
Impact of coral spawning on the biogeochemistry of a Hawaiian reef

In: Oysters ISBN 978-1-62100-518-6
Editor: Jian G. Qin, pp. 181-202 © 2012 Nova Science Publishers, Inc.
Chapter 7
EXPLORING CORRELATIONS BETWEEN
SPAWNING MEDIATED RESPONSE
AND SUMMER MORTALITY IN PACIFIC
OYSTER CRASSOSTREA GIGAS
Yan Li1, Kirsten Benkendorff
2, Jian G. Qin3,
and Xiaoxu Li4
1School of Agriculture and Food Sciences, The University of Queensland,
Brisbane, Queensland 4072, Australia 2School of Environmental Sciences and Management, Southern Cross
University, Lismore, NSW 2480 Australia 3School of Biological Sciences, Flinders University,
Adelaide, SA, Australia 4South Australian Research and Development Institute (SARDI),
Henley Beach, SA, Australia
ABSTRACT
Pacific oyster, Crassostrea gigas, is one of the most important
species for aquaculture and ecological indicators for environmental
changes. However, mass summer mortality has become a widespread
challenge to Pacific oyster aquaculture industry in recent decades. Due to
Corresponding author Email: [email protected].

Yan Li, Kirsten Benkendorff, Jian G. Qin et al. 182
the energy cost for gametogenesis, the reproductive season is considered
a period of high risk for mortality events. This chapter reviews both
immunity and energy metabolism in Pacific oysters, providing an
appropriate approach to evaluate the integrative health condition in
oysters, with a focus on the spawning-dependent stress response via
comparison between pre- and post-spawning oysters under environmental
stress. This review endeavor reflects the likely consequences of oyster
spawning under food deprivation, heat shock and bacterial challenge and
contributes to our understanding of the underlying mechanisms for oyster
summer mortality. As oysters have already been used as important
bioindicators in the environment and are representatives for intertidal
populations, the implications of spawning impacts on resilience to
environmental stress may be applicable to other compatible species in
similar geographic distributions. By drawing on the broader literature
from other commercial species, particularly clams, mussels, cockles and
abalone, we synthesized the current evidence for spawning related
immuosuppression contributing to epidemic disease and mortality in
molluscs. This review provides insights into the understanding of global
warming effects on ocean productivity in a world-wide aquaculture
species whose reproduction is triggered by temperature increases.
Keywords: oyster spawning, immunosuppression, energy depletion, summer
mortality, global warming
INTRODUCTION
Pacific oyster Crassostrea gigas has a competitive advantage and
dominates other molluscan species in global distribution and aquaculture
production (Food and Agriculture Organisation [FAO] of the United Nations,
2002). At present, Pacific oyster has become one of the most important edible
oysters in many parts of the world as a worldwide aquaculture species. In
recent decades, however, mass summer mortality in cultured Pacific oysters
has become a widespread phenomenon in the world [1-4]. It has been
estimated that up to 50% of the harvestable crop can be lost in a given year,
and these losses can be even higher in some areas [2, 5]. Therefore, the
motivation behind this review is a desire to provide insights into the
underlying cause of summer mortality, based on a synthesis of the current
literature.
As a sessile bivalve living in the intertidal zone, oysters are frequently
subject to various environmental stresses. As proposed in fish [6], Le Roux et

Exploring Correlations… 183
al. [7] suggested that oyster mortality could occur as a result of complex
interactions between the host, environmental factors and disease agents (e.g.
Figure 1). Oysters are permanently exposed to various stressful situations and
required physiological adaptations to survive [8]. Under such stressful
conditions immunosuppression can be induced, which in turn will severely
compromise the capacity of defense against parasites and pathogens [9]. This
may result in high bacterial loads, disease outbreaks or subsequent mortalities
[10, 11]. In such a situation, the interaction between environmental conditions
and disease agents can play an important role to induce oyster mortality.
Figure 1. Interactions between the host, pathogen and the environment provide an
explanation for summer mortality in oyster populations.
The abilities of oysters to cope with pathogens, stress, and their interactive
effects change over seasons [12, 13]. It has been reported that summer
mortality is generally associated with water temperature elevation and
coincides with the period of sexual ripeness in oysters (i.e., pre-spawning and
post-spawning) [1, 14-16]. Despite discrepancies among different oyster
farms, oyster spawning usually occurs simultaneously with increasing water
temperature. Both high temperature and spawning impact molluscan
physiology, resulting in high energetic demands. Therefore, Cheney et al. [2]
and Goulletquer et al. [16] conclude that summer mortality in Pacific oysters
results from the synergy of biotic and abiotic processes that are related to
critical developmental stages in the life history of the species.
This review covers (1) oyster reproductive biology and energy
requirements; (2) the response of oysters to nutritional stress under different
spawning conditions [17]; (3) impacts of heat shock and spawning on oyster
physiology and health [19]; and (4) spawning-dependent immune response to

