![[Cool] Gas Chromatography and Lipids](https://static.fdokumen.com/doc/165x107/6325a4b1852a7313b70e98e9/cool-gas-chromatography-and-lipids.jpg)
Enhanced phospholipase B activity and alteration of phospholipids and neutral lipids in...
11
ORIGINAL PAPER Enhanced phospholipase B activity and alteration of phospholipids and neutral lipids in Saccharomyces cerevisiae exposed to N-nitrosonornicotine Panneerselvam Vijayaraj • Jayaraja Sabarirajan • Vasanthi Nachiappan Received: 13 July 2010 / Accepted: 22 October 2010 Ó Springer Science+Business Media B.V. 2010 Abstract A tobacco-specific nitrosamine (TSNA), N-nitrosonornicotine (NNN), is a potent carcinogen present in cigarette smoke, and chronic exposure to it can lead to pulmonary cancer. NNN causes changes in phospholipid metabolism and the mechanism is yet to be elucidated. Exposure of Saccharomyces cerevisiae to 50 lM NNN leads to a substantial decrease in phosphatidylserine (PS) by 63%, phosphatidylcholine (PC) by 42% and phosphatidylethanolamine (PE) by 36% with a concomitant increase in lysophospholipids (LPL) by 25%. The alteration in phospholipid content was dependent on increasing NNN concentration. Reduced phospholipids were accompanied with increased neutral lipid content. Here we report for the first time that NNN exposure, significantly increases phospholipase B (PLB) activity and the preferred substrate is PC, a major phospholipid responsible for a series of metabolic functions. Furthermore, NNN also promotes the alteration of fatty acid (FA) composition; it increases the long chain fatty acid (C18 series) in phospholipids specifically phosphatidylethanolamine (PE) and PS; while on the contrary it increases short chain fatty acids in cardiolipin (CL). NNN mediated degradation of phospholipids is associated with enhanced PLB activity and alteration of phospholipid composition is accompanied with acyl chain remodelling. Understanding the altered phospholipid metabolism produced by NNN exposure is a worth- while pursuit because it will help to understand the toxicity of tobacco smoke. Keywords Tobacco specific nitrosamines Á N-Nitrosonornicotine Á Phospholipid Á Phospholipase B Á Neutral lipid Á Free fatty acid Abbreviations TSNA’s Tobacco specific nitrosamines NNN N-Nitrosonornicotine PC Phosphatidylcholine PE Phosphatidylethanolamine PS Phosphatidylserine PI Phosphatidylinositol PA Phosphatidic acid PG Phosphatidylglycerol CL Cardiolipin LPC Lysophosphatidylcholine LPL Lysophospholipid PLB Phospholipase B PLA 2 Phospholipase A 2 FA Fatty acid MAG Monoacylglycerol DAG Diacylglycerol TAG Triacylglycerol FFA Free fatty acid SE Sterol esters ROS Reactive oxygen species DMSO Dimethyl sulphoxide Present Address: P. Vijayaraj Á J. Sabarirajan Á V. Nachiappan (&) Department of Biochemistry, Bharathidasan University, Tiruchirappalli 24, Tamilnadu, India e-mail: [email protected] 123 Antonie van Leeuwenhoek DOI 10.1007/s10482-010-9526-1
Transcript of Enhanced phospholipase B activity and alteration of phospholipids and neutral lipids in...

ORIGINAL PAPER
Enhanced phospholipase B activity and alterationof phospholipids and neutral lipids in Saccharomycescerevisiae exposed to N-nitrosonornicotine
Panneerselvam Vijayaraj • Jayaraja Sabarirajan •
Vasanthi Nachiappan
Received: 13 July 2010 / Accepted: 22 October 2010
� Springer Science+Business Media B.V. 2010
Abstract A tobacco-specific nitrosamine (TSNA),
N-nitrosonornicotine (NNN), is a potent carcinogen
present in cigarette smoke, and chronic exposure to it
can lead to pulmonary cancer. NNN causes changes in
phospholipid metabolism and the mechanism is yet to
be elucidated. Exposure of Saccharomyces cerevisiae
to 50 lM NNN leads to a substantial decrease in
phosphatidylserine (PS) by 63%, phosphatidylcholine
(PC) by 42% and phosphatidylethanolamine (PE) by
36% with a concomitant increase in lysophospholipids
(LPL) by 25%. The alteration in phospholipid content
was dependent on increasing NNN concentration.
Reduced phospholipids were accompanied with
increased neutral lipid content. Here we report for the
first time that NNN exposure, significantly increases
phospholipase B (PLB) activity and the preferred
substrate is PC, a major phospholipid responsible for a
series of metabolic functions. Furthermore, NNN also
promotes the alteration of fatty acid (FA) composition;
it increases the long chain fatty acid (C18 series) in
phospholipids specifically phosphatidylethanolamine
(PE) and PS; while on the contrary it increases short
chain fatty acids in cardiolipin (CL). NNN mediated
degradation of phospholipids is associated with
enhanced PLB activity and alteration of phospholipid
composition is accompanied with acyl chain
remodelling. Understanding the altered phospholipid
metabolism produced by NNN exposure is a worth-
while pursuit because it will help to understand the
toxicity of tobacco smoke.
Keywords Tobacco specific nitrosamines �N-Nitrosonornicotine � Phospholipid �Phospholipase B � Neutral lipid � Free fatty acid
Abbreviations
TSNA’s Tobacco specific nitrosamines
NNN N-Nitrosonornicotine
PC Phosphatidylcholine
PE Phosphatidylethanolamine
PS Phosphatidylserine
PI Phosphatidylinositol
PA Phosphatidic acid
PG Phosphatidylglycerol
CL Cardiolipin
LPC Lysophosphatidylcholine
LPL Lysophospholipid
PLB Phospholipase B
PLA2 Phospholipase A2
FA Fatty acid
MAG Monoacylglycerol
DAG Diacylglycerol
TAG Triacylglycerol
FFA Free fatty acid
SE Sterol esters
ROS Reactive oxygen species
DMSO Dimethyl sulphoxide
Present Address:P. Vijayaraj � J. Sabarirajan � V. Nachiappan (&)
Department of Biochemistry, Bharathidasan University,
Tiruchirappalli 24, Tamilnadu, India
e-mail: [email protected]
123
Antonie van Leeuwenhoek
DOI 10.1007/s10482-010-9526-1

Introduction
Phospholipids are major components of cellular
membranes that take part in a series of metabolic
events including maintenance of cellular permeability,
regulation of proteins associated with the membrane
and regulation of intracellular signalling by providing
signalling molecules (Yamashita et al. 1997; Carman
and Zeimetz 1996; Exton 1994). The functions of
membrane is inherently linked to the type of phos-
pholipids present in the cell membrane, and their
interactions with each other. Environmentally induced
alterations in phospholipid composition can lead to
changes in membrane integrity, permeability, mem-
brane cell injury and even death (Stenger et al. 2009).
This is especially true with toxic substances because
they easily generate reactive oxygen species (ROS)
which in turn interact with cell membrane (Nachiap-
pan et al. 1994). Cigarette smoke is one of the potent
environmental pollutants, because smoking is highly
addictive and a habit that is difficult to break.
Tobacco-specific nitrosamines (TSNAs) are the main
toxic compounds present in cigarette smoke and
tobacco products (Hecht and Hoffmann 1988).
