 100 pM HSA - Fig. 3. Response of CPB and BVW cysteine proteinases to various combinations of C)CI and HSA. Inhibition values are expressed as the relative inhibitions of cysteine proteinases as com- pared to the E-64 inhibitions (arbitrarily fixed at 100%j. The control values corresponded to the activity measured in samples for which no inhibitor was added. Each datum represents the mean of three values * S.E.

Inhibition of Coleoptera Cysteine Proteinases 459
crude extract GSH-GST/OCI 0 GSH-GST/HSA
OCI HSA OCI HSA
CPB BVW Fig. 4. Inhibition of CPB and BVW proteinases by OCI and HSA used in large excess (100 kM), before and after GSH-GST/OCI (or GSH-GST/HSA) affinity chromatography. Results are expressed as the relative activity as compared to control values for which no inhibitor was added. Each bar represents the mean of three values 2 S.E.
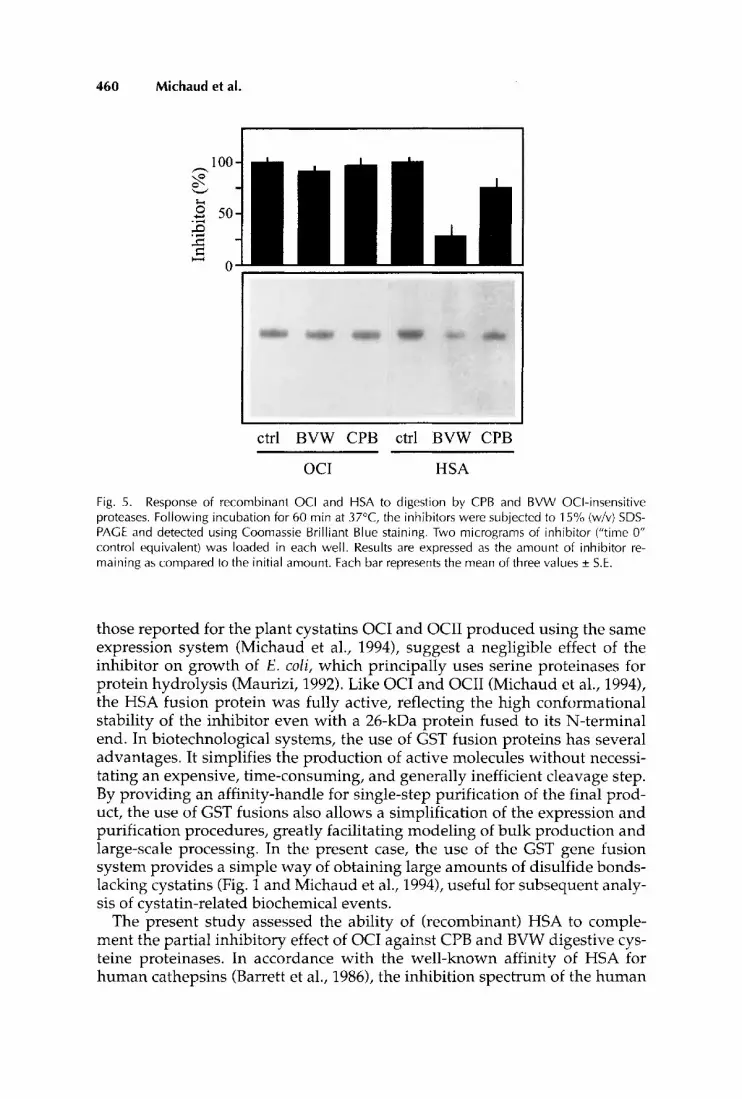
insect extracts with either cystatin for 60 min at 37°C (Fig. 5 ) . While OCI visibly remained unaffected, HSA was subjected to proteolysis by the insect proteases, leading to its disappearance in the enzyme mixture (Fig. 5). In con- trast with a recent study reporting the limited proteolysis of OCII by BVW non-target proteases (Michaud et al., 1995a), no stable intermediate of HSA was detected on gel, suggesting the presence of multiple sensitive cleavage sites susceptible to the action of the insect proteases on this inhibitor. To- gether with its apparently low affinity for a fraction of the insect cysteine proteinases, this apparent susceptibility of HSA to the action of the BVW pro- teases may contribute to the absence of complementary inhibitions between this inhibitor and OCI when low inhibitor concentrations are used (Fig. 3). A similar hydrolytic process might also partially explain the two-step inhibi- tion of CPB cysteine proteinases, although the HSA degradation was not as important as for BVW proteases (Fig. 5 ) . Interestingly, the inhibitions by ei- ther OCI (10 pM) and HSA (10 pM) in both CPB and BVW extracts remained unchanged over a 2 h period (Fig. 61, suggesting stable inhibitory effects for both cystatins. It is not clear whether the proteinase/cystatin complexes were resistant to proteolysis by the non-target proteases, or if both the enzyme and the inhibitor in the complexes were hydrolyzed following inhibition.
