
Aggregation Pheromone of Palm Weevil, Dynamis borassi
11
Journa/ of Chemical Ecology, Vol. 23, No. 10, 1997 1 AGGREGATION PHEROMONE OF PALM WEEVIL, Dynamis borassi 1.* 2 2 - 3 R. M. GIBLIN-DA VIS, R. GRIES, G. GRIES, E. PENA-ROJAS, , 4 - 5 6 7 1. PINZON, J. E. PENA, A. L. PEREZ, H. D. PIERCE, JR., and A. C. OEHLSCHLAGER 7 . 8 I University of Florida, Fort Lauderdale Research and Education Center 3205 College Ave., FortLauderdale , Florida 33314-7799 2Centre for Pest Management, Department of Biological Sciences Simon Fraser University Burnabv, British Columbia V5A IS6, Cunada 'Corpo-ICA, C. 1. El Mira Apartado Aéreo 161, Tumaco, Nariño, Colombia "Palmas de Tumaco LTDA. Plantación Apartado Aéreo 248, Tumaco . Nariño, Colombia 'University of Florida, Tropical Research and Education Center 18905 S. W. 280 Street , Homestead, Florida 33031 "Department of Chemistry, University of Costa Rica 2060 San Pedro, San Jose, Costa Rica 7 Department of Chemistry, Simon Fraser University Burnabv, British Columbia V5A IS6, Canada 8ChemTica Internacional Apdo. 159-2150, San Jase, Costa Rica (Received December 12, 1996; accepted May 25, 1997) ;1 Abstract-4-Methyl-5-nonanol (1) is the male-produced aggregation phero- mone of the palm weevil, Dynamis borassi (F.) from Colombia. The phero- mone was identified by coupled gas chromatographic-electroantennographic detection (GC-EAD) analysis of male- and female-produced volatiles, and comparative GC-mass spectrometry (MS) of weevil-produced 1. In field experiments in Colombia, traps baited with a stereoisomeric mixture of syn- thetic 1 (3 mg/day) plus sugarcane captured more weevils than traps baited with 1 or sugarcane alone, suggesting that pheromone and plant volatiles are synergistically attractive. Use of a chiral, stereoisomer-separating Cyclo- dex-B column in GC-EAD and GC-MS analyses revealed that D. borassi males produce, and antennae of males and females respond to (4S,5S)-1. Previously identified palm weevil (Rhynchophorus spp.) aggregation phero- *To whom correspondence should be addressed. 2287 0098-0.H I 197/1000-2287/$ 12.50/0 © 1997 Plcnum Publishing Corporanon
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Aggregation Pheromone of Palm Weevil, Dynamis borassi

Journa/ of Chemical Ecology, Vol. 23, No. 10, 1997
1AGGREGATION PHEROMONE OF PALM WEEVIL,
Dynamis borassi
1.* 2 2 - 3R. M. GIBLIN-DA VIS, R. GRIES, G. GRIES, E. PENA-ROJAS,, 4 - 5 6 71. PINZON, J. E. PENA, A. L. PEREZ, H. D. PIERCE, JR.,
and A. C. OEHLSCHLAGER7.8
IUniversity of Florida, Fort Lauderdale Research and Education Center3205 College Ave., FortLauderdale , Florida 33314-7799
2Centre for Pest Management, Department of Biological SciencesSimon Fraser University
Burnabv, British Columbia V5A IS6, Cunada
'Corpo-ICA, C. 1. El MiraApartado Aéreo 161, Tumaco, Nariño, Colombia
"Palmas de Tumaco LTDA. PlantaciónApartado Aéreo 248, Tumaco . Nariño, Colombia
'University of Florida, Tropical Research and Education Center18905 S. W. 280 Street , Homestead, Florida 33031
"Department of Chemistry, University of Costa Rica2060 San Pedro, San Jose, Costa Rica
7Department of Chemistry, Simon Fraser UniversityBurnabv, British Columbia V5A IS6, Canada
8ChemTica InternacionalApdo. 159-2150, San Jase, Costa Rica
(Received December 12, 1996; accepted May 25, 1997)
;1
Abstract-4-Methyl-5-nonanol (1) is the male-produced aggregation phero-mone of the palm weevil, Dynamis borassi (F.) from Colombia. The phero-mone was identified by coupled gas chromatographic-electroantennographicdetection (GC-EAD) analysis of male- and female-produced volatiles, andcomparative GC-mass spectrometry (MS) of weevil-produced 1. In fieldexperiments in Colombia, traps baited with a stereoisomeric mixture of syn-thetic 1 (3 mg/day) plus sugarcane captured more weevils than traps baitedwith 1 or sugarcane alone, suggesting that pheromone and plant volatiles aresynergistically attractive. Use of a chiral, stereoisomer-separating Cyclo-dex-B column in GC-EAD and GC-MS analyses revealed that D. borassimales produce, and antennae of males and females respond to (4S,5S)-1.Previously identified palm weevil (Rhynchophorus spp.) aggregation phero-
*To whom correspondence should be addressed.