Specifically N-nitrosonornicotine (NNN) is a potent
pulmonary carcinogen, reported by the International
Agency for Research on Cancer as a human carcinogen
(IARC Monogr 2007) and exerts a wide spectrum of
biological effects on the lung and lung airways,
including oxidative stress, inflammation and DNA
damage (Hecht and Hoffmann 1988; Lin et al. 2010).
Smoking is associated with reduced surfactant phos-
pholipids in the bronchoalveolar lavage particularly
phosphatidylcholine (PC) (Oulton et al. 1991; Finley
and Ladman 1972). NNN primarily targets the lung and
alters the composition of lipid-protein complex specif-
ically in the surfactant and alters its biological functions.
More interestingly interaction of NNN to the membrane
mainly depends on the phospholipid composition and its
fatty acid composition as well (Schuster et al. 1992,
1995; Scott 2004). Carcinogenicity of NNN is well
documented; however impact on lipids particularly
phospholipid metabolism is yet to be elucidated.
The objective of this study was to analyze in detail
the influence of NNN on metabolism of neutral lipids
and phospholipids and to depict the underlying mech-
anisms using yeast as a model system. Assessment of
chemicals and their toxicological data usually requires
a large number of animal species, hence a microbial
system is a useful and an alternative approach for our
objectives. The Saccharomyces cerevisiae, a well
studied eukaryotic model system is used to characterise
the effect of NNN on lipid metabolism. Investigations
were also under taken to study the alterations in the
fatty acid composition of the neutral and phospholip-
ids. Overall results suggest that reduced phospholipid
content is due to activated PLB, resulting in increased
lysophospholipid (LPL) content and also adaptation of
membrane by altered fatty acid composition. Our
studies, depicting the changes in phospholipids and
neutral lipids by NNN, would contribute to a better
understanding of the molecular mechanisms underly-
ing its toxicity.
Materials and methods
Materials
Wild-type strain S. cerevisiae BY4741 [MATa his3D1
leu2D0 met15D0 ura3D0] was procured from Open
Biosystems (AL, USA) and its isogenic plb1D, plb2Dand plb3D were gifted by CIMAP, India. [32P]
orthophosphate (5000 Ci/mmol) was obtained from
Bhabha Atomic Research Centre (Mumbai, India) and
[1-14C] acetate (1000 Ci/mmol) was obtained from
PerkinElmer Life Sciences (USA). Yeast medium,
lipid and fatty acid standards were obtained from
Sigma (India). Silica gel 60F254 TLC plates were
from Merck (India). All chemicals and solvents were
purchased from Sigma unless specifically mentioned.
NNN was obtained from Sigma (India). We performed
purity assessment by liquid chromatography electro-
spray ionization tandem mass spectrometry (LC/ESI-
MS/MS) for NNN and it was about 99% pure (data not
shown). Stock solution was prepared in dimethyl
sulphoxide (DMSO) and stored at -20�C till use.
Maximum DMSO in growth media was 0.05% (v/v), a
concentration that had no detectable effect on yeast
growth (data not shown).
Growth conditions and sensitivity assay
Yeast cells were grown in YPD (1% yeast extract, 2%
peptone, and 2% dextrose) medium (pH 7.0) with
aeration at 30�C. The cells were grown in 5 ml of
YPD medium for 12 h and harvested, transferred to
Antonie van Leeuwenhoek
123

25 ml of YPD media with indicated concentrations of
NNN. Cells were grown at 30�C at 180 rpm and cell
growth was studied by measuring the A600 of the
cultures at frequent time intervals. The sensitivity of
the yeast cells to NNN was observed on solid medium
supplemented by spotting experiments. For this the
cells were serially diluted and spotted on YPD plates
containing 2% agar with or without NNN and
incubated for 48 h at 30�C. The dilutions that are
used are 10-1, 10-2, 10-3, 10-4 and 10-5.
Labelling of yeast lipids
Phospholipid alterations were analysed by [32P] ortho-
phosphate labelling (Ghosh et al. 2008) and neutral
lipids were analysed by [1-14C] acetate labelling
(Rajakumari and Daum 2010). The cells were grown
at 30�C in YPD medium with or without different
concentrations of NNN. To this either 100 lCi of [32P]
orthophosphate (5000 Ci/mmol) or 0.5 lCi of [1-14C]
acetate (1000 Ci/mmol) was added. Cells were grown
for 12 h and harvested by centrifugation. Unused radio
labelled phosphate and acetate were removed by
washing with 2% phosphoric acid and equal numbers
of cells were taken, subjected to lipid extraction.
Lipid extraction and Identification
Total lipids from yeast cells were extracted by the
method of Bligh and Dyer (1959). Briefly, to the cell
pellet 400 ll of methanol and 200 ll of chloroform
were added and vortexed. To this, 400 ll of acidified
water (2% orthophosphoric acid) was added and
vigorously vortexed. Chloroform extracts containing
total lipids were subjected to two-dimensional thin
layer chromatography (TLC) with Silica Gel. In TLC
plates phospholipid separation was done by using the
following solvents, for first dimension chloroform/
methanol/ammonia (65:25:5, v/v) and for the second
dimension chloroform/methanol/acetone/acetic acid/
water (50:10:20:15:5, v/v). Neutral lipids were sep-
arated by using petroleum ether: diethyl ether: acetic
acid (70:30:1, v/v) as the solvent system. Individual
lipids were located by comparing the Rf values of the
unknown with the Rf values of the standard. The
radioactive spots were developed by autoradiography
for 12 h. The spots were scraped from the TLC plate,
and the incorporation was counted in liquid scintil-
lation counter (PerkinElmer Life Sciences, USA).
Fatty acids analysis
Fatty acids of both neutral lipids and phospholipids
were analyzed by gas chromatography. Lipids were
separated by TLC and individual lipids were
extracted from the silica gel with chloroform/meth-
anol (2:1 v/v), and subjected to methanolysis using
BF3/methanol for conversion to methyl esters (Mor-
rison and Smith 1964). Fatty acid methyl esters were
separated by gas chromatography and quantification
was referred to heptadecanoic methyl ester, as an
internal standard.
Preparation of phospholipid substrates
Radioactively labelled phospholipids were prepared
biosynthetically by incubating yeast cells with [32P]
orthophosphate for 12 h, followed by extraction of
lipids and separation of phospholipid by 2D-TLC
(Wagner and Paltauf 1994). Purity was typically
[95%, as checked by TLC. Ethanol-solubilized
phospholipid micelles were prepared by sonication
(Gheriani-Gruszka et al. 1988).
Phospholipase assays
Phospholipase activity was assayed using ethanol-
solubilized [32P] orthophosphate labelled phospho-
lipids. The concentration of PC was 2 mM and the
specific radioactivity was 5000 Ci/mmol (Fisher et al.
1992). The buffer (200 ll) contained 50 mM Tris–
HCl pH 8.0/1 mM-EGTA/10 mM-CaCl2. The reac-
tion was initiated by addition of enzyme sources
(50 lg) from corresponding yeast lysate (wild type
and plbD strains treated with or without NNN) and
incubated at 37�C for 60 min. The assay was also
performed in the presence or absence of calcium. The
reaction were stopped by adding chloroform: meth-
anol: 2% phosphoric acid (1:2:1, v/v) and lipids were
extracted, separated by TLC and quantified. Protein
was determined by the method of Bradford (1976).
