
Annexin expressions are temporally and spatially regulated during rat hepatocyte differentiation
Upload
independentCategory
view
2download
0
IMF YJMBI-64434; No. of pages: 14; 4C: 4, 8
Article
Ann Kari Grind3
0022-2836/© 2014 Elsevi
Please cite this article aNuclear Localisation an
Effect of Serine Phosphorylation and Ser25Phospho-Mimicking Mutations on NuclearLocalisation and Ligand Interactions ofAnnexin A2
heim1, 2, Hanne Hollås1, J
uan Ramirez3, Jaakko Saraste1, 2,Gilles Travé and Anni Vedeler 11 - Department of Biomedicine, University of Bergen, N-5009 Bergen, Norway2 - Molecular Imaging Center (MIC), University of Bergen, N-5009 Bergen, Norway3 - Biotechnologie et Signalisation Cellulaire UMR 7242, Ecole Supérieure de Biotechnologie de Strasbourg, F-67412 Illkirch, France
Correspondence to Anni Vedeler: Jonas Lies Vei 91, N-5009 Bergen, Norway. [email protected]://dx.doi.org/10.1016/j.jmb.2014.04.019Edited by P. Griffin
Abstract
Annexin A2 (AnxA2) interacts with numerous ligands, including calcium, lipids, mRNAs and intracellular andextracellular proteins. Different post-translational modifications participate in the discrimination of thefunctions of AnxA2 by modulating its ligand interactions. Here, phospho-mimicking mutants (AnxA2-S25E andAnxA2-S25D) were employed to investigate the effects of Ser25 phosphorylation on the structure and functionof AnxA2 by using AnxA2-S25A as a control. The overall α-helical structure of AnxA2 is not affected by themutations, since the thermal stabilities and aggregation tendencies of the mutants differ only slightly from thewild-type (wt) protein. Unlike wt AnxA2, all mutants bind the anxA2 3′ untranslated region and β-γ-G-actin withhigh affinity in a Ca2+-independent manner. AnxA2-S25E is not targeted to the nucleus in transfected PC12cells. In vitro phosphorylation of AnxA2 by protein kinase C increases its affinity to mRNA and inhibits its nuclearlocalisation, in accordancewith the data obtainedwith the phospho-mimickingmutants. Ca2+-dependent bindingof wt AnxA2 to phosphatidylinositol, phosphatidylinositol-3-phosphate, phosphatidylinositol-4-phosphate andphosphatidylinositol-5-phosphate, as well as weaker but still Ca2+-dependent binding to phosphatidylserine andphosphatidylinositol-3,5-bisphosphate, was demonstrated by a protein–lipid overlay assay, whereas binding ofAnxA2 to these lipids, aswell as its binding to liposomes, is inhibited by theSer25mutations. Thus, introduction ofa modification (mutation or phosphorylation) at Ser25 appears to induce a conformational change leading toincreased accessibility of the mRNA- and G-actin-binding sites in domain IV independent of Ca2+ levels, whilethe Ca2+-dependent binding of AnxA2 to phospholipids is attenuated.
© 2014 Elsevier Ltd. All rights reserved.
Introduction
Annexin A2 (AnxA2), a multifunctional ~39-kDaprotein of the annexin superfamily, was originallycharacterised by its ability to bind negatively chargedphospholipids in a Ca2+-dependent manner [1–5].Structurally, AnxA2 harbours the conserved annexincore structure containing four domains and adoptingthe shape of a slightly curved disc with a concaveand a convex surface. The N-terminal end of AnxA2,located along the concave surface of the corestructure [6], folds into a structurally separate domainwhen bound to the EF-hand protein S100A10 [7].However, from Pro20 (counting the first Ser as 1throughout the text), it is likely to establish interactions
er Ltd. All rights reserved.
s: Grindheim Ann Kari, et al, Effect of Serind Ligand Interactions of Annexin A2, J Mol
with hydrophobic residues on the surface of the corestructure and not with interior residues as observed inAnxA1 [8,9].In addition to calcium and phospholipids that bind
to the convex surface of the core structure [1,2],interactions of AnxA2with, for example, G-actin [10,11]and/or F-actin [12], specific mRNAs [13], tissueplasminogen activator and plasminogen/plasmin [14]have been reported. S100A10 is an important majorligand. Two copies of AnxA2 can bind to an S100A10homodimer and thus form an AnxA2-S100A10 hetero-tetramer. While monomeric AnxA2 is preferentiallyfound soluble in the cytosol, formation of the hetero-tetrameric complex leads to translocation of AnxA2to the plasma membrane or intracellular membrane
J. Mol. Biol. (2014) xx, xxx–xxx
e Phosphorylation and Ser25 Phospho-Mimicking Mutations onBiol (2014), http://dx.doi.org/10.1016/j.jmb.2014.04.019

2 Serine 25 phosphorylation of Annexin A2
compartments, often to sites of membrane–cytoskel-eton contact [11,15,16]. In addition, AnxA2 has alsobeen observed in the nucleus [17], in the extracellularmatrix [18] and at cell–cell contact sites [19]. The lattermay involve both intracellular and extracellularligands.AnxA2 is a substrate for the Tyr kinase Src [20]
and the Ser/Thr protein kinase C (PKC) [21], as wellas other kinases [1]. The most commonly employedphosphorylation sites appear to be Ser11 [22], Tyr23[23] and Ser25 [21], all located in the N-terminaldomain. Ser25 and Tyr23 phosphorylations appearto be mutually exclusive [21,24].Although the physiological roles of phosphoryla-
tion of AnxA2 are still incompletely understood,several studies have suggested that phosphoryla-tion can affect the localisation of the protein andthereby regulate its cellular functions. Phosphory-lation of AnxA2 by PKC, most likely on Ser25, mayinduce its translocation from the cytoplasm to thecell periphery with concomitant secretion of cate-cholamines, thus suggesting a regulatory role of Serphosphorylation of AnxA2 in Ca2+-dependent exo-cytosis [25–28]. By contrast, Ser phosphorylationprevents the translocation of AnxA2 to the extracel-lular cell surface [24], since Ser phosphorylation, inparticular of Ser11, can dissociate/prevent the forma-tion of the AnxA2-S100A10 heterotetrameric complex[22,24,29,30], and S100A10 is required for thetranslocation of AnxA2 to the cell periphery [24,31].Furthermore, Ser phosphorylation prevents Tyr phos-phorylation, which is a requisite for externalisationof AnxA2. However, another reported effect of Serphosphorylation of AnxA2 is a markedly decreasedability of the protein to aggregate lipid vesicles [32]and chromaffin granules [33]. Phosphorylation ofAnxA2 has also been reported to induce its transportto the nucleus, although it has been debated whetherSer or Tyr phosphorylation is involved [34–36]. AnxA2lacks a nuclear localisation signal. Therefore, nuclearimport of AnxA2may involve its binding to a protein thatcontains nuclear localisation signal [37] after phos-phorylation. Furthermore, it has also been proposedthat phosphorylation may expose the nuclear exportsignal in the N-terminus of AnxA2. This signal is notnormally exposed in the nucleus, most likely due to theinteraction of the N-terminal end with the C-terminus orother AnxA2 ligands [38].Both monomeric and heterotetrameric forms of
AnxA2 are able to bind mRNA [5]. However, only10–20% of the cellular pool of AnxA2 is engagedin mRNA binding [4], indicating regulation of theinteraction. Since Ser phosphorylation by PKC mayinduce the nuclear translocation of AnxA2 [35,37,39],we explored the possibility that Ser25 phosphorylationof AnxA2 could be a negative switch for mRNAbinding. As the phosphorylation state of a protein canbe complex and heterogeneous, phospho-mimickingmutants have been used to investigate the effect of
Please cite this article as: Grindheim Ann Kari, et al, Effect of SerinNuclear Localisation and Ligand Interactions of Annexin A2, J Mol
phosphorylation. Previously, humanAnxA2-S25E (andAnxA2-S25A) mutants have been used to investigatethe effect of Ser25 phosphorylation on nuclear localisa-tion of AnxA2 [35,39], its aggregation and binding toS100A10 [22,30] or its association with membranes[30]. The phospho-mimicking AnxA2-S25Eand AnxA2-S25D mutants, as well as the phospho-defective AnxA2-S25A mutant, have been employedhere to characterise the effect of these mutations onthe structure of AnxA2 and its ability to bind mRNA.Likewise, we investigated the effect of themutations onthe binding of AnxA2 to actin, which represents anotherligand that interacts with domain IV [40] and most likelyalso the N-terminus [41]. Since the Ca2+-binding sitesof AnxA2 are involved in its binding to lipidmembranes,the effect of the mutation of Ser25 on its binding tophospholipids and liposomes was also assessed.These investigations were complemented by localisa-tion studies of the AnxA2-S25E phospho-mimickingmutant and endogenous AnxA2 after activation of PKCby 12-O-tetradecanoylphorbol 13-acetate (TPA).
