
Influence of squid extracts on the triggering of secondary vitellogenesis in Penaeus vannamei
Upload
independentCategory
view
1download
0
Effects of age, size, and light intensity on spawning
performance of pond-reared Penaeus merguiensis
Tung Hoang a,*, S.Y. Lee a, Clive P. Keenan b, Gay E. Marsden c
aSchool of Environmental and Applied Sciences, Griffith University, PMB 50 GCMC, QLD 9726, AustraliabBribie Island Aquaculture Research Centre, QDPI, PO Box 2066, QLD 4507, Australia
cNFIEC, Locked Bag 5, Grafton, NSW 2460, Australia
Received 3 October 2001; received in revised form 20 December 2001; accepted 6 January 2002
Abstract
The effects of prawn age, size, and light intensity on spawning performance of pond-reared
Penaeus merguiensis were studied by inducing spawning of prawn broodstock by eyestalk ablation
under two different light intensities (bright, 1100 lx and dim, 2 lx) at three different known ages (7,
10, and 13 months old). For each age group (except the 13-month-old group), two distinguished size
classes (large and small) were established to reveal the effect of prawn size. Results showed that light
intensity did not affect spawning performance of prawns of different ages and sizes (P > 0.05). In
contrast, age had a significant effect on spawning performance of prawns (P < 0.01). The overall
spawning performance of the prawns increased from 7 to 10 months old and started to decline at 13
months old. Within the 7- and 10-month-old groups, larger prawns produced a superior overall
spawning performance compared with smaller ones (P< 0.01). Comparison across the size range of
the experimental prawns showed that spawning performance improved with the increase of body
size. However, the 13-month-old prawns, although having a similar size as large 10-month-old
prawns, had an inferior spawning performance (P< 0.01). Survival and growth of ablated prawns
significantly decreased with the increase of prawn size and age (P < 0.01). It is suggested that
spawning performance of pond-reared P. merguiensis peaked between 10 and 12 months old, and
broodstock should be bred during this period for hatchery production.
D 2002 Elsevier Science B.V. All rights reserved.
Keywords: Age; Size; Light intensity; Spawning performance; Pond-reared broodstock; Penaeus merguiensis
0044-8486/02/$ - see front matter D 2002 Elsevier Science B.V. All rights reserved.
PII: S0044 -8486 (02 )00019 -4
* Corresponding author. Bribie Island Aquaculture Research Centre, Queensland Department of Primary
Industries (QDPI), PO Box 2066, 144 North Street, Qld 4507, Australia. Tel.: +61-7-3400-2020; fax: +61-7-
3408-3535.
E-mail addresses: [email protected], [email protected] (T. Hoang).
www.elsevier.com/locate/aqua-online
Aquaculture 212 (2002) 373–382

1. Introduction
The effect of broodstock size on spawning performance has been documented for
several penaeid species (Motoh, 1981; Primavera, 1985; Menasveta et al., 1994; Cavalli et
al., 1997; Crocos and Coman, 1997). Generally, larger broodstock tend to produce a
superior spawning performance than do smaller ones. This information, however, is not
available for the banana prawn, Penaeus merguiensis, which is farmed in Southeast Asia
and Australia but not to the same extent as P. monodon. Furthermore, the lack of accurate
methods to determine crustacean age has limited information about the effect of age on
reproductive performance in penaeids. As experimental prawns have been widely
collected from the wild, larger prawns are often assumed to be older ones. However,
prawns that have the same size may not be necessarily of the same age. In crustacean
fisheries management, length–frequency analysis is commonly used to study the age
structure or recruitment of a population of interest (Thomas 1975; Somers and Kirkwood
1991; Guerao and Ribera, 1995; Roa and Ernst, 1996). Based on the age structure of
known populations, Crocos and Coman (1997) and Cavalli et al. (1997) estimated the age
of wild P. semisulcatus and P. paulensis, respectively, and compared spawning perform-
ance among the age groups. Results showed a correlation between spawning performance
and prawn age. Needless to say, such studies require a thorough understanding of the
detailed reproductive biology of the population from which samples are collected.
