
Enzymes active in the areas undergoing cartilage resorption during the development of the secondary...
18
Enzymes Active in the Areas Undergoing Cartilage Resorption During the Development of the Secondary Ossification Center in the Tibiae of Rats Aged 0 –21 Days: II. Two Proteinases, Gelatinase B and Collagenase-3, Are Implicated in the Lysis of Collagen Fibrils MARIA ANTONIETTA DAVOLI, 1 LISA LAMPLUGH, 1 ANNIE BEAUCHEMIN, 1 KEVIN CHAN, 1 SYLVIE MORDIER, 2 JOHN S. MORT, 2,3 GILLIAN MURPHY, 4 ANDREW J.P. DOCHERTY, 5 CHARLES PHILIPPE LEBLOND, 1,6 AND EUNICE R. LEE 1,3 * 1 Electron Microscopy Unit, Shriners Hospitals for Children, Montreal, Quebec, Canada 2 Joint Diseases Laboratory, Shriners Hospitals for Children, Montreal, Quebec, Canada 3 Division of Surgical Research, Department of Surgery, McGill University, Montreal, Quebec, Canada 4 School of Biological Sciences, University of East Anglia, Norwich, England 5 Celltech Group plc., Slough, United Kingdom 6 Department of Anatomy and Cell Biology, McGill University, Montreal, Quebec, Canada ABSTRACT In the transformation of the cartilaginous epiphysis into bone, the first indi- cation of change in the surfaces destined for re- sorption is the cleavage of aggrecan core protein by unidentified matrix metalloproteinases (MMPs) (Lee et al., this issue). In cartilage areas undergoing resorption, the cleavage leaves as su- perficial, 6-mm-thick band of matrix, referred to as “pre-resorptive layer.” This layer harbors G1- fragments of the aggrecan core protein within a framework of collagen-rich fibrils exhibiting var- ious stages of degeneration. Investigation of this layer in every resorption area by gelatin histozy- mography and TIMP-2 histochemistry demon- strates the presence of an MMP whose histozy- mographic activity is inhibited by such a low dose of the inhibitor CT1746 as to identify it as gelatinase A or B. Attempts at blocking the his- tozymographic reactions with neutralizing anti- bodies capable of inhibiting either gelatinase A or B reveals that only those against gelatinase B do so. Immunostaining of sections with anti– ge- latinase B IgG confirms the presence of gelati- nase B in every pre-resorptive layer, that is, at the blind end of excavated canals (stage I; 6-day- old rats), at sites along the walls of the forming marrow space (stage II; 7days), at sites within the walls of this space as it becomes the ossification center (stage III; 9 days) and along the wall of the maturing center (stage IV; 10 –21 days). We also report the presence of collagenase-3 in precisely the same sites, possibly as active enzyme, but this remains to be proven. Because the results reveal that collagenase-3 is present beside gelatinase B in every pre-resorptive layer and, because these sites exhibit various signs of degradation includ- ing fibrillar debris, reduction in fibril number, or overt loss, we propose that gelatinase B and col- lagenase-3 mediate the lysis of this pre-resorp- tive layer—most likely through a cooperative at- tack leading to the disintegration of the collagen fibril framework. © 2001 Wiley-Liss, Inc. Key words: gelatinase B; collagenase-3; matrix metalloproteinases; skeletal devel- opment; cartilage lysis; vascular in- vasion; zymography INTRODUCTION During the transformation of a cartilage model into bone, the secondary ossification center is formed in the epiphysis as the result of complex excavations. Thus in the rat, canals are dug from the surface toward the center of the epiphysis, where their ends fuse to give rise to the initial marrow space which, with the acqui- sition of ossification units, becomes the secondary ossi- fication center (Levene, 1964; Delgado-Baeza et al., 1991a,b). These excavations are the result of cartilage resorptions that have been localized in a series of sites and classified into two types, according to whether the resorption is associated with bone formation or not (Lee et al., 2001, this issue). The sites involved in bone formation are referred to as “ossification-coupled”, whereas those independent of it are called “free” (Table 1). The resorption of cartilage suggests the lysis of the two major cartilage components: the proteoglycan “ag- Grant Sponsor: Shriners of North America; Grant number: 8530. Dr. Sylvie Mordier is currently at INRA THEIX, Unite Nutrition Cellulaire and Moleculaire, 63122 Saint Genes Champanelle, France. *Correspondence to: Dr. Eunice R. Lee, Electron Microscopy Unit, Shriners Hospitals for Children, 1529 Cedar Avenue, Montreal, Que- bec H3G 1A6, Canada. E-mail: [email protected] Received 19 January 2001; Accepted 24 April 2001 Published online 13 July 2001; DOI 10.1002/dvdy.1160 DEVELOPMENTAL DYNAMICS 222:71– 88 (2001) © 2001 WILEY-LISS, INC.
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Enzymes active in the areas undergoing cartilage resorption during the development of the secondary...

Enzymes Active in the Areas Undergoing CartilageResorption During the Development of the SecondaryOssification Center in the Tibiae of Rats Aged 0–21 Days:II. Two Proteinases, Gelatinase B and Collagenase-3, AreImplicated in the Lysis of Collagen FibrilsMARIA ANTONIETTA DAVOLI,1 LISA LAMPLUGH,1 ANNIE BEAUCHEMIN,1 KEVIN CHAN,1
SYLVIE MORDIER,2 JOHN S. MORT,2,3 GILLIAN MURPHY,4 ANDREW J.P. DOCHERTY,5
CHARLES PHILIPPE LEBLOND,1,6 AND EUNICE R. LEE1,3*1Electron Microscopy Unit, Shriners Hospitals for Children, Montreal, Quebec, Canada2Joint Diseases Laboratory, Shriners Hospitals for Children, Montreal, Quebec, Canada3Division of Surgical Research, Department of Surgery, McGill University, Montreal, Quebec, Canada4School of Biological Sciences, University of East Anglia, Norwich, England5Celltech Group plc., Slough, United Kingdom6Department of Anatomy and Cell Biology, McGill University, Montreal, Quebec, Canada
ABSTRACT In the transformation of thecartilaginous epiphysis into bone, the first indi-cation of change in the surfaces destined for re-sorption is the cleavage of aggrecan core proteinby unidentified matrix metalloproteinases(MMPs) (Lee et al., this issue). In cartilage areasundergoing resorption, the cleavage leaves as su-perficial, 6-mm-thick band of matrix, referred toas “pre-resorptive layer.” This layer harbors G1-fragments of the aggrecan core protein within aframework of collagen-rich fibrils exhibiting var-ious stages of degeneration. Investigation of thislayer in every resorption area by gelatin histozy-mography and TIMP-2 histochemistry demon-strates the presence of an MMP whose histozy-mographic activity is inhibited by such a lowdose of the inhibitor CT1746 as to identify it asgelatinase A or B. Attempts at blocking the his-tozymographic reactions with neutralizing anti-bodies capable of inhibiting either gelatinase Aor B reveals that only those against gelatinase Bdo so. Immunostaining of sections with anti–ge-latinase B IgG confirms the presence of gelati-nase B in every pre-resorptive layer, that is, atthe blind end of excavated canals (stage I; 6-day-old rats), at sites along the walls of the formingmarrow space (stage II; 7days), at sites within thewalls of this space as it becomes the ossificationcenter (stage III; 9 days) and along the wall of thematuring center (stage IV; 10–21 days). We alsoreport the presence of collagenase-3 in preciselythe same sites, possibly as active enzyme, but thisremains to be proven. Because the results revealthat collagenase-3 is present beside gelatinase Bin every pre-resorptive layer and, because thesesites exhibit various signs of degradation includ-ing fibrillar debris, reduction in fibril number, orovert loss, we propose that gelatinase B and col-
lagenase-3 mediate the lysis of this pre-resorp-tive layer—most likely through a cooperative at-tack leading to the disintegration of the collagenfibril framework. © 2001 Wiley-Liss, Inc.
Key words: gelatinase B; collagenase-3; matrixmetalloproteinases; skeletal devel-opment; cartilage lysis; vascular in-vasion; zymography
INTRODUCTION
During the transformation of a cartilage model intobone, the secondary ossification center is formed in theepiphysis as the result of complex excavations. Thus inthe rat, canals are dug from the surface toward thecenter of the epiphysis, where their ends fuse to giverise to the initial marrow space which, with the acqui-sition of ossification units, becomes the secondary ossi-fication center (Levene, 1964; Delgado-Baeza et al.,1991a,b). These excavations are the result of cartilageresorptions that have been localized in a series of sitesand classified into two types, according to whether theresorption is associated with bone formation or not(Lee et al., 2001, this issue). The sites involved in boneformation are referred to as “ossification-coupled”,whereas those independent of it are called “free”(Table 1).
The resorption of cartilage suggests the lysis of thetwo major cartilage components: the proteoglycan “ag-
Grant Sponsor: Shriners of North America; Grant number: 8530.Dr. Sylvie Mordier is currently at INRA THEIX, Unite Nutrition
Cellulaire and Moleculaire, 63122 Saint Genes Champanelle, France.*Correspondence to: Dr. Eunice R. Lee, Electron Microscopy Unit,
Shriners Hospitals for Children, 1529 Cedar Avenue, Montreal, Que-bec H3G 1A6, Canada. E-mail: [email protected]
Received 19 January 2001; Accepted 24 April 2001Published online 13 July 2001; DOI 10.1002/dvdy.1160
DEVELOPMENTAL DYNAMICS 222:71–88 (2001)
© 2001 WILEY-LISS, INC.

