
Bioremediation of aflatoxin B1-contaminated maize by king oyster ...
Upload
independentCategory
view
0download
0
mmeoant(pCwaauwiasmvssamriA
pC
tGk
Journal of Invertebrate Pathology 74, 173–183 (1999)Article ID jipa.1999.4879, available online at http://www.idealibrary.com on
Analysis of the Effects of Perkinsus marinus Proteases on PlasmaProteins of the Eastern Oyster (Crassostrea virginica)
and the Pacific Oyster (Crassostrea gigas)
Jacques L. Oliver, Teresa D. Lewis, Mohamed Faisal, and Stephen L. Kaattari1
Department of Environmental Sciences, School of Marine Science, Virginia Institute of Marine Science,The College of William and Mary, Gloucester Point, Virginia 23062
Received October 13, 1998; accepted May 28, 1999
pD(pcaidest
dmmpdddp1phptpnfephtr
r
We employed two in vitro buffer systems to deter-ine the potential pathogenic effects of Perkinsusarinus serine proteases on the plasma proteins of the
astern oyster (Crassostrea virginica) and the Pacificyster (Crassostrea gigas). Specifically, this study char-cterized the oyster plasma protein targets of P. mari-us proteases. Additionally, protease-specific inhibi-ory activity was revealed upon comparison of artificialPBS) and endogenous (plasma-based) diluents em-loyed during protease digestions. It was found that a. virginica plasma protein of approximately 35 kDaas eliminated when a standard buffer (PBS) was useds a diluent; however, this protein was preserved when
low-molecular-weight, plasma-based, diluent wassed. The results strongly indicate that low-molecular-eight inhibitors of P. marinus proteases are present
n oyster plasma. A control (nonparasitic) serine prote-se, a-chymotrypsin, was employed to ascertain thepecificity of the protease inhibitors. Although a-chy-otrypsin possesses ample proteolytic activity for C.
irginica plasma proteins, the anti-proteases couldpecifically inhibit only P. marinus proteases. Suchpecificity of anti-protease activity is not uncommonmong low-molecular-weight serine proteases. The he-olymph target protein was isolated by 2D electropho-
esis and isoelectrically isolated for further character-zation by N-terminal amino acid sequencing. r 1999
cademic Press
Key Words: Perkinsus marinus; proteases; plasmaroteins; protease inhibitors; Crassostrea virginica;rassostrea gigas; pathogenesis.
1 To whom correspondence should be addressed at Virginia Insti-ute of Marine Science, Chesapeake Bay Hall, P. O. Box 1346,loucester Point, Virginia 23062. Fax: (804) 684-7186. E-mail:
173
INTRODUCTION
Perkinsus marinus is a protozoan pathogen (Apicom-lexa; Levine, 1978) that is the causative agent ofermo, a lethal disease afflicting the eastern oyster
Crassostrea virginica) (Andrews, 1988). Although as-ects of the disease process, such as adverse physiologi-al effects on the eastern oyster (Paynter, 1996) andbiotic effects on the susceptibility, transmission, andnfectivity of P. marinus (Chu, 1996), have been ad-ressed, little is known of the pathogenic mechanismsmployed (Anderson, 1996). Such knowledge is neces-ary for designing therapeutic and prophylactic modali-ies.
Successful in vitro propagation of the pathogen, asescribed by La Peyre et al. (1993), has provided theeans to isolate and analyze the virulence factors of P.arinus. Among the in vitro secreted extracellular
roteins of P. marinus, La Peyre and Faisal (1995)iscovered chymotrypsin-like serine proteases. Theyemonstrated that these proteases were capable ofigesting a variety of proteins, including hemolymphroteins. As with many other pathogens (McKerrow,989; McKerrow et al., 1993), the possibility that theseroteases could also facilitate invasion, compromiseost defense mechanisms, and provide the invadingathogen with suitable nutrients has been explored. Inhis context, it was demonstrated that P. marinusroteases can compromise immune defense mecha-isms of the eastern oyster (Garreis et al., 1996) and
avor the protozoan’s propagation in oysters (La Peyret al., 1996). Identification and characterization ofutative molecular targets of P. marinus proteases,owever, has yet to be performed. One impediment tohese studies has been the general dearth of knowledgeegarding the protein composition of oyster plasma.Of additional importance to P. marinus pathogenesis
esearch is the fact that Pacific oysters (Crassostrea
igas) contract only light infections upon exposure.0022-2011/99 $30.00Copyright r 1999 by Academic Press
All rights of reproduction in any form reserved.

MtmvmPMpfpnCt
cptbstwedsbtop
O
mMRpaH(wfdR
fOusq
ma
P
istmpctNCreuact
D
fPws
P
RmPCwfweAwosar
I
smam
tdIt
174 OLIVER ET AL.
eyers et al. (1991) found that prevalences and mortali-ies in diploid and triploid C. gigas exposed to P.arinus were lower than that in diploid and triploid C.
irginica. This suggested a tolerance of C. gigas to P.arinus; however, the exact mode of resistance ofacific oysters to Dermo remains to be elucidated.eyers et al. (1991), however, postulated that the
athogen might be unable to proliferate in C. gigas. Inact, La Peyre (1993) noted that the lower hemolymphrotein concentrations in Pacific oysters may pose autritional limitation for the pathogen. Alternatively,. gigas might possess specific defense mechanisms
hat allow the host to eliminate the pathogen.This study was conducted to determine which plasma
omponents may be the primary targets of P. marinusroteases in C. virginica and C. gigas. Such informa-ion may be critical in revealing the possible molecularasis of Perkinsus pathogenesis. Paramount to suchtudies is the development of a protease assay systemhat simulates in vivo physio-chemical conditions. Itas found that when an artificial buffer, PBS,2 was
mployed in the protease assay system, a distinctlyifferent digestion pattern of plasma proteins is ob-erved than that observed with an endogenous, plasma-ased diluent. Based on these observations, the exis-ence of low-molecular-weight protease inhibitors inyster plasma was revealed and these inhibitors wererotease specific.