DISCUSSION
Recombinant HSA produced in E. coli as a GST/HSA fusion protein was as active against cysteine proteinases as the naturally occurring inhibitor from human skin (Barrett et al., 1986). The high yields obtained, comparable to
460 Michaud et al.
”
ctrl BVW CPB ctrl BVW CPB
OCI HSA
Fig. 5 . Response of recombinanl OCI and HSA to digestion by CPB and BVW OCI-insensitive proteases. Following incubation for 60 rnin at 37”C, the inhibitors were subjected to 15% (w/v) SDS- PAGE and detected using Coomassie Brilliant Blue staining. Two micrograms of inhibitor (“time 0” control equivalent) was loaded in each well. Results are expressed as the amount of inhibitor re- maining a5 compared to the initial amount. Each bar represents: the mean of three values * S.E.
those reported for the plant cystatins OCI and OCII produced using the same expression system (Michaud et al., 1994), suggest a negligible effect of the inhibitor on growth of E . coli, which principally uses serine proteinases for protein hydrolysis (Maurizi, 1992). Like OCI and OCII (Michaud et al., 1994), the HSA fusion protein was fully active, reflecting the high conformational stability of the inhibitor even with a 26-kDa protein fused to its N-terminal end. In biotechnological systems, the use of GST fusion proteins has several advantages. It simplifies the production of active molecules without necessi- tating an expensive, time-consuming, and generally inefficient cleavage step. By providing an affinity-handle for single-step purification of the final prod- uct, the use of GST fusions also allows a simplification of the expression and purification procedures, greatly facilitating modeling of bulk production and large-scale processing. In the present case, the use of the GST gene fusion system provides a simple way of obtaining large amounts of disulfide bonds- lacking cystatins (Fig. 1 and Michaud et al., 1994), useful for subsequent analy- sis of cystatin-related biochemical events.
The present study assessed the ability of (recombinant) HSA to comple- ment the partial inhibitory effect of OCI against CPB and BVW digestive cys- teine proteinases. In accordance with the well-known affinity of HSA for human cathepsins (Barrett et al., 1986), the inhibition spectrum of the human

Inhibition of Coleoptera Cysteine Proteinases 461
.e 20
.- c.' 0 0 rd
BVW 4 ; 100 d -U .A
80
60
40
20
0
2
0 15 60 90 Incubation time (min)
Fig. 6. Inhibition of CPB and BVW proteinase activity by recombinant OCI and HSA after various incubation times. The inhibitors were incubated in excess (10 pM) with the insect crude extracts at 37°C for various incubation time< (0, 15, 60, 90 min). An azocaseinase assay was then performed for 30 min at 37OC, as described in Materials and Methods. Results are ex- pressed as the relative activity compared to control values for which no inhibitor was added. Each bar represents the mean of three values k S.E.
inhibitor against the insect cysteine proteinases was wider than that of OCI, but large amounts of inhibitor were needed to complement the effect of the plant cystatin. This low inhibitory efficiency of HSA against the insect cys- teine proteinases was probably due to a weak affinity between the human cystatin and the OCI-insensitive cysteine proteinases, and to the occurrence of exposed sites susceptible to proteolysis on HSA. Obviously, these observa- tions are based on biochemical (in vitro) data, and they may be of limited physiological significance. Several studies have reported the compartmental- ization of proteases in insect midguts associated with the sequential diges- tion of dietary proteins (Terra and Ferreira, 1994). Hydrolysis of HSA could be initiated by the action of a specific protease not present in the midgut lumen, preventing proteolysis of HSA in vivo. As suggested by the apparent absence of detectable degradation intermediates during hydrolysis of HSA by the non-target proteases from both CPB and BVW, it however appears plausible that the hydrolysis of HSA results from the action of non-specific digestive proteases acting at various sites on the inhibitor. The major protein-

