
Biogas Production from Anchovy Processing Waste after Fish ...
Upload
independentCategory
view
1download
0
Spawning groups of European anchovy: population structureand management implications
Nuria Sanz, Jose-Luis Garcıa-Marın, Jordi Vinas, Marina Roldan, and Carles Pla
Sanz, N., Garcıa-Marın, J-L., Vinas, J., Roldan, M., and Pla, C. 2008. Spawning groups of European anchovy: population structure andmanagement implications. – ICES Journal of Marine Science, 65: 1635–1644.
Samples of Engraulis encrasicolus from the western Mediterranean and the eastern Atlantic spawning groups were examined by proteinelectrophoresis to investigate their relevance in the genetic structure of European anchovy, and how they fit with the current defi-nition of anchovy fisheries. Samples from an earlier study were also included in the analyses to assess the temporal genetic stability ofspawning groups. Gene diversity analysis indicated greater differentiation between spawning groups within seas (GRS ¼ 0.046) thanbetween seas (GSEA ¼ 0.022), and lesser differentiation between locations within spawning groups (GLR ¼ 0.003), as well as geneticstability of spawning groups (GCL ¼ 0.002). The six spawning groups apparently involved three genetic units according to STRUCTURE
results, and the results of other analyses also suggested genetic differences between the two Atlantic spawning groups. Thegenetic substructure detected within the Atlantic agrees with the spawning groups, but does not fit easily with ICES Subdivisions.Based on these results, a revision of the current anchovy management in both Alboran Sea and Bay of Biscay areas is suggested.
Keywords: allozymes, anchovy, fishery stock, population structure.
Received 11 November 2007; accepted 30 June 2008; advance access publication 7 August 2008.
N. Sanz, J-L. Garcıa-Marın, J. Vinas, M. Roldan, and C. Pla: Laboratori d’Ictiologia Genetica, Universitat de Girona, Campus Montilivi s/n, E-17071Girona, Spain. Present address of J. Vinas: Centre Mediterrani d’Investigacions Marines i Ambientals, Passeig Marıtim de la Barceloneta, 37–49,E-08003 Barcelona, Spain. Correspondence to N. Sanz: tel: þ34 972 418168; fax: þ34 972 418277; e-mail: [email protected].
IntroductionThe European anchovy (Engraulis encrasicolus) is found along theeastern Atlantic, from Norway to South Africa, and in the entireMediterranean and Black and Azoz Seas, and is a target commer-cial species. Among small pelagic fish, it is unusual in showing agenetically complex population structure (Spanakis et al., 1989;Bembo et al., 1995, 1996; Magoulas et al., 1996, 2006). For north-western Mediterranean samples, Tudela et al. (1999) showed mor-phological variability among populations, but no genetic structureassociated with this variation, either within or between spawningaggregations. To the southwest, an aggregation in the AlboranSea that may correspond to a distinct population (Pertierra andLleonart, 1996) collapsed during the past decade as a consequenceof overfishing (Irazola et al., 1996). Studies in the Adriatic andBlack Seas revealed spatial and temporal genetic variation withinthe seas (Spanakis et al., 1989; Bembo et al., 1995, 1996). Thehigh level of differentiation among European anchovy populationshas been attributed to present-day and historical processes thathave shaped population structures (Magoulas et al., 1996, 2006;Grant, 2005). Among these processes, the recent review of Grant(2005) related the mosaic pattern of temporal and spatial geneticvariations among Mediterranean populations to the complex inter-action of geography and oceanography regimes of the region. Onthe other hand, based on morphometric, allozyme, and mitochon-drial and nuclear DNA, the studies of Borsa (2002) and Borsa et al.(2004) proposed the existence of a new cryptic anchovy species(Engraulis albidus) along the Mediterranean coast, with its maindistribution restricted to estuaries.
No widespread genetic studies have focused on the spawninggroups located in the Northeast Atlantic, specifically the Bay ofBiscay. There, biological data suggest the presence of two spawninggroups, one located along the eastern Cantabrian coast and asecond off southwestern France (Borja et al., 1998). Spanish andFrench fisheries exploit the Bay of Biscay anchovy, each countryholding a different ICES-proposed stock, separated by the CapeBreton canyon (Figure 1). However, confrontations betweenfishers are frequent when catches decrease. In the past fewdecades, anchovy fisheries in the area have dropped to historicalminima (Borja et al., 1998; AZTI-Tecnalia, 2006), leading to theclosure by the European Commission of the Bay of Biscay fisheriesin 2005 and 2006, in confrontation with the French government.
The aim of our study was to assess the genetic differencesamong spawning groups of E. encrasicolus in the northwesternMediterranean Sea, the Alboran Sea, and the northeasternAtlantic (Palomera et al., 1995; Borja et al., 1998). Data areanalysed to seek improved management and conservation.
Material and methodsSampling locations and genetic polymorphismSampling locations were selected based on putative spawninggroups (A, B, C, D, E, and F; Figure 1) identified by Palomeraet al. (1995) and Borja et al. (1998). For the northwesternMediterranean (spawning areas A, B, and C), we used literaturedata from six samples (1-Elba, 2-Liguria, 3-Lyon 1, 4-Lyon 2,5-Roses, and 7-Tarragona) captured during the PELMED 1993survey (Tudela et al., 1999), and new data from 100 fish from
# 2008 International Council for the Exploration of the Sea. Published by Oxford Journals. All rights reserved.For Permissions, please email: [email protected]
1635
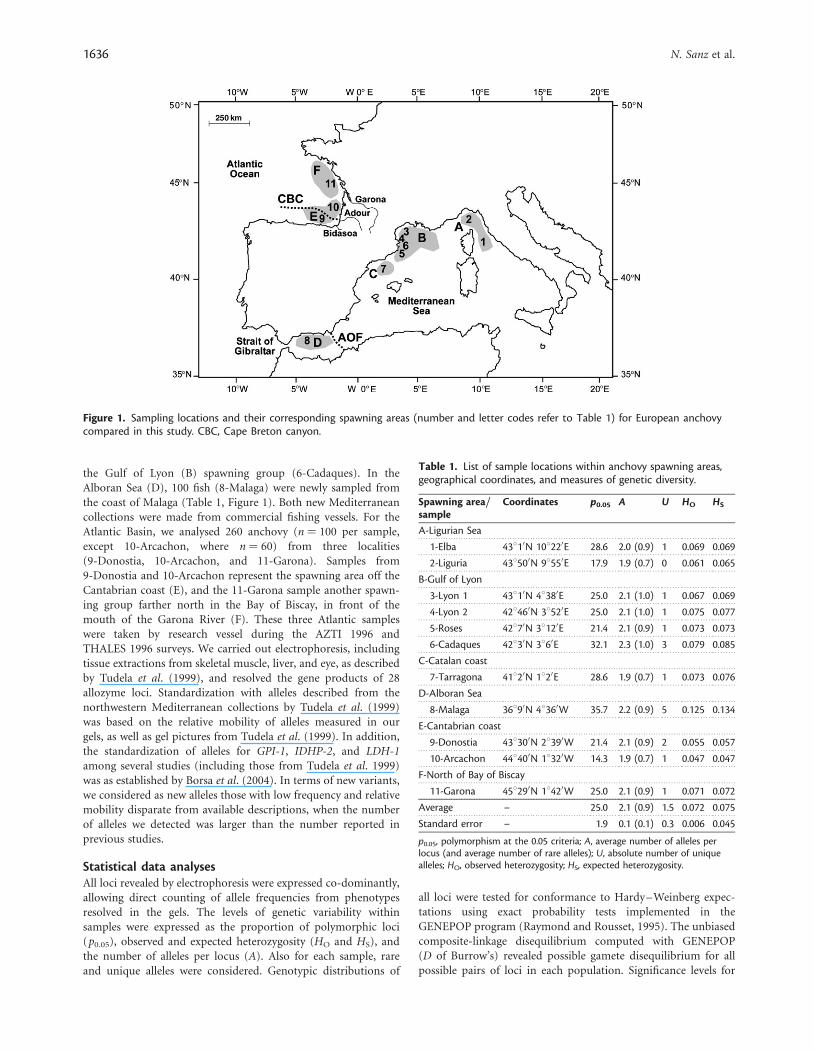
the Gulf of Lyon (B) spawning group (6-Cadaques). In theAlboran Sea (D), 100 fish (8-Malaga) were newly sampled fromthe coast of Malaga (Table 1, Figure 1). Both new Mediterraneancollections were made from commercial fishing vessels. For theAtlantic Basin, we analysed 260 anchovy (n ¼ 100 per sample,except 10-Arcachon, where n ¼ 60) from three localities(9-Donostia, 10-Arcachon, and 11-Garona). Samples from9-Donostia and 10-Arcachon represent the spawning area off theCantabrian coast (E), and the 11-Garona sample another spawn-ing group farther north in the Bay of Biscay, in front of themouth of the Garona River (F). These three Atlantic sampleswere taken by research vessel during the AZTI 1996 andTHALES 1996 surveys. We carried out electrophoresis, includingtissue extractions from skeletal muscle, liver, and eye, as describedby Tudela et al. (1999), and resolved the gene products of 28allozyme loci. Standardization with alleles described from thenorthwestern Mediterranean collections by Tudela et al. (1999)was based on the relative mobility of alleles measured in ourgels, as well as gel pictures from Tudela et al. (1999). In addition,the standardization of alleles for GPI-1, IDHP-2, and LDH-1among several studies (including those from Tudela et al. 1999)was as established by Borsa et al. (2004). In terms of new variants,we considered as new alleles those with low frequency and relativemobility disparate from available descriptions, when the numberof alleles we detected was larger than the number reported inprevious studies.