Results
Effect of NNN on the lipid composition
Cells were treated with NNN and phospholipid
alterations were monitored. We performed the study
Antonie van Leeuwenhoek
123

with different concentrations of NNN (0–400 lM)
and the incorporation of [32P] orthophosphate into
phospholipids was decreased significantly when
compared with control. Figure 1a depicted the con-
centration dependent reduction of total phospholipid
up to 400 lM of NNN; Even though the maximum
reduction (*57%) was in 400 lM of NNN-treated
cells, when compared to control, it was only *17%
greater than 50 lM of NNN treated (*40% reduc-
tion vs control). Hence 50 lM was chosen for further
studies. The impact of 50 lM NNN on individual
phospholipids were analysed (Fig. 1b). There was a
significant (P \ 0.001) reduction in phosphatidylser-
ine (PS) (63%) when compared to control. Apart
from PS, profound decrease was observed in PC
(42%) and phosphatidylethanolamine (PE) (36%).
This might be as a consequence of PS reduction,
since in yeast PS is a precursor for both PC and PE
(Vance and Steenbergen 2005). Observed reduction
of PG leads to the decreased content in CL (37%)
(Chicco and Sparagna 2006). Reduction of phospho-
lipid was accompanied with a simultaneous increase
in LPL (Fig. 1b). Based on the above observations,
we suggest that NNN significantly (P \ 0.001)
decreases the overall phospholipid content.
Declined phospholipid synthesis suggested that
there might be a rapid metabolic switchover from
phospholipid synthesis to neutral lipid leading to its
accumulation. [14C] acetate incorporation into neutral
lipid was increased approximately 30–50% in NNN
treated cells when compared to control (Fig. 1c).
Cells treated with 50 lM of NNN showed a signif-
icant (P \ 0.001) increase in diacylglycerol (DAG)
(threefold) and an increase in TAG (51%) when
compared to control. Increased DAG content might
be due to enhanced hydrolysis of phospholipid. In
addition, both MAG (51%) and sterol (31%) content
were also increased with NNN when compared to
control cells. Similar trend was observed with sterol
esters (SE) also. There was also a significant
(P \ 0.001) increase in FFA (90%) content in NNN
treated cells.
Labelling of yeast cells in the presence of NNN
clearly showed a significant (P \ 0.001) reduction in
phospholipids and a significant increase (P \ 0.001)
in neutral lipids associated with an increase in LPL
(Fig. 1b) and FFA (Fig. 1c). These results have led us
to investigate the involvement of NNN on the
phospholipid degrading enzyme, PLB.
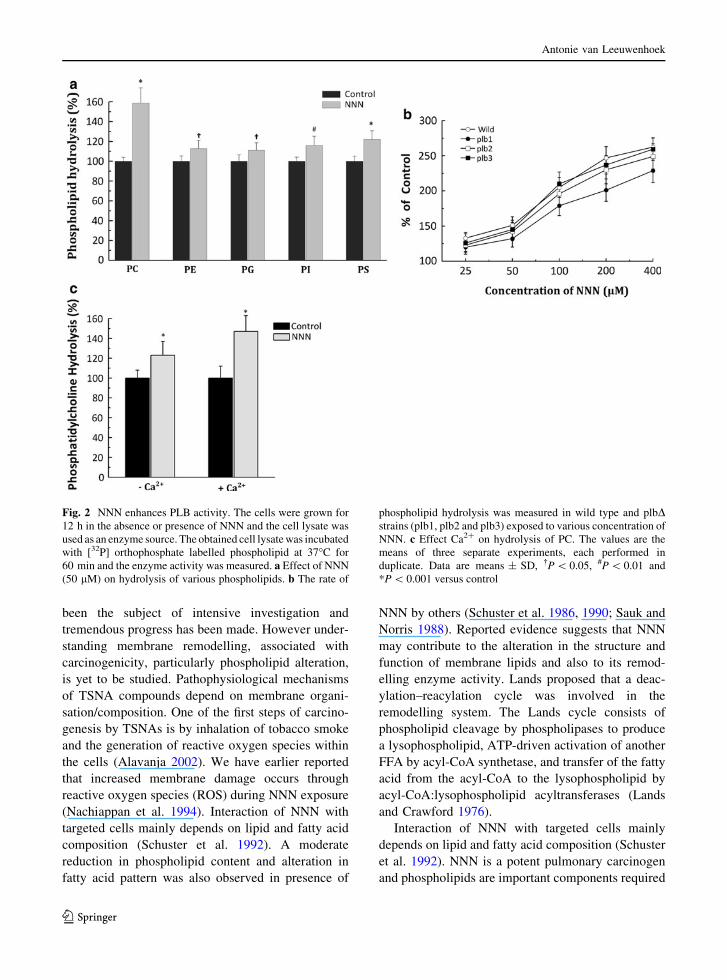
Phospholipase B activity enhanced by NNN
To provide evidence for NNN induced hydrolysis of
phospholipid, we analyzed phospholipase activity using
[32P] orthophosphate labelled phospholipid as substrate
and NNN-treated or untreated yeast lysate as an enzyme
source from both wild type and all three PlbDmaintained uniformity throughout. Figure 2 depicts
the effect of the NNN on hydrolysis of phospholipids by
phospholipase. PC was the preferred substrate and
hydrolysis of PC was substantially (58%) higher with
50 lM NNN treated cells followed by PS (22%) when
compared to other phospholipids (Fig. 2a).
To assess the effect of NNN on hydrolysis of
phospholipid by different PLBs, we used wild type and
three plbD strains (plb1D, plb2D and plb3D) cell lysate
as enzyme source. When the cells were exposed
to NNN, concentration-dependent hydrolysis of
phospholipid was observed in both wild type and
all three mutants (Fig. 2b). The activity was greatly
enhanced by NNN in wild type (*64%), whereas
mutant cells showed diminished activity with
plb1D\ plb2D\ plb3D\ wild type. Both plb2Dand plb3D responded similar to wild type in presence
of NNN and plb1D showed significantly lower enzyme
activity (*16%) as compared to wild type. From these
results we suggest that enhanced phospholipase activ-
ity may be due to plb1 stimulated by NNN exposure.
It is worth assessing the effect of Ca2? on NNN
mediated phospholipid hydrolysis. We performed the
enzyme assay in the presence or absence of Ca2? with
or without NNN (Fig. 2c). NNN enhances both Ca2?
dependent as well as independent phospholipase
activity. The activity was greatly enhanced by the
addition of 10 mM Ca2? (*1.4-fold increase) whereas
in the absence of Ca2? *1.2-fold increase was
observed when compared to their respective controls
(Fig. 2c). Our result suggests that Ca2? partially
activates NNN mediated phospholipid hydrolysis.
