
Glutathione peroxidase3 of Saccharomyces cerevisiae protects phospholipids during cadmium-induced...
11
ORIGINAL PAPER Glutathione peroxidase3 of Saccharomyces cerevisiae protects phospholipids during cadmium-induced oxidative stress Kannan Muthukumar • Selvaraj Rajakumar • Mary Nirmala Sarkar • Vasanthi Nachiappan Received: 22 October 2010 / Accepted: 4 January 2011 / Published online: 13 January 2011 Ó Springer Science+Business Media B.V. 2011 Abstract The present study was undertaken to determine the role of glutathione peroxidase3 (gpx3) in phospholipid protection in cells. Wild-type (WT) cells showed an overall increase in phospho- lipids upon 50 lM cadmium (Cd)-treatment, whereas an untreated gpx3D strain showed a drastic reduction in overall phospholipids which was further reduced with 50 lM Cd. In WT cells, Cd-exposure increased the short chain fatty acids and decreased the unsat- urated fatty acids and the magnitude was high in Cd- treated gpx3D cells. Purified recombinant gpx3p showed higher activity with phospholipid hydroper- oxides than shorter hydroperoxides. An increase in gpx activity was observed in Cd-treated WT cells and no such alteration was observed in gpx3D. WT cells treated with Cd showed an increase in MDA over untreated, while untreated gpx3D cells themselves showed a higher level of MDA which was further enhanced with Cd-treatment. Iron, zinc and calcium levels were significantly altered in WT and gpx3D cells during Cd-treatment. Keywords Glutathione peroxidase Á Phospholipids Á Fatty acids Á Antioxidants Á Cadmium Á Saccharomyces cerevisiae and oxidative stress Abbreviations Cd Cadmium CL Cardiolipin Gpx Glutathione peroxidase IPTG Isopropyl b-D-1-thiogalactopyranoside LPL Lysophospholipids MDA Malondialdehyde Ni 2? NTA nickel nitrilo acetic acid-agarose PC Phosphatidylcholine PE Phosphatidylethanolamine PHGpx Phospholipid hydroperoxide glutathione peroxidase PI Phosphatidylinositol PLOOH Phospholipid hydroperoxide PS Phosphatidylserine ROS Reactive oxygen species Introduction In cells phospholipids play diverse roles that are essential for growth and metabolism. The major phospholipids found in the cell membranes of Saccha- romyces cerevisiae include phosphatidylcholine (PC), phosphatidylethanolamine (PE), phosphatidylinositol K. Muthukumar Á S. Rajakumar Á V. Nachiappan (&) Department of Biochemistry, Bharathidasan University, Tiruchirappalli 620 024, Tamilnadu, India e-mail: [email protected] M. N. Sarkar Department of Biochemistry, Indian Institute of Science, Bangalore 560012, India 123 Antonie van Leeuwenhoek (2011) 99:761–771 DOI 10.1007/s10482-011-9550-9
Transcript of Glutathione peroxidase3 of Saccharomyces cerevisiae protects phospholipids during cadmium-induced...

ORIGINAL PAPER
Glutathione peroxidase3 of Saccharomyces cerevisiaeprotects phospholipids during cadmium-inducedoxidative stress
Kannan Muthukumar • Selvaraj Rajakumar •
Mary Nirmala Sarkar • Vasanthi Nachiappan
Received: 22 October 2010 / Accepted: 4 January 2011 / Published online: 13 January 2011
� Springer Science+Business Media B.V. 2011
Abstract The present study was undertaken to
determine the role of glutathione peroxidase3
(gpx3) in phospholipid protection in cells. Wild-type
(WT) cells showed an overall increase in phospho-
lipids upon 50 lM cadmium (Cd)-treatment, whereas
an untreated gpx3D strain showed a drastic reduction
in overall phospholipids which was further reduced
with 50 lM Cd. In WT cells, Cd-exposure increased
the short chain fatty acids and decreased the unsat-
urated fatty acids and the magnitude was high in Cd-
treated gpx3D cells. Purified recombinant gpx3p
showed higher activity with phospholipid hydroper-
oxides than shorter hydroperoxides. An increase in
gpx activity was observed in Cd-treated WT cells and
no such alteration was observed in gpx3D. WT cells
treated with Cd showed an increase in MDA over
untreated, while untreated gpx3D cells themselves
showed a higher level of MDA which was further
enhanced with Cd-treatment. Iron, zinc and calcium
levels were significantly altered in WT and gpx3Dcells during Cd-treatment.
Keywords Glutathione peroxidase � Phospholipids �Fatty acids � Antioxidants � Cadmium �Saccharomyces cerevisiae and oxidative stress
Abbreviations
Cd Cadmium
CL Cardiolipin
Gpx Glutathione peroxidase
IPTG Isopropyl b-D-1-thiogalactopyranoside
LPL Lysophospholipids
MDA Malondialdehyde
Ni2?NTA nickel nitrilo acetic acid-agarose
PC Phosphatidylcholine
PE Phosphatidylethanolamine
PHGpx Phospholipid hydroperoxide glutathione
peroxidase
PI Phosphatidylinositol
PLOOH Phospholipid hydroperoxide
PS Phosphatidylserine
ROS Reactive oxygen species
Introduction
In cells phospholipids play diverse roles that are
essential for growth and metabolism. The major
phospholipids found in the cell membranes of Saccha-
romyces cerevisiae include phosphatidylcholine (PC),
phosphatidylethanolamine (PE), phosphatidylinositol
K. Muthukumar � S. Rajakumar � V. Nachiappan (&)
Department of Biochemistry, Bharathidasan University,
Tiruchirappalli 620 024, Tamilnadu, India
e-mail: [email protected]
M. N. Sarkar
Department of Biochemistry, Indian Institute of Science,
Bangalore 560012, India
123
Antonie van Leeuwenhoek (2011) 99:761–771
DOI 10.1007/s10482-011-9550-9

(PI), and phosphatidylserine (PS), and the mitochon-
drial membranes also contain a significant amount of
phosphatidylglycerol (PG) and cardiolipin (CL) (Car-
man and Han 2009). Phospholipid biosynthetic
enzymes are regulated to cope with a variety of stress
conditions (Carman and Han 2009; Iwanyshyn et al.
