
Effect of light intensity on maturation and spawning of ablated female Penaeus merguiensis
12
Effect of light intensity on maturation and spawning of ablated female Penaeus merguiensis Tung Hoang a, * , S.Y. Lee a , Clive P. Keenan b , Gay E. Marsden c a School of Environmental and Applied Sciences, Griffith University, PMB 50 GCMC, Queensland 9726, Australia b Department of Primary Industries, Bribie Island Aquaculture Research Centre, PO Box 2066, 144 North Street, Bribie Island, Queensland 4507, Australia c NFIEC, Locked Bag 5, Grafton, NSW 2460, Australia Received 10 September 2001; received in revised form 12 September 2001; accepted 19 September 2001 Abstract The effect of three levels of light intensity (BRIGHT: 1100 lux; DIM: 2 lux; and DARK: 0 lux) on ovarian maturation and spawning of Penaeus merguiensis was investigated. The proportion of spawners, latency period, spawning rate, fecundity, spawning interval and number of days spent at stage IV per spawn were not significantly different ( P > 0.05) among the three light intensity treatments. Compared to dim light, bright light did not adversely affect the maturation and reproductive performance of ablated prawns. However, the percentage of spawning at stage III (31.6%) and mortality (26.7%) were significantly higher in the DARK treatment ( P < 0.05). A trend of increasing fecundity with an increase in light intensity was observed. Also, the hypothesis of whether the prawn eye is the only light receptor involved in ovarian maturation and spawning was tested by covering the remaining eye of ablated prawns with prepared caps made from black heat- shrinkable tubing (polyolefin). These prawns were held under 1100 lux (BRI-COV treatment) and 2 lux (DIM-COV) with the DARK treatment as the control group. The proportion of spawners was lowest in the BRI-COV treatment and highest in the DARK treatment ( P < 0.05), whereas there was no significant difference between these two treatments and the DIM-COV treatment. The DARK treatment, however, had a significantly higher spawning rate ( P < 0.05). The significantly higher percentage of spawning at stage III and less time spent at stage IV per spawn of prawns in the DARK treatment ( P < 0.05) compared with the DIM-COV and BRI-COV treatments suggest that the prawn 0044-8486/02/$ - see front matter D 2002 Elsevier Science B.V. All rights reserved. PII:S0044-8486(01)00814-6 * Correspondence author. Bribie Island Aquaculture Research Centre, Queensland Department of Primary Industries, PO Box 2066 (144 North Street), Bribie Island, Qld 4507, Australia. Tel.: +61-7-3400-2020; fax: +61-7-3408-3535. E-mail address: [email protected] (T. Hoang). www.elsevier.com/locate/aqua-online Aquaculture 209 (2002) 347 – 358
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Effect of light intensity on maturation and spawning of ablated female Penaeus merguiensis

Effect of light intensity on maturation and spawning
of ablated female Penaeus merguiensis
Tung Hoang a,*, S.Y. Lee a, Clive P. Keenan b, Gay E. Marsden c
aSchool of Environmental and Applied Sciences, Griffith University, PMB 50 GCMC,
Queensland 9726, AustraliabDepartment of Primary Industries, Bribie Island Aquaculture Research Centre, PO Box 2066,
144 North Street, Bribie Island, Queensland 4507, AustraliacNFIEC, Locked Bag 5, Grafton, NSW 2460, Australia
Received 10 September 2001; received in revised form 12 September 2001; accepted 19 September 2001
Abstract
The effect of three levels of light intensity (BRIGHT: 1100 lux; DIM: 2 lux; and DARK: 0 lux)
on ovarian maturation and spawning of Penaeus merguiensis was investigated. The proportion of
spawners, latency period, spawning rate, fecundity, spawning interval and number of days spent at
stage IV per spawn were not significantly different (P> 0.05) among the three light intensity
treatments. Compared to dim light, bright light did not adversely affect the maturation and
reproductive performance of ablated prawns. However, the percentage of spawning at stage III
(31.6%) and mortality (26.7%) were significantly higher in the DARK treatment (P< 0.05). A trend
of increasing fecundity with an increase in light intensity was observed. Also, the hypothesis of
whether the prawn eye is the only light receptor involved in ovarian maturation and spawning was
tested by covering the remaining eye of ablated prawns with prepared caps made from black heat-
shrinkable tubing (polyolefin). These prawns were held under 1100 lux (BRI-COV treatment) and 2
lux (DIM-COV) with the DARK treatment as the control group. The proportion of spawners was
lowest in the BRI-COV treatment and highest in the DARK treatment (P < 0.05), whereas there was
no significant difference between these two treatments and the DIM-COV treatment. The DARK
treatment, however, had a significantly higher spawning rate (P< 0.05). The significantly higher
percentage of spawning at stage III and less time spent at stage IV per spawn of prawns in the DARK
treatment (P < 0.05) compared with the DIM-COV and BRI-COV treatments suggest that the prawn
0044-8486/02/$ - see front matter D 2002 Elsevier Science B.V. All rights reserved.
