
Function of the coxsackie and adenovirus receptor (CAR) in ...
Upload
independentCategory
view
5download
0
Chromosoma (Berl) (1986) 93:537-544 CHROMOSOMA �9 Springer-Verlag 1986
Human cellular sequences detectable with adenovirus probes I. Evidence for novel repeat sequences and a possible Eia-like cellular "gene"
Antony W. Braithwaite ~, William P. Diver, Sue Le Jeune, Felice Driver, and Hiroto Naora Molecular Biology Unit, Research School of Biological Sciences, Australian National University, GPO BOX 475, Canberra, ACT 2601, Australia; 1 addressee for reprint requests
Abstract. Previous studies suggesting homology between human cellular DNA and the DNAs from adenovirus types 2 and 5 are extended in the present paper. A clone (ChAdh), isolated from a human genomic DNA library us- ing an adenovirus probe, hybridized to discrete regions of adenovirus 2 DNA, including part of the transforming genes Ela and Elb, as well as to repeated sequences within human DNA. The Ela and Elb genes both hybridize to the same 300 base pair Sau3AI fragment within ChAd h al- though there is no obvious homology between Ela and E1b. The Ad 2 Ela gene was also used as a probe to screen other cellular DNAs to determine whether repeated se- quences detectable with Ad 2 DNA probes were conserved over long evolutionary periods. Hybridization was detected to the genomes of man, rat, mouse and fruit fly, but not to those of yeast and bacteria. In addition to a "smear" hybridization, discrete fragments were detected in both ro- dent and fruit fly DNAs. The experiments reported suggest the existence of two different types of cellular sequences detected by Ad 2 DNA: (1) repeated sequences conserved in a variety of eukaryote genomes and (2) a possible unique sequence detected with an Ela probe different from that responsible for hybridization to repeated sequences. This unique sequence was detected as an EcoRI fragment in mouse DNA and had a molecular size of about 8.8 kb.
Introduction
Nucleotide or amino acid sequence similarities between dif- ferent fragments of DNA or proteins have been used exten- sively to search for similar genes in different organisms. Amongst other things this approach has led to the discovery of cellular nucleotide sequences which have homology with retroviral oncogenes. In some cases this homology is exten- sive (for a review see Bishop 1983).
The transforming genes of human adenoviruses, known as Ela and Elb, also appear to have homology with nucleo- tide sequences from apparently normal human cells (Jones et al. 1979; Arrand et al. 1983; Braithwaite et al. 1984). Ra- dioactively labelled probes prepared from total human cel- lular RNA have been found to hybridize to parts of these transforming genes (Jones et al. 1979; Arrand et al. 1983) as do "high sequence complexity" probes prepared from total cellular DNA (Braithwaite et al. 1984) which hybridize preferentially to repeated nucleotide sequences (Peden et al.
1982). The above results suggest that human adenovirus DNA and human cellular DNA contain one or more nucle- otide sequences in common, and that these sequences are repeated in cellular DNA and transcribed.
The hybridization data presented in the above three re- ports all suggest that the adenoviral/cellular DNA/RNA or DNA/DNA hybrids are thermally stable. Arrand et al. (1983) have reported about 7% mismatch and in approxi- mate agreement with this, our results (Braithwaite et al. 1984) indicate less than 15% mismatch.
In this paper, we report experiments on the characteriza- tion of a genomic DNA clone isolated from a human gene library using adenovirus type 2 (Ad 2) DNA clones as ra- dioactively labelled probes. We show that this clone con- tains a repeated sequence(s) and that it hybridizes to the transforming genes of Ad 2 DNA. We show also that, when the common repeat sequence from the cloned Ela gene o fAd 2 is deleted and used as a probe, a single DNA EcoRI restriction fragment is detected in mouse DNA suggesting that there is an Ela-like "gene" in normal cells.
Materials and methods
Preparation of DNAs. Purified Ad 2 DNA was purchased from the Sigma Chemical Co. (D 3390) and Ad 5 DNA was prepared as previously described (Braithwaite et al. 1981). Celo DNA was a gift from Dr. P. Li and fruit fly (Drosophila melanogaster, Canton-S strain) DNA was a gift from Dr. R. Russell. Other genomic DNAs were extracted by phenol and chloroform after pronase digestions (1.0 rag/ ml) from I 3 days (for details see Braithwaite et al. 1981). Plasmid DNAs were grown and purified using the method of Betlach et al. (1976).
Isolation of specific DNA fragments. DNAs from which fragments were to be isolated were digested with the appro- priate restriction enzyme then electrophoresed in low melt- ,ing point agarose (Sigma Type VII). The desired DNA frag- ment was excised from the gel by slicing. The DNA-agarose composite was melted (at 65 ~ C) then extracted with phenol and chloroform to remove contaminating agarose. This procedure yields pure DNA fragments capable of being in vitro labelled and ligated.
Restriction endonuclease digestions and electrophoresis. De- tails of restriction digestions and electrophoresis are as pre-

538
viously described (Braithwaite et al. 1984) but with minor modifications. Digestions with EcoRI (New England Bio- labs or Pharmacia) were carried out in the presence of 100 mM NaC1 and 100 mM Tris-HC1, pH 7.4, to avoid in- correct cleavage (EcoRI*) which was sometimes noticed under lower ionic strength and 50 mM NaC1 (data to be presented elsewhere). All electrophoresis was through 1% agarose gels.