Statistical data analysesAll loci revealed by electrophoresis were expressed co-dominantly,allowing direct counting of allele frequencies from phenotypesresolved in the gels. The levels of genetic variability withinsamples were expressed as the proportion of polymorphic loci(p0.05), observed and expected heterozygosity (HO and HS), andthe number of alleles per locus (A). Also for each sample, rareand unique alleles were considered. Genotypic distributions of
all loci were tested for conformance to Hardy–Weinberg expec-tations using exact probability tests implemented in theGENEPOP program (Raymond and Rousset, 1995). The unbiasedcomposite-linkage disequilibrium computed with GENEPOP(D of Burrow’s) revealed possible gamete disequilibrium for allpossible pairs of loci in each population. Significance levels for
Figure 1. Sampling locations and their corresponding spawning areas (number and letter codes refer to Table 1) for European anchovycompared in this study. CBC, Cape Breton canyon.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . .. . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . .. . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . .. . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . .. . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . .. . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . .. . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . .. . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . .. . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . .. . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . .. . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . .. . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . .. . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . .. . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . .. . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . .. . . . . . . . . . . .. . . . . . . . .
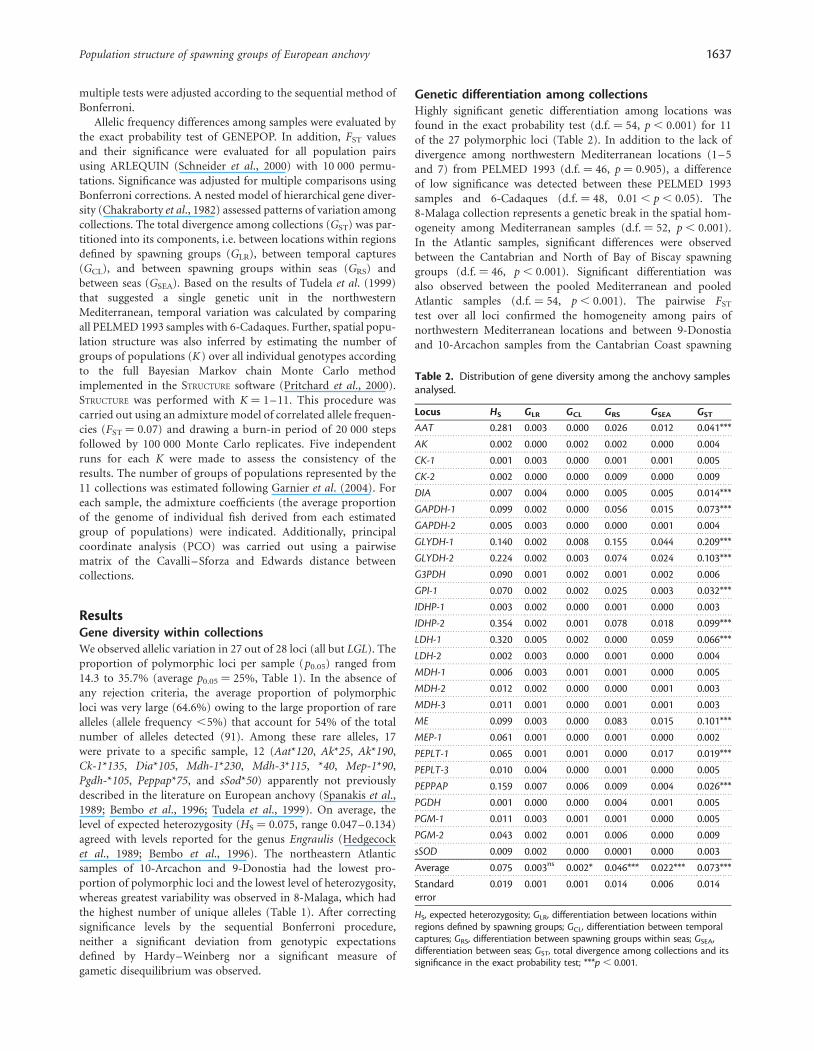
Table 1. List of sample locations within anchovy spawning areas,geographical coordinates, and measures of genetic diversity.
Spawning area/sample
Coordinates p0.05 A U HO HS
A-Ligurian Sea
1-Elba 43810N 108220E 28.6 2.0 (0.9) 1 0.069 0.069
2-Liguria 438500N 98550E 17.9 1.9 (0.7) 0 0.061 0.065
B-Gulf of Lyon
3-Lyon 1 43810N 48380E 25.0 2.1 (1.0) 1 0.067 0.069
4-Lyon 2 428460N 38520E 25.0 2.1 (1.0) 1 0.075 0.077
5-Roses 42870N 38120E 21.4 2.1 (0.9) 1 0.073 0.073
6-Cadaques 42830N 3860E 32.1 2.3 (1.0) 3 0.079 0.085
C-Catalan coast
7-Tarragona 41820N 1820E 28.6 1.9 (0.7) 1 0.073 0.076
D-Alboran Sea
8-Malaga 36890N 48360W 35.7 2.2 (0.9) 5 0.125 0.134
E-Cantabrian coast
9-Donostia 438300N 28390W 21.4 2.1 (0.9) 2 0.055 0.057
10-Arcachon 448400N 18320W 14.3 1.9 (0.7) 1 0.047 0.047
F-North of Bay of Biscay
11-Garona 458290N 18420W 25.0 2.1 (0.9) 1 0.071 0.072
Average – 25.0 2.1 (0.9) 1.5 0.072 0.075
Standard error – 1.9 0.1 (0.1) 0.3 0.006 0.045
p0.05, polymorphism at the 0.05 criteria; A, average number of alleles perlocus (and average number of rare alleles); U, absolute number of uniquealleles; HO, observed heterozygosity; HS, expected heterozygosity.
1636 N. Sanz et al.
multiple tests were adjusted according to the sequential method ofBonferroni.
Allelic frequency differences among samples were evaluated bythe exact probability test of GENEPOP. In addition, FST valuesand their significance were evaluated for all population pairsusing ARLEQUIN (Schneider et al., 2000) with 10 000 permu-tations. Significance was adjusted for multiple comparisons usingBonferroni corrections. A nested model of hierarchical gene diver-sity (Chakraborty et al., 1982) assessed patterns of variation amongcollections. The total divergence among collections (GST) was par-titioned into its components, i.e. between locations within regionsdefined by spawning groups (GLR), between temporal captures(GCL), and between spawning groups within seas (GRS) andbetween seas (GSEA). Based on the results of Tudela et al. (1999)that suggested a single genetic unit in the northwesternMediterranean, temporal variation was calculated by comparingall PELMED 1993 samples with 6-Cadaques. Further, spatial popu-lation structure was also inferred by estimating the number ofgroups of populations (K) over all individual genotypes accordingto the full Bayesian Markov chain Monte Carlo methodimplemented in the STRUCTURE software (Pritchard et al., 2000).STRUCTURE was performed with K ¼ 1–11. This procedure wascarried out using an admixture model of correlated allele frequen-cies (FST ¼ 0.07) and drawing a burn-in period of 20 000 stepsfollowed by 100 000 Monte Carlo replicates. Five independentruns for each K were made to assess the consistency of theresults. The number of groups of populations represented by the11 collections was estimated following Garnier et al. (2004). Foreach sample, the admixture coefficients (the average proportionof the genome of individual fish derived from each estimatedgroup of populations) were indicated. Additionally, principalcoordinate analysis (PCO) was carried out using a pairwisematrix of the Cavalli –Sforza and Edwards distance betweencollections.
