
Enhanced Therapeutic Efficacy for Ovarian Cancer with a Serotype 3 Receptor-Targeted Oncolytic...
10
Enhanced Therapeutic Efficacy for Ovarian Cancer with a Serotype 3 Receptor-Targeted Oncolytic Adenovirus Anna Kanerva, 1,4,5,6 Kurt R. Zinn, 2 Tandra R. Chaudhuri, 2 John T. Lam, 1 Kaori Suzuki, 1,7 Taco G. Uil, 1,8 Tanja Hakkarainen, 1,9 Gerd J. Bauerschmitz, 1 Minghui Wang, 1 Bin Liu, 1 Zhihong Cao, 2 Ronald D. Alvarez, 3 David T. Curiel, 1 and Akseli Hemminki 1,4,5,† 1 Division of Human Gene Therapy, Department of Medicine, Department of Pathology, and Department of Surgery, 2 Department of Radiology, and 3 Department of Obstetrics and Gynecology, University of Alabama at Birmingham, Birmingham, Alabama, USA 4 Rational Drug Design, Biomedicum, University of Helsinki, Finland; 5 Department of Oncology, Helsinki University Central Hospital, 00014 Helsinki, Finland 6 Department of Surgery, Helsinki University Central Hospital–Jorvi Hospital, Espoo, Finland 7 Molecular Medicine Program, Mayo Clinic and Foundation, Rochester, Minnesota 8 Department of Molecular Cell Biology, Leiden University Medical Center, Leiden, The Netherlands 9 Department of Biotechnology and Molecular Medicine, A. I. Virtanen Institute for Molecular Sciences, University of Kuopio, Kuopio, Finland † To whom correspondence and reprint requests should be addressed at the Cancer Gene Therapy Group, Rational Drug Design, Biomedicum Helsinki, Department of Oncology, Helsinki University Central Hospital, P.O. Box 63 (Haartmaninkatu 8), 00014 University of Helsinki, Finland. Fax: 358-9-1912 5155. E-mail: Akseli.Hemminki@Helsinki.fi. Oncolytic viruses that are replication competent in tumor but not in normal cells represent a novel approach for treating neoplastic diseases. However, the oncolytic potency of replicating agents is determined directly by their capability of infecting target cells. Most adenoviruses used for gene therapy or virotherapy have been based on serotype 5 (Ad5). Unfortunately, expression of the primary receptor for Ad5 (the coxsackie–adenovirus receptor, or CAR) is highly variable on ovarian and other cancer cells. By performing genetic fiber pseudotyping, we created Ad5/3-24, a conditionally replicating adenovirus that does not bind CAR but facilitates entry into and killing of ovarian cancer cells. We show replication of Ad5/3-24 and subsequent oncolysis of ovarian adenocarcinoma lines. Replication was also analyzed with quantitative PCR on three-dimensional primary tumor cell spheroids purified from patient samples. Moreover, in a therapeutic orthotopic model of peritoneal carcinomatosis, dramatically enhanced survival was noted. Finally, Ad5/3-24 achieved a significant antitumor effect as assessed by noninvasive, in vivo bioluminescence imaging. Therefore, the preclinical therapeutic efficacy of Ad5/3-24 is improved over the respective CAR- and integrin-binding controls. Taken together with promising biodistribution and toxicity data, this approach could translate into successful clinical interventions for ovarian cancer patients. Key Words: gene therapy, ovarian neoplasms, adenovirus, virus replication, biological therapy INTRODUCTION Adenovirus-mediated gene therapy has been proposed as a treatment alternative for advanced cancers refractory to other therapies. Adenoviruses are attractive vectors for cancer due to their unparalleled capacity for gene transfer, stability in vivo, and easy high-titer production. Conse- quently, replication-deficient first-generation viruses have shown high preclinical transduction rates and good antitumor efficacy in animal models. Most completed clinical trials have been Phase I, and the safety data have been excellent. However, there is little evidence support- ing significant clinical benefits [1]. Nevertheless, support- ing the feasibility of adenoviral gene therapy, positive results were reported in the only randomized trial pub- lished to date, albeit a Phase I [2]. Recently, it has been suggested that the binding of adenovirus to its primary receptor may be an important rate-limiting step for gene transfer [reviewed in 3]. As most adenoviral gene therapy vectors are based on sero- type 5 (Ad5), which binds to the coxsackie–adenovirus receptor (CAR), lack of CAR could make target tissues refractory [3,4]. Indeed, recent data suggest that CAR ex- pression in tumors may be highly variable, resulting in resistance to Ad5 infection [3,5–9]. Specifically, resistance may be due to low expression levels or aberrant localiza- tion at the cellular or tissue level [10]. Consequently, various strategies have been evaluated to modify adeno- ARTICLE doi:10.1016/S1525-0016(03)00200-4 449 MOLECULAR THERAPY Vol. 8, No. 3, September 2003 Copyright © The American Society of Gene Therapy 1525-0016/03 $30.00
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Enhanced Therapeutic Efficacy for Ovarian Cancer with a Serotype 3 Receptor-Targeted Oncolytic...

Enhanced Therapeutic Efficacy for Ovarian Cancer with aSerotype 3 Receptor-Targeted Oncolytic Adenovirus
Anna Kanerva,1,4,5,6 Kurt R. Zinn,2 Tandra R. Chaudhuri,2 John T. Lam,1 Kaori Suzuki,1,7
Taco G. Uil,1,8 Tanja Hakkarainen,1,9 Gerd J. Bauerschmitz,1 Minghui Wang,1 Bin Liu,1
Zhihong Cao,2 Ronald D. Alvarez,3 David T. Curiel,1 and Akseli Hemminki1,4,5,†
1Division of Human Gene Therapy, Department of Medicine, Department of Pathology, and Department of Surgery, 2Department of Radiology, and3Department of Obstetrics and Gynecology, University of Alabama at Birmingham, Birmingham, Alabama, USA
4Rational Drug Design, Biomedicum, University of Helsinki, Finland; 5Department of Oncology, Helsinki University Central Hospital, 00014 Helsinki, Finland6Department of Surgery, Helsinki University Central Hospital–Jorvi Hospital, Espoo, Finland
7Molecular Medicine Program, Mayo Clinic and Foundation, Rochester, Minnesota8Department of Molecular Cell Biology, Leiden University Medical Center, Leiden, The Netherlands
9Department of Biotechnology and Molecular Medicine, A. I. Virtanen Institute for Molecular Sciences, University of Kuopio, Kuopio, Finland
†To whom correspondence and reprint requests should be addressed at the Cancer Gene Therapy Group, Rational Drug Design, Biomedicum Helsinki,Department of Oncology, Helsinki University Central Hospital, P.O. Box 63 (Haartmaninkatu 8), 00014 University of Helsinki, Finland. Fax: �358-9-1912
5155. E-mail: [email protected].
Oncolytic viruses that are replication competent in tumor but not in normal cells represent a novelapproach for treating neoplastic diseases. However, the oncolytic potency of replicating agents isdetermined directly by their capability of infecting target cells. Most adenoviruses used for genetherapy or virotherapy have been based on serotype 5 (Ad5). Unfortunately, expression of theprimary receptor for Ad5 (the coxsackie–adenovirus receptor, or CAR) is highly variable onovarian and other cancer cells. By performing genetic fiber pseudotyping, we created Ad5/3-�24,a conditionally replicating adenovirus that does not bind CAR but facilitates entry into and killingof ovarian cancer cells. We show replication of Ad5/3-�24 and subsequent oncolysis of ovarianadenocarcinoma lines. Replication was also analyzed with quantitative PCR on three-dimensionalprimary tumor cell spheroids purified from patient samples. Moreover, in a therapeutic orthotopicmodel of peritoneal carcinomatosis, dramatically enhanced survival was noted. Finally, Ad5/3-�24achieved a significant antitumor effect as assessed by noninvasive, in vivo bioluminescenceimaging. Therefore, the preclinical therapeutic efficacy of Ad5/3-�24 is improved over therespective CAR- and integrin-binding controls. Taken together with promising biodistribution andtoxicity data, this approach could translate into successful clinical interventions for ovarian cancerpatients.