Preparation of radioactive probes and hybridization. This was as described previously (Braithwaite et al. 1984) but without prior digestion of template D N A with HaeIII . After incubation the reaction mixture was twice precipitated with 2 M ammonium acetate, 70% ethanol and 50 gg yeast t R N A to collect the incorporated fraction (generally around 50%-70% incorporation of added 32p_c~_dATP)" The probe was dissolved, denatured by boiling and used immediately for hybridization.
Filter hybridization. Hybridizations were carried out at 65 ~ C as previously described (Braithwaite et al. 1984) ex- cept that Escherichia coli D N A was substituted for salmon sperm in Figure 6 (M, D, R). Also, for the experiment in Figure 8 hybridization was at 58~ and the " h o t wash" was at 50 ~ C in 2 x SSC (1 x SSC=0.15 M NaC1, 0.015 M Na3 citrate) and 0.1% sodium dodecyl sulphate.
Construction of a human genomic (DNA) library. The hu- man D N A sequence library was constructed by ligation of Sau3AI-cleaved human placental (male) D N A fragments in the range of 10-16 kb on to the arms of BamHI-cleaved 2Charon 30 DNA. Preparation of DNAs, ligation, and in vitro packaging of bacteriophage clones were done essen- tially as described in Maniatis et al. (1978).
Results
Adenovirus DNA hybridizes to "'multiple-copy" human DNA sequences
D N A from uninfected Hela cells and from Hela cells in- fected with Ad 5 was digested with HindII I and the resul- tant fragments separated by electrophoresis through an agarose gel. After transfer to nitrocellulose these DNAs were hybridized to a low specific activity (105 dpm/gg), 3 zp_ labelled Ad 5 (genome length) D N A probe. This experiment shows (Fig. 1 A) that, as expected, Ad 5 D N A hybridizes to adenovirus D N A replicated in the infected cells which is seen as a series of discrete HindII I restriction fragments (lanes 1, 2). Also, this probe detects multiple size classes of cellular D N A from uninfected Hela cells (lane 3). Both large and small Hela cell D N A fragments are detected (lane 3), the largest fragments being larger than the Ad 5 HindIII A fragment (7.96 kb, lane 1). However, no large "cell" D N A fragments are detected in lane 1. Why this is so we are not sure. It is possible that the presence of highly abundant adenovirus D N A molecules in this lane might saturate the probe and so prevent detection of the cellular D N A repeat sequences.
In a second experiment a higher specific activity (2 • 10 7 dpm/gg) genome length Ad 2 probe was hybridized to three human placental D N A preparations digested with EcoRI. With 4 days of exposure to X-ray film the Ad 2 probe was again found to hybridize to human D N A frag-
Fig. 1 A, B. Adenovirus type 2 and 5 DNAs hybridize to repeated sequences in human DNA. 5 10 gg of DNA were digested with restriction endonucleases, electrophoresed in an agarose gel, and transferred to nitrocellulose membrane filters (Southern 1975). A DNAs were HindIII digested and hybridized with 32p-labelled Ad 5 DNA. Lanes: 1 DNA from Ad 5 infected Hela cells; 2 1/10 (about 0.5 gg) of preparation in 1 ; 3 DNA from uninfected Hela cells. Sizes of some Ad 5 specific fragments are indicated. B DNAs were EcoRI digested and hybridized with 32p-labelled Ad 2 DNA. Lanes: 1, 2 human male placental DNAs; 3 human female placen- tal DNA. HindlII-digested 2DNA was used as a size marker. The results in B show clear evidence of hybridization to multiple size classes of DNA, forming a "smear". Shorter exposures however produced no better resolution of the hybridization than presented here
ments essentially as a smear (Fig. IB). Male and female placental DNAs were compared, and in this experiment a minor sex difference was evident. Both male D N A s (lanes 1 and 2) showed a band migrating in front of the smear, which was not apparent in the female D N A (lane 3). This difference between male and female D N A s is not gen- eral and most likely results from a transfer artifact. There are thus no sex differences between human male and female DNAs in terms of their ability to hybridize with an adeno- virus probe. These experiments suggest that Ad 5 and Ad 2 DNAs, which are known to be > 80% homologous (Tooze 1980), contain a stretch of nucleotides which is also found in human D N A as a repeat sequence. These results formed the basis of our original report (Braithwaite et al. 1984) and of this paper.
Computer comparisons of the "adenovirus-human" DNA hybridizing regions with known repeat sequences
Previous results showed that there were three regions on the Ad 2 D N A molecule that hybridized with a human re- petitive D N A probe. These regions are contained within the Ad 2 early genes Ela, Elb and E4 (Braithwaite et al. t984). The Ela and Elb regions showed the strongest hy-

539
bridization and were therefore defined more precisely. The Ela region, located between the XbaI site at 3.8% and the end of the insert at 4.4% in pE la (Braithwaite et al. 1984), is a sequence of about 200 base pairs (bp) at the 3' end of the Ela gene, and within the coding region.