Furthermore, separating old cohorts by size is difficult and may lead to misclassification
(Somers and Kirkwood, 1991). Using the brain pigment, lipofuscin, to estimate the age of
crustaceans is a promising tool. However, the general paucity of lipofuscin in penaeid
prawns has been noted (Sheehy, 1990), suggesting a combination with cohort analysis is
necessary for accurate estimation of prawn age (Sheehy et al., 1995). Alternatively,
comparison can be made between age groups by growing prawns in a captive environment
and inducing them to spawn at different known ages. This approach is feasible with P.
merguiensis, which has been observed to mature and spawn in culture ponds (Lichatowich
et al., 1978; Hoang et al., in press (a)) and will help further understand the effect of age on
reproductive performance of prawns.
Light intensity has been reported to influence ovarian maturation and spawning perform-
ance of penaeid prawns (Primavera, 1985; Bray and Lawrence, 1992; Browdy, 1992). Dim
light has beenwidely used in breeding trials or in commercial prawn hatcheries, where prawn
broodstock are mostly ablated. However, our recent study on P. merguiensis revealed that
strong light intensity (1100 lx) did not affect ovarian maturation and spawning of ablated
prawns, compared with dim light (2 lx) (Hoang et al., in press (b)). It was assumed that the
strong effect of eyestalk ablation had overridden the effect of light intensity in ablated prawns
since strong light (1100 lx) has been reported to inhibit ovarian maturation of unablated P.
merguiensis (Hoang et al., in press (c)). Alternatively, the effect of light intensitymay change
according to prawn age. The ablated prawns used in our previous study were approximately
10–11 months old and may have been ready for reproduction. It is hypothesized that light
intensity may affect ovarian maturation and spawning of younger and/or smaller ablated
prawns, which presumably are less ready for reproduction.
In this paper, the effect of prawn age, size, and light intensity on spawning performance
of pond-reared P. merguiensis is reported.
T. Hoang et al. / Aquaculture 212 (2002) 373–382374

2. Materials and methods
2.1. Broodstock culture conditions
PL25 (postlarvae, 25 days old) were produced from 10 gravid females, originally collected
from local waters in southeast Queensland, Australia and grown to maturity in ponds at the
Bribie Island Aquaculture Research Centre (BIARC). These PL25 were cultured in two 60-
m3 tanks during the first 4 months and then transferred to a 200-m2 pond until the end of the
study. This pond was covered with transparent roofing material to keep pond water warm
during winter. Basic water quality parameters were checked twice daily at 09:00 and 16:00 h
using a U10 Water Checker (Horiba, Japan). During the study, water temperature, pH,
salinity, and dissolved oxygen (DO) varied between 19.9 and 32.7 jC, 7.0 and 9.0, 30.5 and35.6 ppt, and 4.8 and 10.9 mg/l, respectively. Water exchange (approximately 30%) was
conducted twice a week, but limited to 20–30% a week during winter. A good algae bloom
(predominantly diatoms, secchi disk reading < 40 cm) was maintained continually through-
out the study. Prawns were fed with EBI STAR pellets (50% crude protein, Higashimaru,
Japan) at approximately 5% of prawn biomass/day.
2.2. Breeding trials
Female prawns were collected from the culture pond and induced to spawn by eyestalk
ablation at three different ages: 7, 10, and 13 months old (from hatching). For each age
group (except the 13-month-old group), two size classes were distinguished by carapace
length and body weight. Thirty large and 30 small prawns were collected from the pond
and stocked in 12 250-l tanks (measuring 90� 60� 55 cm) at a stocking density of five
prawns/tank. Body size was similar among the 13-month-old prawns. Thus, only 30 13-
month-old prawns were collected from the pond. Two light intensity treatments were
applied in each breeding trial: bright light (1100 lx) and dim light (2 lx). Each combination
of age, size, and light intensity, therefore, had three replicate tanks. Light intensity was
measured at the tank water surface using a photometer (Model 185B, LI-COR, USA).