Yan Li, Kirsten Benkendorff, Jian G. Qin et al. 184
bacterial challenge [21]. Through a systematic review of the impact of
spawning on the physiology and immunology of Pacific oysters Crassostrea
gigas, we demonstrate that the energy expended for spawning compromises
the immune response and physiology of oysters, leaving post-spawning oysters
more susceptible to mortality under environmental stress. These findings have
potential implications for the development of preventive measures to reduce
mortality of post-spawning oysters during the physiologically vulnerable
period.
1. OYSTER REPRODUCTIVE PROCESS
AND ENERGY REQUIREMENTS
Oyster reproductive activities are cyclic and follow distinct seasonal or
annual patterns. In temperate regions, oysters typically follow a seasonal
reproduction cycle with periods of gonad maturation in late spring and
spawning in summer [22]. Spawning typically results in reduction in the
condition index and body weight (Fig 2A) [13]. For this reason, there has been
significant research towards the development of non-spawning triploid oysters
to maximize productivity in oyster aquaculture [23].
The gametogenic stage is classically categorised according to the
reproductive scale reported by Lubet [24]: Stage 0 is the undifferentiated
gonads, characterised by empty gonads (sexual repose). There is no presence
of follicles peripherally to the digestive gland, making sex determination
difficult. Stage I is the gonad in the early-developmental stage, known by the
decrease of the interfollicular connective tissue after gonia multiplication and
expansion of the follicles, and contains oogonia or spermatogonia and primary
oocytes or spermatocytes. Stage II is the gonad at a late-developmental stage
when the oocyte and spermatozoa are observed and secondary oocytes and
spermatocytes predominate in the follicles. Stage III A shows intense
gametogenic activity, characterized by the absence of the interfollicular
connective tissue. Stage III B is advanced with further gonad maturation,
during which the male oyster gonads are present, the homogenous stage after
the disappearance of spermatids running toward the centre of the follicle.
Stage III C is shown in spent gonads, characterised by the presence of ruptured
follicles and residual gametes in the oyster gonads. Then a reproductively
quiescent or subsequent “rest” period is followed, i.e., the post-spawning
period [25]. In this review, we have simplified the stages of gametogenesis
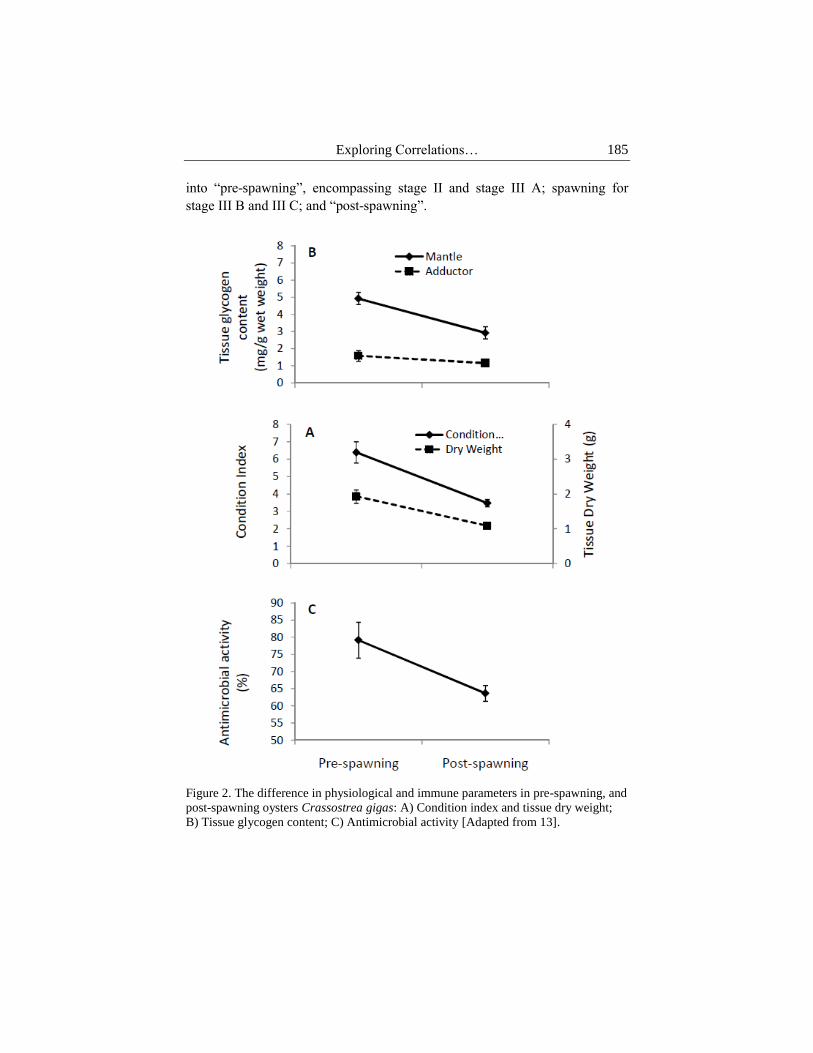
Exploring Correlations… 185
into “pre-spawning”, encompassing stage II and stage III A; spawning for
stage III B and III C; and “post-spawning”.
Figure 2. The difference in physiological and immune parameters in pre-spawning, and
post-spawning oysters Crassostrea gigas: A) Condition index and tissue dry weight;
B) Tissue glycogen content; C) Antimicrobial activity [Adapted from 13].

Yan Li, Kirsten Benkendorff, Jian G. Qin et al. 186
As gonad development is a high energy-demanding process, it not only
requires mobilization of exogenous energy from ingested food, but also
requires endogenous energy reserves from body tissues [26-28], especially
glycogen from mantle tissues (Figure 2). In oysters, the reproductive effort and
costs are similar in the male and female, with 27% of oxygen consumption for
reproductive processes [29]. Being the basic bio-energy fuel, carbohydrates
are stored mainly as glycogen in bivalves [19, 30]. It has been demonstrated
that glycogen plays a central role in energetic and metabolic supply during
gametogenesis [1, 22, 27, 30]. However, the capacity of glycogen storage
reduces sharply and reaches its lowest after spawning [1, 13, 31]. Meanwhile,
glycogen phosphorylase and synthase in oysters are also suppressed by
spawning [32]. As glycogen reserves supply additional energy to mobilise
other energetic reserves [33, 34], low glycogen is often correlated with oyster
mortality events [1, 31, 35, 36].
The low energy reserves available after spawning could result in indirect
effects on oyster immunity. Energy is required for routine immune function
but under stress, all available energy will be diverted away from the immune
system to the maintenance of metabolic activities. This relationship between
the physiological status and immune activity can be observed in spawning
oysters. Concomitant with the drop in the condition index (Fig 2A) and
glycogen content (Fig 2B), there is also a drop in the antibacterial activity (Fig
2C). This combined impact on the metabolic reserves and immune activity
leads oysters to a vulnerable condition, subject to potential infection after
spawning. In a laboratory study on the recovery of oyster physiology and
immunity, we have demonstrated that the first 8 days after spawning is a
critically vulnerable period for oysters [37].
2. SPAWNING-DEPENDENT RESPONSE
TO NUTRITIONAL STRESS
Oysters are likely to encounter low food availability in the ocean due to
heterogeneous distribution of seston particles at both spatial and temporal
scales. Berg and Newell [38] reported that chlorophyll a fluctuated eight fold
from summer to autumn in the same location within a year. Even within a tidal
cycle, both quantity and quality of seston can vary as much as across seasons
[39]. Particularly in nutrient poor areas (i.e., with low terrestrial nutrient input
or upwelling), oysters are frequently subjected to low food supply [40]. Most
Exploring Correlations… 187
nutrition studies on oysters have been focused on the effects of dietary
deficiency on growth [eg., 41, 42, 43], but there has been little consideration of
summer morality as a consequence of starvation concomitant with
reproduction. However, starved spawning oysters provide an ideal model to
study spawning-dependent response to metabolic stress.
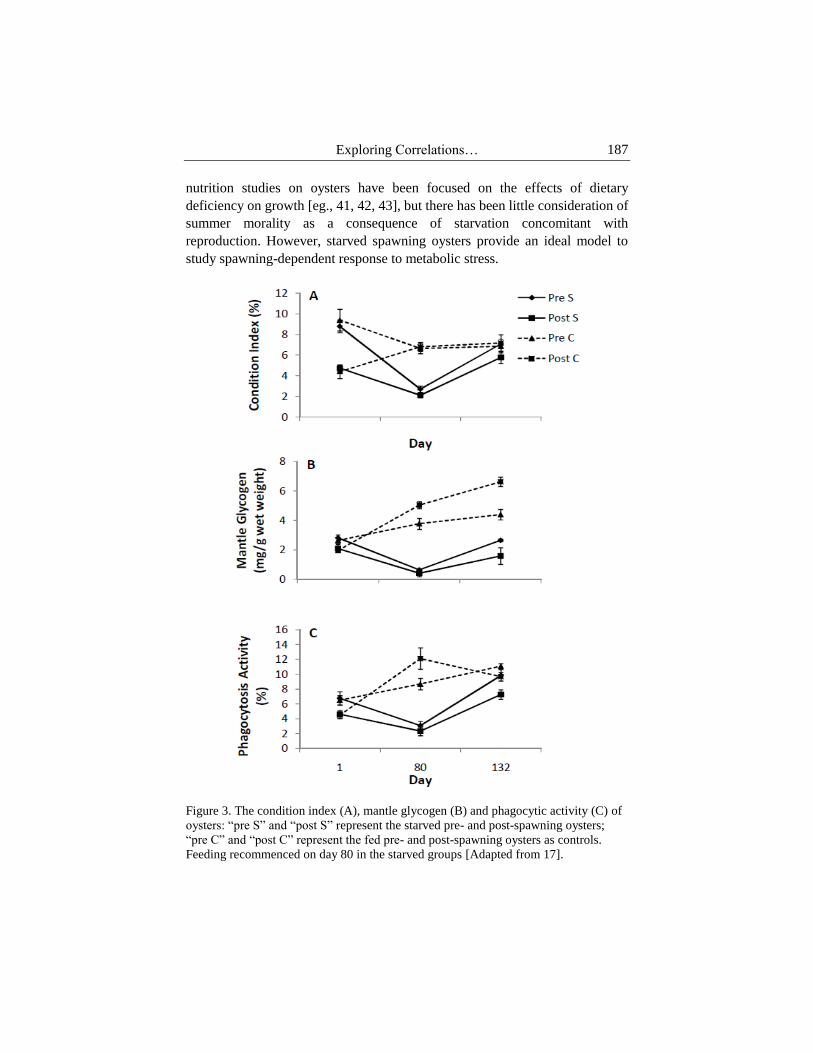
Figure 3. The condition index (A), mantle glycogen (B) and phagocytic activity (C) of
oysters: “pre S” and “post S” represent the starved pre- and post-spawning oysters;
“pre C” and “post C” represent the fed pre- and post-spawning oysters as controls.
Feeding recommenced on day 80 in the starved groups [Adapted from 17].

