
Physiological and pathological changes in the eastern oyster Crassostrea virginica infested with the...
13
1 3 Physiological and pathological changes in the eastern oyster Crassostrea 4 virginica infested with the trematode Bucephalus sp. and exposed 5 to the toxic dinoflagellate Alexandrium fundyense 6 7 8 Malwenn Lassudrie a,⇑ , Gary H. Wikfors b , Inke Sunila c , Jennifer H. Alix b , Mark S. Dixon b , 9 Doriane Combot a , Philippe Soudant a , Caroline Fabioux a , Hélène Hégaret a 10 a Laboratoire des Sciences de l’Environnement Marin (LEMAR), Institut Universitaire Européen de la Mer (IUEM), UBO/CNRS/IRD/IFREMER, rue Dumont d’Urville, 11 technopôle Brest-Iroise, 29280 Plouzané, France 12 b Northeast Fisheries Science Center, NOAA National Marine Fisheries Service, 212 Rogers Avenue, Milford, CT 06460, USA 13 c State of Connecticut, Department of Agriculture, Bureau of Aquaculture, P.O. Box 97, Milford, CT 06460, USA 14 15 16 18 article info 19 Article history: 20 Received 27 June 2014 21 Revised 21 January 2015 22 Accepted 29 January 2015 23 Available online xxxx 24 Keywords: 25 Crassostrea virginica 26 Alexandrium fundyense 27 Bucephalus sp. 28 Perkinsus marinus 29 Host–pathogen interaction 30 31 abstract 32 Effects of experimental exposure to Alexandrium fundyense, a Paralytic Shellfish Toxin (PST) producer 33 known to affect bivalve physiological condition, upon eastern oysters, Crassostrea virginica with a variable 34 natural infestation of the digenetic trematode Bucephalus sp. were determined. 35 After a three-week exposure to cultured A. fundyense or to a control algal treatment with a non-toxic 36 dinoflagellate, adult oysters were assessed for a suite of variables: histopathological condition, hemato- 37 logical variables (total and differential hemocyte counts, morphology), hemocyte functions (Reactive 38 Oxygen Species (ROS) production and mitochondrial membrane potential), and expression in gills of 39 genes involved in immune responses and cellular protection (MnSOD, CAT, GPX, MT-IV, galectin CvGal) 40 or suspected to be (Dominin, Segon). 41 By comparing individual oysters infested heavily with Bucephalus sp. and uninfested individuals, we 42 found altered gonad and digestive gland tissue and an inflammatory response (increased hemocyte 43 concentration in circulating hemolymph and hemocyte infiltrations in tissues) associated with trematode 44 infestation. Exposure to A. fundyense led to a higher weighted prevalence of infection by the protozoan 45 parasite Perkinsus marinus, responsible for Dermo disease. Additionally, exposure to A. fundyense in trem- 46 atode-infested oysters was associated with the highest prevalence of P. marinus infection. These observa- 47 tions suggest that the development of P. marinus infection was advanced by A. fundyense exposure, and 48 that, in trematode-infested oysters, P. marinus risk of infection was higher when exposed to A. fundyense. 49 These effects were associated with suppression of the inflammatory response to trematode infestation by 50 A. fundyense exposure. Additionally, the combination of trematode infestation and A. fundyense exposure 51 caused degeneration of adductor muscle fibers, suggesting alteration of valve movements and catch state, 52 which could increase susceptibility to predation. Altogether, these results suggest that exposure of 53 trematode-infested oysters to A. fundyense can lead to overall physiological weakness that decrease 54 oyster defense mechanisms. 55 Ó 2015 Published by Elsevier Inc. 56 57 58 59 1. Introduction 60 The eastern oyster Crassostrea virginica, native to the east coasts 61 of North and Central America, is a major commercial species with 62 144,012 tons produced in 2011 in the USA, which accounted for 63 75% of worldwide production of this species (FAO, 2014). Infectious 64 diseases of eastern oysters caused by protozoan parasites Perkinsus 65 marinus (Dermo), Haplosporidium nelsoni (MSX) and Haplosporidi- 66 um costale (SSO) are responsible for economic loss from mortality 67 during epizootics (Ford and Tripp, 1996). The protozoan parasite 68 P. marinus has been the primary cause of mortality in cultured C. 69 virginica on the east coast of the US (Ford and Smolowitz, 2007). 70 Other parasites also may present a serious risk to the oyster 71 industry. Among these are the digenetic trematodes, referred to 72 as bucephalids, that in advanced infestations affect gonadal folli- 73 cles causing sterility (Lauckner, 1983). Marine bivalves are the first 74 intermediary hosts for bucephalids, fish being the final hosts 75 (Olsen, 1974). http://dx.doi.org/10.1016/j.jip.2015.01.011 0022-2011/Ó 2015 Published by Elsevier Inc. ⇑ Corresponding author. E-mail address: [email protected] (M. Lassudrie). Journal of Invertebrate Pathology xxx (2015) xxx–xxx Contents lists available at ScienceDirect Journal of Invertebrate Pathology journal homepage: www.elsevier.com/locate/jip YJIPA 6639 No. of Pages 13, Model 5G 11 February 2015 Please cite this article in press as: Lassudrie, M., et al. Physiological and pathological changes in the eastern oyster Crassostrea virginica infested with the trematode Bucephalus sp. and exposed to the toxic dinoflagellate Alexandrium fundyense. J. Invertebr. Pathol. (2015), http://dx.doi.org/10.1016/ j.jip.2015.01.011
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Physiological and pathological changes in the eastern oyster Crassostrea virginica infested with the...

1
3
4
5
6
7
8
9
10111213
141516
1 8
1920212223
24252627282930
3 1
5859
60
61
62
63
64
Journal of Invertebrate Pathology xxx (2015) xxx–xxx
YJIPA 6639 No. of Pages 13, Model 5G
11 February 2015
Contents lists available at ScienceDirect
Journal of Invertebrate Pathology
journal homepage: www.elsevier .com/ locate/ j ip
Physiological and pathological changes in the eastern oyster Crassostreavirginica infested with the trematode Bucephalus sp. and exposedto the toxic dinoflagellate Alexandrium fundyense
http://dx.doi.org/10.1016/j.jip.2015.01.0110022-2011/� 2015 Published by Elsevier Inc.
⇑ Corresponding author.E-mail address: [email protected] (M. Lassudrie).
Please cite this article in press as: Lassudrie, M., et al. Physiological and pathological changes in the eastern oyster Crassostrea virginica infested wtrematode Bucephalus sp. and exposed to the toxic dinoflagellate Alexandrium fundyense. J. Invertebr. Pathol. (2015), http://dx.doi.org/1j.jip.2015.01.011
Malwenn Lassudrie a,⇑, Gary H. Wikfors b, Inke Sunila c, Jennifer H. Alix b, Mark S. Dixon b,Doriane Combot a, Philippe Soudant a, Caroline Fabioux a, Hélène Hégaret a
a Laboratoire des Sciences de l’Environnement Marin (LEMAR), Institut Universitaire Européen de la Mer (IUEM), UBO/CNRS/IRD/IFREMER, rue Dumont d’Urville,technopôle Brest-Iroise, 29280 Plouzané, Franceb Northeast Fisheries Science Center, NOAA National Marine Fisheries Service, 212 Rogers Avenue, Milford, CT 06460, USAc State of Connecticut, Department of Agriculture, Bureau of Aquaculture, P.O. Box 97, Milford, CT 06460, USA
a r t i c l e i n f o a b s t r a c t
323334353637383940414243
Article history:Received 27 June 2014Revised 21 January 2015Accepted 29 January 2015Available online xxxx
Keywords:Crassostrea virginicaAlexandrium fundyenseBucephalus sp.Perkinsus marinusHost–pathogen interaction
44454647484950515253545556
Effects of experimental exposure to Alexandrium fundyense, a Paralytic Shellfish Toxin (PST) producerknown to affect bivalve physiological condition, upon eastern oysters, Crassostrea virginica with a variablenatural infestation of the digenetic trematode Bucephalus sp. were determined.
After a three-week exposure to cultured A. fundyense or to a control algal treatment with a non-toxicdinoflagellate, adult oysters were assessed for a suite of variables: histopathological condition, hemato-logical variables (total and differential hemocyte counts, morphology), hemocyte functions (ReactiveOxygen Species (ROS) production and mitochondrial membrane potential), and expression in gills ofgenes involved in immune responses and cellular protection (MnSOD, CAT, GPX, MT-IV, galectin CvGal)or suspected to be (Dominin, Segon).
By comparing individual oysters infested heavily with Bucephalus sp. and uninfested individuals, wefound altered gonad and digestive gland tissue and an inflammatory response (increased hemocyteconcentration in circulating hemolymph and hemocyte infiltrations in tissues) associated with trematodeinfestation. Exposure to A. fundyense led to a higher weighted prevalence of infection by the protozoanparasite Perkinsus marinus, responsible for Dermo disease. Additionally, exposure to A. fundyense in trem-atode-infested oysters was associated with the highest prevalence of P. marinus infection. These observa-tions suggest that the development of P. marinus infection was advanced by A. fundyense exposure, andthat, in trematode-infested oysters, P. marinus risk of infection was higher when exposed to A. fundyense.These effects were associated with suppression of the inflammatory response to trematode infestation byA. fundyense exposure. Additionally, the combination of trematode infestation and A. fundyense exposurecaused degeneration of adductor muscle fibers, suggesting alteration of valve movements and catch state,which could increase susceptibility to predation. Altogether, these results suggest that exposure oftrematode-infested oysters to A. fundyense can lead to overall physiological weakness that decreaseoyster defense mechanisms.
� 2015 Published by Elsevier Inc.
57
65
1. Introduction marinus (Dermo), Haplosporidium nelsoni (MSX) and Haplosporidi- 6667
68
69
70
71
The eastern oyster Crassostrea virginica, native to the east coastsof North and Central America, is a major commercial species with144,012 tons produced in 2011 in the USA, which accounted for75% of worldwide production of this species (FAO, 2014). Infectiousdiseases of eastern oysters caused by protozoan parasites Perkinsus
72
73
74
75
um costale (SSO) are responsible for economic loss from mortalityduring epizootics (Ford and Tripp, 1996). The protozoan parasiteP. marinus has been the primary cause of mortality in cultured C.virginica on the east coast of the US (Ford and Smolowitz, 2007).Other parasites also may present a serious risk to the oysterindustry. Among these are the digenetic trematodes, referred toas bucephalids, that in advanced infestations affect gonadal folli-cles causing sterility (Lauckner, 1983). Marine bivalves are the firstintermediary hosts for bucephalids, fish being the final hosts(Olsen, 1974).
ith the0.1016/

76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
2 M. Lassudrie et al. / Journal of Invertebrate Pathology xxx (2015) xxx–xxx
YJIPA 6639 No. of Pages 13, Model 5G
11 February 2015
Diseases caused by parasites imply an imbalance in the host-parasite relationship; for example, the host immune response isinadequate to control proliferation. In bivalves, internal defenseis based upon an innate immune system composed primarily ofhemocyte-mediated cellular mechanisms and humoral factorssuch as lectins, lysins, and antimicrobial peptides (Cheng, 1996;Hine, 1999; Song et al., 2010; Soudant et al., 2013). The best knownimmune responses to microbial pathogens involve phagocytosis byindividual hemocytes followed by intracellular degradation(Cheng, 1996; Janeway and Medzhitov, 2002; Song et al., 2010;Soudant et al., 2013). Encapsulation by masses of hemocytes isanother mechanism of isolation and destruction of pathogens toolarge for phagocytosis (Cheng, 1981; Feng, 1988; Fisher, 1986,1988). Cellular immune response to bucephalid trematodes inbivalves usually is low or not detected (Cheng and Burton, 1965;da Silva et al., 2002) and consists mainly, when existent, ofhemocyte infiltration (Cheng and Burton, 1965); however, humoralfactors seem also to be involved (Feng and Canzonier, 1970).