Results
Structure and solubility of theAnxA2-S25mutants
Wild-type (wt) AnxA2 and mutants in which Ser25had been mutated to Glu (E), Asp (D) or Ala (A) wereexpressed as His-tagged proteins in bacteria duringON incubation at 15 °C. To investigate whether themutations affected the solubility of the protein, weprepared Total (T), Supernatant (S) and Pellet (P)fractions from bacteria and subjected the expressedproteins to SDS-PAGE (Fig. 1). Comparison of theproteins showed that wt AnxA2 was mainly recoveredin the supernatant (Fig. 1, compare lanes 1 and 2),although a small fraction was also present in the pellet(Fig. 1, lane 3). All three AnxA2-S25mutants were alsomainly recovered in the supernatant (Fig. 1, comparelanes 4 and 5, 7 and 8 and 10 and 11), although aslightly larger fraction of the mutants appears to bepresent in the pellet as compared to the wt protein(Fig. 1, compare lanes 1–3, 4–6, 7–9 and 10–12).However, the mutants were sufficiently soluble to bepurified in high yields.The effect of the Ser25 mutations on the α-helical
conformation of AnxA2 was analysed by circulardichroism (CD) spectroscopy. Far-UV CD spectra(Fig. 2a) of all the mutants have negative maxima at~208 and ~222 nm and a positive maximum at195 nm, which is typical for proteins dominated byα-helices [42]. The spectra of the mutants are almostidentical with that of wt AnxA2. CD-monitored thermaldenaturation revealed that all three AnxA2-S25mutants, like wt AnxA2, have a two-state cooperativeunfolding, as judged from the S-shape of the thermaltransition curves (Fig. 2b). The apparent transition
e Phosphorylation and Ser25 Phospho-Mimicking Mutations onBiol (2014), http://dx.doi.org/10.1016/j.jmb.2014.04.019
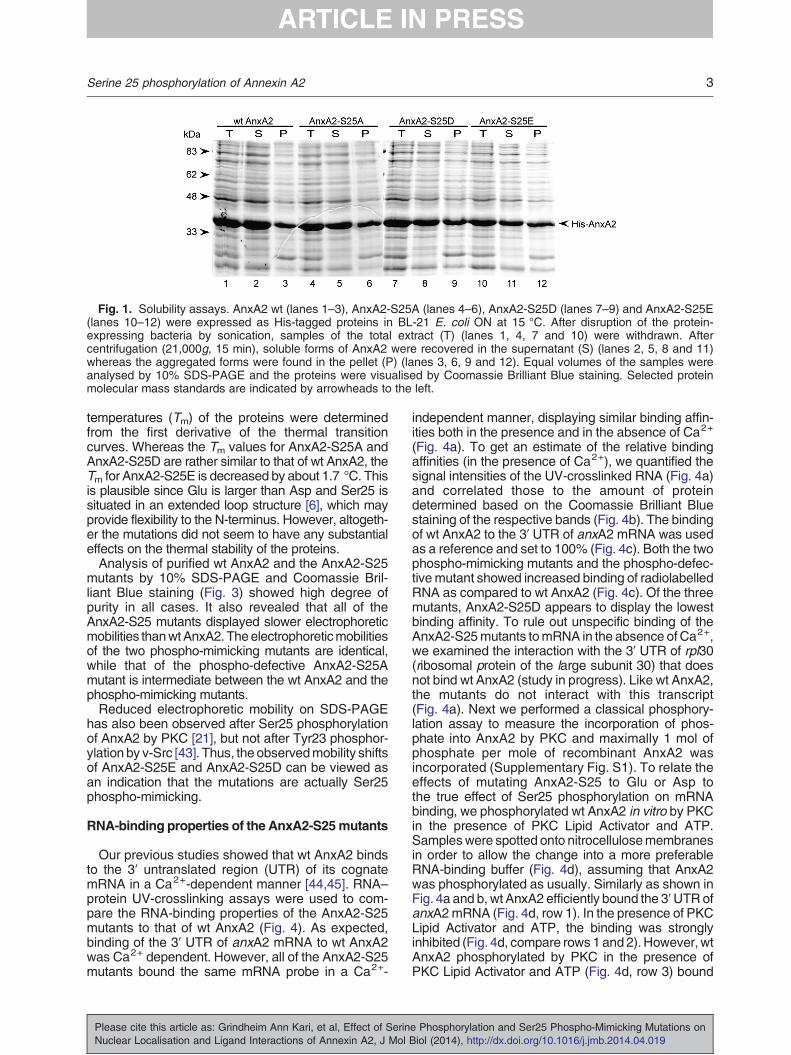
Fig. 1. Solubility assays. AnxA2 wt (lanes 1–3), AnxA2-S25A (lanes 4–6), AnxA2-S25D (lanes 7–9) and AnxA2-S25E(lanes 10–12) were expressed as His-tagged proteins in BL-21 E. coli ON at 15 °C. After disruption of the protein-expressing bacteria by sonication, samples of the total extract (T) (lanes 1, 4, 7 and 10) were withdrawn. Aftercentrifugation (21,000g, 15 min), soluble forms of AnxA2 were recovered in the supernatant (S) (lanes 2, 5, 8 and 11)whereas the aggregated forms were found in the pellet (P) (lanes 3, 6, 9 and 12). Equal volumes of the samples wereanalysed by 10% SDS-PAGE and the proteins were visualised by Coomassie Brilliant Blue staining. Selected proteinmolecular mass standards are indicated by arrowheads to the left.
3Serine 25 phosphorylation of Annexin A2
temperatures (Tm) of the proteins were determinedfrom the first derivative of the thermal transitioncurves. Whereas the Tm values for AnxA2-S25A andAnxA2-S25D are rather similar to that of wt AnxA2, theTm for AnxA2-S25E is decreased by about 1.7 °C. Thisis plausible since Glu is larger than Asp and Ser25 issituated in an extended loop structure [6], which mayprovide flexibility to the N-terminus. However, altogeth-er the mutations did not seem to have any substantialeffects on the thermal stability of the proteins.Analysis of purified wt AnxA2 and the AnxA2-S25
mutants by 10% SDS-PAGE and Coomassie Bril-liant Blue staining (Fig. 3) showed high degree ofpurity in all cases. It also revealed that all of theAnxA2-S25 mutants displayed slower electrophoreticmobilities thanwtAnxA2.Theelectrophoreticmobilitiesof the two phospho-mimicking mutants are identical,while that of the phospho-defective AnxA2-S25Amutant is intermediate between the wt AnxA2 and thephospho-mimicking mutants.Reduced electrophoretic mobility on SDS-PAGE
has also been observed after Ser25 phosphorylationof AnxA2 by PKC [21], but not after Tyr23 phosphor-ylation by v-Src [43]. Thus, the observedmobility shiftsof AnxA2-S25E and AnxA2-S25D can be viewed asan indication that the mutations are actually Ser25phospho-mimicking.