ResultsGene diversity within collectionsWe observed allelic variation in 27 out of 28 loci (all but LGL). Theproportion of polymorphic loci per sample (p0.05) ranged from14.3 to 35.7% (average p0.05 ¼ 25%, Table 1). In the absence ofany rejection criteria, the average proportion of polymorphicloci was very large (64.6%) owing to the large proportion of rarealleles (allele frequency ,5%) that account for 54% of the totalnumber of alleles detected (91). Among these rare alleles, 17were private to a specific sample, 12 (Aat*120, Ak*25, Ak*190,Ck-1*135, Dia*105, Mdh-1*230, Mdh-3*115, *40, Mep-1*90,Pgdh-*105, Peppap*75, and sSod*50) apparently not previouslydescribed in the literature on European anchovy (Spanakis et al.,1989; Bembo et al., 1996; Tudela et al., 1999). On average, thelevel of expected heterozygosity (HS ¼ 0.075, range 0.047–0.134)agreed with levels reported for the genus Engraulis (Hedgecocket al., 1989; Bembo et al., 1996). The northeastern Atlanticsamples of 10-Arcachon and 9-Donostia had the lowest pro-portion of polymorphic loci and the lowest level of heterozygosity,whereas greatest variability was observed in 8-Malaga, which hadthe highest number of unique alleles (Table 1). After correctingsignificance levels by the sequential Bonferroni procedure,neither a significant deviation from genotypic expectationsdefined by Hardy–Weinberg nor a significant measure ofgametic disequilibrium was observed.
Genetic differentiation among collectionsHighly significant genetic differentiation among locations wasfound in the exact probability test (d.f. ¼ 54, p , 0.001) for 11of the 27 polymorphic loci (Table 2). In addition to the lack ofdivergence among northwestern Mediterranean locations (1–5and 7) from PELMED 1993 (d.f. ¼ 46, p ¼ 0.905), a differenceof low significance was detected between these PELMED 1993samples and 6-Cadaques (d.f. ¼ 48, 0.01 , p , 0.05). The8-Malaga collection represents a genetic break in the spatial hom-ogeneity among Mediterranean samples (d.f. ¼ 52, p , 0.001).In the Atlantic samples, significant differences were observedbetween the Cantabrian and North of Bay of Biscay spawninggroups (d.f. ¼ 46, p , 0.001). Significant differentiation wasalso observed between the pooled Mediterranean and pooledAtlantic samples (d.f. ¼ 54, p , 0.001). The pairwise FST
test over all loci confirmed the homogeneity among pairs ofnorthwestern Mediterranean locations and between 9-Donostiaand 10-Arcachon samples from the Cantabrian Coast spawning
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . .
Table 2. Distribution of gene diversity among the anchovy samplesanalysed.
Locus HS GLR GCL GRS GSEA GST
AAT 0.281 0.003 0.000 0.026 0.012 0.041***
AK 0.002 0.000 0.002 0.002 0.000 0.004
CK-1 0.001 0.003 0.000 0.001 0.001 0.005
CK-2 0.002 0.000 0.000 0.009 0.000 0.009
DIA 0.007 0.004 0.000 0.005 0.005 0.014***
GAPDH-1 0.099 0.002 0.000 0.056 0.015 0.073***
GAPDH-2 0.005 0.003 0.000 0.000 0.001 0.004
GLYDH-1 0.140 0.002 0.008 0.155 0.044 0.209***
GLYDH-2 0.224 0.002 0.003 0.074 0.024 0.103***
G3PDH 0.090 0.001 0.002 0.001 0.002 0.006
GPI-1 0.070 0.002 0.002 0.025 0.003 0.032***
IDHP-1 0.003 0.002 0.000 0.001 0.000 0.003
IDHP-2 0.354 0.002 0.001 0.078 0.018 0.099***
LDH-1 0.320 0.005 0.002 0.000 0.059 0.066***
LDH-2 0.002 0.003 0.000 0.001 0.000 0.004
MDH-1 0.006 0.003 0.001 0.001 0.000 0.005
MDH-2 0.012 0.002 0.000 0.000 0.001 0.003
MDH-3 0.011 0.001 0.000 0.001 0.001 0.003
ME 0.099 0.003 0.000 0.083 0.015 0.101***
MEP-1 0.061 0.001 0.000 0.001 0.000 0.002
PEPLT-1 0.065 0.001 0.001 0.000 0.017 0.019***
PEPLT-3 0.010 0.004 0.000 0.001 0.000 0.005
PEPPAP 0.159 0.007 0.006 0.009 0.004 0.026***
PGDH 0.001 0.000 0.000 0.004 0.001 0.005
PGM-1 0.011 0.003 0.001 0.001 0.000 0.005
PGM-2 0.043 0.002 0.001 0.006 0.000 0.009
sSOD 0.009 0.002 0.000 0.0001 0.000 0.003
Average 0.075 0.003ns 0.002* 0.046*** 0.022*** 0.073***
Standarderror
0.019 0.001 0.001 0.014 0.006 0.014
HS, expected heterozygosity; GLR, differentiation between locations withinregions defined by spawning groups; GCL, differentiation between temporalcaptures; GRS, differentiation between spawning groups within seas; GSEA,differentiation between seas; GST, total divergence among collections and itssignificance in the exact probability test; ***p , 0.001.
Population structure of spawning groups of European anchovy 1637
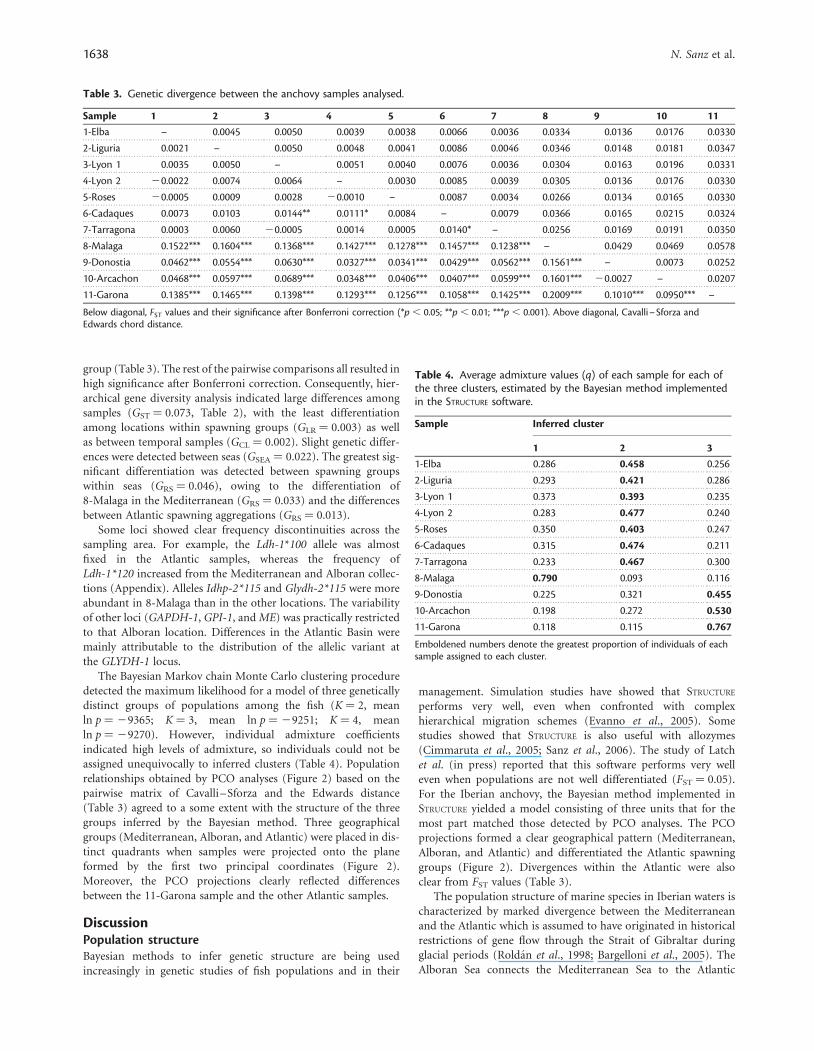
group (Table 3). The rest of the pairwise comparisons all resulted inhigh significance after Bonferroni correction. Consequently, hier-archical gene diversity analysis indicated large differences amongsamples (GST ¼ 0.073, Table 2), with the least differentiationamong locations within spawning groups (GLR ¼ 0.003) as wellas between temporal samples (GCL ¼ 0.002). Slight genetic differ-ences were detected between seas (GSEA ¼ 0.022). The greatest sig-nificant differentiation was detected between spawning groupswithin seas (GRS ¼ 0.046), owing to the differentiation of8-Malaga in the Mediterranean (GRS ¼ 0.033) and the differencesbetween Atlantic spawning aggregations (GRS ¼ 0.013).