Involvement of TSNA’s in phospholipid
molecular species alteration
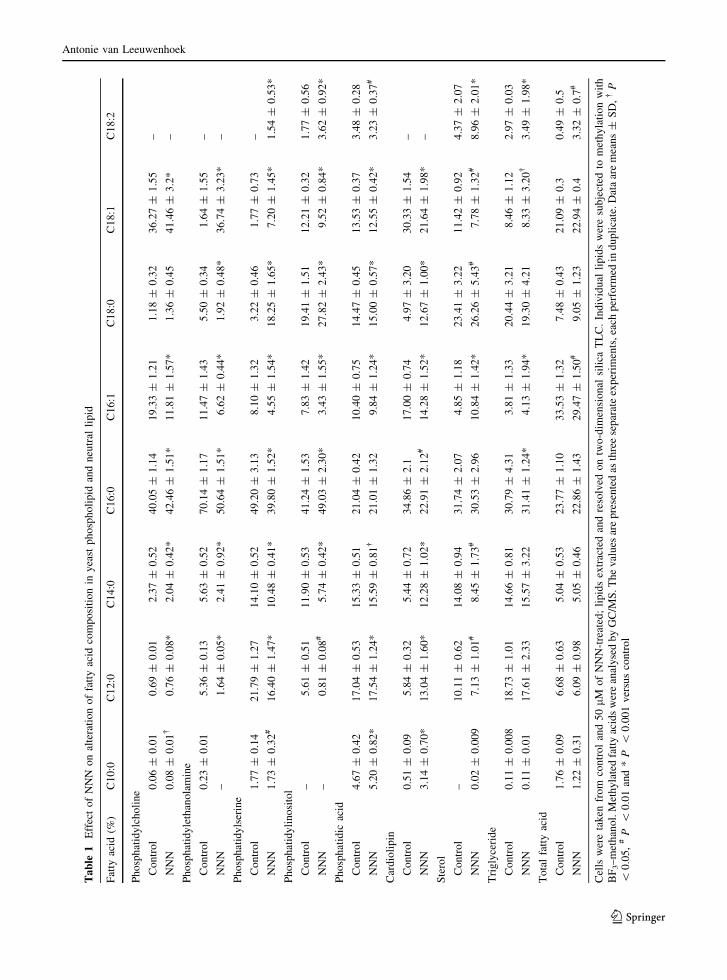
Table 1 shows the changes in FA composition in both
phospholipids and neutral lipids of NNN treated cells.
Cells during adaptation to NNN exposure; showed an
overall decrease in C16 series of fatty acids (C16:0
and C16:1) and an overall increase in C18 series fatty
acids (C18:0 and C18:1), particularly in PE and PS.
Antonie van Leeuwenhoek
123

In this study, alteration of the molecular species in
the presence of NNN were significantly (P \ 0.001)
higher in phospholipids when compared with neutral
lipids (Table 1). In PE monounsaturated FA (C18:1)
was increased significantly where as saturated C16
FA was decreased in NNN treated cells. NNN
exposed cells seemed to increase the saturated FA
chain length in PI. Among the phospholipids, short
chain FA content (C12:0 and C14:0) was increased in
CL in NNN treated cells (Table 1). In sterol there was
a marked increase in C18 series FAs.
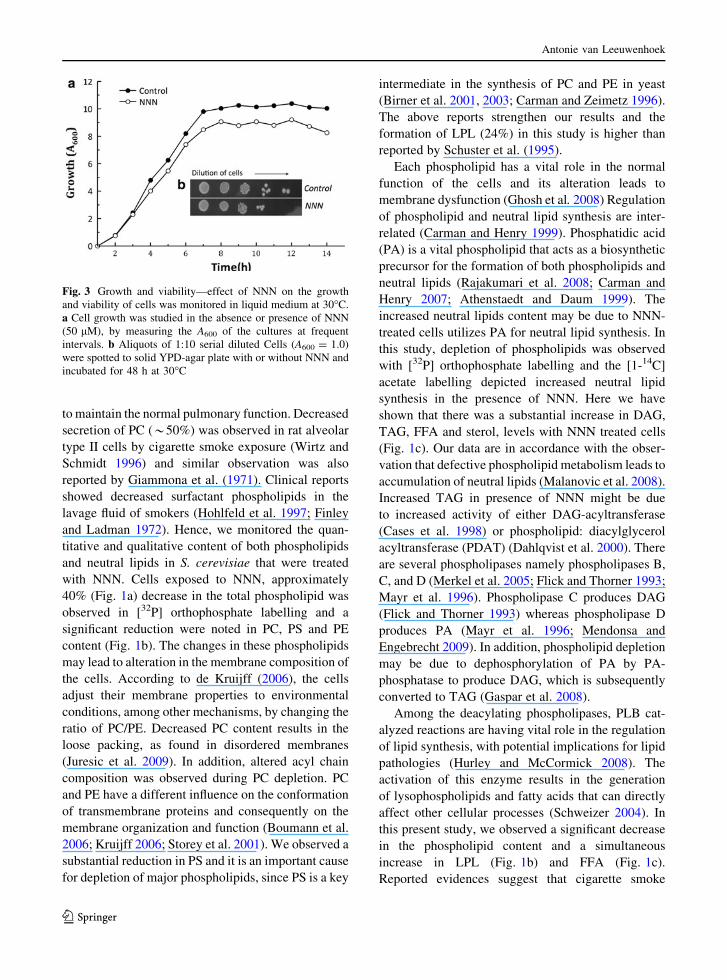
Cell counts
NNN treated cells yielded a lower (*80%) number
of cells than control (Fig. 3a), and it can be due to
cell damage and it was shown in Fig. 3b.
Discussion
Reduction of phospholipid content with NNN exposure
was fully corroborated, using S. cerevisiae as a model
system for investigating membrane lipid homeostasis.
In order to understand the impact of NNN on lipid
metabolism, S. cerevisiae cells were treated over a wide
range of NNN concentration. The alterations in total
phospholipid levels were determined in presence of
NNN and a concentration dependent reduction in total
phospholipid content (Fig. 1a) was observed. Reported
evidences showed that NNN exposure alters the lipid
composition of the membrane during adaptation and it
serves as an important factor for NNN binding
(Schuster et al. 1995; Sauk and Norris 1988).
During the last two decades, carcinogenicity of
TSNA compounds and its associated problems has
Fig. 1 Exposure to NNN alters the lipid composition. Equal
number of cells was taken in each group and at the end of the
experiment, lipids were extracted and resolved on silica TLC
and quantified. a Effect of NNN on incorporation of [32P]
orthophosphate into yeast phospholipid. The cells were grown
in YPD medium containing 100 lCi of [32P] orthophosphate
for 12 h in the absence or presence of different concentrations
(0–400 lM) of NNN as described in ‘‘Materials and Methods’’.
b, c Cells were grown in the absence or presence of optimal
concentration of NNN (50 lM) in the medium. b Phospholipid
content was observed by labelling phospholipids with 100 lCi
of [32P] orthophosphate. c Neutral lipid content was observed
by labelling neutral lipids with 0.5 lCi of [14C] acetate. Values
are the mean of three separate experiments, each performed in
duplicate. Data are means ± SD, �P \ 0.05, #P \ 0.01 and
*P \ 0.001 versus control
Antonie van Leeuwenhoek
123
been the subject of intensive investigation and
tremendous progress has been made. However under-
standing membrane remodelling, associated with
carcinogenicity, particularly phospholipid alteration,
is yet to be studied. Pathophysiological mechanisms
of TSNA compounds depend on membrane organi-
sation/composition. One of the first steps of carcino-
genesis by TSNAs is by inhalation of tobacco smoke
and the generation of reactive oxygen species within
the cells (Alavanja 2002). We have earlier reported
that increased membrane damage occurs through
reactive oxygen species (ROS) during NNN exposure
(Nachiappan et al. 1994). Interaction of NNN with
targeted cells mainly depends on lipid and fatty acid
composition (Schuster et al. 1992). A moderate
reduction in phospholipid content and alteration in
fatty acid pattern was also observed in presence of
NNN by others (Schuster et al. 1986, 1990; Sauk and
Norris 1988). Reported evidence suggests that NNN
may contribute to the alteration in the structure and
function of membrane lipids and also to its remod-
elling enzyme activity. Lands proposed that a deac-
ylation–reacylation cycle was involved in the
remodelling system. The Lands cycle consists of
phospholipid cleavage by phospholipases to produce
a lysophospholipid, ATP-driven activation of another
FFA by acyl-CoA synthetase, and transfer of the fatty
acid from the acyl-CoA to the lysophospholipid by
acyl-CoA:lysophospholipid acyltransferases (Lands
and Crawford 1976).