MATERIALS AND METHODS
ysters
Eastern oysters (shell height approximately 70–80m) were purchased from Permaquid Inc., Waldoboro,aine, and acclimated for 3 months in filtered Yorkiver (Virginia) water. Oysters were notched and ap-roximately 3–4 ml of hemolymph were bled from thedductor sinus using a syringe and 25-gauge needle.emolymph was then filtered using a 0.45-µm filter
Costar Scientific Corp., Cambridge, MA) and aliquotsere stored in sterile microfuge tubes at 280°C until
urther use. Oyster plasma was assayed for proteinetermination using BCA protein assay kit (Pierce,ockford, IL).Pacific oysters, Crassostrea gigas, were obtained
rom Lookout Point Oyster Company (Lookout Point,regon). Oysters were notched and bled immediatelypon arrival as described above. Hemolymph wasterile filtered using 0.45 µm filter (Costar) and ali-uots stored at 280°C until further use. Protein deter-
2 Abbreviations used: PBS, phosphate-buffered saline; ppt, part perhousand; T-TBS, Tween–tris-buffered saline; PVDF, polyvinylideneifluoride; ECL, enhanced chemiluminescence; pI isoelectric point;EF, isoelectric focusing; BCA, bicinchoninic acid; DTT, dithio-
ehrietol.
inations of plasma were determined using the BCAssay (Pierce).
erkinsus marinus Extracellular Protein(ECP) Preparation
Perkinsus marinus-1 cells were cultured for 8 weeksn JL-ODRP-1 medium (La Peyre et al., 1993). Cultureupernatants were obtained by centrifuging the cul-ures at 3600g in 50-ml centrifuge tubes at 4°C for 20in. The supernatants were decanted into Centri-
rep-10 concentrators (Amicon Inc., Beverly, MA) andentrifuged at 3600g for 40 min at 4°C. After centrifuga-ion, cold (4°C) PBS (137 mM NaCl, 3 mM KCl, 8 mMa2HPO4, 2 mM KH2PO4, pH 7.2) was added to theentriprep-10 apparatus and the centrifugation stepepeated. This procedure was performed three times toquilibrate the ECP in PBS. ECP was then filteredsing a 0.22-µm bottle filter system (Costar) and storedt 4°C until further use. Protein determination ofoncentrated ECP was performed using the BCA pro-ein assay (Pierce).
etection of Protease Activity
A qualitative analysis of concentrated ECP was per-ormed to detect and confirm the proteolytic activity of. marinus proteases. The proteolytic activity of ECPas analyzed using substrate (gelatin) gel electrophore-
is as described by La Peyre et al. (1995).
roduction of C. virginica Plasma Protein Antisera
Female New Zealand rabbits (2.27 kg) (Franklinabbitry, Wake Forest, NC) were immunized with 0.45l of 1:1 emulsion of 200 µg C. virginica plasma/mlBS and Freund’s complete adjuvant (Sigma Chemicalo., St. Louis, MO). Fifty microliters of the emulsionas injected intramuscularly into the hind legs and
our 100-µl subcutaneous injections along the backere administered. Rabbit serum was procured fromar vein bleeds at 2 and 4 months post-injection.liquots were frozen at 280°C until used. Antiseraere tested for reactivity with eastern and Pacificyster plasma through immunoblot analysis. Resultshowed cross-reactivity of rabbit antisera to all observ-ble proteins in both species of oysters and no cross-eactivity with P. marinus protein.
ncubation of Oyster Plasma with P. marinus Proteasesand a-Chymotrypsin
Ten micrograms of concentrated ECP was placed in aterile microfuge tube at a volume of 14 µl. Tenicrograms of C. virginica oyster plasma proteins was
dded to the same tube in a volume of 2 µl. The reactionixture was brought to a final volume of 100 µl using
ither PBS or a plasma-based diluent. The latter

da(ar7mPositetf1ass
iTsruetip7tmmIf
itaµvPwc
I
r1uLpHmwC
ablw(Mari(Kairpbwr1dmMagtSrpRwsS
T
dtapmtldcs(dts
N
175EFFECTS OF P. marinus PROTEASES ON PLASMA PROTEINS
iluent was generated by centrifuging oyster plasma inCentricon-10 concentrator (Amicon Inc.). The filtrate
containing ,10-kDa proteins) was retained and useds the plasma-based diluent. The pH of the PBSeaction mixture and the plasma-based diluent were.2 and 7.4, respectively. The osmolarity of the reactionixtures was measured using a Westcor 5500 Vaporressure Osmometer (Westcor, Inc., Logan, UT). Thesmometer was calibrated using 100 and 1000 mmol/kgtandards. Ten microliters of each mixture was loadednto the osmometer and measured. The osmolarity ofhe PBS reaction mixture and the plasma-based dilu-nt was 316 and 476 mmol/kg, respectively. The reac-ion mixture was capped and covered with aluminumoil and placed in an end-over-end rotator for 24 h at7°C. In separate tubes, negative controls consisting ofn aliquot of ECP and an aliquot of oyster plasma wereimilarly placed on the rotator and incubated for theame duration.Ten microliters of C. gigas plasma protein correspond-
ng to 10 µg of protein was added to the microfuge tube.en micrograms of concentrated ECP was placed in aterile microfuge tube at a volume of 13.7 µl. Theeaction mixture was brought to a final volume of 100 µlsing PBS or the C. gigas-derived plasma-based dilu-nt generated as previously described. Negative con-rols were unmixed ECP and C. gigas plasma incubatedn separate microfuge tubes. The pH of the PBS andlasma-based diluent reaction mixtures were 7.2 and.4, respectively. Osmolarity of the two reaction mix-ures was 407 mmol/kg for the PBS diluent and 854mol/kg for the plasma-based diluent. Osmolarityeasurements were made as previously described.
ncubations of the mixture and controls were per-ormed as previously described.
a-Chymotrypsin from bovine stomach (Sigma) wasncubated with eastern oyster plasma proteins usinghe methods previously described. Briefly, 10 µg of-chymotrypsin diluted in PBS was incubated with 10g of eastern oyster plasma proteins. The remainingolume of the reaction mixture was filled with eitherBS or the plasma-based diluent. Analysis of targetsas performed using SDS–PAGE, immunoblotting, and
olloidal gold staining procedures described above.