462 Michaud et al.
ases identified so far in insect midguts belong to the trypsin, chymotrypsin, elastase, cathepsin B, cathepsin D, or cathepsin H proteinase families (Terra and Ferreira, 1994), which all include proteinases with little specificity. Sim- ply put, hydrolysis of HSA would be due to the accessibility of multiple sen- sitive cleavage sites on HSA easily recognized by a variety of insect digestive proteinases, rendering useless the wide inhibitory spectrum noted for this inhibitor against CPB and BVW cysteine proteinases.
The presence of either insensitive or weakly sensitive proteolytic enzymes in the digestive tract of insects leads to question the potential effectiveness of cystatins for the control of Coleoptera. Partial inhibition of digestive cysteine proteinases, notably, indicates the need for additional studies about a pos- sible physiological complementation by the non-target cysteine proteinases, which might allow partial or even complete recovery of inhibited proteolytic functions (Bolter and Jongsma, 1995). The important question of cystatin sta- bility in the insect midgut also needs additional studies (Orr et al., 1994; Michaud et al., 1995a). The presence of potential (sensitive) cleavage sites in the primary structure of certain plant cystatins, recently reported for OCII (Michaud et al., 1995a), could cause their rapid breakdown in the digestive tract of target insects, thereby affecting the global efficiency of the control approach. The interactions between insect non-target proteases and plant cystatins may possibly explain the very limited amount of information now available in the literature about the effective use of cystatins for the control of economically important coleopteran pests like the CPB and the BVW. Work is currently underway to address the putative occurrence of such "satellite" molecular interactions in the digestive tract of both species.
LITERATURE CITED Abe K, Hiroto K, Arai S (1987): Molecular cloning of a cysteine proteinase inhibitor of rice
(oryzacystatin). J Biol Chem 262:16793-16797.
Barrett AJ, Rawlings ND, Davies ME, Machlcidt W, Salvesen G, Turk V (1986): Cysteine pro- teinase inhibitors of the cystatin superfamily. In Barrett AJ, Salvesen G (eds): Proteinase Inhibitors. Amsterdam: Elsevier, pp 515-569.
Benchekroun A, Michaud D, Nguyen-Quoc B, Overney S, Desjardins Y, Yelle S (1995): Synthe- sis of active oryzacystatin I in transgenic potato plants. Plant Cell Rep 14:585-588.
Bolter CJ, Jongsma MA (1995): Colorado potato beetles (Leptinotansa decemliizeatni adapt to pro- teinase inhibitors induced in potato leaves by methyl jasmonate. J Insect Physiol41:1071-1078.
Ronad6-Bottino M (1993): Defense du colza contre les insectes phytophages d6predateurs: Etude d'une stratkgie bas6e sur l'expression d'inhibiteurs de protkases dans la plante. PhD The- sis, Universite de Paris-Sud, Centre d'Orsay.
Bradford MM (1976): A rapid and sensitive method for the quantitation of microgram quanti- ties of protein using the principle of protein-dye binding. Anal Biochem 72:248-254.
Chen M-S, Johnson B, Wen L, Muthukrishnan S, Kramer KJ, Morgan TD, Reeck GR (1992): Rice cystatin: Bacterial expression, purification, cysteine proteinase inhibitory activity, and insect growth suppressing activity of a truncated form of the protein. Protein Expres Purif 3:41-49.

Inhibition of Coleoptera Cysteine Proteinases 463
Dixon M, Webb EC (1979): Enzyme inhibition and activation. In Dixon M, Webb EC (eds): Enzymes. London: Longmans, pp 332-380.
Fong D, Kartasova T, Sloane BF, Chan MM (1989): Bacterial expression of human cysteine proteinase inhibitor stefin A. FEBS Lett 257:55-58.
Gillikin JW, Bevilocqua S, Graham JS (1992): Partial characterization of digestive tract pro- teinases from Western corn rootworm, Diabrotica virgifeva. Arch Insect Biochem Physiol 191285-298.