grecan” and the “collagen” fibrils (Cole and Wezeman,1985, 1987a,b; Cole and Cole, 1989). The lysis of one ofthe two, aggrecan, has been demonstrated in the prox-imal epiphysis of the rat by Lee et al. (2001, this issue).The enzyme responsible for the aggrecan lysis has notbeen directly identified; instead, its presence has beenrevealed by the detection of a neoepitope known toarise from the cleavage of the aggrecan core protein bya so far unidentified matrix metalloproteinase (MMPs).In the “free” sites, a space approximately 6 mm thicklocated behind the surface undergoing resorption andknown as “pre-resorptive layer” contains the smaller ofthe two fragments arising from the aggrecan cleavage,the so-called G1-341 fragment. This fragment carriesthe neoepitope recognition site and is retained in thetissue. Meanwhile, the larger fragment that comprisesthe bulk of the core protein is lost from the tissue, sothat the pre-resorptive layer is essentially composed ofG1 fragments with some collagen fibrils. With the lossof the aggrecan mass, the pre-resorptive layer is weak-ened in preparation for the final resorption (Lee et al.,2001, this issue).
In the present article, the enzymes responsible forthe lysis of the other major cartilage component, colla-gen, are being investigated. Because the cartilage col-lagen is in the form of heterogeneous fibrils (for reviewsee Eyre and Wu, 1995), the goal is to find whichenzymes are involved in the break-up of these collagenfibrils. The approach is based on the use of the histozy-mographic technique (Lee et al., 1999). This techniqueconsists in applying unfixed frozen sections of a tissueonto slides bearing a target substrate that can be di-gested by the enzyme suspected of being present in thetissue, as initially done by Adams and Tuqan (1961)and Daoust (1965). In the present case, the slides havebeen coated with photographic emulsion that is thenblackened by exposure to light, so that the target sub-strate is the emulsion gelatin, whose digestion is madevisible by the loss of its black silver grains and theresulting lightening. The histozymographic techniquehas been used in three successive steps: (1) simplehistozymography, (2) histozymography performed inthe presence of enzyme inhibitors, and (3) histozymog-raphy performed in the presence of “neutralizing” an-tibodies.
In addition, a second approach has been taken toconfirm the presence of an MMP. To do so, antibodiesagainst the candidate enzyme have been used for im-munostaining epiphyseal sections at the various stagesin the hope of finding whether this enzyme is actuallylocated in the areas inducing histozymographic reac-tions.
Lastly, TIMP-2, an inhibitor of MMPs, has beenfound to prevent the appearance of the histozymo-graphic reactions. To find out whether or not thisTIMP-2 binds to the tissue, and if so, to further delin-eate the sites in which it binds, it has been biotinyl-ated, then applied to fresh tissue sections and detectedthrough its biotin label.
By these various approaches, the conclusion hasbeen reached that at least two enzymes are involved inlysing the cartilage collagen in the excavations re-quired for the building of a secondary ossification cen-ter. These enzymes complete the lytic process that isinitiated by the cleavage of aggrecan core protein.
RESULTSChanges in the Tibial Head Architecture inPostnatal Rats
A few days before and up to 5 days after birth, thetibia is composed of an ossifying diaphysis between twosolidly cartilaginous epiphyses (Fig. 1A). At approxi-mately 6 days of age, a series of cartilage resorptionsare initiated in the proximal epiphysis. These pro-cesses are divided into four stages, as described inFigure 2 of Lee et al. (2001, this issue) and summarizedhere in Table 1.
Histozymographic Reactions
Exposure of epiphyseal sections from prenatal rats(E20) to blackened gelatin emulsion for 4-hr results intwo histozymographic reactions seen as light bands,that correspond to the metaphyseal borders, where en-dochondral bone formation is taking place (Fig. 1B). Atstage I, when the emergence of cartilage canals hascommenced, exposure of sections for the same amountof time results in small, more or less intense histozy-mographic reactions (Fig. 2B) which, by comparisonwith the sections (Fig. 2A), are assigned to the blindends of the canals. In addition, an intense band-like
TABLE 1. Stages in the Development of the Rat Proximal Tibial Epiphysis
Stage Description of events Resorption site Resorption modeOnset (range)in days of age
I Canals emerge Canal blind end Free 6 (4–7)II Marrow space arises All marrow space walls Free 7 (6–8)III Hypertrophic chondrocytes appear
on proximal wall of marrow spaceTransverse septae Ossification-coupled 9 (8–10)
Marrow space expands mainly onthe distal wall
Distal wall Free
IV Secondary growth plate isdelineated on the proximal wall
Transverse septae Ossification-coupled 10 (9–11)
Distal wall recedes Epiphyseal border Ossification-coupled(but some free segments)
72 DAVOLI ET AL.

reaction at the base of the figure corresponds to theendochondral bone formation under way at the me-taphyseal border, where active cartilage resorption hasbeen shown to take place (Lee et al., 1998, 1999, 2001[this issue]). At stage II, a histozymographic reaction isseen on all marrow space walls or only on part of themas shown by comparing Figure 2C and D. A similarsituation is observed at stages III and IV when a his-tozymographic reaction is observed between as well asover groups of hypertrophic chondrocytes (Fig. 2E,F).The band-like reaction at the base of the figure, whichis clear-cut in Figure 2D and distinguishable in Figure2F, again corresponds to the area of endochondral boneformation at the metaphyseal border.
When exposure time is reduced to 2 hr (Fig. 3A), theintensity of the histozymographic reactions is muchless than in a serial section exposed 4 hr (Fig. 3B).Thus, it is concluded that a proteinase endowed withgelatinolytic activity is reacting at a time-related ratein the sites defined by histozymography.
Effect of Inhibitors on the HistozymographicReactions
To identify the enzyme responsible for the gelatino-lytic activity detected by histozymography, the firstapproach has been to repeat the procedure in the pres-ence of an inhibitor of known specificity during thereaction incubation. In this way, inhibitors of cysteine,serine, and aspartic proteinases do not interfere withthe production of histozymograms by sections at stagesI–IV (data not shown), but the natural MMP inhibitorTIMP-2 does inhibit the histozymographic reaction(Table 2). Another MMP inhibitor, CT1746, induces apartial inhibition at 0.04 nM (Fig. 4B) and a completeone at 1 nM concentration (Table 2). Anderson et al.(1996) have shown the following inhibition constants(Ki): 0.04 nM for gelatinase A (MMP-2), 0.17 nM forgelatinase B (MMP-9), 10.9 nM for stromelysin-1(MMP-3), 132 nM for interstitial collagenase (MMP-1),and 136 nM for matrilysin (MMP-7); therefore, the 0.04nM range observed by histozymography with this in-hibitor (Table 2) indicates that the reactivities are pro-duced by gelatinase A or B.
Effect of Neutralizing Antibodies on theHistozymographic Reactions
The second approach to the gelatinolytic enzymeidentification has been to expose stage I–IV tibialheads to neutralizing antibodies against gelatinase Aor B before the histozymographic procedure in the hopethat one of the two would prevent the reactions fromappearing. To test for the presence of gelatinase A,three doses of sheep anti-human gelatinase A (0.5, 1.0,5.0 mg/ml) have been applied to rat tibial head sectionsfor 30 min at 37°C, then again during contact withblackened emulsion at room temperature, but the pro-duction of the histozymograms has not been prevented(Table 3). However, when two concentrations of sheepanti-pig gelatinase B (0.5 mg or 1.0 mg/ml of IgG) havebeen applied to sections under the same conditions, thelower dosage has partially prevented the reaction,whereas the higher one has inhibited it completely(Table 3), whether the tibial heads are of stage I (Fig.5A–C), stage II (not shown), stage III (Fig. 5D–F), orstage IV (not shown). Because the gelatinolytic protein-ase is neutralized by the anti–gelatinase B IgG, thereactions are attributed to gelatinase B. The sugges-tion is that gelatinase B is an enzyme located withinthe histozymographically reactive sites of the tissue.
Immunolocalization of Gelatinase B
In the hope of confirming the presence of gelatinaseB in histozymographically reactive sites, the enzymehas been looked for by an immunohistochemicalmethod. To do so, 8-mm-thick sections of stage I–IV rattibial heads have been immunostained with anti–gela-tinase B IgGs — an approach that detects not only theactive enzyme but also its inactive proenzyme and theTIMP-inhibited form of the enzyme. With this reserva-
Fig. 1. Histozymographic reactions (B) produced by a section of thetibia from a prenatal rat at embryonic day 20 (E20). A: The structure ofthe tibia showing two white epiphyses and, between them, a dottedshading indicative of the extent of the primary ossification center. B: Twolightened areas are apparent in the blackened emulsion. These occuradjacent to the metaphyseal interface of the two epiphyses, hereafterreferred to as metaphyseal borders. These borders are identified byarrows in 1A). Both are sites of active cartilage resorption. Scale bar 5250 mm in B (applies to A,B).
73GELATINASE B AND COLLAGENASE-3, AGENTS IN COLLAGEN LYSIS