It was shown previously (Braithwaite et al. 1984 and in Fig. 3) that the Elb hybridizing region remained attached to the pBR322 vector of p E l b - f after cutting with PstI. This occurred despite the fact that the method of isolation, homopolymer tailing of a restriction fragment into the PstI site of pBR322 (Nevins 1981), should have allowed com- plete excision of the Elb region by PstI. Restriction enzyme analysis of this plasmid (unpublished results) suggests that the Elb hybridizing region is a sequence of about 150 bp at the 3' end of the Elb gene.
A detailed computer comparison of the Ela and Elb hybridizing regions showed little significant homology be- tween these genes. This suggested prima facie that Ela and Elb were detecting different repeated sequences in the hu- man genome. To determine exactly which human DNA sequences the adenoviral genes were detecting by hybridiza- tion, both the Ela and Elb hybridizing regions were used to search through the current (1985) GENBANK data base. The human DNA repeat sequences contained in this data base comprise e(105 copies/haploid genome; Weiss et al. 1983); Alu (10 s copies/haploid genome; Schmid and Jelinek 1982); HinfI (102 copies/haploid genome; Shimizu et al. J983; KpnI (Digiovanni et al. 1983); HindIII (Manuelidis 1982) and two unidentified non-Alu repeats (Deininger et al. 1981). Additionally, the Ela and Elb regions were aligned with the non-Alu repeats described by Sun et al. (1984) namely O, K and poly(CA). None of the listed repeat sequences showed significant homology with the Ela or Elb regions. Finally, the conserved sex chromosome asso- ciated GATA repeat, described by Singh et al. (1980), was found by hybridization to have no sequence homology to Ad 2 DNA (data not shown). Taken together, these data suggest that the Ela and Elb hybridizing regions detect one or more as yet undescribed repeat sequences in human DNA.
Isolation of human genomic DNA clones showing homology with Ad 2 DNA
Since little information was gained by computer alignment studies, we attempted to clone the putative repeat sequence. A recombinant library of human male placental DNA was constructed using the vector 2 Charon 30 as outlined in Ma- terials and methods. The library contained mostly small inserts (<10 kb) and was under-represented in low copy number sequences (unpublished observations). This library was screened with a probe prepared from a mixture of Ad 2 DNA clones (pEla, p E l b - f and pEco C containing the E4 gene in pBR322; see Braithwaite et al. 1984) which were known to contain the regions which hybridised with human DNA. More than 100 positive clones were found after an initial hybridization to about 105 plaques. Following a sec- ond screen, the DNAs from ten clones were analysed by Southern blotting and hybridization and four clones which hybridized strongly were selected for further study.
The results of an experiment in which these clones were hybridized to 32p-labelled Ad 2 DNA (Fig. 2) shows that an Ad 2 probe detects a single 3.9 kb EcoRI fragment which is identical in all clones (clone 4 DNA was not shown in
Fig. 2. Human genomic DNA clones contain sequences which hy- bridize to adenovirus DNA. 2Charon 30 DNA and 3 clones were digested with EcoRI. After electrophoresis and transfer to nitrocel- lulose they were hybridized with 3/P-labelled Ad 2 DNA. The Ad 2 probe detects a single 3.9 kb EcoRI DNA fragment in all 3 clones. The other EcoRI fragments from these clones migrated with EcoRI-cut vector DNA. Location of HindIlI-digested DNA markers is indicated
Fig. 2). As the library was amplified, these four clones are likely to be siblings. No hybridization was observed to any other EcoRI fragment, all of which belong to the vector.
To determine whether the library screening had detected the "adenoviral-human" repeat sequence, a parallel experi- ment was carried out. Clone 1 (see Fig. 2), hereafter referred to as ChAd h, and h u m a n " repetitive" probes were prepared and hybridized to Ad 2/5 DNA clones containing Ela (pEla), the 3' half of Elb (pElb-f), and pE2, containing the Ad 5 E2A gene, (see Braithwaite et al. 1984). The results of this experiment (Fig. 3) show that the human '~ tive" and ChAd h probes both have an identical hybridiza- tion pattern, which detects sequences in Ela and Elb, but not in E2A, as reported previously for the repetitive probe (Braithwaite et al. 1984). Differences in hybridization inten- sity are due to differences in specific activity. This experi- ment thus confirms that we have cloned at least part of the human repeat sequences that hybridize with discrete regions of Ad 2 and Ad 5 DNAs.