The immune systems of marine bivalves may be affected byenvironmental factors, such as harmful algal blooms (HABs)(Hégaret et al., 2011). Toxic dinoflagellates, such as Alexandriumspecies that produce neurotoxins, were reported to decrease phago-cytosis, production of Reactive Oxygen Species (ROS), and adhesionby hemocytes of many bivalve species in in vitro assays (Hégaretet al., 2011; Mello et al., 2013), suggesting a direct immuno-suppressive effect. Such an effect was also hypothesized in vivo afterobservation of decreased total hemocyte count (THC) in the bluemussel Mytilus edulis (Galimany et al., 2008) and increase in hemo-cyte mortality in Crassostrea gigas (at 18 �C) (Hégaret et al., 2007)exposed to Alexandrium spp. A positive effect of A. minutum expo-sure (depending upon the season and the oyster ploidy) or of toxinaccumulation, however, was reported upon THC in C. gigas(Haberkorn et al., 2010a; Hégaret et al., 2007), possibly stimulatingthe immune system. Modifications of cellular pro- and anti-oxidantmechanisms, which may participate in the immune response andare involved in cellular protection, also have been reported(Fabioux et al., in preparation; Lassudrie et al., 2014). Whether ornot these effects upon immune-related variables are direct conse-quences of Alexandrium sp. exposure could not be assessed, becauseAlexandrium spp. also affect general physiological processes (Cucciet al., 1985; Gainey and Shumway, 1988a, 1988b; Landsberg,2002; Shumway, 1990) and tissue integrity (Galimany et al.,2008; Haberkorn et al., 2010b; Lassudrie et al., 2014; Medhioubet al., 2012), that could indirectly affect hemocytes.
A hypothesis is that toxic dinoflagellate exposure could increasebivalve susceptibility to disease by altering both defense andphysiological condition. HAB intensity and frequency are reportedto be increasing (Van Dolah, 2000), and HAB co-occurrence withpathogen infections in marine bivalves is recurrent. Only fewstudies, however, have reported the effects of these interactionswhich could lead to modified host–pathogen relationships(Bricelj et al., 2011; da Silva et al., 2008; Hégaret et al., 2009).One such interaction was reported by Bricelj et al. (2011), whoshowed that exposure to Alexandrium tamarense increased mortal-ity of juvenile clams challenged with Vibrio tapetis, the causativeagent of Brown Ring Disease.
The present study was undertaken to investigate possibleinteractions between parasitic diseases and toxic Alexandrium incompromising the health of the eastern oyster C. virginica. Thisstudy was conducted in spring, when Alexandrium fundyenseblooms may occur along the northeastern coast of the USA(Anderson et al., 2005; Hattenrath et al., 2010; Lopez et al., 2014)and when P. marinus intensity and Haplosporidium spp. prevalenceand intensity in oysters are at the lowest (Ford and Smolowitz,2007; Ford, 1985). Experimental oysters were found to containhigh infestations of the trematode Bucephalus sp. We hypothesized
Please cite this article in press as: Lassudrie, M., et al. Physiological and patholtrematode Bucephalus sp. and exposed to the toxic dinoflagellate Alexandj.jip.2015.01.011
that: (1) oyster exposure to A. fundyense could modify susceptibil-ity to parasitic infestations, and (2) the interaction of A. fundyenseand parasites could lead to combined effects upon oyster physio-logical condition. To test these hypotheses, we examined parasiticinfestations and histopathological changes in oysters exposed tocultured A. fundyense. In addition, we assessed immune status bydetermining hematology (total and differential hemocyte counts)and hemocyte functions (ROS production and mitochondrialmembrane potential), as well as gene expression of sequencescoding for proteins associated with immune responses andcellular protection: a galectin, CvGal, involved in recognition ofmicro-organisms prior to phagocytosis and also implicated inproliferation of P. marinus (Tasumi and Vasta, 2007); the twonewly-described C. virginica major plasma proteins Dominin (Itohet al., 2011) and Segon (Xue et al., 2012), for which specific rolesare still under investigation; antioxidant enzymes (MnSOD, CAT,GPX); and a metallothionein family (MT-IV) that responded toimmune challenge (Jenny et al., 2006; Wang et al., 2010) andwas isolated from hemocytes (Jenny et al., 2006). Gene expressionanalysis was conducted in the gill, an organ rich in hemocytes andin direct contact with the external milieu.
2. Materials and methods
2.1. Experimental oysters
Eastern oysters C. virginica of 9.3 ± 0.1 cm length (mean ± SE)were collected from Milford Harbor (CT, USA) on April 20th,2012, and placed the same day in tanks with running, unfilteredseawater.
2.2. Characteristics of the seawater supplied
Throughout the experiment, flow of 30–40 L h�1 of unfilteredseawater (containing natural phytoplankton) from Milford Harborwas distributed to the tanks. Temperature of the seawater was notcontrolled to provide the natural temperature of the local oysterhabitat and varied from 10.5 to 15.8 �C during the 23 days of theexperiment (12.9 ± 0.4 �C, mean ± SE). Salinity was 24.2 ± 0.5 ppt(mean ± SE) over the entire experiment. Light cycle in experimen-tal facilities was controlled, with L:D = 14:10, correspondingroughly to the natural cycle when the experiment was conducted.
Biodiversity of phytoplankton in the seawater supplied to thetanks was surveyed during the experiment. 150 mL was sampledevery 3 days and fixed with 3% Lugol’s solution. Sedimentationcolumns were used to concentrate the samples, and slides wereobserved under an inverted light microscope. Two diatoms,Skeletonema sp. and Thalassionema sp., dominated the phytoplank-ton community during the experiment, and no harmful algalspecies were detected.
Total particulate matter (TPM) and particulate organic matter(POM) in the water supplied to the tanks was quantified every4 days by filtering 300 mL in triplicate through Whatman GF/C fil-ters following the procedure described by Galimany et al. (2011).Briefly, TPM was determined by weighing dried filters (48 h at60 �C). Particulate inorganic matter (PIM) then was obtained byweighing ashed filters (4 h at 450 �C), and POM was calculated asthe difference between POM and TPM. TPM in the seawater sup-plied to the tanks was 13 ± 1 mg L�1 (mean ± SE) during the exper-iment, with POM accounting for 20 ± 1% of the TPM (mean ± SE).
2.3. Algal culture
The non-toxic dinoflagellate Scrippsiella lachrymosa (strainCCMP2666) was used as the control alga for the experiment, and
ogical changes in the eastern oyster Crassostrea virginica infested with therium fundyense. J. Invertebr. Pathol. (2015), http://dx.doi.org/10.1016/

200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242243
245245
246
247
248
249
250
251
252
253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269
270
271
272
273
274
275
276
277
278
279
280
281
282
283
284
285
286
287
288
289
290
291
292
293
294
295
296
297
298
299
300
301
302
303
304
M. Lassudrie et al. / Journal of Invertebrate Pathology xxx (2015) xxx–xxx 3
YJIPA 6639 No. of Pages 13, Model 5G
11 February 2015
the toxic dinoflagellate A. fundyense (BF2 strain, isolated from theGulf of Maine, USA) was used for the toxic-algal exposure. Bothdinoflagellate species were cultured at 20 �C with 24 h light usingf/2-enriched filtered seawater from Milford Harbor (Guillard andRyther, 1962) in 20-L glass carboy assemblies using aseptic tech-nique (Ukeles, 1973). To maintain consistency in culture qualityduring the study, cultures were harvested semi-continuously inlate-log or early-stationary phase. The Paralytic Shellfish Toxins(PSTs) produced by the BF2 strain of A. fundyense, cultured and har-vested under these conditions, were previously reported to occurin the following mass proportions: GTX1,4 17.4%; NEO 29.0%;GTX2,3 5.4%; B1 6.2%; STX 6.6%; and C1,2 35.3% (Galimany et al.,2008). Cultured algal cell densities were determined by flowcytometry (FACScan, BD BioSciences, San Jose, CA, USA).
2.4. Experimental design and sampling
170 oysters were distributed haphazardly into ten 18-L tanks(17 oysters per tank). Oysters were acclimated for 3 days to labora-tory conditions with no addition of algal culture before beingexposed for 23 days (April 23rd to May 17th, 2012) to the two algaltreatments, S. lachrymosa or A. fundyense (5 replicates each). Fourpulses of algal culture per day were provided to each tank tosimulate tidal delivery. Each pulse of algal culture yielded a finalcell count in each tank of 103 cells mL�1 for the two first weeksand 2 � 103 cells mL�1 the last week (9–18 � 106 cells per oyster,per day after the T0 sampling). Water flow was stopped for 1 h fol-lowing algal distribution to allow feeding and prevent immediatewashout of the algae. This procedure added the dinoflagellates tothe existing, background plankton community described above.
80 oysters (8 oysters per tank) were sampled individually at theend of the acclimation period, prior to dinoflagellate addition (T0)and after 23 days exposure (T23). Hemolymph was withdrawn fromthe adductor muscle of each oyster with a syringe before oysterswere shucked, and soft tissues were weighed. Next, the anal-rectalregion was excised for P. marinus quantification, and a tissue cross-section and a transversal section were fixed for histological analy-sis. Remaining digestive gland and remaining gills were dissected,individually frozen immediately in liquid N2, and stored at �80 �Cfor toxin quantification and gene expression analysis, respectively.Oysters at T0 were analyzed for P. marinus only.
2.5. Condition index
Condition index (CI) was determined for 77 oysters at eachsampling time, T0 and T23, as follows (adapted from Bodoy et al.,1986):
CI ¼ wet flesh weightheight�width
� 100
305
Table 1Semi-quantitative scale categorizing intensity of histopathological conditionsobserved (from Lassudrie et al., 2014).
Levelintensity
Occurrence of the pathological condition in the examined tissuearea
0.0 Absence0.5 Very low (1–5 total occurrence)1.0 Low (>5 occurrence/presence in all fields at magnification 10�)1.5 Moderate (presence in all fields at magnification 20�/covering
about 10% of the tissue)2.0 High (presence in all fields at magnification 40�/covering about
20% of the tissue)2.5 Very high (presence in all fields at magnification 60�/covering
about 30% or above of the tissue)
2.6. Analysis of hemocyte variables
77 oysters sampled at T23 were analyzed for hemocyte vari-ables. Hemolymph withdrawn from adductor muscle was storedtemporarily in Eppendorf microcentrifuge tubes held on ice beforeflow-cytometric analysis. Characteristics determined in live, circu-lating hemocytes – total (THC) and differential hemocyte counts(granulocytes and hyalinocytes) (in cell mL�1), size, and internalcomplexity (in arbitrary units a.u.), as well as mortality (percent-age of dead hemocytes) – were assessed following Hégaret et al.(2003). Functional responses, i.e. production of Reactive OxygenSpecies (ROS) (specifically H2O2 and O2
� �) by unstimulatedhemocytes and mitochondrial membrane potential (MMP) weredetermined as described in Delaporte et al. (2003) and Donaghy
Please cite this article in press as: Lassudrie, M., et al. Physiological and patholtrematode Bucephalus sp. and exposed to the toxic dinoflagellate Alexandj.jip.2015.01.011
et al. (2012), respectively. Hemocyte analyses were performedwith a FACScan flow-cytometer, and data were processed usingWinMDI 2.8.
2.7. Histopathological observations
54 oysters sampled at T23 were analyzed for histopathology. A5-mm cross section of soft tissues including gills, mantle, gonadand digestive gland, and a transversal section including adductormuscle, heart and kidney, were excised and fixed in Davidson’s fix-ative for 48 h at 4 �C (Shaw and Battle, 1957). Tissue sections thenwere rinsed in 50% ethanol in filtered seawater and transferred to70% ethanol, dehydrated in ascending ethanol solutions, clearedwith xylene, and embedded in paraffin wax. Six-lm sections werestained with Harris’ hematoxylin and eosin (Howard et al., 2004)and read blind under a light microscope.
Histopathological condition was examined per tissue. Intensityof each histopathological observation was rated using a six-levelsemi-quantitative scale ranging from 0 to 2.5 as described inTable 1, according to Lassudrie et al. (2014). Based upon this scale,mean weighted prevalences of each histopathological conditionwere calculated as the mean intensities of each histopathologicalcondition of all individuals.
In addition, the sum of mean weighted prevalences of eachhistopathological condition was calculated in each organ.
Gonadal maturation stage was determined following an 8-level,semi-quantitative scale (1: sexually undifferentiated, to 8:spawned) adapted from Ford and Figueras (1988) by Powell et al.(1993). Mean maturation stage was calculated and used forstatistical analysis.