2004). Changes in the phospholipid content were
shown to be an important factor in providing tolerance
against organic solvents (Ghosh et al. 2008). Elegant
work on mechanisms of solvent tolerance in Pseudo-
monas putida strain showed that the strain was able to
repair the damaged phospholipids through increased
biosynthesis and this was necessary to stabilize the cell
membrane structure (Pinkart and White 1997). Cell
membranes contain a carefully balanced mixture of
different phospholipid classes, each with a certain fatty
acyl chain composition. These compositions are
important for the physical properties of the membrane
and are altered with variations in the temperature and
other environmental conditions (Aguilar and de Men-
doza 2006; Martin et al. 2007; Falcone et al. 2004). The
most common fatty acids esterified to the glycero-
phosphate backbone of the phospholipids include
palmitic acid (C16:0), palmitoleic acid (C16:1), stearic
acid (C18:0), and oleic acid (C18:1) (Rattray et al.
1975). Unsaturated acyl chains of phospholipids in cell
membranes are prominent targets of oxidant attack.
This can result in lipid peroxidation, a degenerative
process that perturbs structure and function of the
target system, often with cytopathological conse-
quences (Girotti 1985).
A variety of environmental toxicants cause oxida-
tive stress through peroxidation of membrane lipids,
(Steffensen et al. 1994) and heavy metals are
considered to be one of the potent causative agents.
Cadmium (Cd) is one of the most toxic heavy metal
pollutants, inducing a broad spectrum of toxicolog-
ical and biochemical dysfunctions constituting seri-
ous hazard to health. Cd is a multi-target toxicant for
most organisms, causing severe damage to lung,
liver, kidney, testis and placenta (Patrick 2003). The
main routes of poisoning are either through occupa-
tional exposure or through ingestion of contaminated
food and water (Thorsen et al. 2009). Cd has an ionic
radius close to calcium (Ca2?) while its electro
negativity is similar to zinc (Zn2?). These properties
allow Cd to enter the cells and interfere with the
physiological functions of both Ca and Zn (Suwalsky
et al. 2004). It has a high affinity for protein thiol
groups and also induces membrane damage by
interacting with lipids (Girault et al. 1998). Cd
induces the production of a variety of reactive oxygen
species (ROS) including O2•- and •OH. Cytoprotec-
tion against lipid peroxidation and other types of
oxidative damage is accomplished by diverse enzy-
matic and non-enzymatic antioxidants. Saccharomy-
ces cerevisiae has evolved with many antioxidants
that include glutathione (GSH), catalase (CAT),
superoxide dismutase (SOD) (Temple et al. 2005)
and glutathione peroxidase (gpx) (Arthur 2000).
Gpx is an important participant in antioxidant
protection, which can reduce and detoxify various
organic hydroperoxides at the expense of GSH.
Saccharomyces cerevisiae expresses three different
gpx namely gpx1, gpx2 and gpx3 (Inoue et al. 1999)
with phospholipid hydroperoxidase activity (Avery
and Avery 2001). Among them, gpx3 is major
cellular enzyme that prevents the peroxidation of
membrane lipids and it also plays a major role in
enzymatic defence against oxidative damage (Ursini
et al. 1995). It has been reported that gpx3D is
hypersensitive to peroxides, whereas gpx1D or gpx2Dare resistant to oxidative stress (Ursini et al. 1995).
Gpx3 has multiple functions, by adapting to oxidative
stress response, acting as a ROS scavenger and also
as an efficient linker in repairing proteins during
oxidative stress damage (Temple et al. 2005). Two
major forms of gpx have been characterized in cells:
classical gpx and phospholipid hydroperoxide gluta-
thione peroxidase (PHGPx) (Arthur 2000). The
inorganic and organic hydroperoxides are substrates
for both enzymes and PHGPx alone reduces lipid
hydroperoxides esterified to bio membranes. Thus,
PHGPx is often considered the principal cellular
enzyme capable of repairing membrane lipid perox-
idation, the highly damaging process that has been
linked to pathological conditions such as ischemic
injury, atherosclerosis and carcinogenesis (Halliwell
and Gutteridge 1999).
Hence we determined the differences in phospho-
lipid and its fatty acid composition of wild-type (WT)
and gpx3D strains during Cd-treatment. The substrate
preference assay of gpx3 was studied with recombi-
nant purified gpx3p. To assess lipid peroxidation, the
levels of malondialdehyde (MDA) was quantified and
the activity of gpx was also assayed. The levels of
intracellular contents of divalent iron (Fe), Ca, Zn, Cd
and copper (I) (Cu) were analysed. In this study we
762 Antonie van Leeuwenhoek (2011) 99:761–771
123

investigated the possible relationship of gpx3 and
phospholipids in Saccharomyces cerevisiae during
oxidative stress.
Materials and methods
Materials
WT strain of Saccharomyces cerevisiae BY4741
[MATa his3D1 leu2D0 met15D0 ura3D0] was pro-
cured from Open Biosystems, USA and its isogenic
gpx3D strain was a gift from Prof. William A. Prinz,
NIH. The pET14b-gpx3 plasmid was kindly provided
by Prof. Simon V. Avery, University of Nottingham,
UK. Yeast extract and peptone were obtained from
Difco. The nickel-nitrilotriacetic acid (Ni2?NTA)
matrix was from Qiagen India Pvt. Ltd. All chemicals
and solvents were purchased from Sigma unless
specifically mentioned.
Growth conditions
Yeast cells were grown in YPD (1% yeast extract, 2%
peptone, and 2% dextrose) medium (pH 7.0) with
aeration at 30�C. For growth in Cd-containing medium,
the yeast cells were grown in 5 ml of YPD medium for
12 h. The cells were harvested and transferred to 25 ml
of YPD media, so that A600 of 0.1 OD was reached and
cells were grown at 30�C (with or without Cd) till the
final A600 of 1.00 OD was achieved.
Lipid extraction and analysis
Lipids from yeast cells were extracted by the method
of Bligh and Dyer (1959). The individual phospho-
lipids were separated by two-dimensional TLC on
silica plates (Wagner and Paltauf 1994). Individual
phospholipids were identified by comparing the Rf
values of the unknown with the Rf values of the
standard. Spots corresponding to individual phospho-
lipids were scraped off and quantified by estimating
phospholipid phosphorus after perchloric acid diges-
tion by the method of Siakotos et al. (1966).
Fatty acids analysis
Fatty acids of phospholipids were analyzed by gas
chromatography. Phospholipids were separated by
TLC and individual phospholipids were extracted
from the silica gel with chloroform/methanol (2:1
v/v), and subjected to methanolysis using BF3-
methanol for conversion to methyl esters (Morrison
and Smith 1964). Fatty acid methyl esters were
separated by gas chromatography and quantification
was referred to heptadecanoic methyl ester, as an
internal standard.