Light was provided by daylight fluorescent tubes (OSRAM 36 W, Australia) suspended on
top of the tanks. For the dim light treatment, green 70% shade cloth (Dindas Lew Cat No
5c7036 BL) was used to wrap the light tubes to reduce light intensity.
Broodstock selection, eyestalk ablation technique, maturation conditions, feeding,
spawning assessment, and tank management were standardized for all the three breeding
trials. All the selected prawns were healthy in appearance. Injured and post-moult females
were excluded and no prawn introduced to the tanks possessed developed ovaries. Prawns
were acclimated in the experimental tanks for 3 days before being ablated. Eyestalk
ablation involved ligating, cutting the eyeball, and squeezing out all the contents. During
this handling, the prawns were kept in cool seawater (16–18 jC) to prevent stress. Prawns
were tagged with a numbered ring on the remaining eyestalk to facilitate the recording of
spawning performance of each individual.
The study used a flow-through system. Filtered seawater (5 Am) was heated and fed by
gravity to the experimental tanks, exchanging at a rate of 300%/day. Water temperature (in
the experimental tanks) and photoperiod were maintained at 27 jC and 12L:12D, respec-
T. Hoang et al. / Aquaculture 212 (2002) 373–382 375

tively. Salinity and pH varied from 33.5 to 34.6 and 7.9 to 8.2, respectively, and were not
different among the three breeding trials. All tanks were aerated to keep DO above 5 mg/l.
Prawns were fed with the BIARC moist artificial maturation diet (a mixture of fresh,
frozen green mussels, squids, calf liver, and other essential nutrients, 54.6% crude protein;
Marsden et al., 1997) at 10% of prawn biomass and EBI STAR pellets at 2% of prawn
biomass. Feeding was conducted twice a day at 09:00 and 17:00 h. Pellets were only given
in the later feeding. All tanks were cleaned before feeding time.
Ovarian maturation was checked daily at 17:30 h, following the descriptions by Tuma
(1967) and Primavera (1985). Stage-IV females (full ovarian maturation) were individually
transferred to 150-l spawning tanks and returned to the original tanks next morning.
Sampling for fecundity was conducted by vigorously agitating the tank water to ensure a
homogeneous suspension of eggs, then quickly taking four 100-ml aliquot samples and
counting the number of eggs under a stereo microscope. In all the breeding trials, prawns
were kept in tanks for 21 days from eyestalk ablation.
2.3. Statistical analysis
Variables used to assess spawning performance included the proportion of spawners,
latency period (the number of days from eyestalk ablation to the first spawn), spawning
rate (number of spawns/female/day), absolute fecundity (eggs/spawn), relative fecundity
(eggs/g initial body weight), and egg production/female. Survival and growth (specific
growth rate, % weight gain/day) were also taken into account. No attempt was made to
investigate hatching rate and survival of prawn larvae due to the lack of time and larval
rearing facilities required for adequate replication. Nonetheless, viable nauplii were
observed throughout all the breeding trials. Larvae from several spawns of the 10-
month-old prawns were nursed and cultured to broodstock size in ponds at BIARC.
Experimental data were analyzed by Genstat 4.1 for Windows (Payne et al., 1993).
Two-way analysis of variance (ANOVA) was initially employed to separate the effect of
two factors: age and light intensity. The results showed that only age had a significant
effect (P < 0.01) on spawning performance of the prawns. Therefore, data of the two light
intensity treatments were pooled for further examination on the effect of prawn size. One-
way ANOVA with least significant difference procedure (LSD) was employed as the
experiment had an incomplete factorial design (i.e. there was only one size class in the 13-
month-old group). Overall, five age and size combinations were formed for the analysis:
7L (7 months old, large class), 7S (7 months old, small class), 10L (10 months old, large),
10S (10 months old, small), and 13 (13 months old). Generalized linear model (modelling
of binomial proportion by logits) was used to analyze binomial data such as the proportion
of spawners and survival (McCullagh and Nelder, 1989).