2287
0098-0.H I197/1000-2287/$ 12.50/0 © 1997 Plcnum Publishing Corporanon

2288 GIBLlN-DAVIS ET AL.
mones 5-methyl-4-octanol (cruentol) and 6-methyl-2-hepten-4-o1 (rhyncho-phorol) also elicited antennal responses by D. borassi. In field experiments,D. borassi females were captured equally well in traps baited with sugarcaneplus either 1, cruentol or rhynchophorol. In contrast, D. borassi maleswere captured most often in traps baited with sugarcane plus 1. Because D.borassi is a potential vector of the red ring nematode that causes the lethalred ring disease of palms, pherornone-based trapping of D. borassi could aidin monitoring or management of red ring disease in commercial palm plan-tations.
Key Words-Aggregation pheromone, Bursaphelenchus cocophilus, coconut,Cocos nucifera, Coleoptera, Curculionidae, Dvnamis borassi, 4-methyl-5-nonanol, palm weevils, red ring disease, red ring nematode.
INTROOUCTION
The palm weevil, Dynamis borassi (F.) is widely distributed in South America(Wattanapongsiri, 1966). As a major pest of coconut (Cocos nucifera L.), itdamages palm inflorescences and crown and vectors the lethal nematode [Bur-saphelenchus cocophilus (Cobb)J-caused red ring disease (Gerber et al., 1990).Dynamis borassi by itself causes death of Arikury palm, Syagrus schizophylla(Martius), and S. vagans (Bondar) (Bondar, 1940; Landeiro, 1941), and isattracted to maturing fruit of star-nut palm, Astrocaryum standleyanum (Giblin-Davis et al., unpublished observation).
Males of many of the rhynchophorinae produce aggregation pheromonesthat mediate colonization of susceptible host plants (Giblin-Davis et al., 1996).Capture of D. borassi in traps baited with the synthetic aggregation pheromone(±)-6-methyl-2-hepten-4-o1 (rhynchophorol) of the American palm weevil,Rhynchophorus palmarum (L.) and pesticide-treated sugarcane (Giblin-Davis etal., unpublished observation) suggest similarities in the chemical ecology ofthese weevils. We report the identification of the D. borassi aggregation pher-omone and cross-attraction of D. borassi to pheromones of Rhynchophorus palmweevils.
METHOOS ANO MATERIALS
Volatile Collections. Dynamis borassi ofmixed age and sex and ofunknownmating status were field-collected in an African oil palm (Elaeis guineensisJacquin) plantation at Palmas de Tumaco LTDA., Tumaco, Nariño, Colombia,provided with sugarcane, and shipped to Simon Fraser University , Canada, forcapture of male- and fernale-produced volatiles in May 1994 and 1996. Groupsof 5-10 weevils, separated by sex , were placed in modified 9-liter Naglene(Nalge Co., Rochester, New York) desiccators with 0-4-day-old apple slices.

PALM WEEVIL AGGREGATION PHEROMONE 2289
i¿
A charcoal-filtered air stream (l.5 crrr'rmin) was maintained through the des-iccators for five days, collecting insect volatiles on 10 g of Porapak Q (50-80mesh, Waters Associates, Inc., Milford, Massachusetts) packed in Pyrex glasstubing (Oehlschlager et al., 1988, 1992). Volatiles were eluted from the PorapakQ with distilled pentane and concentrated by distillation through a Dufton col-umn (300 X 20 mm) (Quickfit & Quartz Ltd., Staffordshire, England).