dentification of Protease Targets in Oysters
After the protease incubation, an aliquot from eacheaction mixture was electrophoresed in a reducing2% SDS–polyacrylamide gel (Harlow and Lane, 1988)sing a mini-PROTEAN II Cell apparatus (Bio-Radaboratories, Hercules, CA). All samples were pre-ared in reducing (DTT) sample buffer as described byarlow and Lane (1988). The volume of the reactionixture loaded onto the gel was equivalent to thathich would contain 1 µg of oyster plasma protein.
ontrols (plasma incubated without ECP and ECP 2lone) were analyzed in the same manner. Biotinylatedroad-range molecular weight markers (Bio-Rad) wereoaded as a reference. After electrophoresis, proteinsere transblotted at 100 V for 1 h to PVDF membranes
Millipore Corp., Burlington, MA), as described by theini Trans-blot Electrophoretic Cell Manual (Bio-Rad),
nd immunoblotting analysis was performed to moreeadily identify oyster plasma protein targets. Follow-ng transblotting, membranes were blocked with T-TBS0.1% (v/v) Tween 20, 25 mM Tris, 137 mM NaCl, 3 mMCl, 8 mM Na2HPO4, 2 mM KH2PO4, pH 7.2) (Arkooshnd Kaattari, 1990) overnight at 4°C. Membranes werencubated on a shaker at room temperature with aabbit antiserum generated against C. virginica plasmaroteins (diluted 1:5000 in T-TBS) for 1.5 h. Mem-ranes were washed 3 times in T-TBS at 5 min perash. Membranes were incubated with a goat-anti-
abbit-HRPO antibody (Sigma Chemical Co.) (diluted:5000 in T-TBS) and streptavidin–horse radish peroxi-ase (diluted 1:1000 in T-TBS) for molecular weightarker identification for 1 h at room temperature.embranes were washed 5 times in T-TBS. ECL re-
gents (Amersham Life Science, Buckinghamshire, En-land) were used for chemiluminescent development ofhe membranes on ECL Hyperfilm (Amersham Lifecience). Following ECL development, membranes wereinsed 10 times with deionized-distilled water and totalrotein was stained with Colloidal Gold Stain (Bio-ad). Target protein molecular weight determinationas performed via comparison to molecular weight
tandards using densitometric analysis (Integratedeparation Systems Enprotech, Natick, MA).
wo-Dimensional Isoelectric Focusing (2D IEF) andWestern Blot of Eastern Oyster Plasma Proteins
A two-dimensional IEF analysis was performed toetermine potential pI diversity of the 35-kDa target inhe eastern oyster plasma previously identified in thebove analyses. First-dimension IEF tube gels wererepared and samples electrophoresed according to theethods described by Harlow and Lane (1988) using
he Bio-Rad Minigel apparatus. Each tube gel wasayered upon 12% SDS–polyacrylamide gels for second-imension electrophoresis. In order to attain reducingonditions for the tube gel, 200 µl of reducing (DTT)ample buffer prepared according to Harlow and Lane1988) was added on top of the tube gel. After second-imension electrophoresis was performed, proteins wereransblotted to PVDF membranes, immunoblotted, andtained by the methods previously described.
-Terminal Amino Acid Sequencing and Amino AcidComposition of Eastern Oyster Target Proteins
Five micrograms of target protein was isolated via
D IEF as previously described. Preparation of the
mapmP0mtmtrbafnatS(pm
O
prtbc(aacbsdw
eeUmaiog
rpgsh
alalhpm2o3w
E
PPd3o(33
pe3di
T
ttt3g(tIcleo4f
plaaag
176 OLIVER ET AL.
embrane for N-terminal sequencing and amino acidnalysis was performed using a protocol (Gilardi, 1996)rovided by Commonwealth Biotechnologies, Inc. (Rich-ond, VA). Briefly, following transblot of proteins to aVDF membrane, the membrane was stained with.1% (v/v) Coomassie Brilliant Blue R-250 in 50% (v/v)ethanol/deionized-distilled water for 5 min at room
emperature. Membranes were destained with 50%ethanol, 10% acetic acid, 40% deionized-distilled wa-
er for 10 min at room temperature. Membranes wereinsed twice with deionized-distilled water, equili-rated for 10 min in deionized-distilled water, and thenir dried for 1 h. The target protein band was excisedrom the membrane and sent to Commonwealth Biotech-ologies, Inc. for N-terminal amino acid sequencingnd amino acid composition analysis. Sequence informa-ion was compared to known sequences by searchingWISSPROT using BLASTP (2.0.8). The Expect value
E) in the database search was increased to 1000 torovide statistically significant or nonsignificantatches.
RESULTS
yster Plasma Protein Digests Using PBS andPlasma-Based Diluents
PBS diluent. Co-incubation of C. virginica plasmaroteins with P. marinus proteases using a PBS diluentesulted in the complete elimination of a 35-kDa pro-ein band (Figs. 1a and 1b, lane 3, arrow). An immuno-lot using a rabbit anti-C. virginica plasma permitted alearer resolution of the digested oyster ‘‘target’’ proteinFig. 1a, lane 4, arrow). It should be noted that andditional band appears upon digestion (Fig. 1a, lane 3,sterisk). It is unlikely that this band representsross-reactivity with P. marinus ECP since no suchand is apparent in lane 2. The appearance of this banduggests that it may be a product of partial proteolyticigestion of an as yet to be determined high-molecular-eight species by ECP.