3. Results
3.1. Effect of light intensity
Light intensity did not significantly (P>0.05) affect the spawning performance of pond-
reared P. merguiensis. The proportion of spawners, survival of prawns, latency period,
T. Hoang et al. / Aquaculture 212 (2002) 373–382376

number of spawns/female/day, fecundity, egg production, and SGR were similar between
the two light intensity treatments in all the age groups (Fig. 1). No interaction between
light intensity and prawn age was detected.
Fig. 1. Spawning performance, survival, and growth of ablated pond-reared P. merguiensis under different light
intensities at different ages. Data are meansF S.E. Bars that have the same letter are not significantly different
( P>0.05).
T. Hoang et al. / Aquaculture 212 (2002) 373–382 377

3.2. Effect of age
Age had a significant effect (P < 0.01) on the spawning performance of pond-reared P.
merguiensis (Fig. 1). The best spawning performance was achieved with the 10-month-old
group. The proportion of spawners, number of spawns/female/day, fecundity, and egg
production/female were significantly (P < 0.01) higher in the 10- and 13-month-old
groups. Prawns in these groups also had a significantly shorter latency period (8.5 and
6.8 days, respectively) compared with the 7-month-old group (14.0 days), suggesting that
they were more ready for reproduction. In the 7-month-old group, only 35% of the prawns
spawned, producing only 0.026 spawns/female/day. The proportion of spawners and
number of spawns/female/day significantly increased to 76.7% and 0.095, respectively, in
the 10-month-old group before declining to 60% and 0.075 spawns/female/day, respec-
tively, in the 13-month-old group. The absolute fecundity (eggs/spawn) increased
significantly (P < 0.01) as prawn age increased. The absolute fecundity was highest in
the 13-month-old group (116.5� 103), followed by the 10-month-old group (93.3� 103),
and lowest in the 7-month-old group (27.4� 103) (Fig. 1). However, there was no
significant difference (P>0.05) in relative fecundity (eggs/g body weight) between the
13-month-old group (3.38� 103/g) and the 10-month-old group (2.96� 103/g).
Survival and growth of the prawns significantly declined as they grew older (P < 0.01)
(Fig. 1). The 13-month-old group had the lowest survival rate of only 53%. Prawns at this
age, although healthy in appearance, were observed to die either after spawning or a few
days after eyestalk ablation. Positive growth (SGR) was only observed in the 7-month-old
group, significantly higher than the 10- and 13-month-old groups (Fig. 1).
The difference in spawning performance among the age groups was not only attributed
to prawn age, but also prawn size. As the prawns grew older, their body sizes also
increased (Table 1).
3.3. Effect of size
The difference in spawning performance between the two nominal size classes within
each age group was observed for a number of variables (Table 1). Generally, at the same
age, larger prawns had a better overall spawning performance, but lower survival and/or
lower growth rate than smaller prawns. At both 7- and 10-months old, the larger size class
(7L and 10L) had a significantly (P < 0.01) higher proportion of spawners than the small
size class (7S and 10S). The 7L group also produced a significantly (P>0.01) higher
spawning rate (0.044 spawns/female/day) than the 7S group (0.008 spawns/female/day).
At this age, no significant difference was found in the latency period, fecundity (both
absolute and relative), egg production/female, and survival. Smaller prawns, however, had
a significantly higher growth rate (SGR) than larger prawns. Between the 10L and 10S
groups, spawning rate and relative fecundity were not significant (P>0.05). However, the
10L group had a higher absolute fecundity (due to larger size), higher egg production/
female (P < 0.01), but lower survival (P < 0.01) (Table 1).