Yan Li, Kirsten Benkendorff, Jian G. Qin et al. 188
We designed a manipulative experiment where pre- and post-spawning
oysters were deprived of food for 80 days and then refed for 52 days [17]. The
results demonstrated that starvation and spawning had a rapid and dramatic
effect on oyster condition (Figure 3A). During 80-days of food deprivation,
the mantle tissue thinned out and the body became slightly translucent in post-
spawning oysters. Due to a lack of available gonad mass for absorption and the
thinned mantle tissue, post-spawning oysters need to catabolise energy stored
in other tissues different from pre-spawning oysters (Fig 3B shows these
effects on energy storage). Despite reaching a similar low level of condition
index and glycogen after 80-day food deprivation, the recovery process in pre-
spawning oysters was much quicker than that of post-spawning oyster upon
refeeding (Figure 3), suggesting that pre-spawning oysters are more resilient to
food deprivation than post-spawning individuals.
Under food deprivation, the mantle glycogen of post-spawning oysters is
lower than pre-spawning oysters and is further depleted with reduction of
adductor glycogen during food deprivation [17]. This implies that post-
spawning oysters have developed adaptations to use adductor glycogen as a
source of spare energy to maintain their metabolism and to cope with food
deprivation. Mantle and gill proteins in pre- and post-spawning oysters are
also reduced and maintained at a low level upon food deprivation [43]. Pre-
spawning oysters exhibited strong metabolic resilience to food deprivation as
indicated by faster glycogen (Figure 3B) and protein recovery in starved pre-
spawning oysters than in post-spawning oysters upon refeeding. After 80-day
food deprivation, the slower energy restoration in post-spawning oysters
shows that spawning can suppress the metabolic response to feeding after
starvation in oysters. Spawning activity retards energy recovery after having
experienced a period of food shortage.
Although the synergetic effect of food deprivation and spawning does not
directly cause mortality, concurrent energetic and immunological dysfunctions
may possibly lead to mass mortality in post-spawning oysters [17]. Results on
haemocyte phagocytosis and hemolymph antimicrobial activity demonstrate
that post-spawning oysters have low immune resilience under food deprivation
[17]. As reported by Delaporte et al. [44], there would be less energetic supply
for haemocyte phagocytosis due to the cost of spawning. Similarly, the low
energy intake during food deprivation can decrease haemocyte phagocytosis,
as found in our study and others [45, 46]. When feeding was resumed,
phagocytosis in pre-spawning oysters increased much quicker than that in
post-spawning oysters (Figure 3C). Similarly, food resumption enhanced the
antimicrobial activity, but pre-spawning oysters showed a faster recovery than

Exploring Correlations… 189
post-spawning oysters. In the natural environment, therefore, starved post-
spawning oysters are likely to be more sensitive to potentially lethal stressors,
e.g., bacterial challenge and heat shock. In a worst case scenario, the
synergistic effect of spawning and poor nutritional condition could lead to
mass mortality, especially in summer under high temperature stress when
pathogens prevail.
3. IMPACTS OF HEAT SHOCK AND SPAWNING ON OYSTER
PHYSIOLOGICAL AND IMMUNE RESPONSES
Among various environmental conditions, temperature has long been
recognised as a key factor that influences all physiological processes in oysters
[47]. With elevated temperatures, animals need to increase metabolism to
acquire an adequate energetic supply for survival [48]. Pacific oysters can
adapt to a wide range of temperatures from 3 to 35ºC [49]. Furthermore,
oysters can exhibit adaptive plasticity to heat shock, known as "induced
thermotolerance" [8]. Indeed, thermotolerance has been demonstrated in the
laboratory, with exposure to a single sublethal heat shock (e.g., 37ºC) enabling
oysters to transiently tolerate extreme, otherwise lethal temperatures (43ºC) for
an hour [19]. This phenotypic plasticity of the heat-shock response is
associated with the expression of heat-shock proteins in oysters [50, 51].
In Pacific oysters, Hsp70 is the primary family of Hsp that is responsive to
thermal stress [50]. The thermal limits of oysters are correlated with changes
in isoform expression in the Hsp70 family, which can be detected by western
blotting techniques. Thermal adaptation in oysters is known to be dependent
on hydrographic and biological factors, including ambient temperature and
reproductive stress [19, 50]. As the cost of protein synthesis is estimated to be
18~26% of the energy budget of ectothermal organisms [52], the metabolic
adaptation in spawning oysters is also subject to a deficiency in energy supply
[19]. In order to examine the underlying causes leading to this phenomenon,
thermotolerance of the Pacific oyster was assessed using pre- and post-
spawning oysters that were sequentially treated with sublethal (37ºC) and
lethal heat shocks (44ºC) [19].
In comparison, although both pre and post-spawning oysters were
recorded to have some adaptability of induced thermotolerance via the
expression of Hsp69, the ability to resist sublethal and lethal temperature
shock was reduced in post-spawning oysters. A preventative 37ºC exposure