Key Words: gene therapy, ovarian neoplasms, adenovirus, virus replication, biological therapy
INTRODUCTION
Adenovirus-mediated gene therapy has been proposed asa treatment alternative for advanced cancers refractory toother therapies. Adenoviruses are attractive vectors forcancer due to their unparalleled capacity for gene transfer,stability in vivo, and easy high-titer production. Conse-quently, replication-deficient first-generation viruseshave shown high preclinical transduction rates and goodantitumor efficacy in animal models. Most completedclinical trials have been Phase I, and the safety data havebeen excellent. However, there is little evidence support-ing significant clinical benefits [1]. Nevertheless, support-ing the feasibility of adenoviral gene therapy, positive
results were reported in the only randomized trial pub-lished to date, albeit a Phase I [2].
Recently, it has been suggested that the binding ofadenovirus to its primary receptor may be an importantrate-limiting step for gene transfer [reviewed in 3]. Asmost adenoviral gene therapy vectors are based on sero-type 5 (Ad5), which binds to the coxsackie–adenovirusreceptor (CAR), lack of CAR could make target tissuesrefractory [3,4]. Indeed, recent data suggest that CAR ex-pression in tumors may be highly variable, resulting inresistance to Ad5 infection [3,5–9]. Specifically, resistancemay be due to low expression levels or aberrant localiza-tion at the cellular or tissue level [10]. Consequently,various strategies have been evaluated to modify adeno-
ARTICLEdoi:10.1016/S1525-0016(03)00200-4
449MOLECULAR THERAPY Vol. 8, No. 3, September 2003Copyright © The American Society of Gene Therapy1525-0016/03 $30.00

virus tropism to circumvent CAR deficiency, includingretargeting complexes or genetic capsid modifications [4].
In earlier studies, we explored substituting the receptorbinding fiber knob domain of Ad5 with the serotype 3(Ad3) knob. This resulted in CAR-independent virus at-tachment to ovarian and other cancer cells, as Ad3 has adistinct, but unidentified receptor [11]. Further, our pre-vious studies suggest that, in comparison to CAR, the Ad3receptor is expressed at higher levels on ovarian cancercells and that 5/3 serotype chimeras bind to the Ad3receptor but not to CAR as analyzed by competition withfree Ad knob protein [5]. Importantly, Ad5/3 displayedhigher transgene expression in primary ovarian cancercells. Furthermore, the murine safety profile of Ad5/3 wasevaluated by biodistribution, liver toxicity, and bloodclearance analyses and was comparable to that of Ad5 [6].
The main reason for the mostly less than impressiveclinical results obtained with replication-deficient agentsmay be related to inefficient transduction of solid tumormasses [12]. To help overcome this obstacle, selectivelyoncolytic adenoviruses, i.e., conditionally replicating ad-enoviruses (CRAds), have been constructed. Infection oftumor cells results in replication, oncolysis, and subse-quent release of the virus progeny. In combination withchemotherapeutics, CRAds have recently displayed someclinical utility [13]. Normal tissue is spared due to abro-gation of replication, which can be achieved by incorpo-ration of mutations in immediately-early (E1A) or early(E1B) adenoviral genes resulting in mutant E1 proteinsunable to bind cellular proteins necessary for viral repli-cation in normal cells, but not in cancer cells [14–17].Ad5-�24 contains a 24-bp deletion in constant region 2(CR2) of E1A, and the expressed protein is unable to bindretinoblastoma (Rb) protein for induction of S phase [15].Thus, Ad5-�24 is unable to replicate in nondividing nor-mal cells, but replicates in cells inactive in the Rb/p16pathway [15,16]. It has been suggested that all humancancers, including ovarian cancers, may be deficient inthis crucial pathway [18].
Recently, it has been demonstrated that the oncolyticpotency of replicating agents is determined directly by theircapability of infecting target cells [19,20]. Thus, variableCAR expression on cancer cells could hinder CRAd-medi-ated oncolysis. Therefore, methods to circumvent CAR de-ficiency and improve cell killing have been evaluated in thecontext of CRAds. Heretofore, these endeavors include in-fectivity enhancement of the agent by incorporating motifsin fiber, such as the integrin-binding arginine-glycine-aspar-tic acid (RGD-4C) modification into the HI loop of the knob[21,22] or the heparan sulfate-binding polylysine residue atthe COOH-terminus of the fiber [23]. These viruses bindboth CAR and an alternative receptor. In this study, ablationof CAR binding was achieved by genetic pseudotyping ofthe fiber with the knob domain from Ad3 (Ad5/3-�24, Fig.1). This may be a member of a new generation of agents thatutilize a non-CAR pathway, which may be advantageous for
enhanced tumor targeting. Further, the novel agent wascompared to Ad5-�24RGD, a recently described promisingoncolytic agent, which is currently in ovarian cancer clinicaltrials.
RESULTS
Ad5/3-�24 Displays Efficient Killing of OvarianCancer Cells in VitroWe infected monolayers of SKOV3.ip1, OV-4, OV-3, Hey,and ES-2 cells (Fig. 2) with Ad5/3-�24, Ad5-�24E3, andAdCMVHSV-TK. In all cell lines, the quantitative cell kill-ing assay showed oncolysis with Ad5/3-�24, while Ad5-�24E3 (the isogenic control with the Ad5 fiber) causedminimal (OV-4, OV-3, Hey) or no cell killing (SKOV3.ip1,ES-2). The percentage of viable cells remaining with Ad5/3-�24 was 0.5, 5.6, 15, 2.7, and 5.5% for SKOV3.ip1, OV-4,OV-3, Hey, and ES-2, respectively, compared to unin-fected wells. With Ad5-�24E3, viability was 117, 91, 87,85, and 109%, respectively. On all cell lines, oncolysis wassignificantly improved with fiber-modified Ad5/3-�24 incomparison to Ad5-�24E3 (P � 0.0001, P � 0.0001, P �0.0033, P � 0.0011, and P � 0.0001 for SKOV3.ip1, OV-4,OV-3, Hey, and ES-2, respectively). AdCMVHSV-TK wasincluded as an E1-deleted control, and it did not causeoncolysis (P � 0.0018 for SKOV3.ip1, P � 0.0001 for othercell lines, compared to Ad5/3-�24).
Ad5/3-�24 Replicates in Ovarian Cancer Primary CellSpheroidsWe analyzed four purified, unpassaged human ovariancancer samples for adenovirus replication. To measureviral copy number, we collected spheroids and growthmedium at indicated time points and performed quanti-tative PCR for the adenoviral E4 gene (Figs. 3A–3D). Todetermine the relative increase in copies, we normalized
FIG. 1. Structure of Ad5/3-�24. Ad5/3-�24 has a 24-bp deletion in constantregion 2 (CR2) of the E1A gene, corresponding to the region utilized for Rbprotein binding. This results in an agent replication competent and oncolyticin cells defective in the Rb/p16 pathway, such as ovarian cancer cells. The fiberis modified to incorporate the serotype 3 knob, while retaining the Ad5 shaftand tail.
