
Quasi-Optimized Overlapping Schwarz Waveform Relaxation Algorithm for PDEs with Time-Delay

Journal of Physiology - Paris xxx (2014) xxx–xxx
Contents lists available at ScienceDirect
Journal of Physiology - Paris
journal homepage: www.elsevier .com/locate / jphyspar is
Maternal and affective behaviors of lactating rats reared in overlappinglitters
http://dx.doi.org/10.1016/j.jphysparis.2014.04.0010928-4257/� 2014 Elsevier Ltd. All rights reserved.
⇑ Corresponding author at: Laboratorio de Neurociencias, Sección Biomatemática,Facultad de Ciencias, Universidad de la República, Iguá 4225, CP 11400 Montevideo,Uruguay. Tel.: +598 25258618x7151; fax: +598 25258617.
E-mail address: [email protected] (N. Uriarte).
Please cite this article in press as: Uriarte, N., et al. Maternal and affective behaviors of lactating rats reared in overlapping litters. J. Physiol. (2014)dx.doi.org/10.1016/j.jphysparis.2014.04.001
Natalia Uriarte a,c,⇑, María Victoria Fernández a,c, Daniella Agrati b,c, María José Zuluaga b,c,Marcela Ferreño b,c, Annabel Ferreira b,c
a Laboratorio de Neurociencias, Sección Biomatemática, Facultad de Ciencias, Universidad de la República, Uruguayb Sección Fisiología y Nutrición, Facultad de Ciencias, Universidad de la República, Uruguayc Laboratorio de Experimentación Animal, Facultad de Ciencias, Universidad de la República, Uruguay
a r t i c l e i n f o
Article history:Received 30 January 2014Received in revised form 26 March 2014Accepted 2 April 2014Available online xxxx
Keywords:Maternal behaviorMaternal aggressionExperimental anxietyEarly experienceLactationOverlapping littersCommunal nestingRat
a b s t r a c t
Postpartum mating in rats gives rise to complex family units consisting of the mother and two overlappinglitters. As a consequence, newborn pups of the second litter, since the moment they are born, acquireexperience not only from interaction with the mother and age-matched littermates but also frominteraction with older siblings. Newborn pups reared in overlapping litters (OLs) receive a differentpattern of maternal stimulation compared to those reared in single litters (SL: one litter of same agedpups), as the mothers reduce some maternal behavior components and juvenile pups from the first litterdevelop maternal behavior. Since there is strong evidence showing that variations in maternal behaviorare transmitted throughout generations, we hypothesized that the altered pattern of maternal stimulationreceived by OL reared females would modify their behavior during motherhood. To test this hypothesismaternal behavior, maternal aggression and experimental anxiety of dams reared under OL and SLconditions during the first postpartum week were compared. No differences were found between thegroups in their maternal behavior and aggression. This result may be explained by the maternal behaviorof the juveniles that could compensate for the deficits in the caregiving behaviors received by OL litters.However, a subtle temporal reorganization of the licking behavior was found in OL reared mothers,together with an increased anxiety-related behavior in the plus maze test. These results suggestdissociation in the effects provoked by early environmental alterations on different behavioral systems,and more importantly, that independently of their early family composition, both groups can copeeffectively with the changing demands of the pups.
� 2014 Elsevier Ltd. All rights reserved.
1. Introduction questioned based on their artificial nature (Wurbel, 2001; Macri
In mammals maternal behavior is essential for the survival ofthe offspring and represents the most relevant source of sensorystimulation for altricial pups, sculpting the development of theirnervous system (Suchecki et al., 1993; Anisman et al., 1998;Caldji et al., 1998). The long term physiological and behavioralconsequences that can arise from disruptions of this early mater-nal environment have been studied extensively employing experi-mental interventions like the neonatal handling and maternalseparation procedures (Weinberg et al., 1978; Pryce et al., 2001;Cirulli et al., 2003; Benetti et al., 2007). However, the ethologicalrelevance of the information provided by these models has been
et al., 2004; McLeod et al., 2007; Zuluaga et al., 2014).On the other hand, several studies have shown that natural
variations in maternal behavior provoke long-term changes in thebehavioral and neuroendocrine responses of the pups (Caldji et al.,1998; Champagne et al., 2003; Uriarte et al., 2007). In this line ofthought, we validated a new model to study the effects of variationsin early social environment on the individuals’ development underan ethologically relevant context: the overlapping of litters (OLs)(Uriarte et al., 2008, 2009). OL occurs in natural conditions when,following the successful mating during the postpartum estrus, twosuccessive litters coexist within the maternal burrow (Calhoun,1963; Gilbert et al., 1983). As a consequence, mother rats will raisepups from different ages with different sensory traits, physiologicaldemands, and behavioral capabilities at the same time.
Being reared in this complex early environment profoundlyaffects the neurobehavioral phenotype of adult rats. Thus, com-pared to animals reared in single litters (SLs), OL reared females
, http://

2 N. Uriarte et al. / Journal of Physiology - Paris xxx (2014) xxx–xxx
show a decrease in their behavioral inhibition assessed in the openfield test, a blunted corticosterone secretion in response torestraint stress and a reduction in their sexual receptiveness duringproestrus (Uriarte et al., 2009). These behavioral changes may bedue to the altered maternal care received, since during the overlap-ping of the two litters mothers modify their maternal behaviorwhen compared to dams taking care of a single litter. AlthoughOL mothers adapt their behavior according to the characteristicsand needs of the two different-aged litters and direct most of theirattention to neonates, pups from the second litter receive signifi-cantly less licking from the mother than pups raised in a single lit-ter (Uriarte et al., 2008).
Taking into account these results where mothers of OL altertheir maternal behavior, and previous studies demonstrating thatvariations in maternal behavior can be transmitted throughoutgenerations (Francis et al., 1999; Champagne and Meaney, 2001;Gonzalez et al., 2001; Fleming et al., 2002), we hypothesized thatfemales reared in OL would modify their own maternal behaviorwhen adults.
Maternal behavior consists of a complex collection of activities,which can dynamically change over time and also adapt to the con-text. Thus, pup care taking activities fluctuate across the light/darkcycle and through the course of the postpartum period (Smithet al., 1976; Leon et al., 1984; Pereira et al., 2008; Jensen Peñaand Champagne, 2013). Interestingly, it has been shown that inter-ference of early mother–pup interactions alter the temporal char-acteristics of maternal behavior rather than the overall frequencyof its components (Leon et al., 1984; Macri et al., 2004), highlight-ing the importance of analyzing the temporal dimension of thisbehavior.
Maternal behavior can be assessed by different methods thatprovide valuable and complementary information. While the con-tinuous observation of maternal behavior following a brief periodof separation from the litter is more suitable for analyzing activemotivational components in a short period of time (Pereira andFerreira, 2006), the periodic observation (spot-checks) of themother’s undisturbed behavior allows a deeper analysis of its tem-poral dynamic characteristics (Lonstein and Fleming, 2002;Champagne et al., 2007).
Motherhood does not only imply the development of care tak-ing activities directed to the pups, but also involves changes inthe affective responses to other environmental stimuli or situa-tions, such as the development of maternal aggression and mater-nal anxiolysis (Ferreira et al., 2002). Lactating females vigorouslyattack intruders in their home cages (Erskine et al., 1978;Ferreira and Hansen, 1986; Mayer and Rosenblatt, 1987) and exhi-bit a reduction of their anxiety-related behaviors in several exper-imental models (Fleming and Luebke, 1981; Hard and Hansen,1985; Ferreira et al., 1989, 2002; Bitran et al., 1991; Lonsteinet al., 19982). These behavioral adaptations complement directpup-caring behavior, as they provide additional protection to thealtricial pups from potential predators or dangerous situations(Pereira et al., 2005).