Volatile Analyses. Porapak Q extracts were analyzed by coupled gas chro-matographic-electroantennographic detection (GC-EAD) (Am et al., 1975),using a Hewlett Packard (HP) 5885B gas chromatograph equipped with aDB-5-coated, fused silica column (30 m x 0.25 mm ID) (J&W Scientific,Folsom, California). For GC-EAD recordings, an antenna was removed froma live weevil and suspended between two glass capillary electrodes with thescape inserted into one and the olfactory club impaled by the other. To obtainmas s spectra of EAD-active compounds, a coupled HP 5985B GC-mass spec-trometer (MS) equipped with an SP-I000-coated, fused silica column (30 m X
0.25 mm ID) (Supelco lnc., Bellefonte, Pennsylvania) was used in both electronimpact (El) (70 eV) and chemical ionization (CI) mode. To determine the wee-vil-produced stereoisomer(s) of 4-methyl-5-nonanol (1), weevil-produced 1 wasisolated from the Porapak Q extract by high-performance liquid chromatography(HPLC), employing a Waters LC 626 high-performance Iiquid chromatographequipped with a Waters 486 variable-wavelength UV-visible detector set at 210nm, a Waters 746 data module integrator, and a Nova-Pak silica HPLC cartridgecolumn (3.9 x 150 mm) with a 1 ml/min ftow of isocratic hexane (96 %)-ether(4%). HPLC-isolated 1 was then analyzed by GC-MS, employing a VarianSaturn GC-MS equipped with a fused silica Cyclodex-B-coated column (30 mX 0.25 mm ID) (J&W Scientific, Folsom, California). The temperature pro-gram was 80°C for 1 min, then 3°C/min to 95°C, then 95°C for 30 mino Todetermine the EAD-active stereoisomer of 1, synthetic 1 was analyzed by GC-EAD with the Cyclodex-B column.
To study olfactory recognition of congeneric aggregation pheromones, apentane solution was prepared for GC-EAD analyses containing the followingRhynchophorus pheromones at 5 ng each: (1) 1 (ferrugineol) (Hallett et al.,1993; Oehlschlager et al., 1995), (2) (± )-6-methyl-2-hepten-4-ol (rhynchopho-rol) (Oehlschlager et al., 1992; Rochat et al., 1991), (3) 5-methyl-4-octanol(cruentol) (Weissling et al., 1994), (4) 3-methyl-4-octanol (phoenicol) (Gries etal., 1993), and (5) 2-methyl-4-heptanol (Perez et al., 1997). AII compoundswere available in our laboratories from previous work (cited aboye) and were> 95 % chemically pureo Stereoisomers of 1 were synthesized using the methodsdescribed by Perez et al. (1996, 1997).
Field Experiments. Two field experiments were conducted near the bordersof an African oil palm plantation at Palmas de Tumaco LTDA., Tumaco, Nariño,Colombia, in a completely randomized block design with traps at about 50-m

2290 GIBLIN-DA VIS ET AL.
intervals and blocks > 100 m aparto Lethal traps were constructed from closed4-liter white plastic containers, similarly shaped to herbicide containers (30.0high X 16.5 wide X 10.0 cm deep) with small holes in the bottom and two 6X 13-cm entry slots cut into the sides. Each trap was suspended 1.5-1.8 maboye ground from an African oil palm.
The first four-treatment experiment tested unbaited control traps and trapsbaited with 1 or fresh sugarcane alone (about I kg; ten lO-cm long pieces offreshly cut stem) and in combination. All traps contained a water-moistenedrag. The sugarcane and/or rag was immersed in a suspension of liquid Sevin[21.3 % carbaryl; I-naphthyl N-methyl-carbamate (1.11 mi active ingredient/liter water)] to make each trap lethal to incoming weevils. About 90 mg ofsynthetic 1and an indicator dye were hermetically sealed in a polymer membranerelease device (ChemTica Intemational, San lose, Costa Rica) and hung insidethe trap for pheromone dissemination at 3 mg/day at 25°C. The dye was usedto indicate when the pheromone release device was empty.