ARTICLE doi:10.1016/S1525-0016(03)00200-4
450 MOLECULAR THERAPY Vol. 8, No. 3, September 2003Copyright © The American Society of Gene Therapy

E4 copy number at each time point to the copy numberobtained with AdCMVHSV-TK at 12 h. On day 4 afterinfection, Ad5/3-�24 copy number had increased 378-,149-, 562-, and 212-fold in patient samples 1–4, respec-tively, compared to 27.3-, 71.1-, 11.6-, and 104-fold in-creases with Ad5-�24E3. On day 12, Ad5/3-�24 copynumber had increased to 406-, 267-, 572-, and 73.8-fold,while these figures were 252-, 115-, 172-, and 131-fold forAd5-�24E3. Thus, replication of Ad5/3-�24 occurredmore rapidly than that of Ad5-�24E3 in all cases and to ahigher degree in two of four patients (Figs. 3A and 3C).
To estimate total virus production and allow visualcomparison of replication dynamics, we calculated cumu-
lative virus copy numbers (Figs. 3E–3H). With patients1–3 the highest cumulative amount of E4 copies wasmeasured with Ad5/3-�24. When total cumulative virusproduction was compared, Ad5/3-�24 produced 241-,63.1-, 450-, and 54.9-fold more virus than AdCMV-HSV-TK (patients 1–4, respectively). Ad5-�24E3 achieved124-, 35.5-, 116-, and 57.8-fold higher total virus produc-tion than AdCMVHSV-TK.
Ad5/3-�24 in an Orthotopic Murine Model ofOvarian CancerWe tested the efficacy of Ad5/3-�24 in an ip model ofdisseminated ovarian cancer. We allowed advanced carci-
FIG. 2. Ad5/3-�24 displays increased cellkilling of ovarian cancer cells. Cells wereinfected with 0, 0.1, 1, or 10 vp/cell ofAd5/3-�24, Ad5-�24E3 (non-fiber-modi-fied isogenic control virus), or AdCMV-HSV-TK (E1-deleted control virus). On day 6(SKOV3.ip1), day 13 (OV-4), day 6 (OV-3),day 10 (Hey), or day 10 (ES-2) cell viabilitywas measured with the MTS assay. TheOD490 values of uninfected cells were set as100%. Data are expressed as means � SD ofquadruplicate experiments. With all celllines, oncolysis was significantly improvedwith Ad5/3-�24 compared to Ad5-�24E3 orAdCMVHSV-TK (all P values � 0.004).
ARTICLEdoi:10.1016/S1525-0016(03)00200-4
451MOLECULAR THERAPY Vol. 8, No. 3, September 2003Copyright © The American Society of Gene Therapy

nomatosis to develop for 10 days and then treated themice with a single injection of Ad5/3-�24, Ad5-�24E3,AdCMVHSV-TK, or no virus (Fig. 4A). Ad5-�24E3 is anisogenic control virus with the Ad5 fiber. AdCMVHSV-TKwas included to control for the effect of virus not capableof significant replication. The median survival was notreached for Ad5/3-�24, and 50% of mice were alive at theend of the experiment on day 135. For Ad5-�24E3, AdCMV-HSV-TK, and no virus, the median survivals of mice were96.5, 32 and 33 days, respectively. In comparison to the
other groups, the overall survival of mice treated withAd5/3-�24 was statistically significantly improved (log-rank test P � 0.0001). In pair-wise comparisons survivalwith Ad5/3-�24 was significantly enhanced (versus Ad5-�24E3, P � 0.0001; versus AdCMVHSV-TK, P � 0.0001;versus no virus, P � 0.0001).
Injection on 3 consecutive days was also tested (Fig.4B). Again, the median survival of mice treated with Ad5/3-�24 was not reached, and 50% of mice were alive on day135. For Ad5-�24E3, AdCMVHSV-TK, and no virus, the
FIG. 3. Ad5/3-�24 replicates in three-dimensional human ovarian primarycancer cell spheroids. Purified, unpas-saged cancer cells were allowed toform spheroids, which were infectedwith 1000 vp/surface cell of Ad5/3-�24, Ad5-�24E3 (non-fiber-modifiedisogenic control virus), or AdCMV-HSV-TK (E1-deleted control virus).Spheroids and growth medium wereharvested at indicated time points, andvirus copy number was measured withquantitative PCR. (A–D) The increase invirus copy number with Ad5/3-�24and Ad5-�24E3 in patient samples1–4, respectively. (E–H) Cumulativevalues from patients 1–4 allow com-parison of total virus production andreplication kinetics. The backgroundvalues (uninfected spheroids) weresubtracted from the data at each timepoint.
ARTICLE doi:10.1016/S1525-0016(03)00200-4
452 MOLECULAR THERAPY Vol. 8, No. 3, September 2003Copyright © The American Society of Gene Therapy
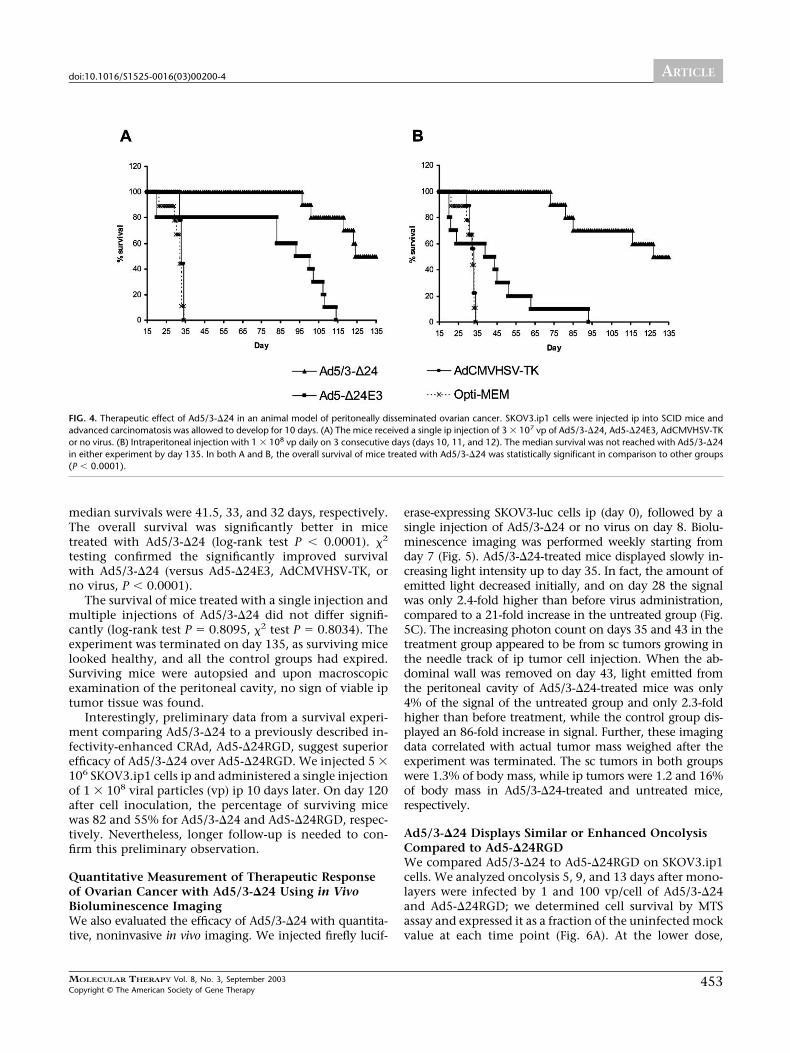
median survivals were 41.5, 33, and 32 days, respectively.The overall survival was significantly better in micetreated with Ad5/3-�24 (log-rank test P � 0.0001). �2
testing confirmed the significantly improved survivalwith Ad5/3-�24 (versus Ad5-�24E3, AdCMVHSV-TK, orno virus, P � 0.0001).