Based on the previous evidence exposed, this study aims to ana-lyze the maternal behavior, maternal aggression and anxiety-related behavior of lactating female rats reared in OL during thelight/dark periods of the photoperiodic cycle in the first weekpostpartum.
2. Materials and methods
2.1. Animals
Male and female rats (Rattus norvegicus, Wistar strain, from 0 to120 days old) were used for this study. The animals were housed
Please cite this article in press as: Uriarte, N., et al. Maternal and affective behavdx.doi.org/10.1016/j.jphysparis.2014.04.001
under controlled temperature (22 ± 1 �C) and humidity (65%) in a12 h light–dark cycle (lights on at 0500 h) with free access to foodand water. Animal care and experimental procedures were inaccordance with Uruguayan law (N�18 611) for the care and useof laboratory animals. Experimental protocol was approved bythe local Ethical Committee on Animal Care and Protocols.
2.2. Experimental groups and general procedure
To assess the changes in maternal and affective behaviors ofpostpartum females raised in OL, two groups were used:
Single litter (SL) reared dams, n = 9: lactating female rats rearedin single litters by multiparous mothers.Overlapping litter (OL) reared dams, n = 8: lactating female ratsreared in overlapping litters (a 10-day cohabitation period withtheir mothers and two juvenile siblings from the first litter).
To synchronize parturitions, adult females reared in SL or OLwere monitored for estrous cycle and mated overnight during theproestrous phase. One week prior to expected parturitions, femaleswere housed individually. On postpartum day (PPD) 0, litters werecounted, sexed, and culled to eight pups (with usually 4 or at least 3pups of each sex). Lactating females were left undisturbed and theirmaternal behavior was recorded by a spot-check observational pro-cedure in the breeding room from PPD1 to PPD7. During the lightphase of PPD8 the animals were transported to an adjacent testingroom with the same temperature and photoperiod conditions of thebreeding room and left to acclimatize at least one hour beforestarting the tests. Afterwards, a 30-min maternal behavior testwas performed followed by a 10-min aggression test in the females’nest-cage. Finally, one hour after the beginning of the dark phasethe dams were tested in the elevated plus maze for 5 min.
2.3. Breeding procedure to obtain experimental animals
To obtain adult females reared in single or overlapping littersthe following procedures were carried out:
Single littersFemales reared in single litters were obtained from multiparous
mothers in their second parturition, as the overlapping litterfemales were reared by mothers who were in their second lacta-tion. Approximately seven days before delivery (gestational day16, GD16), pregnant primiparous females were individually housedand, starting at GD20, the presence of pups was checked twice aday. On the day of birth litters were culled to eight individuals(four of each sex) (postpartum day (PPD) 0). Litters remained withtheir mothers until weaning on PPD21.
Overlapping littersOn GD16, pregnant nulliparous females were housed until par-
turition alongside the same males they had previously mated with,in order to copulate at postpartum estrus. After the postpartumestrus, vaginal smears were performed to check for the presenceof spermatozoids to confirm successful mating; and the maleswere removed. On GD21 of the second pregnancy the juvenilesfrom the first litter, except one male and one female, were weaned.These two juveniles remained in the maternal cage for 10 daysafter the birth of the second litter. Starting on GD22 of the secondpregnancy the presence of pups was checked twice a day.Newborns were weighed and culled in the same manner as singlelitter animals in both parturitions. Once the 10-day-period ofcohabitation with the juveniles ended, the mother and the secondlitter were left undisturbed until weaning on PPD21. The OL rearedfemales were obtained from these second litters.
iors of lactating rats reared in overlapping litters. J. Physiol. (2014), http://

N. Uriarte et al. / Journal of Physiology - Paris xxx (2014) xxx–xxx 3
Both groups of weaned female rats were housed in same-sex,same-litter groups of three to four animals per cage until matingand testing at adulthood (90 days old). To control for possible littereffects, individuals from different litters were used in all experi-mental groups.
2.4. Undisturbed maternal behavior observations
The behavior of lactating females was scored daily during four60-min observation sessions from PPD1 to PPD7. Observationswere performed at regular times, with three sessions during thelight phase (1000, 1300, 1600 h) and one during the dark phase(1830 h) of the light–dark cycle. Within each session, the behaviorof each subject was scored every 3 min (21 observations per peri-od � 4 periods per day = 84 observations/subject/day) (Champagneet al., 2003).
The following behaviors were recorded: Mother off pups (themother is away from the nest and without physical contact withthe pups); Anogenital licking (the mother licks the pups’ anogenitalregion), Body licking (the mother licks pups’ body surface); Nursing(the mother nurses the pups in either a high crouch posture, a lowcrouch posture in which the mother lays over the pups, or a supineposture in which the mother is lying either on her back or sidewhile the pups nurse); Retrieving and mouthing (retrieval of the dis-placed pups or oral repositioning of the pups into the nest); Nestbuilding (the female pushes or picks up nest material in hermouth).
2.5. Maternal behavior test
On PPD8 mothers were transported to the testing room and fol-lowing one hour of habituation the entire litter was removed. Tenminutes later five pups were scattered in the home cage oppositeto the female’s nest. The number of occurrences of the followingmaternal behavioral components was continuously recorded for30 min: retrieval of pups into the nest, mouthing, sniffing, bodyand anogenital licking, and nest building. In addition we alsorecorded the total duration of huddling activities, including lyingin contact with pups and hovering over the pups in the nest whileactively carrying out other behaviors (i.e. licking of pups or selfgrooming), and the high crouching, low crouching and supine nursingpostures. Total time (s) in contact with pups was the summed dura-tions of huddling plus nursing behaviors, the time that the motherspent off nest was calculated subtracting the total time in contactfrom the duration of the test (1800 s). Latency time was alsorecorded (i.e. time from the introduction of the pups to the firstbehavioral response from the female) to begin retrieving pups, toreunion of the entire litter into the nest, and to begin hovering overand nursing the litter. A latency of 1800 s was assigned to any cat-egory of behavior that was not initiated (or completed, i.e. reunionof the litter) within the 30 min observation period (Pereira andMorrell, 2009).
2.6. Maternal aggression test
The aggression test consisted in placing a male rat in the corneropposite to the nest in the female’s home cage, and in the presenceof all pups. The intruders were smaller sized young males than thefemales and were used only once to assure high levels of aggres-sion (Mos et al., 1987). Behavioral tests were recorded on a digitalvideo camera for ten minutes and analyzed later by a trainedobserver blind to experimental conditions, using JWatcher com-puter software.
The following behaviors were analyzed: sniffing the intruder(female investigates the intruder and sniffs its body and/or geni-tals), frontal attack (female launches towards the neck or head of
Please cite this article in press as: Uriarte, N., et al. Maternal and affective behavdx.doi.org/10.1016/j.jphysparis.2014.04.001
the intruder), lateral attack (female moves laterally towards theintruder, usually in association with piloerection and biting theback of the intruder), head bite (female bites the head of the intru-der), body bite (female bites any part of the intruder’s body exclud-ing the head), boxing (both rats facing one another while standingupright, typically accompanied by the female striking the intru-der’s face with its forepaws), lateral threat (female stands with backarched laterally oriented to the intruder, but with no attack againstthe intruder), intruder submission posture (sitting on haunches, withcurved back, forepaws raised off the ground; or lying on back withthe resident on top), and pup care (female interacts with her off-spring by crouching, retrieving, sniffing or licking them while theintruder is present) (Ferreira and Hansen, 1986).
2.7. Elevated plus maze test
The apparatus consists of an elevated (50 cm above the floor)plus-shaped maze with two opposite enclosed arms (50 cm-highwalls) and two open arms, each measuring 50 cm long and 10 cmwide. Each animal was placed in the center of the maze facing aclosed arm and allowed to freely explore the maze for 5 min. Thebehavioral parameters assessed were: frequency of total entries(arm entry, all four paws into an arm), frequency of open armsentries, time spent in the open arms and frequency of groomingand rearing (Severino et al., 2004).