RNA-binding properties of the AnxA2-S25mutants
Our previous studies showed that wt AnxA2 bindsto the 3′ untranslated region (UTR) of its cognatemRNA in a Ca2+-dependent manner [44,45]. RNA–protein UV-crosslinking assays were used to com-pare the RNA-binding properties of the AnxA2-S25mutants to that of wt AnxA2 (Fig. 4). As expected,binding of the 3′ UTR of anxA2 mRNA to wt AnxA2was Ca2+ dependent. However, all of the AnxA2-S25mutants bound the same mRNA probe in a Ca2+-
Please cite this article as: Grindheim Ann Kari, et al, Effect of SerinNuclear Localisation and Ligand Interactions of Annexin A2, J Mol
independent manner, displaying similar binding affin-ities both in the presence and in the absence of Ca2+
(Fig. 4a). To get an estimate of the relative bindingaffinities (in the presence of Ca2+), we quantified thesignal intensities of the UV-crosslinked RNA (Fig. 4a)and correlated those to the amount of proteindetermined based on the Coomassie Brilliant Bluestaining of the respective bands (Fig. 4b). The bindingof wt AnxA2 to the 3′ UTR of anxA2 mRNA was usedas a reference and set to 100% (Fig. 4c). Both the twophospho-mimicking mutants and the phospho-defec-tivemutant showed increased binding of radiolabelledRNA as compared to wt AnxA2 (Fig. 4c). Of the threemutants, AnxA2-S25D appears to display the lowestbinding affinity. To rule out unspecific binding of theAnxA2-S25mutants tomRNA in the absence ofCa2+,we examined the interaction with the 3′ UTR of rpl30(ribosomal protein of the large subunit 30) that doesnot bind wt AnxA2 (study in progress). Like wt AnxA2,the mutants do not interact with this transcript(Fig. 4a). Next we performed a classical phosphory-lation assay to measure the incorporation of phos-phate into AnxA2 by PKC and maximally 1 mol ofphosphate per mole of recombinant AnxA2 wasincorporated (Supplementary Fig. S1). To relate theeffects of mutating AnxA2-S25 to Glu or Asp tothe true effect of Ser25 phosphorylation on mRNAbinding, we phosphorylated wt AnxA2 in vitro by PKCin the presence of PKC Lipid Activator and ATP.Sampleswere spotted onto nitrocellulosemembranesin order to allow the change into a more preferableRNA-binding buffer (Fig. 4d), assuming that AnxA2was phosphorylated as usually. Similarly as shown inFig. 4a and b, wt AnxA2 efficiently bound the 3′UTRofanxA2mRNA (Fig. 4d, row 1). In the presence of PKCLipid Activator and ATP, the binding was stronglyinhibited (Fig. 4d, compare rows1 and2). However,wtAnxA2 phosphorylated by PKC in the presence ofPKC Lipid Activator and ATP (Fig. 4d, row 3) bound
e Phosphorylation and Ser25 Phospho-Mimicking Mutations onBiol (2014), http://dx.doi.org/10.1016/j.jmb.2014.04.019

Fig. 2. CD of wt AnxA2 and the AnxA2-S25 mutants.(a) Far-UV spectra were obtained at 195–260 at a proteinconcentration of 20 μM in 20 mM Tris buffer (pH 8.0) at20 °C nm, using a light path of 1 mm. The shown spectrarepresent the average of four scans from which a bufferscan was subtracted. (b) CD-monitored thermal denatur-ation shows the disruption of the α-helicity at a proteinconcentration of 20 μM. The change in ellipticity at 222 nmwas measured at pH 8.0 at a heating rate of 40 °C/h from20 to 70 °C. The spectra shown are smoothed and theobservedoptical activities are expressedas themean residuemolar ellipticity [θ]MRW (deg cm2 dmol−1).
Fig. 3. Electrophoretic mobility of purified wt AnxA2 andthe AnxA2-S25 mutants. 2 μg of purified wt AnxA2 (lane1), AnxA2-S25E (lane 2), AnxA2-S25D (lane 3) andAnxA2-S25A (lane 4) were analysed by 10% SDS-PAGEand visualised by Coomassie Brilliant Blue staining.AnxA2 wt and the AnxA2-S25 mutants are indicated byarrowheads. Selected protein molecular mass standardsare indicated by arrowheads to the left.
4 Serine 25 phosphorylation of Annexin A2
radiolabelled RNA stronger than the correspondingnonphosphorylated wt AnxA2 under the same condi-tions (Fig. 4d, compare rows 2 and 3).
Actin binding of the AnxA2-S25 mutants
To investigate the binding of wt AnxA2 and thethree AnxA2-S25 mutants to actin, we incubatedWestern blots containing the renatured proteins withβ-γ-actin. Subsequently, actin binding was detectedusing specific antibodies. To verify the detection, theblots also contained β-γ-actin as a positive control.Coomassie Brilliant Blue staining confirmed that equalamounts of wt AnxA2 and each of the AnxA2-S25mutant proteins were loaded in the 10% SDS-PAGE(results not shown). The results showed thatwt AnxA2,as well as all three AnxA2-S25 mutants, boundβ-γ-actin in the monomeric form (Fig. 5). Moreover, allthree mutants also appeared to have an increasedaffinity for monomeric β-γ-actin as compared to wtAnxA2.
Please cite this article as: Grindheim Ann Kari, et al, Effect of SerinNuclear Localisation and Ligand Interactions of Annexin A2, J Mol
Lipid binding of the AnxA2-S25 mutants
The effect of the Ser25 mutations on lipid-bindingproperties of AnxA2 was assayed by using a protein–lipid overlay assay (Fig. 6). The wt protein displayedbinding to phosphatidylinositol (PI), phosphatidylino-sitol-3-phosphate (PI3P), phosphatidylinositol-4-pho-sphate (PI4P) and phosphatidylinositol-5-phosphate(PI5P), as well as weaker binding to phosphatidylser-ine (PS) and phosphatidylinositol-3,5-bisphosphate(PI35P2) in the presence of 1 mM Ca2+. Binding wasonly detected in the presence of Ca2+. Mutations ofSer25 exerted a dramatic effect on the lipid-bindingproperties of AnxA2 as the binding of the differentlipids to all the mutant proteins was almost completelyabolished under the same conditions. However,AnxA2-S25D displayed weak binding to PI5P, whileonly very weak binding of the two other mutants,AnxA2-S25E and AnxA2-S25A, was detected. Evenweaker binding of AnxA2-S25D and AnxA2-S25A toPI4P could also be detected.To approach in vivo conditions, we investigated the
effect of Ser25 mutations on the binding of AnxA2to PS/PC/cholesterol liposomes (1/1/1 molar ratio)[phosphatidylcholine (PC)] (Fig. 7a–d; with quantita-tions shown in Fig. 7e). Since AnxA2 is known to self-associate in the presence of Ca2+ [46], the tendencyof wt AnxA2 and the mutants to self-associate in thepresence of 1 mM Ca2+ in the absence of liposomesduring the 1-h incubation period was also examined.The phospho-mimicking AnxA2 mutants show anenhanced tendency to self-associate (Fig. 7c and d,lanes 1 and 2) as compared to wt AnxA2 andAnxA2-S25A (Fig. 7a and b, lanes 1 and 2).AnxA2-S25A and wt AnxA2 showed increased
e Phosphorylation and Ser25 Phospho-Mimicking Mutations onBiol (2014), http://dx.doi.org/10.1016/j.jmb.2014.04.019

Fig. 4. AnxA2–mRNA interaction. UV-crosslinking was carried out in the presence of 12 fmol of the 3′ UTR of anxA2 orrpl30 mRNA and 3 μM wt AnxA2 (lanes 1, 5 and 9), AnxA2-S25E (lanes 2, 6 and 12), AnxA2-S25D (lanes 3, 7 and 11) orAnxA2-S25A (lanes 4, 8, and 10) in the presence (lanes 1–4) or in the absence (lanes 5–12) of 70 μM calcium. Sampleswere analysed by 10% SDS-PAGE, where after RNA binding was determined by the detection of bound radioactivity usinga phosphoimager (a) and the proteins were visualised by Coomassie Brilliant Blue staining (b). Relative binding affinities ofwt AnxA2 and the AnxA2-S25 mutants to the 3′ UTR of anxA2 mRNA (c). Following subtraction of the background signals,the signal intensities of the UV-crosslinked samples were quantified and correlated to the amount of protein quantifiedbased on the Coomassie Brilliant Blue staining. The binding of wt AnxA2 to the 3′ UTR of anxA2 mRNA in the presence ofcalcium was defined as 100% (n = 4 independent experiments). The effect of PKC phosphorylation of AnxA2 on RNAbinding (d). A nitrocellulose membrane was spotted in triplicate with 1.5 μg of wt AnxA2 alone (row 1), in the presence of PKCLipid Activator and ATP (row 2), or phosphorylated by PKC in the presence of PKC Lipid Activator and ATP (row 3). Themembrane was incubated with 50,000 cpm/ml of the 3′UTR of anxA2mRNA in binding buffer containing 1 mM calcium. Themembrane was exposed to a radiosensitive screen, which was subsequently developed in the phosphoimager.