The survival of mice treated with a single injection andmultiple injections of Ad5/3-�24 did not differ signifi-cantly (log-rank test P � 0.8095, �2 test P � 0.8034). Theexperiment was terminated on day 135, as surviving micelooked healthy, and all the control groups had expired.Surviving mice were autopsied and upon macroscopicexamination of the peritoneal cavity, no sign of viable iptumor tissue was found.
Interestingly, preliminary data from a survival experi-ment comparing Ad5/3-�24 to a previously described in-fectivity-enhanced CRAd, Ad5-�24RGD, suggest superiorefficacy of Ad5/3-�24 over Ad5-�24RGD. We injected 5 �106 SKOV3.ip1 cells ip and administered a single injectionof 1 � 108 viral particles (vp) ip 10 days later. On day 120after cell inoculation, the percentage of surviving micewas 82 and 55% for Ad5/3-�24 and Ad5-�24RGD, respec-tively. Nevertheless, longer follow-up is needed to con-firm this preliminary observation.
Quantitative Measurement of Therapeutic Responseof Ovarian Cancer with Ad5/3-�24 Using in VivoBioluminescence ImagingWe also evaluated the efficacy of Ad5/3-�24 with quantita-tive, noninvasive in vivo imaging. We injected firefly lucif-
erase-expressing SKOV3-luc cells ip (day 0), followed by asingle injection of Ad5/3-�24 or no virus on day 8. Biolu-minescence imaging was performed weekly starting fromday 7 (Fig. 5). Ad5/3-�24-treated mice displayed slowly in-creasing light intensity up to day 35. In fact, the amount ofemitted light decreased initially, and on day 28 the signalwas only 2.4-fold higher than before virus administration,compared to a 21-fold increase in the untreated group (Fig.5C). The increasing photon count on days 35 and 43 in thetreatment group appeared to be from sc tumors growing inthe needle track of ip tumor cell injection. When the ab-dominal wall was removed on day 43, light emitted fromthe peritoneal cavity of Ad5/3-�24-treated mice was only4% of the signal of the untreated group and only 2.3-foldhigher than before treatment, while the control group dis-played an 86-fold increase in signal. Further, these imagingdata correlated with actual tumor mass weighed after theexperiment was terminated. The sc tumors in both groupswere 1.3% of body mass, while ip tumors were 1.2 and 16%of body mass in Ad5/3-�24-treated and untreated mice,respectively.
Ad5/3-�24 Displays Similar or Enhanced OncolysisCompared to Ad5-�24RGDWe compared Ad5/3-�24 to Ad5-�24RGD on SKOV3.ip1cells. We analyzed oncolysis 5, 9, and 13 days after mono-layers were infected by 1 and 100 vp/cell of Ad5/3-�24and Ad5-�24RGD; we determined cell survival by MTSassay and expressed it as a fraction of the uninfected mockvalue at each time point (Fig. 6A). At the lower dose,
FIG. 4. Therapeutic effect of Ad5/3-�24 in an animal model of peritoneally disseminated ovarian cancer. SKOV3.ip1 cells were injected ip into SCID mice andadvanced carcinomatosis was allowed to develop for 10 days. (A) The mice received a single ip injection of 3 � 107 vp of Ad5/3-�24, Ad5-�24E3, AdCMVHSV-TKor no virus. (B) Intraperitoneal injection with 1 � 108 vp daily on 3 consecutive days (days 10, 11, and 12). The median survival was not reached with Ad5/3-�24in either experiment by day 135. In both A and B, the overall survival of mice treated with Ad5/3-�24 was statistically significant in comparison to other groups(P � 0.0001).
ARTICLEdoi:10.1016/S1525-0016(03)00200-4
453MOLECULAR THERAPY Vol. 8, No. 3, September 2003Copyright © The American Society of Gene Therapy

Ad5-�24RGD did not cause significant cell killing, whilethe percentage of viable cells remaining with Ad5/3-�24was 76, 12, and 1.7% on days 5, 9, and 13 after infection,respectively, compared to uninfected wells. At the higherviral dose, the viability with Ad5/3-�24 was 17, 0.3, and0.5%, while with Ad5-�24RGD these percentages were 92,35, and 1.2%, respectively. At 1 vp/cell dose, the differ-ence was significant at every time point (P � 0.0048, P �0.0001, and P � 0.0001 for days 5, 9, and 13, respectively).When we compared the higher doses, statistically signif-icant differences were found at the earlier time points (P �0.0001, P � 0.0022) but not on day 13 (P � 0.2652).
We also evaluated oncolytic efficacy on two humanprimary ovarian cancer patient samples (Fig. 6B). We in-fected spheroids and measured cell viability at two time
points and compared it to that of uninfected cells. Withpatient sample A, cell viability was 71 and 59% for Ad5/3-�24 and 60 and 47% for Ad5-�24RGD on days 9 and 13,respectively. There was no statistical significance (P �0.4022 and P � 0.5312, respectively). On patient sampleB, Ad5/3-�24 showed earlier cell killing than Ad5-�24RGD, and the viabilities were 66% versus 84% (P �0.0020) on day 8. However, on day 14 the oncolysis wascomparable (cell viability of 62 and 59% for Ad5/3-�24and Ad5-�24RGD, respectively, P � 0.0904).
DISCUSSION
Despite exciting preclinical data, cancer gene therapy ap-proaches utilizing various strategies have yet to display
FIG. 5. Noninvasive assessmentof Ad5/3-�24 efficacy in vivo.SKOV3-luc cells were injectedip into SCID mice (day 0) andadvanced carcinomatosis wasallowed to develop. The micereceived a single ip injection of3 � 107 vp of Ad5/3-�24 or novirus on day 8. Biolumines-cence was imaged weekly start-ing on day 7. Location andmagnitude of light captured byoptical CCD imaging after ipinjection of D-luciferin on day21, Ad5/3-�24 treated (A), novirus (B). A pseudocolor imagerepresenting light intensity(blue, least intense, and red,most intense) and gray-scalereference images were super-imposed. (C) Signal from theentire abdominal region ofeach mouse were quantified,and the mean photon countsare shown. SKOV3-luc ovariancancer cells express firefly lucif-erase, which reacts with D-lu-ciferin in a bioluminescence re-action. Therefore, reduction insignal indicates killing of cancercells by Ad5/3-�24. Bars indi-cate SD. *Imaging performedafter removal of the abdominalwall.
ARTICLE doi:10.1016/S1525-0016(03)00200-4
454 MOLECULAR THERAPY Vol. 8, No. 3, September 2003Copyright © The American Society of Gene Therapy

definitive clinical breakthroughs. In general, this resulthas been attributed to insufficient transduction of tumorcells [12]. An important factor reducing transduction ef-ficacy with adenoviruses may have been variable expres-sion of the primary receptor, CAR, on human primarycancer cells [3,5–9]. Consequently, efforts to enhance in-fectivity or retarget Ad5 have been endeavored. We dem-onstrated recently that swapping the primary receptorbinding knob domain for a serotype 3 knob allows in-creased transgene expression in the most stringent avail-able preclinical ovarian cancer substrates, purified humanprimary cancer cells, and in an animal model. This 5/3chimeric virus binds to the distinct Ad3 receptor. In fact,Ad3 pseudotyping was superior to RGD-4C modification[6]. Also, with regard to preclinical evaluation of safety,we showed that the murine biodistribution, liver toxicity,and blood clearance of an Ad5/3 chimera were not signif-icantly different from those of Ad5 [6]. As Kupffer cell uptakeof adenovirus in liver is not CAR mediated, it is not surpris-ing that liver uptake of chimeric and other retargeted viruseswould not differ from the parental virus.