Interaction of NNN with targeted cells mainly
depends on lipid and fatty acid composition (Schuster
et al. 1992). NNN is a potent pulmonary carcinogen
and phospholipids are important components required
Fig. 2 NNN enhances PLB activity. The cells were grown for
12 h in the absence or presence of NNN and the cell lysate was
used as an enzyme source. The obtained cell lysate was incubated
with [32P] orthophosphate labelled phospholipid at 37�C for
60 min and the enzyme activity was measured. a Effect of NNN
(50 lM) on hydrolysis of various phospholipids. b The rate of
phospholipid hydrolysis was measured in wild type and plbDstrains (plb1, plb2 and plb3) exposed to various concentration of
NNN. c Effect Ca2? on hydrolysis of PC. The values are the
means of three separate experiments, each performed in
duplicate. Data are means ± SD, �P \ 0.05, #P \ 0.01 and
*P \ 0.001 versus control
Antonie van Leeuwenhoek
123
Ta
ble
1E
ffec
to
fN
NN
on
alte
rati
on
of
fatt
yac
idco
mp
osi
tio
nin
yea
stp
ho
sph
oli
pid
and
neu
tral
lip
id
Fat
tyac
id(%
)C
10
:0C
12
:0C
14
:0C
16
:0C
16
:1C
18
:0C
18
:1C
18
:2
Ph
osp
hat
idy
lch
oli
ne
Co
ntr
ol
0.0
6±
0.0
10
.69
±0
.01
2.3
7±
0.5
24
0.0
5±
1.1
41
9.3
3±
1.2
11
.18
±0
.32
36
.27
±1
.55
–
NN
N0
.08
±0
.01�
0.7
6±
0.0
8*
2.0
4±
0.4
2*
42
.46
±1
.51
*1
1.8
1±
1.5
7*
1.3
6±
0.4
54
1.4
6±
3.2
*–
Ph
osp
hat
idy
leth
ano
lam
ine
Co
ntr
ol
0.2
3±
0.0
15
.36
±0
.13
5.6
3±
0.5
27
0.1
4±
1.1
71
1.4
7±
1.4
35
.50
±0
.34
1.6
4±
1.5
5–
NN
N–
1.6
4±
0.0
5*
2.4
1±
0.9
2*
50
.64
±1
.51
*6
.62
±0
.44
*1
.92
±0
.48
*3
6.7
4±
3.2
3*
–
Ph
osp
hat
idy
lser
ine
Co
ntr
ol
1.7
7±
0.1
42
1.7
9±
1.2
71
4.1
0±
0.5
24
9.2
0±
3.1
38
.10
±1
.32
3.2
2±
0.4
61
.77
±0
.73
–
NN
N1
.73
±0
.32
#1
6.4
0±
1.4
7*
10
.48
±0
.41
*3
9.8
0±
1.5
2*
4.5
5±
1.5
4*
18
.25
±1
.65
*7
.20
±1
.45
*1
.54
±0
.53
*
Ph
osp
hat
idy
lin
osi
tol
Co
ntr
ol
–5
.61
±0
.51
11
.90
±0
.53
41
.24
±1
.53
7.8
3±
1.4
21
9.4
1±
1.5
11
2.2
1±
0.3
21
.77
±0
.56
NN
N–
0.8
1±
0.0
8#
5.7
4±
0.4
2*
49
.03
±2
.30
*3
.43
±1
.55
*2
7.8
2±
2.4
3*
9.5
2±
0.8
4*
3.6
2±
0.9
2*
Ph
osp
hat
idic
acid
Co
ntr
ol
4.6
7±
0.4
21
7.0
4±
0.5
31
5.3
3±
0.5
12
1.0
4±
0.4
21
0.4
0±
0.7
51
4.4
7±
0.4
51
3.5
3±
0.3
73
.48
±0
.28
NN
N5
.20
±0
.82
*1
7.5
4±
1.2
4*
15
.59
±0
.81�
21
.01
±1
.32
9.8
4±
1.2
4*
15
.00
±0
.57
*1
2.5
5±
0.4
2*
3.2
3±
0.3
7#
Car
dio
lip
in
Co
ntr
ol
0.5
1±
0.0
95
.84
±0
.32
5.4
4±
0.7
23
4.8
6±
2.1
17
.00
±0
.74
4.9
7±
3.2
03
0.3
3±
1.5
4–
NN
N3
.14
±0
.70
*1
3.0
4±
1.6
0*
12
.28
±1
.02
*2
2.9
1±
2.1
2#
14
.28
±1
.52
*1
2.6
7±
1.0
0*
21
.64
±1
.98
*–
Ste
rol
Co
ntr
ol
–1
0.1
1±
0.6
21
4.0
8±
0.9
43
1.7
4±
2.0
74
.85
±1
.18
23
.41
±3
.22
11
.42
±0
.92
4.3
7±
2.0
7
NN
N0
.02
±0
.00
97
.13
±1
.01
#8
.45
±1
.73
#3
0.5
3±
2.9
61
0.8
4±
1.4
2*
26
.26
±5
.43
#7
.78
±1
.32
#8
.96
±2
.01
*
Tri
gly
ceri
de
Co
ntr
ol
0.1
1±
0.0
08
18
.73
±1
.01
14
.66
±0
.81
30
.79
±4
.31
3.8
1±
1.3
32
0.4
4±
3.2
18
.46
±1
.12
2.9
7±
0.0
3
NN
N0
.11
±0
.01
17
.61
±2
.33
15
.57
±3
.22
31
.41
±1
.24
*4
.13
±1
.94
*1
9.3
0±
4.2
18
.33
±3
.20
�3
.49
±1
.98
*
To
tal
fatt
yac
id
Co
ntr
ol
1.7
6±
0.0
96
.68
±0
.63
5.0
4±
0.5
32
3.7
7±
1.1
03
3.5
3±
1.3
27
.48
±0
.43
21
.09
±0
.30
.49
±0
.5
NN
N1
.22
±0
.31
6.0
9±
0.9
85
.05
±0
.46
22
.86
±1
.43
29
.47
±1
.50
#9
.05
±1
.23
22
.94
±0
.43
.32
±0
.7#
Cel
lsw
ere
tak
enfr
om
con
tro
lan
d5
0l
Mo
fN
NN
-tre
ated
;li
pid
sex
trac
ted
and
reso
lved
on
two
-dim
ensi
on
alsi
lica
TL
C.
Ind
ivid
ual
lip
ids
wer
esu
bje
cted
tom
eth
yla
tio
nw
ith
BF
3–
met
han
ol.