Some loci showed clear frequency discontinuities across thesampling area. For example, the Ldh-1*100 allele was almostfixed in the Atlantic samples, whereas the frequency ofLdh-1*120 increased from the Mediterranean and Alboran collec-tions (Appendix). Alleles Idhp-2*115 and Glydh-2*115 were moreabundant in 8-Malaga than in the other locations. The variabilityof other loci (GAPDH-1, GPI-1, and ME) was practically restrictedto that Alboran location. Differences in the Atlantic Basin weremainly attributable to the distribution of the allelic variant atthe GLYDH-1 locus.
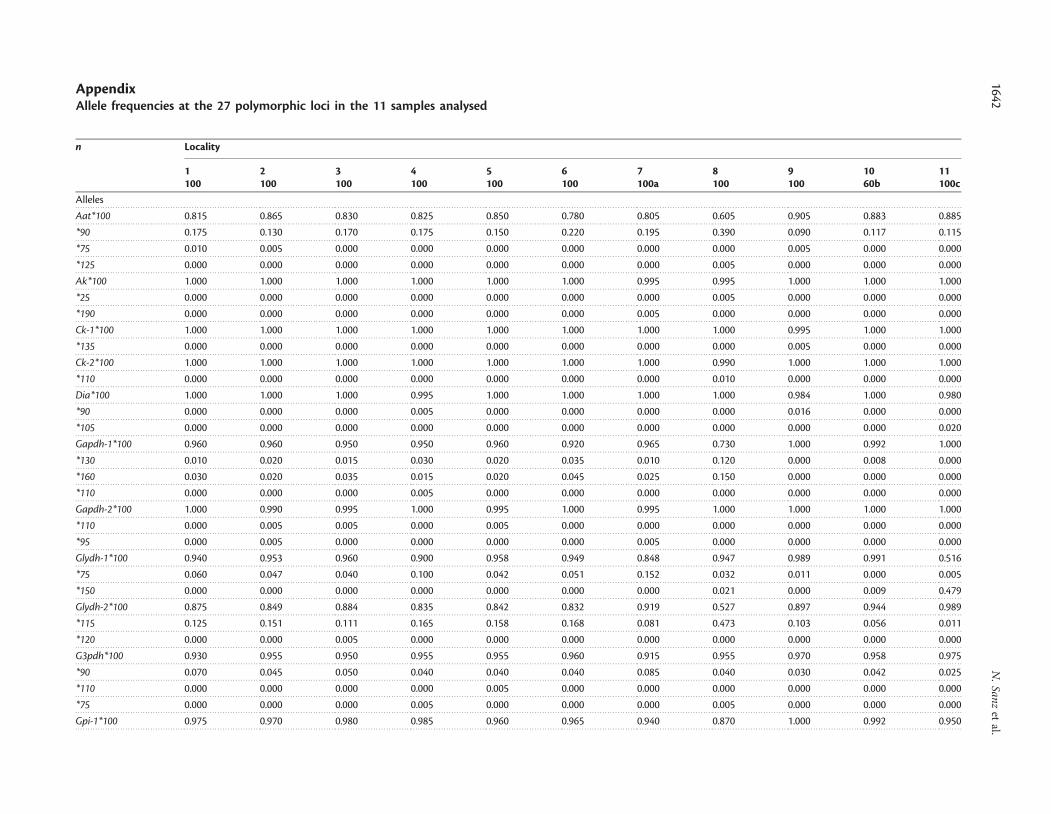
The Bayesian Markov chain Monte Carlo clustering proceduredetected the maximum likelihood for a model of three geneticallydistinct groups of populations among the fish (K ¼ 2, meanln p ¼ 29365; K ¼ 3, mean ln p ¼ 29251; K ¼ 4, meanln p ¼ 29270). However, individual admixture coefficientsindicated high levels of admixture, so individuals could not beassigned unequivocally to inferred clusters (Table 4). Populationrelationships obtained by PCO analyses (Figure 2) based on thepairwise matrix of Cavalli–Sforza and the Edwards distance(Table 3) agreed to a some extent with the structure of the threegroups inferred by the Bayesian method. Three geographicalgroups (Mediterranean, Alboran, and Atlantic) were placed in dis-tinct quadrants when samples were projected onto the planeformed by the first two principal coordinates (Figure 2).Moreover, the PCO projections clearly reflected differencesbetween the 11-Garona sample and the other Atlantic samples.
DiscussionPopulation structureBayesian methods to infer genetic structure are being usedincreasingly in genetic studies of fish populations and in their
management. Simulation studies have showed that STRUCTURE
performs very well, even when confronted with complexhierarchical migration schemes (Evanno et al., 2005). Somestudies showed that STRUCTURE is also useful with allozymes(Cimmaruta et al., 2005; Sanz et al., 2006). The study of Latchet al. (in press) reported that this software performs very welleven when populations are not well differentiated (FST ¼ 0.05).For the Iberian anchovy, the Bayesian method implemented inSTRUCTURE yielded a model consisting of three units that for themost part matched those detected by PCO analyses. The PCOprojections formed a clear geographical pattern (Mediterranean,Alboran, and Atlantic) and differentiated the Atlantic spawninggroups (Figure 2). Divergences within the Atlantic were alsoclear from FST values (Table 3).
The population structure of marine species in Iberian waters ischaracterized by marked divergence between the Mediterraneanand the Atlantic which is assumed to have originated in historicalrestrictions of gene flow through the Strait of Gibraltar duringglacial periods (Roldan et al., 1998; Bargelloni et al., 2005). TheAlboran Sea connects the Mediterranean Sea to the Atlantic
. . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . .
Table 3. Genetic divergence between the anchovy samples analysed.
Sample 1 2 3 4 5 6 7 8 9 10 11
1-Elba – 0.0045 0.0050 0.0039 0.0038 0.0066 0.0036 0.0334 0.0136 0.0176 0.0330
2-Liguria 0.0021 – 0.0050 0.0048 0.0041 0.0086 0.0046 0.0346 0.0148 0.0181 0.0347
3-Lyon 1 0.0035 0.0050 – 0.0051 0.0040 0.0076 0.0036 0.0304 0.0163 0.0196 0.0331
4-Lyon 2 20.0022 0.0074 0.0064 – 0.0030 0.0085 0.0039 0.0305 0.0136 0.0176 0.0330
5-Roses 20.0005 0.0009 0.0028 20.0010 – 0.0087 0.0034 0.0266 0.0134 0.0165 0.0330
6-Cadaques 0.0073 0.0103 0.0144** 0.0111* 0.0084 – 0.0079 0.0366 0.0165 0.0215 0.0324
7-Tarragona 0.0003 0.0060 20.0005 0.0014 0.0005 0.0140* – 0.0256 0.0169 0.0191 0.0350
8-Malaga 0.1522*** 0.1604*** 0.1368*** 0.1427*** 0.1278*** 0.1457*** 0.1238*** – 0.0429 0.0469 0.0578
9-Donostia 0.0462*** 0.0554*** 0.0630*** 0.0327*** 0.0341*** 0.0429*** 0.0562*** 0.1561*** – 0.0073 0.0252
10-Arcachon 0.0468*** 0.0597*** 0.0689*** 0.0348*** 0.0406*** 0.0407*** 0.0599*** 0.1601*** 20.0027 – 0.0207
11-Garona 0.1385*** 0.1465*** 0.1398*** 0.1293*** 0.1256*** 0.1058*** 0.1425*** 0.2009*** 0.1010*** 0.0950*** –
Below diagonal, FST values and their significance after Bonferroni correction (*p , 0.05; **p , 0.01; ***p , 0.001). Above diagonal, Cavalli–Sforza andEdwards chord distance.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . .
Table 4. Average admixture values (q) of each sample for each ofthe three clusters, estimated by the Bayesian method implementedin the STRUCTURE software.
Sample Inferred cluster
1 2 3
1-Elba 0.286 0.458 0.256
2-Liguria 0.293 0.421 0.286
3-Lyon 1 0.373 0.393 0.235
4-Lyon 2 0.283 0.477 0.240
5-Roses 0.350 0.403 0.247
6-Cadaques 0.315 0.474 0.211
7-Tarragona 0.233 0.467 0.300
8-Malaga 0.790 0.093 0.116
9-Donostia 0.225 0.321 0.455
10-Arcachon 0.198 0.272 0.530
11-Garona 0.118 0.115 0.767
Emboldened numbers denote the greatest proportion of individuals of eachsample assigned to each cluster.
1638 N. Sanz et al.

Ocean and is bounded by the Iberian margin to the north and theNorth African margin to the south. Surface circulation in theAlboran Sea is characterized by the entrance of a jet of Atlanticsurface water 200–300-m thick. The interaction of this Atlanticjet with the denser, more-saline Mediterranean water results inthe formation of a quasi-continuous front, known as theAlmeria–Oran front (AOF). As a result of the AOF, surfaceAtlantic water entering the Mediterranean Basin is deflectedfrom Almeria on the Spanish coast south towards Oran on theAlgerian coast of Africa (Tintore et al., 1988). This water circula-tion apparently defines the current isolation of Atlantic andnorthwestern Mediterranean populations. Studies on Mytilus gal-loprovincialis (Quesada et al., 1995), Dicentrarchus labrax (Naciriet al., 1999), and Merluccius merluccius (Roldan et al., 1998;Cimmaruta et al., 2005) have reported high genetic similaritybetween samples from the Alboran Sea and those from adjacentAtlantic populations, with a genetic break at the eastern marginof the Alboran Sea. Mitochondrial DNA diversity in Europeananchovy also agrees with this pattern of population structure(Magoulas et al., 2006). However, the shallow between-basindivergence of anchovy populations (GSEA ¼ 0.022) that we havedetected and the admixture levels indicated by the Bayesianmethods of STRUCTURE (Table 4) suggests that the signals ofcurrent gene flow and ancestral isolation are overimposed. Genediversity analysis indicated that isolation among spawninggroups (GRS ¼ 0.046) resulted in greater genetic divergence thanhistorical isolation through the Gibraltar Strait and AOF.
In the Atlantic Basin, the significant genetic differencesdetected between the two spawning aggregations contrasted withthe homogeneity observed previously among northwesternMediterranean groups (Tudela et al., 1999). Bembo et al. (1996)suggested that physical and hydrographic barriers were responsiblefor the population structures observed. The two spawning
aggregations in the Atlantic (Figure 1) are related to feedingareas located at the mouths of the Garona River (North of Bayof Biscay spawning group) and the Adour and Cantabrian Rivers(Cantabrian coast spawning group), respectively (Masse et al.,1995). Acoustic surveys indicate that in years with strong recruit-ment, dispersal of eggs, larvae, and young-of-the-year anchovythroughout the entire Bay of Biscay (Motos et al., 1996) couldlead to high levels of gene flow that would tend to homogenizedistinct populations. However, in years with reduced spawningbiomass, the concentration of anchovy eggs near the mouths ofthe Garona and Adour–Bidasoa would leave a gap betweenthese primary spawning areas (Motos et al., 1996). Evidencesuggesting contraction and expansion events has been documen-ted already for other areas inhabited by the European anchovy,and also for the East Pacific anchovy (Engraulis mordax), theIberian sardine (Sardina pilchardus), and the Pacific sardine(Sardinops sagax), and the resulting patterns are best understoodin light of metapopulation dynamics (Carrera and Porteiro,2003; Lecomte et al., 2004; Laurent et al., 2006).
All the analyses have demonstrated the uniqueness of the8-Malaga collection from the Alboran Sea. According toMagoulas et al. (2006), anchovy in the Alboran Sea have aprofile of mtDNA typical of Atlantic populations in the CanaryCurrent. Therefore, the recently collapsed fishery in the AlboranSea could recover by the immigration of Canary Currentanchovy into the Alboran Sea. Nevertheless, without Atlantic col-lections from adjacent areas, we cannot rule out alternativehypotheses involving either putative endogenous recovery or dis-persal from other sources. Endogenous recovery in a situation oflittle competition and relaxed selection allows an increase in themutation load and the accumulation of rare and new alleles suchas those detected in the 8-Malaga sample (Appendix). On theother hand, the allele frequencies at loci IDH-2 and LDH-1 inthe 8-Malaga collection agree with the frequencies distinguishingthe newly described species, E. albidus (Borsa, 2002). Despite theprimary habitat of this species being restricted to estuaries,Borsa et al. (2004) reported that E. albidus could leave inshorehabitats and enter nearby coastal marine waters. Therefore, athird hypothesis involving the presence of this cryptic species inthe Alboran Sea cannot be excluded. If this was indeed the case,the high divergence reported between Alboran and northwesternMediterranean populations would be based on the differencesbetween the two species. Moreover, it would demonstrateE. albidus to be more widespread than previously thought.
Management implicationsThe pattern of population structure defined for the northeasternAtlantic anchovy did not fit with ICES Subdivisions separated bythe Cape Breton canyon (Figure 1). The current managementpolicy could, therefore, be revised in view of the incomplete iso-lation between anchovy spawning groups and the critical situationof these Atlantic anchovy stocks. Because boundaries betweenstocks can change annually, we suggest that both genetic groupsbe managed together, as proposed by the European Commissionwhen closing the ICES-based anchovy fisheries in the area.
The population structure we have observed amongMediterranean populations invalidates the current managementpolicy that groups the Alboran Sea and northwesternMediterranean anchovy populations into a single stock. Ourresults strongly point to the existence of a different stock in theAlboran Sea and would justify modification of the present
Figure 2. PCO based on the Cavalli–Sforza and Edwards distancebetween anchovy collections. Samples are projected onto the planeformed by the two principal coordinate axes. Ellipsoids groupingsimilar populations are depicted. Spawning areas are indicated bytheir letter identifiers between parentheses (number and letter codesrefer to Table 1).
Population structure of spawning groups of European anchovy 1639

management approach in the area. Until broader ranging studiesconfirm the similarity of anchovy from the Alboran Sea and theadjacent Atlantic, a management strategy based on the assumedisolation of the Alboran Sea would warrant conservation of thisstock. Moreover, although the high genetic diversity in the8-Malaga sample could be interpreted as a sign of recovery ofthe Alboran fishery, this area was classified as containing a stockoutside Safe Biological Limits in 2001 (ICES, FAO and GFCM,SAC Report, 2002).
Finally, the current overfishing of most anchovy stocks couldwell lead not only to commercial collapse but also to the biologicalextinction of some populations. In contrast to the perception thatmarine fish are highly resilient to large population reductions,Hutchings (2001) showed little empirical support for a rapidrecovery from prolonged declines, even for some clupeids. Inour view, and given the considerable commercial value ofanchovy fisheries, genetic monitoring needs to be implementedso that permanent, accurate, geographic, and temporal assess-ments of the state of anchovy stocks, at least those consideredoutside safe biological limits, can be obtained.
AcknowledgementsThis research was carried out within the objectives of the researchproject AMB95-0339 of the Spanish CICYT. NS was a FIAP-fellowof the Autonomous Government of Catalunya. We thankA. Giraldez (IEO-Spain), A. Uriarte (AZTI-Spain), and J. Masse(IFREMER-France) for their help in collecting the samples.
ReferencesAZTI-Tecnalia. 2006. Situacion del Stock de la Anchoa tras la
Campana BIOMAN 06 llevada a cabo por AZTI-Tecnalia. Spain.Cited 2 May 2007. http://www.azti.es/muestracontenido.asp?idcontenido=687&content=6&nodo1=0&nodo2=0.