Fig. 2. Comparison between tibial head section at left, and the his-tozymograms produced when placed in contact for 4 hr with blackenedemulsion slides, at right, shown at stage I (A,B), stage II (C,D), and earlystage IV (E,F). A: The section separated from the emulsion and stainedwith toluidine blue exhibits a large canal and two small ones, all openingat the free surface of the cartilaginous epiphysis. B: The emulsion revealsthree discrete histozymographic reactions (arrows), which can be as-signed to the blind ends of the three canals, whereas the band-likereaction at the base of the figure (arrows, B) corresponds to the metaph-yseal border of the epiphysis. C: The section exhibits one narrow canal,which opens above at the free surface of the cartilaginous epiphysis andis continuous below with the marrow space (MS). D: The emulsion
reveals a histozymographic reaction (arrow) which is assigned to thelateral wall of the marrow space (MS) while the canal is unreactive.E: The section exhibits one canal and a marrow space around whichgroups of hypertrophic chondrocytes can be distinguished (larger ar-rows). F: The emulsion reveals a circular shaped lightening of the emul-sion, which is assigned to the interface of the marrow space walls. In B,D, and F, the band-like reaction at the base of the figure, which iscomplete in the first two and interrupted in F, corresponds to the metaph-yseal border of the epiphysis. The interpretation is that tissue sites facingthe lightened emulsion contain a gelatinolytic enzyme. Scale bar 5 500mm in A (applies to A–F).

tion in mind, three immunoreactive sites have beenidentified: the canal walls with emphasis on the blindends at stage I (Fig. 6B,C), segments of the wall in theforming marrow space at stage II (Fig. 6D), and alongthe edges of the wall containing groups of hypertrophicchondrocytes at stages III and IV (not shown). Hence,these sites include gelatinase B. Finally, because theimmunoreactivity coincides with sites shown to havegelatinolytic activity by histozymography, it is con-cluded that these sites contain the active form of theenzyme.
For the localization of gelatinase B at the electron mi-croscopic (EM) level, thin sections of stage I epiphyseshave been immunostained with 10 mg/ml of anti–gelati-nase B IgGs. The blind ends of the canals display areaction visualized as an accumulation of dark dots (Fig.7A). The reaction in the depicted photomicrograph ex-tends from the free surface of the canal down into thecartilage matrix to a depth of 4.0 6 0.85 mm. This thick-ness varies in other sites. We consider that Figures 7Aand B correspond to the region mentioned above underthe name “pre-resorptive layer.” In contrast, the rest ofthe matrix is generally free of dark dots (lower rightcorner of Fig. 7A) and so are the sections exposed topreimmune IgG for control (Fig. 7B).
To examine the structure of the pre-resorptive layer,EM sections of the canal blind end have been preparedby routine methods, as shown in Figure 7C, in whichthe lumen of the canal is seen at left. At the rightextremity of the figure, normal cartilage matrix (CM)displays intact collagen fibrils between which fine pro-teoglycan particles are scattered. The rest of the figureexhibits a decreasing gradient of fibril length andstaining density, whereas the fine proteoglycan parti-cles are rare or absent. The matrix close to the freesurface of the canal (indicated by three parallel arrows)appears pale with a few unidentifiable elements.
Immunolocalization of TIMP-2
A second approach has been to use TIMP histochem-istry to more precisely pin down the location of activegelatinase B in the tissue. Since TIMP-2 was known toform complexes with active MMPs in vitro resulting intheir inhibition (Butler et al., 1999), and since histozy-mography revealed that TIMP-2 can block the gelatino-
lytic activity of gelatinase B (Table 2), we have at-tempted to use TIMP-2 to localize the enzyme intissues. To do so, recombinant TIMP-2 labeled withbiotin has been applied to sections of stage I–IV rattibial heads. The labeled TIMP-2 retained within thesections has then been detected by treating them firstwith the avidin/biotin horseradish peroxidase complex,then with 3,39-diaminobenzidine (DAB). In practice,adjacent serial sections have been cut in pairs at 20mm. The biotinylated TIMP-2 has been applied to onesection of each pair for its localization, whereas his-tozymography has been used to detect gelatinolyticsites in the other. For the localization, control wasprovided by replacing the biotinylated TIMP-2 eitherwith TIMP-2 carrying no biotin label (data not shown)or with biotinylated recombinant, noninhibitory hu-man TIMP-1, produced as either a glycosylated or non-glycosylated form. The TIMP-1 control has allowed usto assess the amount of staining on the tissue sectionsthat could be attributed to nonspecific protein binding,which occurs, but is low (Fig. 8C,G). In contrast, thebiotinylated TIMP-2 is visualized as a brown reactionband. It is, thus, bound at stage I to canal blind ends(Fig. 8A,B) and metaphyseal border (Fig. 8A). At stageIII, TIMP-2 is seen to bind to the walls of the marrowspace (Fig. 8D) either in association with hypertrophicchondrocytes (Fig. 8E, small arrows) or in the siteslacking these cells (Fig. 8E, large arrows), as well as atthe metaphyseal border (Fig. 8D). At stage IV, theproximal wall of the marrow space, associated withhypertrophic chondrocytes is still reactive, as are theepiphyseal (EB) and metaphyseal borders (MB) of theprimary growth plate as shown in Figure 8F. Thus, the
Fig. 3. Histozymographic reactions produced byplacing adjacent serial sections of stage I tibial head incontact with the blackened emulsion for either 2 hr (A) or4 hr (B). After 2 hr, the reactions are approximately halfor less than half as intense as after 4 hr, as shown by thedegree of lightening produced at the blind end of thecanals (large arrows) or the metaphyseal border of theepiphysis (small arrows) versus time. Scale bar 5 500mm in A (applies to A,B).
TABLE 2. Assay of MMP Inhibitorsby Histozymographya
Inhibitor Concentration InhibitionCT1746 0.04 nM Partial
1 nM CompleteTIMP-2 0.023 mM None
0.23 mM Partial2.3 mM Complete
aMMP, matrix metalloproteinase.
75GELATINASE B AND COLLAGENASE-3, AGENTS IN COLLAGEN LYSIS

TIMP-2 bound to all of the pre-resorptive cartilagelayers and, in so doing, confirmed the presence of activegelatinase B therein.
The second section of each serial pair processed byhistozymography showed reactions in exactly the sametissue locations as the staining produced with the bio-tinylated TIMP-2 (compare Fig. 9A with 8A or Fig. 9Bwith 8D). Briefly, the tissue sites of gelatinolytic activ-ity coincide with those of TIMP-2 binding.
Immunolocalization of Collagenase-3
By using antibodies specific for collagenase-3, thelocalization of this enzyme has been examined in young
rat tibial heads. Although preimmune antibodies usedfor control yield negative results (not shown), the anti–collagenase-3 antibodies induce an immunostaining re-stricted to canal blind ends at stage I (Fig. 6E) and tosegments of the wall of the marrow space, particularlythe distal wall, at stage II (Fig. 6F). In addition, thereis weak, diffuse immunostaining throughout the carti-lage matrix of the epiphysis. When compared with sim-ilar sections immunostained for gelatinase B (Fig.6C,D), the localizations are comparable, although therestriction to the canal blind end is more definite withanti–collagenase-3 than with anti–gelatinase B anti-bodies, while the weak matrix staining is lacking withthe latter (compare Fig. 6E with C). Hence, only colla-genase-3 is distributed throughout the matrix, yet thisproteinase is concentrated at the distal cartilage wallbelonging to the canal blind end and to the distal wallof the marrow space, that is, the very sites where thecartilage is resorbed.
DISCUSSION
The development of the skeleton is the result of abalance between two opposite processes: the depositionof new bone and cartilage components in specific sitesand, in contrast, the lysis of these components at othersites. Indeed, it is the combination of these two pro-cesses that cause the small cartilage models of thefetus to gradually evolve into the long bones of theadult. As a continuation of our interest in lytic phenom-ena, we searched for activated enzymes during thetransformation of the cartilaginous epiphysis into asecondary ossification center. Histozymography re-vealed that some gelatin-cleaving proteinase waspresent during the tunneling in progress at the blindend of the canals (stage I) and during the excavation ofthe marrow space wall (stages II, III, and IV). Threeapproaches were used to identify the proteinase: his-tozymographic reactions were completely ablated bythe anti–gelatinase B neutralizing IgG; routine immu-nohistochemistry showed that gelatinase B residedwhere the activity of the enzyme was assigned by his-tozymography; and TIMP-2 histochemistry showedTIMP-2 (capable of blocking the histozymography re-actions) to bind to the very sites in the tissue wheregelatinase B was revealed by the other methods. Re-cently, a new application of neoepitope antibodies tothe localization of “activated” gelatinase B confirmedits presence in the sites revealed by histozymography
Fig. 4. Comparison of histograms prepared in the absence (A) andpresence (B) of low concentration of the MMP inhibitor CT1746. In theabsence of the inhibitor, the histozymogram shows reactions at the canalblind end and metaphyseal border (arrows in A), whereas the serialsection exposed to the inhibitor shows absent or weak reactivity in theseareas (arrows in B). (A higher concentration of CT1746 completely inhib-its the reactions; Table 2). Scale bar 5 500 mm in A (applies to A,B).
TABLE 3. Assay of Neutralizing Antibodies by Histozymographya
Target Purified IgG Amount (in mg/ml) InhibitionGelatinase A (MMP-2) Sheep anti-human MMP-2 0.5 None
1.0 None5.0 None
Gelatinase B (MMP-9) Sheep anti-pig (MMP-9) 0.5 Partial1.0 Complete
aMMP, matrix metalloproteinase; IgG, immunoglobulin G.
76 DAVOLI ET AL.