Plasma-based diluent. Use of a physiologically rel-vant plasma-based diluent revealed a strikingly differ-nt digestion profile from that observed with PBS.nder these conditions the 35-kDa protein band re-ains substantively intact (Figs. 1c and 1d, lane 3,
rrow). Although the 35-kDa protein appears protectedn this diluent, some partial digestion appears to beccurring, as new, low-molecular-weight species areenerated (Figs. 1c and 1d, lane 3, asterisks).Plasma proteins from the relatively P. marinus-
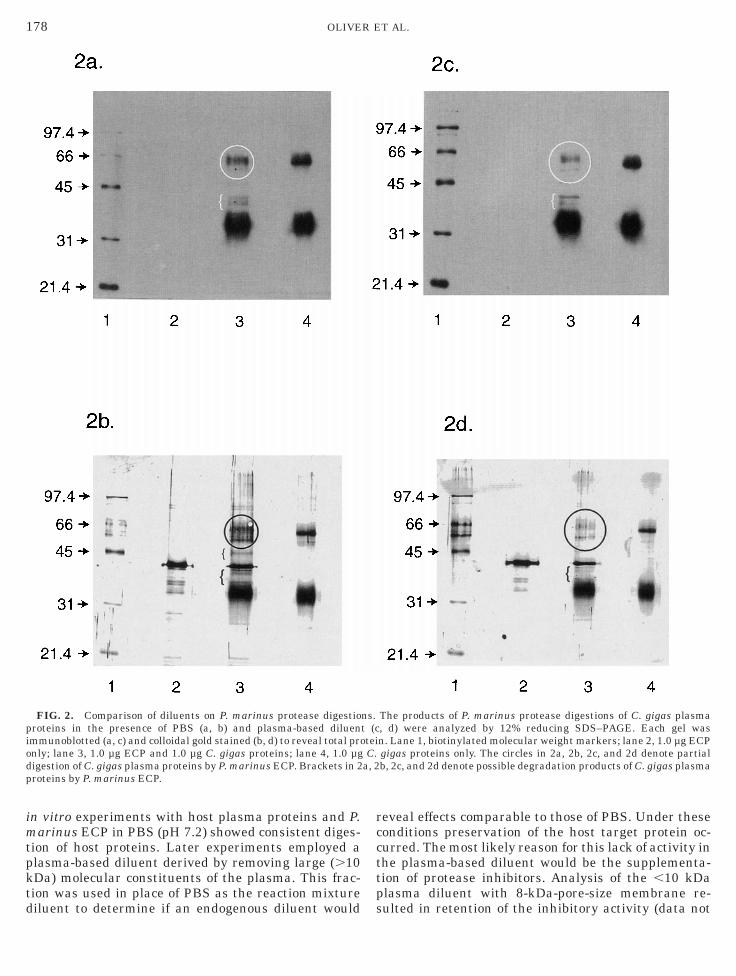
esistant oyster, C. gigas, were exposed to P. marinusroteases under identical conditions used with C. vir-inica (Fig. 2). In this case, the PBS diluent did notupport the complete digestion of any plasma proteins;
owever, partial digestion appears to be occurring with pprotein of approximately 66 kDa (Figs. 2a and 2b,ane 3, circle). In addition, some breakdown products ofpproximately 35 kDa are observed (Figs. 2a and 2b,ane 3, brackets) which may be due to degradation ofigh-molecular-weight proteins. In the presence of thelasma diluent, a more intense digestion of the higher-olecular-weight species (approximately 66 kDa) (Figs.
c and 2d, lane 3, circle) occurs, leaving an assortmentf lower-molecular-weight species (Figs. 2c and 2d, lane, bracket). However, digestion is not as thorough asith C. virginica proteins.
ffects of a-Chymotrypsin on Eastern Oyster PlasmaProtein Substrates
Incubation of eastern oyster plasma with a non-erkinsus (bovine) serine protease, a-chymotrypsin, inBS resulted in proteolysis of the 35-kDa target, as wasemonstrated with P. marinus serine proteases (Figs.a and 3b, lane 3, arrow). In addition to the eliminationf the 35-kDa target, a 30-kDa band becomes apparentFigs. 3a and 3b, lane 3, asterisk). It is likely that the0-kDa band represents degradation products of the5-kDa target protein.When the diluent was switched from PBS to the
lasma-based diluent, the result was unchanged. East-rn oyster plasma proteins were digested (Figs. 3c andd, lane 3, arrow). In addition, there appear to beegradation products at 30 kDa (Fig. 3c, lane 3, aster-sk), as seen in Figs. 3a and 3b, lane 3.
wo-Dimensional Isoelectric Focusing of EasternOyster Plasma Proteins
Further characterization of the plasma proteins bywo-dimensional isoelectric focusing and immunoblot-ing revealed a rather restricted pI heterogeneity forhe 35-kDa target protein (Fig. 4a). Specifically, the5-kDa target of the eastern oyster shows no pI hetero-eneity and the target is composed of a single proteinFigs. 4a and 4b, circle/arrow). Identification of thearget can be determined upon comparison of the 2DEF of oyster plasma exposed to ECP (Figs. 4c and 4d,ircle/arrow) and the oyster plasma control (Fig. 4c,ane 2, arrow). Comparison of Figs. 4a and 4c revealslimination of a second target near 66 kDa not previ-usly seen in one-dimensional immunoblotting (Figs.a and 4c, circle/asterisk). The isoelectric point rangeor the 35-kDa target is 5.9–6.6.
A second 2D IEF of oyster plasma proteins waserformed. Proteins were transblotted and noncova-ently stained. N-terminal amino acid sequencing andmino acid composition analysis revealed the first 10mino acid residues (Table 1). Amino acid compositionnalysis revealed the target to be abundant in serine,lycine, and alanine (Table 2). Database searches were
erformed to match the obtained sequence with other
km(eE
ppC
EOatapec
pe
piEt
177EFFECTS OF P. marinus PROTEASES ON PLASMA PROTEINS
nown amino acid sequences. Results revealed the bestatch to be with plant plasma membrane ATPases
50% identity); however, these matches were consid-red to be statistically nonsignificant due to the highxpect values of the returned matches (data not shown).
DISCUSSION
This study was conducted to analyze the in vitroathogenic effects of P. marinus proteases on plasmaroteins of two oysters, C. virginica and C. gigas. When
FIG. 1. Comparison of diluents on P. marinus protease digestionsroteins in the presence of PBS (a, b) and plasma-based diluentmmunoblotted (a, c) and colloidal gold stained (b, d) to reveal the totaCP only; lane 3, 1.0 µg ECP and 1.0 µg C. virginica proteins; lane 4, 1
he 35-kDa plasma protein target in C. virginica. Asterisks in 1a, 1c, a
. virginica proteins were incubated with P. marinus n
CP, degradation of a 35-kDa protein was observed.ne-dimensional SDS–PAGE of host plasma proteinsllowed us to characterize the molecular weight of thearget; however, two-dimensional isoelectric focusingnd electrophoresis was required to determine theossible isoelectric heterogeneity of proteins. The iso-lectric diversity of the target appears to be limited andonsisting of one or possibly two proteins.Our experiments were designed to determine any
lasma protein targets of P. marinus proteases; how-ver, we were able to additionally identify a potentially
he products of P. marinus protease digestions of C. virginica plasmac, d) were analyzed by 12% reducing SDS–PAGE. Each gel wasrotein. Lane 1, biotinylated molecular weight markers; lane 2, 1.0 µgg C. virginica proteins only. The arrows in 1a, 1b, 1c, and 1d indicate
1d denote possible degradation products of P. marinus ECP digestion.