Determination of MDA
Lipid peroxidation was measured by the content of
MDA as described by Steels et al. (1994).
Preparation of cell-free extract
Cells were centrifuged (6,000 rpm for 5 min),
washed and sonicated in buffer (100 mM Tris–HCl
pH 7.4, 5 mM MgCl2, 1% glycerol, 1 mM phenyl
methyl sulfonyl fluoride, and 0.03 mM leupeptin).
The homogenate was then centrifuged at 10,000 rpm
for 15 min at 4�C and the supernatant was used to
determine enzyme activity. Total protein was deter-
mined by the method of Bradford (1976).
Expression and purification of recombinant gpx3p
The pET14b-gpx3 construct was transformed into E.
coli BL21 (DE3) cells and the transformed cells were
induced with 0.5 mM IPTG for 2 h. Briefly, IPTG
induced cells were centrifuged and the cell pellet was
resuspended in lysis buffer containing 50 mM Tris–
HCl (pH 8.0) and 300 mM NaCl (buffer A). Cells
were disrupted by sonication. The supernatant was
allowed to bind to the Ni2?-NTA matrix. The column
was washed with buffer A containing 50 mM imid-
azole. The bound protein was eluted with 250 mM
imidazole in buffer A. One-millilitre fractions were
collected and checked for activity. The fractions
showing activity were analyzed for the presence of
protein in 10% SDS-PAGE (Laemmli 1970) followed
by Coomassie Blue staining.
Synthesis and quantification of phospholipid
hydroperoxide
Phospholipid hydroperoxide (PLOOH) was synthesised
using soybean type IV lipoxygenase (Maiorino et al.
1990). The separation of peroxidized phospholipids was
Antonie van Leeuwenhoek (2011) 99:761–771 763
123
done by TLC and identified using tetramethyl-P-
phenylenediamine detection (Kriska and Girotti
2004). The concentration of PLOOH was estimated
by a colorimetric reaction using the ferrous oxidation of
xylenol orange reagent (FOX 2) with reference to a
calibration curve produced with known H2O2 concen-
trations (Wolff 1994).
Assay of gpx
Enzymatic activities of gpx in crude cell extracts or
purified gpx3p were assayed at 25�C with PLOOH or
other substrates according to standard protocols
(Inoue et al. 1999).
Determination of intracellular metals
Cells were harvested by centrifugation at 7,000 rpm
for 10 min. Crude extracts were prepared by glass
bead lysis and then resuspended in nitric acid and
heated at 200�C for 4 h. After complete digestion,
1 ml was diluted to 10 ml in metal-free water and
subjected to analysis using an inductively coupled
plasma atomic emission spectrometer (ICP-OES)
(Perkin Elmer 3300). This analysis was done at
Central Institute of Medicinal and Aromatic Plants
(CIMAP), Lucknow, India following the method of
Doner and Ege (2004).
Statistics
Data were analyzed using the programs of SPSS 10.0
(Statistical Package for the Social Science for Win-
dows 10.0). All the values reported in the paper were
the mean of three replicates. Statistical analysis was
carried out by analysis of variance (ANOVA) test. A
significant difference among means was determined
at P \ 0.05.
Results
Exposure of Cd alter phospholipid composition
in WT and gpx3D cells
To analyze the alteration of phospholipids under Cd-
stress, WT and gpx3D cells were grown in the
presence or absence of 50 lM Cd up to mid log
phase. It was observed that on exposure of WT cells
to Cd significantly (P \ 0.05) increases the levels of
total and individual phospholipids (PC, PE, PS and
LPL) when compared to WT control cells (Fig. 1).
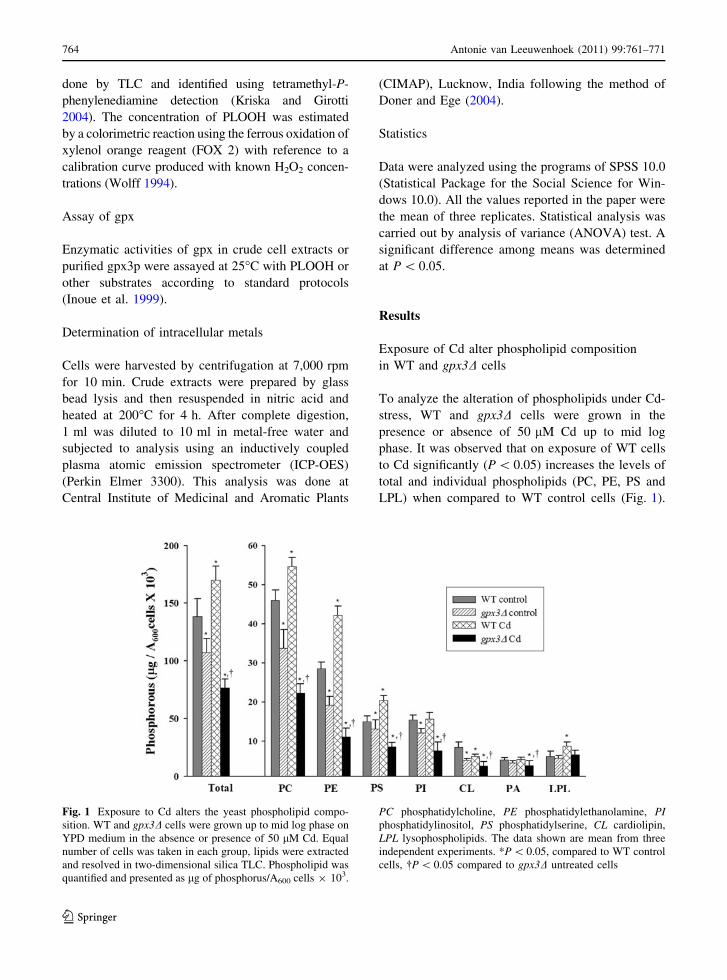
Fig. 1 Exposure to Cd alters the yeast phospholipid compo-
sition. WT and gpx3D cells were grown up to mid log phase on
YPD medium in the absence or presence of 50 lM Cd. Equal
number of cells was taken in each group, lipids were extracted
and resolved in two-dimensional silica TLC. Phospholipid was
quantified and presented as lg of phosphorus/A600 cells 9 103.