Met
hy
late
dfa
tty
acid
sw
ere
anal
yse
db
yG
C/M
S.T
he
val
ues
are
pre
sen
ted
asth
ree
sep
arat
eex
per
imen
ts,ea
chp
erfo
rmed
ind
up
lica
te.D
ata
are
mea
ns
±S
D,�
P\
0.0
5,
#P
\0
.01
and
*P
\0
.00
1v
ersu
sco
ntr
ol
Antonie van Leeuwenhoek
123
to maintain the normal pulmonary function. Decreased
secretion of PC (*50%) was observed in rat alveolar
type II cells by cigarette smoke exposure (Wirtz and
Schmidt 1996) and similar observation was also
reported by Giammona et al. (1971). Clinical reports
showed decreased surfactant phospholipids in the
lavage fluid of smokers (Hohlfeld et al. 1997; Finley
and Ladman 1972). Hence, we monitored the quan-
titative and qualitative content of both phospholipids
and neutral lipids in S. cerevisiae that were treated
with NNN. Cells exposed to NNN, approximately
40% (Fig. 1a) decrease in the total phospholipid was
observed in [32P] orthophosphate labelling and a
significant reduction were noted in PC, PS and PE
content (Fig. 1b). The changes in these phospholipids
may lead to alteration in the membrane composition of
the cells. According to de Kruijff (2006), the cells
adjust their membrane properties to environmental
conditions, among other mechanisms, by changing the
ratio of PC/PE. Decreased PC content results in the
loose packing, as found in disordered membranes
(Juresic et al. 2009). In addition, altered acyl chain
composition was observed during PC depletion. PC
and PE have a different influence on the conformation
of transmembrane proteins and consequently on the
membrane organization and function (Boumann et al.
2006; Kruijff 2006; Storey et al. 2001). We observed a
substantial reduction in PS and it is an important cause
for depletion of major phospholipids, since PS is a key
intermediate in the synthesis of PC and PE in yeast
(Birner et al. 2001, 2003; Carman and Zeimetz 1996).
The above reports strengthen our results and the
formation of LPL (24%) in this study is higher than
reported by Schuster et al. (1995).
Each phospholipid has a vital role in the normal
function of the cells and its alteration leads to
membrane dysfunction (Ghosh et al. 2008) Regulation
of phospholipid and neutral lipid synthesis are inter-
related (Carman and Henry 1999). Phosphatidic acid
(PA) is a vital phospholipid that acts as a biosynthetic
precursor for the formation of both phospholipids and
neutral lipids (Rajakumari et al. 2008; Carman and
Henry 2007; Athenstaedt and Daum 1999). The
increased neutral lipids content may be due to NNN-
treated cells utilizes PA for neutral lipid synthesis. In
this study, depletion of phospholipids was observed
with [32P] orthophosphate labelling and the [1-14C]
acetate labelling depicted increased neutral lipid
synthesis in the presence of NNN. Here we have
shown that there was a substantial increase in DAG,
TAG, FFA and sterol, levels with NNN treated cells
(Fig. 1c). Our data are in accordance with the obser-
vation that defective phospholipid metabolism leads to
accumulation of neutral lipids (Malanovic et al. 2008).
Increased TAG in presence of NNN might be due
to increased activity of either DAG-acyltransferase
(Cases et al. 1998) or phospholipid: diacylglycerol
acyltransferase (PDAT) (Dahlqvist et al. 2000). There
are several phospholipases namely phospholipases B,
C, and D (Merkel et al. 2005; Flick and Thorner 1993;
Mayr et al. 1996). Phospholipase C produces DAG
(Flick and Thorner 1993) whereas phospholipase D
produces PA (Mayr et al. 1996; Mendonsa and
Engebrecht 2009). In addition, phospholipid depletion
may be due to dephosphorylation of PA by PA-
phosphatase to produce DAG, which is subsequently
converted to TAG (Gaspar et al. 2008).
Among the deacylating phospholipases, PLB cat-
alyzed reactions are having vital role in the regulation
of lipid synthesis, with potential implications for lipid
pathologies (Hurley and McCormick 2008). The
activation of this enzyme results in the generation
of lysophospholipids and fatty acids that can directly
affect other cellular processes (Schweizer 2004). In
this present study, we observed a significant decrease
in the phospholipid content and a simultaneous
increase in LPL (Fig. 1b) and FFA (Fig. 1c).
Reported evidences suggest that cigarette smoke
Fig. 3 Growth and viability—effect of NNN on the growth
and viability of cells was monitored in liquid medium at 30�C.
a Cell growth was studied in the absence or presence of NNN
(50 lM), by measuring the A600 of the cultures at frequent
intervals. b Aliquots of 1:10 serial diluted Cells (A600 = 1.0)
were spotted to solid YPD-agar plate with or without NNN and
incubated for 48 h at 30�C
Antonie van Leeuwenhoek
123

reduces the phospholipid content (Scott 2004) by
activated phospholipase A2 (Oulton et al. 1991).
Reduction of phospholipids may be due to increased
degradation and this was confirmed by enzymatic
hydrolysis of phospholipids by phospholipase assay.
We found that the enhanced phospholipase activity in
the presence of NNN, and the hydrolysis of PC was
significantly higher when compared to other phos-
pholipids (Fig. 2a). Current study suggested that the
reduction of phospholipids is mainly the result of
NNN mediated phospholipase activity and PC is a
preferred substrate. Apart from PC, marked hydroly-
sis was also observed in PS, PE, and PG in the
presence of NNN (Fig. 2a).
In S. cerevisiae PLBs are encoded by plb1, plb2,
and plb3 and are associated with the plasma mem-
brane, cell wall, and periplasmic space, respectively.
plb1 and plb2 preferred to hydrolyse PC, PE, PS and PI
whereas Plb3 hydrolyzes only PI and PS (Merkel et al.
2005). To identify the role NNN on these enzyme
activities, we performed phospholipase assay with all
three mutants and wild type cells. Our results suggest
that NNN significantly enhances the plb1 activity as
compared to plb2 and plb3 (Fig. 2b). PC hydrolysis
was markedly decreased when compared with wild-
type cells. This reveals the existence of other PC
degrading enzyme(s) other than plb1 that are activated
(Merkel et al. 2005). plb1 appears to be primarily
responsible for the degradation of mainly PC and to
some extent PI (Lee et al. 1994; Henry and Patton-
Vogt 1998) and we also observed similar results.
Interestingly, over expression of Plb2 does not result
in degradation of PC or PI, while Plb3 hydrolysed PI,
but not PC (Merkel et al. 2005). In addition, increased
plb1 activity leads to a significant increase in TAG and
a concomitant decrease in cellular phospholipids
(Merkel et al. 2005), suggesting that fatty acids used
for TAG synthesis are derived partly from PLB
catalysed phospholipid degradation.
Phospholipases are activated either at low Ca2?
concentrations or at high concentrations. To check
whether Ca2? is required for NNN mediated
phospholipid breakdown in vitro, we tested for
phospholipase assay with or without Ca2? in wild
type cells treated with or without NNN. In the
presences of NNN, the phospholipase activity was
greatly enhanced by the addition of Ca2?. In addition,
we have observed the increased enzyme activity in
NNN-treated cell even in the absence of Ca2?. It may
be due to PC degrading enzyme(s) other than Ca2?
dependent phospholipase that are activated under
certain conditions. Reported evidence showed that
PLB from yeast can be either Ca2? or other cations
dependent (Merkel et al. 2005; Fyrst et al. 1999; De
Kroon 2007). Overall results suggest that NNN
decreases the phospholipid content by enhancing
PLB activity and is associated with an increase in
LPL and free fatty acid content.