2.8. Gene expression analysis in gills
77 oysters sampled at T23 were analyzed for gene expressionfollowing the procedure described by Béguel et al. (2013) andcomplying with Bustin et al. (2009) recommendations. Briefly,RNA was extracted from the gills and DNase-treated. The quantityand quality of total RNA were checked before reverse-transcrip-tion. The transcript levels of targeted genes were determined byqPCR relative to a constitutively-expressed reference gene, the18S rRNA (GenBank accession AB199895.1). Primers for 18S ribo-somal, Dominin (AB199895.1), Segon (JQ235755.1), MnSOD(EH646608.1), CAT (CV087224.1), GPX (CD650160.1) and all theknown MT-IV subtypes, A, B and C (DQ117912.1, DQ117913.1,DQ117914.1) were picked from the literature (Brulle et al., 2012;Itoh et al., 2011; Xue et al., 2012), or designed using Primer 3web v. 4.0.0 (Koressaar and Remm, 2007; Untergasser et al.,2012) from nucleotide sequences published in GenBank forCvGal (DQ779197.1) (forward: 50-GAACCGTGTAATGGCGAAGT-30,reverse: 50-TACATTCCCGAGGTCCAGTC-30). When non-annotatedsequences from C. virginica were used, gene identification was
ogical changes in the eastern oyster Crassostrea virginica infested with therium fundyense. J. Invertebr. Pathol. (2015), http://dx.doi.org/10.1016/

306
307
308
309
310
311
312
313
314
315
316
317
318
319
320
321
322
323
324
325
326
327
328
329
330
331
332
333
334
335
336
337
338
339
340
341
342
343
344
345
346
347
348
349
350
351
352
353
354
355
356
357
358359
360
361
362
363
364
365
366
367
368
369
370
371
372
373
374
375
376
377
378
379
380
381
382
383
384
385
386
387
388
389
390
391
392
393
394
395
396
397
398
399
400
401
402
403
404
405
406
407
408
409
410
411
412
413
414
415
416
417
418
419
420
421
422
423
424
425
426
427
428
4 M. Lassudrie et al. / Journal of Invertebrate Pathology xxx (2015) xxx–xxx
YJIPA 6639 No. of Pages 13, Model 5G
11 February 2015
performed with BLASTX (Altschul et al., 1997), accepting the resultshowing the lowest E-value. Primer pairs amplified a singlesequence and sequencing of each amplicon (performed by GATCBiotech) ensured they corresponded to the expected nucleotidesequence. No Perkinsea and Bucephalidea nucleotide sequenceswas amplified according to sequence comparison in GenBankdatabase using BLASTN (Altschul et al., 1997). PCR efficiency ofall primer pairs, calculated based upon amplification of a standardcurve from a serial dilution of control cDNA, was constrainedbetween 95% and 105%. The relative expression ratio (R) of eachtarget gene was calculated following the Pfaffl (2001) formula.
2.9. Parasitic infestations
77 oysters for each sampling time, T0 and T23, were analyzed forP. marinus. A sample of the anal-rectal region from each oyster wasincubated for 8 days in the dark at room temperature in Ray’s FluidThioglycollate Medium (RFTM) as modified by Bushek et al. (1994).Hypnospores of P. marinus in tissue squashes then were stained withLugol’s solution and examined under the light microscope. Intensityof infection was determined using the Mackin scale (Mackin, 1962)(0: no parasite detected, to 5: heavy infection). Prevalence of infec-tion, mean weighted prevalence (intensity of the infection/totalnumber of oyster examined), and mean intensity (intensity of theinfection/number of infected oysters) were calculated.
Prevalence (n = 77) and intensity (n = 10) of trematode Buceph-alus sp. infestation were assessed by histological observations atT23. Intensity of infestation was determined as the mean percent-age of the area of Bucephalus sp. sporocyst occupation calculatedfrom 10 random fields per oyster (including gills, digestive gland,gonad and mantle) at total magnification 150�, using Image Jsoftware.
Other parasites were detected by examination of histologicalslides on 54 oysters at T23. Prevalence of Haplosporidium spp. wasdetermined, and weighted prevalence and intensity of the infectionwere assessed using a semi-quantitative scale followed by acomposite rating matrix as published by Kim et al. (2006) for afinal rating on a 4-level scale. Intensity of infestation with otherparasites was rated on the six-level semi-quantitative scale rang-ing from 0 to 2.5 described in Table 1. Mean weighted prevalenceof each infestation was calculated (intensity of the infestation/number of oyster examined) and used for statistical analysis.
2.10. Toxin detection
77 oysters sampled at T23 were analyzed for toxins. Digestivegland tissue was ground using a ‘‘Dangoumeau’’-like ball grinderand frozen in liquid nitrogen. Paralytic Shellfish Toxin (PST) accu-mulation in the digestive gland was estimated using a PSP ELISAkit (Abraxis), after being extracted from ground digestive glandin HCl 0.1 M (1:1, w:v) and boiled for 5 min, according to manufac-turer instructions, leading to acid hydrolysis that can induce chem-ical conversion of some PST analogues to STX (Vale et al., 2008).This ELISA assay recognizes mostly STX, and other PSTs only tovarying degrees (cross-reactivities of 100% for STX and from 29%to 0% for other PSTs). Consequently, toxicity was expressed aslg STX kg�1 of wet digestive gland weight. The principle of thisassay is different from bioassay or HPLC methods; therefore,results obtained with the Abraxis PSP ELISA kit should not becompared directly to toxicity obtained with other methods.
2.11. Statistical analysis
Considering the low number of oysters infected with Haplospo-ridium spp., no statistical analyses was performed to assess effectof this infection upon pathological and physiological variables.
Please cite this article in press as: Lassudrie, M., et al. Physiological and patholtrematode Bucephalus sp. and exposed to the toxic dinoflagellate Alexandj.jip.2015.01.011
Effects of algal treatment and trematode infestation were testedupon prevalence of P. marinus with non-parametric, Chi-squaretests performed independently in each Bucephalus sp.-infested orBucephalus sp.-uninfested group, to assess effect of A. fundyenseexposure. Differences in P. marinus infection weighted prevalencewere assessed using non-parametric tests: the Mann–Whitney test(effect of algal treatment or trematode infestation, independently)and the Kruskal–Wallis test (non-nested – comparison of fourgroups categorized by both algal treatment and trematode infesta-tion) followed by the Nemenyi–Damico–Wolfe–Dunn (NDWD)post hoc test, to detect differences between the four groups definedaccording to algal exposure and trematode status.
Effects of Bucephalus sp. and P. marinus infestation status uponPST accumulation were tested with a two-way ANOVA (the factorswere infestation with Bucephalus sp. and with P. marinus), andeffect of the intensity level of each infection upon PST accumula-tion was tested using the Kruskal–Wallis test.
Two-way ANOVA was used to analyze effects of trematodeinfestation, algal treatment, and the interaction, and effects ofP. marinus infection, algal treatment, and the interaction, uponhemocyte variables and gene expression, followed by the LeastSignificant Difference (LSD) post hoc test, to detect differencesbetween the four groups defined according to algal exposure andtrematode status, or algal exposure and P. marinus status.
For histopathological changes and other parasitic infestations,the Mann–Whitney test was used to assess the effect of algaltreatment, Bucephalus sp., and P. marinus infestation individually,and the Kruskal–Wallis test followed by the NDWD post hoc test,to detect differences between the four groups defined accordingto algal exposure and trematode status, or algal exposure andP. marinus status.
Data were log or 1/X transformed, when needed, to meetnormality and homoscedasticity assumptions, and percentage datawere transformed as Arc-sin of square root. Differences wereconsidered significant when p < 0.05. Statistics were performedwith Statgraphics Plus statistical software (Manugistics, Inc.,Rockville, MD, USA) and R version 2.15.1 (R Core Team, 2012).
3. Results
3.1. Parasitic infestations
Infestation with the trematode Bucephalus sp. was detected inhistological sections. The microscopic presentation of Bucephalussp. in the tissues consisted of sporocysts containing germ ballsand different developmental stages of cercariae (Fig. 2A, B, E).Although trematode sporocyst branches were present mainly inthe gonad, they could be observed in connective tissues in allorgans and in hemolymph vessels and were sometimes observedbreaking the hemolymph-vessel walls. After 23 days of algalexposure, 34% of experimental oysters were infested with thetrematode Bucephalus sp. (n = 77). Prevalence was not significantlydifferent at T23 between the A. fundyense-treated group (29%;n = 38) and the S. lachrymosa-treated group (38%; n = 39). Trema-tode sporocyst occupation accounted for 22 ± 2% of the tissues inoysters (mean ± SE, n = 10 oysters), and no significant differencebetween algal treatments was detected.
Prevalence of infection with P. marinus in oysters at the begin-ning of the experiment was 65% (n = 77) and decreased signifi-cantly to 38% after 23 days exposure to the control dinoflagellate(p < 0.01) (n = 39), whereas, it remained at 61% in A. fundyense-exposed oysters (n = 38) (Fig. 1A), but the difference between algaltreatments at T23 was not significant.
At the beginning of the experiment, P. marinus weighted preva-lence was 0.7 ± 0.1 (mean ± SE; n = 77) (Fig. 1A), and decreasedsignificantly after 23 days in S. lachrymosa-exposed oysters
ogical changes in the eastern oyster Crassostrea virginica infested with therium fundyense. J. Invertebr. Pathol. (2015), http://dx.doi.org/10.1016/

429
430
431
432
433
434
435
436
437
438
439
440
441
442
443
444
A B
Fig. 1. (A) Prevalence (bars) and weighted prevalence (lines) (mean ± SE) of Perkinsus marinus in oysters Crassostrea virginica at the beginning of the experiment (T0) and after23 days (T23) of algal exposure to the non-toxic Scrippsiella lachrymosa (dashed line and gray bar) or toxic Alexandrium fundyense (solid line and black bar). Letters indicatesignificant differences in prevalence (upper case; chi-square test, p < 0.01) and in weighted prevalence (lower case; Kruskal–Wallis test followed by NDWD post hoc test,comparing the four groups, p < 0.05). At T0, n = 77; at T23, n = 38–39 per group. (B) Prevalence of infection with P. marinus in oysters sampled at T23 according to algaltreatment and infestation with the trematode Bucephalus sp. ⁄ indicates significant difference among infested oysters between the two groups (chi-square test, p < 0.05). Nper group is indicated inside each bar.
M. Lassudrie et al. / Journal of Invertebrate Pathology xxx (2015) xxx–xxx 5
YJIPA 6639 No. of Pages 13, Model 5G
11 February 2015
(p < 0.05) to 0.4 ± 0.1 (mean ± SE; n = 39). In A. fundyense-exposedoysters, weighted prevalence of P. marinus after 23 days exposure,however, did not vary significantly compared to day 0, remainingat 0.7 ± 0.1 (mean ± SE; n = 38), and was thus significantly highercompared to S. lachrymosa-exposed oysters (p < 0.05).
Trematode infestation (not determined at day 0) had no signif-icant effect upon prevalence and weighted prevalence of P. marinusat T23.
s
sss
s *
A
D
G H
*
50 µm
50 µm
25 µm
B
E
c c
h
hh
d
d*
*
s
sl
s
F
G
s
hG
s
F
h
s
sl
c
s
Fig. 2. Histopathological conditions in experimental oysters Crassostrea virginica inducexposure for 23 days. (A) Bucephalus sp. sporocysts (s) in gill intraplical sinus inducing disassociated with hemocyte infiltration (⁄) and hemocytes (h) in gonaducts (G) and folliclfollicles (F) and visible in gonoducts (G); (D) heavy hemocyte infiltration (⁄) associated wduct and tubule degeneration in a digestive gland infested with Bucephalus sp. sporocystsand hemocyte infiltration (⁄) and presence of ceroid bodies (c); (F) normal digestive ductducts and undergoing diapedesis (h) associated with ceroid bodies (c) in the digestive glanBucephalus sp.-infested oyster exposed to A. fundyense; (I) normal adductor muscle.
Please cite this article in press as: Lassudrie, M., et al. Physiological and patholtrematode Bucephalus sp. and exposed to the toxic dinoflagellate Alexandj.jip.2015.01.011
A significant effect of algal exposure was found upon prevalenceof P. marinus among Bucephalus sp.-infested oysters at T23. Indeed,82% of Bucephalus sp.-infested oysters exposed to A. fundyense wereinfected with P. marinus (n = 11), which was significantly highercompared to prevalence of Bucephalus sp.-infested oysters exposedto the control dinoflagellate (40%; n = 15) (p < 0.05) (Fig. 1B). Such adifference was not detected within oysters uninfested withBucephalus sp. (45% of P. marinus prevalence).