2.8. Statistical analysis
Data are expressed as means ± standard error of the mean(SEM).
The litter size and sex ratio as well as behavioral data attainedin the maternal behavior, maternal aggression and plus maze testsof SL and OL reared dams were compared by the Unpaired Stu-dent’s t-tests.
The undisturbed maternal behavior of SL and OL mothers wasanalyzed within and across the seven postpartum days:
1. To evaluate fluctuations of maternal behavior across the firstpostpartum week, the sum of the frequencies of the four dailyobservation sessions along the first seven postpartum dayswas analyzed. A two-way ANOVA examining the effects ofgroup (SL and OL) and postpartum day (repeated measures:Day 1 to Day 7) was performed for each of the behavioralcomponents.
2. To assess variations in maternal behavior within the time ofday, the accumulated frequencies of the first seven postpartumdays on each of the four daily observation sessions were ana-lyzed. Therefore, a two-way ANOVA examining the effects ofgroup (SL and OL) and the time of day (repeated measures: at1000, 1300, 1500 and 1830 h) was performed for each of thebehavioral components.
When appropriate, post hoc comparisons were carried out usingNewman–Keuls test. In all cases the level of significance was set atp < 0.05.
3. Results
3.1. Pregnancy rate and litter size
All (nine of nine) dams reared in SL and eight out of ninefemales reared in OL became pregnant and successfully deliveredtheir litters. As shown in Table 1, the number of pups per litterand the pups’ sex ratio did not differ between SL and OL reareddams at birth.
iors of lactating rats reared in overlapping litters. J. Physiol. (2014), http://

Table 1Litter size and sex ratio of SL (n = 9) and OL (n = 8) reared dams’ litters (mean ± SEM).
Groups
SL OL p
Litter size 14.2 ± 0.7 12.9 ± 0.9 0.38Litter sex ratio 0.9 ± 0.1 1.0 ± 0.2 0.27
4 N. Uriarte et al. / Journal of Physiology - Paris xxx (2014) xxx–xxx
3.2. Maternal behavior during the first seven postpartum days
The total frequency of maternal behaviors between OL and SLreared dams during the first seven postpartum days are shown inFig. 1. The two-way ANOVA did not reveal a main effect of groupon: nursing (F[1,15] = 1.36, p = 0.26), time that mothers remainedoutside the nest (F[1,15] = 1.05, p = 0.32) and frequency of anogeni-tal (F[1,15] = 0.14, p = 0.72) or body licking (F[1,15] = 2.50, p = 0.14).
However, a main effect of days was detected on the frequenciesthat mothers spent outside the nest (F[6,90] = 10.65, p < 0.001) andengaged in anogenital (F[6,90] = 2.61, p = 0.022) and body licking(F[6,90] = 2.40, p = 0.033). By contrast, nursing behavior did not dif-fer between the groups across the first seven postpartum days(F[6,90] = 0.29, p = 0.94). No interaction was detected between thetwo factors.
3.3. Maternal behavior within daily observations
The two-way ANOVA did not show a significant effect of thegroup in the total time that mothers spent nursing (F[1,15] = 0.02,p = 0.89) or outside the nest (F[1,15] = 1.27, p = 0.27). However, aneffect of the time of day along the four observational periods wasdetected in nursing (F[3,45] = 25.33, p < 0.001), and mother off nest
Fig. 1. Mean ± SEM of the sum of the four daily frequencies for each of the first seven posand (D) body licking performed by SL (n = 9) and OL (n = 8) reared dams.
Please cite this article in press as: Uriarte, N., et al. Maternal and affective behavdx.doi.org/10.1016/j.jphysparis.2014.04.001
(F[3,45] = 58.92, p = 0 < 0.001). Thus, as shown in Fig. 2, both groupsof mothers spent more time outside the nest during the dark phase(18:30 h) with a concomitant decrease of their nursing behavior.No interaction between group and time of day was detected.
In addition, a main effect of time (F[3,45] = 4.89, p = 0.005) and asignificant interaction between time of observation and group(F[3,45] = 3.42, p = 0.025) was detected in the frequency of anogeni-tal licking. However, no effect of the group was found (F[1,15] = 0.10,p = 0.75) on this behavior. Post hoc analysis showed that for SLdams, the frequency of this behavior remained stable during theday and significantly decreased during the dark phase (the lastdaily observation session). On the other hand, the anogenital lick-ing behavior performed by OL reared dams exhibited an invertedU-shaped pattern with a significant increase in this behavior dur-ing the second and third observation sessions (see Fig. 2). By con-trast, the two-way ANOVA analysis did not detect any group(F[1,15) = 2.09, p = 0.67) or time of day (F[3,45] = 1.11, p = 0.354)effect in the total frequency of body licking.
3.4. Nest attendance
A more detailed analysis of the mother’s nest attendance, com-paring the number of bouts that dams spent outside the nest,revealed that independently of their duration, the number of off-nest bouts was statistically higher (t = 2.75, p = 0.015) in OL com-pared to SL reared dams during the dark period observation session(see Fig. 3A). However, no differences were detected during thelight phase (data not shown). Fig. 3B depicts the pattern of nestattendance during the night from one representative dam in eachgroup, illustrating that even when the total duration of nest atten-dance did not differ between these females, the frequency in whichthe OL reared dams left the nest was higher.
tpartum days of (A) total nursing, (B) mother outside the nest, (C) anogenital licking
iors of lactating rats reared in overlapping litters. J. Physiol. (2014), http://
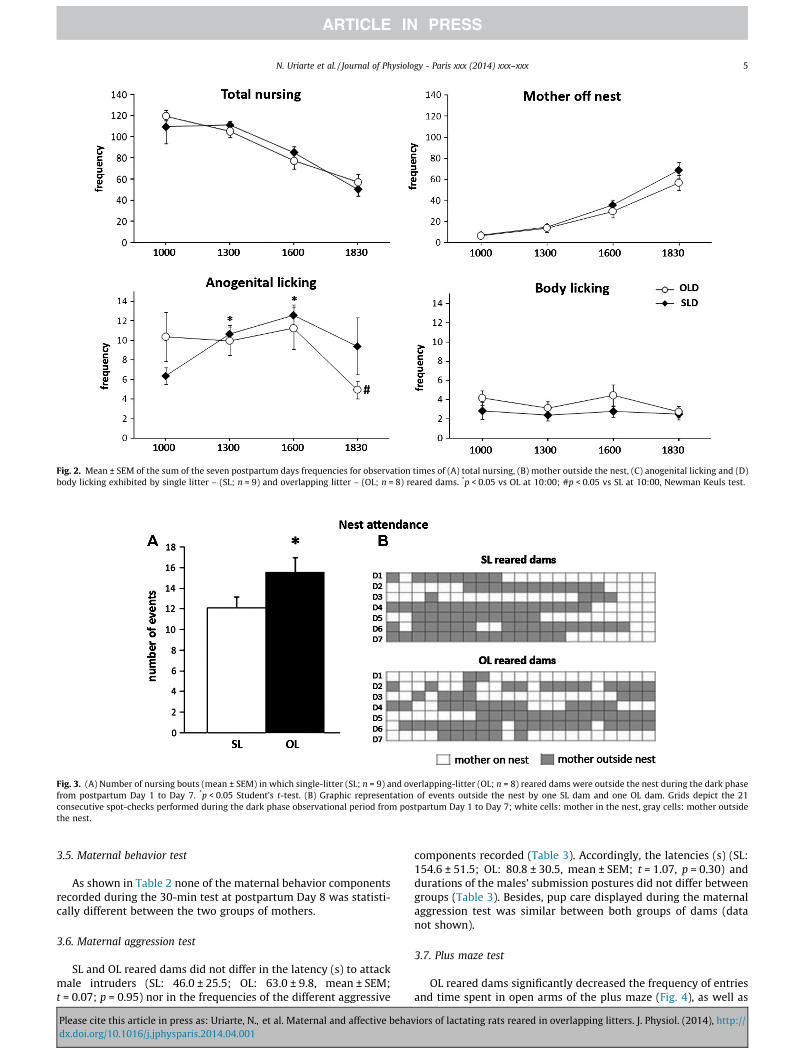
Fig. 2. Mean ± SEM of the sum of the seven postpartum days frequencies for observation times of (A) total nursing, (B) mother outside the nest, (C) anogenital licking and (D)body licking exhibited by single litter – (SL; n = 9) and overlapping litter – (OL; n = 8) reared dams. *p < 0.05 vs OL at 10:00; #p < 0.05 vs SL at 10:00, Newman Keuls test.