Restriction endonuclease mapping and partial characterization of ChAd h
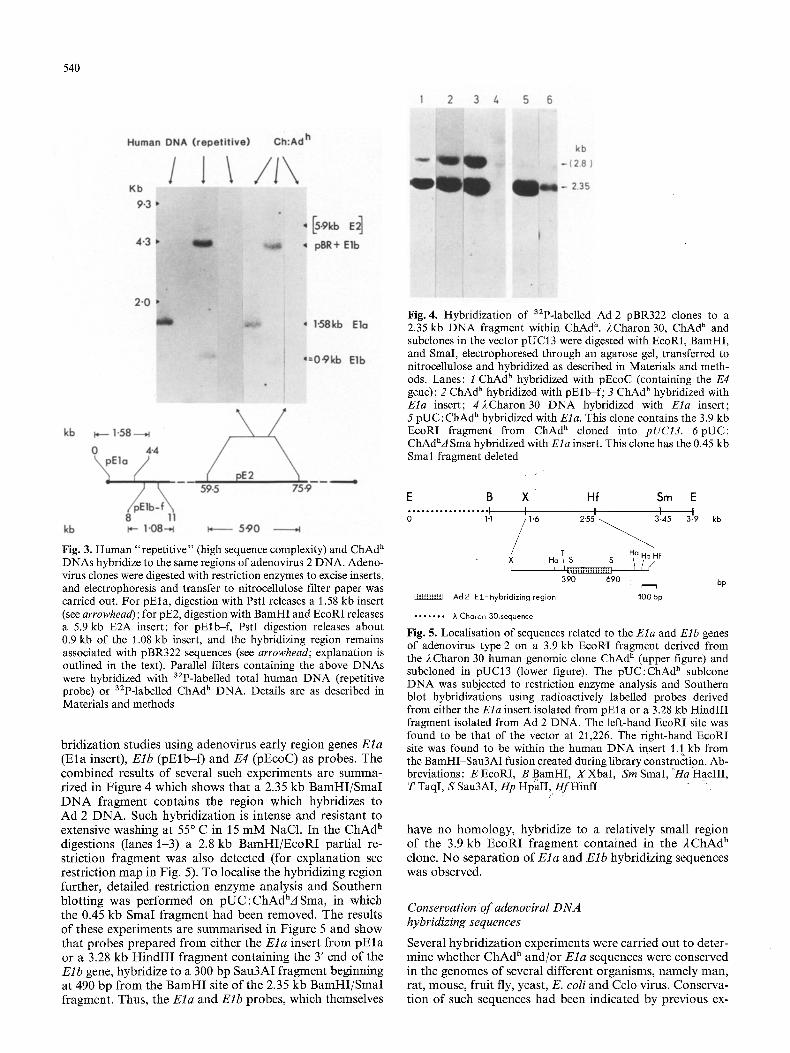
Preliminary characterization of ChAd h and two subclones in the vector pUCI3 was possible by restriction enzyme analysis with EcoRI, BamHI and Sinai, combined with hy-
540
Fig. 4. Hybridization of s2p-labelled Ad 2 pBR322 clones to a 2.35 kb DNA fragment within ChAd h. 2Charon 30, ChAd h and subclones in the vector pUC13 were digested with EcoRI, BamHI, and SmaI, electrophoresed through an agarose gel, transferred to nitrocellulose and hybridized as described in Materials and meth- ods. Lanes: 1 ChAd h hybridized with pEcoC (containing the E4 gene); 2 ChAd h hybridized with pElb-f; 3 ChAd h hybridized with Ela insert; 42Charon30 DNA hybridized with Ela insert; 5 pUC:ChAd h hybridized with Ela. This clone contains the 3.9 kb EcoRI fragment from ChAd ~ cloned into pUC13. 6pUC: ChAdhA Sma hybridized with Ela insert. This clone has the 0.45 kb Smal fragment deleted
Fig. 3. Human "repetitive" (high sequence complexity) and ChAd h DNAs hybridize to the same regions of adenovirus 2 DNA. Adeno- virus clones were digested with restriction enzymes to excise inserts, and electrophoresis and transfer to nitrocellulose filter paper was carried out. For pEla, digestion with PstI releases a 1.58 kb insert (see arrowhead) ; for pE2, digestion with BamHI and EcoRI releases a 5.9 kb E2A insert; for pElb-f, PstI digestion releases about 0.9 kb of the 1.08 kb insert, and the hybridizing region remains associated with pBR322 sequences (see arrowhead; explanation is outlined in the text). Parallel filters containing the above DNAs were hybridized with 32p-labelled total human DNA (repetitive probe) or 32p-labelled ChAd h DNA. Details are as described in Materials and methods
bridization studies using adenovirus early region genes E l a (Ela insert), E l b (pElb- f ) and E4 (pEcoC) as probes. The combined results of several such experiments are summa- rized in Figure 4 which shows that a 2.35 kb BamHI/SmaI D N A fragment contains the region which hybridizes to Ad 2 DNA. Such hybridization is intense and resistant to extensive washing at 55 ~ C in 15 mM NaC1. In the ChAd h digestions (lanes 1-3) a 2.8 kb BamHI/EcoRI partial re- striction fragment was also detected (for explanation see restriction map in Fig. 5). To localise the hybridizing region further, detailed restriction enzyme analysis and Southern blotting was performed on pUC: ChAdhA Sma, in which the 0.45 kb SmaI fragment had been removed. The results of these experiments are summarised in Figure 5 and show that probes prepared from either the E l a insert from p E l a or a 3.28 kb HindII I fragment containing the 3' end of the Elb gene, hybridize to a 300 bp Sau3AI fragment beginning at 490 bp from the BamHI site of the 2.35 kb BamHI/SmaI fragment. Thus, the E l a and Elb probes, which themselves
E B X Hf Sm E . . . . . . . . . . . . . . . . . . I I I l I 0 1"1 / 1 "6 2 " 5 5 ~ 3 . 4 5 3"9 kb
/ T Ha w~ la
X Ha S S \ i ' W 'f I / I'iiii:iiiiiiiiii.I / / r
390 690 bp
IIHH!,L'*HI Ad2 El-hybridizingreglon lO0bp
. . . . . . . X Charon 30~sequence
Fig. 5. Localisation of sequences related to the Ela and Elb genes of adenovirus type 2 on a 3.9 kb EcoRI fragment derived from the 2Charon 30 human genomic clone ChAd h (upper figure) and subcloned in pUC13 (lower figure). The pUC:ChAd h sublcone DNA was subjected to restriction enzyme analysis and Southern blot hybridizations using radioactively labelled probes derived from either the Ela insert isolated from pEla O r a 3.28 kb HindIII fragment isolated from Ad 2 DNA. The left-hand EcoRI site was found to be that of the vector at 21,226. The right-hand EcoRI site was found to be within the human DNA insert 1A kb from the BamHI-Sau3AI fusion created during library construction. Ab- breviations: E EcoRI, B BamHI, X XbaI, Sm SmaI, H a ~aeIII, T TaqI, S Sau3AI, Hp HpaII, Hf Hinfl
have no homology, hybridize to a relatively small region of the 3.9 kb EcoRI fragment contained in the 2ChAd h clone. No separation of E l a and Elb hybridizing sequences was observed.