G
a
25 µm 25 µm
50 µm 50 µm
C
F
Id
d
d
F
G
F
F
F
F
* *
h
G
s
F
100 µm100 µm
sl
ed by the trematode Bucephalus sp. or its interaction with Alexandrium fundyenseruption of normal gill structure; (B) gonad infested with Bucephalus sp. sporocysts (s)es (F) lumina; (C) normal gonad of an uninfested oyster, with gametes maturing inith destruction of Bucephalus sp. sporocysts (s) in the digestive gland; (E) digestive(s): epithelium atrophy (a) associated with epithelium sloughing into the lumen (sl)s and tubules in an uninfested oyster; (G) hemocyte infiltrated (⁄) around digestived of Bucephalus sp.-infested oysters; (H) degenerated adductor muscle fibers (d) in a
ogical changes in the eastern oyster Crassostrea virginica infested with therium fundyense. J. Invertebr. Pathol. (2015), http://dx.doi.org/10.1016/

445
446
447
448
449
450
451
452
453
454
455
456
457
458
459
460
461
462
463
464
465
466
467
468
469
470
471
472
473
474
475
476
477
478
479
480
481
482
483
484
485
486
487
488
489
490
491
492
493
494
495
496
497
498
499
500
501
502
503
504
505
506
507
508
509
510
511
512
513
514
515
516
517
518
519
520
521
522
523
524
525
526
527
528
529
530
531
532
533
534
535
536
537
538
539
540
541
542
543
544
545
546
547
548
549
550
551
552
553
554
555
556
557
558
559
560
561
562
563
564
565
566
6 M. Lassudrie et al. / Journal of Invertebrate Pathology xxx (2015) xxx–xxx
YJIPA 6639 No. of Pages 13, Model 5G
11 February 2015
Intensity of P. marinus infection (calculated for infected oysters)was 1.02 ± 0.08 (mean ± SE; n = 88) and did not vary significantlyduring the experiment or with algal treatment or trematode status.
Haplosporidium spp. was observed in 9% of the examined oys-ters (5 of 54 oysters), at intensities ranging from 1 (4 oysters) to3 (1 oyster) and with a weighted prevalence of 0.13/4. Other micro-organisms and parasites were observed on histological slides:Nematopsis ostrearum oocysts in all examined organs, RLO (Rickett-sia-like organisms) in digestive tract epithelium, platyhelmintheTurbellaria sp. in intestinal lumen, and ciliates attached to the epi-thelia of pericardium, gills, mantle or in digestive tract lumina. Allof these organisms were observed at very low intensities and prev-alence: mean weighted prevalence per condition was less than 0.6/2.5, and no statistical difference was detected attributable to algalexposure, Bucephalus sp. or P. marinus infection (n = 54 oysters).
3.2. PST accumulation in the digestive gland
PST accumulation, as measured with the Abraxis ELISA assay,was 1,030 ± 124 lg STX kg�1 of wet digestive gland (mean ± SE,n = 38) in A. fundyense-exposed oysters after 23 days of exposure;no PST was detected in the control group. No significant differencein PST measurement was detected according to Bucephalus sp. orP. marinus infection status or level.
3.3. Condition index
No significant effect upon condition index after 23 days ofexperimental exposure was detected according to algal treatmentor P. marinus infection; whereas, infestation with the trematodeBucephalus sp. was associated with a significantly higher(p < 0.01) condition index (0.28 ± 0.01, mean ± SE; n = 26)compared to uninfested oysters (0.24 ± 0.01, mean ± SE; n = 51).
3.4. Histopathology
Statistical differences related to A. fundyense exposure, trema-tode Bucephalus sp., and the algal treatment – Bucephalus sp. inter-action, P. marinus, and the algal treatment – P. marinus interaction,upon each histopathological observation in experimental oystersafter 23 days of algal exposure, are presented in Table 2.
Intact and partially-lysed cells of A. fundyense or S. lachrymosaand of other algae from the seawater supplied were observed inintestine lumina of the oysters, regardless of trematode- or P. mari-nus-status.
Growth of Bucephalus sp. sporocysts in intraplical sinuses andconnective tissue of gills caused disruption of the normal configu-ration of gill filaments in all infested oysters (Fig. 2A). In addition,in the digestive gland hemolymph sinus of one oyster (of 54oysters analyzed), heavy focal infiltration with hemocytes wasobserved around the trematode sporocysts, associated with tissuedestruction (Fig. 2D).
Oysters infested with the trematode Bucephalus sp. had moreintense hemocyte infiltration in the mantle (p < 0.001), gills(p < 0.001), gonads (p < 0.001) (Fig. 2B, C) and digestive gland(p < 0.001), compared to oysters with no visible trematodes.Bucephalus sp. infestation also caused higher weighted prevalenceof ceroid bodies in gonadal tissues (p < 0.01) and in digestive-glandconnective tissue (p < 0.01) (Fig. 2E, G). In the digestive gland,trematode infestation was associated also with increased epithe-lium sloughing into digestive ducts (p < 0.05) and tubules(p < 0.001) (Fig. 2E, F), mucus in digestive duct lumina(p < 0.001), and hemocytes in digestive-tubule lumina (p < 0.05).Atrophy of the digestive-tubule epithelium (Fig. 2E, F) was associ-ated with both trematode infestation (p < 0.01) and P. marinusinfection (p < 0.05).
Please cite this article in press as: Lassudrie, M., et al. Physiological and patholtrematode Bucephalus sp. and exposed to the toxic dinoflagellate Alexandj.jip.2015.01.011
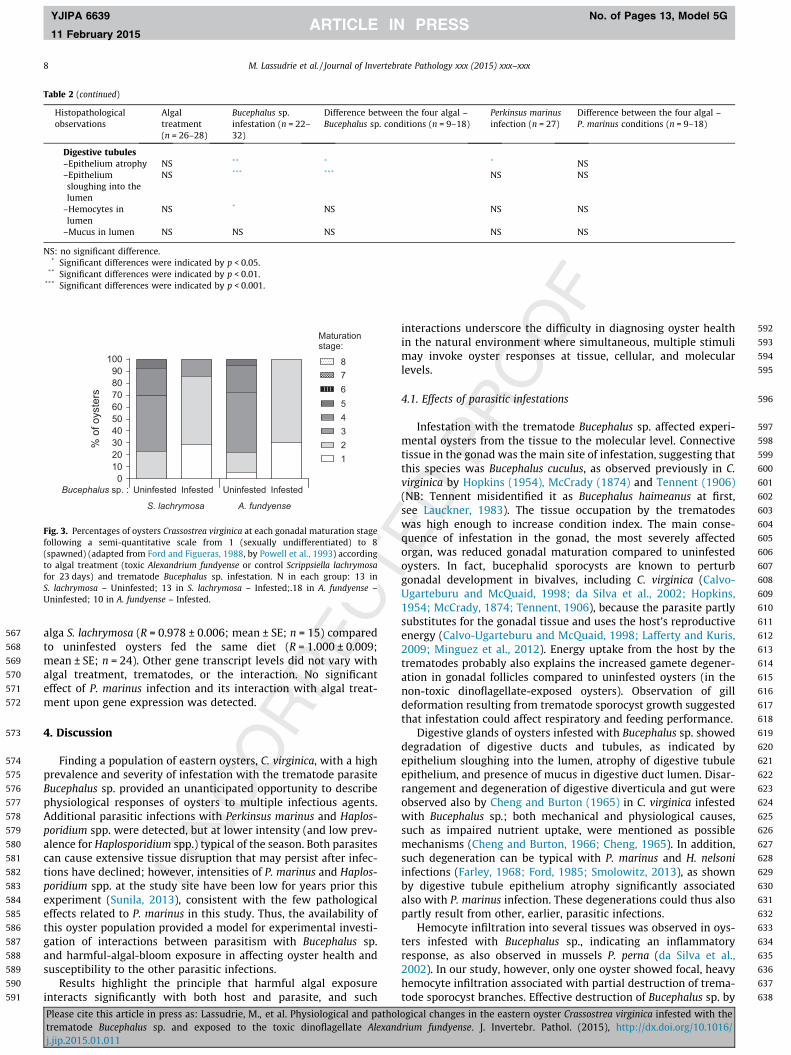
A reduction in the average gonadal maturation stage (p < 0.001)was detected in Bucephalus sp.-infested oysters (Fig. 2B, C, andFig. 3), with a mean maturation stage of 3.0 ± 0.2 in Bucephalussp.-uninfested oysters, corresponding to a mid-gonadal develop-ment stage, vs. 1.8 ± 0.1 in infested oysters, corresponding to anearly developmental stage.
Some histopathological conditions were observed more inten-sely in response to trematode infestation, but only in oystersexposed to the control alga S. lachrymosa: hemocyte infiltrationaround digestive ducts and tubules (p < 0.05) (Fig. 2E, F, G, andFig. 4B) or within gonoduct and follicle lumina (p < 0.05) (Fig. 2B,C, and Fig. 4D), and degeneration of gametes within follicles(p < 0.01) (Fig. 2B, C, and Fig. 4E).
In the kidney, hemocyte infiltration was lower in control oys-ters (exposed to S. lachrymosa and without trematodes) than inoysters exposed to A. fundyense or with Bucephalus sp. (p < 0.01)(Fig. 4C).
Trematode infestation (p < 0.01), P. marinus infection (p < 0.05)and A. fundyense-exposure (p < 0.05) led to higher weighted preva-lence of hemocyte diapedesis in digestive ducts (Fig. 2G). As aconsequence, trematode-infested oysters exposed to A. fundyensehad the highest weighted prevalence of hemocyte diapedesis indigestive ducts (p < 0.05) (Fig. 4A) compared to the other algaltreatment – Bucephalus sp. groups. The same result was observedin P. marinus-infected oysters exposed to A. fundyense comparedto the other algal treatment – P. marinus status groups (p < 0.05).
Degeneration of muscle fibers in the adductor muscle was sig-nificantly more intense in trematode-infested oysters also exposedto A. fundyense (p < 0.01) than in oysters in other groups (Fig. 2H, Iand Fig. 4F).
Other histopathological conditions observed did not vary signif-icantly with algal treatment, trematode or P. marinus status(Table 2).
The sum of the mean weighted prevalence for each type ofhemocyte response (infiltration, diapedesis, presence in lumina),per organ, is presented in Fig. 5A. The following general patternis apparent: A. fundyense exposure, and even more so Bucephalussp. infestation, increased hemocyte responses. Hemocyteresponses to the interaction, however, were lower compared tothe response to Bucephalus sp. alone.
The global pattern for ceroidosis in each organ is presented inFig. 5B as the sum of the mean, weighted prevalence per organ,illustrating that trematodes Bucephalus sp. infestation causedhigher intensity of ceroidosis.
The sum of the mean weighted prevalence for each type ofdegenerative condition (sloughing epithelium, cellular degenera-tions, mucus in digestive lumina) per organ shows that degenera-tion was associated principally with Bucephalus sp. (Fig. 5C).
3.5. Hemocyte variables
Percentage of dead hemocytes was lower in trematode-infestedoysters than in uninfested oysters (p < 0.05). Total hemocyte countwas significantly higher in oysters with trematodes and alsoexposed to the control alga S. lachrymosa (p < 0.05) (Fig. 6). Otherhemocyte variables were not affected significantly by algal treat-ment, trematodes, or the interaction. When considering the inter-action of algal treatment with P. marinus infection, MMP of bothgranulocytes and hyalinocytes was significantly higher in circulat-ing hemolymph of uninfected oysters exposed to the control alga(p < 0.05), but other hemocyte variables were not affected.
3.6. Gene expression
A significantly lower MnSOD gene expression level (p < 0.05)was found in trematode-infested oysters exposed to the control
ogical changes in the eastern oyster Crassostrea virginica infested with therium fundyense. J. Invertebr. Pathol. (2015), http://dx.doi.org/10.1016/

Table 2Statistical differences in weighted prevalence of histopathological conditions in experimental oysters Crassostrea virginica according to exposure to either toxic Alexandriumfundyense or the control dinoflagellate Scrippsiella lachrymosa for 23 days (Mann–Whitney test), infestation with trematodes Bucephalus sp. (Mann–Whitney test), between thefour conditions defined by algal treatment and Bucephalus sp. infestation (Kruskal–Wallis test), according to Perkinsus marinus infection (Mann–Whitney test), and between thefour conditions defined by algal treatment and P. marinus infection (Kruskal–Wallis test).