5Serine 25 phosphorylation of Annexin A2
association with the pelleted liposomes and werereleased by ethylene glycol bis(β-aminoethyl ether)N,N´-tetraacetic acid (EGTA) treatment (Fig. 7aand b, lanes 3–6), while the phospho-mimickingAnxA2 mutants displayed a much lower affinity forthe liposomes as compared to the respective self-associated AnxA2 proteins (Fig. 7c and d, comparelane 2 with lane 4).
Localisation of the AnxA2-S25E mutant andSer25 phosphorylated AnxA2
In order to be able to extrapolate the results obtainedwith the phospho-mimicking AnxA2 variant with the invivo Ser phosphorylation of AnxA2, the localisation ofAnxA2-S25E (which bound mRNA most efficiently of
Please cite this article as: Grindheim Ann Kari, et al, Effect of SerinNuclear Localisation and Ligand Interactions of Annexin A2, J Mol
the two phospho-mimicking AnxA2 mutants) and Serphosphorylated AnxA2was investigated (Fig. 8). Sincethese forms all bound mRNA with increased affinitycompared to wt AnxA2, the experiments were per-formed in the absence and presence of leptomycin B(LmB), which block CMR1-mediated export of RNAfrom the nucleus [47], since mRNA is not transportedinto the nucleus. Transfected wt AnxA2-GFP wasfound predominantly in the cytoplasm and some in thenucleus (Fig. 8a), while transfected AnxA2-S25E wasfound almost exclusively in the cytoplasm (Fig. 8c).Although the degree of transfection was poor, it isevident that LmB inhibited only the export of nuclear wtAnxA2-GFP as expected (compare Fig. 8b and d).The level of endogenous AnxA2 increased in the
nucleus after LmB treatment in serum-starved cells
e Phosphorylation and Ser25 Phospho-Mimicking Mutations onBiol (2014), http://dx.doi.org/10.1016/j.jmb.2014.04.019

Fig. 5. Binding of wt AnxA2 and AnxA2-S25 mutantsto β-γ-actin. 13 μM of wt AnxA2 (lane 2), AnxA2-S25E(lane 3), AnxA2-S25D (lane 4) and AnxA2-S25A (lane 5)were separated by 10% SDS-PAGE and transferred to anitrocellulose membrane that was incubated with mono-meric β-γ-actin after renaturation of the proteins on theblot. Actin binding was detected by monoclonal actinantibodies. Lane 1 contains 3 μM β-γ-actin. The positionsof actin, AnxA2-S25 mutants and wt AnxA2 are indicatedby arrowheads.
6 Serine 25 phosphorylation of Annexin A2
(Fig. 8e and f), although not to the same degree as innon-starved cells (results not shown). TPA treatmentof PC12 cells to activate PKC did not increase theamount of AnxA2 in the nucleus (compare Fig. 8eand g), while treatment with the PKC inhibitor did(compareFig. 8eand i). Furthermore, the localisationofendogenous AnxA2 in cells where PKC was activatedby TPA (Fig. 8g and h) or inhibited by a myristoylatedPKC peptide inhibitor (Fig. 8i and j) [48] seems to beunaffected by LmB treatment (Fig. 8h and j). ThemajorPKC phosphorylation site of AnxA2 is Ser25 as theSer11 site has only been shown to be phosphorylatedin vitro [49], although this does not rule out Ser11phosphorylation in vivo.
Fig. 6. Binding of wt AnxA2 and AnxA2-S25 mutants tolipids. PIP Strips™ were incubated with 5 μg/ml of wtAnxA2, AnxA2-S25E, AnxA2-S25D or AnxA2-S25A in theabsence or presence of 1 mM Ca2+. Binding of protein tothe lipid spots was detected by subsequent incubationwithmonoclonalAnxA2primaryantibodies. The lipids spottedon the PIP Strips™ are lysophosphatidic acid (LPA), lysopho-sphatidylcholine (LPC), PI, PI3P, PI4P, PI5P, phosphatidyl-ethanolamine (PE), PC, sphingosine-1-phosphate (S1P),phosphatidylinositol-3,4-bisphosphate (PI34P2), PI35P2,PI45P2, phosphatidylinositol-3,4,5-trisphosphate (PI345P3),PA, PS and blank as indicated.
Discussion
Expression, purification and characterisation ofAnxA2- S25 mutants
To investigate the effect of Ser25 phosphoryla-tion on the ability of AnxA2 to bind mRNA, we haveproduced two phospho-mimicking mutants,AnxA2-S25E and AnxA2-S25D, and a phospho-defective mutant, AnxA2-S25A, of the protein. Toinvestigate the structural effects of the mutations,we performed initial characterisations of the mutantproteins. Although the sizes (including length of sidegroup) and charges of Asp (D) andGlu (E) (−1) are notidentical with a phosphorylated Ser (S) (−2 at pH 7.5),these phospho-mimicking mutants may contributeto the understanding of how phosphorylation of aspecific Ser may result in conformational changesaffecting the ligand-binding properties of the protein. Inaddition, they provide important tools, since AnxA2 isphosphorylated on Ser by conventional PKCs thatrequire lipids for their activity [24] and the presence oflipids may obscure other AnxA2–ligand interactions.Although all three Ser25 mutations were expected todestabilise the overall structure ofAnxA2, as predicted
Please cite this article as: Grindheim Ann Kari, et al, Effect of SerinNuclear Localisation and Ligand Interactions of Annexin A2, J Mol
by the Cupsat software [50], the mutant proteinswere soluble and could be successfully purified,although they exhibited a slightly increased tenden-cy to aggregate (Fig. 1). Furthermore, the thermalstabilities of the mutants were largely unaffected asevidenced by the CD-monitored thermal denaturationcurves (Fig. 2). However, SDS-PAGE of the purifiedproteins showed that all three AnxA2-S25 mutantsdisplayed reduced electrophoretic mobilities as com-pared to wt AnxA2 (Fig. 3). The mobility of AnxA2-S25A was slightly increased as compared to the two
e Phosphorylation and Ser25 Phospho-Mimicking Mutations onBiol (2014), http://dx.doi.org/10.1016/j.jmb.2014.04.019

Fig. 7. Binding of wt AnxA2 and AnxA2-S25 mutants to PS/PC/cholesterol liposomes (1/1/1 molar ratio; 4 mg/ml) in thepresence of 1 mM Ca2+. 10 μg of wt AnxA2 (a), AnxA2-S25A (b), AnxA2-S25D (c) or AnxA2-S25E (d) were incubatedalone (lanes 1 and 2) or together with 50 μl of liposomes (lanes 3–6) in the absence (lanes 3–4) or presence of 10 mMEGTA (lanes 5–6) for 1 h at 4 °C. The samples were centrifuged for 30 min at 13,000g to obtain supernatant (S) and pellet(P) fractions. Equal volumes of the samples (S and P) were analysed by 10% SDS-PAGE and the proteins were visualisedby Coomassie Brilliant Blue staining. The distribution of protein in P (dark grey) and S (light grey) fractions were quantitated(e) and standard errors are given (n = 3 separate experiments).