Comparisons across the five combinations of age and size showed that the overall
spawning performance of prawns increased with the increase in prawn size (from the 7S to
the 10L). The 13-month-old group had a similar size as the 10L group, but had a
T. Hoang et al. / Aquaculture 212 (2002) 373–382378
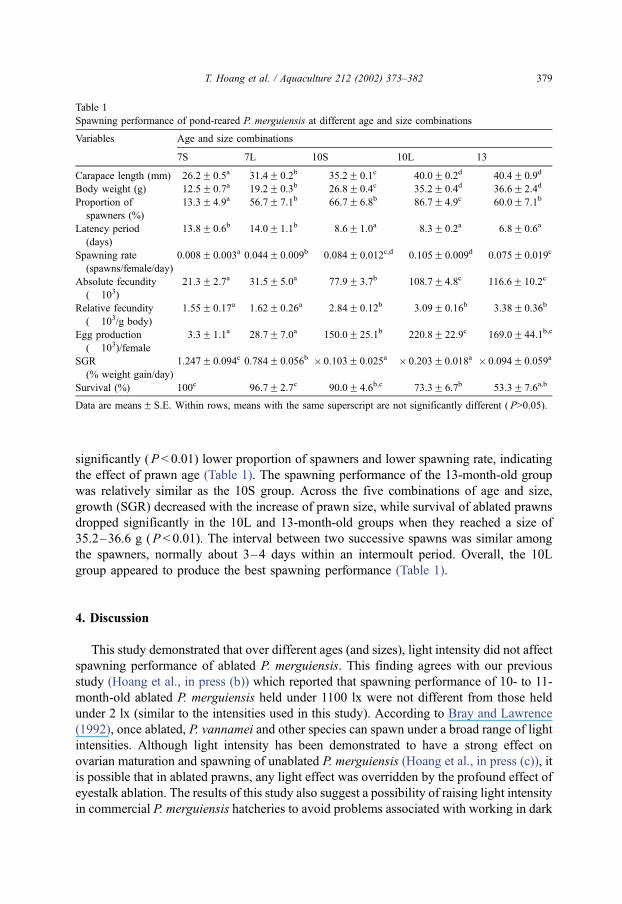
significantly (P < 0.01) lower proportion of spawners and lower spawning rate, indicating
the effect of prawn age (Table 1). The spawning performance of the 13-month-old group
was relatively similar as the 10S group. Across the five combinations of age and size,
growth (SGR) decreased with the increase of prawn size, while survival of ablated prawns
dropped significantly in the 10L and 13-month-old groups when they reached a size of
35.2–36.6 g (P < 0.01). The interval between two successive spawns was similar among
the spawners, normally about 3–4 days within an intermoult period. Overall, the 10L
group appeared to produce the best spawning performance (Table 1).
4. Discussion
This study demonstrated that over different ages (and sizes), light intensity did not affect
spawning performance of ablated P. merguiensis. This finding agrees with our previous
study (Hoang et al., in press (b)) which reported that spawning performance of 10- to 11-
month-old ablated P. merguiensis held under 1100 lx were not different from those held
under 2 lx (similar to the intensities used in this study). According to Bray and Lawrence
(1992), once ablated, P. vannamei and other species can spawn under a broad range of light
intensities. Although light intensity has been demonstrated to have a strong effect on
ovarian maturation and spawning of unablated P. merguiensis (Hoang et al., in press (c)), it
is possible that in ablated prawns, any light effect was overridden by the profound effect of
eyestalk ablation. The results of this study also suggest a possibility of raising light intensity
in commercial P. merguiensis hatcheries to avoid problems associated with working in dark
Table 1
Spawning performance of pond-reared P. merguiensis at different age and size combinations
Variables Age and size combinations
7S 7L 10S 10L 13
Carapace length (mm) 26.2F 0.5a 31.4F 0.2b 35.2F 0.1c 40.0F 0.2d 40.4F 0.9d
Body weight (g) 12.5F 0.7a 19.2F 0.3b 26.8F 0.4c 35.2F 0.4d 36.6F 2.4d
Proportion of
spawners (%)
13.3F 4.9a 56.7F 7.1b 66.7F 6.8b 86.7F 4.9c 60.0F 7.1b
Latency period
(days)
13.8F 0.6b 14.0F 1.1b 8.6F 1.0a 8.3F 0.2a 6.8F 0.6a
Spawning rate
(spawns/female/day)
0.008F 0.003a 0.044F 0.009b 0.084F 0.012c,d 0.105F 0.009d 0.075F 0.019c
Absolute fecundity
(� 103)
21.3F 2.7a 31.5F 5.0a 77.9F 3.7b 108.7F 4.8c 116.6F 10.2c
Relative fecundity
(� 103/g body)
1.55F 0.17a 1.62F 0.26a 2.84F 0.12b 3.09F 0.16b 3.38F 0.36b
Egg production
(� 103)/female
3.3F 1.1a 28.7F 7.0a 150.0F 25.1b 220.8F 22.9c 169.0F 44.1b,c
SGR
(% weight gain/day)
1.247F 0.094c 0.784F 0.056b � 0.103F 0.025a � 0.203F 0.018a � 0.094F 0.059a
Survival (%) 100c 96.7F 2.7c 90.0F 4.6b,c 73.3F 6.7b 53.3F 7.6a,b
Data are meansF S.E. Within rows, means with the same superscript are not significantly different ( P>0.05).