. Ts (l p.0 µnd
ovel host defense factor in C. virginica plasma. Initial
imtpktd
rccttp
piodp
178 OLIVER ET AL.
n vitro experiments with host plasma proteins and P.arinus ECP in PBS (pH 7.2) showed consistent diges-
ion of host proteins. Later experiments employed alasma-based diluent derived by removing large (.10Da) molecular constituents of the plasma. This frac-ion was used in place of PBS as the reaction mixture
FIG. 2. Comparison of diluents on P. marinus protease digestioroteins in the presence of PBS (a, b) and plasma-based diluenmmunoblotted (a, c) and colloidal gold stained (b, d) to reveal total pronly; lane 3, 1.0 µg ECP and 1.0 µg C. gigas proteins; lane 4, 1.0 µgigestion of C. gigas plasma proteins by P. marinus ECP. Brackets in 2roteins by P. marinus ECP.
iluent to determine if an endogenous diluent would s
eveal effects comparable to those of PBS. Under theseonditions preservation of the host target protein oc-urred. The most likely reason for this lack of activity inhe plasma-based diluent would be the supplementa-ion of protease inhibitors. Analysis of the ,10 kDalasma diluent with 8-kDa-pore-size membrane re-
The products of P. marinus protease digestions of C. gigas plasma, d) were analyzed by 12% reducing SDS–PAGE. Each gel wasn. Lane 1, biotinylated molecular weight markers; lane 2, 1.0 µg ECPgigas proteins only. The circles in 2a, 2b, 2c, and 2d denote partialb, 2c, and 2d denote possible degradation products of C. gigas plasma
ns.t (cteiC.
a, 2
ulted in retention of the inhibitory activity (data not

srbowlgaowaf
mwaoetpti1i
Cwm13 di
179EFFECTS OF P. marinus PROTEASES ON PLASMA PROTEINS
hown). This indicates that the inhibitory activityesides with 8- to 10-kDa-sized molecule(s) and cannote due to factors such as minor differences in pH,smolarity, or other aspects of ion or low-molecular-eight composition. Furthermore, evidence that osmo-
arity of each diluent (PBS, C. virginica-derived or C.igas-derived) was not a factor affecting proteolyticctivity is the observation that expression and activityf P. marinus proteases occurs in a culture mediumith elevated osmolarity (650 mmol/kg) (La Peyre etl., 1993, 1995). Further evidence that pH was not a
FIG. 3. Comparison of diluents on a-chymotrypsin digestion of C.. virginica plasma proteins in the presence of PBS (a, b) and plasma-as immunoblotted (a, c) to reveal the plasma proteins and colloidolecular weight markers; lane 2, 1.0 µg a-chymotrypsin only; lane 3
.0 µg C. virginica plasma proteins only. The arrows in 3a, 3b, 3c, and 3a, 3b, and 3c denote possible degradation products of a-chymotrypsin
actor affecting proteolytic activity is the finding that P. i
arinus protease activity occurs in a variety of buffersith varying pH (5.0–9.5), including PBS (La Peyre etl., 1995). Thus, if plasma protease inhibitors areperative, it may be that dilution of plasma in PBS mayffectively reduce endogenous inhibitors to a concentra-ion insufficient to block the activity of P. marinusroteases. Protease inhibitors are known to be impor-ant regulatory molecules as well as having importancen humoral defense (Ellis, 1987; Calkins and Sloane,995; Wiedow et al., 1998). Furthermore, proteasenhibitors have been identified in a number of marine
ginica plasma proteins. The products of a-chymotrypsin digestions ofed diluent (c, d) were analyzed by 12% reducing SDS–PAGE. Each gelgold stained (b, d) to reveal the total protein. Lane 1, biotinylated0 µg a-chymotrypsin and 1.0 µg C. virginica plasma proteins; lane 4,ndicate the 35-kDa plasma protein target in C. virginica. Asterisks ingestion.
virbasal, 1.d i
nvertebrates (Donovan and Laue, 1991; Sousa et al.,

1T
mbpsfioRp
TsniasapPp3
Crbap
180 OLIVER ET AL.
992) and specifically in molluscs (Bender et al., 1992;horgersen et al., 1992; Armstrong and Quigley, 1992).La Peyre et al. (1995) noted the similarity of P.arinus proteases to chymotrypsin-like enzymes. Incu-
ations of eastern oyster plasma with another serinerotease, a-chymotrypsin, were undertaken to comparepecificity of P. marinus proteases and a-chymotrypsinor plasma proteins. Additionally, the specificity of thenhibitory activity in the ,10-kDa fraction of easternyster plasma against a-chymotrypsin was examined.esults showed that a-chymotrypsin can recognize and
FIG. 4. Two-dimensional electrophoretic analysis of C. virginica p. virginica plasma protein samples were analyzed by two-dimensio
educing SDS–PAGE, second dimension). Each gel was immunoblotteiotinylated molecular weight markers, 2D gel, 1.0 µg C. virginica plan arrow in 4a, 4b, 4c, and 4d denotes the 35-kDa plasma protein taossible additional plasma protein target.
roteolytically digest eastern oyster plasma proteins. (
hus, P. marinus serine proteases and a-chymotrypsinhare substrate specificity. However, whereas P. mari-us proteases were successfully inhibited from digest-
ng oyster proteins with the endogenous inhibitor,-chymotrypsin was uninhibited. This differential sen-itivity among serine proteases is common. For ex-mple, La Peyre et al. (1995) showed that P. marinusroteases exhibited low activity in the presence ofMSF and chymostatin but were quite active in theresence of other serine protease inhibitors, such as,4-DCI and elastatinal. Additionally, Faisal et al.