Fig. 5. Comparison of the histozymograms prepared in the absence(B,E) and presence of anti–gelatinase B immunoglobulin G (IgG) anti-bodies (C,F). At top are reference sections at stage I on the left, showingthe blind end of a canal (arrow in A) and at stage III on the right, showingthe marrow space wall, which is plain on one side (arrow 2 in D) butassociated with hypertrophic chondrocytes on the other side (arrow 1).Below are the corresponding histozymogram controls showing at stage I
reactions of canal blind end (arrow in B) and metaphyseal border, and atstage III reactions over the plain side of the marrow space wall (arrow 2in E) and the hypertrophic chondrocyte associated side (arrow 1) as wellas over the two canals and the metaphyseal border. Finally, after expo-sure to anti–gelatinase B IgG, the serial sections at stage I (C) and III (F)are unreactive. Scale bar 5 500 mm in A (applies to A–F).

Fig. 6. Tibial head sections immunostained by using either preim-mune IgG for control (A) or anti–gelatinase B IgG (B,C,D) or, for com-parison, collagenase-3 antiserum (E and F, being respectively serialsections of C and D). Although the control shows no reaction (A), theanti–gelatinase B IgG produces a reaction predominantly at the canalblind ends (at stage I), as indicated by the arrowheads in B and C and onthe wall of the forming marrow space at stage II (D), thus confirming thepresence of gelatinase B in sites to which it has been ascribed byhistozymography. Finally, the panels of the lowest row show the pres-
ence of collagenase-3 in canal blind end at stage I (E) and marrow spacewalls at stage II (F). Comparisons with gelatinase B in serial sections (Cand D, respectively) demonstrate some overlap of the two antibodies atthe base of the canals (arrows, C and E) and the distal border of themarrow space (arrow, D and F). However, the collagenase-3 is restrictedto the center of the canal blind end (E), whereas gelatinase B extends tothe nearby portions of the canal side walls (C). Scale bar 5 200 mm in A(applies to A–F).
78 DAVOLI ET AL.
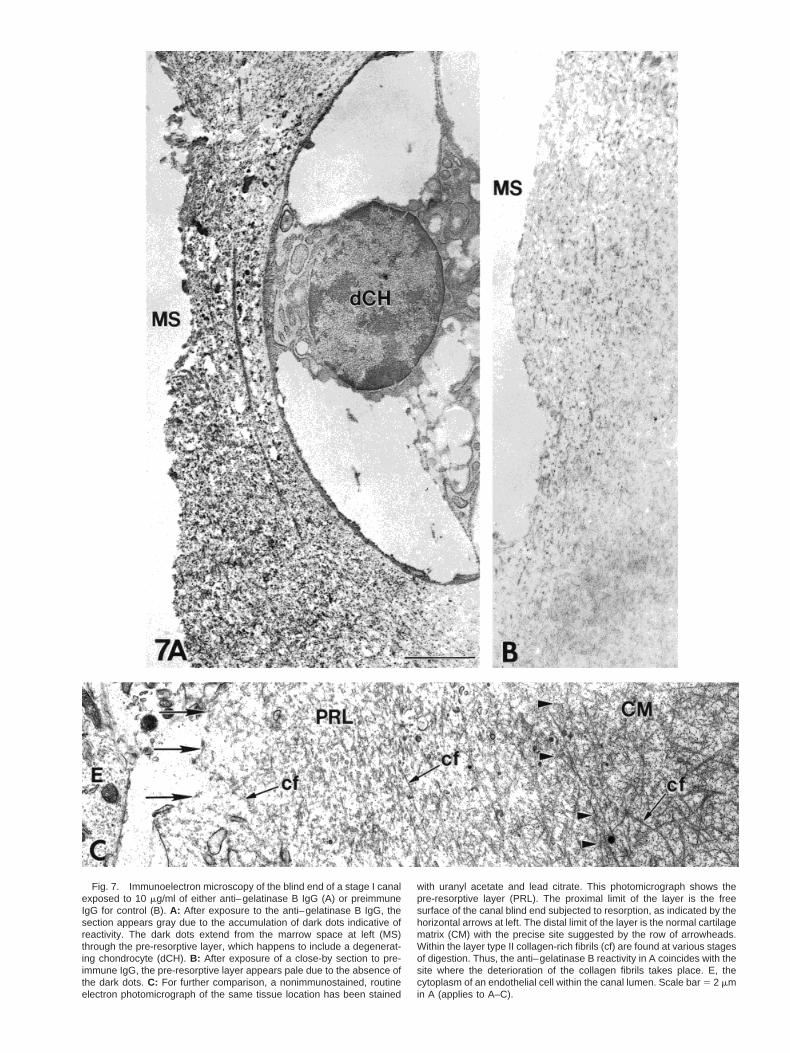
Fig. 7. Immunoelectron microscopy of the blind end of a stage I canalexposed to 10 mg/ml of either anti–gelatinase B IgG (A) or preimmuneIgG for control (B). A: After exposure to the anti–gelatinase B IgG, thesection appears gray due to the accumulation of dark dots indicative ofreactivity. The dark dots extend from the marrow space at left (MS)through the pre-resorptive layer, which happens to include a degenerat-ing chondrocyte (dCH). B: After exposure of a close-by section to pre-immune IgG, the pre-resorptive layer appears pale due to the absence ofthe dark dots. C: For further comparison, a nonimmunostained, routineelectron photomicrograph of the same tissue location has been stained
with uranyl acetate and lead citrate. This photomicrograph shows thepre-resorptive layer (PRL). The proximal limit of the layer is the freesurface of the canal blind end subjected to resorption, as indicated by thehorizontal arrows at left. The distal limit of the layer is the normal cartilagematrix (CM) with the precise site suggested by the row of arrowheads.Within the layer type II collagen-rich fibrils (cf) are found at various stagesof digestion. Thus, the anti–gelatinase B reactivity in A coincides with thesite where the deterioration of the collagen fibrils takes place. E, thecytoplasm of an endothelial cell within the canal lumen. Scale bar 5 2 mmin A (applies to A–C).

(Lee et al., 2000). Finally, the gelatinase B sites were ofthe two types defined by Lee et al. (2001, this issue):“ossification-coupled” and “free.” The prominent ossifi-
cation coupled site was the epiphysis metaphyseal bor-der which, between stages I and IV, became the growthplate metaphyseal border. A critical role for gelatinase
Figure 8.
80 DAVOLI ET AL.

B in this site was also reported by Vu et al. (1998) who,however, did not think that gelatinase B played a sig-nificant role in the development of the secondary ossi-fication center, whereas our observations showed it inabundance at “free” sites. Indeed, the various gelati-nase B sites comprised all the resorption sites definedin the epiphysis by Lee et al. (2001, this issue), includ-ing those in which aggrecan was cleaved and the one inwhich it was not, that is, the transverse cartilage par-titions of the secondary growth plate.
Localization of Collagenase-3
Immunostaining also revealed that collagenase-3was associated with gelatinase B in every pre-resorp-tive layer. The immunohistochemical approach did notindicate whether the detected collagenase-3 was func-tional, but because it was present in sites of cartilageresorption, it was possible that, like gelatinase B, itwas in an active form and, therefore, played a role inthese resorptions. Hence, the pre-resorptive layer in-cluded the two identified matrix metalloproteinases.
Collagen as a Likely Target of Gelatinase B andCollagenase-3 in the Pre-resorptive Layer
The pre-resorptive layer underlying the canal blindend was depicted by routine electron microscopy inFigure 7C. At the extreme right, normal cartilage ma-trix displayed abundant collagen fibrils, but in the restof the picture, the collagen fibrils gradually decreased
in width, length, and density to finally disintegrate asthe lumen of the canal was approached. Thus, thecompartment was the site of gradual deterioration ofcollagen fibrils culminating in their digestion. Thepresence of both gelatinase B and collagenase-3 in thepre-resorptive layer suggested that these enzymeswere responsible for the demise of the collagen fibrils.What was known of the collagenolytic potential of thesetwo enzymes will now be examined.
Mechanism of collagen lysis.Of the various com-ponents of the collagen fibrils present in cartilage, he-lical type XI collagen was known to be degraded bygelatinase B (Murphy et al., 1981; Pourmotabbed et al.,1994; Niyibizi et al., 1994). As for the more abundanttype II collagen (approximately 80% of total), gelati-
Fig. 9. Histozymographic reactions produced by sections from stageI (A) and III (B) tibial heads neighboring the sections exposed to biotin-ylated TIMP-2 depicted in Figure 8A and D, respectively). The histozy-mograms reveal a lightening of the emulsion at the same sites as thosestained with the biotinylated TIMP-2 (compare the reactions indicatedwith large and small arrows in Fig. 8A and 9A at stage I, and in Figs. 8Dand 9B at stage III). Thus, TIMP-2 binds at the sites that contain activegelatinase B. Scale bar 5 500 mm in A (applies to A,B).
Fig. 8. Outcomes of the treatments of unfixed tissue sections fromtibial heads at stages I, III, and IV with mutant noninhibitory TIMP-1 forcontrol (C,G) or normal biotinylated TIMP-2 (A,B,D,E,F). Reactivity isvisualized with avidin/biotin horseradish peroxidase, then 3,39-diamino-benzidine (DAB). In A, the section exhibits a large central canal (arrow),which is enlarged in B to reveal a brown reactivity, assigned to thepre-resorptive layer underlying the surface of the canal blind end. C: Afterexposure of a serial section to mutant TIMP-1, the pre-resorptive layerappears pale due to a low insignificant brown reaction. D: A stage III tibialhead, the marrow space has formed through the merger of two canals;the one at left is fully in view, the one at right, is only partially visible.E: The left canal and adjoining marrow space, are enlarged, where abrown reaction is observed along plain segments of the wall of themarrow space (MS) described as sites of “free” cartilage resorption (largearrows) and a weaker reaction on groups of hypertrophic chondrocytesassociated with “ossification-coupled” resorption (small arrows). G: Aneighboring section treated with mutant TIMP-1 lacks the reactivity re-vealed in E. F: In a stage IV tibial head, the primary (1°GP) and second-ary (2°GP) growth plates are delineated and spots of reactivity, moreintense than those found at stage III (D), are revealed in the formingsecondary ossification center (small arrows). Moreover, the epiphysealborder (EB) of the primary growth plate (mostly a “free” mode of resorp-tion) is intensely reactive. Furthermore, some reactivity is also observedat the bases of two emerging canals (arrows, upper left). Finally, in A, D,and F, the small arrows at the base of the photomicrographs identify abrown colored reaction line found along the metaphyseal border locatedbetween epiphysis and metaphysis. At stage IV (F), other reactivity alsoappears along the lateral (Lat B) and medial (Med B) borders of theprimary growth plate where lysis was detected previously (Lee et al.,1998). The overall interpretation is that the TIMP-2 horseradish peroxi-dase DAB reactions coincide with the sites of cartilage lysis. Scale bars 5500 mm in A (applies to A,D,F), 200 mm in B (applies to B,C,E,G).
81GELATINASE B AND COLLAGENASE-3, AGENTS IN COLLAGEN LYSIS