PII: S0044 -8486 (01 )00814 -6
* Correspondence author. Bribie Island Aquaculture Research Centre, Queensland Department of Primary
Industries, PO Box 2066 (144 North Street), Bribie Island, Qld 4507, Australia. Tel.: +61-7-3400-2020; fax:
+61-7-3408-3535.
E-mail address: [email protected] (T. Hoang).
www.elsevier.com/locate/aqua-online
Aquaculture 209 (2002) 347–358

eye may not be the only light receptor involving in ovarian maturation and spawning in P.
merguiensis. D 2002 Elsevier Science B.V. All rights reserved.
Keywords: Light intensity; Reproduction; Penaeus merguiensis; Light receptors; Fecundity
1. Introduction
Further understanding of the mechanism by which environmental factors control
ovarian maturation in prawns is necessary not only for the improvement of induction
technique in hatcheries, but also for the formulation of alternative techniques, e.g.
environmental manipulation, to replace the current traumatic eyestalk ablation technique
for inducing spawning of penaeids. Among the potentially important environmental
factors, light seems to be an important one. Its importance has been implicated by a
number of studies on the effects of light intensity, light quality and photoperiod on
maturation in Penaeus monodon (Pudadera and Primavera, 1981), P. stylirostris, P.
vannamei (Chamberlain and Lawrence, 1981; Wyban et al., 1987), and P. indicus
(Emmerson et al., 1983). Similar studies, however, have not been conducted on the
banana prawn P. merguiensis, a commercially cultured species.
Although unilateral eyestalk ablation is a reliable method of inducing maturation,
ablated prawns still need a suitable environment to reach full maturation and spawn.
Research on different penaeid species has shown that dim light is an important prerequisite
for maturation (Primavera, 1985; Browdy, 1992). In contrast, bright light is considered
unfavourable for maturation (Chamberlain and Lawrence, 1981). Our recent studies
revealed that strong light (1100 lux) inhibits ovarian maturation of unablated P. mer-
guiensis (Hoang et al., in press(a)). Since gonadal inhibitory hormone (GIH) is synthesized
in the eyestalk of crustacean decapods (see Fingerman, 1987 for a review) and the prawn’s
eyes are the visual organ, the production of GIH may be proportional to light intensity.
This speculation is supported by the results of research on ocypodid crabs. Nadarajalingam
and Subramoniam (1987) reported that neurosecretory cells on the eyestalk of Ocypoda
platytarsis and O. macrocera showed higher secretory activity under elevated light
intensity. In ablated prawns kept under strong light, the production of GIH in the re-
maining eye, although not high enough to inhibit ovarian maturation, may possibly retard
the process.
On the other hand, light seems to be essential for the maturation process in prawns.
Chamberlain and Lawrence (1981) reported that continuous dark treatment produced
lower survival and lower maturation success in P. stylirostris than dim light treatment. A
continually dark environment is believed to interfere with photoperiod-dependent pro-
cesses, which may regulate reproduction of nonhuman animals (Reiter, 1991). Under
natural conditions, wild prawns mature and spawn in deep waters (Chong, 1980; Motoh,
1981). Light intensity at the maturation and spawning grounds, therefore, is probably
weak, but not completely absent (Wurts and Stickney, 1984). These studies in
combination suggest that ovarian maturation in penaeids requires the presence of light
at low intensity. Furthermore, since dim light favours maturation, whereas bright light or
T. Hoang et al. / Aquaculture 209 (2002) 347–358348

continuous dark condition are unfavourable for ovarian maturation, it is suggested that
the prawn eyes may not be the only light receptor related to the maturation process in
prawns.
If the eyes are the only receptor for light, maturation and spawning performance of
unilaterally ablated prawns that have their remaining eye covered and kept under dim or
bright light should not be different from those kept in continuous dark condition. This
study investigated the effect of light intensity on maturation of unilaterally ablated banana
prawn females, P. merguiensis, and examined whether the remaining eye of ablated
prawns is the only receptor for light.
2. Methods and materials
2.1. Broodstock source
Wild banana prawns were collected from local waters in southeast Queensland, Australia
in May 1999. The prawns were then stocked in a 200-m2 covered pond over winter, and
fed with EBI STAR pellet (50% protein; Higashimaru, Japan) twice a day at a rate of
5% of prawn biomass. Prawn females were randomly selected from the covered pond in
November 1999. The prawns were acclimatized in a 12-m3 maturation tank for a week
before the experiment started. From the acclimatization tank, prawn females were
selected for the experiment. All prawns were healthy in appearance. Injured and
postmoult prawns were excluded. Prawn size was similar among the treatments (Table
1).