Bargelloni, L., Alarcon, J. A., Alvarez, M. C., Penzo, E., Magoulas, A.,Palma, J., and Patarnello, T. 2005. The Atlantic–Mediterraneantransition: discordant genetic patterns in two seabream species,Diplodus puntazzo (Cetti) and Diplodus sargus (L.). MolecularPhylogenetics and Evolution, 36: 523–535.
Bembo, D. G., Carvalho, G. R., Cingolani, N., and Pitcher, T. J. 1996.Electrophoretic analysis of stock structure in northernMediterranean anchovies, Engraulis encrasicolus. ICES Journal ofMarine Science, 53: 115–128.
Bembo, D. G., Carvalho, G. R., Snow, M., Cingolani, N., and Pitcher,T. J. 1995. Stock discrimination among European anchovies,Engraulis encrasicolus, by means of PCR-amplified mitochondrialDNA analysis. Fishery Bulletin US, 94: 31–40.
Borja, A., Uriarte, A., Egana, J., Motos, L., and Valencia, V. 1998.Relationships between anchovy (Engraulis encrasicolus) recruit-ment and environment in the Bay of Biscay (1967–1996).Fisheries Oceanography, 7: 375–380.
Borsa, P. 2002. Allozyme, mitochondrial-DNA, and morphometricvariability indicate cryptic species of anchovy (Engraulis encrasico-lus). Biological Journal of the Linnean Society, 75: 261–269.
Borsa, P., Collet, A., and Durand, J. D. 2004. Nuclear-DNA markersconfirm the presence of two anchovy species in theMediterranean. Comptes Rendus Biologies, 327: 1113–1123.
Carrera, P., and Porteiro, C. 2003. Stock dynamic of the Iberiansardine (Sardina pilchardus, W.) and its implication on thefishery off Galicia (NW Spain). Scientia Marina, 67: 245–258.
Chakraborty, R., Haag, M., Ryman, N., and Stahl, G. 1982.Hierarchical gene diversity analysis and its application to browntrout population data. Hereditas, 97: 17–21.
Cimmaruta, R., Bondanelli, P., and Nascetti, G. 2005. Genetic struc-ture and environmental heterogeneity in the European hake(Merluccius merluccius). Molecular Ecology, 14: 2577–2591.
Evanno, G., Regnaut, S., and Goudet, J. 2005. Detecting the number ofclusters of individuals using the software STRUCTURE: a simulationstudy. Molecular Ecology, 14: 2611–2620.
Garnier, S., Alibert, P., Audiot, P., Prieur, B., and Rasplus, J. Y. 2004.Isolation by distance and sharp discontinuities in gene frequencies:implications for the phylogeography of an alpine insect species,Carabus solieri. Molecular Ecology, 13: 1883–1897.
Grant, W. S. 2005. A second look at mitochondrial DNA variability inEuropean anchovy (Engraulis encrasicolus): assessing models ofpopulation structure and the Black Sea isolation hypothesis.Genetica, 125: 293–309.
Hedgecock, D., Hutchinson, E. S., Li, G., Sly, F. L., and Nelson, K.1989. Genetic and morphometric variation in the Pacific sardine,Sardinops sagax caerulea: comparisons and contrasts with historicaldata and with variability in the northern anchovy, Engraulismordax. Fishery Bulletin US, 87: 653–671.
Hutchings, J. A. 2001. Conservation biology of marine fishes: perceptionsand caveats regarding assignment of extinction risk. CanadianJournal of Fisheries and Aquatic Sciences, 58: 108–121.
ICES, FAO and GFCM, SAC Report. 2002. Compiled by ETC Water.# 1993–2006, European Environment Agency. EEA. Cited 2May 2007. http://themes.eea.europa.eu/Sectors_and_activities/fishery/indicators/FISH01a,2004.05.
Irazola, M., Lucchetti, A., Lleonart, J., Ocana, A., Tapia, J. M., andTudela, S. 1996. La pesca en el siglo XXI. CCOO, Federacion delTransporte, Barcelona, Spain.
Latch, E. K., Dharmarajan, G., Glaubitz, J. C., and Rhodes, O. E. 2006.Relative performance of Bayesian clustering software for inferringpopulation substructure and individual assignment at low levelsof population differentiation. Conservation Genetics, 7: 295–302.
Laurent, V., Voisin, M., and Planes, S. 2006. Genetic clines in the Bayof Biscay provide estimates of migration for Sardina pilchardus.Journal of Heredity, 97: 81–88.
Lecomte, F., Grant, W. S., Dodson, J. J., Rodrıguez-Sanchez, R., andBowen, B. W. 2004. Living with uncertainty: genetic imprintsof climate shifts in East Pacific anchovy (Engraulis mordax) andsardine (Sardinops sagax). Molecular Ecology, 13: 2169–2182.
Magoulas, A., Castilho, R., Caetano, S., Marcato, S., and Patarnello, T.2006. Mitochondrial DNA reveals a mosaic pattern of phylogeogra-phical structure in Atlantic and Mediterranean populations ofanchovy (Engraulis encrasicolus). Molecular Phylogenetics andEvolution, 39: 734–746.
Magoulas, A., Tsimenides, N., and Zouros, E. 1996. MitochondrialDNA phylogeny and the reconstruction of the population historyof a species: the case of the European anchovy (Engraulis encrasico-lus). Molecular Biology and Evolution, 13: 178–190.
Masse, J., Leroy, C., Halgand, D., and Beillois, P. 1995. Anchovy(Engraulis encrasicolus, L.) adult and egg distribution in the Bayof Biscay, in relation to environmental conditions, as observedduring acoustic surveys (1990, 1991, 1992). Actas del IVColoquio Internacional sobre Oceanografıa del Golfo de Vizcaya:281–283.
Motos, L., Uriarte, A., and Valencia, V. 1996. The spawning environ-ment of the Bay of Biscay anchovy (Engraulis encrasicolus L.).Scientia Marina, 60: 117–140.
Naciri, M., Lemaire, C., Borsa, P., and Bonhomme, F. 1999. Geneticstudy of the Atlantic/Mediterranean transition in sea bass(Dicentrarchus labrax). Journal of Heredity, 90: 591–596.
Palomera, I., Garcıa, A., and Giovanardi, O. 1995. NorthwesternMediterranean anchovy spawning grounds off the Catalan Sea,Gulf of Lyons and the Ligurian Sea during 1992 and 1993.Rapports CIESM, 34: 252.
Pertierra, J. P., and Lleonart, J. 1996. NW Mediterranean anchovyfisheries. Scientia Marina, 60: 257–267.
1640 N. Sanz et al.

Pritchard, J. K., Stephens, M., and Donnelly, P. 2000. Inference ofpopulation structure using multilocus genotype data. Genetics,155: 945–959.
Quesada, H., Beynon, C. M., and Skibinski, D. O. F. 1995. A mito-chondrial DNA discontinuity in the mussel Mytilus gallopro-vincialis Lmk: Pleistocene vicariance biogeography and secondaryintergradation. Molecular Biology and Evolution, 12: 521–524.
Raymond, M., and Rousset, F. 1995. GENEPOP (version 1.2): a popu-lation genetics software for exact test and ecumenicist. Journal ofHeredity, 86: 248–249.
Roldan, M. I., Garcıa-Marın, J. L., Utter, F. M., and Pla, C. 1998.Population genetic structure of European hake, Merluccius merluc-cius. Heredity, 81: 327–334.
Sanz, N., Araguas, R. M., Fernandez, R., Vera, M., and Garcıa-Marın, J. L. Efficiency of markers and methods for detectinghybrids and introgression in stocked populations. ConservationGenetics (in press).
Schneider, S., Roessli, D., and Excoffier, L. 2000. Arlequin Ver. 2.000: aSoftware for Population Genetic Data Analysis. Genetics andBiometry Laboratory, University of Geneva, Switzerland.
Spanakis, E., Tsimenides, N., and Zouros, E. 1989. Genetic differencesbetween populations of sardine, Sardina pilchardus, and anchovy,Engraulis encrasicolus, in the Aegean and Ionian seas. Journal ofFish Biology, 35: 417–437.
Tintore, J., LaViolette, P. E., Blade, I., and Cruzado, A. 1988. Astudy of an intense density front in the eastern Alboran Sea: theAlmeria–Oran front. Journal of Physical Oceanography, 18:1384–1397.