Conservation o f adenoviral D N A hybridizing sequences
Several hybridization experiments were carried out to deter- mine whether ChAd ~ and/or E l a sequences were conserved in the genomes of several different organisms, namely man, rat, mouse, fruit fly, yeast, E. coli and Celo virus. Conserva- tion of such sequences had been indicated by previous ex-

Fig. 6. Conservation of Ela hybridizing sequences in the genomes of higher eukaryotes. 5 10 lag of cellular DNAs were digested with EcoRI and subjected to electrophoresis and transfer to nitrocellu- lose. Human (H) DNA was hybridized to 32p-labelted pUC: ChAd h and the other DNAs were hybridized to azp-labelled Ela. Mouse (M), rat (R) and Drosophila (D) DNAs were electrophoresed on the same get and hybridized to the same probe. HindIII-digested 2DNA was used as a size marker
periments (man and chicken; Braithwaite et al. 1984; Ar- rand et al. 1983).
Human DNA. EcoRI-digested human placental DNA was hybridized with the subclone pUC : ChAdhA Sma. The result from this experiment (Fig. 6H) is indistinguishable from results previously obtained with probes prepared from Ad 5, Ad 2 and the adenovirus clones. Such a result shows that the sequences present in ChAd h are repeated to high copy number in human DNA. This cloned repeat sequence might be the sequence we screened for with Ad 2 probes, or a possible alternative interpretation is that ChAd h con- tains a repeat sequence(s) other than that which was se- lected during the original plaque screening. For this reason, the conservation experiments we describe in this section were done with probes prepared from the adenovirus Ela gene. Nevertheless, preliminary data did indicate that ChAd h probes do detect multiple-copy sequences in differ- ent genomes (data not shown).
Under normal conditions, which contained salmon sperm DNA as a non-specific competitor, Ela hybridiza- tions to DNAs of different species were often not reproduc- ible and generally weak. These results suggested that Ela- hybridizing sequences might be present at a much lower copy number, It was observed that hybridization was in part blocked by similar sequences in salmon sperm DNA
541
during pre-hybridization. When E. coli DNA was used in the hybridizing solutions instead of salmon sperm DNA, clear and reproducible results were obtained.
Rat DNA. EcoRI-digested rat liver DNA (Wistar Outbred) was hybridized to a probe prepared from Ela DNA (from pEla). Strong hybridization was detected to many DNA size classes (Fig. 6R) and some hybridizing bands were quite discrete. These discrete bands had molecular sizes of 1.7 kb to 19 kb, with two bands around 3.9 kb and 7.6 kb. These two latter hybridizing bands might partly correspond to those reported by Frolova and Georgiev (1979) when Ad 5 DNA was hybridized to normal rat DNA. In their experiments, an Ad 5 probe detected a cluster of poorly resolved DNA fragments migrating between 4 and 8 kb.
Mouse DNA. EcoRI-digested DNA from C57/BI.6J mouse embryo fibroblasts was hybridized to Ela (Fig. 6M). The result was similar to rat DNA in that some discrete DNA fragments were detected by Ela, together with a weak smear. The hybridization intensity overall is much weaker than for rat.
Fruit f ly DNA. D. melanogaster DNA was also examined for the presence of Ela-like sequences. Results showed a unique sequence of about 4.0 kb and a faint background smear (Fig. 6 D).
The hybridization results for rodent and fruit fly DNAs suggest that an Ela probe detects low-copy sequences of greater homology as well as repeated sequences in these DNAs (see below).
Yeast DNA, E. coli DNA. Yeast (Saccharomyces cerevisiae) DNA and E. coli DNA (from RR1 cells) were examined for hybridization to Ela and pUC:ChAd h DNAs prepared as 32P-labelled probes. No hybridization was observed to these DNAs (data not shown).