2.2. Experimental design and conditions
The experiment consisted of five treatments, as described in Table 1, with three
replicate tanks; each tank stocked with five prawns. Light was provided by daylight
fluorescent tubes (OSRAM 36 W, Australia), suspended above the experimental tanks. For
the DIM treatment, the light tubes were wrapped in green 70% ‘‘shade-cloth’’ (Dindas
Lew Cat. No. 5c7036 BL) to reduce light intensity. Light intensity was measured at the
water surface by a photometer (Model 185B, LI-COR, USA). Water depth was 40 cm in
Table 1
Summary of prawn size at stocking and light treatments
Treatments CL (mm) W (g) Descriptions
DIM 41.6F 0.4 42.3F 0.1 2 lux
DIM-COV 41.0F 0.1 42.9F 0.9 2 lux + the remaining eye covered
BRI 41.1F 0.2 41.2F 0.6 1100 lux
BRI-COV 41.4F 0.4 42.7F 1.5 1100 lux + the remaining eye covered
DARK 40.9F 0.6 41.4F 0.7 0 lux
Data are meansF 1 S.E. No significant difference ( P > 0.5) in size was found among treatments.
T. Hoang et al. / Aquaculture 209 (2002) 347–358 349

the experimental tanks, which were 1-m3 fiberglass tanks, with the inside painted in
black. For the DARK treatment, the experimental tanks had no overhead lights and were
covered by 200-Am-thick black plastic sheet. Except for the DARK treatment, photo-
period was 12 h light:12 h dark, controlled by a digital timer. Light went on at 5:00 h
and off at 17:00 h with 15 min of ramping. The experiment used a flow-through system.
Filtered (5 Am) seawater was heated to 27 jC and exchanged at a rate of 350% per 24 h.
Water quality was checked daily with the Horiba U-10 Water Checker (Horiba, Japan).
During the 18-day experimental period, the mean values (F 1S.D.) of temperature,
salinity, pH and dissolved oxygen were 27.1F 0.2 jC, 34.2F 0.3 ppt, 8.1F 0.1, and
5.2F 0.3 mg/l, respectively.
Eyestalk ablation involved ligating (tying a thread around the base of the eyestalk),
slitting the eyeball and squeezing out the contents. The remaining eye of ablated prawns in
the DIM-COVand BRI-COV treatments was then covered with a prepared cap made from
black heatshrinkable tubing (polyolefin, US Mil Spec. MIL-I-23053/5, Dicksmith,
Australia) (Fig. 1). The cap was first filled with cold (16–18 jC) seawater. This was toprovide a buffer preventing stress or possible damage to the prawn eye from heating (to
shrink the opening of the cap). Then, the remaining eye of the prawns was inserted into
the cap. Heat generated by a small-tipped, temperature-controlled soldering iron (RS 545-
008, Germany) was quickly applied to shrink the opening of the cap around the eyestalk.
The whole process took approximately 30 s, 15 s for eyestalk ablation and 15 s for
covering the remaining eye. During this handling process, the prawn body was kept in
cold seawater (16–18 jC) to reduce stress. Prawns that had the remaining eye intact were
kept in cold seawater for an addition of 15 s. Preliminary tests in our laboratory showed
Fig. 1. Prawn with the remaining eye covered by the eye cap.
T. Hoang et al. / Aquaculture 209 (2002) 347–358350

that the prepared cap effectively blocked light from the prawn eye. When their covered
eyes were illuminated by a light source, the prawns did not flick nor move away like
those with the remaining eye intact. Further, for this study, prawns that showed abnormal
behaviour (being sluggish or swimming spirally after their eyes were covered) were
excluded from the experiment and replaced by another one.
Prawns were fed at a feeding rate of 10% of prawn biomass with the BIARC moist
maturation diet (a mixture of fresh frozen green mussels, squids, calf liver and other essential
nutrients, 54.6% crude protein; Marsden et al., 1997) twice a day at 10:00 and 17:00 h. This
was supplemented at a rate of 2% of prawn biomass by EBI STAR pellets, fed at 17:00 h
only. Unused feed and faeces were siphoned out of tanks prior to feeding.
Ovarian maturation was examined externally, using a submersible flashlight, following
the descriptions by Tuma (1967) and Primavera (1985). Since all prawns were maturing,
ovarian maturation included four stages: early maturing (stage II), late maturing (stage III),
ripe (stage IV) and spent (stage V). To avoid undue disturbance to the prawns, the
flashlight was wrapped with red cellophane sheet. Prawns were checked daily at 17:30 h.
Prawn females at stage IV were moved to 150-l spawning tanks kept totally dark and
returned to the corresponding experimental tanks early the following morning. In the
DARK treatment, spawning at stage III was fortunately detected right at the beginning of
the experiment. Therefore, prawn females at stage III in this treatment were also removed
to the spawning tanks. Fecundity was estimated by vigorously agitating the tank water to
ensure a homogenous suspension of eggs, then immediately taking four 100-ml sub-
samples of water from the spawning tank and counting the number of eggs under a stereo
microscope. The experiment continued for 18 days from eyestalk ablation.
2.3. Statistical analysis
The response variables for statistical analyses included latency period (number of days
from ablation to spawning. Prawns that did not spawn were assigned a latency period of 18
days, i.e. the length of the experiment to facilitate statistical analysis), proportion of
spawners, number of spawns produced per female per prawn * day (prawn * day is the
number of days that the prawn was held under the treatment conditions, presumably
capable of spawning), fecundity (number of eggs per spawn), ovarian stage at spawning,
number of days spent at stage IV per spawn at this final stage, mortality, specific growth
rate (SGR), number of moults and moult interval.