ma proteins. Undigested (a, b) and P. marinus protease-digested (c, d)electrophoretic analysis (isoelectric focusing—first dimension, 12%, c) and colloidal gold stained (b, d) to reveal the total protein. Lane 1,
a proteins; lane 2, 1.0 µg C. virginica plasma proteins. The circle witht in C. virginica. The circle with an asterisk in 4a and 4c denotes a
lasnald (asmrge
1998) suggested that the ability of C. gigas to strongly

iwsta
teiPtdtetpteaPtcttssemtmmpt
wagirraqcpdiptfio
5s
fomaKiEpcea
atmarSOk(fiAt
AGSHGTAATCVMTPILLPHEBC
v
181EFFECTS OF P. marinus PROTEASES ON PLASMA PROTEINS
nhibit P. marinus proteases yet weakly inhibit trypsinas due to specificity of protease inhibitors in Cras-
ostrea spp. Thus, it appears that inhibitory activity ofhe eastern oyster is specific and may have developednd been directed against P. marinus proteases.Pacific oysters have been of particular interest due to
heir ability to resist P. marinus upon challenge (Mey-rs, 1991). Having observed the patterns of susceptibil-ty and resistance of eastern oyster plasma proteins to. marinus proteases, it was of interest to determinehe effects on Pacific oyster plasma proteins. Resultsemonstrated that C. gigas plasma proteins are suscep-ible to proteolysis in both diluents. Despite the appar-nt partial digestion of an approximately 66-kDa pro-ein in Pacific oyster plasma, the sensitivity of thisrotein appears to be less than the sensitivity of the p35arget in the eastern oyster, as p35 was completelyliminated under identical conditions. Although thereppears to be a target of P. marinus proteases withinacific oyster plasma, there are a number of possibili-ies that may preclude disease susceptibility beingontingent upon the loss of this target. The 66-kDaarget may be inconsequential to any vital process inhe Pacific oyster.Alternatively, the incomplete proteoly-is of the protein suggests that the protein is onlylightly sensitive to P. marinus proteases and fulllimination of this protein does not occur. Finally, thereay be mechanisms utilized by Pacific oysters outside
he scope of this in vitro experiment that prevent P.arinus from exerting its virulence via proteases. Thisay occur by suppression of protease expression or
erhaps rapid clearance of the pathogen before replica-ion can occur.
The possible function of the target molecule (p35)ithin C. virginica hemolymph may be most readilyscertained by identification of structurally homolo-ous molecules. Therefore, as the p35 protein wassoelectrically restricted, its isolation and analysis waseadily accomplished. Amino acid composition analysisevealed p35 to be abundant in serine, glycine, andlanine. Furthermore, the N-terminal amino acid se-uence of the first 10 residues of p35 was obtained. Thelosest sequence identity (50%) was found with plantlasma membrane ATPases when a search was con-ucted to ascertain homologous sequences; thus, thenformation gained from the database searches hasrovided limited insight into the possible function ofhe p35 target. Although sequence matches were identi-ed, it is likely that such matches were merely randomccurrences. This is evident by the high E values (428,
TABLE 1N-Terminal Amino Acid Sequence of the p35 Target
from the Eastern Oyster
H2N—Tyr—Pro—Leu—Glu—His—Asn—Asn—Tyr—Gln—Asp
p61, and 735) of each match, which signified lowtatistical significance.Additional information, albeit limited, can be gleaned
rom the amino acid composition of p35. The abundancef alanine, an uncharged amino acid with a nonaro-atic side chain, in p35 is evidence that supports p35
s a specific target of P. marinus serine proteases.raut (1977) described elastase, another common ser-
ne protease, as having preference for alanine residues.lastase was 4000 times more efficient in hydrolyzingeptide substrates with amino acid sequences thatontained consecutive alanine residues. Further infer-nces about the function and identity of p35 throughmino acid composition cannot be made.Despite the limited effectiveness of the amino acid
nalyses for protein identification, the unique combina-ion of molecular weight and pI are suggestive that p35ight be a subunit of a serum lectin. Vasta et al. (1984)
ffinity purified a serum lectin of C. virginica using rated blood cells (rRBC) and analyzed it via reducingDS–PAGE. Their results showed three subunit bands,SLS-I (67 kDa), OSLS-II (34 kDa), and OSLS-III (25Da). It is interesting to note the presence of a 34-kDaOSLS-II) protein identified by Vasta et al. and ourndings of a 35-kDa target in oyster hemolymph.dditionally, Vasta et al. (1984) isoelectrically focused
he serum lectin and found that the lectin had a broad
TABLE 2Amino Acid Analysis of 35-kDa Plasma Protein Target
(p35) from C. virginica
nmolDetected
Total nmolin sample
Equivalentµg Res/mol
s(x) 0.0231 1.848 0.21252 20.71733l(x) 0.018 1.44 0.18576 16.14337er 0.0397 3.176 0.276312 35.60511is 0.0059 0.472 0.064664 5.291439ly 0.1307 10.456 0.595992 117.2188hr 0.0113 0.904 0.091304 10.13445la 0.0881 7.048 0.500408 79.01285rg 0.015 1.2 0.1884 13.45281yr 0.0156 1.248 0.203424 13.99092ystine 0 0 0 0al 0.0367 2.936 0.290664 32.91455et 0.003 0.24 0.03144 2.690562
rp 0 0 0 0he 0.0107 0.856 0.125832 9.596339le 0.0066 0.528 0.59664 5.919237eu 0.0103 0.824 0.093112 9.237597ys 0.0025 0.2 0.0256 2.242135ro 0.0228 1.824 0.176928 20.44827yPro 0 0 0 0ACA 0 0 0 0-Ala 0 0 0 0ys 0 0 0 0
Note. Results indicate abundance of glycine, alanine, serine, andaline in p35. MGS in sample: 0.003122.