Tudela, S., Garcıa-Marın, J. L., and Pla, C. 1999. Genetic structure ofthe European anchovy, Engraulis encrasicolus L., in the north-westMediterranean. Journal of Experimental Marine Biology andEcology, 234: 95–109.
doi:10.1093/icesjms/fsn128
Population structure of spawning groups of European anchovy 1641
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
n Locality
1 2 3 4 5 6 7 8 9 10 11100 100 100 100 100 100 100a 100 100 60b 100c
Alleles
Aat*100 0.815 0.865 0.830 0.825 0.850 0.780 0.805 0.605 0.905 0.883 0.885
*90 0.175 0.130 0.170 0.175 0.150 0.220 0.195 0.390 0.090 0.117 0.115
*75 0.010 0.005 0.000 0.000 0.000 0.000 0.000 0.000 0.005 0.000 0.000
*125 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.005 0.000 0.000 0.000
Ak*100 1.000 1.000 1.000 1.000 1.000 1.000 0.995 0.995 1.000 1.000 1.000
*25 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.005 0.000 0.000 0.000
*190 0.000 0.000 0.000 0.000 0.000 0.000 0.005 0.000 0.000 0.000 0.000
Ck-1*100 1.000 1.000 1.000 1.000 1.000 1.000 1.000 1.000 0.995 1.000 1.000
*135 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.005 0.000 0.000
Ck-2*100 1.000 1.000 1.000 1.000 1.000 1.000 1.000 0.990 1.000 1.000 1.000
*110 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.010 0.000 0.000 0.000
Dia*100 1.000 1.000 1.000 0.995 1.000 1.000 1.000 1.000 0.984 1.000 0.980
*90 0.000 0.000 0.000 0.005 0.000 0.000 0.000 0.000 0.016 0.000 0.000
*105 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.020
Gapdh-1*100 0.960 0.960 0.950 0.950 0.960 0.920 0.965 0.730 1.000 0.992 1.000
*130 0.010 0.020 0.015 0.030 0.020 0.035 0.010 0.120 0.000 0.008 0.000
*160 0.030 0.020 0.035 0.015 0.020 0.045 0.025 0.150 0.000 0.000 0.000
*110 0.000 0.000 0.000 0.005 0.000 0.000 0.000 0.000 0.000 0.000 0.000
Gapdh-2*100 1.000 0.990 0.995 1.000 0.995 1.000 0.995 1.000 1.000 1.000 1.000
*110 0.000 0.005 0.005 0.000 0.005 0.000 0.000 0.000 0.000 0.000 0.000
*95 0.000 0.005 0.000 0.000 0.000 0.000 0.005 0.000 0.000 0.000 0.000
Glydh-1*100 0.940 0.953 0.960 0.900 0.958 0.949 0.848 0.947 0.989 0.991 0.516
*75 0.060 0.047 0.040 0.100 0.042 0.051 0.152 0.032 0.011 0.000 0.005
*150 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.021 0.000 0.009 0.479
Glydh-2*100 0.875 0.849 0.884 0.835 0.842 0.832 0.919 0.527 0.897 0.944 0.989
*115 0.125 0.151 0.111 0.165 0.158 0.168 0.081 0.473 0.103 0.056 0.011
*120 0.000 0.000 0.005 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000
G3pdh*100 0.930 0.955 0.950 0.955 0.955 0.960 0.915 0.955 0.970 0.958 0.975
*90 0.070 0.045 0.050 0.040 0.040 0.040 0.085 0.040 0.030 0.042 0.025
*110 0.000 0.000 0.000 0.000 0.005 0.000 0.000 0.000 0.000 0.000 0.000
*75 0.000 0.000 0.000 0.005 0.000 0.000 0.000 0.005 0.000 0.000 0.000
Gpi-1*100 0.975 0.970 0.980 0.985 0.960 0.965 0.940 0.870 1.000 0.992 0.950
AppendixAllele frequencies at the 27 polymorphic loci in the 11 samples analysed
1642N
.San
zet
al.
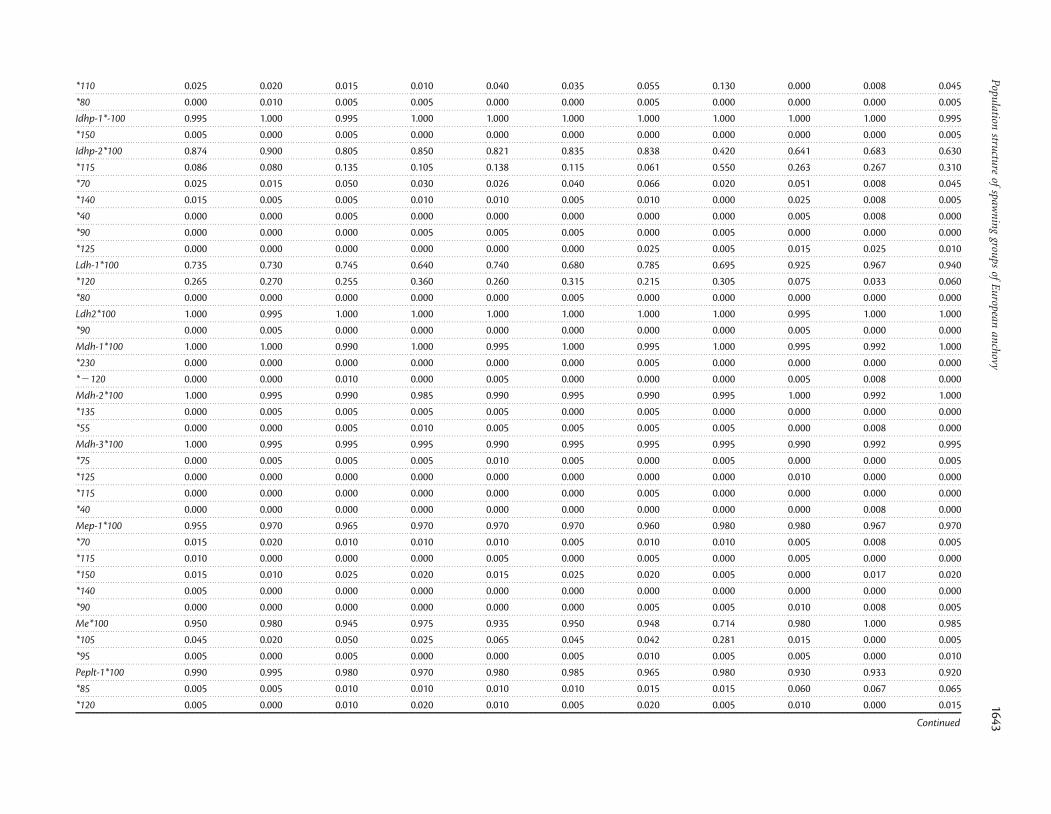
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
*110 0.025 0.020 0.015 0.010 0.040 0.035 0.055 0.130 0.000 0.008 0.045
*80 0.000 0.010 0.005 0.005 0.000 0.000 0.005 0.000 0.000 0.000 0.005
Idhp-1*-100 0.995 1.000 0.995 1.000 1.000 1.000 1.000 1.000 1.000 1.000 0.995
*150 0.005 0.000 0.005 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.005
Idhp-2*100 0.874 0.900 0.805 0.850 0.821 0.835 0.838 0.420 0.641 0.683 0.630
*115 0.086 0.080 0.135 0.105 0.138 0.115 0.061 0.550 0.263 0.267 0.310
*70 0.025 0.015 0.050 0.030 0.026 0.040 0.066 0.020 0.051 0.008 0.045
*140 0.015 0.005 0.005 0.010 0.010 0.005 0.010 0.000 0.025 0.008 0.005
*40 0.000 0.000 0.005 0.000 0.000 0.000 0.000 0.000 0.005 0.008 0.000
*90 0.000 0.000 0.000 0.005 0.005 0.005 0.000 0.005 0.000 0.000 0.000
*125 0.000 0.000 0.000 0.000 0.000 0.000 0.025 0.005 0.015 0.025 0.010
Ldh-1*100 0.735 0.730 0.745 0.640 0.740 0.680 0.785 0.695 0.925 0.967 0.940