Experimental data were analyzed with Genstat (version 4.1 for Windows) (Payne et al.,
1993), for one-way analysis of variance (ANOVA) with the least significant difference
(LSD) multiple-range procedure. All data were checked for the assumptions of normality
and homoscedasticity before the application of the procedure. Generalized linear models
(McCullagh and Nelder, 1989) were used to analyze binomial data such as proportion of
spawners and mortality. Tanks were used as observation units for statistical analysis. For
each variable, two major tests were conducted. The first test was conducted with data from
DARK, DIM and BRI treatments to reveal the effect of light intensity. Data from BRI-
COV, DIM-COV and DARK treatments were used in the second test to examine whether
the remaining eye of the ablated prawns is the only receptor for light related to the
maturation process.
T. Hoang et al. / Aquaculture 209 (2002) 347–358 351

3. Results
3.1. Effects of light intensity on maturation of ablated prawns
Light intensity did not affect the maturation and spawning performance of ablated
prawns. No significant difference (P < 0.05) was found among the BRI, DIM and DARK
treatments in terms of proportion of spawners, latency period, number of spawn per female
per prawn * day, fecundity and spawning interval (Table 2). The interval between two
successive spawns within an intermoult period was usually 72 h. When moulting occurred
in between successive spawning events, spawning interval extended to 5–9 days and was
not significantly different among the treatments. Although not significantly (P>0.05)
increased, fecundity showed an improvement with the increase of light intensity. Mean
fecundity of prawns in the DARK treatment was only 95.4� 103, while it was
111.3� 103 and 132.6� 103 in the DIM and the BRI treatments, respectively (Table 2).
A significantly higher percentage of spawning at stage III of ovarian maturation and
higher mortality were observed in the DARK treatment (Table 2). Of the total number of
spawns produced in this treatment, 31.6% occurred at stage III of ovarian maturation,
which was unusually early. In addition, the number of days spent at stage IV of ovarian
maturation for each spawning at this final stage was smaller in the DARK treatment (Table
2). Generally, it was observed that prawns in the DARK treatment did not necessarily
develop their ovaries to full size (i.e. the ovary at the first abdominal segment did not
exhibit a diamond shape with two wings expanding laterally) to spawn as did in the BRI
and DIM treatments. The DARK treatment also had the highest mortality (P < 0.05).
During the experimental period, 26.7% of prawns in the DARK treatment died, while only
6.7% and nil died in the BRI and DIM treatments, respectively.
3.2. The eye as the only receptor for light?
The second hypothesis was tested by comparing results from the DARK, BRI-COVand
DIM-COV treatments. Results (Table 3) showed that the remaining eye of ablated prawn
Table 2
Reproductive performance of ablated prawns under different light intensities
Response variable Treatment (lux)
BRI (1100) DIM (2) DARK (0)
Proportion of spawners (%) 93.3F 5.7 93.3F 5.7 100.0
Latency period (days) 6.7F 0.7 8.0F 0.7 6.4F 0.3
Number of spawns per female per prawn * day 0.124F 0.005 0.107F 0.013 0.146F 0.013
Fecundity (� 103 eggs) 132.6F 11.7 111.3F 8.5 95.4F 18.2
Percentage of spawning at stage III (%) 0.0b 0.0b 31.6F 3.8a
Number of days at stage IV per spawn 1.6F 0.26 1.7F 0.03 1.1F 0.10
Interval between spawns (days) 4.0F 0.5 4.6F 0.5 4.4F 0.9
Mortality (%) 6.7F 6.6a 0.0a 26.7F 11.7b
Data are meansF 1 S.E. Within row means that have different superscripts are significantly different ( P < 0.05).