Fig. 3. (A) Number of nursing bouts (mean ± SEM) in which single-litter (SL; n = 9) and overlapping-litter (OL; n = 8) reared dams were outside the nest during the dark phasefrom postpartum Day 1 to Day 7. *p < 0.05 Student’s t-test. (B) Graphic representation of events outside the nest by one SL dam and one OL dam. Grids depict the 21consecutive spot-checks performed during the dark phase observational period from postpartum Day 1 to Day 7; white cells: mother in the nest, gray cells: mother outsidethe nest.
N. Uriarte et al. / Journal of Physiology - Paris xxx (2014) xxx–xxx 5
3.5. Maternal behavior test
As shown in Table 2 none of the maternal behavior componentsrecorded during the 30-min test at postpartum Day 8 was statisti-cally different between the two groups of mothers.
3.6. Maternal aggression test
SL and OL reared dams did not differ in the latency (s) to attackmale intruders (SL: 46.0 ± 25.5; OL: 63.0 ± 9.8, mean ± SEM;t = 0.07; p = 0.95) nor in the frequencies of the different aggressive
Please cite this article in press as: Uriarte, N., et al. Maternal and affective behavdx.doi.org/10.1016/j.jphysparis.2014.04.001
components recorded (Table 3). Accordingly, the latencies (s) (SL:154.6 ± 51.5; OL: 80.8 ± 30.5, mean ± SEM; t = 1.07, p = 0.30) anddurations of the males’ submission postures did not differ betweengroups (Table 3). Besides, pup care displayed during the maternalaggression test was similar between both groups of dams (datanot shown).
3.7. Plus maze test
OL reared dams significantly decreased the frequency of entriesand time spent in open arms of the plus maze (Fig. 4), as well as
iors of lactating rats reared in overlapping litters. J. Physiol. (2014), http://

Table 2Maternal behavior components displayed by SL (n = 9) and OL (n = 8) reared lactatingfemales (mean ± SEM) in a 30-min test.
Behaviors Groups
SL OL p
FrequenciesRetrieving 9.2 ± 2.0 8.4 ± 0.9 0.72Anogenital licking 17.6 ± 1.9 17.1 ± 3.2 0.90Body licking 20.1 ± 2.7 17.0 ± 2.9 0.45Nest building 10.0 ± 3.2 17.6 ± 5.9 0.26Mother off nest 10.3 ± 1.6 9.1 ± 1.6 0.61
Duration (s)Total nursing 266.6 ± 134.7 361.5 ± 158.1 0.66Mother off nest 1157.4 ± 189.5 105.1 ± 171.8 0.85
Table 3Maternal aggression components displayed by SL (n = 9) and OL (n = 7) rearedlactating females and males’ submission postures during the 10-min aggression test(mean ± SEM).
Behaviors Groups
SL OL p
FrequenciesAttack 4.9 ± 1.1 4.6 ± 0.9 0.84Lateral posture 1.3 ± 0.4 1.4 ± 0.9 0.92Boxing 1.3 ± 0.5 1.4 ± 0.7 0.91On top 2.2 ± 0.8 1.4 ± 0.6 0.45Upright posture 2.2 ± 0.7 2.3 ± 0.9 0.96Head bite 1.8 ± 0.6 1.6 ± 0.5 0.81Body bite 1.6 ± 1.0 1.9 ± 0.1 0.74Rear kick 4.8 ± 1.7 2.4 ± 1.0 0.28
Durations (s)Attack posture 125.8 ± 44.74 152.8 ± 47.4 0.49Male submission posture 295.7 ± 68.84 388.6 ± 73.1 0.73
6 N. Uriarte et al. / Journal of Physiology - Paris xxx (2014) xxx–xxx
their grooming (SL: 2.7 ± 0.6, OL: 0.5 ± 0.3, mean ± SEM; t = 3.05,p < 0.01), while they increased the frequency of entries in theclosed arms and rearing postures (SL: 13.8 ± 1.9, OL: 24.1 ± 2.6,mean ± SEM; t = 3.23, p < 0.01) compared to SL reared lactatingfemales. The total frequency of arm entries (closed + open) didnot differ between the groups (SL: 17.1 ± 0.8, OL: 17.0 ± 2.0,mean ± SEM; t = 0.28, p < 0.53).
Fig. 4. Behavior of single litter reared dams (SL; n = 9) and overlapping litter reared
Please cite this article in press as: Uriarte, N., et al. Maternal and affective behavdx.doi.org/10.1016/j.jphysparis.2014.04.001
4. Discussion
The main findings of the present study are:
1. Females reared in OL had similar pregnancy rates, litter sizesand composition of litters to those reared in standardconditions.
2. The maternal behavior of mothers reared in SL and OL did notdiffer; however some differences were found in the temporalorganization of this behavior within the day.
3. Dams reared in OL displayed similar levels of maternal aggres-sion as control mothers, but exhibited greater experimentalanxiety in the plus maze test.
4.1. Single- and overlapping-litters reared dams showed similarpregnancy rates and litter sizes
The pregnancy rates and litter sizes of mothers reared in OL andSL did not differ, suggesting that being raised in these differentenvironments does not affect the reproductive success of females.In accordance, we previously reported that despite the reduction intheir lordosis quotient, females reared in OL do not diminish theintensity of lordosis nor the number of oocytes released duringproestrus (Uriarte et al., 2009). Moreover, the high pregnancy ratesof OL reared rats could be also related to the mating systememployed (copula ad libitum with experienced males), that allowsmales to complete several ejaculatory series. In this sense, it hasbeen shown that under standard laboratory conditions only oneejaculatory series is sufficient to induce high rates of pregnancyin cycling females (Adler, 1969).
4.2. Maternal behavior of single- and overlapping-litter rearedmothers
Contrary to our prediction, we found no differences in the caretaking activities towards pups between SL and OL reared mothersduring the first postpartum week, indicating that, independently oftheir early family composition, both groups can cope effectivelywith the changing demands of the pups during lactation.
Reinforcing this concept, the results of the maternal behaviortest showed no differences between SL and OL dams. This type of
dams (OL; n = 8) in the plus maze test. Mean ± SEM. *p < 0.05 Student’s t-test.
iors of lactating rats reared in overlapping litters. J. Physiol. (2014), http://

N. Uriarte et al. / Journal of Physiology - Paris xxx (2014) xxx–xxx 7
test includes a brief mother-litter separation period which stimu-lates the active components of maternal behavior (Pereira andFerreira, 2006), and therefore the latencies to begin pups’ retrievaland to group the litter into the nest are indicative of the females’capacity to organize maternal behavior and of their motivationalstate (Ferreira et al., 2012). Thus, under this testing condition, bothgroups had a similar maternal motivation and capacity to organizetheir maternal behavior. It would be interesting to evaluate thematernal motivation of SL and OL reared mothers in more chal-lenging situations, such as the retrieval of pups in a T-maze(Bridges et al., 1972; Stern and Mackinnon, 1976), or in modelsdesigned to quantify the strength of the reinforcing value of pups,such as place preference associated with pup’s cues (Fleming et al.,1994; Pereira and Morrell, 2010, 2011) and bar-pressing to gainaccess to pups (Lee et al., 2000).