Because of very low densities of D. borassi, we used the six replicate trapseach week as subsamples that were pooled for each treatment. Each treatmentwas replicated in time (eight replicates) from February 7 to April 4, 1995.Captured weevils were removed from traps weekly, sexed, and counted. Cleantraps were rebaited and rerandomized within each of the six blocks.
The second field experiment compared attraction of D. borassi to variousRhynchophorus pheromones. Traps were baited with sugarcane plus either syn-thetic 1, rhynchophorol, cruentol, or phoenicol. Each pheromone was dissem-inated at 3 mg/day at 25°C. The experimental design was as in experiment 1except that each treatment was replicated 12 times from April 4 to June 20,1995.
Statistical Analyses. Data from field tests were square root-transformed (x+ 0.5) to eliminate heteroscedasticity and analyzed by analysis of variance usingPROC ANOV A (SAS Institute, 1985). Least significant difference tests (SASInstitute, 1985) were used for mean separation where significant (P < 0.05)statistical differences occurred.
RESULTS AND DISCUSSION
GC-EAD analysis of volatiles from male and female D. borassi revealedone male-specific compound that elicited strong antennal responses from maleand female weevils (Figure 1). Mass spectral and retention characteristics ofthis compound were consistent with those of 4-methyl-5-nonanol (1), the mainaggregation pheromone of R. ferrugineus (Oliv.) (Hallett et al., 1993), R. bil-ineatus (Montr.) (Oehlschlager et al., 1995), and Metamasius hemipterus seri-
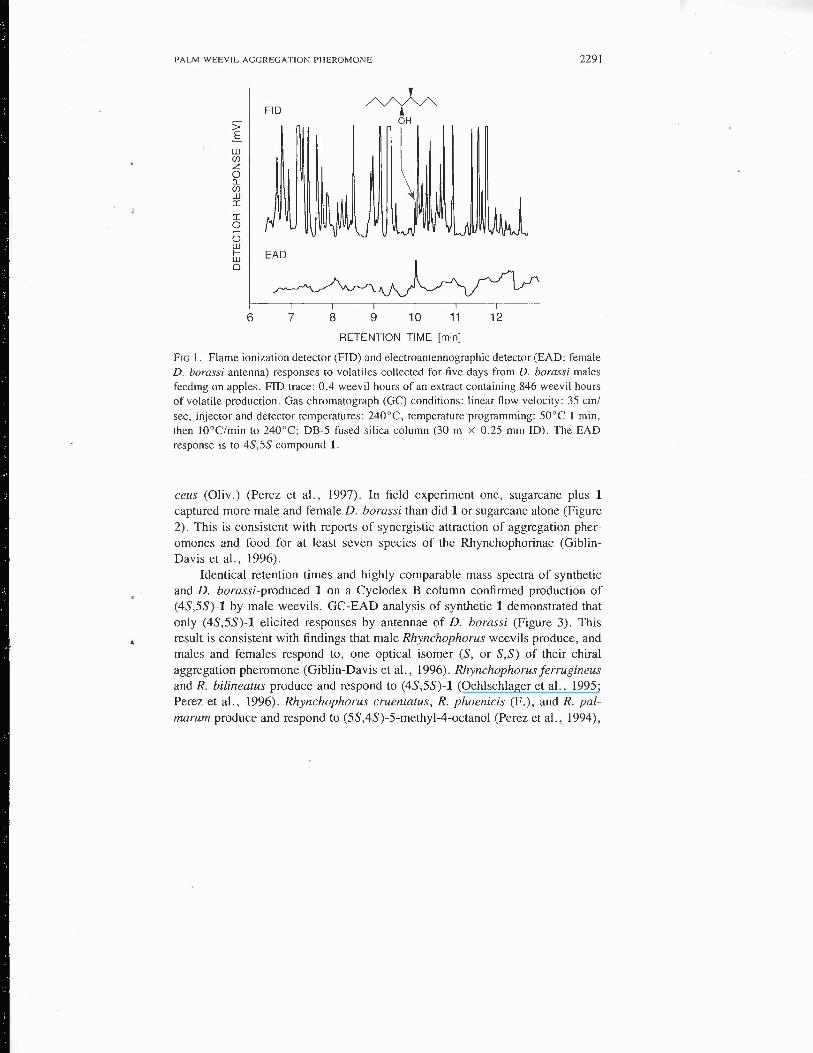
PALM WEEVIL AGGREGATION PHEROMONE 2291
FID ~:> OH
.s \w(f)zOe,(f)wa:a:Of-OWf- EADwO
6 7 8 9 10 11 12
RETENTION TIME [min]
FIG l. Flame ionization detector (FID) and electroantennographic detector (EAD: femaleD. borassi antenna) responses to volatiles collected for five days from D. borassi malesfeeding on apples. FID trace: 0.4 weevil hours of an extract containing 846 weevil hoursof volatile production. Gas chromatograph (GC) conditions: linear flow velocity: 35 cm/sec, injector and detector temperatures: 240°C, temperature programming: 50°C 1 min,then tO°C/min to 240°C; DB-5 fused silica column (30 m x 0.25 mm ID). The EADresponse is to 4S,5S compound 1.