PC phosphatidylcholine, PE phosphatidylethanolamine, PIphosphatidylinositol, PS phosphatidylserine, CL cardiolipin,
LPL lysophospholipids. The data shown are mean from three
independent experiments. *P \ 0.05, compared to WT control
cells, �P \ 0.05 compared to gpx3D untreated cells
764 Antonie van Leeuwenhoek (2011) 99:761–771
123

On the contrary to WT cells, Cd-treated gpx3D cells
showed a significant (P \ 0.05) reduction in both
total and individual phospholipids when compared to
gpx3D untreated cells (Fig. 1). Control gpx3D cells
themselves showed a significant (P \ 0.05) decrease
in PC, PE, PI and CL when compared to WT control
cells (Fig. 1).
Fatty acid profile of Cd-treated WT and gpx3Dcells
To elucidate the changes in the acyl composition of
individual phospholipids, fractions were analyzed by
GC–MS. Table 1 shows the changes in acyl compo-
sition of phospholipids in WT and gpx3D cells. In
contrast to phospholipid results not much difference
was observed in fatty acid content of gpx3D control
cells when compared to WT control cells. Exposure
of yeast cells to Cd, showed an overall increase in
short chain fatty acids (C 10:0) and decrease in
unsaturated fatty acids (16:1 and 18:1) in all phos-
pholipids (Table 1). In WT cells Cd-treatment sig-
nificantly increases the level of short chain and
medium chain (C12:0 and C14:0) fatty acids in PC,
PE, PS and CL. The levels of saturated fatty acids
(C16:0 and 18:0) were also increased in PC, PE, PS,
PI and CL when compared to WT control cells.
Similar results was found with Cd-treated gpx3D cells
and the magnitude was high when compared to WT
cells (Table 1).
Determination of MDA in Cd-treated WT
and gpx3D cells
The role of gpx3 in elimination of lipid peroxides was
studied in WT and gpx3D cells. The level of MDA in
Cd-treated and untreated conditions are shown in
Fig. 2. In response to Cd, WT and gpx3D cells
accumulated (P \ 0.05) more MDA than its control
cells. The untreated gpx3D cells themselves showed
an increase in MDA levels and the levels were closer
to Cd-treated WT cells. The gpx3D cells showed a
further increase in MDA upon Cd- treatment (Fig. 2).
Expression and purification of gpx3p
The recombinant gpx3p was expressed in E. coli and
this protein was purified by Ni2?NTA affinity
chromatography. Protein bands corresponding to the
recombinant product were evident from 10% SDS-
PAGE as 18.6 kDa, respectively (Fig. 3).
Substrate preference of the gpx3p
To determine the substrate specificity, the gpx activ-
ities were measured in WT, gpx3D cell extracts and
with purified recombinant gpx3 using different sub-
strates (Fig. 4). Functionally active purified recombi-
nant gpx3 protein showed activity against PCOOH,
PEOOH, PSOOH, LPCOOH, t-BHP and H2O2
(Fig. 4). However higher activity was noted in
PCOOH, PEOOH followed by PSOOH and a
decreased activity was observed against t-BHP, H2O2
and LPCOOH. A similar trend was also observed with
WT cell extract. A significant (P \ 0.05) decrease in
the gpx activity was observed in gpx3D cells against
hydroperoxides when compared to WT cells (Fig. 4).
Activity of gpx with H2O2 and PCOOH
The activity of gpx in WT and gpx3D cells exposed to
Cd was studied. Increased activity was observed in
Cd-treated WT cells and PCOOH was a preferred
substrate over H2O2 (Fig. 5). A decrease in activity
was observed in gpx3D as compared to WT control
cells. No significant changes were observed between
Cd-treated or untreated gpx3D cells (Fig. 5).
Determination of intracellular metals
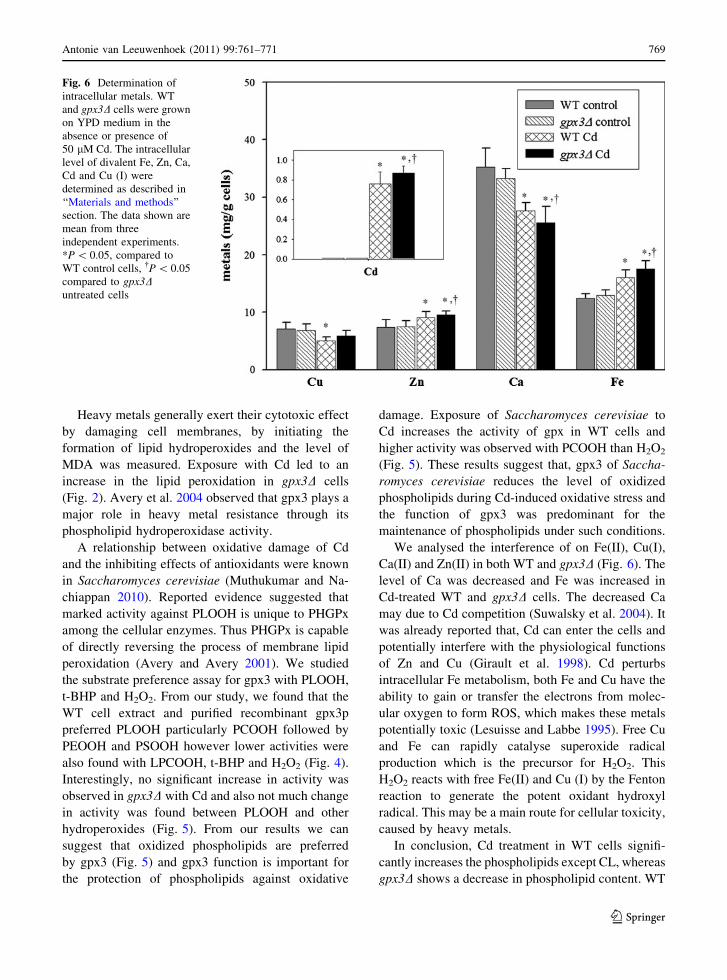
Figure 6 shows the levels of biologically important
metal ions in WT and gpx3D cells. Cd-treated WT
and gpx3D cells showed significant (P \ 0.05)
increase in Fe and Zn. Exposure of WT and gpx3Dcells to Cd showed a decrease in Cu and Ca content
(Fig. 6). WT and gpx3D cells absorb Cd equally.