7Serine 25 phosphorylation of Annexin A2
phospho-mimicking mutants (AnxA2-S25E andAnxA2-S25D) but still slower than wt AnxA2. Themobility shifts typical for the purified proteins were notevident when the proteins present in the total,supernatant and pellet fractions were analysed(Fig. 1), most likely because large amounts of proteinsloaded onto the gel affects the electrophoreticmobilities of the individual proteins. The alteredmobilityof the AnxA2-S25E mutant relative to wt AnxA2 hasalso previously been observed, while AnxA2-S25Awas reported to have the same electrophoretic mobilityas the wt protein [30]. This discrepancy could be due tothe different conditions for SDS-PAGE (polyacrylamideconcentrations, separation/running conditions andgel length). In this connection, we noticed that EGTAtreatment of AnxA2 during binding to liposomes oftenresulted in a slightly increased migration of the proteinas detected by SDS-PAGE (Fig. 7a–d, lane 5).
Please cite this article as: Grindheim Ann Kari, et al, Effect of SerinNuclear Localisation and Ligand Interactions of Annexin A2, J Mol
Secondary structure predictions of wt AnxA2 andthe AnxA2-S25 mutants were performed using fivedifferent methods: Porter [51], Sable [52], Psipred,jnet and sspro [53], allowing comparison of thepredicted structures. All the algorithms predicted onlyminor changes in the protein secondary structure, andnone of them could be used to explain the observedfunctional effects of the AnxA2-S25 mutations(Figs. 4–8). NMRanalysiswas not conducted becauseof our previous experience showing that structuralchanges are difficult to detect by NMR in case ofmutant proteins with very similar CD spectra.A previous finding showing that AnxA2-
S11A-S25E and AnxA2-S11E-S25A mutants haveincreased sensitivity to cleavage by chymotrypsin ascompared to wt AnxA2 indicates that Ser25 muta-tions affect the structure of the N-terminal end andthat the cleavage site between Tyr29 and Thr30 in
e Phosphorylation and Ser25 Phospho-Mimicking Mutations onBiol (2014), http://dx.doi.org/10.1016/j.jmb.2014.04.019

8 Serine 25 phosphorylation of Annexin A2
the N-terminal end is more exposed in the mutants[30]. The solvent accessibility of Ser25 is predictedto be ~18% (Cupsat), indicating that the residuemight also be involved in non-covalent intramolec-ular interactions.The chymotrypsin data [30] together with the similar
functional properties and electrophoretic mobilities ofthe phospho-defective and phospho-mimicking mu-tants of AnxA2,which differ from those of thewt AnxA2,
Fig. 8. The Ser25 phospho-mimicking AnxA2-S25Eand Ser phosphorylated AnxA2 is not targeted to thenucleus. PC12 cells transfected with GFP-AnxA2 wt (a andb) or GFP-AnxA2-S25E (c and d) were treated with 20 nMLmB for 2 h (b and d) or left untreated (a and c).4′,6-diamidino-2-phenylindole is shown in blue in overlayimages in the insets. Non-transfected cells (e–j) wereuntreated (e) or treated with 200 nM LmB (f), 900 nM TPA(g), 900 nM TPA in combination with 200 nM LmB (h),100 μM PKC pseudosubstrate (i) or 100 μM PKC pseu-dosubstrate in combination with 200 nM LmB (j) for 30 minand were subsequently subjected to immunofluorescencestaining using polyclonal AnxA2 antibodies. Images wereobtained by confocal imaging. Scale bars are 10 μm.
Please cite this article as: Grindheim Ann Kari, et al, Effect ofNuclear Localisation and Ligand Interactions of Annexin A2, J
SerinMol
open the possibility that a conformational change/increased flexibility of the protein is induced as ageneral consequence of the modification of Ser25.Because the total α-helical content of the AnxA2-S25mutants seems to be unaffected, a structural changemight involve the N-terminal ends (tails) of the mutantproteins as indicated for AnxA1 [54]. In the crystalstructure of AnxA2, Pro20 is the first visible amino acid,indicating that the N-terminal end of AnxA2 is veryflexible. The crystal structure also shows that Ser25 islocated in an extended loop [6].
RNA binding
AnxA2 binds directly and selectively to a subgroupof mRNAs translated on cytoskeletal polysomes[13]. Several AnxA2-binding mRNAs have beenidentified, including c-myc [55,56], anxA2 [44,45],andN-methyl-D-aspartateR1 (NMDAR1) [57]mRNAsand infectious bronchitis viral RNA [58]. The bindinghas been mapped to be to the 3′ UTRs of anxA2 [44],c-myc [55] and NMDA R1 [57] mRNAs. Since it hasbeen proposed that Ser25 phosphorylation targetsAnxA2 to the nucleus [35,39], we anticipated that thephospho-mimicking Ser25 mutants would not bindRNA. Thus, to investigate this hypothesis, we ana-lysed the binding of wt AnxA2 and the AnxA2-S25mutants to the 3′ UTR of anxA2 mRNA by UV-cros-slinking assays. Surprisingly, all mutants exhibitedincreased binding of radiolabelledRNAas compared towt AnxA2 (Fig. 4c). As another approach, AnxA2 wasphosphorylated in vitro by PKC and used in RNA-bind-ing assays. The phosphorylation by PKC requires thepresence of lipids, which are difficult to subsequentlyremove from the reactionmixture. As the lipids interferewith the AnxA2–RNA interaction (Fig. 4d), the resultscan be difficult to interpret. However, Ser phosphory-lation of AnxA2 clearly increased its binding to RNA ascompared to wt AnxA2 in the presence of lipids. Themain in vitro phosphorylation site of the recombinantAnxA2 is Ser25, since a truncated form of AnxA2(Δ19AnxA2) containing only the Ser25 site is about80–90% phosphorylated by PKC, as compared to wt
e Phosphorylation and Ser25 Phospho-Mimicking Mutations onBiol (2014), http://dx.doi.org/10.1016/j.jmb.2014.04.019

9Serine 25 phosphorylation of Annexin A2
AnxA2, which contains both Ser11 and Ser25 sites(Supplementary Fig. S1).The interaction between wt AnxA2 and its cognate
mRNA has been previously shown to be Ca2+
dependent [45] (see also Fig. 4) and specific, since acontrol AnxA2 protein, in which residues involved inRNA binding have been mutated, does not interactwith RNA [45]. It has been hypothesised that Ca2+
binding induces a conformational change [59], whichmay unmask an RNA-binding site in domain IV of theprotein [45]. By contrast, both the phospho-mimickingmutants and the phospho-defective mutant bound the3′ UTR of anxA2 mRNA in a Ca2+-independentmanner. Introducing a negative charge at position 25of AnxA2, by exchanging the Ser to Asp or Glu or byintroducing a phosphate group, cannot directly en-hance RNA binding as the negative charge of thecarboxyl or phosphate group is expected to repel thephosphate backbone of the RNA. Thus, the Ca2+--independent binding of the phospho-mimicking AnxA2mutants is most likely due to a conformational changerendering the RNA-binding site in domain IV accessi-ble. This is supported by the observations that thephospho-defectivemutant and the phospho-mimickingmutants share the same RNA-binding properties(Fig. 4) and that a soluble domain IV of AnxA2, whichharbours the RNA-binding site in helices C and D [45],as well as two N-terminally truncated forms of AnxA2,Δ19AnxA2 and Δ33AnxA2, also bind RNA in theabsence of Ca2+ (Supplementary Fig. S2). Thus,Ca2+ and/or Ser phosphorylation appears to induceconformational changes that unmask the mRNA-bind-ing site of AnxA2.