Replication-competent agents such as CRAds haveemerged as a tool for overcoming low tumor transduc-tion. Importantly, they have been tested in a number ofclinical trials, demonstrating excellent safety with somepreliminary evidence of efficacy. In a phase II study of anintratumorally administered E1B 55K-deleted CRAd in 40patients with head and neck cancer, three complete andtwo partial responses were reported [24]. Further, CRAdshave been administered ip, iv, intra-arterially, and intra-tumorally in large doses without significant toxicity [25].
The oncolytic potency of replicating agents is deter-
mined directly by the capability of infecting target cells[19,20]. Consequently, infectivity-enhanced CRAds havebeen constructed, and in preclinical studies, their onco-lytic potency has been increased, when additional cellularreceptors have been used [23]. Ad5-�24RGD features anRGD-4C modification of the HI loop of the knob, whichallows binding to �v integrins, which are regularly ex-pressed and often overexpressed on ovarian cancer cells[26] and tumor vasculature [27]. In the context of ovariancancer, Ad5-�24RGD demonstrated impressive oncolyticpotential and significantly increased survival in an animalmodel [22]. Consequently, clinical trials utilizing this vi-rus for ovarian cancer and glioma are in development.Nevertheless, Ad5-�24RGD continues to bind CAR and istherefore not retargeted but infectivity enhanced instead.
In this study, we used fiber chimerism as a retargetingstrategy. The Ad5/3-�24 fiber features the knob from Ad3,and this chimerism results also in enhanced infectivity,which translated into increased oncolysis of target cells(Fig. 2). While the isogenic agent without the fiber mod-ification displayed minimal or no cell killing, Ad5/3-�24achieved dose-dependent cell lysis. Further, this virus wasable to replicate in ovarian cancer primary cell spheroidsas measured with quantitative PCR (Fig. 3). The in vitroassay used here is probably not very sensitive to the effectsof infectivity enhancement, as in vitro all viruses presentin the supernatant are expected to enter cells. Neverthe-less, Ad5/3-�24 was superior to control viruses in all pa-tient samples. Therefore, the novel CRAd effectively rep-licates in primary cancer cells and the replication kineticsare not adversely affected by the genetic fiber modifica-tion.
FIG. 6. The oncolytic efficacy of Ad5/3-�24 is comparable or superior to that of Ad5-�24RGD. (A) SKOV3.ip1 monolayers were infected with 0, 1, or 100 vp/cellof Ad5/3-�24 or Ad5-�24RGD (isogenic control virus with an RGD-4C capsid modification). Cell viability was measured with MTS assay on days 5, 9, and 13.The OD490 values of uninfected cells were set as 100% at each time point. Data are expressed as means � SD of quadruplicate experiments. Oncolysis wassignificantly improved with Ad5/3-�24 compared to Ad5-�24RGD (on day 13, 100 vp/cell, P � 0.2652; all other P � 0.005). (B) Human primary ovarianadenocarcinoma cells were purified and cultured as spheroids, and CRAd infections were performed. Cell killing was analyzed with MTS assay. With patientsample A, Ad5/3-�24 displayed faster oncolysis (P � 0.0020 on day 8).
ARTICLEdoi:10.1016/S1525-0016(03)00200-4
455MOLECULAR THERAPY Vol. 8, No. 3, September 2003Copyright © The American Society of Gene Therapy

All ovarian cancer gene therapy trials published so farhave relied on ip administration of the agent to patientswith peritoneally disseminated disease [4]. To mimic aclinical situation, we inoculated advanced ip carcinoma-tosis, followed by ip administration of viruses (Fig. 4).With both doses, Ad5/3-�24 demonstrated significantlyimproved survival. In fact, with a single injection, Ad5/3-�24 allowed 80% survival on day 114, when all the con-trol mice were dead. Direct comparisons with other stud-ies utilizing this model are difficult due to differences inexperimental setup. Nevertheless, other studies reportsmaller cell inoculum and earlier or more numerous in-jections of virus [22,28,29]. Therefore, the results ob-tained here could compare favorably to other reports.Interestingly, the survival with Ad5/3-�24 using singleinjection versus multiple injections did not differ signifi-cantly. However, the larger dose of Ad5-�24E3 seemed toresult in less antitumor efficacy in addition to givingincreased toxicity (Fig. 4B). This might be due to livertoxicity, as we have experienced same phenomenon witha wild-type Ad5 (A. Kanerva, work in progress).
There are possible obstacles to ip administration ofCRAds. First, most ovarian cancer patients have malig-nant ascites fluid, which has neutralizing anti-adenovirusantibodies [26,30]. These antibodies could hamper theinitial infection of tumor cells and spreading of new viri-ons. Fortunately, previous studies have suggested partialescape of Ad5/3 chimeric viruses from preexisting neutral-izing anti-adenovirus antibodies, perhaps due to the chi-meric nature of the fiber [6]. Another possible problemcould be related to local toxicity, specifically, replicationin the peritoneal lining. An interesting finding in murinebiodistribution studies was a lower peritoneal uptake ofAd5/3 [6]. This could be beneficial in the context of ipadministration of Ad5/3-�24. Converted weight/weightinto humans, the smaller dose used here would equal ca.9 � 1010 vp. This is well below the 2 � 1012 vp daily for5 consecutive days used in a CRAd trial in which themaximal tolerated dose was not reached [31]. In vectortrials up to 7.5 � 1013 vp have been administered daily for5 consecutive days, without dose-limiting toxicity [32].
Imaging techniques can provide fundamental safetyand efficacy information on experimental therapy ap-proaches [33]. Specifically, utilizing an orthotopic animalmodel for monitoring CRAd efficacy is advantageous as itmay resemble the clinical picture more closely than sctumors, but this approach is also problematic as tumorsare not accessible to diameter measurements. Further,another important feature of noninvasive imaging is thepossibility of performing repeated measurements. There-fore, various noninvasive imaging systems have beenevaluated [33]. For example, expression of somatostatinreceptor subtype 2, coded by an adenovirus vector, can beimaged with radioisotope gamma camera after adminis-tration of the somatostatin analogue 99mTc-P2045 [29].Also, optical charge-coupled device (CCD) imaging has
been used to detect bioluminescence emitted from D-luciferin reacting with firefly luciferase, coded by an ade-novirus vector [34]. Other approaches include magneticresonance and positron-emission tomography imaging ofpositron-emitting ligands [33]. Cancer cells expressing re-porter genes such as firefly luciferase and green fluores-cence protein are useful means of following tumor growth[35,36].
We used an orthotopic ovarian cancer model ofSKOV3-luc cells, which emit light after ip administrationof D-luciferin. Therefore, oncolytic killing of tumor cellscorresponds with reduction of signal in comparison tocontrol animals. Kinetics of tumor growth with or with-out virus injection was followed weekly. Ad5/3-�24-treated mice initially responded, but then relapsed with sctumors in the needle tract, an intrinsic defect of thisanimal model [22] (Fig. 5C). This has the potential forconfounding ip imaging data as even small sc tumors cangive a strong signal due to their proximity to the camera.Nevertheless, the sensitivity of the assays was sufficient toallow detection of ip cell killing.