Decreased C16 series fatty acids (C16:0 and C16:1)
and increased C18 series fatty acids (C18:1) were
observed in NNN treated cells. The changes were more
predominant in PE the most abundant phospholipid
during PC depletion. Our results were strengthened by
Ferreira et al. (2004) who also found that the excess
saturated FAs are stored mainly in specific phospho-
lipids and not in TAG and steryl esters. The acyl chain
composition of CL is important for its function and we
investigated the CL profile in the presence of NNN.
The average length of the fatty acids is decreased,
specifically with increase in C12:0 and C14:0 and a
decrease in C16 series. The increase in myristic acid
might have a vital role in protein modification, an
essential step in signaling reactions such as the Ras
pathway, thus fatty acid modification alters cellular
proliferation and function (Tehlivets et al. 2007). Cell
viability was assessed with 50 lM of NNN and a 20%
reduction in growth was depicted. Possibly, the low PC
levels interfere with the diauxic shift; leading to
increased cell death (Boumann et al. 2006).
To summarize, we demonstrated the changes in
lipids by NNN in S. cerevisiae. Our results suggest
the decreased phospholipid content may be due to
enhanced PLB activity associated with increased
generation of lysophospholipids and free fatty acids.
We also observed accumulation of neutral lipids and
substantial changes in fatty acid composition. PLB/
PLA2 is a key enzyme involved in the pathology of
many clinically distinct lipid dysfunctions (Hurley
and McCormick 2008). Further studies using mutants
are required to clarify the mechanisms of the NNN in
both phospholipid and neutral lipid metabolism.
Acknowledgments The financial support from Bharathidasan
University, Tiruchirappalli, India is gratefully acknowledged.
We also acknowledge Department of Biochemistry, Indian
Institute of Science, Bangalore, India for helping us to conduct
the radioactive study and also providing the radioactive material.
Conflict of interest None.
Antonie van Leeuwenhoek
123

References
Alavanja MC (2002) Biologic damage resulting from exposure
to tobacco smoke and from radon: implication for pre-
ventive interventions. Oncogene 21:7365–7375
Athenstaedt K, Daum G (1999) Phosphatidic acid, a key
intermediate in lipid metabolism. Eur J Biochem 266:
1–16
Birner R, Burgermeister M, Schneiter R, Daum G (2001) Roles
of phosphatidylethanolamine and of its several biosyn-
thetic pathways in Saccharomyces cerevisiae. Mol Biol
Cell 12:997–1007
Birner R, Nebauer R, Schneiter R, Daum G (2003) Synthetic
lethal interaction of the mitochondrial phosphatidyletha-
nolamine biosynthetic machinery with the prohibition
complex of Saccharomyces cerevisiae. Mol Biol Cell
14:370–383
Bligh G, Dyer WJ (1959) A rapid method of total lipid
extraction and purification. Can J Biochem Physiol
37:911–917
Boumann HA, Gubbens J, Koorengevel MC, Oh CS, Martin
CE, Heck AJ, Patton-Vogt J, Henry SA, de Kruijff B, de
Kroon AI (2006) Depletion of phosphatidylcholine in
yeast induces shortening and increased saturation of the
lipid acyl chains: evidence for regulation of intrinsic
membrane curvature in a eukaryote. Mol Biol Cell
17:1006–1017
Bradford MM (1976) A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing
the principle of protein–dye binding. Anal Biochem
72:248–254
Carman GM, Henry SA (1999) Phospholipid biosynthesis in the
yeast Saccharomyces cerevisiae and interrelationship with
other metabolic processes. Prog Lipid Res 38:361–399
Carman GM, Henry SA (2007) Phosphatidic acid plays a
central role in the transcriptional regulation of glycero-
phospholipid synthesis in Saccharomyces cerevisiae.
J Biol Chem 282:37293–37297
Carman GM, Zeimetz GM (1996) Regulation of phospholipid
biosynthesis in the yeast Saccharomyces cerevisiae. J Biol
Chem 271:13293–13296
Cases S, Smith SJ, Zheng YW, Myers HM, Lear SR, Sande E,
Novak S, Collins C, Welch CB, Lusis AJ, Erickson SK,
Farese RV Jr (1998) Identification of a gene encoding an
acyl CoA:diacylglycerol acyltransferase, a key enzyme in
triacylglycerol synthesis. Proc Natl Acad Sci 95:
13018–13023
Chicco AJ, Sparagna GC (2006) Role of cardiolipin alterations
in mitochondrial dysfunction and disease. Am J Physiol
Cell Physiol 292:C33–C44
Dahlqvist A, Stahl U, Lenman M, Banas A, Lee M, Sandager
L, Ronne H, Stymne S (2000) Phospholipid:diacylglyc-
erol acyltransferase: an enzyme that catalyzes the acyl-
CoA-independent formation of TAG in yeast and plants.
Proc Natl Acad Sci 97:6487–6492
De Kroon AI (2007) Metabolism of phosphatidylcholine and its
implications for lipid acyl chain composition in Saccha-romyces cerevisiae. Biochim Biophys Acta 1771:343–352
Exton JH (1994) Phosphatidylcholine breakdown and signal
transduction. Biochim Biophys Acta 1212:26–42
Ferreira T, Regnacq M, Alimardani P, Moreau-Vauzelle C,
Berges T (2004) Lipid dynamics in yeast under haem-
induced unsaturated fatty acid and/or sterol depletion.
Biochem J 378:899–908
Finley TN, Ladman AJ (1972) Low yield of pulmonary sur-
factant in cigarette smokers. N Engl J Med 286:223–227
Fisher AB, Dodia C, Chander A, Jain M (1992) A competitive
inhibitor of phospholipase A2 decreases surfactant phos-
phatidylcholine degradation by the rat lung. Biochem J
288:407–411
Flick JS, Thorner J (1993) Genetic and biochemical charac-
terization of a phosphatidylinositol-specific phospholipase
C in Saccharomyces cerevisiae. Mol Cell Biol 13:
5861–5876
Fyrst H, Oskouian B, Kuypers FA, Saba JD (1999) The PLB2
gene of Saccharomyces cerevisiae confers resistance to
lysophosphatidylcholine and encodes a phospholipase B/
lysophospholipase. Biochemistry 38:5864–5871
Gaspar ML, Jesch SA, Viswanatha R, Antosh AL, Brown WJ,
Kohlwein SD, Henry SA (2008) A block in endoplasmic
reticulum-to-Golgi trafficking inhibits phospholipid syn-
thesis and induces neutral lipid accumulation. J Biol
Chem 283:25735–25751
Gheriani-Gruszka N, Almog S, Biltonen RL, Lichtenberg D
(1988) Hydrolysis of phosphatidylcholine in phosphati-
dylcholine-cholate mixtures by porcine pancreatic phos-
pholipase A2. J Biol Chem 263:11808–11813
Ghosh AK, Ramakrishnan G, Rajasekharan R (2008)
YLR099C (ICT1) encodes a soluble Acyl-CoA-dependent
lysophosphatidic acid acyltransferase responsible for
enhanced phospholipid synthesis on organic solvent stress
in Saccharomyces cerevisiae. J Biol Chem 283:
9768–9775
Giammona ST, Tocci P, Webb WR (1971) Effects of cigarette
smoke on incorporation of radioisotopically labeled pal-
mitic acid into pulmonary surfactant and on surface
activity of canine lung extracts. Am Rev Respir Dis
104:358–367
Hecht SS, Hoffmann D (1988) Tobacco-specific nitrosamines,
an important group of carcinogens in tobacco and tobacco
smoke. Carcinogenesis 9:875–884
Henry SA, Patton-Vogt JL (1998) Genetic regulation of
phospholipid metabolism: yeast as a model eukaryote.