Actin binding
To investigate the interaction of AnxA2 with anotherligand binding to its domain IV, we assayed the bindingof wt AnxA2 and the Ser25 mutants to actin. Bothmonomeric and heterotetrameric forms of AnxA2are associated with dynamic actin structures in thevicinity to cellular membranes and can bind and bundleactin filaments (F-actin) in a Ca2+-dependent manner[11,59,60]. Furthermore, monomeric AnxA2 is alsoable to bindG-actin andbinds to thebarbedendof actinfilaments [10]. To investigate actin binding, we used aprotein overlay assay where wt AnxA2 and theAnxA2-S25 mutants were blotted onto a nitrocellulosemembrane. The AnxA2 proteins were first denaturedand then renatured since denatured AnxA2 does notbind actin (results not shown). Following renaturation,wt AnxA2 and all three AnxA2-S25 mutants boundG-actin in the absence of Ca2+ (Fig. 5). Also, themutants typically displayed a higher affinity forβ-γ-actinas compared towt AnxA2. It has been reported that thebinding of AnxA2 to F-actin is a Ca2+-dependent event[12,61]. However, previous studies also showed that apool of AnxA2 binds tightly to cholesterol-rich mem-branes in a Ca2+-independent manner. This pool of
Please cite this article as: Grindheim Ann Kari, et al, Effect of SerinNuclear Localisation and Ligand Interactions of Annexin A2, J Mol
AnxA2 forms a complex that also contains severalactin-binding proteins and actin and could be isolatedin the presence of 2 mM EGTA, indicating Ca2+-independent interactions [62]. Tyr phosphorylation hasbeen shown to abolish the actin-binding and actin-bundling activities of the AnxA2 tetramer [63], as wellas induce actin re-modelling, altered cell morphol-ogy, cell detachment and cell scattering [64–66]. Bycontrast, mutation of Tyr23 to Asp (AnxA2-Y23D)leads to a large increase in actin binding comparedto both wt AnxA2 and AnxA2-Y23A [67]. Here wereport, to our knowledge, a novel finding that mutationsof Ser25 can promote the binding of AnxA2 to G-actin,in a Ca2+-independent manner.
Lipid binding
Lipids interact with the convex side of the AnxA2core structure in a Ca2+-dependent and Ca2+-independent manner [2,68]. A protein–lipid overlayassay demonstrated Ca2+-dependent binding of wtAnxA2 to PI, PI3P, PI4P and PI5P. Weaker but stillCa2+-dependent binding to PS and PI35P2 wasalso evident (Fig. 6). A similar lipid strip assay waspreviously used to determine the lipid-binding proper-ties of wt AnxA2 [69] with results that are not fullyconsistent with that of ours. In contrast to Hayes et al.,we did not detect binding of wt AnxA2 to phosphati-dylinositol-3,4-bisphosphate, phosphatidic acid (PA),phosphatidylinositol-4,5-bisphosphate (PI45P2) orphosphatidylinositol-3,4,5-trisphosphate (Fig. 6). Pro-longed exposure resulted in detection of the binding ofwt AnxA2 to PA as well (results not shown).Furthermore, we detected binding to PI3P and PS,whereas Hayes et al. did not. In line with our data, aconsensus sequence for PS recognition was found indomain II of AnxA2 [70]. These discrepancies couldbe caused by differences in the procedures, related tothe protein concentrations, buffer conditions andincubation times that were used, for example. Itshould also be noted that we used a Ca2+ concen-tration (1 mM) two times higher as compared toHayeset al. [69]. Furthermore, the heterotetrameric form ofAnxA2 in complex with S100A10 has at least a 10-foldhigher affinity for PI45P2 than the monomeric form, asdetermined by the surface plasmon resonancetechnique [71].Ser25mutants of AnxA2 appeared to still retain the
Ca2+ dependency for lipid binding (results not shown),whereas their binding to lipids in the presence of 1 mMCa2+ was dramatically affected. AnxA2-S25D andAnxA2-S25A showed weak binding to only PI5P andPI4P in the presence of Ca2+, and AnxA2-S25E alsoshowedweakbinding toPI5P. This is in linewith resultsshowing that Ser phosphorylation of AnxA2 by PKCinhibits its ability to aggregate phospholipid vesicles[32].Since the analysis of the lipid-binding properties of
the three AnxA2-S25 mutants is based on
e Phosphorylation and Ser25 Phospho-Mimicking Mutations onBiol (2014), http://dx.doi.org/10.1016/j.jmb.2014.04.019

10 Serine 25 phosphorylation of Annexin A2
immunodetection, it should be taken into consider-ation that the monoclonal AnxA2 antibody evidentlydoes not bind with equal affinity to wt AnxA2 and thedifferent mutant proteins. As compared to wt AnxA2,the antibody has a stronger affinity for AnxA2-S25Eand an even stronger affinity for AnxA2-S25D. Bycontrast, antibody affinity for AnxA2-S25A wasclearly weaker than for the wt protein (Supplemen-tary Fig. S3). Thus, it appears that AnxA2-S25D andAnxA2-S25A have approximately equal bindingaffinities for PI5P and PI4P.Ayala-Sanmartin et al. have also used the
AnxA2-S25E and AnxA2-S25A mutants to studyAnxA2–lipid interactions [30]. The binding of mono-meric or heterotetrameric AnxA2 to chromaffingranules and liposomes consisting of PC and PSwas not affected by the two mutations [30]. Relatedto this, phosphorylation of heterotetrameric AnxA2by PKC has been shown not to affect the binding tolipid vesicles [32], while the affinity but not capacityof binding to chromaffin granules was reduced [33].The results from the liposome-binding experiments(Fig. 7) corroborate the data obtained from the purelipid-binding assays (Fig. 6) in that the phospho-mi-micking mutations decrease the affinity of AnxA2 forthe PS/PC/cholesterol liposomes, while in contrastAnxA2-S25A behaves similar to the wt protein andbinds the liposomes in a Ca2+-dependent manner.Since the liposome association of AnxA2-Y23D waspreviously shown to significantly increase as com-pared to wt AnxA2 [67], the liposome association ofAnxA2-Y23E and AnxA2-Y23F was tested under thesame assay conditions as used for the AnxA2-S25mutants; AnxA2-Y23F behaved similar to wt AnxA2,while AnxA2-Y23E showed increased liposome bind-ing as compared to wt AnxA2 (results not shown).Concerning AnxA2, Ayala-Sanmartin et al. have
previously pointed to the possibility that phospho-mimicking amino acids do not fully mimic a phos-phorylation event [30]. Namely, they observed thatthe calcium sensitivity for chromaffin granule aggre-gation was different for phospho-mimicking mutants,as compared to earlier results obtained with phosphor-ylated AnxA2 [33]. However, our localisation studies ofthe phospho-mimicking AnxA2-S25Emutant and PKCphosphorylatedAnxA2 showed that they localise to thesame cellular sites, excluding the nucleus. Further-more, inhibition of PKC increased the signal of AnxA2in the nucleus. Combined with the findings that Ser25and Tyr23 phosphorylations are mutually exclusive[24], these results suggest that Tyr23 phosphorylation,perhaps in combination with other post-translationalmodifications, may target AnxA2 to nucleus in line withprevious published results [36].Ser phosphorylated AnxA2 and the Ser25 phos-
pho-mimicking mutants all show an increased affinityfor mRNA and share the same cellular localisation,indicating that the phospho-mimicking mutants can beused for more detailed molecular studies. It appears
Please cite this article as: Grindheim Ann Kari, et al, Effect of SerinNuclear Localisation and Ligand Interactions of Annexin A2, J Mol
that any perturbation at Ser25 changes protein functionof AnxA2 by leading to a conformational change thatexposes the actin and/or mRNA-binding sites. Thismay also explain why AnxA2 is able to interact withinfectious bronchitis virus RNA in the absence of Ca2+
[58]. Thus, the phosphorylation of AnxA2 Ser25 couldhave different outcomes: (a) the negative charge of thephosphate group could affect electrostatic attraction orrepulsion to nearby amino acids in the tertiary structureof AnxA2. Thereby, it could mediate changes in AnxA2conformation, in particular, affecting the flexibility of theN-terminus leading to modulation of the accessibility ofcertain ligand-binding sites. (b) Alternatively (or inaddition), phosphorylation of Ser25 couldmodulate thebinding affinities of ligands (such as liposomes).In conclusion, mutation or phosphorylation of AnxA2
Ser25 has minor effects on its secondary structure butinducessmall tertiary conformational changesaffectingsomeof itsmany functions. Thus,modulation of AnxA2Ser25 increases RNA and G-actin binding, decreaseslipid binding and reduces nuclear localisation.