Histopathologicalobservations
Algaltreatment(n = 26–28)
Bucephalus sp.infestation (n = 22–32)
Difference between the four algal –Bucephalus sp. conditions (n = 9–18)
Perkinsus marinusinfection (n = 27)
Difference between the four algal –P. marinus conditions (n = 9–18)
Pericardium–Ceroid bodies in
auricleNS NS NS NS NS
Kidney–Hemocyte infiltration ** NS ** NS NS–Ceroid bodies NS NS NS NS NS
Adductor muscle–Hemocyte infiltration NS NS NS NS NS–Muscle fiber
degeneration
* * ** NS NS
–Ceroid bodies NS NS NS NS NS
Gills–Hemocyte infiltration NS *** *** NS NS–Ceroid bodies NS NS NS NS NS
Mantle–Hemocyte infiltration NS *** ** NS NS–Ceroid bodies NS NS NS NS NS
Gonad–Maturation NS *** *** NS NS–Hemocyte infiltration NS *** *** NS NS–Ceroid bodies NS ** * NS NS–Gamete degeneration
in gonoductsNS NS NS NS NS
–Gamete degenerationin follicles
NS ** ** NS NS
–Hemocytes ingonoduct andfollicle lumina
NS * * NS NS
Digestive gland connective tissue–Hemocyte infiltration NS *** *** NS NS–Ceroid bodies NS ** * NS NS
Stomach–Epithelium sloughing
into the lumenNS NS NS NS NS
–Hemocytes indiapedesis
NS NS NS NS NS
–Hemocytes in lumen NS NS NS NS NS–Dinoflagellate cells in
lumenNS NS NS NS NS
–Other microalgal cellsin lumen
NS NS NS NS NS
–Hemorhage NS NS NS NS NS
Intestine–Hemocytes in
diapedesisNS NS NS NS NS
–Hemocytes in lumen NS NS NS NS NS–Dinoflagellate cells in
lumenNS NS NS NS NS
–Other microalgal cellsin lumen
NS NS NS NS NS
Digestive ducts and tubules–Hemocyte infiltration
surroundingNS ** * NS NS
Digestive ducts–Epitheliumsloughing into thelumen
NS * NS NS NS
–Hemocytediapedesis
* ** ** * *
–Hemocytes inlumen
NS NS NS NS NS
–Mucus in lumen NS * ** NS NS
(continued on next page)
M. Lassudrie et al. / Journal of Invertebrate Pathology xxx (2015) xxx–xxx 7
YJIPA 6639 No. of Pages 13, Model 5G
11 February 2015
Please cite this article in press as: Lassudrie, M., et al. Physiological and pathological changes in the eastern oyster Crassostrea virginica infested with thetrematode Bucephalus sp. and exposed to the toxic dinoflagellate Alexandrium fundyense. J. Invertebr. Pathol. (2015), http://dx.doi.org/10.1016/j.jip.2015.01.011
567
568
569
570
571
572
573
574
575
576
577
578
579
580
581
582
583
584
585
586
587
588
589
590
591
592
593
594
595
596
597
598
599
600
601
602
603
604
605
606
607
608
609
610
611
612
613
614
615
616
617
618
619
620
621
622
623
624
625
626
627
628
629
630
631
632
633
634
635
636
637
638
Table 2 (continued)
Histopathologicalobservations
Algaltreatment(n = 26–28)
Bucephalus sp.infestation (n = 22–32)
Difference between the four algal –Bucephalus sp. conditions (n = 9–18)
Perkinsus marinusinfection (n = 27)
Difference between the four algal –P. marinus conditions (n = 9–18)
Digestive tubules–Epithelium atrophy NS ** * * NS–Epitheliumsloughing into thelumen
NS *** *** NS NS
–Hemocytes inlumen
NS * NS NS NS
–Mucus in lumen NS NS NS NS NS
NS: no significant difference.* Significant differences were indicated by p < 0.05.
** Significant differences were indicated by p < 0.01.*** Significant differences were indicated by p < 0.001.
Bucephalus sp. : ScripsTr0 ScripsTr1 AlexTr0 AlexTr10
102030405060708090
100
12345
Uninfested Infested Infested
Maturationstage:
S. lachrymosa A. fundyense
Uninfested
78
6
% o
f oys
ters
Fig. 3. Percentages of oysters Crassostrea virginica at each gonadal maturation stagefollowing a semi-quantitative scale from 1 (sexually undifferentiated) to 8(spawned) (adapted from Ford and Figueras, 1988, by Powell et al., 1993) accordingto algal treatment (toxic Alexandrium fundyense or control Scrippsiella lachrymosafor 23 days) and trematode Bucephalus sp. infestation. N in each group: 13 inS. lachrymosa – Uninfested; 13 in S. lachrymosa – Infested;.18 in A. fundyense –Uninfested; 10 in A. fundyense – Infested.
8 M. Lassudrie et al. / Journal of Invertebrate Pathology xxx (2015) xxx–xxx
YJIPA 6639 No. of Pages 13, Model 5G
11 February 2015
alga S. lachrymosa (R = 0.978 ± 0.006; mean ± SE; n = 15) comparedto uninfested oysters fed the same diet (R = 1.000 ± 0.009;mean ± SE; n = 24). Other gene transcript levels did not vary withalgal treatment, trematodes, or the interaction. No significanteffect of P. marinus infection and its interaction with algal treat-ment upon gene expression was detected.
4. Discussion
Finding a population of eastern oysters, C. virginica, with a highprevalence and severity of infestation with the trematode parasiteBucephalus sp. provided an unanticipated opportunity to describephysiological responses of oysters to multiple infectious agents.Additional parasitic infections with Perkinsus marinus and Haplos-poridium spp. were detected, but at lower intensity (and low prev-alence for Haplosporidium spp.) typical of the season. Both parasitescan cause extensive tissue disruption that may persist after infec-tions have declined; however, intensities of P. marinus and Haplos-poridium spp. at the study site have been low for years prior thisexperiment (Sunila, 2013), consistent with the few pathologicaleffects related to P. marinus in this study. Thus, the availability ofthis oyster population provided a model for experimental investi-gation of interactions between parasitism with Bucephalus sp.and harmful-algal-bloom exposure in affecting oyster health andsusceptibility to the other parasitic infections.
Results highlight the principle that harmful algal exposureinteracts significantly with both host and parasite, and such
Please cite this article in press as: Lassudrie, M., et al. Physiological and patholtrematode Bucephalus sp. and exposed to the toxic dinoflagellate Alexandj.jip.2015.01.011
interactions underscore the difficulty in diagnosing oyster healthin the natural environment where simultaneous, multiple stimulimay invoke oyster responses at tissue, cellular, and molecularlevels.
4.1. Effects of parasitic infestations
Infestation with the trematode Bucephalus sp. affected experi-mental oysters from the tissue to the molecular level. Connectivetissue in the gonad was the main site of infestation, suggesting thatthis species was Bucephalus cuculus, as observed previously in C.virginica by Hopkins (1954), McCrady (1874) and Tennent (1906)(NB: Tennent misidentified it as Bucephalus haimeanus at first,see Lauckner, 1983). The tissue occupation by the trematodeswas high enough to increase condition index. The main conse-quence of infestation in the gonad, the most severely affectedorgan, was reduced gonadal maturation compared to uninfestedoysters. In fact, bucephalid sporocysts are known to perturbgonadal development in bivalves, including C. virginica (Calvo-Ugarteburu and McQuaid, 1998; da Silva et al., 2002; Hopkins,1954; McCrady, 1874; Tennent, 1906), because the parasite partlysubstitutes for the gonadal tissue and uses the host’s reproductiveenergy (Calvo-Ugarteburu and McQuaid, 1998; Lafferty and Kuris,2009; Minguez et al., 2012). Energy uptake from the host by thetrematodes probably also explains the increased gamete degener-ation in gonadal follicles compared to uninfested oysters (in thenon-toxic dinoflagellate-exposed oysters). Observation of gilldeformation resulting from trematode sporocyst growth suggestedthat infestation could affect respiratory and feeding performance.
Digestive glands of oysters infested with Bucephalus sp. showeddegradation of digestive ducts and tubules, as indicated byepithelium sloughing into the lumen, atrophy of digestive tubuleepithelium, and presence of mucus in digestive duct lumen. Disar-rangement and degeneration of digestive diverticula and gut wereobserved also by Cheng and Burton (1965) in C. virginica infestedwith Bucephalus sp.; both mechanical and physiological causes,such as impaired nutrient uptake, were mentioned as possiblemechanisms (Cheng and Burton, 1966; Cheng, 1965). In addition,such degeneration can be typical with P. marinus and H. nelsoniinfections (Farley, 1968; Ford, 1985; Smolowitz, 2013), as shownby digestive tubule epithelium atrophy significantly associatedalso with P. marinus infection. These degenerations could thus alsopartly result from other, earlier, parasitic infections.
Hemocyte infiltration into several tissues was observed in oys-ters infested with Bucephalus sp., indicating an inflammatoryresponse, as also observed in mussels P. perna (da Silva et al.,2002). In our study, however, only one oyster showed focal, heavyhemocyte infiltration associated with partial destruction of trema-tode sporocyst branches. Effective destruction of Bucephalus sp. by
ogical changes in the eastern oyster Crassostrea virginica infested with therium fundyense. J. Invertebr. Pathol. (2015), http://dx.doi.org/10.1016/

639
640
641
642
643
644
645
646
647
648
649
650
651
652
653
654
655
656
657
658
659
660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
678
679
680
681
682
0.0
0.5
1.0
1.5
2.0
2.5
aab ab
bH
emoc
yte
diap
edes
isin
dig
estiv
e du
cts
(a.u
.)
0.0
0.5
1.0
1.5
2.0
2.5
a
b
abab
UninfestedInfested
Bucephalus sp. :
Hem
ocyt
e in
filtra
tion
surro
undi
ngdi
gest
ive
duct
s an
d tu
bule
s (a
.u.)
0.0
0.5
1.0
1.5
2.0
2.5
a
b b b
Hem
ocyt
e in
filtra
tion
in k
idne
y (a
.u.)
0.0
0.5
1.0
1.5
2.0
2.5
a
b
a ab
Hem
ocyt
es in
gon
oduc
tan
d fo
llicle
lum
ina
(a.u
.)
S. lachrymosa A. fundyense0.0
0.5
1.0
1.5
2.0
2.5
a
b
aab
Gam
ete
dege
nera
tion
in g
onad
al fo
llicle
s (a
.u.)
S. lachrymosa A. fundyense0.0
0.5
1.0
1.5
2.0
2.5
a a a
b
Mus
cle
fiber
deg
ener
atio
nin
the
addu
ctor
mus
cle
(a.u
.)
A
C D
B
E F
Fig. 4. Mean weighted prevalences of histopathological conditions modified by the interaction of algal treatment (toxic Alexandrium fundyense or control Scrippsiellalachrymosa) and infestation with trematodes Bucephalus sp. in oysters Crassostrea virginica after 23 days of experimental algal exposure. (A) hemocyte diapedesis in digestiveducts; (B) hemocyte infiltration surrounding digestive ducts and tubules; (C) hemocyte infiltration in kidney; (D) hemocytes in gonoduct and follicle lumina; (E) degenerationof gametes in gonadal follicles; (F) muscle fiber degeneration in the adductor muscle. Letters indicate significant differences between conditions (Kruskal–Wallis test followedby post hoc NDWD test, p < 0.05). Error bars represent standard errors. N per group: 13 in S. lachrymosa – Uninfested; 13 in S. lachrymosa – Infested; 18 in A. fundyense –Uninfested; 10 in A. fundyense – Infested.
M. Lassudrie et al. / Journal of Invertebrate Pathology xxx (2015) xxx–xxx 9
YJIPA 6639 No. of Pages 13, Model 5G
11 February 2015
hemocytic defense responses is usually not observed in bivalvesand has only been reported rarely (Cheng and Burton, 1965; daSilva et al., 2002; Douglass, 1975); whereas, other digenetic trema-todes, such as Proctoeces maculatus infesting mussels Mytilus gallo-provincialis can be encapsulated (Villalba et al., 1997). In addition,hemocytes also migrated in response to lesions caused by the par-asitic infestations. Indeed, degeneration of gametes in follicles wasassociated with infiltration and presence of hemocytes in follicleand gonaduct lumina, probably to resorb degenerated material. Indigestive glands of oysters exposed to the non-toxic dinoflagellate,degeneration caused by parasitic infections was associated withmore intense hemocyte infiltration. This also was associated withmore intense hemocyte diapedesis in the digestive-duct epitheliaof P. marinus- and in Bucephalus sp.-infested oysters, and presenceof hemocytes in digestive tubule lumina of Bucephalus sp.-infestedoysters. In hemolymph, the increase in hemocyte concentration intrematode-infested oysters exposed to the non-toxic alga wasaccompanied by a decrease in dead hemocytes within trematode-infested oysters. This may result from enhancement of de novohemocyte production, as proposed in previous studies (Hégaretand Wikfors, 2005; Hégaret et al., 2004, 2007; Soudant et al.,2004) and may help to sustain the inflammatory response.