T. Hoang et al. / Aquaculture 209 (2002) 347–358352
might not be the only light receptor involved in the reproduction of prawn. The proportion
of spawners number of spawns per female per prawn * day, percentage of spawning at
stage III and number of days at stage IV per spawn were significantly different among the
Table 3
Reproductive performance of prawns that had the remaining eye blocked from light under different light
intensities. Data are meansF 1 S.E. Within row means that have different superscripts are significantly different
( P< 0.05)
Response variable Treatment (lux)
DARK (0) DIM-COV (2) BRI-COV (1100)
Proportion of spawners (%) 100.0a 88.2F 6.6ab 66.7F 10.2b
Latency period (days) 6.4F 0.3 8.1F1.3 9.0F 2.0
Number of spawns per female per prawn * day 0.146F 0.013a 0.099F 0.010b 0.083F 0.014b
Fecundity (� 103 eggs) 95.4F 18.2 128.1F 3.5 123.5F 13.9
Interval between spawns (days) 4.4F 0.9 3.3F 0.1 4.4F 1.3
Percentage of spawning at stage III (%) 31.6F 3.8a 0.0b 0.0b
Number of days at stage IV per spawn 1.1 F 0.1 a 2.8F 0.7b 2.1F 0.3b
Mortality (%) 26.7F 13.9 17.6F 11.2 40.0F 15.4
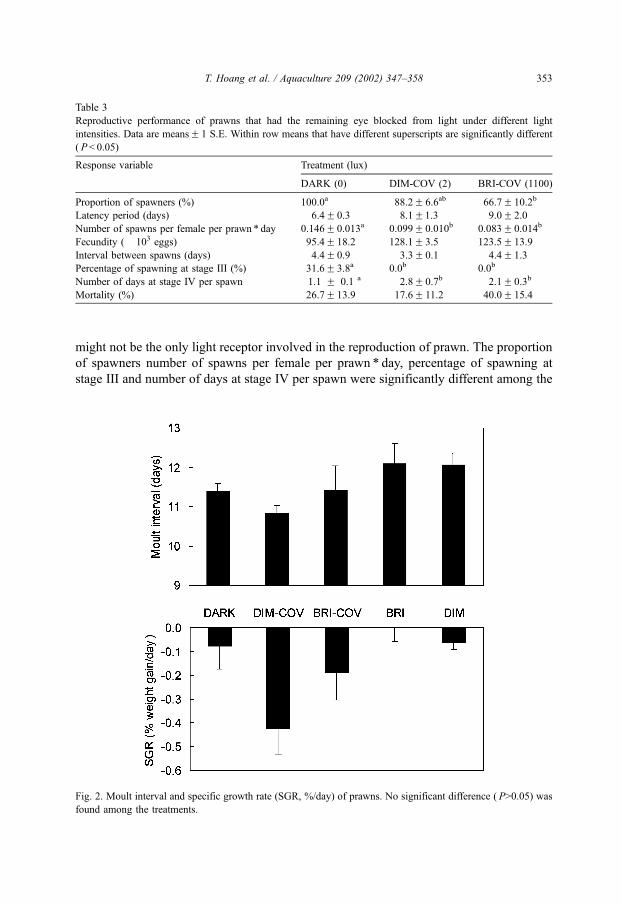
Fig. 2. Moult interval and specific growth rate (SGR, %/day) of prawns. No significant difference ( P>0.05) was
found among the treatments.
T. Hoang et al. / Aquaculture 209 (2002) 347–358 353

three treatments (P < 0.05). The proportion of spawners and mean number of spawns per
female per prawn * day were highest in the DARK treatment and lowest in the BRI-COV
treatment, whereas there was no significant difference between these treatments and the
DIM-COV treatment. Again, prawns in the DARK treatment had a higher percentage of
spawning at stage III of ovarian maturation and spent significantly less time at stage IV for
each spawning at this final stage (P < 0.05). The latency period, fecundity, interval
between spawns and mortality were not significantly different among the three treatments
(P < 0.05) (Table 3).
3.3. Moulting and growth of prawns
Moulting activity was similar across the five treatments. The mean numbers of moults
in the DARK, DIM, BRI, DIM-COV and BRI-COV treatments were 1.5, 1.7, 1.7, 1.6 and
1.6, respectively. Moult interval varied from 9 to 15 days and was not significantly (P>
0.05) different among the treatments (Fig. 2). Specific growth rate (SGR) was negative in
all treatments except the BRI treatment, and were not significantly (P>0.05) different
among the treatments (Fig. 2).
4. Discussion
4.1. The effects of light intensity on spawning and maturation of ablated prawns
This current study demonstrated that light intensity did not affect ovarian maturation
rate and spawning performance of ablated female P. merguiensis (Table 2). Bray and
Lawrence (1992) also reported that, when acclimated to a particular light level, ablated P.
vannamei as well as other species will reproduce over a broad range of light levels. In
contrast, our recent studies on unablated female P. merguiensis (conducted in the same
conditions, using the same source of broodstock) showed that light intensity has strong
effects on all these variables (Hoang et al., in press (a)). For unablated prawns, bright light
(1100 lux) inhibited the progression of ovarian maturation to the final stage (stage IV),
therefore halting spawning. The relevance of these light intensities to the natural environ-
ments of prawn have been discussed in that paper. The results of this study indicate that
when other critical environmental factors, e.g. temperature, salinity and pH, are favour-
able, the effect of light intensity is secondary to eyestalk ablation in controlling most
aspects of reproduction of ablated prawns. The light intensities tested in this study (0, 2
and 1100 lux) are more relevant to the light conditions in commercial hatcheries rather
than those in the natural environment since all prawns were ablated. The fact that bright
light did not adversely affect ovarian maturation and spawning of ablated P. merguiensis
may indicate a possibility to raise light intensity (which is usually kept very dim) in P.
merguiensis hatcheries to avoid problems associated with working in dark or dim
conditions. However, further research is needed to look at larval quality, which was not
assessed in this study, before the application of this finding.