Yan Li, Kirsten Benkendorff, Jian G. Qin et al. 190
significantly reduced oyster mortality after exposure to a second heat shock of
44ºC, but in post-spawning oysters, mortality remained at 80%, compared to
less than 10% in pre-spawning oysters [19]. As suggested by Kregel [53]
morbidity and mortality from thermal shock is due to the dysfunction of some
critical target tissues that are heat sensitive and vital to the animal. We found
that both Hsp72 and Hsp69 had heat shock-dependent stimulation in the gills
of pre-spawning oysters, but not in post-spawning oysters indicating that
spawning can reduce heat shock protein synthesis [19]. Impairment of heat
shock protein synthesis in the gill and the consequent protein denaturation is
likely to lead to hyperthermic killing. Ultimately, dysfunction in the gill, the
most important respiratory organ of marine animals, will cause a mismatch
between oxygen delivery and the ability to respond to heat shock, which
finally leads to the collapse of physiological function [54, 55].
Besides depletion of glycogen in post-spawning oysters, spawning and
heat shock were concomitant with decreasing adenylate energy charge (AEC),
indicative of lower cellular energy for metabolic activity. After heat shock, the
AEC value in pre-spawning oysters was maintained within the range of 0.5 -
0.7 (Table 1), which is the sub-optimal stress range according to Cattani et al.
[57]. This indicates that pre-spawning oysters can maintain normal energy
metabolism at a cellular level irrespective of induced thermotolerance.
Table 1. Effects of heat shock and spawning on adenylate energy
charge in oyster mantle tissue (Mean ± SE). “Pre HS” and “Post HS”
represent the heat shocked pre- and post-spawning oysters; “
Pre Con” and “Post Con” represent non-heat treated
pre- and post-spawning oysters [Source from 19]
Day Pre HS Post HS Pre Con Post Con
0 Oysters were heated at 37ºC for 1 hour (sublethal heat shock)
0.64 ± 0.01 0.55 ± 0.01 0.71 ± 0.04 0.61 ± 0.01
1 0.69 ± 0.02 0.54 ± 0.01 0.76 ± 0.07 0.63 ± 0.03
2 0.64 ± 0.03 0.58 ± 0.01 0.70 ± 0.04 0.63 ± 0.01
3 0.65 ± 0.02 0.54 ± 0.03 0.70 ± 0.02 0.63 ± 0.01
5 Oysters were reheated at 44ºC for 1 hour(lethal heat shock)
6 0.59 ± 0.01 0.39 ± 0.02 0.76 ± 0.06 0.66 ± 0.02
10 0.63 ± 0.04 0.32 ± 0.02 0.71 ± 0.01 0.67 ± 0.02
* The bold italic numbers are less than 0.5 which is the critical threshold for oyster
AEC values based on Shofer and Tjeerdema [56].

Exploring Correlations… 191
However, post-spawning oysters had much lower AEC levels, especially
after lethal heat shock (Table 1). The low AEC values and high mortality
recorded in post-spawning oysters subjected to lethal heat shock are consistent
with a study by Shofer and Tjeerdema [56], where AEC values between 0.3 -
0.5 were suggested to be within a critical range from which the capacity to
recover from stress is impossible. This critical limit may be attributed to a
transition to anaerobic metabolism when temperature is beyond
thermotolerance, which leads to insufficient cellular energy supply [54]. In this
stressful situation, the physiological trade-offs will divert energy reserves
towards essential maintenance [47], although the residual energy reserves
(glycogen) seem not to be adequate for energy modulation in post-spawning
oysters.
A cumulative effect of spawning and heat shock was observed on the
immunocompetence of oysters, demonstrated by reduced haemocyte
phagocytosis and hemolymph antimicrobial activity [19]. These results
support the hypothesis that the energy expended during reproduction
compromises the thermotolerance and immune status of oysters, leaving them
easily subject to mortality if heat stress occurs in the post-spawning stage. This
reflects the likely consequences of spawning under thermal stress, thus
contributing our understanding of oyster summer mortality. Consequently,
these results have implications for the long-term persistence of molluscs under
the influence of global warming [19].
4. SPAWNING-DEPENDENT IMMUNE RESPONSE
TO BACTERIAL CHALLENGE
Although oysters have developed physiological adaptation for survival
[8], stress can suppress their immunocompetence and substantially
compromise the ability to defend against parasites and pathogens [9, 19].
There is some evidence that reduction of defense efficiency can lead to high
bacterial loads, disease outbreaks and mass mortality in oysters [10, 11]. We
have already demonstrated that oyster immunity is impacted by the
physiological stress of spawning, as well as environmental stressors such as
low nutrient availability and high temperature. However, infection by
pathogens can be regarded as another stressor. Typically an animal will
respond to infection by mounting an immune response. Therefore, the question
remains as to whether spawned oysters can respond effectively to bacterial