Please cite this article in press as: Lassudrie, M., et al. Physiological and patholtrematode Bucephalus sp. and exposed to the toxic dinoflagellate Alexandj.jip.2015.01.011
In the two most infested tissues, gonad and digestive gland,presence of Bucephalus sp. also was associated with more numer-ous ceroid bodies (also referred to as brown cells, serous cells, porecells, or rhogocytes in the literature), as previously observed byCheng and Burton (1965). Ceroid, which is believed to play a rolein detoxication and in the excretory system in bivalves(Haszprunar, 1996; Zaroogian and Voyer, 1995), is constituted ofnon-degradable material resulting mainly from the accumulationof oxidized proteins and lipids (Yin, 1996). The increased intensityof ceroidosis in affected oysters may indicate oxidative stressinduced by the inflammatory response upon trematode infestation.In hemocytes of P. marinus-infected oysters exposed to the controldinoflagellate, MMP was lower compared to uninfected oysters,suggesting that P. marinus affects cellular metabolic functions ofits host. This effect could be a consequence of antioxidantcompounds produced by P. marinus that protect the parasite fromthe oxidative burst associated to phagocytosis and enables it todevelop within C. virginica hemocytes (Anderson, 1999; Schottet al., 2003; Smolowitz, 2013; Volety and Chu, 1995; and seereview of Soudant et al., 2013) or be associated with hemocyteapoptosis repression by P. marinus (Goedken et al., 2005; Hugheset al., 2010; Ly et al., 2003).
ogical changes in the eastern oyster Crassostrea virginica infested with therium fundyense. J. Invertebr. Pathol. (2015), http://dx.doi.org/10.1016/

683
684
685
686
687
688
689
690
691
692
693
694
695
696
697
698
699
700
701
702
703
704
705
706
707
708
709
710
711
712
713
714
715
716
717
718
719
720
721
722
723
724
725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
747
748
749
750
751
752
753
754
755
756
757
758
759
0
5
10
15
Uninfested
Hem
ocyt
e re
spon
ses
(wei
ghte
d pr
eval
ence
, a.u
.)
InfestedBucephalus sp.:0
2
4
6
8
Cer
oïdo
sis
(wei
ghte
d pr
eval
ence
, a.u
.)
Uninfested Infested0
2
4
6
8
Deg
ener
atio
ns(w
eigh
ted
prev
alen
ce, a
.u.)
Uninfested InfestedS. lachrymosa A. fundyense
Uninfested InfestedS. lachrymosa A. fundyense
Uninfested InfestedS. lachrymosa A. fundyense
Uninfested Infested
MantleKidneyGonadGills
Muscle
Digestive glandHeart auricleA B C
Fig. 5. Sum of the mean weighted prevalences of each different histopathological conditions, per organ, corresponding to the following categories: (A) hemocyte responses;(B) ceroidosis; (C) degenerations in experimental oysters Crassostrea virginica after 23 days of experiment according to algal treatment (toxic Alexandrium fundyense or controlScrippsiella lachrymosa) and infestation with trematodes Bucephalus sp. N per group: 13 in S. lachrymosa – Uninfested; 13 in S. lachrymosa – Infested; 18 in A. fundyense –Uninfested; 10 in A. fundyense – Infested.
S. lachrymosa A. fundyense0
1
2
3
4
5
a
b
abab
THC
(10 5
cel
l mL-1
)
UninfestedInfested
Bucephalus sp. :
Fig. 6. Total hemocyte count (THC) from circulating hemolymph of oysterCrassostrea virginica after exposure to toxic Alexandrium fundyense or controlScrippsiella lachrymosa for 23 days, and trematode Bucephalus sp. infestation. Lettersindicate significant differences (2-way ANOVA followed by LSD post hoc test,p < 0.05). N per group: 24 in S. lachrymosa – Uninfested; 15 in S. lachrymosa –Infested; 27 in A. fundyense – Uninfested; 11 in A. fundyense – Infested.
10 M. Lassudrie et al. / Journal of Invertebrate Pathology xxx (2015) xxx–xxx
YJIPA 6639 No. of Pages 13, Model 5G
11 February 2015
4.2. Effect of A. fundyense exposure
In all oysters exposed for three weeks to A. fundyense, histolog-ical observations of intact or lysed A. fundyense cells in intestinelumen, as well as PST accumulation in the digestive gland, demon-strated that oysters ingested and digested this alga. In oystersuninfested by trematodes, increased hemocyte infiltration inresponse to A. fundyense exposure also was detected in connectivetissues of kidneys, which form an excretory complex together withthe pericardial gland (Morse, 1987). This inflammatory response,which was specific to this organ, may indicate that algal toxinsare carried, presumably by hemocytes, toward kidneys to beexcreted. Galimany et al. (2008) also suggested that hemocytescould transport PST from tissues to feces by diapedesis throughthe digestive epithelium. In fact, hemocyte diapedesis is a typicalresponse of bivalves exposed to toxic Alexandrium spp.(Haberkorn et al., 2010b; Lassudrie et al., 2014; Medhioub et al.,2012). Similarly, we observed that A. fundyense exposure induceddiapedesis of hemocytes through the digestive duct epithelia inoysters uninfested by trematodes.
In addition, A. fundyense exposure affected oyster susceptibilityto Dermo disease, caused by P. marinus. Indeed, althoughweighted prevalence decreased during the time course of theexperiment in oysters exposed to the control alga, no decreasewas observed in A. fundyense-exposed oysters, resulting in ahigher weighted prevalence than in control oysters. In contrast,da Silva et al. (2008) described lower intensity of infection withPerkinsus olseni in Manila clams exposed to the toxic dinoflagellateKarenia selliformis, which was attributed to direct toxicity of thealga to the parasite. Similarly, another dinoflagellate, Prorocentrum
Please cite this article in press as: Lassudrie, M., et al. Physiological and patholtrematode Bucephalus sp. and exposed to the toxic dinoflagellate Alexandj.jip.2015.01.011
minimum, was reported to be toxic to P. olseni (Hégaret et al.,2009). Such direct interaction between A. fundyense and P. marinuswould not explain the results of the present study. Thus, inabilityto repress P. marinus infection could have been caused by physio-logical alteration to the host induced by A. fundyense exposure,either by decreasing P. marinus excretion rate, or by decreasingdefense mechanisms. The last hypothesis was suggested byGalimany et al. (2008) after observation of increased trematode(Gymnophallidae) prevalence after A. fundyense exposure inmussels. In the present study, effects of A. fundyense alone uponhemocytes were minimal, suggesting that other mechanisms areinvolved.
4.3. Effects of the interaction between parasitic infestations andA. fundyense exposure
In oysters infested with the trematode Bucephalus sp., theexposure to A. fundyense was associated with a higher prevalenceof P. marinus, suggesting that the combined effects of A. fundyenseand trematodes may promote P. marinus infection or at leastprevent regression.
The responses of hemocytes to the trematodes that were sup-pressed by A. fundyense exposure (THC, infiltrations in kidneyand in digestive gland, hemocytes in gonoduct and follicle lumina)may have impaired immune functions. In fact, deficiencies inimmune functions could explain why infection and proliferationof P. marinus was enhanced.
Another response to combined Bucephalus sp. infestation andA. fundyense exposure was degeneration of adductor muscle fibers.Paralytic Shellfish Toxins (PSTs) and bucephalids individually areknown to affect adductor muscles of bivalves (Calvo-Ugarteburuand McQuaid, 1998; Haberkorn et al., 2010b; Hégaret et al.,2012; Howell, 1967); however, in the present experiment, onlythe interaction of both led to a significant increase in myodegener-ation. Again, suppression of hemocyte responses could have a rolein this pathology. A first hypothesis considers the potential role ofhemocytes in carrying and detoxifying PSTs, which would be lessefficient in parasitized oysters. Although the PST content in thedigestive gland remained unchanged between oysters with orwithout trematodes, PST concentration in other tissues may havevaried. Another hypothesis considers the role of hemocytes innutrient digestion and transport, combined with trematode energyuptake, resulting in depressed nutrient supply to the muscle andenergy depletion. The degeneration of adductor muscle fiberscould have deleterious consequences upon valve movements andthe catch state. Adductor muscle dysfunction also could affectfiltration, interfering with respiration and nutrition functions,and could increase susceptibility of oysters to predation andexposure to environmental variations that otherwise could bemoderated by shell closure.
ogical changes in the eastern oyster Crassostrea virginica infested with therium fundyense. J. Invertebr. Pathol. (2015), http://dx.doi.org/10.1016/

760
761
762
763
764
765
766
767
768
769
770
771
772
773
774
775
776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
793
794
795
796797798799800801802803804805806807808809810811812813814815816817818819820821822823824825826
827828829830831832833834835836837838839840841842843844845846847848849850851852853854855856
M. Lassudrie et al. / Journal of Invertebrate Pathology xxx (2015) xxx–xxx 11
YJIPA 6639 No. of Pages 13, Model 5G
11 February 2015
5. Conclusions
Infestation with trematodes, Bucephalus sp., affected oyster, C.virginica, physiological functions including reproduction andimmunity. In addition, considering histopathological lesions ingills and digestive tubules, respiration and digestion could bealtered. Neither A. fundyense blooms nor Bucephalus sp. infestationhave been reported to be lethal for C. virginica. The results of thepresent study suggest, however, that an A. fundyense bloom couldintensify the severity of a potentially-lethal disease, Dermo,especially in trematode-infested oysters that would be more sus-ceptible to Dermo infections. In addition, oysters with trematodeinfestations exposed to blooms of PST-producing microalgae maybe more susceptible to predation as a consequence of muscle fiberdegeneration that could compromise valve-closure movementsand the catch state of the adductor muscle. There is increasingevidence of HAB relationships with climate change (temperature,eutrophication, etc.) (Anderson et al., 2008; Hallegraeff et al.,2003), and HABs can modify host–pathogen interactions, for exam-ple by aggravating disease status as demonstrated in the presentstudy. In this context, a better understanding of complex bioticinteractions is needed to predict and possibly prevent new diseaseoutbreaks in bivalve populations.
857858859860861862863864865866867868869870871872873874875876877
Acknowledgments
This work was supported by Université de Bretagne Occiden-tale, the NOAA Fisheries Service Aquaculture Program, and from‘‘Laboratoire d’Excellence’’ LabexMER (ANR-10-LABX-19), whichwas co-funded by a grant from the French government under theprogram ‘‘Investissements d’Avenir’’. Authors are grateful to EveGalimany, Barry Smith, Yaqin Li, Christophe Lambert, Nelly LeGoïc, Anne-Laure Cassone and Marc Long for technical assistanceand advice, to Milford laboratory staff for their participation indissections, to Joseph DeCrescenzo for help with the RFTM assayand to Sébastion Hervé for graphical abstract realization. Authorsare also grateful to the anonymous reviewers who helped improvethe manuscript.
878879880881882883884885886887888889890891892893894895896897898899900901902903904905906907908909910911912
References
Altschul, S.F., Madden, T.L., Schäffer, A.A., Zhang, J., Zhang, Z., Miller, W., Lipman, D.J.,1997. Gapped BLAST and PSI-BLAST: a new generation of protein databasesearch programs. Nucleic Acids Res. 25, 3389–3402.
Anderson, R.S., 1999. Perkinsus marinus secretory products modulate superoxideanion production by oyster (Crassostrea virginica) haemocytes. Fish ShellfishImmunol. 9, 51–60.
Anderson, D.M., Keafer, B.A., McGillicuddy, D.J., Mickelson, M.J., Keay, K.E., Libby,P.S., Manning, J.P., Mayo, C.A., Whittaker, D.K., Hickey, J.M., He, R., Lynch, D.R.,Smith, K.W., 2005. Initial observations of the 2005 Alexandrium fundyense bloomin southern New England: General patterns and mechanisms. Deep Sea Res. PartII Top. Stud. Oceanogr. 52, 2856–2876.
Anderson, D.M., Burkholder, J.M., Cochlan, W.P., Glibert, P.M., Gobler, C.J., Heil, C.A.,Kudela, R., Parsons, M.L., Rensel, J.E.J., Townsend, D.W., Trainer, V.L., Vargo, G.A.,2008. Harmful algal blooms and eutrophication: Examining linkages fromselected coastal regions of the United States. Harmful Algae 8, 39–53.