Chicken embryo lethal orphan (Celo) virus DNA. Celo virus is an avian adenovirus (Laver et al. 1971), the DNA of which has very little sequence homology with human adeno- viruses. A recent report has shown two regions of homology between Celo and Ad 2 DNA which are between 12% and 20% and between 41% and 49% of Celo DNA (Alestr6m et al. 1982). These are detected under high stringency. We hybridized 32p-labelled ChAO to EcoRI- and HindIII-di- gested Celo DNA (Fig. 7; lanes 3, 4) and found that ChAO sequences are contained within the HindIII (B) fragment, being 61.7% to 77.8% on the Celo DNA map. This region is outside the homologous Ad 2 and Celo sequences referred to above.
Detection of a possible Ela-like "gene" in mouse DNA
Results from the previous section showed that for rodent and fruit fly DNAs an Ela probe strongly detects discrete DNA restriction fragments and hybridization smearing is weak. This is in contrast to human DNA where smearing is prominent, and discrete fragments are not detectable. Discrete fragments detected in this manner represent re- gions of either greater abundance (clustered repeats) or greater homology. If the latter is the case, the discrete hy- bridizing sequences might represent a putative Ela-like

542
Fig. 7. Conservation of ChAd h sequences in the genome of a chicken adenovirus (Celo). Celo DNA was digested with EcoRI (lane 1) and HindIII (lane 2). After electrophoresis and transfer these DNAs were hybridized to 3zP-labelled ChAd h. Hybridization detected the EcoRI (A; lane 3) and the HindIII (B; lane4) frag- ments of Celo DNA, which are indicated by arrows. The positions of these fragments on the Celo genome map are shown. Restriction map is as outlined in Alestr6m et al. (1982). Abbreviations: E EcoRI, H HindIII, A EcoRI hybridizing fragment; B HindIII hy- bridizing fragment. The indicated numbers are given in % genetic map units
Fig. 8. Detection of an Ela-like sequence in mouse DNA. 5 gg of mouse DNA was digested with EcoRI, electrophoresed through agarose and transferred to nitrocellulose paper. Mouse DNAs were hybridized with Ela DNA in which the 3" XbaI fragment from pEla (1,336 bp to the end of pEla) was deleted. Lanes I and 2 represent results from two separate experiments, indicating that an 8.8 kb EcoRI fragment was reproducibly detected in mouse DNA. The arrows indicated by 9.4 kb and 6.6 kb represent posi- tions of 2HindIII markers
"gene" or "gene family". We know from previous experi- ments that the bulk of the homology between human geno- mic DNA and adenovirus DNA is located between the XbaI site at 1,336 on the Ad 2 map and the end of the Ela gene contained within the pEla clone (see Fig. 3 and Braithwaite et al. 1984). If this region of the viral E l a gene is deleted and a probe prepared from the remaining Ela DNA, then a positive hybridization result to genomic DNA
would indicate a region of greater homology rather than greater abundance. In Figure 8 the results of two indepen- dent experiments are shown in which EcoRI-digested mouse DNAs were hybridized with Ela probes prepared from Ela DNA deleted for the sequences Y to the above XbaI site. These hybridizations weakly detect a single DNA fragment of about 8.8 kb (lane 1) and a similar result in lane 2, but with a little smearing. This 8.8 kb fragment was also ob- served when the complete Ela gene was used as a p r o b e (Fig. 6M). This result shows that the 8.8 kb fragment hy- bridises to another region of Ela and having regard for the above argument, the 8.8 kb fragment has greater homol- ogy with Ela DNA. A little smearing was observed in one experiment (lane 2) which suggests we may not have deleted the entire repetitive hybridizing region, or the Xba-deleted El a probe is still contaminated with part of the repeat hy- bridizing region. We are currently examining mouse DNA for discrete hybridizing sequences using probes prepared from different regions of the Ela gene.
Preliminary results indicate similar discrete "Ela- l ike" sequences in human DNA when the 3' end deletion Ela is used as a probe (data not shown).
Discussion
The results described above can be summarized as follows: (1) A human genomic clone (ChAd h) isolated from a
2Charon 30 library showed strong hybridization to the Ela and L i b genes of Ad 2 and to multiple size classes (as a smear) of human DNAs digested with EcoR1. ChAd h thus contains at least some of the putative repeated human DNA sequence which is detectable with Ad 2 DNA probes.
(2) ChAd h was characterized by restriction endonucle- ase mapping and hybridization experiments (Figs. 4, 5), and the region detected by Ad 2 probes was localized to a 300 bp Sau3AI fragment (see Fig. 5).
(3) Mouse (and probably human) DNA contains a dis- crete EcoRI fragment detected with part of the Ad 2 Ela gene which does not hybridize to repeated sequences. This might represent an Ela-like sequence.