nase B was known to attack its C-telopeptide domain(Eyre, 2000; unpublished communication), but not tocleave its triple helix (Murphy et al., 1982). Cleavagesof the helix could only be achieved by collagenases,such as collagenase-3 (MMP-13), the interstitial colla-genase (MMP-1), and neutrophil collagenase (MMP-8),which all cleaved type II collagen in vitro at the G775-L776 bond, yielding two fragments whose length wasthree-quarters and one-quarter that of the whole mol-ecule (Miller et al., 1976; Hasty et al., 1990; Mitchell etal., 1996). Thereafter, the fragments were known to besusceptible to degradation by gelatinases (Harris andKrane, 1972; Murphy et al., 1982). In the young rattibial head, collagenase-3 was detected in every sitewhere gelatinase B was present and, even though itsactivity had not been demonstrated, its associationwith gelatinase B in resorption sites made it likely thatthe two enzymes functioned in tandem to break downthe collagen. Engsig et al. (2000) demonstrated thatgelatinase B had a weak, and collagenase-3 a strong,ability to lyse type II collagen in vitro, and that in thepresence of both enzymes, collagenolytic activity morethan doubled that achieved by collagenase-3 alone.Hence, gelatinase B potentiated collagenase-3 activity.
However, because other components were presentwithin the matrix under attack, the question naturallyarose as to whether or not gelatinase B or collagenase-3might also lyse the aggrecan core protein. Althoughgelatinase B was not efficient at cleaving either iso-lated aggrecan or aggrecan aggregates in vitro (Mur-phy et al., 1982, 1991a), collagenase-3 could not bediscounted as a candidate for this lysis. Finally, othercomponents were present in minor amounts, but theirsusceptibility to either gelatinase B or collagenase-3was not known (Handler et al., 1997; Gerber et al.,1999; Aszodi et al., 2000; Carlevaro et al., 2000; Engsiget al., 2000).
Behavior of collagen and aggrecan in the pre-resorptive layer.The events taking place in the pre-resorptive layer were summarized in Figure 10. Be-cause there was a progressive deterioration of collagenfibrils throughout the layer (Fig. 7C), the amount ofthese fibrils, which was considered 100% in normalcartilage, was assumed to decrease regularly to 0%from the distal to the proximal limit, presumably underthe joint action of the enzymes collagenase-3 and gela-tinase B detected in the area. As for the aggrecan coreprotein, its amount was assigned to 100% in normalcartilage, but was believed to drop to none at the distallimit, as a result of cleavage at this site (Lee et al.,2001, this issue). Finally, this cleavage yielded G1-341fragments, whose amount was thought to remain es-sentially unchanged from the time they arose at thedistal limit to the time they were resorbed at the prox-imal limit.
Thus, the pre-resorptive layer was a dynamic entity.Moreover, that the lysis of either collagen or aggrecanwas observed whenever looked for by the availablemethods suggested that the lysis of both substances
was taking place continuously. This conclusion sug-gested the existence of a steady state not only for col-lagen and aggrecan, but also for the whole compart-ment, that is, for instance during the digging of thecanals, the loss of substance by resorption at the prox-imal limit should be balanced by expansion of the distallimit into normal cartilage. Lee et al. (2001, this issue)had estimated, for example, that the turnover time ofthe compartment was 42 min at the canal blind end.Hence, the events occurring at the distal limit of thelayer preceded those occurring at the proximal limit by42 min and, therefore, Figure 10 represented eventsdeveloping in time from right to left. Accordingly, theG1-341 fragments produced at the distal limit spent 42min in the layer before being lost at the proximal limit.Finally, the time taken for the degradation of collagenfibrils from normal to complete disintegration shouldalso be of the order of 42 min.
Sequence of Lytic Events in the CartilageResorption Required for the Formation of theSecondary Ossification Center
Lee et al. (1999) have previously proposed that car-tilage resorptions take place in three main steps,namely, aggrecan lysis, collagen II cleavage by collage-nase-3, collagen fragment lysis by gelatinase B. Al-though the present results and those of Lee et al. (2001,this issue) essentially support this sequence, findingsthat have recently come to light suggest modifications.Our tentative proposal for the role of the three involvedenzymes, i.e., the unidentified MMP, gelatinase B, andcollagenase-3, in the lysis of the pre-resorptive layer isas follows.
(1) It is first proposed that these enzymes originate inthe cells facing the surface about to be resorbed andthen migrate from there to their site of actions.Besides recent evidence pointing to the origin ofgelatinase B and collagenase-3 in canal lumen cells(Lee E.R., unpublished data), indirect support forthis view has been derived from what is known ofthe enzyme MT1-MMP. This enzyme, which ispresent at the surface of canal cells (Lee E.R., Mc-Quibban G.A., and Overall C.M., unpublished)where it is presumably anchored in the plasmamembrane (Sato et al., 1997), has been investigatedby knocking-out the controlling gene, in which casethe formation of canals in the epiphysis is pre-vented (Holmbeck et al., 1999; Zhou et al., 2000).The likely mechanism of this prevention is that thecanals cell lack MT1-MMP—a potent activator ofother MMPs (for review, see Murphy and Knauper,1997). As a result, the three enzymes involved incartilage resorption would not be activated, andcanals would not form. It is tentatively concludedthat the first step in the cartilage resorption se-quence was the production of the unidentifiedMMP, gelatinase B, and collagenase-3 within thecells facing the resorbed areas, followed by the
82 DAVOLI ET AL.