Yan Li, Kirsten Benkendorff, Jian G. Qin et al. 192
challenge or if their immune systems are too compromised from the low
energy reserves.
To investigate this issue further, we developed two models to simulate a
bacterial challenge [47]. The first model involved the injection of extracellular
products from Vibrio harveyi into the oysters to establish whether they were
more susceptible to virulence factors after spawning. The second model
involved the injection of heat killed Vibrio harveyi into the oysters to examine
the sublethal immune responses to a challenge. The injection of heat-killed
bacteria caused less than 6% mortality in both pre- and post- spawning oysters.
However, the injection of bacterial extracellular products resulted in ~ 30%
mortality in pre-spawned oysters. Mortality was significantly high at ~50% in
post-spawning oysters, indicating that they are less able to resist the toxic
factors secreted by bacterial pathogens.
In the sublethal bacterial challenge model, no significant differences were
observed between pre- and post-spawning oysters in the total haemocyte
abundance and the phagocytic rate, suggesting that spawning does not have a
direct effect on cell-mediated immunity [21]. Nevertheless, we did observe
lower phagocytosis in post-spawning oysters than pre-spawning oysters in the
control groups throughout the study period (Figure 4A). Despite seasonal
variation, a reduction of phagocytosis has also been observed during spawning
in field populations [58]. However, the effect of spawning might be over-
ridden by the impact of simulated bacterial challenge, as all challenged oysters
had double the phagocytic rate of the controls, irrespective of spawning status.
Consequently, post-spawning oysters appear to be capable of eliciting a
similar cellular immune response as pre-spawning oysters, to cope with
invasion by foreign particles. This finding is coincident with previous
observations that haemocyte phagocytosis is similar between summer
mortality-susceptible oysters and summer mortality-resistant oysters [59].
Therefore, Lambert et al. [60] concluded that relevant indicators other than
haemocyte profiles are required to help explain oyster survival during summer
mortality events.
Unlike the cellular immune response, the humoral immune function was
not only influenced by simulated bacterial challenge, but was also suppressed
by spawning (Figure 4B). Antimicrobial agents are synthesized by the
haemocytes [61] and stored in the cells as reserves, then released only after
stimulation by microbial infection or other similar challenges [10, 62].
Consequently, the nonviable bacterial injection was expected to increase the
antimicrobial activity in comparison to seawater injection, by stimulating the
release of active factors. However, the opposite effect was observed, whereby
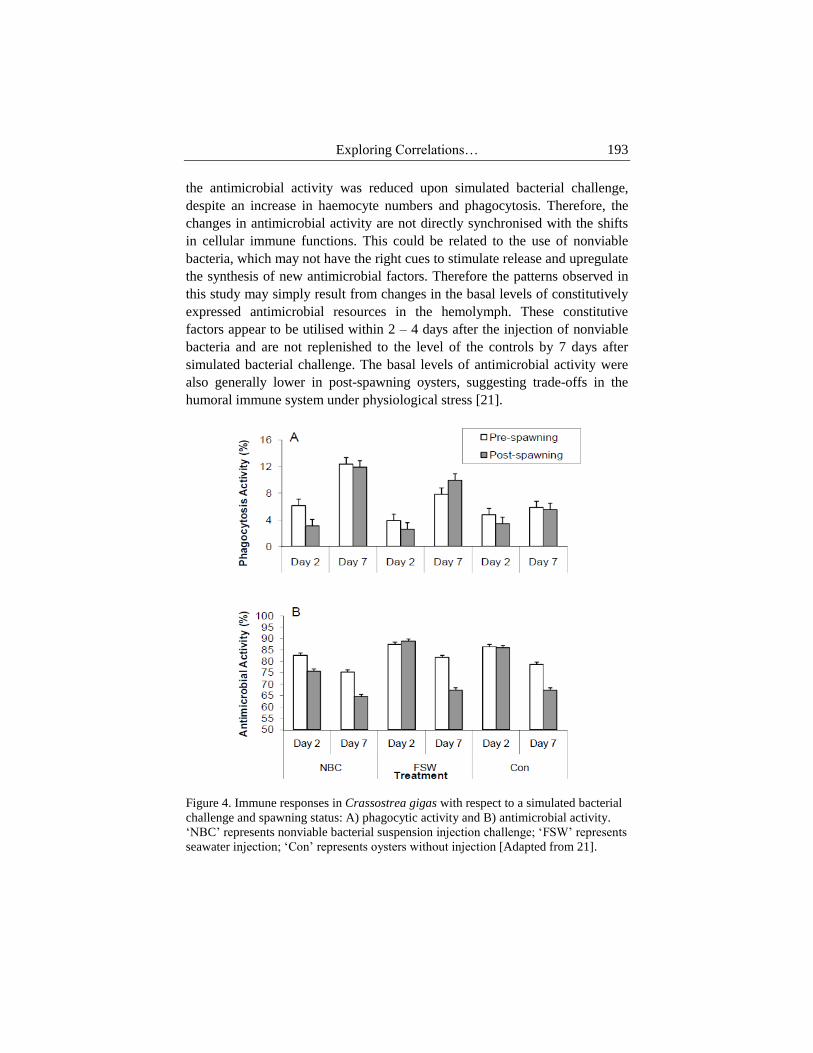
Exploring Correlations… 193
the antimicrobial activity was reduced upon simulated bacterial challenge,
despite an increase in haemocyte numbers and phagocytosis. Therefore, the
changes in antimicrobial activity are not directly synchronised with the shifts
in cellular immune functions. This could be related to the use of nonviable
bacteria, which may not have the right cues to stimulate release and upregulate
the synthesis of new antimicrobial factors. Therefore the patterns observed in
this study may simply result from changes in the basal levels of constitutively
expressed antimicrobial resources in the hemolymph. These constitutive
factors appear to be utilised within 2 – 4 days after the injection of nonviable
bacteria and are not replenished to the level of the controls by 7 days after
simulated bacterial challenge. The basal levels of antimicrobial activity were
also generally lower in post-spawning oysters, suggesting trade-offs in the
humoral immune system under physiological stress [21].
Figure 4. Immune responses in Crassostrea gigas with respect to a simulated bacterial
challenge and spawning status: A) phagocytic activity and B) antimicrobial activity.
‘NBC’ represents nonviable bacterial suspension injection challenge; ‘FSW’ represents
seawater injection; ‘Con’ represents oysters without injection [Adapted from 21].

Yan Li, Kirsten Benkendorff, Jian G. Qin et al. 194
Concomitant with the decrease in antimicrobial activity, an initial
reduction in hemolymph protein was found upon simulated bacterial challenge
[21], suggesting the participation of hemolymph protein in antimicrobial
activity. In post-spawning oysters, the hemolymph protein declined over time
in controls and although it increased slightly after injection, this occurred at a
much slower rate than observed in pre-spawning oysters. As a range of plasma
antibacterial peptides and bacteriolytic enzymes are involved in bivalve
immune defense [62-64], it is likely that these proteinaceous substances were
rapidly mobilized in response to simulated wounding by injection in both of
pre- and post-spawning oysters. However, the ability to recover humoral
immune function after simulated bacterial challenge is clearly impacted by
spawning (Figure 4B). This implies that after exhausting the basal levels of
humoral antimicrobial proteins, post-spawning oysters have insufficient
metabolic resources to initiate the expression of new proteins and peptides to
continue fighting infection. Therefore, the humoral immune function may not
be maintained effectively in post-spawning oysters exposed to prolonged or
repeated infections. Coupled with the degradation of host proteins by bacterial
extracellular products, this may weaken the oysters to a point of no recovery.
Therefore, despite the apparently normal cellular immune function in post-
spawning oysters, the spawning-suppressed humoral immune function could
be fatal to oysters when facing opportunistic pathogens in the environment,
particularly in warm environments where the growth rate of Vibrio spp. is high
[65].
5. POTENTIAL IMPLICATIONS
The published literature and anecdotal reports suggest that many of the
mortalities in Pacific oysters are the result of multiple factors or stressors [2, 3,
66]. Although there was no massive mortality in summer during our field
study, the coupled relationship of low antibacterial activity and energy
reserves is a strong indication of the fragile condition of spawned oysters [13].
In the present study, the synergistic impacts of environmental stress and
spawning on physiology and immunocompetence provide one plausible
explanation for the phenomenon of summer mortality in the Pacific oyster.
This research review reflects the likely consequences for post-spawning
oysters under food deprivation [17], thermal stress [19] and bacterial challenge
[21], thus contributing to our understanding of the underlying biological
reasons for mass oyster mortality in the post-spawning period. Meanwhile, this