Discussion
Yeast cells respond to environmental toxicant by
altering the composition of cellular phospholipids
(Vijayaraj et al. 2010). The main aim of this study is to
elucidate the role of gpx3 in protecting phospholipids
against Cd-induced oxidative stress in Saccharomy-
ces cerevisiae. Previously the PHGPx activity was
reported in gpx1, gpx2, and gpx3 of Saccharomy-
ces cerevisiae (Avery and Avery 2001). Gpx3 has
Antonie van Leeuwenhoek (2011) 99:761–771 765
123

Ta
ble
1E
ffec
to
fC
do
nal
tera
tio
no
ffa
tty
acid
com
po
siti
on
iny
east
ph
osp
ho
lip
id
Fat
tyac
ids
(%)
C1
0:0
C1
2:0
C1
4:0
C1
6:0
C1
6:1
C1
8:0
C1
8:1
18
:16
UF
A/S
FA
PC W
Tco
ntr
ol
3.6
8±
0.1
96
.78
±0
.24
13
.64
±0
.86
17
.61
±0
.71
29
.62
±1
.44
17
.83
±1
.56
12
.46
±1
.10
1.0
71
.22
WT
Cd
9.5
3±
0.5
7*
10
.10
±0
.61
*1
8.3
0±
1.1
2*
19
.43
±1
.16
*1
4.6
4±
0.9
3*
22
.31
±1
.24
*6
.51
±0
.11
*1
.14
0.5
0
Gp
x3
con
tro
l4
.21
±0
.36
*7
.13
±0
.42
12
.27
±0
.61
17
.26
±0
.59
28
.13
±1
.12
19
.28
±1
.17
11
.22
±0
.54
*1
.09
1.0
1
Gp
x3
Cd
8.6
2±
0.4
1*
,�
13
.76
±0
.94
*,
�2
1.3
1±
0.5
8*
,�
21
.26
±1
.21
*,
�6
.52
±0
.08
*,
�2
5.7
6±
1.3
1*
,�
4.1
9±
0.1
5*
,�
1.2
10
.22
PE W
Tco
ntr
ol
2.6
0±
0.1
14
.15
±0
.23
8.2
3±
0.4
93
4.6
9±
2.7
21
4.4
2±
2.8
72
1.5
7±
1.6
41
2.1
4±
1.5
40
.62
0.4
7
WT
Cd
4.8
4±
0.2
9*
7.2
1±
0.3
8*
8.7
4±
0.5
64
0.7
1±
3.4
3*
5.6
2±
0.4
6*
26
.61
±1
.45
*5
.36
±0
.31
*0
.65
0.1
6
Gp
x3
con
tro
l3
.11
±0
.16
*5
.42
±0
.36
9.6
8±
0.3
23
5.5
8±
2.1
21
5.7
9±
0.7
12
0.4
3±
1.2
01
1.7
1±
1.2
60
.57
0.4
9
Gp
x3
Cd
6.8
7±
0.3
2*
,�
8.7
4±
0.4
2*
,�
9.2
1±
0.4
14
3.8
3±
2.8
1*
,�
4.4
0±
0.2
5*
,�
25
.79
±1
.71
*,
�4
.36
±0
.23
*,
�0
.58
0.1
2
PS W
Tco
ntr
ol
4.2
1±
0.1
2–
9.1
3±
0.5
13
9.1
5±
2.2
69
.74
±0
.34
17
.25
±1
.35
19
.69
±1
.31
0.4
40
.52
WT
Cd
6.3
4±
0.3
1*
–1
3.6
2±
1.1
2*
43
.91
±3
.42
*5
.31
±0
.12
*2
6.9
3±
2.1
2*
6.2
6±
0.3
4*
0.6
10
.16
Gp
x3
con
tro
l4
.82
±0
.17
–9
.87
±0
.36
38
.72
±2
.34
10
.52
±0
.61
*1
6.6
2±
1.2
11
8.4
8±
1.8
00
.42
0.6
2
Gp
x3
Cd
7.9
6±
0.3
1*
,�
–1
5.3
2±
1.1
5*
,�
44
.73
±2
.69
*,
�3
.84
±1
.22
*,
�2
1.8
4±
1.6
8*
,�
6.7
3±
0.3
5*
,�
0.4
80
.15
PI W
Tco
ntr
ol
–5
.75
±0
.35
18
.33
±1
.24
37
.67
±2
.08
4.2
9±
0.6
92
4.9
6±
1.3
29
.21
±0
.43
0.6
80
.22
WT
Cd
–8
.21
±0
.42
*1
9.2
7±
1.6
73
8.7
6±
2.4
13
.19
±0
.17
*2
6.4
7±
1.5
65
.11
±0
.21
*0
.68
0.1
2
Gp
x3
con
tro
l–
6.2
2±
0.2
81
7.4
5±
1.2
83
6.8
4±
2.3
63
.71
±0
.13
23
.84
±1
.94
10
.79
±0
.62
*0
.59
0.2
6
Gp
x3
Cd
–9
.87
±0
.39
*,
�1
7.8
1±
1.1
64
1.9
6±
2.1
2*
,�
1.5
9±
0.1
1�
22
.74
±2
.11
7.7
3±
0.3
50
.54
0.1
4
CL W
Tco
ntr
ol
1.3
8±
0.1
12
.38
±0
.14
8.2
1±
0.3
42
2.8
7±
1.3
11
7.0
8±
1.1
92
8.1
4±
1.2
52
0.3
4±
1.9
11
.27
0.7
1
WT
Cd
4.6
1±
0.3
6*
4.0
2±
0.2
1*
10
.24
±0
.66
*2
9.6
4±
1.2
8*
5.3
5±
0.2
4*
34
.73
±2
.32
*1
3.8
9±
0.8
2*
1.1
70
.29
Gp
x3
con
tro
l1
.74
±0
.18
2.6
7±
0.1
67
.32
±0
.43
23
.31
±1
.47
*1
6.1
1±
0.7
32
7.2
9±
1.6
41
8.1
3±
1.5
11
.07
0.6
5
Gp
x3
Cd
6.1
2±
0.3
3*
,�
6.9
3±
0.4
3*
,�
12
.73
±0
.82
*,
�3
1.6
7±
1.2
9*
,�
5.2
9±
0.3
1*
,�
32
.53
±1
.42
*,
�5
.72
±0
.33
*,
�1
.27
0.1
7
Ph
osp
ho
lip
ids
wer
eex
trac
ted
fro
mb
oth
con
tro
lan
dC
d-t
reat
edce
lls
and
sub
ject
edto
met
hy
lati
on
wit
hB
F3
-met
han
ol.
Met
hy
late
dfa
tty
acid
sw
ere
anal
yse
db
yG
C/M
S.