We are currently sequencing the Sau3AI fragment to determine exactly the nature of the human repetitive se- quence contained in ChAd h. On the basis of our results so far, which do not show any homology with adenovirus DNA, and from a number of other lines of evidence we predict that the sequence will be unusual. Firstly, this Sau3AI fragment hybridizes to portions of the Ela and Elb genes (see Results) and the E4 gene of Ad 2 DNA, yet there is no obvious nucleotide "consensus" sequence among these three genes (determined by computer sequence alignment analysis). In addition, radioactive probes pre- pared from parts of the El a gene do not hybridize to any other Ad 2 gene apart from El a under the usual stringency conditions (data not shown). Secondly, the Sau3AI frag- ment hybridizes to part of the Celo virus genome (see Fig. 7) but outside the regions that have homology with Ad 2 DNA. Thirdly, Arrand et al. (1983) have shown that tran- scripts from normal human lymphoid and fibroblastic cells hybridize to regions of Ad 2 DNA (group C adenoviruses), Ad 3 DNA (group B) and Ad 12 DNA (groupA) (see Norrby et al. 1976), yet these viruses show remarkable nu- cleotide sequence variation (Green 1970; Garon et at. 1973). Furthermore, comparison of two hybridizing regions of Ad 2 DNA (EcoRI-F fragment) and Ad 12 DNA (HindIII-

543
I fragment) failed to reveal any sequence homology (Arrand et al. 1983). Four th ly , similar " repe t i t ious" hybr idizat ion between cellular R N A s and D N A with the Epstein-Barr virus genome (Arrand et al. 1983; Peden et al. 1982) has been repor ted and, once again, the hybridizing regions show no homology sufficient to account for the hybr idizat ion data.
One possible explanat ion of our data, and that of others (see references cited above), is that the hybr idizat ion is due to G + C bonding alone. We do not suppor t this interpreta- t ion for reasons that are largely detailed by Ar rand et al. (1983) and because the E2A genes of A d 2 and A d 5 which do not hybridize to ChAd h (see Results) or high sequence complexity human D N A probes (Braithwaite et al. 1984) are about 65% G + C rich.
We do not know the function o f the repetitive sequences contained within ChAd h, but as pointed out previously (Ar- rand et al. 1983) it is of interest that the D N A sequences a round the adenovirus D N A integrat ion sites contain some repeated sequences (see for example Sambrook et al. 1979; Stabel and Doerfler 1982). We note that the adenovirus genes that become stably integrated are the t ransforming Ela and Elb genes, the same genes involved in the hybrid- ization repor ted in this paper. The E4 gene also integrates in some cases (Sambrook et al. 1974) and this gene too hybridizes though less strongly with a high sequence com- plexity human D N A probe (Braithwaite et al. 1984). It will be interesting to compare the sequence present in the Sau3AI f ragment with known repeated sequences near adenovirus D N A integrat ion sites.
We have also examined whether A d 2 viral " r e p e a t " sequences are conserved in a variety of eukaryote and pro- karyote genomes. Ela was used as a probe in these experi- ments to avoid the problem of ChAd h containing other repeat sequences besides those selected for with A d 2 probes. The results of these experiments (Fig. 6) showed that Ela did hybridize to several cellular D N A s including those from rodents and the fruit fly. In this connection, several discrete hybridizing fragments were detected in ro- dent DNAs , and a single fragment in fruit fly D N A . The results shown in Figure 8 indicate that the A d 2 Ela-hybrid- izing sequence(s) present in these discrete fragments is dif- ferent from those responsible for hybr idizat ion with re- peated sequences. This discrete region, the da ta argue, must have a greater degree o f homology with Ela than the repeat sequences and thus suggest the presence of an "Ela-like gene" in mouse D N A . This must remain a tentative conclu- sion until more detailed investigations have been carried out, but the observat ion is interesting because it provides the first evidence (albeit prel iminary) for a cellular ho- molgue of an oncogene from a D N A turnout virus. I f this were the case it suggests a mechanism through which adeno- virus Ela gene products are able to modify the growth cycle propert ies of cells (Braithwaite et al. 1983 ; Braithwaite 1986; Bellett et al. 1985), namely the putat ive Ela homo- logue may be a cell cycle regulatory gene which A d 2 Ela proteins are able to mimic. A cellular Ela-like gene has also been suggested recently by others, based on indirect experiments (Kao et al. 1985).
Acknowledgements. We thank Dr. P. Li for Celt DNA, Dr. R. Russell for Drosophila DNA and Dr. J. Nevins for the Ad 2/5 clones. This work was partly supported by a grant from the A.C.T. Cancer Society.