Exploring Correlations… 195
review greatly improves our knowledge of the sublethal changes that occur in
the immunological and physiological responses of oysters under stress, as well
as the natural temporal variation. Results from the references cited provide
good baseline information for future monitoring of oyster health.
Since the mid-1970s, large-scale episodic events, such as disease
epidemics, mass mortalities and biological population explosions have been
occurring in marine environments with increasing frequency, intensity and
range [67]. Notable examples are the polar-ward expansion of oyster diseases
and concerns of escalating summer mortality in European and Australian
bivalve aquaculture. Oscillations in the earth’s climate also lead to associated
fluctuations in the temperature regimes of many marine and terrestrial
ecosystems. Consequences in marine ecosystems include changes in the timing
of reproduction, as well as changes in reproductive success, recruitment,
growth performance and mortality of species and finally, changes in their
geographical distribution [54]. These factors will be the major threat to
aquaculture and the economic and ecological sustainability of marine fisheries
in the near future.
The Pacific oyster offers a good opportunity to assess the impacts of
climate change on ocean productivity, as an important world-wide aquaculture
species whose reproduction is triggered by temperature increases. Meanwhile,
it is possible that the incidents of mass mortality in marine organisms,
including species such as abalone, mussels and clams that also have great
economic value, will increase globally, coincident with unpredictable
environmental patterns [68]. For Pacific oyster aquaculture, although the post-
spawning period falls into the out-of-season seafood marketing, husbandry
management during this period is critical to reduce mortality. To prevent
mortality at this vulnerable period, it is recommended that farmers try to
reduce or avoid any anthropogenic disturbance to spawned animals. In
intertidal farming areas, submerging spawned oysters into deeper seawater
could reduce stress from exposure to high air temperature during low tide.
Removing any seen morbid and moribund individuals will help reduce fouling
of the water and stop the spread of infection. The impact of physio-chemical
aspects of seawater in culture areas, as well as culture practices should also be
taken into account. Food supplementation for spawned populations should be
considered if natural food availability is low during the recovery period (eg.,
cultivation area shift), particularly in lean farming areas.
As oysters are already used as important bioindicators in the environment
and can be representative for intertidal populations [69], the implications of
spawning impact on resilience to environmental stress should be compatible to

Yan Li, Kirsten Benkendorff, Jian G. Qin et al. 196
other species in similar geographic distributions with a similar reproduction
pattern. This review highlights the sensitivity of spawned molluscs to
environmental stressors and therefore provides vital information for the
development of future adaptation strategies in a changing global environment.
An improved understanding of the interactions between the host, pathogen and
environment will facilitate long-term economic and environmental
sustainability in marine biological resources.
REFERENCES
[1] Perdue J, Beattie JH, Chew KK: Some relationships between
gametogenentic cycle and summer mortality phenomenon in the Pacific
oyster Crassostrea gigas in the Washington state. J. Shellfish Res. 1981,
1:9-16.
[2] Cheney DP, Macdonald EF, Elston RA: Summer mortality of Pacific
Oysters, Crassostrea gigas (Thunberg): initial findings on multiple
environmental stressors in Puget Sound, Washington, 1998. J. Shell Res.
2000, 19(1):353-359.
[3] Huvert A, Herpin A, Dégremont L, Labreuche Y, Samain J-F,
Cunningham C: The identification of genes from the oyster Crassostrea
gigas that are differentially expressed in progeny exhibiting opposed
susceptibility to summer mortality. Gene 2004, 343:211-220.
[4] Garnier M, Labreuche Y, Garcia C, Robert M, Nicolas J-L: Evidence for
the involvement of pathogenic bacteria in summer mortalities of the
Pacific oyster Crassostrea gigas. Microbial Ecol. 2007, 53:187-196.
[5] Renault T, Cochennec N: Rickettsia-like organisms in the cytoplasm of
gill epithelial cells of the Pacific oyster Crassostrea gigas. J. Invertebr.
Pathol. 1994, 64(2):160-162.
[6] Snieszko SF: The effects of environmental stress on outbreaks of
infectious diseases of fishes. J. Fish Biol. 1974, 6:197-208.
[7] Le Roux F, Gay M, Lambert C, Waechter M, Poubalanne S, Chollet B,
Nicolas J-L, Berthe F: Comparative analysis of Vibrio splendidus-related
strains isolated during Crassostrea gigas mortality events. Aquat. Living
Resour. 2002, 15(4):251-258.
[8] Colombo L, Pickering AD, Belvedere P, Schreck CB: Stress inducing
factors and stress reaction in aquaculture. In: Aquaculture Europe'89–
Business Joins Science: 1990; Bredene, Belgium; 1990: 93-121.

Exploring Correlations… 197
[9] Harvell CD, Kim K, Burkholder JM, Colwell RR, Epstein PR, Grimes
DJ: Emerging marine diseases-climate links and anthropogenic factors.
Science 1999, 285:1505-1510.
[10] Cheng TC: Cellular defense mechanisms in oyster. In: Recent Advances
in Marine Biotechnology, Immunology and Pathology: 2000; Enfied
(NH), USA: Science Publishers, Inc; 2000: 43-83.
[11] Chu F-L: Defense mechanisms of marine bivalves. In: Recent Advances
in Marine Biotechnology: Immunology and Pathology: 2000; Enfied,
NH, USA: Science Publishers, Inc.; 2000: 1-42.
[12] Fisher WS, Oliver LM, Winstead JT, Long ER: A survey of oysters
Crassostrea virginica from Tampa Bay, Florida: associations of internal
defense measurements with contaminant burdens. Aquat. Toxicol. 2000,
51(1):115-138.
[13] Li Y, Qin JG, Li X, Benkendorff K: Monthly variation of condition
index, energy reserves and antibacterial activity in Pacific oysters,
Crassostrea gigas, in Stansbury (South Australia) Aquaculture 2009,
286:64-71.
[14] Lipoveshy V, Chew KK: Mortality of Pacific oyster Crassostrea gigas:
the influence of temperature and enriched seawater on oyster survival.
Proc. Nat. Shellfish Assoc. 1972, 62:72-82.
[15] Beattie JH, Chew KK, Hershberger WK: Differential survival of
selected strains of Pacific oysters Crassostrea gigas during summer
mortality. Proc. Nat. Shellfish Assoc. 1980, 70:184-189.
[16] Goulletquer P, Soletchnick P, Le Moine O, Razet D, Gaeairon P, Faury
N, Tailade S: Summer mortality of the Pacific cupped yster Crassostrea
gigas in the Bay of Marennes-Oleron (France). In: Mariculture
Committee CM: 1998; 1998: 1-20.
[17] Li Y, Qin JG, Li X, Benkendorff K: Spawning-dependent stress
response to food deprivation in Pacific oyster Crassostrea gigas.
Aquaculture 2009, 286:309-317.
[18] Malacological Society of London. Centenary Symposium (1993 :
Natural History Museum), Taylor JD: Origin and evolutionary radiation
of the Mollusca. Oxford ; New York: Oxford University Press; 1996.
[19] Li Y, Qin JG, Abbot CA, Li X, Benkendorff K: Synergistic impacts of
heat shock and spawning on the physiology and immune health of
Crassostrea gigas: An explanation for summer mortality in Pacific
oysters. Am. J. Physiol.-Reg. I 2007, 293:2353-2362.