Th
e
dat
ash
ow
nar
em
ean
fro
mth
ree
ind
epen
den
tex
per
imen
ts
*P
\0
.05
,co
mp
ared
toW
Tco
ntr
ol
cell
s,�
P\
0.0
5co
mp
ared
tog
px3
Du
ntr
eate
dce
lls.
UF
A-u
nsa
tura
ted
fatt
yac
ids,
SF
A-s
atu
rate
dfa
tty
acid
s
766 Antonie van Leeuwenhoek (2011) 99:761–771
123

been shown to play a crucial role in resistance to
oxidative stress by efficiently reducing the PLOOH
(Avery et al. 2004), however the relationship of gpx3
with phospholipids under oxidative stress conditions is
yet to be studied. Hence we studied here the alteration
of phospholipid levels during gpx3 deficiency and Cd-
induced oxidative stress conditions.
Increased major phospholipid levels were
observed in WT cells when treated with Cd
(Fig. 1). Inversely reduction was observed in gpx3Dcells and further decreased with Cd, This suggests the
role of gpx3 in the maintenance of phospholipids
under oxidative stress. Previously it was reported that
gpx3D was hypersensitive to Cd (Avery et al. 2004).
We observed that gpx3D cells showed a decrease in
growth rate when exposed to Cd (data not shown).
Reported evidence suggested that increase in the
phospholipid content was shown to be an important
factor in providing tolerance against organic solvents
(Ghosh et al. 2008). Our results suggest that increased
phospholipid levels in WT cells (Fig. 1) may be
required to resist the Cd-induced oxidative stress.
Stress tolerance mechanism is known to be associated
with increased phospholipid biosynthesis in Saccha-
romyces cerevisiae (Ghosh et al. 2008). Phospholip-
ids were substantially decreased in gpx3D control
cells, thus indicating the partial loss or damage of
phospholipids in the absence of gpx3 and this was
further reduced on Cd-treatment thereby, leading to
major compositional changes (Fig. 1). Previously it
was reported that the deficiency of antioxidants, leads
to marked decrease in the levels of PC and also
leading to mitochondrial damage (Jain et al. 1992).
When treated with Cd the levels of CL were
decreased in both WT and gpx3D cells. Mitochondria
is an important cellular source of O2•- and H2O2
(Lambert and Brand 2009) and are intracellular
targets for different stressors including Cd (Sanni
et al. 2008). Mechanisms of metal-induced mito-
chondrial damage are not fully understood. From our
observation we can suggest that, in the absence of
gpx3, cells were unable to protect phospholipids in
Cd-treated and untreated conditions, as a result
phospholipid content was decreased.
Our phospholipid data suggested that gpx3 is linked
with maintenance of cellular phospholipids during Cd-
induced oxidative stress. The functions of phospholip-
ids mainly depend on its fatty acyl composition.
Previously it was reported that the fatty acid
Fig. 2 Effect of Cd on formation of MDA. WT and gpx3Dcells were grown up to midlog phase on YPD medium in the
absence or presence of 50 lM Cd. The level of MDA was
determined in WT and gpx3D cells as described in ‘‘Materials
and methods’’ section. The data shown are mean from three
independent experiments. *P \ 0.05, compared to WT control
cells, �P \ 0.05 compared to gpx3D untreated cells
Fig. 3 Purification of recombinant gpx3p. The gpx3p from E.coli BL21 (DE3) was purified using nickel-nitrilotriacetic acid
affinity column. Induced gpx3p was visualized by coomassie
blue-staining of 10% SDS-polyacrylamide gel. M molecular
weight marker, Lane 1 isopropyl b-D-1-thiogalactopyranoside
(IPTG) uninduced, Lane 2 induced with 0.4 mM IPTG, Lane3 Ni2?–NTA purified gpx3p
Antonie van Leeuwenhoek (2011) 99:761–771 767
123

composition of CL plays an important role in the
function of the lipid, as aberrant CL remodelling causes
Barth syndrome (Schlame et al. 2005). To understand
molecular species alterations in the absence of gpx3,
we studied the alterations of fatty acyl composition of
phospholipids. Exposure of WT and gpx3D cells to Cd
significantly increases short chain (C10:0), and satu-
rated fatty acids (C16:0, C18:0) in all phospholipids
(Table 1). Oxidative degradation of unsaturated fatty
acids was supported by the lower cellular fatty acid
unsaturation indices evident during exponential
growth in the presence of Cd; depletion of unsaturated
fatty acids is a useful marker of lipid peroxidation (De
Vos et al. 1993). We also found higher reduction in
unsaturated fatty acids (C16:1 and C18:1) (Table 1).
Previous studies with higher organisms have indicated
that susceptibility to heavy-metal toxicity may be
partly dependent on the lipid composition and physical
properties of cellular membranes (e.g. Vossen et al.
1995; De Vos et al. 1993). Cd is suspected to induce the
production of ROS which can attack and damage
biological molecules, including DNA, proteins and
lipids (Esterbauer et al. 1991). Polyunsaturated fatty
acids are esterified in membrane or storage lipids are
subjected to ROS-induced peroxidation and may
yield cytotoxic aldehydes, like 4-hydroxy-2-nonenal
(HNE), MDA and acrolein (Esterbauer et al. 1991).
Fig. 4 Substrate preference of the gpx3p The activities of gpx
from WT, gpx3D cells extract and purified gpx3p are expressed
as U/mg protein. One unit of activity is defined as the amount
of protein required to oxidize 1 lMol of NADPH/min at 25�C.
PCOOH oxidized phosphatidylcholine; PEOOH oxidized
phosphatidylethanolamine, PSOOH oxidized phosphatidylser-
ine, LPCOOH oxidized lysophosphotidylcholine, t-BHP tert
butyl hydro peroxide, H2O2 hydrogen peroxide. The data
shown are mean from three independent experiments.
*P \ 0.05, compared to WT control cells
Fig. 5 Gpx3 activities in Saccharomyces cerevisiae. The
enzyme activity was assayed with PCOOH or H2O2 in WT
and gpx3D cells that were grown on YPD medium in the
absence or presence of 50 lM Cd. Activity of gpx was
expressed as U/mg protein. One unit of activity was defined as
the amount of protein required to oxidize 1 lMol of NADPH/
min at 25�C. The data shown are mean from three independent
experiments. *P \ 0.05, compared to WT control cells,�P \ 0.05 compared to gpx3D untreated cells
768 Antonie van Leeuwenhoek (2011) 99:761–771
123
Heavy metals generally exert their cytotoxic effect
by damaging cell membranes, by initiating the
formation of lipid hydroperoxides and the level of
MDA was measured. Exposure with Cd led to an
increase in the lipid peroxidation in gpx3D cells
(Fig. 2). Avery et al. 2004 observed that gpx3 plays a
major role in heavy metal resistance through its
phospholipid hydroperoxidase activity.