Materials and Methods
Construction of the AnxA2-S25E, AnxA2-S25D andAnxA2-S25A mutants
The bovine wt anxA2 cDNA [72] was used as a templatefor the construction of the three AnxA2 mutants(AnxA2-S25E, AnxA2-S25D and AnxA2-S25A) by usingthe PCR/BbsI seamless cloning method [73] and specificprimers obtained from Sigma (Supplementary Table S1).Each of the PCR fragments was ligated into the TOPOvector (Invitrogen) using TA cloning for amplification of thefragments in bacteria, followed by their digestion with theBbsI restriction enzymeand ligation of the fragments into theNcoI andAcc65I sites of the pETMvector (kindly providedbyGunter Stier, University of Heidelberg, Germany). Thevector is derived from the pET24d expression vector(Novagen) and contains a 6His-tag. Human GFP-AnxA2wt (kindly provided by Prof. Volker Gerke, University ofMünster, Germany) was used as a template for theconstruction of human GFP-AnxA2-S25E using the Quik-Change Site-Directed Mutagenesis Kit (Stratagene) andspecific primers obtained fromSigma (Supplementary TableS1). The final cDNA constructs were verified by DNAsequencing.
Protein expression and purification
The expression and purification of the His-AnxA2 wt andmutant proteins have been described in detail elsewhere[44,74], with the exception that the proteins in this studywere not expressed as His-MBP (maltose-binding protein)fusion proteins but only with a 6His-tag. Protein concen-trations were determined by the Bradford method [75] or byUV spectroscopy at 280 nm using an extinction coefficient of33,810 M−1 cm−1 for AnxA2 wt, AnxA2-S25E, AnxA2-S25Dand AnxA2-S25A, calculated using the ExPASy ProtPram
e Phosphorylation and Ser25 Phospho-Mimicking Mutations onBiol (2014), http://dx.doi.org/10.1016/j.jmb.2014.04.019

11Serine 25 phosphorylation of Annexin A2
tool, assuming that all Cys residues are reduced and basedon using a protein mass of 39 kDa.
Solubility assays
After expression of the wt AnxA2 and mutant proteins inBL-21Escherichia coli, 1 ml of the respective bacterial cultureswas collected and centrifuged for 10 min at 2000g at 4 °C. Thesupernatant was discarded and the pellet resuspended in500 μl Breakage buffer [50 mMNa2PO4 × 2H2O, 0.5 MNaCl,10 mM imidazole, 5% glycerol, 0.5 μ/ml DNase I, 0.25 μg/mlRNase A, 1× protease inhibitor cocktail (Roche) and 1 mMDTT]. Cells were disrupted by ~20 sonication cycles, using anamplitude of 60 and 1–2 s pulses for 1 min at 4 °C. Wewithdrew 100 μl of the homogenate (Total extract, T). Another100 μl aliquot of each extract was transferred to a new tubeand centrifuged for 15 min at 21,000gat 4 °C. The supernatantwas withdrawn (Supernatant, S). The pellet (Pellet, P) wasresuspended in 100 μl of Breakage buffer. Finally, 33 μl of 4×loading buffer (240 mM Tris base, pH 6.8, 8% SDS, 5%2-mercaptoethanol, 40% glycerol and 0.02% bromophenolblue) was added to each sample (T, S, P), where after equalvolumes (30 μl) of the samples were analysed by SDS-PAGE.
CD measurements
Far-UV CD spectra (195–260 nm; light path, 1 mm)were recorded at 20 °C in a Jasco J-810 spectropolarim-eter equipped with a Peltier temperature control unit. Priorto CD measurements, the protein solutions were centri-fuged for 15 min at 200,000gav at 4 °C to eliminate largeaggregates. The proteins were diluted in 20 mM Tris buffer(pH 8) to a final concentration of 20 μM. The spectraobtained represent the average of four scans. Buffer scanswere subtracted from protein spectra. Thermal transitioncurves (20–70 °C) were determined by monitoring thedecrease in ellipticity at 222 nm at a heating rate of40 °C/h.
In vitro transcription
The 3′ UTR of full-length bovine anxA2 cDNA [72] inan EcoRI linearised pGEM3Zf+ vector was used asa template for the SP6-directed in vitro transcription of[α-32P]UTP-labelled (3000 Ci/mmol, 10 μCi/μl; PerkinElmer) 3′ UTR of anxA2 mRNA by previously describedstandard procedures [44], obtaining a specific activity of 7to 8 × 107 cpm/μg RNA. Likewise, the rpl30 3′ UTR wastranscribed from the T7 promoter. Uniformly labelled RNAprobes were analysed for integrity and purity by agarosegel electrophoresis.
UV-crosslinking RNA-binding assays
Purified radiolabelled RNA that had been heated at72 °C for 3 min (and subsequently slowly cooled to RT)was incubated with wt AnxA2 or mutants in RNA-bindingbuffer (50 mM KCl, 1 mM DTT, 2 mM MgSO4 and10 mM triethanolamine, pH 7.4) with or without 70 μMCa2+ as previously described [44], with the exceptionthat nucleotide–protein complexes were visualised using a
Please cite this article as: Grindheim Ann Kari, et al, Effect of SerinNuclear Localisation and Ligand Interactions of Annexin A2, J Mol
Fujifilm Image Reader BAS-5000 or a BioRad MolecularImager® FX.
Spot blot RNA-binding assays
Samples (1.5 μg in triplicate) of nonphosphorylated [withor without PKC Lipid Activator (Upstate-Millipore) and ATP]or PKC phosphorylated wt AnxA2 were spotted onto a0.2-μm nitrocellulose membrane and were air-dried. Subse-quently, the membrane was incubated in RNA-binding buffercontaining 1 mM Ca2+, 1 mg/ml yeast tRNA and Denhardt'ssolution [0.02% each of Ficoll, polyvinylpyrrolidone andbovine serum albumin (BSA)] ON at 4 °C and then for 1 hat RT to block unspecific binding. The RNA was heated andcooled as described above. Themembrane was incubated inRNA-binding buffer containing 1 μl/ml RNasin (40 U/μl) and50,000 cpm/ml of the 3′ UTR of anxA2 mRNA for 20 min atRT. Subsequently, the membrane was quickly rinsed threetimes with RNA-binding buffer without tRNA to removeexcess RNA and thereafter washed 4 × 15 min with thesame buffer. wt AnxA2 (0.5 μg/μl) was phosphorylated byPKC (10 ng/μl; Upstate-Millipore) for 2 h at 30 °C in thepresence of 45 mM ATP and 0.5× PKC Lipid Activator(0.5 mg/ml PS and 0.05 mg/ml diacylglycerol in 20 mMMops, pH 7.2, 25 mM β-glycerolphosphate, 1 mM sodiumorthovanadate, 1 mM DTT and 1 mM CaCl2).
Actin-binding assays
Samples (5 μg) of wt AnxA2 and mutants weresubjected to 10% SDS-PAGE and the proteins weretransferred to nitrocellulose membranes (Schleicher &Schuell). Proteins on the nitrocellulose membranes weredenatured for 2 h with blotting denaturation buffer (7 Mguanidine hydrochloride, 2 mM ethylenediaminetetraaceticacid, 50 mMDTTand 50 mMTris, pH 8.3). Themembraneswere washed for 2 × 5 min with TBS (Tris, pH 7.4, 140 mMNaCl) and the proteins were subsequently renatured byincubation with renaturation buffer (140 mM NaCl, 10 mMTris, pH 7.4, 2 mM ethylenediaminetetraacetic acid, 1%BSA, 0.1% Tween-20 and 2 mM DTT) ON at 4 °C [76]. Themembranes were washed for 2 × 5 min with TBS andblocked for 2 h with TBS/Tween buffer (20 mM Tris, pH 8.0,150 mMNaCl and0.05%Tween-20) containing 5%defatteddry milk (Marvel) and 1% glycine employing gentle agitationat RT. Subsequently, the membranes were incubatedwith 10 μg/ml β-γ-actin (Cytoskeleton) in the monomericform in TBS/Tween buffer containing 3% defatted drymilk and 1% glycine for ~24 h with gentle agitation. Actinbinding was assayed by primary monoclonal mouseanti-actin antibody (ICN Biomedicals) (ON), horseradish-peroxidase-conjugated goat anti-mouse secondary antibod-ies (BioRad) (2 h) employing ECL detection (Thermo).