As pups reared in overlapping litters received less maternallicking stimulation than those reared in single litters (Uriarteet al., 2008), the absence of differences between both groupscontrasts with evidence showing that maternal behavior ismainly shaped by the maternal licking received during the firstdays of life, and that variations in the maternal behavior aretransmitted throughout generations (Francis and Meaney, 1999;Champagne and Meaney, 2001; Gonzalez et al., 2001; Fleminget al., 2002).
A possible explanation for the lack of differences between thegroups may rely on the fact that the maternal stimulation of thenewborns reared in OL comes both from the mother and their sib-lings. As aforementioned, maternal licking received by the pupsdecreased in the OL rearing condition; however, the juveniles fromthe first litter develop maternal-like behaviors, licking and groom-ing their newborn siblings and spending significant amounts oftime in the nest. We have previously shown that the licking behav-ior performed by juveniles is sufficient to compensate the quantityof maternal stimulation of OL pups, allowing them to receive sim-ilar levels of stimulation than those of control pups (Uriarte et al.,2008). Therefore, it is possible that this extra source of stimulationprovided by juveniles is effective in overriding the deficits in thematernal stimulation of OL female’s pups.
Another putative explanation for the lack of differences inmaternal behavior between groups, even when the OL pupsreceived less maternal stimulation, could rely on the social com-plexity of this environment, which may represent an enrichedexperience for the pups. For instance, the exposure to sensoryand social cues from littermates counteracts the effects of maternalisolation after birth on the maternal behavior of females duringadulthood (Gonzalez et al., 2001; Melo et al., 2006, 2009). Also,in the asynchronous communal nesting model in mice, peer stim-ulation, in addition to mother–infant interactions, contributes toshape developmental trajectories of individuals (Branchi et al.,2013). Therefore, the augmented sensory stimulation and interac-tions provided by the juvenile siblings in our model seem to coun-teract the effects of less maternal stimulation, leading to a normaldevelopment of the newborns.
In agreement with previous studies (Grota and Ader, 1969;Croskerry et al., 1976; Ivy et al., 2008; Pereira et al., 2008; JensenPeña and Champagne, 2013) both groups of mothers exhibited areduction in their nest attendance, while their nursing behaviorremained stable during the first postpartum week. However, con-versely to that found by Jensen Peña and Champagne (2013), thetotal amount of licking did not decrease during the progression ofthe first postpartum week (data not shown). Moreover, the patternof anogenital licking seemed to differ from that of body licking, aresult that may be related to their different characteristics (Moore,1984; Gubernick and Alberts, 1985; Moore and Chadwick-Dias,1986). For instance, anogenital licking is controlled by separatedsensory and motivational mechanisms than those controlling body
Please cite this article in press as: Uriarte, N., et al. Maternal and affective behavdx.doi.org/10.1016/j.jphysparis.2014.04.001
licking (Friedman et al., 1981; Gubernick and Alberts, 1985; Moore,1992).
4.3. Daily pattern of maternal behavior of single- and overlapping-litter reared mothers
In accordance with the work of Jensen Peña and Champagne(2013), SL and OL reared dams increased their time spent outsidethe nest during the dark phase of the photoperiodic cycle, whichwas also reflected in a reduction of their nursing activity. Despitethis similar circadian fluctuations in the maternal behavior of bothgroups, OL reared mothers left their pups and the nest area morefrequently than SL reared dams during the dark phase, a result thatmay be interpreted as a greater fragmentation of their maternalbehavior pattern (Plotsky et al., 2005; Ivy et al., 2008, 2010). Inagreement, some studies have shown that not only the totalamount of maternal behavior but also its temporal distribution ismodified as a consequence of alterations of the mother-litter bond(Macri et al., 2004; Ivy et al., 2008). However, in our study, the totalamount of time off the nest did not differ between both groups,since OL mothers left their pups unattended for shorter periodsof time, questioning the putative impact of this change in the careof the litter, and therefore in its development.
The daily pattern of anogenital licking, but not that of corporallicking, differed between SL and OL reared rats, stressing theimportance of analyzing both behaviors separately, and pointingtoward a selective alteration of anogenital licking by this earlyrearing experience. As dams lick the anogenital region of malesmore frequently than females, being this stimulation essential forthe normal sexual differentiation (Moore, 1984), this differentialpattern could affect the development of sexual behavior in males.However, this seems unlikely because no differences were found inour previous studies (Uriarte et al., 2009) in the sexual behavior ofmales reared in OL or SL.
The difference in the expression of some maternal componentsof SL and OL reared mothers according to the time of the day sug-gests an influence of being reared in overlapping litters in the dailyorganization of pups’ care taking activities. However, the lack ofquantitative differences in these and other maternal componentswithin and across the postpartum days minimizes the relevanceof this change for the organization of maternal behavior, as wellas for the development of the pups. In this sense, Ivy et al.(2008) proposed that temporal variations in maternal behaviorwould impact the normal development of pups if they wereaccompanied by quantitative alterations. Therefore, future studiesneed to be done to clarify the importance of these temporalchanges of maternal behavior according to the early familycomposition.
4.4. Maternal aggression and anxiety-related behavior of single- andoverlapping-litter reared dams
Similarly to what was shown for maternal behavior, SL and OLreared dams displayed high levels of aggression toward intruders,indicating that being reared in overlapping litters does not affectthe capacity of the females to protect the pups against potentiallyhostile conspecifics. This result could be expected considering thesimilarities in maternal behavior of SL and OL reared females.Although maternal behavior and aggression have differences intheir subjacent mechanisms, maternal aggression is strongly mod-ulated by the maternal state of the female and both behaviorsshare many sensory, hormonal and motivational properties(Mayer and Rosenblatt, 1993; Rosenblatt et al., 1994; Ferreiraet al., 2002; Lonstein and Gammie, 2002).
Few studies have analyzed the effects of early environment onthe affective behavior of females during motherhood, obtaining
iors of lactating rats reared in overlapping litters. J. Physiol. (2014), http://

8 N. Uriarte et al. / Journal of Physiology - Paris xxx (2014) xxx–xxx
different results. While some found that daily events of maternalseparation increase maternal aggression in mice (Veenema et al.,2007) and rats (Padoin et al., 2001; Giovenardi et al., 2005), othersreported a reduction (Boccia and Pedersen, 2001) or even no effect(Boccia and Pedersen, 2001) of these interventions on dams’aggressiveness. In the same line, mothers artificially reared insocial isolation show augmented aggression toward intruders(Melo et al., 2009). However, unlike these models of early life expe-riences, in overlapping litters neonates do not experience a reduc-tion of the sensory stimulation received, but rather a modification.In this sense, Melo et al. (2009) reported that the effects of socialisolation during artificial rearing on maternal aggression are pre-vented by permitting the contact with two peers.