MT1-MMP activation of these enzymes and theirmigration to the site of action.
(2) Because, as indicated in Figure 10, aggrecan lysisprecedes collagen lysis, the next step is the cleav-age of aggrecan core protein by a so far unidentifiedMMP, as shown by the presence of G1 fragmentsterminating in the amino acid residues . . .FV-DIPEN in the walls and blind end of the canals, thewalls of the marrow space, and the metaphysealborder (Lee et al., 2001, this issue). In addition,aggrecanase participates in the cleavage of the ag-grecan on the epiphyseal border of the primarygrowth plate.
(3) The third step in the overall lysis is the initiation ofcollagen fibril digestion by a collagenase. The evi-dence indicates that gelatinase B only attacks he-lical type II collagen after it has been cleaved bycollagenase (Murphy et al., 1982). We have de-tected collagenase-3 in the resorption sites, al-though the activity of the collagenase has not beendemonstrated so far. We nevertheless postulate the
cleavage of type II collagen by collagenase-3 as thethird step in the sequence.
(4) The fourth step is the completion of collagen fibrildigestion by gelatinase B. The role of this enzyme isthe degradation of the type II fragments producedby collagenase (for review see, Murphy and Crabbe,1995). In addition, the type XI collagen present incartilage can be directly attacked by gelatinase B(Murphy et al., 1981, 1982; Pourmotabbed et al.,1994; Niyibizi et al., 1994). Moreover, digestionmay be enhanced by the joint action of collagenasesand gelatinases, which together have been shownto facilitate the collagenolytic process (Harris andKrane, 1972; Murphy et al., 1982; Engsig et al.,2000).
(5) What is the final step in the resorption of the wallpresent at the proximal limit of the compartment?A priori, the demise of collagen would seem to besufficient to insure the collapse of the hyaluronate-attached G1341 fragments and other cartilage rem-nants making up the wall. This does not seem to be
Fig. 10. Semigraphic depiction of the lytic events occurring within thepre-resorptive layer, within which the gelatinase B is located. The X-axisdenotes the position of the proximal limit of the pre-resorptive layer on theleft and the position of the distal limit on the right. The Y-axis indicateshow the percentage of the substances present varies with the location.The distal limit marks the transition from the pre-resorptive layer into thenormal cartilage where the content of collagen and aggrecan is complete(100%). At the junction between the distal limit and the normal cartilage,aggrecan core protein is cleaved at the Asn341-Phe342 (Lee et al., 2001),as indicated by the arrow. Thus, the solid black line depicts a rapid
decline in overall aggrecan content. So, although the G1-341 contentincreases rapidly to 100% in the pre-resorptive layer, the bulk of theaggrecan molecule, that is the G2-G3 fragments containing the attachedglycosaminoglycan chains, have essentially been lost. In contrast, thedotted line depicts a steady loss of fibrils beginning at the distal limit of thelayer and continuing throughout the pre-resorptive layer until completiontoward the free surface undergoing resorption (the proximal limit). Thus,in contrast to aggrecan cleavage, the lysis of the fibrils is perceived totake place in all parts of the pre-resorptive layer.
83GELATINASE B AND COLLAGENASE-3, AGENTS IN COLLAGEN LYSIS

the case, however, because examination of theproximal limit wall in the electron microscope (asseen in Fig. 5C,D of Lee et al., 2001, this issue)indicates that the cytoplasmic expansions from in-tracanal cells are seen breaking through the wall.Presumably, enzymes on the cell surface, perhapsMT1-MMP attached to the cell membrane, makepossible the final cell invasion, which culminates inthe complete resorption of the proximal limit of thepre-resorptive layer.
EXPERIMENTAL PROCEDURESAnimals and Preparation of Tissue
Male Sprague-Dawley rats, from birth to 12 days ofage, housed and handled according to the recommen-dations made by the Canadian Council on AnimalCare, have been anesthetized with sodium pentobarbi-tal (Somnotol, MTC Pharmaceuticals, Cambridge, On-tario; I.P., 50 mg/kg) before proximal tibiae were har-vested and prepared for gelatin histozymography,peroxidase immunohistochemistry, or TIMP histo-chemistry.
Gelatin Histozymography
The first step is the preparation of blackened emul-sion slides in the dark room, when clean glass slidesare dipped in NTB2 radioautographic emulsion (East-man Kodak Company, Rochester, NY) and, after dry-ing, are exposed to room light for 5 min, developed inKodak D-170 for 7 min, rinsed, and fixed in 24% so-dium thiosulfate for 10 min. They are then stored at4°C until use. For histozymography, 20-mm-thick sec-tions of unfixed tibiae are cut in a Bright cryostat andcollected on tape. A 3-ml drop of PBS is placed on thetape-mounted sections, which are then inverted andplaced in contact with a blackened emulsion slide. Theslide emulsion complex is incubated for 4 hr at roomtemperature in a sealed humidified Petri dish. Afterpeeling the section off the emulsion, it is stained in 1%aqueous toluidine blue, and both section and emulsionare then photographed (see Lee et al., 1999, for furtherdetails).
Inhibitors. To identify the proteinases responsiblefor the emulsion digestion, inhibitors are applied onsections destined to histozymography. A 3-ml volume ofeach solution at the concentration indicated in Table 2is added to a section before it is placed on the emulsion.For control, the same volume but lacking the inhibitor,is added to a neighbouring section before applying it tothe emulsion. The results reported in Tables 2 and 3are the outcome of six trials per inhibitor tested. Theeffects of the inhibitors used to characterize proteinaseactivity are summarized in Table 2. The recombinanthuman TIMP-2 has been expressed in NSO myelomacells and purified as described previously (Murphy etal., 1991b; Willenbrock et al., 1993). The concentration-dependant gelatinase hydroxysuccinamide, CT1746,has also been described previously (Anderson et al.,1996). The neutralizing immunoglobulins, which have
been purified from anti-human gelatinase A (MMP-2)and anti-pig gelatinase B (MMP-9) sheep antisera,have been described before (Murphy et al., 1989; Hippset al., 1991, Lelongt et al., 1997). Cross-reactivity ofthese sheep IgGs with rat gelatinases has been shownpreviously by Lee et al. (1999). Although the specificityof the IgGs is an integral part of targeting the appro-priate proteinase, their ability to bind and inactivate,that is neutralize, the mature (active) proteinase hasalso been shown by Lelongt et al. (1997). Thus, theseIgGs are used to neutralize enzymatic activity of gela-tinase A and gelatinase B, respectively.
Peroxidase Immunohistochemistry
Tibial sections processed as described below are pre-pared for staining with sheep anti-pig gelatinase B IgGor rabbit anti-rat collagenase-3 antiserum. The latteris an anti-peptide antiserum prepared according to themethod described in the preceding article (Lee et al.,2001, this issue). Briefly, three peptides have beensynthesized corresponding to the residues 127-136(CGSEVEKAFRKA), 267-278 (DPNPKHPKTPEKC),and 369-379 (CGFPKEVKRLSA) found on rat collage-nase-3; each includes a G and C residue added to the Nor the C-terminus (Quinn et al., 1990). The peptide iscoupled to ovalbumin through the terminal cysteineresidue, and a mixture of the three conjugated peptideshas been injected into a New Zealand White rabbit.Antiserum specificity has been tested against a par-tially activated recombinant human collagenase-3 pre-pared in transfected NSO mouse myeloma cells (Knau-per et al., 1996). Reactivity of the antiserum with therecombinant proteinase is demonstrated by standardsodium dodecyl sulfate-polyacrylamide gel electro-phoresis (SDS-PAGE) and immunoblotting techniquesin Figure 11.
Immunostaining for Light Microscopy
Proximal tibial epiphyses are immersed in a perio-date-lysine-paraformaldehyde fixative (including 2%formaldehyde; McLean and Nakane, 1974) and, then,placed at 4°C overnight. After rinsing in azide-freephosphate buffered saline (PBS), the tibial epiphysesare split in half along the sagittal plane and decalcifiedin 10% EDTA in 0.1 M Tris, pH 7.4, at 4°C. The tissueis infiltrated first with 20% sucrose in PBS and then intwo parts of this solution and one part of OCT com-pound (VWR-Canlab, Ville Mont-Royal, Quebec). Em-bedding is done in this final mixture by using standardspecimen cryomolds (Miles, Elkhart, IN), and tissue isfrozen within the block over dry ice. Eight-micrometersections are cut in a Bright’s OTF cryostat, collected ongelatin-coated slides, and stored at 220°C.
For immunolocalization of gelatinase B with thesheep anti-pig gelatinase B immunoglobulin, the fixed,slide-mounted sections are taken to room temperature,immersed in 4% formaldehyde, and rinsed in PBS be-fore a 1 hr digestion at 37°C with 0.25 U/ml of chon-droitinase ABC (Proteus vulgaris; ICN Biomedical,
84 DAVOLI ET AL.