ceus (Oliv.) (Perez et al., 1997). In field experiment one, sugarcane plus 1captured more mal e and female D. borassi than did 1 or sugarcane alone (Figure2). This is consistent with reports of synergistic attraction of aggregation pher-omones and food for at least seven species of the Rhynchophorinae (Giblin-Davis et al., 1996).
Identical retention times and highly comparable mass spectra of syntheticand D. borassi-produced 1 on a Cyclodex B column confirmed production of(4S,5S)-1 by male weevils. GC-EAD analysis of synthetic 1 demonstrated thatonly (4S,5S)-1 elicited responses by antennae of D. borassi (Figure 3). Thisresult is consistent with findings that male Rhynchophorus weevils produce, andmales and females respond to, one optical isomer (S, or S,S) of their chiralaggregation pheromone (Giblin-Davis et al., 1996). Rhynchophorus ferrugineusand R. bilineatus produce and respond to (4S,5S)-1 (Oehlschlager et al., 1995;Perez et al., 1996). Rhynchophorus cruenta tus , R. phoenicis (F.), and R. pal-marum produce and respond to (5S,4S)-5-methyl-4-octanol (Perez et al., 1994),

2292 GIBLIN-DAVIS ET AL.
B
bn Females
• MalesUnbaited
4-methyl-5-nonanol [3mg/day] Bb
Sugarcane [1 Kg]
I-------¡ASugarcane [1 Kg]4-methyl-5-nonanol [3mg/day]
1 2
MEAN (+SE) NUMBER OFD. borassi / WEEK
FIG. 2. Mean numbers (± SE) of D. borassi captured in lethal traps baited with sug-arcane or 4-methyl-5-nonanol singly and in combination. Wet rags were used in trapswithout sugarcane. Rags or sugarcane were treated with pesticide to prevent weevilescape. The experiment (eight relicates) was conducted in an African oil palm plantation,Palmas de Tumaco LTDA., Tumaco, Colombia, from February 7 to April 4, 1995. Datatransformed to approximate homogeneity are presented untransformed. Means within asex with the same letter superscript are not significantly different (P > 0.05; leastsignificant difference), ANOVA treatment effects-males: F = 8.15; df = 3, 10; P <0.001; females: F = 7.85; df = 3, 10; P < 0.002.
3TREATMENT
(3S,4S)-3-methyl-4-octanol (Perez et al., 1994), and (4S,2E)-6-methyl-2-hep-ten-4-o1 (Oehlschlager et al., 1992), respectively. In field trapping experimentswith the preceding Rhynchophorus weevils, nonweevil produced stereoisomersin synthetic pheromones did not reduce attractiveness of the S or S, S isomers,indicating that stereoisomeric blends can be used for trapping weevils (Giblin-Davis et al., 1996). Low population densities of D. borassi did not allow forfield tests of stereoisomers of 1. Because chemical communication of D. borassiand Rhynchophorus weevils are similar, we hypothesize that nonnatural isomers •of 1 in synthetic pheromone lures will not prevent optimal attraction of D.borassi to pheromone-baited traps.