The DARK treatment in this study had significantly higher mortality (P < 0.05). This
higher mortality was probably due to reproductive exhaustion (Primavera, 1985; Harrison,
T. Hoang et al. / Aquaculture 209 (2002) 347–358354

1990; Palacios et al., 1999). On average prawns in this treatment produced 2.53 spawns
over 18 days; some spawned up to five times. Further, mortality of ablated prawns is
common (for review, see Primavera, 1985; Bray and Lawrence, 1992). Chamberlain and
Lawrence (1981) also reported higher mortality (up to 60%) and lower maturation success
of ablated P. stylirostris and P. vannamei kept in a continuous dark environment compared
with their sky light and dim treatments. This study confirmed their finding on survivor-
ship, but not maturation in ablated P. merguiensis. The proportion of spawners was 100%
in the DARK treatment. However, higher percentage of spawning at stage III, which
probably affects the viability of the offspring, higher mortality and inferior fecundity were
the problems with the continuous dark treatment. In natural conditions, wild prawns
mature and spawn in deep waters (Chong, 1980; Motoh, 1981). Light intensity at the
maturation and spawning grounds is weak, but not completely absent (Wurts and Stickney,
1984). It is suggested that ovarian maturation in penaeids requires the presence of light at
low intensity (or the existence of a photoperiod) since continuous darkness is unnatural
and may interfere with photoperiod-dependent processes.
Fecundity of prawns in the DARK treatment, although not significantly lower, was
inferior (95.4� 103) compared to the DIM and BRI treatments (111.3� 103 and
132.6� 103, respectively). This might relate to the significantly higher percentage of
spawning at stage III (31.6%) the DARK treatment. According to Yano (1995), ovarian
maturation of penaeids consists of two major processes: vitellogenesis and final maturation
of oocytes. Prawn ovaries usually must develop to full maturation (stage IV) to spawn.
Stage III of ovarian maturation is equivalent to the late vitellogenic stage (Tan-Fermin and
Pudadera, 1989; Medina et al., 1996). Since no histological work was conducted in this
current study, it is uncertain whether vitellogenesis was completed when ovulation took
place in these early spawning events. Another possibility is that final maturation of prawns
in the DARK treatment was accelerated, taking only a few hours from checking time
(17:00 h) to spawning time (usually between 19:00 and 22:00 h). However, this possibility
is unlikely in P. merguiensis, since the spawning interval was not significantly different
among the treatments. In P. japonicus, final maturation of oocytes occurs after dark,
immediately followed by spawning (Yano, 1995).
Excluding spawns that occurred at stage III, the number of days that prawn ovaries
remained at stage IV per ‘‘normal’’ spawn (spawn at stage IV) was shorter in the DARK
treatment (1.1 day) than in the DIM and BRIGHT treatments (1.6–1.7 days). Prawns held
longer at stage IV by the presence of light, spawned with favourable fecundity. These
observations suggest a possible controlling effect of light on the final maturation of
oocytes or ovulation. Our recent study (Hoang et al., in press (b)) on the effect of light
intensity on spawning of P. merguiensis supports this hypothesis. Spawning of stage-IV
prawns held under 1100 lux (since stocked in spawning tanks at 18:00 h) was delayed for
up to 21 min and produced favourable fecundity (111.2� 103) compared with the control
group (completely dark; 95.0� 103). Thus, the presence of light at the final stages of
ovarian maturation seemed to have retarded the onset of ovulation, allowing more eggs to
ripen. However, the controlling mechanism is unknown. If this hypothesis is true, the role
of light receptor(s) is critical in conveying the signal of light to the central nervous
system. In this regard, the visual organ, i.e. the eyes are expected to play an important
role.
T. Hoang et al. / Aquaculture 209 (2002) 347–358 355

4.2. The existence of additional light receptors?
As mentioned before, if the remaining eye of ablated prawns is the only receptor for
light, results obtained from the DARK, DIM-COV and BRI-COV treatments should be
similar. However, the proportion of spawners, spawning rate and number of days at stage
IV per spawn were significantly different among the treatments, indicating that the eye
may not be the only receptor for light involved in ovarian maturation. Although the eye
cap and the covering procedure might have contributed to the lower proportion of
spawners and lower spawning rate of prawns in the DIM-COV and BRI-COV treatments,
nonsignificant differences in mortality among the three treatments suggest that any
confounding effect of the eye cap, if it existed, was marginal. Mortality in the DIM-COV
and BRI-COV treatments occurred mainly at moulting. It is unknown whether vigorous
flicking action of the prawn body (including the eyestalk) during moulting or the absence
of light from the prawn eye (where the source of moult inhibiting hormones is located)
was the cause of these mortalities. Since bilateral eyestalk ablation often results in high
mortality (up to 100%, for review, see Primavera 1985; Bray and Lawrence, 1992),
covering the prawn eye is obviously a better option. Although the prepared eye cap used
in this study is not yet an ideal means, it provided an opportunity to preliminarily test the
hypothesis of additional light receptor(s). Further improvement of the eye cap or the