Costa Mesa, CA) in the presence of proteinase inhibi-tors. Thereafter, the slide-mounted sections are ex-posed to 0.3% H2O2 (v/v in methanol) to quench endog-enous peroxidase reactivity before blocking in 1.5%normal goat serum in PBS for 20 min and incubationwith anti–gelatinase B immunoglobulins (IgGs) for 30min. After two PBS washes, the bound IgG is localizedby using the Vectastain Elite ABC kit (Vector Labora-tories, Burlingame, CA). Briefly, the tissue-bound IgGrecognizes an avidin-biotin-horseradish peroxidasecomplex, which is detected by a 5-min exposure toDAB, which, over the areas containing gelatinase B,yields a precipitate that appears brown in the lightmicroscope. After rinsing, the sections are dried andcounterstained with Gill’s hematoxylin for 1 min on ahot plate, rinsed in three changes of distilled water,and mounted under a coverslip.
Immunoelectron Microscopy
The procedure is similar to the one described for lightmicroscopy with several differences. First, the sectionsare cut at a 25 mm thickness. Second, after immuno-staining through the same steps as for light micros-copy, the DAB-exposed sections are stored overnight inPBS at 4°C and, the next morning, are post-fixed by
using a droplet of 1% osmium tetroxide in 0.1 M sodiumcacodylate buffer. The sections are then dehydrated inincreasing gradients of acetone and then acetone-Eponup to pure Epon, a drop of which is finally placed on thesection to be cured overnight at 60°C. A polymerizedEpon block is then inverted onto the section, and thetwo are held together by a fresh drop of resin. While thetwo are held together, they are cured overnight at60°C. The slide is then immersed in liquid nitrogen,causing the block to contract and snap away from it.Thin sections are prepared and examined in a Philips400 EM in the absence of counterstaining.
Routine Electron Microscopy
Tibiae are dissected from rats that have been per-fused through the heart with a mixture containing2.5% glutaraldehyde (highly purified, J.B. EM Servic-es; Pointe Claire, Quebec) and 2.0% formaldehyde(freshly prepared from paraformaldehyde), both in 0.1M cacodylate buffer (pH 7.3). All samples are post-fixedin potassium ferrocyanide–reduced osmium tetroxide,dehydrated in acetone, and embedded in JEMbed ep-oxy resin before examination in a Philips 400 ElectronMicroscope at 80 kV.
TIMP HistochemistryBiotinylation of the TIMPs. Although recombi-
nant human TIMP-2 is used as such in inhibition tests(Table 2) as described before, a biotinylated form isused to localize its binding sites in epiphyseal sectionsand an inactive biotinylated recombinant humanTIMP-1 is used for control. Biotinylation of the twoTIMPs has been achieved with EZ Link Sulfo-NHS-LC-Biotin, according to the directions of the manufacturer(Pierce Chemical Co., Rockford, IL). Briefly, the recom-binant TIMPs (0.2 mg/ml in 20 mM Tris-HCl, pH 7.6)are dialyzed overnight at 4°C against a 50-mM sodiumbicarbonate buffer (pH 8.5), before EZ Link Sulfo-NHS-LC biotin is added at a 4 M excess. The reactionis allowed to continue for 3 hr at room temperatureafter which the biotinylated TIMPs are dialyzed intoazide-free PBS and stored at 220°C until needed. Toconfirm that the proteins are adequately labeled withbiotin, they have been exposed to standard SDS-PAGEand blotting analysis as shown in Figure 12. For theinactivation of TIMP-1 (which has been expressed inPichia pastoris either as glycosylated or nonglycosy-lated form), both forms have been rendered inactive bythe addition of a four–amino acid residue extension atthe N-terminus. To test whether or not the biotinylatedTIMP-2 still retains its ability to inhibit active enzymeafter biotinylation, the biotinylated TIMP-2 has beenexamined in a gelatinase A reverse zymogram. Therecombinant human gelatinase A is mixed with porcinegelatin and both are polymerized into the gel as de-scribed by Oliver et al. (1997). EDTA fully prevents thedigestion of the gelatin by the enzyme (Fig. 13A), asdoes the biotinylated TIMP-2 (Fig. 13B), showing that
Fig. 11. Immunoblot of recombinant human collagenase-3 immuno-stained with anti–collagenase-3 antiserum (A) or preimmune serum (B).The recombinant proteinase (0.5 mg per lane) was electrophoresed on areduced 8% sodium dodecyl sulfate-polyacrylamide gel electrophoresisgel before transfer to nitrocellulose paper. The immunoblot in lane Areveals reactivity of the antiserum with the 60-kDa proenzyme form andthe processed 48-kDa active form (Knauper et al., 1996). No reactionsare observed in lane B stained with the preimmune serum. Reactivity wasvisualized by exposing the membrane to alkaline phosphatase conju-gated anti-rabbit immunoglobulin (Promega, Madison, WI) and to Ni-troBlue Tetrazolium and 5-bromo-4-chloro-3-indolyl phosphate (Pro-mega). The position of two molecular weight (MW) standards is indicatedat left.
85GELATINASE B AND COLLAGENASE-3, AGENTS IN COLLAGEN LYSIS

the biotinylated form of the inhibitor has retained thecapacity to inhibit an active MMP.
TIMP peroxidase histochemistry. The TIMPshave been applied to frozen tape-mounted sections pre-
pared in the same way as described under histozymog-raphy. Obviously, when MMPs are active, they canbind TIMP and be blocked by it. Hence, it is likely thatthe TIMPs will be bound to histologic sections at sitescontaining MMPs. Therefore, to visualize active MMPsin the sections, TIMPs are applied as a 3-ml droplet (40mg/ml in azide-free PBS), which is rapidly spread bycover-slipping, before the slide-section complex istransferred to a humidified chamber for 1 hr at roomtemperature. The complex is then disassembled and, towash away the unbound TIMP, the section is trans-ferred (tissue side down) successively on the surface offour wells filled with azide-free PBS. Bound TIMP isrevealed by exposing the section for 30 min to a pre-formed avidin-biotin horseradish peroxidase complexmade according to the directions of the manufacturer(Vector Laboratories, Burlington, Ontario). After wash-ing away all unbound ABC complex, a reddish brownstain is produced at reactive sites by exposing the sec-tion for 2.5 min to the chromogen 3,39-diaminobenzi-dine (DAB) in the presence of H2O2. The tape-attachedsection is then rinsed in distilled water and wetmounted on a glass slide with coverslip by using asaturated aqueous solution of polyvinyl pyrrolidone(Sigma) and photographed with a Zeiss Axiovert-35inverted microscope.
ACKNOWLEDGMENTS
The authors thank Dr. Mohamed El-Alfy, currentlyat the Medical Research Group in Molecular Endocri-nology, CHUL Research Centre, Laval, Quebec, forteaching M.A. Davoli the technique of histozymogra-phy. We also thank Dr. Anneliese Recklies (Joint Dis-eases Laboratory, Shriners Hospital for Children) for
Fig. 12. Blot of active TIMP-2 in (A) (lanes 1 and 2) and inactiveTIMP-1 in (B) (lanes 3–6). TIMP-1 is the glycosylated form in lanes 3 and5 and the nonglycosylated form in lanes 4 and 6. The TIMP in lanes 2, 5,and 6 was reacted with sulfosuccinimidobiotin before electrophoresiswhereas the TIMP in lanes 1, 3, and 4 was not. The samples, electro-phoresed on reduced sodium dodecyl sulfate-polyacrylamide gel electro-
phoresis gels before transfer to nitrocellulose paper, were then stainedwith streptavidin-alkaline phosphatase and visualized by exposing themembrane to NitroBlue Tetazolium and 5-bromo-4-chloro-3-indolyl phos-phate (Promega). All TIMPS treated with sulfosuccinimidobiotin arestained. Molecular weight (MW) standards are indicated to the right ofeach panel.
Fig. 13. Reverse zymograms in which biotinylated TIMP-2 has beenelectrophoresed in a 12% sodium dodecyl sulfate-polyacrylamide gelelectrophoresis (SDS-PAGE) containing porcine gelatin (2.25 mg/ml) andrecombinant human gelatinase A (640 ng/ml). Thereafter, the gel hasbeen incubated in Triton X-100 buffer either in the presence (A) orabsence of EDTA (B). After the incubation is completed, the gels havebeen stained with Coomassie blue (G-250) to reveal the digestion of thegelatin by gelatinase A, which is indicated by a lightening of the gel. Whengelatinase A is inhibited and unable to degrade the gelatin (as occurs inthe presence of EDTA), the gel is darkly stained with Coomassie blue.Note the increased inhibition of gelatinase A (indicated by arrow) as theconcentration of the biotinylated TIMP-2 is increased in the lanes from leftto right (200 pg, 830 pg, 3 ng, 7 ng, and 13 ng, respectively). Therefore,it is concluded that the biotinylated TIMP-2 has retained its inhibitorproperties after biotinylation.
86 DAVOLI ET AL.