D. borassi-produced 1, and also cruentol, rhynchophorol, and 2-methyl-4-heptanol elicited responses by D. borassi antennae (Figure 4), suggesting olfac-

PALM WEEVIL AGGREGATION PHEROMONE 2293
FID: Synthetic 4-methyl-5-nonanol
::>.sllJ(/)ZOa,(/)llJa:a:Of-ÜllJf-llJO
EAD: Antenna of a female D. borassí
22 24 26 28RETENTION TIME [min]
FIG.3. Flame ionization detector (FID) and electroantennographic detector (EAD: femaleD. borassi antenna) responses to stereoisomers of 4-methyl-5-nonanol. Cyclodex-B col-urnn, isothermal IOO°C, linear ftow velocity of carrier gas 35 cm/sec; split injection;injector temperature 220°C.
tory recognition of aggregation pheromones of other weevils in the Rhyncho-phorinae (Giblin-Davis et al., 1996). This is similar to M. hemipterus , whereantennal responses were elicited by pheromones of other weevils belonging tothe same subfamily (Ramirez-Lucas et al., 1996). In field experiment 2, trapsbaited with sugarcane plus either cruentol, rhynchophorol, or 1were equallyattractive to D. borassi females and more attractive than traps baited with sug-arcane plus phoenicol (Figure 5). More males were captured in traps baited withsugarcane plus 1 than in traps baited with sugarcane plus any of the other threepheromones (Figure 5). The attraction of D. borassi to cruentol produced byallopatric North American R. cruentatus is not understood. However, cross-attraction of D. borassi females to rhynchophorol, the aggregation pheromoneof sympatric R. palmarum, may be adaptive in overcoming a palm's defenseand in time-efficient use of a temporarily suitable resource. Sympatric West

2294 GIBLIN-DA VIS ET AL.
:>.sw(J)zOQ__(J)wa:a:Of-OWf-WO
FID
OH~JOH f' OH ~lOH
~~t ~OH
EAD
6 7 8 9 10 11
RETENTION TIME [min]
FIG. 4. Flame ionization detector (FID) and electroantennographic detector (EAD: femaleD. borassi antenna) responses to (Ieft to right on figure) 2-methyl-4-heptanol, 6-methyl-2-hepten-4-01 (rhynchophorol), 5-methyl-4-octanol (cruentol), 3-methyl-4-octanol(phoenicol) and 4-methyl-5-nonanol (ferrugineol) (aggregation pheromones in the Rhyn-chophorinae). Column and chromatographic conditions as in Figure l.
Indian sugarcane weevil, M. h. sericeus, which uses 1as its major aggregationpheromone (Perez et al., 1997; Ramirez-Lucas et al., 1996) is also cross-attractedto rhynchophorol (Chinchilla et al., unpublished data). Dynamis borassi and R.palmarum are known vectors and M. h. sericeus is a suspected vector of thered ring nematode (Bursaphelenchus cocophilus) (Giblin-Davis et al., 1996).Because D. borassi, R. palmarum, and M. h. sericeus can coinhabit the crownof red ring diseased coconut palms (Gerber et al., 1990; Giblin-Davis et al.,unpublished observation) these weevils together may overwhelm the palm'sdefense and beco me associated with red ring nematodes. As a result, they mayincrease their virulence, successfully occupying more hosts and enhancing theirreproductive potential. Niche divergence and cannibalism of weevil larvae mayserve to reduce interspecific competition in an occupied host (Giblin-Davis etal., 1996).
Knowledge of the chemical ecology of D. borassi can now help determineits population dynamics in pa1m plantations and in the wild and its relativeimportance in red ring nematode transmission.