A relationship between oxidative damage of Cd
and the inhibiting effects of antioxidants were known
in Saccharomyces cerevisiae (Muthukumar and Na-
chiappan 2010). Reported evidence suggested that
marked activity against PLOOH is unique to PHGPx
among the cellular enzymes. Thus PHGPx is capable
of directly reversing the process of membrane lipid
peroxidation (Avery and Avery 2001). We studied
the substrate preference assay for gpx3 with PLOOH,
t-BHP and H2O2. From our study, we found that the
WT cell extract and purified recombinant gpx3p
preferred PLOOH particularly PCOOH followed by
PEOOH and PSOOH however lower activities were
also found with LPCOOH, t-BHP and H2O2 (Fig. 4).
Interestingly, no significant increase in activity was
observed in gpx3D with Cd and also not much change
in activity was found between PLOOH and other
hydroperoxides (Fig. 5). From our results we can
suggest that oxidized phospholipids are preferred
by gpx3 (Fig. 5) and gpx3 function is important for
the protection of phospholipids against oxidative
damage. Exposure of Saccharomyces cerevisiae to
Cd increases the activity of gpx in WT cells and
higher activity was observed with PCOOH than H2O2
(Fig. 5). These results suggest that, gpx3 of Saccha-
romyces cerevisiae reduces the level of oxidized
phospholipids during Cd-induced oxidative stress and
the function of gpx3 was predominant for the
maintenance of phospholipids under such conditions.
We analysed the interference of on Fe(II), Cu(I),
Ca(II) and Zn(II) in both WT and gpx3D (Fig. 6). The
level of Ca was decreased and Fe was increased in
Cd-treated WT and gpx3D cells. The decreased Ca
may due to Cd competition (Suwalsky et al. 2004). It
was already reported that, Cd can enter the cells and
potentially interfere with the physiological functions
of Zn and Cu (Girault et al. 1998). Cd perturbs
intracellular Fe metabolism, both Fe and Cu have the
ability to gain or transfer the electrons from molec-
ular oxygen to form ROS, which makes these metals
potentially toxic (Lesuisse and Labbe 1995). Free Cu
and Fe can rapidly catalyse superoxide radical
production which is the precursor for H2O2. This
H2O2 reacts with free Fe(II) and Cu (I) by the Fenton
reaction to generate the potent oxidant hydroxyl
radical. This may be a main route for cellular toxicity,
caused by heavy metals.
In conclusion, Cd treatment in WT cells signifi-
cantly increases the phospholipids except CL, whereas
gpx3D shows a decrease in phospholipid content. WT
Fig. 6 Determination of
intracellular metals. WT
and gpx3D cells were grown
on YPD medium in the
absence or presence of
50 lM Cd. The intracellular
level of divalent Fe, Zn, Ca,
Cd and Cu (I) were
determined as described in
‘‘Materials and methods’’
section. The data shown are
mean from three
independent experiments.
*P \ 0.05, compared to
WT control cells, �P \ 0.05
compared to gpx3Duntreated cells
Antonie van Leeuwenhoek (2011) 99:761–771 769
123

and gpx3D cells showed an increase in short chain fatty
acids, at the same time the levels of unsaturated fatty
acids were decreased. Gpx3 might be required for the
protection of phospholipids during oxidative stress.
Cd-treatment increases the gpx activity in WT cells
whereas no such change was observed in gpx3D cells.
The results obtained from the study indicate that gpx3
might protect the phospholipids from Cd-induced
oxidative damage in Saccharomyces cerevisiae. Fur-
ther investigations must be carried out to understand
the relationship between phospholipid biosynthetic/
degradative enzymes and gpx3 during Cd-induced
oxidative stress.
Acknowledgement The financial support from Bharathidasan
University, Tiruchirappalli, Tamilnadu, India is gratefully
acknowledged.
Conflict of interest None.
References
Aguilar PS, de Mendoza D (2006) Control of fatty acid
desaturation: a mechanism conserved from bacteria to
humans. Mol Microbiol 62:1507–1514
Arthur JR (2000) The glutathione peroxidises. Cell Mol Life
Sci 57:1825–1835
Avery AM, Avery SV (2001) Saccharomyces cerevisiaeexpresses three phospholipid hydroperoxide glutathione
peroxidases. J Biol Chem 276:33730–33735
Avery AM, Willetts SA, Avery SV (2004) Genetic dissection
of the phospholipid hydroperoxidase activity of yeast
gpx3 reveals its functional importance. J Biol Chem
279:46652–46658
Bligh EG, Dyer WJ (1959) A rapid method of total lipid
extraction and purification. Can J Biochem Physiol 37:
911–917
Bradford MM (1976) A rapid and sensitive method for the
quantitation of microgram quantities of protein utilizing
the principle of protein-dye binding. Anal Biochem
72:248–254
Carman GM, Han GS (2009) Regulation of phospholipid bio-
synthesis in yeast. J Lipid Res 50:69–73
De Vos CHR, Bookum WMT, Vooijs R, Schat H, De Kok LJ
(1993) Effects of copper on fatty acid composition and
peroxidation of lipids in the roots of copper tolerant and
sensitive Silene cucubalas. Plant Physiol Biochem 31:
151–158
Doner G, Ege A (2004) Evaluation of digestion procedures of the
determination of iron and zinc in biscuits by flame atomic
absorption spectrometry. Anal Chim Acta 520:217–222
Esterbauer H, Schaur RJ, Zollner H (1991) Chemistry and
biochemistry of 4-hydroxy nonenal, malonaldehyde and
related aldehydes. Free Radic Biol Med 11:81–128
Falcone DL, Ogas JP, Somerville CR (2004) Regulation of
membrane fatty acid composition by temperature in
mutants of Arabidopsis with alterations in membrane lipid
composition. BMC Plant Biol 4:17
Ghosh AK, Ramakrishnan G, Rajasekharan R (2008)
YLR099C (ICT1) encodes a soluble acyl-CoA-dependent
lysophosphatidic acid acyltransferase responsible for
enhanced phospholipid synthesis on organic solvent
stress in Saccharomyces cerevisiae. J Biol Chem 283:
9768–9775
Girault L, Boudou A, Dufourc E (1998) 113Cd-, 31P-NMR and
fluorescence polarization studies of cadmium (II) inter-
actions with phospholipids in model membranes. Biochim
Biophys Acta 1414:140–154
Girotti W (1985) Mechanisms of lipid peroxidation. Free Radic
Biol Med 1:87–95
Halliwell B, Gutteridge JMC (1999) Free radicals in biology
and medicine, 3rd edn. Oxford University Press, Oxford
Iwanyshyn WM, Han GS, Carman GM (2004) Regulation of
phospholipid synthesis in Saccharomyces cerevisiae by
zinc. J Biol Chem 279:21976–21983
Jain A, Martensson J, Mehta T, Krauss AN, Auld PA, Meister
A (1992) Ascorbic acid prevents oxidative stress in glu-
tathione-deficient mice: effects on lung type 2 cell
lamellar bodies, lung surfactant, and skeletal muscle.