Prog Nucleic Acid Res Mol Biol 61:133–179
Hohlfeld J, Fabel H, Hamm H (1997) The role of pulmonary
surfactant in obstructive airways disease. Eur Respir J
10:482–491
Hurley BP, McCormick BA (2008) Multiple roles of phos-
pholipase A2 during lung infection and inflammation.
Infect Immun 76:2259–2272
IARC Monogr (2007) Smokeless tobacco and some tobacco-
specific N-nitrosamines. Eval Carcinog Risks Hum
89:1–592
Juresic GC, Blagovic B, Rupcic J (2009) Alterations in phos-
phatidylcholine and phosphatidylethanolamine content
during fermentative metabolism in Saccharomyces cere-visiae Brewer’s yeast. Food Technol Biotechnol
47:246–252
Kruijff BD (2006) Membranes, where lipids and proteins meet.
Chem Phys Lipids 143:41–42
Antonie van Leeuwenhoek
123

Lands WEM, Crawford CG (1976) Enzymes of membrane
phospholipid metabolism in animals. In: Martonosi A (ed)
The enzymes of biological membranes, vol 2. Plenum
Press, New York, pp 3–85
Lee KS, Patton JL, Fido M, Hines LK, Kohlwein SD, Paltauf F,
Henry SA, Levin DE (1994) The Saccharomyces cerevi-siae PLB1 gene encodes a protein required for lysophos-
pholipase and phospholipase B activity. J Biol Chem
269:19725–19730
Lin RK, Hsieh YS, Lin PH, Hsu S, Chen CY, Tang YA, Lee
CF, Wang YC (2010) The tobacco-specific carcinogen
NNK induces DNA methyltransferase 1 accumulation and
tumor suppressor gene hypermethylation in mice and lung
cancer patients. J Clin Invest 120:521–532
Malanovic N, Streith I, Wolinski H, Rechberger G, Kohlwein
SD, Tehlivets O (2008) S-adenosyl-L-homocysteine
hydrolase, key enzyme of methylation metabolism, regu-
lates phosphatidylcholine synthesis and TAG homeostasis
in yeast: implications for homocysteine as a risk factor of
atherosclerosis. J Biol Chem 283:23989–23999
Mayr JA, Kohlwein SD, Paltauf F (1996) Identification of a
novel, Ca(2?)-dependent phospholipase D with preference
for phosphatidylserine and phosphatidylethanolamine in
Saccharomyces cerevisiae. FEBS Lett 393:236–240
Mendonsa R, Engebrecht J (2009) Phospholipase D function in
Saccharomyces cerevisiae. Biochim Biophys Acta 1791:
970–974
Merkel O, Oskolkova OV, Raab F, El-Toukhy R, Paltauf F
(2005) Regulation of activity in vitro and in vivo of three
phospholipases B from Saccharomyces cerevisiae. Bio-
chem J 387:489–496
Morrison WR, Smith LM (1964) Preparation of fatty acid
methyl esters and dimethylacetals from lipids with boron
fluoridemethanol. J Lipid Res 5:600–608
Nachiappan V, Mufti SI, Chakravarti A, Eskelson CD, Raja-
sekharan R (1994) Lipid peroxidation and ethanol-related
tumor promotion in Fischer-344 rats treated with tobacco-
specific nitrosamines. Alcohol Alcohol 29:565–574
Oulton M, Moores HK, Scott JE, Janigan DT, Hajela R (1991)
Effects of smoke inhalation on surfactant phospholipids
and phospholipase A2 activity in the mouse lung. Am J
Pathol 138:195–202
Rajakumari S, Daum G (2010) Janus-faced enzymes yeast
Tgl3p and Tgl5p catalyze lipase and acyltransferase
reactions. Mol Biol Cell 21:501–510
Rajakumari S, Grillitsch K, Daum G (2008) Synthesis and
turnover of non-polar lipids in yeast. Prog Lipid Res
47:157–171
Sauk JJ, Norris K (1988) Tobacco components induce altera-
tions in keratinocyte lipid membrane fluidity and inhibit
cell proliferation in culture. J Oral Pathol 17:30–33
Schuster GS, Erblad JF, Wyrick SD, Singh BB (1986) Oral
epithelial cell lipid synthesis in the presence of retinoic
acid or nitrosonornicotine. J Oral Pathol 15:430–433
Schuster GS, Lubas S, Erbland JF, Singh BB (1990) Binding
and uptake of N-nitrosonornicotine by oral epithelial cells.
J Oral Pathol Med 19:114–118
Schuster GS, Lubas S, Dirksen TR, Erbland JF (1992) Lipids as
factors in the cell response to tobacco components. Natl
Cancer Inst Monogr 2:191–201
Schuster GS, Caughman GB, Dirksen TR (1995) Alteration of
epithelial cell lipid synthesis by N-nitrosonornicotine. In
Vitro Cell Dev Biol Anim 31:295–299
Schweizer M (2004) Lipids and membranes. In: Schweizer M,
Dickinson JR (eds) The metabolism and molecular
physiology of Saccharomyces cerevisiae. CRC Press,
London, pp 140–223
Scott JE (2004) The pulmonary surfactant: impact of tobacco
smoke and related compounds on surfactant and lung
development. Tob Induc Dis 2:3–25
Stenger PC, Alonso C, Zasadzinski JA, Waring AJ, Jung CL,
Pinkerton KE (2009) Environmental tobacco smoke
effects on lung surfactant film organization. Biochim
Biophys Acta 1788:358–370
Storey MK, Clay KL, Kutateladze T, Murphy RC, Overduin M,
Voelker DR (2001) Phosphatidylethanolamine has an
essential role in Saccharomyces cerevisiae that is inde-
pendent of its ability to form hexagonal phase structures.
J Biol Chem 276:48539–48548
Tehlivets O, Scheuringer K, Kohlwein SD (2007) Fatty acid
synthesis and elongation in yeast. Biochim Biophys Acta
1771:255–270
Vance JE, Steenbergen R (2005) Metabolism and functions of
phosphatidylserine. Prog Lipid Res 44:207–234
Wagner S, Paltauf F (1994) Generation of glycerophospholipid
molecular species in the yeast Saccharomyces cerevisiae.
Fatty acid pattern of phospholipid classes and selective
acyl turnover at sn-1 and sn-2 positions. Yeast 10:
1429–1437
Wirtz HR, Schmidt M (1996) Acute influence of cigarette
smoke on secretion of pulmonary surfactant in rat alveolar
type II cells in culture. Eur Respir J 9:24–32
Yamashita A, Sugiura T, Waku K (1997) Acyltransferases and
transacylases involved in fatty acid remodeling of phos-
pholipids and metabolism of bioactive lipids in mamma-
lian cells. J Biochem 122:1–16
Antonie van Leeuwenhoek
123




