development of a better means of manipulating light intensity reaching the intact eye
deserves more attention. Moreover, the fecundity of prawns in the DIM-COV and BRI-
COV treatments was comparable to the DIM and BRI treatments (the DIM-COV
treatment even had a favourable fecundity compared with the DIM treatment). This
trend suggests that the absence of light (in the DARK treatment) was probably the main
cause of unfavourable fecundity. Furthermore, with light being blocked from the
remaining eye, prawns in the DIM-COV and BRI-COV treatments spent significantly
more time at stage IV per spawn than the DARK treatment. Since fecundity and the
number of days at stage IV per spawns were comparable among the DIM, BRI, DIM-
COV and BRI-COV treatments, there is possibly another light receptor or receptors apart
from the remaining eye, that is involved in conveying light signals and controlling the
onset of ovulation. The existence of additional photoreceptors, apart from the compound
eyes, in crustaceans has been reviewed by Wiersma et al. (1982). According to these
authors, possible additional photoreceptors include the bilaterally paired caudal photo-
receptors of the sixth abdominal ganglion, certain regions of the body, e.g. the cephalic
region or the uropods. These photoreceptors relate to locomotory movement of crusta-
ceans. No information on possible linkage between these and ovarian maturation is
available. Nadarajalingam and Subramoniam (1987) also reported that neurosecretory
cells in both the eyestalk and thoracic brain of O. platytarsis and O. macrocera showed
higher secretory activities under elevated light intensity. The slightly longer time spent at
stage IV in the DIM-COV and BRI-COV treatments compared to the DIM and BRI
treatments may have resulted from a decrease of production of some essential hormones
(originated from the eyestalk), which may play an important role in regulating the transfer
of nutrients from storage tissues to the ovary. In prawns, the hormones that control final
maturation are still under investigation. This research suggests possible existence of a
hormone that controls spawning when vitellogenesis is completed and the action of that
T. Hoang et al. / Aquaculture 209 (2002) 347–358356

hormone may be triggered by dark conditions. This would explain why prawns only
spawn at night.
The negative growth of ablated prawns observed in this study was possibly a result of
excessive maternal mobilization of nutrients and energy for the formation of eggs
(Harrison, 1990). In nature, nutrients must be available for first, metabolism, second,
growth, and third, reproduction (Bray and Lawrence, 1992); and the process of ovarian
maturation is likely to be gradual (Harrison, 1990). However, since eyestalk ablation
promotes rapid ovarian maturation and spawning, available nutrients are directed into
reproduction and/or maintenance, not growth. Very short rematuration periods (i.e. 3 days
between two successive spawns within an intermoult period of 4 days on average in this
study), high spawning rate and high metabolism require the ablated prawns to utilize their
energy reserves (for review, see Harrison, 1990). Thus, it is understandable that somatic
growth was severely compromised by repeated frequent spawning. Negative growth has
been reported in ablated P. monodon (Emmerson, 1983) and ablated P. indicus (Emmer-
son, 1980; Read and Coulton, 1980, cited by Quinitio and Millamena, 1992; Emmerson et
al., 1983), and in P. monodon commercial hatcheries, where ablated P. monodon were
induced to produce multiple spawnings (Ta Khac Thuong, Nguyen Dinh Phuc, personal
communication).
The controlling mechanism of light on maturation and ovulation needs to be inves-
tigated in relation to possible receptors and/or neurotransmitters that are involved in the
process. Further understanding in this area will then help to develop alternative methods to
induce maturation and spawning in penaeids.
Acknowledgements
We would like to thank Mrs. Nguyen Thi Anh Tuyet for her untiring assistance in the
laboratory; Dr. David Mayer, QDPI statistician, for his comments on the experimental
design and statistical analyses. This study was supported by Griffith University and Brbie
Island Aquaculture Research Centre, Department of Primary Industries, Australia.
References
Bray, W.A., Lawrence, A.L., 1992. Reproduction of Penaeus species in captivity. In: Fast, A.W., Lester, L.J.
(Eds.), Marine Shrimp Culture: Principles and Practices. Elsevier, Amsterdam, pp. 93–170.
Browdy, C.L., 1992. A review of the reproductive biology of Penaeus species: perspectives on controlled shrimp
maturation systems for high quality nauplii production. In: Wyban, J. (Ed.), Proceedings of the Special
Session on Shrimp Farming, 22–25 May 1992, Orlando, FL, USA. World Aquaculture Society, LA, USA,
pp. 22–51.
Chamberlain, G.W., Lawrence, A.L., 1981. Effect of light intensity and male and female eyestalk ablation on
reproduction of Penaeus stylirostris and P. vannamei. J. World Maric. Soc. 12, 357–372.
Chong, V.C., 1980. Maturation and breeding ecology of the white prawn Penaeus merguiensis de Man. In:
Furtado, J.I. (Ed.), Tropical Ecology and Development. Proceedings of the 5th International Symposium of
Tropical Ecology, 16–21 April 1979, Kuala Lumpur, Malaysia, pp. 1175–1184.
Emmerson, W.D., 1980. Induced maturation of prawn Penaeus indicus. Mar. Ecol.: Prog. Ser. 2, 121–131.
Emmerson, W.D., 1983. Muturation and growth of ablated and unablated Penaeus monodon Fabricius. Aqua-
culture 32, 235–241.