T. Hoang et al. / Aquaculture 212 (2002) 373–382 379

or very dim conditions. The hypothesis that the effect of light intensity varies with ages (and
sizes) of the prawns in this study was, therefore, rejected.
This study demonstrated a significant effect of prawn age and size on spawning
performance of pond-reared P. merguiensis. Older and larger prawns were more ready
for reproduction than smaller and younger prawns as indicated by the shorter latency
periods and higher proportion of spawners in these groups. They also spawned more
frequently and produced more eggs. The overall spawning performance of the 7-month-old
group was inferior compared with the 10- and 13-month-old groups. Although not
statistically significant, lower overall spawning performance of the 13-month-old group
relative to the 10-month-old group suggests that spawning performance of these pond-
reared P. merguiensismight have peaked between 10 and 13 months. It was observed in the
maturation pond (where the prawns were sampled for these breeding trials) that full ovarian
maturation and spontaneous spawning occurred commonly when prawns were 11–12
months old. Multiple spawning was possible since some tagged prawns were caught at
stage IVof ovarian maturation (i.e. ready to spawn) a few times over these 2 months. This
observation may help explain the lower survival of prawns in the 13-month-old group. As
these prawns might have undergone several spawnings (in the pond), they may have been
reproductively exhausted and were, therefore, more susceptible to stress from eyestalk
ablation. Similar information has been reported for P. semisulcatus (Crocos and Coman,
1997), P. paulensis (Cavalli et al. 1997), and P. monodon (Niamado and Kane 1993). The
spawning performance of ablated wild P. semisulcatus collected from Albatross Bay, Gulf
of Carpentaria, Australia was found to increase from the time of first maturity (presumably
6 months old) to 12 months old and declined as the prawns reached 14 months old (Crocos
and Coman, 1997). Cavalli et al. (1997) reported a significant drop in spawning perform-
ance of 16.5-month-old P. paulensis broodstock. They also reported an increasing mortality
and negative growth in broodstock older than 15 months despite their larger sizes, which
agrees with our finding on pond-reared P. merguiensis in this study. Negative growth of
ablated P. merguiensis broodstock has been discussed in Hoang et al. (in press (b)). It is
likely due to excessive mobilization of maternal materials and energy into the formation of
eggs over a short time frame (3–4 days between two successive spawns within an
intermoult period). In wild P. monodon, age has been also suspected to influence spawning
performance (Hansford and Marsden, 1995). Pond-reared P. monodon older than 20 months
had higher mortality, lower fecundity, and lower fertility (Niamadio and Kane, 1993).
It has been generally accepted that larger prawns tend to produce a superior spawning
performance than do smaller ones. However, since the age of wild broodstock is often
unknown, it is difficult to separate the effect of prawn age from size. It is common but not
necessarily the case that larger prawns are older ones. If the age of the prawns used in this
study had been unknown, it would have been hard to explain why the overall spawning
performance of the 13-month-old groups was different from the 10-month-old group.
Therefore, the development of a convenient and accurate method to estimate prawn age is
needed to further investigate the effect of prawn age on spawning performance. This study
demonstrated that at the same age, larger P. merguiensis tended to spawn better than
smaller ones. Our results are in agreement with those of Cavalli et al. (1997).