her suggestion to use recombinant TIMP to localizeactive MMPs in tissue prepared for histology. The pep-tides in this study were made by Ms. Elisa de Miguel inthe Core Biotechnology Facility of the Shriners Hospi-tal, and the figures and art work were prepared by Mr.Mark Lepik and Ms. Guylaine Bedard. Dr. GillianMurphy is a Senior Fellow of the Arthritis ResearchCampaign, U.K.
REFERENCES
Adams CWM, Tuqan NA. 1961. The histochemical demonstration ofprotease by a gelatin-silver film substrate. Nature 189:469–472.
Anderson IC, Shipp M, Docherty AJP, Teicher BA. 1996. Combinationtherapy including a gelatinase inhibitor and cytotoxic agent reduceslocal invasion and metastasis of murine Lewis lung carcinoma.Cancer Res 56:715–718.
Aszodi A, Bateman JF, Gustafsson E, Boot-Handford R, Fassler R.2000. Mammalian skeletogenesis and extracellular matrix: whatcan we learn from knockout mice? Cell Struct Funct 25:73–84.
Butler G, Hutton M, Wattam BA, Williamson RA, Knauper V, Wil-lenbrock F, Murphy G. 1999. The specificity of TIMP-2 for matrixmetalloproteinases can be modified by single amino acid mutations.J Biol Chem 274:20391–20396.
Carlevaro MF, Cermelli S, Cancedda R, Cancedda FD. 2000. Vascularendothelial growth factor (VEGF) in cartilage neovascularizationand chondrocyte differentiation: auto-paracrine role during endo-chondral bone formation. J Cell Sci 113:59–69.
Cole AA, Cole MB Jr. 1989. Are perivascular cells in cartilage canalschondrocytes? J Anat 165:1–8
Cole AA, Wezeman FH. 1985. Perivascular cells in cartilage canals ofthe developing mouse epiphysis. Am J Anat 174:119–129
Cole AA, Wezeman FH. 1987a. Cytochemical localization of tartrate-resistant acid phosphatase, alkaline phosphatase, and nonspecificesterase in perivascular cells of cartilage canals in the developingmouse epiphysis. Am J Anat 180:237–242
Cole AA, Wezeman FH. 1987b. Morphometric analysis of cartilagecanals in the developing mouse epiphysis. Acta Anat 128:93–97
Daoust R. 1965. Histochemical localization of enzyme activities bysubstrate film methods: ribonucleases, deoxyribonucleases, pro-teases, amylase and hyaluronidase. Int Rev Cytol 18:191–221.
Delgado-Baeza E, Gimenez-Ribotta M, Miralles-Flores C, Nieto-Chaguaceda A, Santos-Alvarez I. 1991a. Morphogenesis of cartilagecanals: experimental approach in the rat tibia. Acta Anat 142:132–137.
Delgado-Baeza E, Gimenez-Ribotta M, Miralles-Flores C, Nieto-Chaguaceda A, Santos-Alvarez I. 1991b. Relationship between thecartilage canal and the perichondrium in the rat proximal tibialepiphysis. Acta Anat 141:31–35.
Engsig MT, Chen Q-J, Vu TH, Pedersen A-C, Therkidsen B, Lund LR,Henriksen K, Lenhard T, Foged NT, Werb Z, Delaisse J-M. 2000.Matrix metalloproteinase 9 and vascular endothelial growth factorare essential for osteoclast recruitment into developing long bones.J Cell Biol 151:879–889.
Eyre DR, Wu J-J. 1995. Collagen structure and cartilage matrixintegrity. J Rheumatol (Suppl) 21:82–85.
Gerber HP, Vu TH, Ryan AM, Kowalski J, Werb Z, Ferrara N. 1999.VEGF couples hypertrophic cartilage remodeling, ossification andangiogenesis during endochondral bone formation. Nat Med 5:623–628.
Handler M, Yurchenco PD, Iozzo RV. 1997. Developmental expressionof perlecan during murine embryogenesis. Dev Dyn 210:130–145.
Harris ED, Krane SD. 1972. An endopeptidase from rheumatoid sy-novial tissue culture. Biochim Biophys Acta 258:566–576.
Hasty KA , Pourmotabbed TF, Goldberg GI, Thompson JP, SpinellaDG, Stevens RM, Mainardi CL. 1990. Human neutrophil collage-nase: a distinct gene product with homology to other matrix metal-loproteinases. J Biol Chem 265:11421–11424.
Hipps DS, Hembry RM, Docherty AJ, Reynolds JJ, Murphy G. 1991.Purification and characterization of human 72kDa gelatinase (typeIV collagenase). Biol Chem Hoppe-Seyler 372:287–296.
Holmbeck K, Bianco P, Caterina J, Yamada S, Kromer M, KuznetsovSA, Mankani M, Robey PG, Poole AR, Pidoux I, Ward JM, Birkedal-Hansen H. 1999. MT1-MMP-deficient mice develop dwarfism, os-teopenia, arthritis, and connective tissue disease due to inadequatecollagen turnover. Cell 99:81–92.
Knauper V, Lopez-Otin C, Smith B, Knight G, Murphy G. 1996.Biochemical characterization of human collagenase-3. J Biol Chem271:1544–1550.
Lee ER, Lamplugh L, Leblond CP, Mordier S, Magny M-C, Mort JS.1998. Immunolocalization of the cleavage of the aggrecan core pro-tein at the Asn341-Phe342 bond, as an indicator of the location of themetalloproteinases active in the lysis of the rat growth plate. AnatRec 252:117–132.
Lee ER, Murphy G, El-Alfy M, Davoli MA, Lamplugh L, Docherty AJ,Leblond CP. 1999. Active gelatinase B is identified by histozymog-raphy in the cartilage resorption sites of developing long bones. DevDyn 215:190–205.
Lee ER, Lamplugh L, Mort JS, Murphy G, Leblond CP. 2000. Neo-epitope antibodies and immunolocalization techniques reveal thatactive gelatinase B is involved in the lysis required for the devel-opment of the rat long bone epiphysis. J Bone Miner Res 15:S349.
Lee ER, Lamplugh L, Davoli MA, Beauchemin, Chan K, Mort JS,Leblond CP. 2001. Enzymes active in the areas undergoing carti-lage resorption during development of the secondary ossificationcenter in the tibiae of rats aged 0–21 days: I. Two groups of pro-teinases cleave the core protein of aggrecan. Dev Dyn (this issue).
Lelongt B, Trugnan G, Murphy G, Ronco PM. 1997. Matrix metallo-proteinases MMP2 and MMP9 are produced in early stages ofkidney morphogenesis but only MMP9 is required for renal orga-nogenesis in vitro. J Cell Biol 136:1363–1373.
Levene C. 1964. The patterns of cartilage canals. J Anat 98:515–538.McLean I, Nakane PI. 1974. Periodate-lysine-paraformaldehyde fix-
ative. A new fixative for immunoelectron microscopy. J HistochemCytochem 22:1077–1083.
Miller EJ, Harris EC, Chung E, Finch JE, McCroskery PA, ButlerWT. 1976. Cleavage of type II and III collagen with mammaliancollagenase: site of cleavage and primary structure at the NH2-terminal portion of the smaller fragment released from both colla-gens. Biochemistry 15:787–792.
Mitchell PG, Magna HA, Reeves LM Lopresti-Morrow LL, Yocum SA,Rosner FJ, Geoghegan KF, Hambor JE. 1996. Cloning, expression,and type II collagenolytic activity of matrix metalloproteinase-13from human osteoarthritic cartilage. J Clin Invest 97:761–768.
Murphy G, Crabbe T. 1995. Gelatinases A and B. Methods Enzymol248:470–484.
Murphy G, Knauper V. 1997. Relating matrix metalloproteinasestructure to function: Why the “Hemopexin” domain? Matrix Biol15:511–518.
Murphy G, Cawston TE, Galloway WA, Barnes MJ, Bunning RAD,Mercer E, Reynolds JJ, Burgeson RE. 1981. Metalloproteinasesfrom rabbit bone culture medium degrades types IV and V collag-ens, laminin, and fibronectin. Biochem J 199:807–811.
Murphy G, Reynolds JJ, Bretz U, Baggiolini M. 1982. Partial purifi-cation of collagenase and gelatinase from human polymorphonu-clear leucocytes. Analysis of their actions on soluble and insolublecollagens. Biochem J 203:209–221.
Murphy G, Ward R, Hembry RM, Reynolds JJ, Kuhn K, TryggvasonK. 1989. Characterization of gelatinase from pig polymorphonu-clear leucocytes. Biochem J 258:463–472.
Murphy G, Cockett MI, Ward RV, Docherty JP. 1991a. Matrix met-alloproteinase degradation of elastin, type IV collagen and proteo-glycan. Biochem J 277:277–279.
Murphy G, Houbrechts A, Cockett MI, Williamson RA, O’Shea M,Docherty AJP. 1991b. The N-terminal domain of tissue inhibitor ofmetalloproteinases retains metalloproteinase inhibitory activity.Biochemistry 30:8097–8102.
87GELATINASE B AND COLLAGENASE-3, AGENTS IN COLLAGEN LYSIS

Niyibizi C, Chan R, Wu J-J, Eyre D. 1994. A 92 kDa gelatinase(MMP-9) cleavage site in native type V collagen. Biochem BiophysRes Commun 202:328–333.
Oliver GW, Leferson JD, Stetler-Stevenson WG, Kleiner DE. 1997.Quantitative reverse zymography: analysis of picogram amounts ofmetalloproteinase inhibitors using gelatinase A and B reverse zy-mograms. Anal Biochem 244:161–166.
Pourmotabbed T, Solomon TL, Hasty KA, Mainaardi CL. 1994.Characteristics of 92 kDa type IV collagenase/gelatinase pro-duced by granulocytic leukemia cells: structure, expression ofcDNA in E. coli and enzymatic properties. Biochim Biophys Acta1204:97–107.
Quinn CO, Scott DK, Brinckerhoff CE, Matrisian LM, Jeffrey JJ,Partridge NC. 1990. Rat collagenase. Cloning, amino acid sequencecomparison, and parathyroid hormone regulation in osteoblasticcells. J Biol Chem 265:22342–22347.
Sato H, Okada Y, Seiki M. 1997. Membrane-type matrix metallopro-teinases (MT-MMPs) in cell invasion. Thromb Haemost 78:497–500.
Vu TH, Shipley JM, Bergers G, Berger JE, Helms JA, Hanahan D,Shapiro SD, Senior RM, Werb Z. 1998. MMP-9/gelatinase B is a keyregulator of growth plate angiogenesis and apoptosis of hypertro-phic chondrocytes. Cell 93:411–422.
Willenbrock F, Crabbe T, Slocombe PM, Sutton CW, Docherty AJP,Cockett MI, O’Shea M, Brocklehurst K, Phillips IR, Murphy G.1993. The activity of the tissue inhibitors of metalloproteinasesis regulated by C-terminal domain interactions: a kineticanalysis of the inhibition of gelatinase A. Biochemistry 32:4330 –4337.
Zhou Z, Apte SS, Soininen R, Cao R, Baaklini GY, Rauser RW, WangJ, Yihai C, Tryggvason K. 2000. Impaired endochondral ossificationand angiogenesis in mice deficient in membrane-type matrix met-alloproteinase I. Proc Natl Acad Sci USA 97:4052–4057.
88 DAVOLI ET AL.




