Yan Li, Kirsten Benkendorff, Jian G. Qin et al. 198
[20] Biochemical Society (Great Britain). Symposium (64th : 1997 :
University of Liverpool), Downes CP, Wolf CR, Lane DP: Cellular
responses to stress. Princeton, N.J.: Princeton University Press; 1999.
[21] Li Y, Qin JG, Li X, Benkendorff K: Spawning-dependent stress
responses in Pacific oysters Crassostrea gigas: a simulated bacterial
challenge in oysters. Aquaculture 2009, 293:164-171.
[22] Mathieu M, Lubet P: Storage tissue metabolism and reproduction in
marine bivalves-a brief review. Invertebr. Reprod. Develop. 1993,
23:123-129.
[23] Li Y, Li X, Qin JG: Triploidy induction in Australian greenlip abalone
Haliotis laevigata (Donovan) with cytochalasin B. Aquaculture Res.
2007, 38:487-492.
[24] Lubet P: Recherches sur le cycle sexuel et l'emission des gamètes chez
les mytilidés et les pectinidés (mollusques bivalves). Rev. Trav. Inst.
pêches Marit. 1959, 23:387-542.
[25] Giese AC, Pearse V, Pearse JS: Reproduction of marine invertebrates,
vol. 1. New York, USA: Academic Press; 1974.
[26] Gabbott PA: Developmental and seasonal metabolic activities in marine
molluscs. In: The Mollusca: Environmental Biochemistry and
Physiology: 1983; Academic Press, New York.; 1983: 165-217.
[27] Berthelin C, Kellner K, Mathieu M: Histological characterisation and
glucose incorporation into glycogen of the Pacific oyster Crassostrea
gigas storage cells. Mar. Biotechnol. 2000, 2:136-145.
[28] Lodeiros C, Rengel JJ, Guderley HE, Nusetti O, JHimmelman JH:
Biochemical composition and energy allocation in the tropical scallop
Lyropecten (Nodipecten) nodosus during the months leading up to and
following the development of gonads. Aquaculture 2001, 199:63-72.
[29] Honkoop PJC, Bayne BL, Drent J: Flexibility of size of gills and palps
in the Sydney rock oyster Saccostrea glomerata (Gould, 1850) and the
Pacific oyster Crassostrea gigas (Thunberg, 1793). J. Exp. Mar. Biol.
Ecol. 2003, 282(1-2):113-133.
[30] Li Q, Osada M, Mori K: Seasonal biochemical variations in Pacific
oyster gonadal tissue during sexual maturation. Fisheries Sci. 2000,
66:502-508.
[31] Mori K, Tamate HK, Imai TK, Itikawa O: Studies on the mass mortality
of the oyster in Matsushima Bay. V. Changes in the metabolism of lipids
and glycogen of the oyster during the stages of sexual maturation and
spawning. Bull. Tohoko Reg. Fish Res. Lab. 1965, 25:66-68.

Exploring Correlations… 199
[32] Bacca H, Huvet A, Fabioux C, Daniel J-Y, Delaporte M, Pouvreau S,
Van Wormhoudt A, Moal J: Molecular cloning and seasonal expression
of oyster glycogen phosphorylase and glycogen synthase genes. Comp.
Biochem. Physiol. B 2005, 140:635-646.
[33] Ruiz C, Abad M, Sedano F, Garcia-Martin LO, Sanchez-Lopez JL:
Influence of seasonal environmental changes on the gametes production
and biochemical composition of Crassostrea gigas (T.) in suspended
culture in E1 Grove, Galicia, Spain. J. Exp. Mar. Biol. Ecol. 1992,
155:249-262.
[34] Takuji O, Toshiko N, Ikuo H, Yoshinori S: Effects of starvation on
RNA: DNA ratio, glycogen content, and C: N ratio columellar muscle of
the Japanese turban shell Turbo (Botilus) cornutus (Gastropoda).
Fisheries Sci. 2002, 68:306-312.
[35] Berthelin C, Kellner K, Mathieu M: Storage metabolism in the Pacific
oyster (Crassostrea gigas) in relation to summer mortalities and
reproductive cycle (West Coast of France). Comp. Biochem. Physiol. B
2000, 125(3):359-369.
[36] Patrick S, Faury N, Goulletquer P: Seasonal changes in carbohydrate
metabolism and its relationship with summer mortality of Pacific oyster
Crassostrea gigas (Thunberg) in Marennes-Oleron bay (France).
Aquaculture 2006, 252(2-4):328-338.
[37] Li Y, Qin J, Li X, Benkendorff K: Assessment of metabolic and immune
changes in post-spawning Pacific oyster Crassostrea gigas:
identification of a critical period of vulnerability after spawning.
Aquaculture Res. 2010, 41:e155-e165 (DOI: 110.1111/j.1365-
2109.2010.02489.x).
[38] Berg JA, Newell RIE: Temporal and spatial variation in the composition
of seston available to the suspension feeder Crassostrea virginica.
Estuar. Coast Shelf S 1986, 23:375-386.
[39] Peterson CH, Black R: Resource depletion by active suspension feeders
on tidal flats: influence of local density and tidal elevation. Limnol.
Oceanogr. 1987, 32:142-166.
[40] Zhang C, Xie L, Huang J, Chen L, Zhang R: A novel putative tyrosinase
involved in periostracum formation from the pearl oyster (Pinctada
fucata). Biochem. Biophys. Res. Commun. 2006, 342(2):632-639.
[41] Whyte JNC, Englar JR, Carswell BL: Biochemical composition and
energy reserves in Crassostrea gigas exposed to different levels of
nutrition. Aquaculture 1990, 90:157-172.