Béguel, J.-P., Huvet, A., Quillien, V., Lambert, C., Fabioux, C., 2013. Study of theantioxidant capacity in gills of the Pacific oyster Crassostrea gigas in link with itsreproductive investment. Comp. Biochem. Physiol. Part C 157, 63–71.
Bodoy, A., Prou, J., Berthome, J.-P., 1986. Etude comparative des différents indices deconditions chez l’huître creuse (Crassostrea gigas). Haliotis 15, 173–182.
Bricelj, V.M., Ford, S.E., Lambert, C., Barbou, A., Paillard, C., 2011. Effects of toxicAlexandrium tamarense on behavior, hemocyte responses and development ofbrown ring disease in Manila clams. Mar. Ecol. Prog. Ser. 430, 35–48.
Brulle, F., Sunila, I., Wikfors, G., 2012. ‘‘Clinton line’’ oysters have an overall low andnarrow range of expression of stress- and immunity-related genes compared to‘‘EGP line’’ oysters. Abstracts of Technical Papers, Presented at the 32nd Annual,Milford Aquaculture Seminar, Milford, Connecticut, March 12–14, 2012. J.Shellfish Res. 31 (1), 201–230.
Bushek, D., Ford, S.E., Allen Jr, S.K., 1994. Evaluation of methods using ray’s fluidthioglycollate medium for diagnosis of Perkinsus marinus infection in theeastern oyster, Crassostrea virginica. Annu. Rev. Fish Dis. 4, 201–217.
Please cite this article in press as: Lassudrie, M., et al. Physiological and patholtrematode Bucephalus sp. and exposed to the toxic dinoflagellate Alexandj.jip.2015.01.011
Bustin, S.A., Benes, V., Garson, J.A., Hellemans, J., Huggett, J., Kubista, M., Mueller, R.,Nolan, T., Pfaffl, M.W., Shipley, G.L., Vandesompele, J., Wittwer, C.T., 2009. TheMIQE guidelines: minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 55, 611–622.
Calvo-Ugarteburu, G., McQuaid, C.D., 1998. Parasitism and invasive species: effectsof digenetic trematodes on mussels. Mar. Ecol. Prog. Ser. 169, 149–163.
Cheng, T.C., 1965. Histochemical observations on changes in the lipid compositionof the American oyster, Crassostrea virginica (Gmelin), parasitized by thetrematode Bucephalus sp. J. Invertebr. Pathol. 7, 398–407.
Cheng, T.C., 1981. Bivalves. In: Ratcliffe, N.A., Rowley, A.F. (Eds.), Invertebrate BloodCells, vol. 1. Academic Press, London, pp. 233–300.
Cheng, T.C., 1996. Haemocytes: forms and functions. In: Kennedy, V., Newell, R.,Eble, A. (Eds.), The Eastern Oyster Crassostrea virginica. Maryland Sea Grant,College Park, pp. 299–333.
Cheng, T.C., Burton, R.W., 1965. Relationships between Bucephalus sp. Crassostreavirginica: histopathology and sites of infection. Chesap. Sci. 6, 3–16.
Cheng, T.C., Burton, R.W., 1966. Relationships between Bucephalus sp. andCrassostrea virginica: a histochemical study of some carbohydrates andcarbohydrate complexes occurring in the host and parasite. Parasitology 56,111–122.
Cucci, T.L., Shumway, S.E., Newell, R.C., Yentsch, M., 1985. A preliminary study of theeffects of Gonyaulax tamarensis on feeding in bivalve molluscs. In: Anderson,D.M., White, A.W., Baden, D.G. (Eds.), Toxic Dinoflagellates. Elsevier/North-Holland, Amsterdam, pp. 395–400.
da Silva, P.M., Magalhães, A.R.M., Barracco, M.A., 2002. Effects of Bucephalus sp.(Trematoda: Bucephalida) on Perna perna mussels from a culture station inRatones Grande Island, Brazil. J. Invertebr. Pathol. 79, 154–162.
da Silva, P.M., Hégaret, H., Lambert, C., Wikfors, G.H., Le Goïc, N., Shumway, S.E.,Soudant, P., 2008. Immunological responses of the Manila clam (Ruditapesphilippinarum) with varying parasite (Perkinsus olseni) burden, during a long-term exposure to the harmful alga, Karenia selliformis, and possible interactions.Toxicon 51, 563–573.
Delaporte, M., Soudant, P., Moal, J., Lambert, C., Quéré, C., Miner, P., Choquet, G.,Paillard, C., Samain, J.F., 2003. Effect of a mono-specific algal diet on immunefunctions in two bivalve species – Crassostrea gigas and Ruditapes philippinarum.J. Exp. Biol. 206, 3053–3064.
Donaghy, L., Kraffe, E., Le Goïc, N., Lambert, C., Volety, A.K., Soudant, P., 2012.Reactive oxygen species in unstimulated hemocytes of the Pacific oysterCrassostrea gigas: a mitochondrial involvement. PLoS ONE 7, 1–10.
Douglass, W.R., 1975. Host response to infection with Bucephalus in Crassostreavirginica. In: Proceedings of the National Shellfisheries Association 65. pp. 1.
Fabioux, C., Sulistiyani, Y., Haberkorn, H., Hégaret, H., Soudant, P. Exposure to toxicAlexandrium minutum activates the antioxidant and detoxifying systems of theoyster Crassostrea gigas. (in preparation).
FAO, 2014. Fisheries and Aquaculture Information and Statistics Service, GlobalProduction Statistics 1950–2012 [WWW Document]. URL <http://www.fao.org/fishery/statistics/global-production/query/en> (accessed 03.04.14).
Farley, C.A., 1968. Minchinia nelsoni (Haplosporida) disease dyndrome in theAmerican oyster Crassostrea virginica. J. Protozool. 15, 585–599.
Feng, S.Y., 1988. Cellular defence mechanisms of oysters and mussels. Am. Fish. Soc.Spec. Publ. 18, 153–158.
Feng, S.Y., Canzonier, W.J., 1970. Humoral responses in the American oyster(Crassostrea virginica) infected with Bucephalus sp. and Minchinia nelsoni. Am.Fish. Soc. Spec. Publ., 497–510
Fisher, W.S., 1986. Structure and functions of oyster haemocytes. In: Brehélin, M.(Ed.), Immunity in Invertebrates. Springer-Verlag, Heidelberg, Berlin, pp. 25–35.
Fisher, W.S., 1988. Environmental influence on bivalve hemocyte function. Am. Fish.Soc. Spec. Publ. 18, 225–237.
Ford, S.E., 1985. Chronic infections of Haplosporidium nelsoni (MSX) in the oysterCrassostrea virginica. J. Invertebr. Pathol. 45, 94–107.
Ford, S.E., Figueras, A.J., 1988. Effects of sublethal infection by the parasiteHaplosporidium nelsoni (MSX) on gametogenesis, spawning, and sex ratios ofoysters in Delaware Bay, USA. Dis. Aquat. Organ. 4, 121–133.
Ford, S.E., Smolowitz, R., 2007. Infection dynamics of an oyster parasite in its newlyexpanded range. Mar. Biol. 151, 119–133.
Ford, S.E., Tripp, M.R., 1996. Disease and defense mechanisms. In: Kennedy, V.S.,Newell, R.I.E., Eble, A.F. (Eds.), The Eastern Oyster Crassostrea virginica.Maryland Sea Grant Book, College Park, MD, USA, pp. 581–660.
Gainey, L.F., Shumway, S.E., 1988a. Physiological effects of Protogonyaulaxtamarensis on cardiac activity in bivalves molluscs. Comp. Biochem. Physiol.Part C 91, 159–164.
Gainey, L.F., Shumway, S.E., 1988b. A compendium of the responses of bivalvemolluscs to toxic dinoflagellates. J. Shellfish Res. 7, 623–628.
Galimany, E., Sunila, I., Hégaret, H., Ramón, M., Wikfors, G.H., 2008. Experimentalexposure of the blue mussel (Mytilus edulis, L.) to the toxic dinoflagellateAlexandrium fundyense: Histopathology, immune responses, and recovery.Harmful Algae 7, 702–711.
Galimany, E., Ramón, M., Ibarrola, I., 2011. Feeding behavior of the mussel Mytilusgalloprovincialis (L.) in a Mediterranean estuary: a field study. Aquaculture 314,236–243.
Goedken, M., Morsey, B., Sunila, I., De Guise, S., 2005. Immunomodulation ofCrassostrea gigas and Crassostrea virginica cellular defense mechanisms byPerkinsus marinus. J. Shellfish Res. 24, 487–496.
Guillard, R.R.L., Ryther, J.H., 1962. Studies of marine planktonic diatoms. I.Cyclotella nana Hustedt, and Detonula confervacea Cleve. Can. J. Microbiol. 8,229–239.
ogical changes in the eastern oyster Crassostrea virginica infested with therium fundyense. J. Invertebr. Pathol. (2015), http://dx.doi.org/10.1016/

913914915916917918919920921922923924925926927928929930931932933934935936937938939940941942943944945946947948949950951952953954955956957958959960961962963964965966967968969970971972973974975976977978979980981982983984985986987988989990991992993994995996997998
9991000100110021003100410051006100710081009101010111012101310141015101610171018101910201021102210231024102510261027102810291030103110321033103410351036103710381039104010411042104310441045104610471048104910501051105210531054105510561057105810591060106110621063106410651066106710681069107010711072107310741075107610771078107910801081108210831084
12 M. Lassudrie et al. / Journal of Invertebrate Pathology xxx (2015) xxx–xxx
YJIPA 6639 No. of Pages 13, Model 5G
11 February 2015
Haberkorn, H., Lambert, C., Le Goïc, N., Guéguen, M., Moal, J., Palacios, E., Lassus, P.,Soudant, P., 2010a. Effects of Alexandrium minutum exposure upon physiologicaland hematological variables of diploid and triploid oysters, Crassostrea gigas.Aquat. Toxicol. 97, 96–108.
Haberkorn, H., Lambert, C., Le Goïc, N., Moal, J., Suquet, M., Guéguen, M., Sunila, I.,Soudant, P., 2010b. Effects of Alexandrium minutum exposure on nutrition-related processes and reproductive output in oysters Crassostrea gigas. HarmfulAlgae 9, 427–439.
Hallegraeff, G.M., Anderson, D.M., Cembella, A.D. (Eds.), 2003. Manual on harmfulmarine microalgae. Monographs on Oceanographic Methodology, vol. 11.UNESCO Publishing, Paris.
Haszprunar, G., 1996. The molluscan rhogocyte (pore-cell, blasenzelle, cellulenucale), and its significance for ideas on nephridial evolution. J. Molluscan Stud.62, 185–211.
Hattenrath, T.K., Anderson, D.M., Gobler, C.J., 2010. The influence of anthropogenicnitrogen loading and meteorological conditions on the dynamics and toxicity ofAlexandrium fundyense blooms in a New York (USA) estuary. Harmful Algae 9,402–412.
Hégaret, H., Wikfors, G.H., 2005. Time-dependent changes in hemocytes of easternoysters, Crassostrea virginica, and northern bay scallops, Argopecten irradiansirradians, exposed to a cultured strain of Prorocentrum minimum. Harmful Algae4, 187–199.
Hégaret, H., Wikfors, G.H., Soudant, P., 2003. Flow-cytometric analysis ofhaemocytes from eastern oysters, Crassostrea virginica, subjected to a suddentemperature elevation. I. Haemocyte types and morphology. J. Exp. Mar. Bio.Ecol. 293, 237–248.
Hégaret, H., Wikfors, G., Soudant, P., Delaporte, M., Alix, J., Smith, B., Dixon, M.,Quéré, C., Le Coz, J., Paillard, C., Moal, J., Samain, J.-F., 2004. Immunologicalcompetence of eastern oysters, Crassostrea virginica, fed different microalgaldiets and challenged with a temperature elevation. Aquaculture 234, 541–560.
Hégaret, H., Wikfors, G.H., Soudant, P., Lambert, C., Shumway, S.E., Bérard, J.B.,Lassus, P., 2007. Toxic dinoflagellates (Alexandrium fundyense and A. catenella)have minimal apparent effects on oyster hemocytes. Mar. Biol. 152, 441–447.
Hégaret, H., da Silva, P.M., Sunila, I., Shumway, S.E., Dixon, M.S., Alix, J., Wikfors,G.H., Soudant, P., 2009. Perkinsosis in the Manila clam Ruditapes philippinarumaffects responses to the harmful-alga, Prorocentrum minimum. J. Exp. Mar. Bio.Ecol. 371, 112–120.