The pattern of spawning performance over a range of ages and sizes observed in this
study implies some application in breeding pond-reared P. merguiensis. It appears that the
T. Hoang et al. / Aquaculture 212 (2002) 373–382380

spawning performance of the 10-month-old prawns (>26.8 g or 35.6 mm CL) is good for
hatchery production. Considering that full ovarian maturation and spontaneous spawning
were observed commonly when prawns reached 11–12 months old and the decline in
spawning performance was observed with the 13-month-old group (36.6 g or 40.4 mm
CL), it is suggested that pond-reared P. merguiensis broodstock should be induced to
spawn when they reach either 27 g or 10–12 months old. This study did not assess
spawning quality such as hatching rate, survival of larvae, and postlarvae. However, these
prawns started to mate at 6 months old and viable nauplii were produced throughout the
three breeding trials. Some of the nauplii (from the 10-month-old group) were nursed to
postlarvae and grown to broodstock size at BIARC. The spawning performance of the
larger prawns (7L) in the 7-month-old group (19.2 g or 31.4 mm CL) is, however,
acceptable and may indicate a potential for selection for early maturity in P. merguiensis.
Once the domestication or selective breeding of P. merguiensis is achieved, the selection of
pond-reared broodstock that mature at younger ages (and large enough sizes to ensure a
good growth of their offspring) will reduce generation time and allow the selected strains
to be commercially available in the shortest time frame. This study, although providing
preliminary information on the effect of age, size, and light intensity on pond-reared P.
merguiensis, did not cover a wide range of culture conditions, which intuitively may vary
the spawning pattern observed. Further research in this field is needed to confirm these
findings and draw more conclusive results.
Acknowledgements
Thanks are given to Mrs. Nguyen Thi Anh Tuyet for her assistance in the laboratory, Ms.
Jan Rose for providing the algae for larval rearing, Dr. David Mayer for his advice on the
statistical analysis, and Mr. Michael Cosgrove for proofreading. This study was supported
by Griffith University and Queensland Department of Primary Industries, Australia.
References
Bray, W.A., Lawrence, A.L., 1992. Reproduction of Penaeus species in captivity. In: Fast, A.W., Lester, L.J.
(Eds.), Marine Shrimp Culture: Principles and Practices. Elsevier, Amsterdam, pp. 93–170.
Browdy, C.L., 1992. A review of the reproductive biology of Penaeus species: perspectives on controlled shrimp
maturation systems for high quality nauplii production. In: Wyban, J. (Ed.), Proceedings of the Special
Session on Shrimp Farming, 22–25 May 1992, Orlando, Florida, USA. World Aquaculture Society, Baton
Rouge, LA, USA, pp. 22–51.
Cavalli, R.O., Scardua, M.P., Wasielesky, W.J., 1997. Reproductive performance of different sized wild and pond-
reared Penaeus paulensis females. J. World Aquacult. Soc. 28, 260–267.
Crocos, P.J., Coman, G.J., 1997. Seasonal and age variability in the reproductive performance of Penaeus semi-
sulcatus broodstock: optimising broodstock selection. Aquaculture 155, 55–67.
Guerao, G., Ribera, C., 1995. Growth and reproductive ecology of Palaemon adspersus (Decapoda, Palaemo-
nidae) in the western Mediterranean. Ophelia 43, 203–215.
Hansford, S.W., Marsden, G.E., 1995. Temporal variation in egg and larval productivity of eyestalk ablated
spawners of the prawn Penaeus monodon from Cook Bay, Australia. J. World Aquacult. Soc. 26, 396–405.
Hoang, T., Lee, S.Y., Keenan, C.P., Marsden, G.E., in press (a). Observations on growth, sexual maturity, and
spawning performance of pond-reared Penaeus merguiensis. Aquacult. Res.
T. Hoang et al. / Aquaculture 212 (2002) 373–382 381

Hoang, T., Lee, S.Y., Keenan, C.P., Marsden, G.E., in press (b). Effect of light intensity maturation and spawning
of ablated females Penaeus merguiensis. Aquaculture.