I range (pH 5–6 and pH 7–8) with slightly greater

hpnImattmavawmhFeOat
igpcopa
potirP1itptgpppihmthlAagat
ipsigpcTmTvatpurm
simDipp
ABrNGNsI
A
A
A
A
A
A
A
182 OLIVER ET AL.
eterogeneity than our 35-kDa target. The affinity-urified lectin was also tested for its ability to aggluti-ate protease-treated rat, horse, and human O RBC.mmediate comparisons can be made between the twoolecules. They both have a similar molecular weight
nd have similar pI ranges. However, caution should beaken before any conclusions can be drawn between thewo molecules. It is possible that OSLS-II and p35erely share physical characteristics but are function-
lly distinct. Additionally, Vasta et al. (1984) did noterify other proteins in the hemolymph which were notffinity purified and may also share a similar moleculareight. To confirm OSLS-II and p35 as the sameolecule, an N-terminal sequence of OSLS-II wouldave to be determined and compared with that of p35.urther evidence to support homology would be toxpose the serum lectin to P. marinus ECP. Digestion ofSLS-II would support N-terminus results that OSLS-IInd p35 are the same protein and would establish thathe serum lectin is a target of P. marinus proteases.
Confirmation of OSLS-II/p35 homology would havemportant implications in terms of the molecular patho-enesis of P. marinus. It is widely known that lectinslay an important role in invertebrate humoral andellular defense (Arason, 1996). Elimination of anyster defense molecule used in non-self recognition byathogen proteases would represent a pathogenic mech-nism not previously described in the eastern oyster.There is a general dearth of knowledge concerning
rotease inhibitors in the hemolymph of oysters. Previ-us studies on the serological and hemolymph composi-ion of eastern oysters have focused on lysozyme activ-ty, agglutinins, and hemocyte characterization andesponses (Cheng and Rodrick, 1975; Chu and Laeyre, 1989; Li and Flemming, 1967; Chintala et al.,994; Anderson et al., 1995; Anderson, 1996). However,n a recent study Faisal et al. (1998) provided evidencehat C. virginica and C. gigas were capable of producingrotease inhibitors. These findings support our observa-ions that protease inhibitors are present in C. vir-inica, although we could not infer the presence ofrotease inhibitors in C. gigas. Our study does notreclude the possibility that C. gigas can producerotease inhibitors, particularly high-molecular-weightnhibitors. In fact, Faisal et al. (1998) observed muchigher protease inhibitory activity in C. gigas toward P.arinus proteases than C. virginica and speculated
hat this inhibitory activity could be attributed toigh-molecular-weight species, such as a2-macroglobu-
in-like molecules. In addition, previous studies bydham and Faisal (1997) suggested the presence of2-macroglobulin-like activity in C. virginica and C.igas plasmas. This difference in protease inhibitoryctivity between the two species may be important inhe higher susceptibility of C. virginica over C. gigas.
Our findings implicate P. marinus proteases and p35
n the molecular pathogenesis of the disease. It isossible that the p35 target is vitally important to theurvival of the oyster and its elimination may bemportant during infection. Although p35 is not di-ested by P. marinus proteases when in the presence oflasma-based diluent, it is possible that p35 can be-ome susceptible during the progression of the disease.his might occur if the pathogen is capable of compro-ising host anti-proteolytic activity observed herein.he work presented here, however, is reflective of initro conditions only. In vitro experiments have solelyllowed us to identify and partially characterize thearget. Our data suggest a dynamic interplay amongroteases, target proteins, and inhibitors. A fullernderstanding of these potential pathogenic proteasesequires in vivo correlation of target protein loss with P.arinus infections.The experiments performed here are important first
teps in understanding the role of P. marinus proteasesn the pathogenesis of Dermo. It seems unlikely that P.
arinus proteases are the exclusive virulence factor inermo; however, they may very well play a critical role
n disease initiation or progression. The existence ofrotease inhibitors within eastern oysters suggests areviously undescribed defense mechanism.
ACKNOWLEDGMENTS
The authors thank Mr. John Klemer, Ms. Haili Zhang, and Dr.ndrew Dacanay for critical review of the manuscript. Mr. Haroldurrell of VIMS Publications provided assistance with figures. Thisesearch was funded by a grant/cooperative agreement from theational Oceanic and Atmospheric Administration–Virginia Searant Association, Oyster Disease Research Program, Grant No.A56RGO141. The work conducted by J. L. Oliver was further
upported by a National Science Foundation Fellowship. Virginianstitute of Marine Science contribution number 2236.
REFERENCES
dham, K., and Faisal, M. 1997. Involvement of protease inhibitorsin defense mechanisms of oyster: A model to study acquiredimmunodeficiency syndromes. Dev. Comp. Immunol. 21, 101.
nderson, R. S. 1996. Interactions of Perkinsus marinus with hu-moral factors and hemocytes of Crassostrea virginica. J. ShellfishRes. 15, 127–134.nderson, R. S., Burreson, E. M., and Paynter, K. T. 1995. Defenseresponses of hemocytes withdrawn from Crassostrea virginicainfected with Perkinsus marinus. J. Invertebr. Pathol. 66, 82–89.ndrews, J. D. 1988. Epizootiology of the disease caused by the oysterpathogen Perkinsus marinus and its effect on the oyster industry.Am. Fish. Soc. Spec. Publ. 18, 47–63.
rkoosh, M. R., and Kaattari, S. L. 1990. Quantitation of fishantibody to a specific antigen by an enzyme-linked immunosorbentassay (ELISA). In ‘‘Techniques in Fish Immunology’’ (J. S. Stolen,T. C. Fletcher, D. P. Anderson, B. S. Roberson, and W. B. VanMuiswinkel, Eds.), pp. 15–24. SOS Publications, Fair Haven, NJ.rason, G. J. 1996. Lectins as defence molecules in vertebrates andinvertebrates. Fish Shellfish Immunol. 6, 277–289.
rmstrong, P. B., and Quigley, J. P. 1992. Humoral immunity:a2-Macroglobulin activity in the plasma of mollusks. Veliger 35,
161–164.