PNAS 89:5093–5097
Kriska T, Girotti A (2004) Separation and quantitation of
peroxidized phospholipids using high-performance thin-
layer chromatography with tetramethyl-P-phenylenedi-
amine detection. Anal Biochem 327:97–106
Laemmli UK (1970) Cleavage of structural proteins during the
assembly of the head of bacteriophage T4. Nature 227:
680–685
Lambert AJ, Brand MD (2009) Reactive oxygen species pro-
duction by mitochondria. Methods Mol Biol 554:165–181
Lesuisse E, Labbe P (1995) Effects of cadmium and of YAP1and CAD1/YAP2 genes on iron metabolism in the yeast
Saccharomyces cerevisiae. Microbiology 141:2937–2943
Maiorino M, Gregolin C, Ursini F (1990) Phospholipid
hydroperoxide glutathione peroxidase. Methods Enzymol
186:448–457
Martin CE, Oh CS, Jiang Y (2007) Regulation of long chain
unsaturated fatty acid synthesis in yeast. Biochim Biophys
Acta 1771:271–285
Morrison WR, Smith LM (1964) Preparation of fatty acid
methyl esters and dimethylacetals from lipids with boron
fluoride methanol. J Lipid Res 5:600–608
Muthukumar K, Nachiappan V (2010) Cadmium-induced
oxidative stress in Saccharomyces cerevisiae. Indian J
Biochem Biophys 47:383–387
Patrick L (2003) Toxic metals and antioxidants: part II. The
role of antioxidants in arsenic and cadmium toxicity.
Altern Med Rev 8:106–128
Pinkart HC, White DC (1997) Phospholipid biosynthesis and
solvent tolerance in Pseudomonas putida strains. J Bacte-
riol 179:4219–4226
Rattray JB, Schibeci A, Kidby DK (1975) Lipids of yeast.
Bacteriol Rev 39:197–231
Sanni B, Williams K, Sokolov EP, Sokolova IM (2008) Effects
of acclimation temperature and cadmium exposure on
mitochondrial aconitase and LON protease from a model
770 Antonie van Leeuwenhoek (2011) 99:761–771
123

marine ectotherm, Crassostrea virginica. Comp Biochem
Physiol 147:101–112
Schlame M, Ren M, Xu Y, Greenberg ML, Haller I (2005)
Molecular symmetry in mitochondrial cardiolipins. Chem
Phys Lipids 138:38–49
Siakotos AN, Rouser G, Fleischer S (1966) Phospholipid
composition of human, bovine and frog myelin isolated on
a large scale from brain and spinal cord. Lipids 1:85–86
Steels EL, Learmonth RP, Watson K (1994) Stress tolerance
and membrane lipid unsaturation in Saccharomyces ce-revisiae grown aerobically or anaerobically. Microbiology
140:569–576
Steffensen L, Mesna OJ, Andruchow E, Namork E, Hylland K,
Andersen RA (1994) Cytotoxicity and accumulation of
Hg, Ag, Cd, Cu, Pb and Zn in human peripheral T and B
lymphocytes and monocytes in vitro. Gen Pharmacol
251:621–1633
Sumner ER, Shanmuganathan A, Sideri TC, Willetts SA,
Houghton JE, Avery SV (2005) Oxidative protein damage
causes chromium toxicity in yeast. Microbiology 151:
1939–1948
Suwalsky M, Villena F, Norris B, Cuevas F, Sotomayor CP
(2004) Cadmium-induced changes in the membrane of
human erythrocytes and molecular models. J Inorg Bio-
chem 98:1061–1066
Temple MD, Perrone GG, Dawes IW (2005) Complex cellular
responses to reactive oxygen species. Trends Cell Biol
15:319–326
Thorsen M, Perrone GG, Kristiansson E, Traini M, Yel T,
Dawes IW, Nerman O, Tamas MJ (2009) Genetic basis of
arsenite and cadmium tolerance in Saccharomyces cere-visiae. BMC Genomics 10:105–120
Ursini F, Maiorino M, Brigelius-Flohe R, Aumann KD, Roveri
A, Schomburg D, Flohe L (1995) Diversity of glutathione
peroxidises. Methods Enzymol 252:38–53
Vijayaraj P, Sabarirajan J, Nachiappan V (2010) Enhanced
phospholipase B activity and alteration of phospholipids
and neutral lipids in Saccharomyces cerevisiae exposed to
N-nitrosonornicotine. Antonie van Leeuwenhoek. doi:
10.1007/s10482-010-9526-1
Vossen RCRM, Dam-Mieras MCE, van Hornstra G, Zwaal
RFA (1995) Differential effects of endothelial cell fatty
acid modification on the sensitivity of their membrane
phospholipids to peroxidation. Prostaglandins Leukot
Essent Fatty Acids 52:341–347
Wagner S, Paltauf F (1994) Generation of glycerophospholipid
molecular species in the yeast Saccharomyces cerevisiae.
Fatty acid pattern of phospholipid classes and selective acyl
turnover at sn-1 and sn-2 positions. Yeast 10:1429–1437
Wolff SP (1994) Ferrous oxidation in presence of ferric ion
indicator xylenol orange for measurement of hydroper-
oxides. Methods Enzymol 233:182–189
Inoue Yoshiharu, Matsuda T, Sugiyama K-I, Izawa S, Kimura
A (1999) Genetic analysis of glutathione peroxidase in
oxidative stress response of Saccharomyces cerevisiae.
J Biol Chem 274:27002–27009
Antonie van Leeuwenhoek (2011) 99:761–771 771
123




