Yan Li, Kirsten Benkendorff, Jian G. Qin et al. 200
[42] Garcia-Esquivel Z, Bricelj VM, Felbeck H: Metabolic depression and
whole-body response to enforced starvation by Crassostrea gigas
postlarvae. Comp. Biochem. Physiol. A 2002, 133(1):63-77.
[43] Butt D, Aladaileh S, O' Connor WA, Raftos D: Effect of starvation on
biological factors related to immunological defence in the Sydney rock
oysters (Saccostrea glomerata). Aquaculture 2007, 264:82-91.
[44] Delaporte M, Soudant P, Lambert C, Moal J, Pouvreau S, Samain J-F:
Impact of food availability on energy storage and defense related
hemocyte parameters of the Pacific oyster Crassostrea gigas during an
experimental reproductive cycle. Aquaculture 2006, 254(1-4):571-582.
[45] Funakoshi S: Studies on the classification, structure and function of
hemocytes in bivalves. In: Bulletin of National Research Institute of
Aquaculture. National Research Institute of Aquaculture (NRIA) and
Fisheries Agency, Japan; 2000: 1-103.
[46] Hégaret H, Wikfors GH, Soudant P, Delaporte M, Alix JH, Smith BC,
Dixon MS, Quere C, Le Coz JR, Paillard C: Immunological competence
of eastern oysters, Crassostrea virginica, fed different microalgal diets
and challenged with a temperature elevation. Aquaculture 2004, 234(1-
4):541-560.
[47] Lanning G, Flores JF, Sokolova IM: Temperature-dependent stress
response in oysters, Crassostrea virginica: Pollution reduces
temperature tolerance in oysters. Aquat. Toxicol. 2006, 79:278-287.
[48] Mora C, Maya MF: Effect of the rate of temperature increase of the
dynamic method on the heat tolerance of fishes. J. Therm. Biol. 2006,
31:337-341.
[49] Shaktin G, Shumway SE, Hawes R: Considerations regarding the
possible introduction of the Pacific oyster (Crassostrea gigas) to the
Gulf of Marine: a review of global experience. J. Shellfish Res. 1997,
16:463-477.
[50] Clegg J, Uhlinger K, Jackson S, Cherr G, Rifkin E, Friedman C: Induced
thermotolerance and the heat-shock protein-70 family in the Pacific
oyster Crassostrea gigas. Mol. Mar. Biol. Biotechnol. 1998, 7:21-30.
[51] Hamdoun AM, Cheney. D.O., Cherr GN: Phenotypic plasticity of HSP
70 and HSP 70 gene expression in the Pacific oyster (Crassostrea
gigas): Implications for thermal limits and induction of thermal
tolerance. Biol. Bull. 2003, 205(2):160-169.
[52] Houlihua DF: Protein turnover in ectotherms and its relationships to
energetics. Adv. Comp. Env. Physiol. 1991, 7:1-43.

Exploring Correlations… 201
[53] Kregel KC: Heat shock proteins: modifying factors in physiological
stress responses and acquired thermotolerance. J. Appl. Physiol. 2002,
99:2177-2186.
[54] Pörtner HO: Climate variations and the physiological basis of
temperature dependent biogeography: systemic to molecular hierarchy
of thermal tolerance in animals. Comp. Biochem. Physiol. A 2002,
132:739-761.
[55] Pörtner HO: Climate change and temperature dependent biogeography:
Oxygen limitation of thermal tolerance in animals. Naturwissenschaften
2001, 88:137-146.
[56] Shofer SL, Tjeerdema RS: Effects of hypoxia and toxicant exposure on
adenylate energy charge and cytosolic ADP concentrations in abalone.
Comp. Biochem. Physiol. C 1998, 119(1):51-57.
[57] Cattani O, Serra R, Isani G, Raggi G, Cortesi P, Carpene E: Correlation
between metallothionein and energy metabolism in sea bass,
Dicentrarchus labrax, exposed to Cadmium. Comp. Biochem. Physiol. C
1996, 113(2):193-199.
[58] Duchemin MB, Fournier M, Auffret M: Seasonal variation of immune
parameters in diploid and triploid Pacific oysters, Crassostrea gigas
(Thunberg). Aquaculture 2007, 264:73-81.
[59] Delaporte M, Soudant P, Lambert C, Jegaden M, Moal J, Pouvreau S,
Dégremont L, Boudry P, Samain JF: Characterisation of physiological
and immunological differences between Pacific oysters (Crassostrea
gigas) genetically selected for high or low survival to summer
mortalities and fed different rations under controlled conditions. J. Exp.
Mar. Biol. Ecol. 2007, 353:45-57
[60] Lambert C, Soudant P, Dégremont L, Delaporte M, Moal J, Boudry P,
Jean F, Huvet A, Samain JF: Hemocyte characteristics in families of
oysters, Crassostrea gigas, selected for differential survival during
summer and reared in three sites Aquaculture 2007, 270:276-288
[61] Pipe RK, Coles JA: Environmental contaminations influencing immune
function in marine bivalve molluscs. Fish Shellfish Immunol. 1995,
5:581-595.
[62] Anderson RS, Beaven AE: Antibacterial activities of oyster (Crassostrea
virginica) and mussel (Mytilus edulis and Geukensia demissa) plasma.
Aquat. Living Resour. 2001, 14(6):343-349.
[63] Montes JF, Del Rio JA, Durfort M, Garcia-Valero J: The protozoan
parasite Perkinsus atlanticus elicits a unique defense response in the lam
Tapes semidecussatus. Parasitol. 1997, 114:339-350.

Yan Li, Kirsten Benkendorff, Jian G. Qin et al. 202
[64] Montes JF, Durfort M, Garcia-Valero J: Cellular defence mechanism of
the clam Tapes semidecussatus against infection by the protozoan
Perkinsus sp. Cell Tissue Res. 1995, 279:529-538.
[65] Origosa M, SEsteve C, Pujalte MJ: Vibrio species in seawater and
mussels: abundance and numerical taxonomy. Syst. Appl. Microbiol.
1989, 12:316-325.
[66] Pouvreau S, Enriquez-Diaz M, Le Souchu P, Connan JP, Le Roy B,
Mingant C, Moal J, Delaporte M, Le Coz JR, Samain JF: Reproduction,
bioenergetic and summer mortality of Crassostrea gigas: experimental
approach. J. Shellfish Res. 2003, 22:351.
[67] Hayes ML, Bonaventura J, Mitchell TP, Prospero JM, Shinn EA, Dolah
FV, Barber RT: How are climate and marine biological outbreaks
functionally linked? Hydrobiologia 2001, 460:213-220.
[68] Li Y: Spawning mediated responses of Pacific oyster Crassostrea gigas
to environmental stress: toward an explanation of oyster summer
mortality Adelaide: Flinders University; 2008.
[69] Scanes P: 'Oyster watch': Monitoring trace metal and organochlorine
concentrations in Sydney's coastal waters. Mar. Pollut. Bull. 1996, 33(7-
12):226-238.
Copyright © 2022 FDOKUMEN




