B
C
C
C
C
C
D
E
F
G
G
H
K
L
L
L
L
L
L
L
M
M
M
P
S
S
T
V
W
183EFFECTS OF P. marinus PROTEASES ON PLASMA PROTEINS
ender, R. C., Fryer, S. E., and Bayne, C. J. 1992. Proteinaseinhibitory activity in the plasma of a mollusc: Evidence for thepresence of a-macroglobulin in Biomphalaria glabrata. Comp.Biochem. Physiol. 102B, 821–824.
alkins, C., and Sloane, B. 1995. Mammalian cysteine proteaseinhibitors: Biochemical properties and possible role in tumourprogression. Biol. Chem. Hoppe-Seyler 376, 71–80.
heng, T. C., and Rodrick, G. E. 1975. Lysosomal and other enzymesin the hemolymph of Crassostrea virginica and Mercenaria merece-naria. Comp. Biochem. Physiol. 52B, 443–447.
hintala, M. M., Ford, S. E., Fisher, W. S., and Ashton-Alcox, K. A.1994. Oyster serum agglutinins and resistance to protozoan para-sites. J. Shellfish Res. 13, 115–121.hu, F. L. 1996. Laboratory investigations of susceptibility, infectiv-ity and transmission of Perkinsus marinus in oysters. J. ShellfishRes. 15, 57–66.
hu, F. L., and La Peyre, J. F. 1989. Effect of environmental factorsand parasitism on hemolymph lysozyme and protein of Americanoysters (Crassostrea virginica). J. Invertebr. Pathol. 54, 224–232.onovan, M. A., and Laue, T. M. 1991. A novel trypsin inhibitor fromthe hemolymph of the horseshoe crab, Limulus polyphemus. J.Biol. Chem. 266, 2121–2125.
llis, A. 1987. Inhibition of the Aeromonas salmonicida extracellularprotease by a2-macroglobulin in the serum of rainbow trout.Microbial Pathogenesis 3, 167–177.
aisal, M., MacIntyre, E. A., Adham, K. G., Tall, B. D., Kothary,M. H., and La Peyre, J. F. 1998. Evidence for the presence ofprotease inhibitors in eastern (Crassostrea virginica) and Pacific(Crassostrea gigas) oysters. Comp. Biochem. Physiol. 121B, 161–168.arreis, K. A., La Peyre, J. F., and Faisal, M. 1996. The effects ofPerkinsus marinus extracellular products and purified proteaseson oyster defence parameters in vitro. Fish Shellfish Immunol. 6,581–597.ilardi, M. L. 1996. Transfer of protein/peptides from SDS–PAGE toPVDF or similar inert nylon membranes. SOP GENPROC 00003.Commonwealth Biotechnologies, Inc. Richmond, VA.arlow, E., and Lane, D. 1988. ‘‘Antibodies: A Laboratory Manual.’’Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.raut, J. 1977. Serine proteases: Structure and mechanism ofcatalysis. Annu. Rev. Biochem. 46, 331–358.
a Peyre, J. F. 1993. ‘‘Studies on the Oyster Pathogen Perkinsusmarinus (Apicomplexa): Interactions with Host Defenses of Cras-sostrea virginica and Crassostrea gigas and in Vitro Propagation.’’Ph.D. dissertation, College of William and Mary, VA.
a Peyre, J. F., Faisal, M., and Burreson, E. M. 1993. In vitropropagation of the protozoan Perkinsus marinus, a pathogen of the
eastern oyster, Crassostrea virginica. J. Euk. Microbiol. 40, 305–310.
a Peyre, J. F., and Faisal, M. 1995. Perkinsus marinus producesextracellular proteolytic factor(s). Bull. Eur. Assoc. Fish Pathol. 15,1–4.
a Peyre, J. F., Schafhauser, D. Y., Rizkalla, E. H., and Faisal, M.1995. Production of serine proteases by the oyster pathogenPerkinsus marinus (Apicomplexa) in vitro. J. Euk. Microbiol. 42,544–551.
a Peyre, J. F., Yarnall, H. A., and Faisal, M. 1996. Contribution ofPerkinsus marinus extracellular products in the infection of East-ern oysters (Crassostrea virginica). J. Invertebr. Pathol. 68, 312–313.
evine, N. D. 1978. Perkinsus gen. N. and other new taxa in theprotozoan phylum Apicomplexa. J. Parasitol. 64, 549.
i, M. F., and Fleming, C. 1967. Hemagglutinins from oyster hemo-lymph. Can. J. Zool. 45, 1225–1233.cKerrow, J. H. 1989. Parasite proteases. Exp. Parasitol. 68, 111–115.cKerrow, J. H., Sun, E., Rosenthal, P. J., and Bouvier, J. 1993. Theproteases and pathogenecity of parasitic protozoa. Annu. Rev.Microbiol. 47, 821–853.eyers, J. A., Burreson, E. M., Barber, B. J., and Mann, R. 1991.Susceptibility of diploid and triploid pacific oysters, Crassostreagigas, and eastern oysters, Crassostrea virginica, to Perkinsusmarinus. J. Shellfish Res. 10, 433–437.
aynter, K. T. 1996. The effects of Perkinsus marinus infection onphysiological processes in the eastern oyster, Crassostrea virginica.J. Shellfish Res. 15, 119–125.
humway, S. E. 1996. Natural environmental factors. In ‘‘TheEastern Oyster, Crassostrea virginica’’ (V. S. Kennedy, R. I. E.Newell, and A. F. Eble, Eds.), pp. 581–660. Maryland Sea GrantCollege, College Park, MD.
ousa, M., Moradas-Ferreira, P., and Azevedo, C. 1992. Presence of atrypsin-like protease in starfish sperm acrosome. J. Exp. Zool. 261,349–354.
horgersen, I. B., Salvesan, G., Brucato, F. H., Pizzo, S. V., andEnghlid, J. J. 1992. Purification and characterization of andalpha-macroglobulin proteinase inhibitor from the mollusc Octo-pus vulgaris. Biochem. J. 285 (Part 2), 521–527.
asta, G. R., Cheng, T. C., and Marchalonis, J. J. 1984. A lectin on thehemocyte membrane of the oyster (Crassostrea virginica). Cell.Immunol. 99, 475–488.iedow, O., Harder, J., Bartels, J., Streit, V., and Christophers, E.1998. Antileukoprotease in human skin: An antibiotic peptideconstitutively produced by keratinocytes. Biochem. Biophys. Res.Commun. 248, 904–909.
Copyright © 2022 FDOKUMEN




