A further round of imaging was performed without theabdominal wall to distinguish the antitumor without theconfounding sc tumors, and we found that light emittedfrom the peritoneal cavity was only 4% of the untreatedgroup, suggesting effective killing of tumor cells by thevirus. Of note, the survival of the untreated mice wasbetter than in Fig. 4. This might be due to the moremalignant version of SKOV3 cells, i.e., SKOV3.ip1, in thesurvival experiment [37]. Interestingly, we could not findsign of viable ip tumor tissue in the treated mice in thesurvival experiment, though there was some signal left inthe imaging. The susceptibility of the cells to the treat-ment agent could be different, or their viability during theinjection procedure could be different. Also, it is possiblethat SKOV3-luc cells allow less virus production thanSKOV3.ip1 cells, due to concomitant luciferase produc-tion. Further, although genetically similar in theory, dif-ferent sets of mice were used and could contribute to theresults.
We have previously reported promising results with an-other CRAd (Ad5-�24RGD) for ovarian cancer gene therapy[22]. While it is possible that a number of CRAds could beuseful for treatment of patients with advanced disease, itmay be of interest to obtain comparative preclinical efficacydata. Therefore, we compared Ad5-�24RGD to Ad5/3-�24 incell lines, in clinical tumor specimens, and in an orthotopicmurine model of peritoneally disseminated ovarian cancer.Preliminary evidence suggests superiority of the latter ineach of the test systems.
Nevertheless, there are a number of applications inwhich it would be important to have effective oncolyticagents with distinct capsids. For example, if neutralizingantibodies prove to be limiting to clinical efficacy, “seros-witching” could be a useful way to retain antitumor effi-cacy. Preliminary studies have indicated that modifica-
ARTICLE doi:10.1016/S1525-0016(03)00200-4
456 MOLECULAR THERAPY Vol. 8, No. 3, September 2003Copyright © The American Society of Gene Therapy

tion of the fiber knob is sufficient to allow escape frompreexisting neutralizing antibodies [6,30]. Further, se-quential utilization of a number of alternative bindingroutes (CAR, integrins, serotype 3 receptor) could be help-ful in avoiding selection-mediated resistance to treat-ment. This could be tested by randomizing animals (orpatients) into treatment arms receiving either Ad5/3-�24or sequentially alternating Ad5/3-�24, Ad5-�24RGD orAd5-�24E3, all of which have antitumor efficacy but fea-ture distinct capsids and entry routes.
In conclusion, we have demonstrated that retargetinga selectively oncolytic adenovirus to the Ad3 receptorresults in improved infectivity of ovarian cancer cells andovercomes the CAR deficiency on primary cancer cells.Consequently, therapeutic efficacy was dramatically in-creased in vitro and in vivo. Further, we have demonstratedthat Ad5/3-�24 allows cell killing comparable or superiorto previously described Ad5-�24RGD. Thus, Ad5/3-�24could be an effective agent for treatment of ovarian can-cer and other tumors with an inactive Rb/p16 pathwayand high expression of the Ad3 receptor. Clinical trialswill ultimately show if preclinical advances such as re-ported here can be translated in to similar progress incancer gene therapy.
MATERIALS AND METHODSCell culture. 293 cells were purchased from Microbix (Toronto, Canada)while 911 were courtesy of Dr. van der Eb (University of Leiden, TheNetherlands). Lung adenocarcinoma cell line A549 and ovarian adenocar-cinoma cell lines ES-2 and OV-3 were obtained from ATCC (Manassas, VA).Ovarian adenocarcinoma cell lines SKOV3.ip1, Hey, and OV-4 were ob-tained from Dr. Price, Dr. Wolf (both M. D. Anderson Cancer Center,Houston, TX), and Dr. Eberlein (Harvard Medical School, Boston, MA).Firefly luciferase-expressing ovarian adenocarcinoma cell line SKOV3-lucwas kindly provided by Dr. Negrin (Stanford Medical School, Stanford,CA). All cell lines were cultured under recommended conditions.
Primary ovarian adenocarcinoma cells were purified by a previously de-scribed immunomagnetic-based method [38] from malignant ascites fluidsamples from patients undergoing a procedure for ovarian cancer at theUniversity of Alabama at Birmingham Hospital. Briefly, ovarian cancer cellswere bound with a murine anti-TAG-72 antibody and then collected withmagnetic beads coated with anti-mouse IgG. To create three-dimensionalspheroids, cells were suspended in growth medium in 3% agar-coated flasksand incubated overnight at 37°C with 5% CO2 on a rocker [39].
Construction of Ad5/3-�24. A fiber shuttle vector, pNEB.PK.F5/3, contain-ing an Ad5 tail and shaft and Ad3 knob was digested with PacI and KpnI,followed by cotransformation into Escherichia coli for homologous recom-bination with a SwaI-linearized rescue plasmid containing an E1 deletion.This resulted in a plasmid with the E1-deleted Ad5 genome with thechimeric fiber domain (pTU.5/3) [40]. To create Ad5/3-�24, a shuttlevector containing a 24-bp deletion in CR2 of E1A (pShuttle�24) was used[41]. pShuttle�24 was linearized with PacI/PmeI and cotransfected into 911cells with PacI-linearized pTU.5/3 to rescue Ad5/3-�24 (Fig. 1). Propaga-tion was performed on A549 cells.
The presence of the 24-bp deletion in E1A and the absence of thewild-type E1A were confirmed with PCR (�24L1, 5-GTCCGGTTTCTAT-GCCAAAC-3; �24R1, 5-TCACCCTCTTCATCCTCGTC-3). The presenceof the E3 region and the Ad3 knob in the fiber was analyzed withPCR (E3L2, 5-CCTGAAACACCTGGTCCACT-3; E3R2, 5-GCCACACT-TAGGGCTTCTGA-3; knob3(S), 5-GCACAGGTGCCATTACAGTAG-3;knob3(AS), 5-TCATTATGTGTTCCCGCATTA-3).
Other adenoviruses. Ad5-�24E3 was created by homologous recombina-tion between pShuttle�24 linearized by PmeI/EcoRI and PacI-linearizedpTG3602, resulting in an otherwise wild-type Ad5 virus but with the 24-bpdeletion in E1A [41]. The presence of a 24-bp deletion and the E3 regionwas confirmed as above. AdCMVHSV-TK is an E1/E3-deleted adenovirusexpressing herpes simplex virus thymidine kinase [42]. The absence ofwild-type E1 was confirmed with PCR (CRAdITR(S), 5-GATAAT-GAGGGGGTGGAGTTTGTG-3; CRAdWt(AS), 5-GAAAACTCTACTCGCT-GGCACTCA-3). Ad5-�24RGD [21] has been described previously.
All viruses were purified on cesium chloride gradients. The vp concen-tration was determined at 260 nm, and standard plaque assay on 293 cellswas performed to determine infectious particles. The ratio of vp/infectiousparticles was 4.7, 3.2, 9.8, and 50 for Ad5/3-�24, Ad5-�24E3, AdCMVHSV-TK, and Ad5-�24RGD, respectively.
In vitro cytotoxicity assay. Cells in quadruplicate were infected for 1 h at37°C in 50 �l of growth medium with 2% FBS. Thereafter, cells wereincubated with 5% growth medium. For Fig. 2, cell viability was measuredusing the CellTiter 96 AQueous One Solution Cell Proliferation Assay (MTSassay; Promega, Madison, WI) on day 6 (SKOV3.ip1, OV-3), day 10 (Hey,ES-2), or day 13 (OV-4). The results with the Ad5/3-�24 group werecompared to those of the other groups using two-tailed t test (SAS v.8.2,SAS Institute, Cary, NY). For Fig. 6, SKOV3.ip1 monolayers were infectedwith no virus, Ad5/3-�24 (1 and 100 vp/cell) or Ad5-�24RGD (1 and 100vp/cell). On days 5, 9, and 13, cell viability was measured as above.Primary ovarian adenocarcinoma cells were purified and cultured as sphe-roids overnight. After overnight incubation, spheroids were infected with1 vp/cell (patient A) or 10,000 vp/cell (patient B) of Ad5/3-�24 or Ad5-�24RGD or no virus in 2% growth medium for 1 h at 37°C [39]. Then, FBSwas added in cell suspension to achieve 5% concentration, and the sphe-roids were divided into aliquots of 105 cells in Costar 96-well ultralowattachment plates (Corning, Inc., Corning, NY). MTS assay was performedon days 8 and 13 (Patient A) or days 9 and 14 (Patient B). All cell viabilityresults are expressed as percentage of viable cells compared to uninfectedcontrol at each time point.