T. Hoang et al. / Aquaculture 209 (2002) 347–358 357

Emmerson, W.D., Hayes, D.P., Ngonyame, M., 1983. Growth and maturation of Penaeus indicus under blue and
green light. S. Afr. J. Zool. 18, 71–75.
Fingerman, M., 1987. The endocrine mechanism of crustaceans. J. Crustacean Biol. 7, 1–24.
Harrison, K.E., 1990. The role of nutrition in maturation, reproduction and embryonic development of decapod
crustaceans: a review. J. Shellfish Res. 9, 1–28.
Hoang, T., Lee, S.Y., Keenan, C.P., Marsden, G.E, in press (a). Ovarian maturation of the banana prawn, Penaeus
merguiensis de Man under different light intensities. Aquaculture.
Hoang, T., Lee, S.Y., Keenan, C.P., Marsden, G.E., in press (b). Spawning behaviour of Penaeus (Fennerope-
naeus) merguiensis de Man and the effect of light intensity on spawning. Aquacult. Res.
Marsden, G.E., McGuren, J.J., Hansford, S.W., Burke, M.J., 1997. A moist artificial diet for prawn broodstock: its
effect on the variable reproductive performance of wild caught Penaeus monodon. Aquaculture 149,
145–156.
McCullagh, P., Nelder, J.A., 1989. Generalized Linear Models. 2nd edn. Chapman & Hall, London, 551 pp.
Medina, A., Vila, Y., Mourente, G., Rodrıguez, A., 1996. A comparative study of the ovarian development in wild
and pond-reared shrimp, Penaeus kerathurus (Forskal, 1775). Aquaculture 148, 63–75.
Motoh, H., 1981. Studies on the fisheries biology of the giant tiger prawn, Penaeus monodon in the Philippines,
Technical Report, No.7. SEAFDEC Aquaculture Department, Tigbauan, Iloilo, 128 pp.
Nadarajalingam, K., Subramoniam, T., 1987. Influence of light on endocrine system and ovarian activity in the
ocypodid crabs Ocypoda platytarsis and O. macrocera. Mar. Ecol.: Prog. Ser. 36, 43–53.
Palacios, E., Perez-Rostro, C.I., Ramirez, J.L., Ibarra, A.M., Racotta, I.S., 1999. Reproductive exhaustion in
shrimp (Penaeus vannamei) reflected in larval biochemical composition, survival and growth. Aquaculture
171, 309–321.
Payne, R.W., Lane, P.W., Digby, P.G.N., Harding, S.A., Leech, P.K., Morgan, G.W., Todd, A.D., Thompson, R.,
Tunnicliffe, W.G., Welham, S.J., White, R.P., 1993. Genstat 5 Release 3 Reference Manual. Clarendon Press,
Oxford, 796 pp.
Primavera, J.H., 1985. A review of maturation and reproduction in closed thelycum penaeids. In: Taki, Y.,
Primavera, J.H., Llobrera, J.A. (Eds.), Proceedings of the First International Conference on the Culture of
Penaeid Prawns/Shrimps; 1984 December 4–7; Illoilo City, Philippines, pp. 47–64.
Pudadera, R.A., Primavera, J.H., 1981. Effect of light quality and eyestalk ablation on ovarian maturation in
Penaeus monodon. Kalikasan, Philipp. J. Biol. 10, 231–241.
Quinitio, E.T., Millamena, O.M., 1992. Ovarian changes and female specific protein levels during sexual matu-
ration of the white shrimp Penaeus indicus. Isr. J. Aquacult.-Bamidgeh 44, 7–12.
Reiter, R.J., 1991. Pineal gland: interface between the photoperiodic environment and the endocrine system.
Trends Endocrinol. Metab. 2, 13–19.
Tan-Fermin, J.D., Pudadera, R.A., 1989. Ovarian maturation stages of the giant tiger prawn, Penaeus monodon
Fabricius. Aquaculture 77, 229–242.
Tuma, D.J., 1967. A description of the development of primary and secondary sexual characters in the banana
prawn, Penaeus merguiensis de Man (Crustacea: Decapoda: Penaeinae). Aust. J. Mar. Freshwater Res. 18,
73–88.
Wiersma, C.A.G., Roach, J.L.M., Glantz, R.M., 1982. Neural integration in the optic system. In: Sandeman, D.C.,
Atwood, H.L. (Eds.), The Biology of Crustacea: Vol. 4. Neural Integration and Behavior. Academic Press,
New York, pp. 1–31.
Wurts, W., Stickney, R., 1984. An hypothesis on the light requirements for spawning penaeid shrimp, with
emphasis on Penaeus setiferus. Aquaculture 41, 93–98.
Wyban, J.A., Lee, C.S., Sweeney, J.N., Richards, W.K., 1987. Observations on development of a maturation
system for Penaeus vannamei. J. World Aquacult. Soc. 18, 198–200.
Yano, I., 1995. Final oocyte maturation, spawning and mating in penaeid shrimp. J. Exp. Mar. Biol. Ecol. 193,
113–118.
T. Hoang et al. / Aquaculture 209 (2002) 347–358358




