In contrast to maternal aggression, OL reared dams exhibited agreater anxiety-related behaviors in the plus maze test comparedto control reared dams, which suggests that variations in earlyrearing conditions could be predictors of mothers’ responses innovel and aversive environments. Interestingly, OL reared cyclingfemales display reduced levels of anxiety-related behaviors(Uriarte et al., 2009) in contrast to the high levels shown by OLreared dams, indicating that the long lasting effects of early lifeexperience specifically depend on the behavioral system and onthe reproductive stage analyzed. Accordingly, Giovenardi et al.(2005) showed that adult virgin females that had experienced neo-natal handling diminish their fear-like behavior compared to con-trol rats, while lactating handled females exhibit an increase in thisresponse during the postpartum period. Similarly, communal nest-ing experience modifies emotional behavior in a context-depen-dent manner. While anxiety-related behavior is reduced in socialinteraction tests (Branchi et al., 2006a), it increases in plus-mazeand open field tests (Branchi et al., 2006b).
These high levels of anxiety-related behavior of OL dams did notinterfere with the expression of their maternal and aggressivebehaviors. In this sense, although maternal anxiolysis has beenproposed as a key component of the maternal repertoire (Ferreiraet al., 1989; Pereira et al., 2005), it has differential control mecha-nisms and regulatory factors (Ferreira et al., 2002; Lonstein, 2005a,2007). Besides, it has been shown that lactating rats (Bosch et al.,2006; Neumann and Bosch, 2008) and mice (Kessler et al., 2011)selectively bred for high anxiety-like behavior, nurse pups morefrequently, leave the nest less often, and are more motivated toretrieve pups than rats selected for low anxiety-like behavior.
The fact that being reared in overlapped litters modified post-partum females’ anxiety but not their maternal aggression, stressesthe independence in the regulation of both affective behaviors. Inthis line of thought, although a reduction of fear- and anxiety-related behaviors has been strongly related to the display of mater-nal aggression (Lonstein et al., 1998; Lonstein and Gammie, 2002),interventions that affect one of these affective behaviors do notnecessarily modify the other (Hansen and Ferreira, 1986b,a;Ferreira et al., 1987). Bosch and co-workers showed that lactatingfemales bred for high levels of anxiety-like behavior also presentaugmented levels of maternal aggression (Bosch et al., 2005). Thus,present results confirm the complexity of the relationship betweenmaternal care, maternal aggression and anxiolysis during the post-partum period (Lonstein, 2005b).
Previous results show that overlapping of litters, resulting fromthe mothers’ mating at postpartum estrus, gives rise to a morecomplex family unit which enriches early environment in such away that it impacts pup development. Our present study demon-strates that being reared in this early social environment doesnot significantly affect the reproductive success, or the maternaland aggressive behaviors of the females, but changes their emo-tional response in a novel situation. Together, these results showthat although being reared in overlapping litters modifies someaffective responses of the females, it does not interfere with their
Please cite this article in press as: Uriarte, N., et al. Maternal and affective behavdx.doi.org/10.1016/j.jphysparis.2014.04.001
capability to effectively cope with the pups’ imminent demands,ensuring their survival.
The overlapping-litters model in the rat, as the communal nest-ing model in mice (Branchi et al., 2011) or, more recently, the asyn-chronous communal nesting model (Branchi et al., 2013), representnaturally occurring rearing conditions, which are part of a new andmore ethological approach to study the influence of the early socialenvironments on individual’s development. Together these modelsreveal the natural plasticity of the developmental processes andemphasize the idea that multiple developmental trajectories arisefrom the diversity of early social environments. This phenomenon,that seems to be the rule rather than the expectation, may underliethe behavioral variability observed in natural populations.
Acknowledgements
The authors wish to thank Mr. Héctor Rodríguez and Mr. JoelGonzález for their excellent technical assistance. This researchwas supported by CSIC-DT, PEDECIBA, and ANII (SNI, SNB).
References
Adler, N.T., 1969. Effects of the male’s copulatory behavior on successful pregnancyof the female rat. J. Comp. Physiol. Psychol. 69, 613–622.
Anisman, H., Zaharia, M.D., Meaney, M.J., Merali, Z., 1998. Do early-life eventspermanently alter behavioral and hormonal responses to stressors? Int. J. Dev.Neurosci. 16, 149–164.
Benetti, F., Andrade de Araujo, P., Sanvitto, G.L., Lucion, A.B., 2007. Effects ofneonatal novelty exposure on sexual behavior, fear, and stress-response in adultrats. Dev. Psychobiol. 49, 258–264.
Bitran, D., Hilvers, R.J., Kellogg, C.K., 1991. Ovarian endocrine status modulates theanxiolytic potency of diazepam and the efficacy of gamma-aminobutyric acid-benzodiazepine receptor-mediated chloride ion transport. Behav. Neurosci. 105,653–662.
Boccia, M.L., Pedersen, C.A., 2001. Brief vs. long maternal separations in infancy:contrasting relationships with adult maternal behavior and lactation levels ofaggression and anxiety. Psychoneuroendocrinology 26, 657–672.
Bosch, O.J., Meddle, S.L., Beiderbeck, D.I., Douglas, A.J., Neumann, I.D., 2005. Brainoxytocin correlates with maternal aggression: link to anxiety. J. Neurosci. 25,6807–6815.
Bosch, O.J., Kromer, S.A., Neumann, I.D., 2006. Prenatal stress: opposite effects onanxiety and hypothalamic expression of vasopressin and corticotropin-releasing hormone in rats selectively bred for high and low anxiety. Eur. J.Neurosci. 23, 541–551.
Branchi, I., D’Andrea, I., Sietzema, J., Fiore, M., Di Fausto, V., Aloe, L., Alleva, E., 2006a.Early social enrichment augments adult hippocampal BDNF levels and survivalof BrdU-positive cells while increasing anxiety- and ‘‘depression’’-like behavior.J. Neurosci. Res. 83, 965–973.
Branchi, I., D’Andrea, I., Fiore, M., Di Fausto, V., Aloe, L., Alleva, E., 2006b. Early socialenrichment shapes social behavior and nerve growth factor and brain-derivedneurotrophic factor levels in the adult mouse brain. Biol. Psychiatry 60, 690–696.
Branchi, I., D’Andrea, I., Santarelli, S., Bonsignore, L.T., Alleva, E., 2011. The richnessof social stimuli shapes developmental trajectories: are laboratory mouse pupsimpoverished? Prog. Neuropsychopharmacol. Biol. Psychiatry 35, 1452–1460.
Branchi, I., Curley, J.P., D’Andrea, I., Cirulli, F., Champagne, F.A., Alleva, E., 2013. Earlyinteractions with mother and peers independently build adult social skills andshape BDNF and oxytocin receptor brain levels. Psychoneuroendocrinology 38,522–532.
Bridges, R., Zarrow, M.X., Gandelman, R., Denenberg, V.H., 1972. Differences inmaternal responsiveness between lactating and sensitized rats. Dev. Psychobiol.5, 123–127.
Caldji, C., Tannenbaum, B., Sharma, S., Francis, D., Plotsky, P.M., Meaney, M.J., 1998.Maternal care during infancy regulates the development of neural systemsmediating the expression of fearfulness in the rat. Proc. Natl. Acad. Sci. USA 95,5335–5340.
Calhoun, J.B., 1963. The Ecology and Sociology of the Norway Rat. Washington: U.S.Dept. of Health, Education, and Welfare, Public Health Service.
Champagne, F., Meaney, M.J., 2001. Like mother, like daughter: evidence for non-genomic transmission of parental behavior and stress responsivity. Prog. BrainRes. 133, 287–302.
Champagne, F.A., Francis, D., Mar, A., Meaney, M.J., 2003. Variations in maternal carein the rat as a mediating influence for the effects of environment ondevelopment. Physiol. Behav. 79, 359–371.
Champagne, F.A., Curley, J.P., Keverne, E.B., Bateson, P.P., 2007. Natural variations inpostpartum maternal care in inbred and outbred mice. Physiol. Behav. 91, 325–334.
iors of lactating rats reared in overlapping litters. J. Physiol. (2014), http://

N. Uriarte et al. / Journal of Physiology - Paris xxx (2014) xxx–xxx 9
Cirulli, F., Berry, A., Alleva, E., 2003. Early disruption of the mother–infantrelationship: effects on brain plasticity and implications for psychopathology.Neurosci. Biobehav. Rev. 27, 73–82.