Quantitating virus replication. Primary ovarian adenocarcinoma cell sphe-roids were cultured as above. Next day, spheroids were infected with 1000vp/surface cell of Ad5/3-�24, Ad5-�24E3, AdCMVHSV-TK, or no virus asabove [39]. After 1 h infection, FBS was added in to achieve 10% concentra-tion, and aliquots of 105 cells were seeded in Costar 96-well ultralow attach-ment plates. Cells and growth medium were harvested and frozen at 1⁄2 , 1, 2,4, 6, 8, 12, and 16 days after infection. To quantitate the total E4 copynumber, DNA was purified from spheroid suspension (cellular and growthmedium fractions together) using a DNeasy Tissue Kit (Qiagen, Valencia,CA). The primers used for amplifying the E4 were forward, 5-GGAGT-GCGCCGAGACAAC-3, and reverse, 5-ACTACGTCCGGCGTTCCAT-3,and detected with the probe 5-TGGCATGACACTACGACCAACAC-GATCT-3. Human -actin was amplified to control for the amount of cellsas described [26]. The background values (uninfected spheroids) weresubtracted at each time point. To estimate total virus production by thespheroids, cumulative virus copy number was calculated.
Animals. Mice were obtained at 3–4 weeks age and quarantined for 2weeks. Mice were kept under pathogen-free conditions according to theAmerican Association for Accreditation of Laboratory Animal Care guide-lines. Animal protocols were reviewed and approved by the InstitutionalAnimal Care and Use Committee of UAB.
Therapeutic ovarian cancer model. Female CB17 SCID mice (University ofAlabama at Birmingham CFAR SCID Mouse Core Facility) were injected ipwith 1 � 107 SKOV3.ip1 cells on day 0. On day 10 mice were injected ip with3 � 107 vp of Ad5/3-�24 (n � 10 mice), Ad5-�24E3 (n � 10), or AdCM-VHSV-TK (n � 9) in 1 ml of Opti-MEM (Mediatech, Herndon, VA). In anotherexperiment, mice were injected ip on days 10, 11, and 12 with 1 � 108 vp ofAd5/3-�24 (n � 10 mice), Ad5-�24E3 (n � 10), AdCMVHSV-TK (n � 9), or novirus (n � 9) in 1 ml of Opti-MEM. Mice were followed daily and killed whenthere was any evidence of pain or distress. Survival data were plotted on aKaplan–Meier curve, and the Ad5/3-�24 group was compared with the other
ARTICLEdoi:10.1016/S1525-0016(03)00200-4
457MOLECULAR THERAPY Vol. 8, No. 3, September 2003Copyright © The American Society of Gene Therapy

groups with log-rank analysis and �2 testing (LIFETEST procedure in SAS v8.2,the Weibull model of distribution).
Bioluminescence imaging to detect in vivo luciferase expression. The iptumors were established by injecting 107 SKOV3-luc cells ip into femaleCB17 SCID mice (Charles River Laboratories, Wilmington, MA; n �4/group) on day 0. On day 8, 3 � 107 vp of Ad5/3-�24 or no virus wasinjected ip in a 1-ml volume of Opti-MEM. Imaging was performed beforetreatment on day 7 and then weekly until untreated mice showed signs ofdistress (day 43). On day 43, images were also collected after removing theabdominal wall that contained sc tumors. Briefly, 150 mg/kg D-luciferinwas injected ip, and mice were anesthetized with enflurane gas anesthesiaand placed in a light-tight chamber. The photographic (gray-scale) refer-ence image was obtained at 15 min after D-luciferin injection, and thebioluminescent image was collected immediately thereafter. Images wereobtained with a CCD cooled to �120°C, using the IVIS Imaging System(Xenogen Corp., Alameda, CA), with the field of view set at 25 cm height.The photographic images used a 0.2-s exposure, 8 f/stop, 2 binning (res-olution), and open filter. The bioluminescent images used exposures rang-ing from 0.5 to 10 s, 1 f/stop, 2 binning, and open filter. The biolumines-cent and gray-scale images were overlaid using LivingImage software(Xenogen Corp.). Igor image analyses software (Wavemetrics, Lake Os-wego, OR) was also used to obtain a pseudocolor image representingbioluminescence intensity (blue, least intense, and red, most intense).Regions of interest were drawn around the ip tumors, and the total counts(photons) were summed in the entire tumor areas. The total counts werenormalized to image acquisition time.
ACKNOWLEDGMENTS
This work was supported by the NIH (R01 CA94084, R01 CA83821, P50CA83591), Lustgarten Foundation, Susan B. Komen Foundation, Deutsche For-schungsgemeinschaft (BA2076/1-2), Sigrid Juselius Foundation, Emil AaltonenFoundation, Maud Kuistila Foundation, Finnish Medical Foundation, Academyof Finland, Finnish Cancer Society, Biocentrum Helsinki, and University ofHelsinki Internal Funds.
RECEIVED FOR PUBLICATION APRIL 9, 2003; ACCEPTED MAY 25, 2003.
REFERENCES1. Vorburger, S. A., and Hunt, K. K. (2002). Adenoviral gene therapy. Oncologist 7: 46–59.2. Sandmair, A. M., et al. (2000). Thymidine kinase gene therapy for human malignant
glioma, using replication-deficient retroviruses or adenoviruses. Hum. Gene Ther. 11:2197–2205.
3. Bauerschmitz, G. J., Barker, S. D., and Hemminki, A. (2002). Adenoviral gene therapyfor cancer: From vectors to targeted and replication competent agents. Int. J. Oncol. 21:1161–1174.
4. Hemminki, A., and Alvarez, R. D. (2002). Adenoviruses in oncology: A viable option?BioDrugs 16: 77–87.
5. Kanerva, A., et al. (2002). Targeting adenovirus to the serotype 3 receptor increasesgene transfer efficiency to ovarian cancer cells. Clin. Cancer Res. 8: 275–280.
6. Kanerva, A., et al. (2002). Gene transfer to ovarian cancer versus normal tissues withfiber-modified adenoviruses. Mol. Ther. 5: 695–704.
7. Miller, C. R., et al. (1998). Differential susceptibility of primary and established humanglioma cells to adenovirus infection: Targeting via the epidermal growth factor receptorachieves fiber receptor-independent gene transfer. Cancer Res. 58: 5738–5748.
8. Cripe, T. P., et al. (2001). Fiber knob modifications overcome low, heterogeneousexpression of the coxsackievirus–adenovirus receptor that limits adenovirus gene trans-fer and oncolysis for human rhabdomyosarcoma cells. Cancer Res. 61: 2951–2960.
9. Hemmi, S., Geertsen, R., Mezzacasa, A., Peter, I., and Dummer, R. (1998). The presenceof human coxsackievirus and adenovirus receptor is associated with efficient adenovi-rus-mediated transgene expression in human melanoma cell cultures. Hum. Gene Ther.9: 2363–2373.