Hoang, T., Lee, S.Y., Keenan, C.P., Marsden, G.E., in press (c). Ovarian maturation of the banana prawn, Penaeus
merguiensis de Man under different light intensities. Aquaculture.
Lichatowich, T., Smalley, T., Mate, F.D., 1978. The natural reproduction of Penaeus merguiensis (de Man, 1888)
in an earthen pond in Fiji. Aquaculture 15, 377–378.
Marsden, G.E., McGuren, J.J., Hansford, S.W., Burke, M.J., 1997. A moist artificial diet for prawn broodstock: its
effect on the variable reproductive performance of wild caught Penaeus monodon. Aquaculture 149, 145–
156.
McCullagh, P., Nelder, J.A., 1989. Generalized Linear Models. Chapman & Hall, London, 511 pp.
Menasveta, P., Sangpradub, S., Piyatiratitivorakul, S., Fast, A.W., 1994. Effects of broodstock size and source on
ovarian maturation and spawning of Penaeus monodon Fabricius from the Gulf of Thailand. J. World
Aquacult. Soc. 25, 41–49.
Motoh, H., 1981. Studies on the fisheries biology of the giant tiger prawn, Penaeus monodon in the Philippines.
Technical Report no. 7. Aquaculture Department, Southeast Asian Fisheries Development Center, Tigbauan,
Philippines, 128 pp.
Niamadio, I., Kane, A., 1993. The maturing of giant tiger shrimps (Penaeus monodon Fabricius) in Senegal (West
Africa): assessment of optimum reproduction age in hatchery. In: Carrillo, M., Dahle, L., Morales, J.,
Sorgeloos, P., Svennevig, N., Wyban, J. (Eds.), From Discovery to Commercialization. Special Publication
of European Aquaculture Society, European Aquaculture Society, Oostende, Belgium, p. 154.
Payne, R.W., Lane, P.W., Digby, P.G.N., Harding, S.A., Leech, P.K., Morgan, G.W., Todd, A.D., Thompson, R.,
Tunnicliffe Wilson, G., Welham, S.J., White, R.P., 1993. Genstat 5 Release 3 Reference Manual Clarendon
Press, Oxford, 796 pp.
Primavera, J.H., 1985. A review of maturation and reproduction in closed thelycum penaeids. In: Taki, Y.P.,
Primavera, J.H., Llobrera, J.A. (Eds.), Proceedings of the First International Conference on the Culture of
Penaeid Prawns/Shrimps. SEAFDEC Aquaculture Department, Iloilo City, Philippines, pp. 47–64.
Roa, R., Ernst, B., 1996. Age structure, annual growth, and variance of size-at-age of the shrimp Heterocarpus
reedi. Mar. Ecol.: Prog. Ser. 137, 59–70.
Sheehy, M.R.J., 1990. The widespread occurrence of fluorescent morphological lipofuscin in the crustacean brain.
J. Crustacean Biol. 10, 613–622.
Sheehy, M., Cameron, M., Marsden, G., McGrath, J., 1995. Age structure of female giant tiger prawns Penaeus
monodon as indicated by neuronal lipofuscin concentration. Mar. Ecol.: Prog. Ser. 117, 59–63.
Somers, I.F., Kirkwood, G.P., 1991. Population ecology of the grooved tiger prawn, Penaeus semisulcatus, in the
northwestern Gulf of Carpentaria, Australia: growth, movement, age structure, and infestation by the bopyrid
parasite Epipenaeon ingens. Aust. J. Mar. Freshwater Res. 42, 349–367.
Thomas, M.M., 1975. Age and growth, length–weight relationship, and relative condition factor of Penaeus
semisulcatus De Haan. Indian J. Fish. 22, 133–142.
Tuma, D.J., 1967. A description of the development of primary and secondary sexual characters in the banana
prawn, Penaeus merguiensis de Man (Crustacea: Decapoda: Penaeinae). Aust. J. Mar. Freshwater Res. 18,
73–88.
T. Hoang et al. / Aquaculture 212 (2002) 373–382382
Copyright © 2022 FDOKUMEN




