Croskerry, P.G., Smith, G.K., Leon, L.N., Mitchell, E.A., 1976. An inexpensive systemfor continuously recording maternal behavior in the laboratory rat. Physiol.Behav. 16, 223–225.
Erskine, M.S., Barfield, R.J., Goldman, B.D., 1978. Intraspecific fighting during latepregnancy and lactation in rats and effects of litter removal. Behav. Biol. 23,206–218.
Ferreira, A., Hansen, S., 1986. Sensory control of maternal aggression in Rattusnorvegicus. J. Comp. Psychol. 100, 173–177.
Ferreira, A., Dahlof, L.G., Hansen, S., 1987. Olfactory mechanisms in the control ofmaternal aggression, appetite, and fearfulness: effects of lesions to olfactoryreceptors, mediodorsal thalamic nucleus, and insular prefrontal cortex. Behav.Neurosci. 101 (709–717), 746.
Ferreira, A., Hansen, S., Nielsen, M., Archer, T., Minor, B.G., 1989. Behavior of motherrats in conflict tests sensitive to antianxiety agents. Behav. Neurosci. 103, 193–201.
Ferreira, A., Pereira, M., Agrati, D., Uriarte, N., Fernandez-Guasti, A., 2002. Role ofmaternal behavior on aggression, fear and anxiety. Physiol. Behav. 77, 197–204.
Ferreira, A., Agrati, D., Uriarte, N., Pereira, M., Zuluaga, M.J., 2012. The rat as a modelfor studying maternal behavior. In: Cruz- Morales, S.E., Arriaga-Ramírez, J.C.P.(Eds.), Behavioral Animal Models. Research Signpost, Kerala, pp. 73–88.
Fleming, A.S., Luebke, C., 1981. Timidity prevents the virgin female rat from being agood mother: emotionality differences between nulliparous and parturientfemales. Physiol. Behav. 27, 863–868.
Fleming, A.S., Korsmit, M., Deller, M., 1994. Rat pups are potent reinforcers to thematernal animal: effects of experience, parity, hormones, and dopaminefunction. Psychobiology 22, 44–53.
Fleming, A.S., Kraemer, G.W., Gonzalez, A., Lovic, V., Rees, S., Melo, A., 2002.Mothering begets mothering: the transmission of behavior and its neurobiologyacross generations. Pharmacol. Biochem. Behav. 73, 61–75.
Francis, D., Meaney, M.J., 1999. Maternal care and the development of stressresponses. Curr. Opin. Neurobiol. 9, 128–134.
Francis, D., Diorio, J., Liu, D., Meaney, M.J., 1999. Nongenomic transmission acrossgenerations of maternal behavior and stress responses in the rat. Science 286,1155–1158.
Friedman, M.I., Bruno, J.P., Alberts, J.R., 1981. Physiological and behavioralconsequences in rats of water recycling during lactation. J. Comp. Physiol.Psychol. 95, 26–35.
Gilbert, A.N., Burgoon, D.A., Sullivan, K.A., Adler, N.T., 1983. Mother-weanlinginteractions in Norway rats in the presence of a successive litter produced bypostpartum mating. Physiol. Behav. 30, 267–271.
Giovenardi, M., de Azevedo, M.S., da Silva, S.P., Hermel Edo, E., Gomes, C.M., Lucion,A.B., 2005. Neonatal handling increases fear and aggression in lactating rats.Physiol. Behav. 86, 209–217.
Gonzalez, A., Lovic, V., Ward, G.R., Wainwright, P.E., Fleming, A.S., 2001.Intergenerational effects of complete maternal deprivation and replacementstimulation on maternal behavior and emotionality in female rats. Dev.Psychobiol. 38, 11–32.
Grota, L.J., Ader, R., 1969. Continuous recording of maternal behaviour in Rattusnorvegicus. Anim. Behav. 17, 722–729.
Gubernick, D.J., Alberts, J.R., 1985. Maternal licking by virgin and lactating rats:water transfer from pups. Physiol. Behav. 34, 501–506.
Hansen, S., Ferreira, A., 1986a. Effects of bicuculline infusions in the ventromedialhypothalamus and amygdaloid complex on food intake and affective behaviorin mother rats. Behav. Neurosci. 100, 410–415.
Hansen, S., Ferreira, A., 1986b. Food intake, aggression, and fear behavior in themother rat: control by neural systems concerned with milk ejection andmaternal behavior. Behav. Neurosci. 100, 64–70.
Hard, E., Hansen, S., 1985. Reduced fearfulness in the lactating rat. Physiol. Behav.35, 641–643.
Ivy, A.S., Brunson, K.L., Sandman, C., Baram, T.Z., 2008. Dysfunctional nurturingbehavior in rat dams with limited access to nesting material: a clinicallyrelevant model for early-life stress. Neuroscience 154, 1132–1142.
Ivy, A.S., Rex, C.S., Chen, Y., Dube, C., Maras, P.M., Grigoriadis, D.E., Gall, C.M., Lynch,G., Baram, T.Z., 2010. Hippocampal dysfunction and cognitive impairmentsprovoked by chronic early-life stress involve excessive activation of CRHreceptors. J. Neurosci. 30, 13005–13015.
Jensen Peña, C., Champagne, F.A., 2013. Implications of temporal variation inmaternal care for the prediction of neurobiological and behavioral outcomes inoffspring. Behav. Neurosci. 127, 33–46.
Kessler, M.S., Bosch, O.J., Bunck, M., Landgraf, R., Neumann, I.D., 2011. Maternal carediffers in mice bred for high vs. low trait anxiety: impact of brain vasopressinand cross-fostering. Soc. Neurosci. 6, 156–168.
Lee, A., Clancy, S., Fleming, A.S., 2000. Mother rats bar-press for pups: effects oflesions of the mpoa and limbic sites on maternal behavior and operantresponding for pup-reinforcement. Behav. Brain Res. 108, 215–231.
Leon, M., Adels, L., Coopersmith, R., Woodside, B., 1984. Diurnal cycle of mother-young contact in Norway rats. Physiol. Behav. 32, 999–1003.
Lonstein, J.S., 2005a. Reduced anxiety in postpartum rats requires recent physicalinteractions with pups, but is independent of suckling and peripheral sources ofhormones. Horm. Behav. 47, 241–255.
Lonstein, J.S., 2005b. Resolving apparent contradictions concerning therelationships among fear or anxiety and aggression during lactation:
Please cite this article in press as: Uriarte, N., et al. Maternal and affective behavdx.doi.org/10.1016/j.jphysparis.2014.04.001
theoretical comment on D’Anna, Stevenson, and Gammie (2005). Behav.Neurosci. 119, 1165–1168.
Lonstein, J.S., 2007. Regulation of anxiety during the postpartum period. Front.Neuroendocrinol. 28, 115–141.
Lonstein, J.S., Fleming, A.S., 2002. Parental behaviors in rats and mice. Curr. Protoc.Neurosci. 1, 1, Chapter 8:Unit 8 15.
Lonstein, J.S., Gammie, S.C., 2002. Sensory, hormonal, and neural control of maternalaggression in laboratory rodents. Neurosci. Biobehav. Rev. 26, 869–888.
Lonstein, J.S., Simmons, D.A., Stern, J.M., 1998. Functions of the caudalperiaqueductal gray in lactating rats: kyphosis, lordosis, maternal aggression,and fearfulness. Behav. Neurosci. 112, 1502–1518.