10. Anders, M., et al. (2002). Expression and regulation of the coxsackie-and-adenovirusreceptor (CAR) in premalignant lesions and advanced neoplasms. Mol. Ther. 5: S214.
11. Stevenson, S. C., Rollence, M., White, B., Weaver, L., and McClelland, A. (1995).Human adenovirus serotypes 3 and 5 bind to two different cellular receptors via thefiber head domain. J. Virol. 69: 2850–2857.
12. Kirn, D., Martuza, R. L., and Zwiebel, J. (2001). Replication-selective virotherapy forcancer: Biological principles, risk management and future directions. Nat. Med. 7:781–787.
13. Khuri, F. R., et al. (2000). A controlled trial of intratumoral ONYX-015, a selectively-
replicating adenovirus, in combination with cisplatin and 5-fluorouracil in patients withrecurrent head and neck cancer. Nat. Med. 6: 879–885.
14. Bischoff, J. R., et al. (1996). An adenovirus mutant that replicates selectively in p53-deficient human tumor cells. Science 274: 373–376.
15. Fueyo, J., et al. (2000). A mutant oncolytic adenovirus targeting the Rb pathwayproduces anti-glioma effect in vivo. Oncogene 19: 2–12.
16. Heise, C., et al. (2000). An adenovirus E1A mutant that demonstrates potent andselective systemic anti-tumoral efficacy. Nat. Med. 6: 1134–1139.
17. Heise, C., Sampson-Johannes, A., Williams, A., McCormick, F., Von Hoff, D. D., andKirn, D. H. (1997). ONYX-015, an E1B gene-attenuated adenovirus, causes tumor-specific cytolysis and antitumoral efficacy that can be augmented by standard chemo-therapeutic agents. Nat. Med. 3: 639–645.
18. Sherr, C. J. (1996). Cancer cell cycles. Science 274: 1672–1677.19. Douglas, J. T., Kim, M., Sumerel, L. A., Carey, D. E., and Curiel, D. T. (2001). Efficient
oncolysis by a replicating adenovirus (ad) in vivo is critically dependent on tumorexpression of primary ad receptors. Cancer Res. 61: 813–817.
20. Hemminki, A., Dmitriev, I., Liu, B., Desmond, R. A., Alemany, R., and Curiel, D. T.(2001). Targeting oncolytic adenoviral agents to the epidermal growth factor pathwaywith a secretory fusion molecule. Cancer Res. 61: 6377–6381.
21. Suzuki, K., Fueyo, J., Krasnykh, V., Reynolds, P. N., Curiel, D. T., and Alemany, R.(2001). A conditionally replicative adenovirus with enhanced infectivity shows im-proved oncolytic potency. Clin. Cancer Res. 7: 120–126.
22. Bauerschmitz, G. J., et al. (2002). Treatment of ovarian cancer with a tropism modifiedoncolytic adenovirus. Cancer Res. 62: 1266–1270.
23. Shinoura, N., et al. (1999). Highly augmented cytopathic effect of a fiber-mutantE1B-defective adenovirus for gene therapy of gliomas. Cancer Res. 59: 3411–3416.
24. Nemunaitis, J., et al. (2001). Phase II trial of intratumoral administration of ONYX-015,a replication-selective adenovirus, in patients with refractory head and neck cancer.J. Clin. Oncol. 19: 289–298.
25. Kirn, D. (2001). Clinical research results with dl1520 (Onyx-015), a replication-selectiveadenovirus for the treatment of cancer: What have we learned? Gene Ther. 8: 89–98.
26. Hemminki, A., et al. (2001). An adenovirus with enhanced infectivity mediates molec-ular chemotherapy of ovarian cancer cells and allows imaging of gene expression. Mol.Ther. 4: 223–231.
27. Arap, W., Pasqualini, R., and Ruoslahti, E. (1998). Cancer treatment by targeted drugdelivery to tumor vasculature in a mouse model. Science 279: 377–380.
28. Peng, K.-W., TenEyck, C. J., Galanis, E., Kalli, K. R., Hartmann, L. C., and Russell, S. J.(2002). Intraperitoneal therapy of ovarian cancer using an engineered measles virus.Cancer Res. 62: 4656–4662.
29. Hemminki, A., et al. (2002). In vivo molecular chemotherapy and noninvasive imagingwith an infectivity-enhanced adenovirus. J. Natl. Cancer Inst. 94: 741–749.
30. Hemminki, A., Wang, M., Desmond, R. A., Strong, T. V., Alvarez, R. D., and Curiel, D. T.(2002). Serum and ascites neutralizing antibodies in ovarian cancer patients treatedwith intraperitoneal adenoviral gene therapy. Hum. Gene Ther. 13: 1505–1514.
31. Vasey, P. A., et al. (2002). Phase I trial of intraperitoneal injection of the E1B-55-kD-gene-deleted adenovirus ONYX-015 (dl1520) given on days 1 through 5 every 3 weeks inpatients with recurrent/refractory epithelial ovarian cancer. J. Clin. Oncol. 20: 1562–1569.
32. Buller, R. E., et al. (2002). A phase I/II trial of rAd/p53 (SCH 58500) gene replacementin recurrent ovarian cancer. Cancer Gene Ther. 9: 553–566.
33. Gambhir, S. S., Barrio, J. R., Herschman, H. R., and Phelps, M. E. (1999). Assays fornoninvasive imaging of reporter gene expression. Nucl. Med. Biol. 26: 481–490.
34. Adams, J. Y., et al. (2002). Visualization of advanced human prostate cancer lesions inliving mice by a targeted gene transfer vector and optical imaging. Nat. Med. 8:891–897.
35. Rehemtulla, A., et al. (2000). Rapid and quantitative assessment of cancer treatmentresponse using in vivo bioluminescence imaging. Neoplasia 2: 491–495.
36. Nyati, M. K., et al. (2002). The potential of 5-fluorocytosine/cytosine deaminaseenzyme prodrug gene therapy in an intrahepatic colon cancer model. Gene Ther. 9:844–849.
37. Yu, D., Wolf, J. K., Scanlon, M., Price, J. E., and Hung, M. C. (1993). Enhancedc-erbB-2/neu expression in human ovarian cancer cells correlates with more severemalignancy that can be suppressed by E1A. Cancer Res. 53: 891–898.
38. Barker, S. D., et al. (2001). An immunomagnetic-based method for the purification ofovarian cancer cells from patient-derived ascites. Gynecol. Oncol. 82: 57–63.
39. Lam, J. T., et al. (2003). Replication of an integrin targeted conditionally replicatingadenovirus on primary ovarian cancer spheroids. Cancer Gene Ther. 10: 377–387.
40. Uil, T. G., et al. (2003). Generation of an adenoviral vector containing an addition of aheterologous ligand to the serotype 3 fiber knob. Cancer Gene Ther. 10: 121–124.
41. Suzuki, K., Alemany, R., Yamamoto, M., and Curiel, D. T. (2002). The presence of theadenovirus E3 region improves the oncolytic potency of conditionally replicative ad-enoviruses. Clin. Cancer Res. 8: 3348–3359.
42. Rosenfeld, M. E., Feng, M., Michael, S. I., Siegal, G. P., Alvarez, R. D., and Curiel, D. T.(1995). Adenoviral-mediated delivery of the herpes simplex virus thymidine kinasegene selectively sensitizes human ovarian carcinoma cells to ganciclovir. Clin. CancerRes. 1: 1571–1580.
ARTICLE doi:10.1016/S1525-0016(03)00200-4
458 MOLECULAR THERAPY Vol. 8, No. 3, September 2003Copyright © The American Society of Gene Therapy




