Hégaret, H., da Silva, P.M., Wikfors, G.H., Haberkorn, H., Shumway, S.E., Soudant, P.,2011. In vitro interactions between several species of harmful algae andhaemocytes of bivalve molluscs. Cell Biol. Toxicol. 27, 249–266.
Hégaret, H., Brokordt, K.B., Gaymer, C.F., Lohrmann, K.B., Garcia, C., Varela, D., 2012.Effects of the toxic dinoflagellate Alexandrium catenella on histopathogical andescape responses of the Northern scallop Argopecten purpuratus. Harmful Algae18, 74–83.
Hine, P.M., 1999. The inter-relationships of bivalve haemocytes. Fish ShellfishImmunol. 9, 367–385.
Hopkins, S.H., 1954. The American species of trematode confused with Bucephalus(Bucephalopsis) haimeanus. Parasitology 44, 353–370.
Howard, D.W., Lewis, E.J., Keller, B.J., Smith, C.S., 2004. Histological techniques formarine bivalve mollusks and crustaceans. NOAA Tech. Memo. NOS NCCOS 5,218.
Howell, M., 1967. The trematode, Bucephalus longicornutus (Manter, 1954), in theNew Zealand mud-oyster, Ostrea lutaria. Trans. Proc. R. Soc. New Zeal. 8, 221–237.
Hughes, F.M., Foster, B., Grewal, S., Sokolova, I.M., 2010. Apoptosis as a host defensemechanism in Crassostrea virginica and its modulation by Perkinsus marinus.Fish Shellfish Immunol. 29, 247–257.
Itoh, N., Xue, Q.-G., Schey, K.L., Li, Y., Cooper, R.K., La Peyre, J.F., 2011.Characterization of the major plasma protein of the eastern oyster,Crassostrea virginica, and a proposed role in host defense. Comp. Biochem.Physiol. Part B 158, 9–22.
Janeway, C.a., Medzhitov, R., 2002. Innate immune recognition. Annu. Rev.Immunol. 20, 197–216.
Jenny, M.J., Warr, G.W., Ringwood, A.H., Baltzegar, D.a., Chapman, R.W., 2006.Regulation of metallothionein genes in the American oyster (Crassostreavirginica): ontogeny and differential expression in response to differentstressors. Gene 379, 156–165.
Kim, Y., Ashton-Alcox, K.A., Powell, E.N., 2006. Histological techniques for marinebivalve molluscs: update. NOAA Tech. Memo. NOS NCCOS 27, 76.
Koressaar, T., Remm, M., 2007. Enhancements and modifications of primer designprogram Primer3. Bioinformatics 23, 1289–1291.
Lafferty, K.D., Kuris, A.M., 2009. Parasitic castration: the evolution and ecology ofbody snatchers. Trends Parasitol. 25, 564–572.
Landsberg, J.H., 2002. The effects of harmful algal blooms on aquatic organisms. Rev.Fish. Sci. 10, 113–390.
Lassudrie, M., Soudant, P., Henry, N., Medhioub, W., da Silva, P.M., Donval, A., Bunel,M., Le Goïc, N., Lambert, C., de Montaudouin, X., Fabioux, C., Hégaret, H., 2014.Physiological responses of Manila clams Venerupis (=Ruditapes) philippinarumwith varying parasite Perkinsus olseni burden to toxic algal Alexandriumostenfeldii exposure. Aquat. Toxicol. 154, 27–38.
Lauckner, G., 1983. Diseases of mollusca: bivalvia. In: Kinne, O. (Ed.), Diseases ofMarine Animals. Biologische Anstalt Helgoland, Hamburg, pp. 477–879.
Lopez, G., Carey, D., Carlton, J.T., Cerrato, R., Dam, H., DiGiovanni, R., Elphick, C.,Frisk, M., Gobler, C., Hice, L., Howell, P., Jordaan, A., Lin, S., Liu, S., Lonsdale, D.,Mcenroe, M., Mckown, K., Mcmanus, G., Orson, R., Peterson, B., Pickerell, C.,Rozsa, R., Siuda, A., Thomas, E., Taylor, G., Shumway, S., Talmage, S., Patten, M.
Please cite this article in press as: Lassudrie, M., et al. Physiological and patholtrematode Bucephalus sp. and exposed to the toxic dinoflagellate Alexandj.jip.2015.01.011
Van, Vaudrey, J., Wikfors, G., Yarish, C., Zajac, R., 2014. Biology and ecology oflong island sound. In: Latimer, J.S., Tedesco, M.A., Swanson, R.L., Yarish, C.,Stacey, P.E., Garza, C. (Eds.), Long Island Sound: Prospects for the Urban Sea.Springer, pp. 285–479.
Ly, J.D., Grubb, D.R., Lawen, A., 2003. The mitochondrial membrane potential (Dwm)in apoptosis; an update. Apoptosis 8, 115–128.
Mackin, J.G., 1962. Oyster disease caused by Dermocystidium marinum and othermicroorganisms in Louisiana. Publ. Inst. Mar. Sci. Univ. Texas 7, 132–229.
McCrady, J., 1874. Observations on the food and the reproductive organs of Ostreavirginica, with some account of Bucephalus cuculus nov. spec. Proc. Bost. Soc. nat.Hist. 16, 170–192.
Medhioub, W., Lassus, P., Truquet, P., Bardouil, M., Amzil, Z., Sechet, V., Sibat, M.,Soudant, P., 2012. Spirolide uptake and detoxification by Crassostrea gigasexposed to the toxic dinoflagellate Alexandrium ostenfeldii. Aquaculture 358–359, 108–115.
Mello, D.F., da Silva, P.M., Barracco, M.A., Soudant, P., Hégaret, H., 2013. Effects ofthe dinoflagellate Alexandrium minutum and its toxin (saxitoxin) on thefunctional activity and gene expression of Crassostrea gigas hemocytes.Harmful Algae 26, 45–51.
Minguez, L., Buronfosse, T., Giambérini, L., 2012. Different host exploitationstrategies in two zebra mussel-trematode systems: adjustments of host lifehistory traits. PLoS ONE 7, 1–8.
Morse, M.P., 1987. Comparative functional morphology of the bivalve excretorysystem. Am. Zool. 27, 737–746.
Olsen, O.W., 1974. Animal Parasites: Their Life Cycles and Ecology. University ParkPress, Baltimore.
Pfaffl, M.W., 2001. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 29, 2002–2007.
Powell, E.N., Wilson-Ormond, E.A., Choi, K.S., 1993. Gonadal analysis – Crassostreavirginica. In: Sampling and Analytical Methods of the National Status andTrends Program National Benthic Surveillance and Mussel Watch Projects1984–1992. Vol. II Comprehensive Descriptions of ComplementaryMeasurements. NOAA Tech. Mem. NOS ORCA 71. NOAA/NOS/ORCA, SilverSpring, MD, pp. 55–62.
Core Team, R., 2012. R: A Language and Environment for Statistical Computing. RFoundation for Statistical Computing, Vienna, Austria, ISBN 3-900051-07-0,URL: http://www.r-project.org/.
Schott, E.J., Pecher, W.T., Okafor, F., Vasta, G.R., 2003. The protistan parasitePerkinsus marinus is resistant to selected reactive oxygen species. Exp. Parasitol.105, 232–240.
Shaw, B.L., Battle, H.I., 1957. The gross and microscopic anatomy of the digestivetract of the oyster Crassostrea virginica (Gmelin). Can. J. Zool. 35, 325–347.
Shumway, S.E., 1990. A review of the effects of algal blooms on shellfish andaquaculture. J. World Aquac. Soc. 21, 65–104.
Smolowitz, R.M., 2013. A review of current state of knowledge concerning Perkinsusmarinus effects on Crassostrea virginica (Gmelin) (the eastern oyster). Vet.Pathol. 50, 404–411.
Song, L., Wang, L., Qiu, L., Zhang, H., 2010. Bivalve immunity. In: Söderhäll, K. (Ed.),Invertrebrate Immunity – Advances in Experimental Medicine and Biology 708.Landes Bioscience and Springer Science+Business Media, LLC, New York, NY,USA, pp. 44–65.
Soudant, P., Paillard, C., Choquet, G., Lambert, C., Reid, H.I., Marhic, A., Donaghy, L.,Birbeck, T.H., 2004. Impact of season and rearing site on the physiological andimmunological parameters of the Manila clam Venerupis (=Tapes, = Ruditapes)philippinarum. Aquaculture 229, 401–418.
Soudant, P., Chu, F.L., Volety, A., 2013. Host-parasite interactions: marine bivalvemolluscs and protozoan parasites, Perkinsus species. J. Invertebr. Pathol. 114,196–216.
Sunila, I., 2013. Status of oyster diseases in Connecticut. J. Shellfish Res. 32, 607.Tasumi, S., Vasta, G.R., 2007. A galectin of unique domain organization from
hemocytes of the eastern oyster (Crassostrea virginica) is a receptor for theprotistan parasite Perkinsus marinus. J. Immunol. 179, 3086–3098.
Tennent, D.H., 1906. A study of the life-history of Bucephalus haimeanus; a parasiteof the oyster. Q. J. Microsc. Sci., 635–690
Ukeles, R., 1973. Continuous culture—a method for the production of unicellularalgal foods. In: Stein, J.R. (Ed.), Handbook of Phycological Methods: CultureMethods and Growth Measurements. Cambridge University Press, Cambridge,pp. 233–255.
Untergasser, A., Cutcutache, I., Koressaar, T., Ye, J., Faircloth, B.C., Remm, M., Rozen,S.G., 2012. Primer3—new capabilities and interfaces. Nucleic Acids Res. 40,e115.
Vale, C., Alfonso, A., Vieytes, M.R., Romarís, X.M., Arévalo, F., Botana, A.M., Botana,L.M., 2008. In vitro and in vivo evaluation of paralytic shellfish poisoningtoxin potency and the influence of the pH of extraction. Anal. Chem. 80, 1770–1776.
Van Dolah, F.M., 2000. Marine algal toxins: origins, health effects, and theirincreased occurrence. Environ. Health Perspect. 108, 133–141.
Villalba, A., Mourelle, S.G., Carballal, M.J., López, C., 1997. Symbionts and diseases offarmed mussels Mytilus galloprovincialis throughout the culture process in theRías of Galicia (NW Spain). Dis. Aquat. Organ. 31, 127–139.
Volety, A.K., Chu, F.L.E., 1995. Suppression of chemiluminescence of eastern oyster(Crassostrea virginica) hemocytes by the protozoan parasite Perkinsus marinus.Dev. Comp. Immunol. 19, 135–142.
Wang, S., Peatman, E., Liu, H., Bushek, D., Ford, S.E., Kucuktas, H., Quilang, J., Li, P.,Wallace, R., Wang, Y., Guo, X., Liu, Z., 2010. Microarray analysis of geneexpression in eastern oyster (Crassostrea virginica) reveals a novel combination
ogical changes in the eastern oyster Crassostrea virginica infested with therium fundyense. J. Invertebr. Pathol. (2015), http://dx.doi.org/10.1016/

10851086108710881089
10901091109210931094
M. Lassudrie et al. / Journal of Invertebrate Pathology xxx (2015) xxx–xxx 13
YJIPA 6639 No. of Pages 13, Model 5G
11 February 2015
of antimicrobial and oxidative stress host responses after Dermo (Perkinsusmarinus) challenge. Fish Shellfish Immunol. 29, 921–929.
Xue, Q.G., Gauthier, J., Schey, K., Li, Y., Cooper, R., Anderson, R., La Peyre, J.F., 2012.Identification of a novel metal binding protein, Segon, in plasma of the easternoyster, Crassostrea virginica. Comp. Biochem. Physiol. Part B 163, 74–85.
1095
Please cite this article in press as: Lassudrie, M., et al. Physiological and patholtrematode Bucephalus sp. and exposed to the toxic dinoflagellate Alexandj.jip.2015.01.011
Yin, D., 1996. Biochemical basis of lipofuscin, ceroid, and age pigment-likefluorophores. Free Radic. Biol. Med. 21, 871–888.
Zaroogian, G., Voyer, R.A., 1995. Interactive cytotoxicities of selected organic andinorganic substances to brown cells of Mercenaria mercenaria. Cell Biol. Toxicol.11, 263–271.
ogical changes in the eastern oyster Crassostrea virginica infested with therium fundyense. J. Invertebr. Pathol. (2015), http://dx.doi.org/10.1016/