Macri, S., Mason, G.J., Wurbel, H., 2004. Dissociation in the effects of neonatalmaternal separations on maternal care and the offspring’s HPA and fearresponses in rats. Eur. J. Neurosci. 20, 1017–1024.
Mayer, A.D., Rosenblatt, J.S., 1987. Hormonal factors influence the onset of maternalaggression in laboratory rats. Horm. Behav. 21, 253–267.
Mayer, A.D., Rosenblatt, J.S., 1993. Persistent effects on maternal aggression ofpregnancy but not of estrogen/progesterone treatment of nonpregnantovariectomized rats revealed when initiation of maternal behavior is delayed.Horm. Behav. 27, 132–155.
McLeod, J., Sinal, C.J., Perrot-Sinal, T.S., 2007. Evidence for non-genomictransmission of ecological information via maternal behavior in female rats.Genes, Brain Behav. 6, 19–29.
Melo, A.I., Lovic, V., Gonzalez, A., Madden, M., Sinopoli, K., Fleming, A.S., 2006.Maternal and littermate deprivation disrupts maternal behavior and social-learning of food preference in adulthood: tactile stimulation, nest odor, andsocial rearing prevent these effects. Dev. Psychobiol. 48, 209–219.
Melo, A.I., Hernández-Curiel, M., Hoffman, K.L., 2009. Maternal and peer contactduring the postnatal period participate in the normal development of maternalaggression, maternal behavior, and the behavioral response to novelty. Behav.Brain Res. 201, 14–21.
Moore, C.L., 1984. Maternal contributions to the development of masculine sexualbehavior in laboratory rats. Dev. Psychobiol. 17, 347–356.
Moore, C.L., 1992. The role of maternal stimulation in the development of sexualbehavior and its neural basis. Ann. N. Y. Acad. Sci. 662, 160–177.
Moore, C.L., Chadwick-Dias, A.M., 1986. Behavioral responses of infant rats tomaternal licking: variations with age and sex. Dev. Psychobiol. 19, 427–438.
Mos, J., Olivier, B., van Oorschot, R., 1987. Maternal aggression towards differentsized male opponents: effect of chlordiazepoxide treatment of the mothers andd-amphetamine treatment of the intruders. Pharmacol. Biochem. Behav. 26,577–584.
Neumann, I.D., Bosch, O.J., 2008. Maternal stress adaptations peripartum: Mom’sinnate anxiety determines maternal care and aggression. In: Bridges, R.S. (Ed.),Neurobiology of the Parental Brain. Academic Press, San Diego, pp. 115–130(Chapter 8).
Padoin, M.J., Cadore, L.P., Gomes, C.M., Barros, H.M., Lucion, A.B., 2001. Long-lastingeffects of neonatal stimulation on the behavior of rats. Behav. Neurosci. 115,1332–1340.
Pereira, M., Ferreira, A., 2006. Demanding pups improve maternal behavioralimpairments in sensitized and haloperidol-treated lactating female rats. Behav.Brain Res. 175, 139–148.
Pereira, M., Morrell, J.I., 2009. The changing role of the medial preoptic area in theregulation of maternal behavior across the postpartum period: facilitationfollowed by inhibition. Behav. Brain Res. 205, 238–248.
Pereira, M., Morrell, J.I., 2010. The medial preoptic area is necessary for motivatedchoice of pup- over cocaine-associated environments by early postpartum rats.Neuroscience 167, 216–231.
Pereira, M., Morrell, J.I., 2011. Functional mapping of the neural circuitry of ratmaternal motivation: effects of site-specific transient neural inactivation. J.Neuroendocrinol. 23, 1020–1035.
Pereira, M., Uriarte, N., Agrati, D., Zuluaga, M.J., Ferreira, A., 2005. Motivationalaspects of maternal anxiolysis in lactating rats. Psychopharmacology (Berl.)180, 241–248.
Pereira, M., Seip, K.M., Morrell, J.I., 2008. Maternal motivation and its neuralsubstrate across the postpartum period. In: Bridges, R.S. (Ed.), Neurobiology ofthe Parental Brain. Academic Press, San Diego, pp. 39–59 (Chapter 3).
Plotsky, P.M., Thrivikraman, K.V., Nemeroff, C.B., Caldji, C., Sharma, S., Meaney, M.J.,2005. Long-term consequences of neonatal rearing on central corticotropin-releasing factor systems in adult male rat offspring. Neuropsychopharmacology30, 2192–2204.
Pryce, C.R., Bettschen, D., Feldon, J., 2001. Comparison of the effects of earlyhandling and early deprivation on maternal care in the rat. Dev. Psychobiol. 38,239–251.
Rosenblatt, J.S., Factor, E.M., Mayer, A.D., 1994. Relationship between maternalaggression and maternal care in the rat. Aggressive Behav. 20, 243–255.
Severino, G.S., Fossati, I.A., Padoin, M.J., Gomes, C.M., Trevizan, L., Sanvitto, G.L.,Franci, C.R., Anselmo-Franci, J.A., Lucion, A.B., 2004. Effects of neonatal handlingon the behavior and prolactin stress response in male and female rats at variousages and estrous cycle phases of females. Physiol. Behav. 81, 489–498.
Smith, G.K., Leon, L.N., Mitchell, E.A., 1976. An inexpensive system forcontinuously recording maternal behavior in the laboratory rat. Physiol.Behav. 16, 223–225.
Stern, J.M., Mackinnon, D.A., 1976. Postpartum, hormonal, and nonhormonalinduction of maternal behavior in rats: effects on T-maze retrieval of pups.Horm. Behav. 7, 305–316.
iors of lactating rats reared in overlapping litters. J. Physiol. (2014), http://

10 N. Uriarte et al. / Journal of Physiology - Paris xxx (2014) xxx–xxx
Suchecki, D., Rosenfeld, P., Levine, S., 1993. Maternal regulation of thehypothalamic-pituitary-adrenal axis in the infant rat: the role of feeding andstroking. Dev. Brain Res. 75, 192–195.
Uriarte, N., Breigeiron, M.K., Benetti, F., Rosa, X.F., Lucion, A.B., 2007. Effects ofmaternal care on the development, emotionality, and reproductive functions inmale and female rats. Dev. Psychobiol. 49, 451–462.
Uriarte, N., Ferreira, A., Rosa, X.F., Sebben, V., Lucion, A.B., 2008. Overlapping littersin rats: effects on maternal behavior and offspring emotionality. Physiol. Behav.93, 1061–1070.
Uriarte, N., Ferreira, A., Rosa, X.F., Lucion, A.B., 2009. Effects of litter-overlapping onemotionality, stress response, and reproductive functions in male and femalerats. Dev. Psychobiol. 51, 259–267.
Please cite this article in press as: Uriarte, N., et al. Maternal and affective behavdx.doi.org/10.1016/j.jphysparis.2014.04.001
Veenema, A.H., Bredewold, R., Neumann, I.D., 2007. Opposite effects ofmaternal separation on intermale and maternal aggression in C57BL/6mice: link to hypothalamic vasopressin and oxytocin immunoreactivity.Psychoneuroendocrinology 32, 437–450.
Weinberg, J., Krahn, E.A., Levine, S., 1978. Differential effects of handling onexploration in male and female rats. Dev. Psychobiol. 11, 251–259.
Wurbel, H., 2001. Ideal homes? Housing effects on rodent brain and behaviour.Trends Neurosci. 24, 207–211.
Zuluaga, M.J., Agrati, D., Uriarte, N., Ferreira, A., 2014. Social aversive stimulipresented to the mother produce the precocious expression of fear in rat pups.Dev. Psychobiol. http://dx.doi.org/10.1002/dev.21199.
iors of lactating rats reared in overlapping litters. J. Physiol. (2014), http://
Copyright © 2022 FDOKUMEN




