*120 0.265 0.270 0.255 0.360 0.260 0.315 0.215 0.305 0.075 0.033 0.060
*80 0.000 0.000 0.000 0.000 0.000 0.005 0.000 0.000 0.000 0.000 0.000
Ldh2*100 1.000 0.995 1.000 1.000 1.000 1.000 1.000 1.000 0.995 1.000 1.000
*90 0.000 0.005 0.000 0.000 0.000 0.000 0.000 0.000 0.005 0.000 0.000
Mdh-1*100 1.000 1.000 0.990 1.000 0.995 1.000 0.995 1.000 0.995 0.992 1.000
*230 0.000 0.000 0.000 0.000 0.000 0.000 0.005 0.000 0.000 0.000 0.000
*2120 0.000 0.000 0.010 0.000 0.005 0.000 0.000 0.000 0.005 0.008 0.000
Mdh-2*100 1.000 0.995 0.990 0.985 0.990 0.995 0.990 0.995 1.000 0.992 1.000
*135 0.000 0.005 0.005 0.005 0.005 0.000 0.005 0.000 0.000 0.000 0.000
*55 0.000 0.000 0.005 0.010 0.005 0.005 0.005 0.005 0.000 0.008 0.000
Mdh-3*100 1.000 0.995 0.995 0.995 0.990 0.995 0.995 0.995 0.990 0.992 0.995
*75 0.000 0.005 0.005 0.005 0.010 0.005 0.000 0.005 0.000 0.000 0.005
*125 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.010 0.000 0.000
*115 0.000 0.000 0.000 0.000 0.000 0.000 0.005 0.000 0.000 0.000 0.000
*40 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.008 0.000
Mep-1*100 0.955 0.970 0.965 0.970 0.970 0.970 0.960 0.980 0.980 0.967 0.970
*70 0.015 0.020 0.010 0.010 0.010 0.005 0.010 0.010 0.005 0.008 0.005
*115 0.010 0.000 0.000 0.000 0.005 0.000 0.005 0.000 0.005 0.000 0.000
*150 0.015 0.010 0.025 0.020 0.015 0.025 0.020 0.005 0.000 0.017 0.020
*140 0.005 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.000
*90 0.000 0.000 0.000 0.000 0.000 0.000 0.005 0.005 0.010 0.008 0.005
Me*100 0.950 0.980 0.945 0.975 0.935 0.950 0.948 0.714 0.980 1.000 0.985
*105 0.045 0.020 0.050 0.025 0.065 0.045 0.042 0.281 0.015 0.000 0.005
*95 0.005 0.000 0.005 0.000 0.000 0.005 0.010 0.005 0.005 0.000 0.010
Peplt-1*100 0.990 0.995 0.980 0.970 0.980 0.985 0.965 0.980 0.930 0.933 0.920
*85 0.005 0.005 0.010 0.010 0.010 0.010 0.015 0.015 0.060 0.067 0.065
*120 0.005 0.000 0.010 0.020 0.010 0.005 0.020 0.005 0.010 0.000 0.015
Continued
Popu
lationstru
cture
ofspaw
nin
ggrou
psof
Eu
ropeanan
chovy
1643
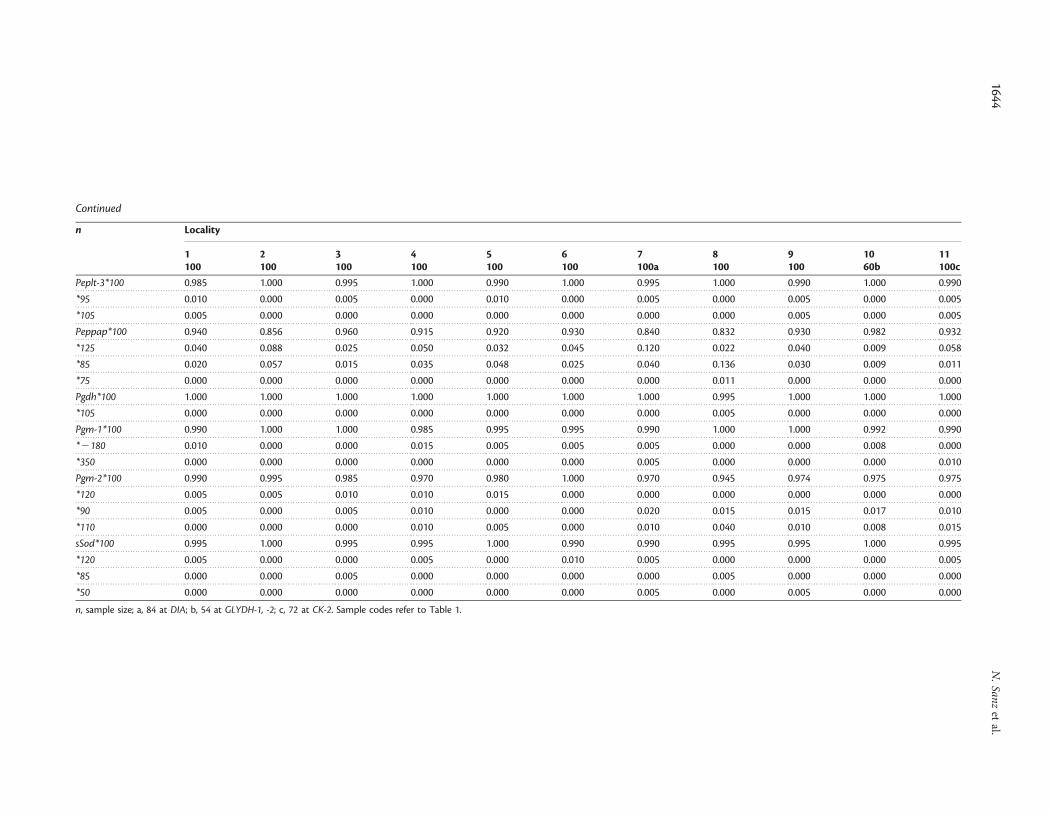
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . .
Continued
n Locality
1 2 3 4 5 6 7 8 9 10 11100 100 100 100 100 100 100a 100 100 60b 100c
Peplt-3*100 0.985 1.000 0.995 1.000 0.990 1.000 0.995 1.000 0.990 1.000 0.990
*95 0.010 0.000 0.005 0.000 0.010 0.000 0.005 0.000 0.005 0.000 0.005
*105 0.005 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.005 0.000 0.005
Peppap*100 0.940 0.856 0.960 0.915 0.920 0.930 0.840 0.832 0.930 0.982 0.932
*125 0.040 0.088 0.025 0.050 0.032 0.045 0.120 0.022 0.040 0.009 0.058
*85 0.020 0.057 0.015 0.035 0.048 0.025 0.040 0.136 0.030 0.009 0.011
*75 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.011 0.000 0.000 0.000
Pgdh*100 1.000 1.000 1.000 1.000 1.000 1.000 1.000 0.995 1.000 1.000 1.000
*105 0.000 0.000 0.000 0.000 0.000 0.000 0.000 0.005 0.000 0.000 0.000
Pgm-1*100 0.990 1.000 1.000 0.985 0.995 0.995 0.990 1.000 1.000 0.992 0.990
*2180 0.010 0.000 0.000 0.015 0.005 0.005 0.005 0.000 0.000 0.008 0.000
*350 0.000 0.000 0.000 0.000 0.000 0.000 0.005 0.000 0.000 0.000 0.010
Pgm-2*100 0.990 0.995 0.985 0.970 0.980 1.000 0.970 0.945 0.974 0.975 0.975
*120 0.005 0.005 0.010 0.010 0.015 0.000 0.000 0.000 0.000 0.000 0.000
*90 0.005 0.000 0.005 0.010 0.000 0.000 0.020 0.015 0.015 0.017 0.010
*110 0.000 0.000 0.000 0.010 0.005 0.000 0.010 0.040 0.010 0.008 0.015
sSod*100 0.995 1.000 0.995 0.995 1.000 0.990 0.990 0.995 0.995 1.000 0.995
*120 0.005 0.000 0.000 0.005 0.000 0.010 0.005 0.000 0.000 0.000 0.005
*85 0.000 0.000 0.005 0.000 0.000 0.000 0.000 0.005 0.000 0.000 0.000
*50 0.000 0.000 0.000 0.000 0.000 0.000 0.005 0.000 0.005 0.000 0.000
n, sample size; a, 84 at DIA; b, 54 at GLYDH-1, -2; c, 72 at CK-2. Sample codes refer to Table 1.
1644N
.San
zet
al.
Copyright © 2022 FDOKUMEN




