Guan KL, Dixon JE (1991): Eukaryotic proteins expressed in Eschevichia coli: An improved thrombin cleavage and purification procedure of fusion proteins with glutathione S-trans- ferase. Anal Biochem 192:262-267.
Hilder VA, Gatehouse AMR, Boulter D (1993): Proteinase inhibitor approach. In Kung S-D, Wu R (eds): Transgenic Plants: Engineering and Utilization, Vol. 1. New York Academic Press, pp 317-338.
Hines ME, Osuala CI, Nielsen SS (1991): Isolation and partial characterization of a soybean cystatin cysteine proteinase inhibitor of Colropteran digestive proteolytic activity. J Agric Food Chem 39:1515-1520.
Hosoyama H, Irie K, Abe K, Arai S (1994): Oryzacystatin exogenously introduced into proto- plasts and regeneration of transgenic rice. Biosci Biotechnol Biochem 58:1500-1505.
Kartasova T, Cornelissen BJC, Belt P, Van de Putte P (1987): Effects of LV, 4-NQO and TPA on gene expression in cultured human epidermal keratinocytes. Nucleic Acids Res 15:59455962.
K o n d o H, Abr K, Nishimura I, Watanabe H, Emori Y, Arai S (1990): Two distinct cystatin species in rice seed with different specificities against cysteine proteinases. Molecular clon- ing, expression, and biochemical studies on oryzacystatin-11. J Biol Chem 26515832-15837.
Laemmli UK (1970): Cleavage of structural proteins during the assembly of the head of bacte- riophage T4. Nature 227:680-685.
Leple J-C, Bonade-Bottino M, Augustin S, Delplanque A, Dumanois V, Pilate G, Cornu D, Jouanin L (1995): Toxicity to Chrysomrla fremulae (Coleoptera: Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor. Mol Breed 1:319-328.
Liang C, Brookhart G, Feng GH, Reeck GR, Kramer KJ (1991): Inhibition of digestive protein- ase of stored grain Coleoptcra by oryzacystatin, a cysteine proteinase inhibitor from rice seeds. FEBS Lett 278:139-142.
Masoud SA, Johnson LB, White FF, Reeck GR (1993): Expression of a cysteine proteinase in- hibitor (oryzacystatin-I) in transgenic tobacco plants. Plant Mol Biol21:655-663.
Maurizi MR (1992): Proteases and protein degradation in Eschrrichia coli. Experientia 48:17%201.
Michaud D, Nguyen-Quoc B, Yelle S (1993): Selective inhibition of Colorado potato beetle cathepsin H by oryzacystatins 1 and 11. FEBS Lett 331:173-176.
Michaud D, Nguyen-Quoc B, Yelle S (1994): Production of oryzacystatins I and I1 in Eschevi- chia coli using the glutathione S-transferase gene fusion system. Biotechnol Frog 10:155-159.
Michaud D, Cantin L, Vrain TC (1995a): Carboxy-terminal truncation of oryzacystatin I1 by oryzacystatin-insensitive insect digestive proteinases. Arch Biochem Biophys 322: 469-474.

464 Michaud et al.
Michaud D, Bernier-Vadnais N, Overney S, Yelle S (1995b): Constitutive expression of diges- tive cysteine proteinase forms during development of the Colorado potato beetle, Leptinutarsa decemlineata Say (Coleoptera: Chrysomelidae). Insect Biochem Mol Biol25:1041-1048.
Nagai K, Thogersen C (1984): Generation of P-globin by sequence-specific proteolysis of a hybrid protein produced in Escherichia coli. Nature 309:810-812.
Orr GL, Strickland JA, Walsh TA (1994): Inhibition of Diabrutica larval growth by a multicystatin from potato tubers. J Insect Physiol40:893-900.
Smith DB, Johnson KS (1988): Single-step purification of polypeptides expressed in Escheri- chiu coli as fusions with glutathione S-transferase. Gene 6:31-40.
Terra WR, Ferreira C (1994): Review. Insect digestive enzymes: Properties, compartmentaliza- tion and function. Comp Biochem Physiol 109B:1-62.
Thie NM, Houseman JG (1990): Identification of cathepsin B, D, and H in the larval midgut o f Colorado potato beetle, Leptinutarsa decemlinea ta Say (Coleoptera: Chrysomelidae). Insect Biochem 20:313-318.




