PALM WEEVIL AGGREGATION PHEROMONE 2295
Sugarcane [1Kg]3-methyl-4-octanol [3mg/day]
DFemales_Males
Sugarcane [1 Kg]5-methyl-4-octanol [3mg/day]
Sugarcane [1 Kg]6-methyl-2-hepten-4-01 [3mg/day]
A
Sugarcane [1 Kg]4-methyl-5-nonanol [3mg/day]
1 234
MEAN (+SE) NUMBER OFD. borassi / WEEK
FIG. 5. Mean numbers (± SE) of D. borassi captured in lethal traps baited with sug-arcane plus either 4-methyl-5-nonanol (ferrugineol), ó-rnethyl-z-hepten-a-ol (rhyncho-phorol), 5-methyl-4-octanol (cruentol), or 3-methyl-4-octanol (phoenicol). Sugarcane wastreated with pesticide to prevent weevil escape. The experiment (12 replicates) wasconducted in an African oil palm plantation, Palmas de Tumaco LTOA., Turnaco, Co-lombia, from April 4 to June 20, 1995. Data transformed to approximate homogeneityare presented untransformed. Bars for male and female weevils with the same lettersuperscript are not significantly different (P > 0.05; least significant difference), ANOVAtreatment effects-males; F = 22.22; df = 3, 14; P < 0.0001; females: F = 6.67; df= 3, 14; P = 0.002.
TREATMENT
Acknowledgments=we thank Dr. C. W. O'Brien for confirmation of the identification ofDynamis borassi, Dr. A. Bellotti for assistance in shipping weevils out of Colornbia, L. M. Gonzalezfor packaging the pheromones, and C. Hubbuch for identification of Astrocaryum standleyanum.We also thank Drs. T. J. Weissling and F. W. Howard for critically reviewing an earlier versionof this manuscript. This research was supported in part by grants from the Natural Sciences andResearch Council of Canada and the Presidenr's Research Council of Canada to A. C. O. and aUSDA Special Grant in Tropical and Subtropical Agriculture CRSR-95-34135-1763 to R.M.G.-D.and J. E. P. This manuscript is Florida Agricultural Research Station Joumal Series R-05529.
REFERENCES
ARN, H., STADLER,E., and RAUSCHER,S. 1975. The electroantennographic detector-a selectiveand sensitive tool in the gas chromatographic analysis al' insect pheromones. Z. Naturforsch.
30c:722-725.

2296 GIBLIN-DAVIS ET AL.
BONDAR, G. 1940. Insectos nocivos e molestias do coqueiro (Cocos nuciferai no Brazil. lnst. Cent.Fom. Econ. Bahia, Bol. 8: 1-160.
GERBER, K., GIBLlN-DAvls, R. M., and ESCOBAR-GOYES, J. 1990. Association of the red ringnematode, Rhadinaphelenchus cocophilus, with weevils from Ecuador and Trinidad. Nema-tropica 20:39-49.
GIBLlN-DAvls, R. M., OEHLSCHLAGER, A. C., PEREZ, A. L., GRIES, G., GRIES, R., WEISSLlNG, T.
J., CHINCHILLA, C. M., PEÑA, J. E., HALLETT, R. H., PIERCE, H. D., JR., and GONZALEZ,L. M. 1996. Chemical and behavioral ecology of palm weevils (Curculionidae: Rhyncho-phorinae). Fla. Entomol. 79: 153-167.
GRIES, G., GRIES, R., PEREZ, A. L., OEHLSCHLAGER, A. C., GONZALES, L. M., PIERCE, H. D.,JR., KOUDA, M., ZEBEYOU, M., and NANOU, N. 1993. Aggregation pheromone of the Africanpalm weevil, Rhynchophorus phoenicis F. Naturwissenschaften 80:90-91.
HALLETT, R. H., GRIES, G., GRIES, R., BORDEN, J. H., CZYZEWSKA, E., OEHLSCHLAGER, A. c..PIERCE, H. D., JR., ANGERILLI, N. P. D., and RANF, A. 1993. Aggregation pheromone oftwo Asian palm weevils, Rhynchophorus ferrugineus and R. vulneratus. Naturwissenschaften80:328-331.
LANDEIRO, R. 1941. Dynamis politus Gyll. iRhvnchophorus politus Gyll.) Urna nova praga do
coqueiro. Bol. Minist. Agric. Indo Com.; Rio de Janeiro 30: 1-6.OEHLSCHLAGER, A. C., PIERCE, A. M., PIERCE, H. D., JR., and BORDEN, J. H. 1988. Chemical
communication in cucujid grain beetles. 1. Chem. Ecol. 14:2071-2098.OEHLSCHLAGER, A. c.. PIERCE, H. D., JR., MORGAN, B., WIMALARATNE, P. D. C., SLESSOR, K.