References
Alestrtm P, Stenlund A, Li P, Bellett A, Petterson U (1982) Se- quence homology between avian and human adenoviruses. J Virol 42:306-310
Arrand JR, Walsh-Arrand JE, Rymo L (1983) Cytoplasmic RNA from normal and malignant human cells shows homology to the DNAs of Epstein-Barr virus and human adenovirus. EMBO J 2:1673-1683
Bellett AJD, Li P, David ET, Mackey EJ, Braithwaite AW, Cutt JR (1985) Control functions of adenovirus transformation re- gion E1A gene products in rat and human cells. Mol Cell Biol 5:1933-1939
Betlach MC, Hershfield V, Chow L, Brown W, Goodman HM, Boyer HW (1976) A restriction endonuclease analysis of bacte- rial plasmid controlling the EcoRI restriction and modification of DNA. Fed Proc 35 : 2037-2043
Braithwaite AW, Murray JD, Bellett AJD (1981) Alterations to controls of cellular DNA synthesis by adenovirus infection. J Virol 39 : 331-340
Braithwaite AW, Cheetham BF, Li P, Parish CR, Waldron-Stevens LK, Bellett AJD (1983) Adenovirus-induced alterations of the cell growth cycle: a requirement for expression of Ela but not of Elb. J Virol 45:192-199
Braithwaite AW, Le Jeune S, Naora H (1984) Adenoviruses have homology with a reiterated sequence in genomic DNA. DNA 3 : 223-230
Braithwaite AW (1986) Semipermissive replication of adenovirus 5 in rat brain cells and evidence for an induction of cellular DNA replication in vivo. J Gen Virol 67 : 391-396
Deininger PL, Jolly D J, Rubin CM, Friedmann T, Schmid CW (1981) Base sequence studies of 300 nucleotide renatured re- peated human DNA clones. J Mol Biol 151 : 17-33
Digiovanni L, Haynes SR, Misra R, Jelinek WR (1983) Kpn I family of long-dispersed repeated DNA sequences of man: evi- dence for entry into genomic DNA of DNA copies of poly(CA)- terminated KpnI RNAs. Proc Natl Acad Sci USA 80:6533-6537
Frolova EI, Georgiev GP (1979) The existence of sequence homolo- gies to adenovirus 5 DNA in the genome of normal rat cells. Nucleic Acids Res 7:1419-1428
Garon CF, Berry K, Hierholzer JC, Rose J (1973) Mapping of base sequence heterologies between genomes from different adenovirus serotypes. Virology 54:414-426
Green M (1970) Oncogenic viruses. Annu Rev Biochem 39: 701-756
Jones KW, Kinross J, Maitland N, Norval M (1979) Normal hu- man tissues contain RNA and antigens related to human infec- tious adenovirus type 2. Nature 277:274-279
Kao H-T, Capasso O, Heintz N, Nevins JR (1985) Cell cycle con- trol of the human HSP70 gene: implications for the role of a cellular Ela-like function. Mol Cell Biol 5:628-633
Laver WG, Younghusband HB, Wrigley NG (1971) Purification and properties of chicken embryo lethan orphan virus (an avian adenovirus). Virology 45:598-614
Maniatis T, Hardison RC, Lacy E, Lauer J, O'Connell C, Quon D, Sire GK, Efstratiadis A (1978) The isolation of structural genes from libraries of eucaryotic DNA. Cell 15:687-701
Manuelidis L (1982) Nucleotide sequence definition of a major human repeated DNA, the Hind III 1.9 kb family. Nucleic Acids Res 10:3211 3219
Nevins JR (1981) Mechanism of activation of early viral transcrip- tion by adenovirus Ela product. Cell 26:213-220
Norrby E, Bartha A, Boulanger P, Dreizin RS, Ginsberg HS, Kalter SS, Kawamura H, Rowe WP, Russell WC, Schlesinger RW, Wigand R (1976) Adenoviridae. Intervirology 7:117-125
Peden K, Mounts P, Hayward GS (1982) Homology between mam- malian cell DNA sequences and human herpes virus genomes detected by a hybridization procedure with high-complexity probe. Cell 31:71-80
Sambrook J, Botchan M, Gallimore P, Ozanne B, Petterson U,

544
Williams J, Sharp PA (1974) Viral DNA sequences in cells transformed by Simian Virus 40, Adenovirus type 2 and Adeno- virus type 5. Cold Spring Harbor Symp Quant Biol 39:615-632
Sambrook J, Greene R, Stringer J, Mitchison T, Hu S-L, Botchan M (1979) Analysis of the sites of integration of viral DNA sequences in rat cells transformed by Adenovirus 2 or SV40. Cold Spring Harbor Symp Quant Biol 44: 56%584
Schmid CW, Jelinek WR (1982) The Alu family of dispersed repeti- tive sequences. Science 216:1065-1070
Shimizu Yu, Yoshida K, Ren C-S, Fujinaga K, Rajagopalan S, Chinnandurai G (1983) Hinf family: a novel repeated DNA family of the human genome. Nature 302: 587-590
Singh L, Purdom F, Jones KW (1980) Conserved sex-chromosome- associated nucleotide sequences in Eukaryotes. Cold Spring Harbor Symp Quant Biol 45:805-814
Sun L, Paulson KE, Schmid CW, Kadyk L, Leinwand L (1984) Non-alu family interspersed repeats in human DNA and their transcriptional activity. Nucleic Acids Res 12:2669~690
Southern EM (1975) Detection of specific sequences among DNA fragments separated by gel electrophoresis. J Mol Biol 98:503-517
Stabel S, Doerfler W (1982) Nucleotide sequence at the site of junction between adenovirus type 12 DNA and repetitive ham- ster cell DNA in transformed cell line CLAC1. Nucleic Acids Res 10:8007-8203
Tooze J (ed) (1981) Molecular biology of tumor viruses, DNA tumor viruses, 2nd edition, Part 2, revised. Cold Spring Harbor Laboratory Press NY
Weiss RB, Mineura K, Henderson EE, Duker NJ, Deriel JK (1983) Enzymic detection of uracil in a cloned and sequenced deoxyri- bonucleic acid segment. Biochemistry 22:4501-4507
Received September 9, 1985 / in revised form January 17, i986 Accepted by B. John
Copyright © 2022 FDOKUMEN




