N., KING, G. G. S., GRIES, G., GRIES, R., BORDEN., J. H.,·JIRON, L. F., CHINCHILLA, C.
M., and MEXZAN, R. G. 1992. Chirality and field activity 01' rhynchophorol, the aggregationpheromone of the American palm weevil. Naturwissenschaften 79: 134-135.
OEHLSCHLAGER, A. C., PRIOR, R. N. B., PEREZ, A. L., GRIES, R., GRIES, G., PIERCE, H. D., JR.,and LAuP, S. 1995. Structure, chirality, and field testing of a male-produced aggregationpheromone of Asian palm weevil Rhynchophorus bilineatus (Montr.) (Coleoptera: Curculion-idae). J. Chem. Ecol. 21:1619-1629.
PEREZ, A. L., GRIES, G., GRIES, R., GIBLlN-DAVIS, R. M., and OEHLSCHLAGER, A. C. 1994.Pheromone chirality of the African palm weevil, Rhynchophorus phoenicis (F.) and the pal-
metto weevil, Rhynchophorus cruenta/lis (F.) (Coleóptera: Curculionidae). 1. Chem. Ecol.20:2653-2671.
PEREZ, A. L., HALLETT, R., GRIES, R., GRIES, G., OEHLSCHLAGER, A. C., and BORDEN, J. H.
1996. Pheromone chirality of Asian palm weevils, Rhynchophorus ferrugineus (Oliv.) andRhynchophorus vulneratus (Panz.) (Coleoptera: Curculionidae). 1. Chem. Ecol. 22:357-368.
PEREZ, A. L., CAMPOS-PIEDRA, Y., CHINCHILLA, C. M., OEHLSCHLAGER, A. C., GRIES, G., GRIES,R., CASTRILLO, G., GIBLlN-DAVIS, R. M., PEÑA, 1. E., DUNCAN, R. E., GONZALEZ, L. M.,
PIERCE, H. D., JR., McDoNALD, R., and ANDRADE, R. 1997. Aggregation pheromones andhost kairomones of the West Indian sugarcane weevil, Metamasius hemipterus sericeus (Oliv.)(Coleóptera: Curculionidae). 1. Chem. Ecol. 23:869-888.
RAMIREZ-LuCAS, P., MALOSSE, c., DUCROT, P. H., LETTERE, M., and ZAGATTI, P. 1996. Chemicalidentification, electrophysiological and behavioral activities of the pheromone of Metamasiushemipterus (Coleoptera: Curculionidae). Bioorg. Med. Chem. 4:323-368.
RAMIREz-LuCAs, P., ROCHAT, D., and ZAGATTI, P. 1996. Field trapping of Metamasius hemipteruswith synthetic aggregation pheromone. Entomol. Exp. Appl. 80:453-460.
ROCHAT, D., MALOSSE, c., LETTERE, M., DUCROT, P. H., ZAGATTI, P., RENOU, M., and DESCOINS,C. 1991. Male-produced aggregation pheromone of the American palm weevil, Rhynchophoruspalmarum (L.) (Coleoptera: Curculionidae): Collection, identification, electrophysiologicalactivity, and laboratory bioassay. 1. Chem. Ecol. 17:2127-2140.

PALM WEEVIL AGGREGATION PHEROMONE 2297
SAS 1 STITUTE. 1985. SAS User's Guide: Statistics, 5th ed. SAS Institute, Cary, North Carolina.WATTA APONGSIRI, A. 1966. A revision of the genera Rhynchophorus and Dynamis (Coleoptera:
Curculionidae). Departrnent of Agricultural Science Bulletin, Bangkok 1: 1-328.WEISSLlNG, T. J., GIBLlN-DAvls, R. M., GRIES, G., GRIES, R., PEREZ, A. L., PIERCE, H. D., JR.,
and.OEHLSCHLAGER, A. C. 1994. Aggregation pherornone of palmetto weevil Rhynchophoruscruentatus (F.) (Coleoptera: Curculionidae). 1. Chem. Eco!. 20:505-515.




