Lipid-binding assays
Lipid-binding properties of wt AnxA2 or themutant proteinswereassayedusingPIPStrips™ (EchelonBiosciences). Theprotocol for PIP Strip™ was used, with some modifications:The membranes were blocked with TBS (0.05% Tween)containing 5% defatted dry milk and 1% glycine ON at 4 °C.The proteins were used at a concentration of 5 μg/ml and
e Phosphorylation and Ser25 Phospho-Mimicking Mutations onBiol (2014), http://dx.doi.org/10.1016/j.jmb.2014.04.019

12 Serine 25 phosphorylation of Annexin A2
incubated in TBS (0.05% Tween) containing 3% defatted drymilk and 1%glycine, with or without 1 mMCa2+ as indicated,for 1.5 h at RT. Protein binding was detected using purifiedmonoclonal AnxA2 primary antibodies (BD TransductionLaboratories) (ON), horseradish-peroxidase-conjugatedgoat anti-mouse secondary antibodies (BioRad) (2 h) andECL detection (Thermo).
Liposome-binding assays
Liposomes containing PS/PC/cholesterol in 1/1/1molar ratioat a final concentration of 4 mg/ml were prepared as previouslydescribed [77]. Since initial binding assays with liposomes andwt AnxA2 at 70 μMCa2+ did not result in binding, 1 mM Ca2+
was used in all assays. Liposome-binding assays using 10 μgof purified recombinant protein were also performed aspreviously described [77], with modifications that lipid–proteincomplexes were allowed to form during a 1-h incubationperiod at 4 °C, and centrifugation was at 13,000g for 30 min.
Cell culture
PC12 cells (sub-clone [78] kindly provided byProf. EyvindRødahl, Haukeland Hospital, Bergen, Norway) were grownin RPMI medium supplemented with 10% heat-inactivatedhorse serum (PAA), 5% heat-inactivated fetal bovine serum(Sigma), L-glutamine (2 mM), penicillin (25 U/ml) andstreptomycin (25 μg/ml). Cells were grown on poly-L-lysinecoated coverslips in completemedium or starved inmediumwith 1%horse serumand 0.5% fetal bovine serum for 6 h, asindicated, in a humidified 5% CO2 atmosphere at 37 °C.Cells were treated with 900 nM TPA (Sigma) or 10 μMMyristoylated PKC Peptide Inhibitor (Promega) for 30 minand/or 200 nM (30 min) or 20 nM (2 h) LmB (Sigma).Transient transfection of cells was carried out usingMetafectene Pro (Biontex).
Immunofluorescence
Cells grown on coverslips were fixed for 20 min with 3%paraformaldehyde in 0.1 M phosphate buffer and subse-quently blocked and permeabilised with blocking buffer(5% goat serum, 0.2% saponin, 0.2% BSA and 0.02%Na-azide). Cells were stained with rabbit polyclonal AnxA2antibodies (1:125) (Abcam) ON and subsequently withDyLight488 conjugated goat anti-rabbit antibodies. Thecoverslips were inverted and mounted in Vectashieldmounting medium containing 4′,6-diamidino-2-phenylin-dole (Vector laboratories). Images were obtained using aLeica SP5 AOBS confocal laser scanning microscope,63×/1.4 NA HCX Plan-Apochromat oil-immersion objec-tives and ~1 Airy unit pinhole aperture. Fluorescenceimages were acquired with 405 Diode and Argon lasers.
Acknowledgements
We thank Prof. Torgeir Flatmark (University ofBergen, Norway) for valuable comments and discus-sions.We also thank Prof. Volker Gerke (University of
Please cite this article as: Grindheim Ann Kari, et al, Effect of SerinNuclear Localisation and Ligand Interactions of Annexin A2, J Mol
Münster, Germany) for the pEGFP-AnxA2 wt plas-mid and Prof. Gunter Stier (University of Heidelberg,Germany) for the pETM vector. This study wassupported by theUniversity of Bergen, HelseVest andResearch Council of Norway (AURORA programme).
Appendix A. Supplementary data
Supplementary data to this article can be foundonline at http://dx.doi.org/10.1016/j.jmb.2014.04.019.
Received 17 January 2014;Received in revised form 9 April 2014;
Accepted 19 April 2014Available online xxxx
Keywords:secondary structure;
G-actin;phospholipid;
mRNA;3′ untranslated region
Abbreviations used:AnxA2, annexin A2; BSA, bovine serum albumin; EGTA,ethylene glycol bis(β-aminoethyl ether) N,N´-tetraaceticacid; LmB, leptomycin B; PC, phosphatidylcholine; PA,phosphatidic acid; PI, phosphatidylinositol; PI3P, phos-phatidylinositol-3-phosphate; PI45P2, phosphatidylinosi-
tol-4,5-bisphosphate; PI4P, phosphatidylinositol-4-phosphate; PI5P, phosphatidylinositol-5-phosphate;
TPA, 12-O-tetradecanoylphorbol 13-acetate; UTR, un-translated region; wt, wild type.
References
[1] Gerke V, Moss SE. Annexins: from structure to function. PhysiolRev 2002;82:331–71.
[2] Gerke V, Creutz CE,Moss SE. Annexins: linkingCa2+ signallingto membrane dynamics. Nat Rev Mol Cell Biol 2005;6:449–61.
[3] Singh P. Role of annexin-II in GI cancers: interaction withgastrins/progastrins. Cancer Lett 2007;252:19–35.
[4] Bharadwaj A, Bydoun M, Holloway R, Waisman D. AnnexinA2 heterotetramer: structure and function. Int J Mol Sci2013;14:6259–305.
[5] Vedeler A, Hollås H, Grindheim AK, Raddum AM. Multipleroles of annexin A2 in post-transcriptional regulation of geneexpression. Curr Protein Pept Sci 2012;13:401–12.
[6] Rosengarth A, LueckeH. Annexin A2: does it inducemembraneaggregation by a newmultimeric state of the protein? Annexins2004;1:129–36.
[7] Rety S, Sopkova J, RenouardM, Osterloh D, Gerke V, TabariesS, et al. The crystal structure of a complex of p11 with theannexin II N-terminal peptide. Nat Struct Biol 1999;6:89–95.
[8] Cordier-Ochsenbein F, Guerois R, Russo-Marie F,Neumann JM, Sanson A. Exploring the folding pathwaysof annexin I, a multidomain protein II. Hierarchy in domainfolding propensities may govern the folding process. J Mol Biol1998;279:1177–85.
e Phosphorylation and Ser25 Phospho-Mimicking Mutations onBiol (2014), http://dx.doi.org/10.1016/j.jmb.2014.04.019

13Serine 25 phosphorylation of Annexin A2
[9] Rosengarth A, Gerke V, Luecke H. X-ray structure of full-length annexin 1 and implications for membrane aggregation.J Mol Biol 2001;306:489–98.
[10] Hayes MJ, Shao D, Bailly M, Moss SE. Regulation of actindynamics by annexin 2. EMBO J 2006;25:1816–26.
[11] Grieve AG, Moss SE, Hayes MJ. Annexin A2 at theinterface of actin and membrane dynamics: a focus on itsroles in endocytosis and cell polarization. Int J Cell Biol2012;2012:852430.
[12] Gerke V, Weber K. Identity of p36K phosphorylated uponRous sarcoma virus transformation with a protein purifiedfrom brush borders; calcium-dependent binding to non-erythroid spectrin and F-actin. EMBO J 1984;3:227–33.
[13] Vedeler A, Hollas H. Annexin II is associated with mRNAswhich may constitute a distinct subpopulation. Biochem J2000;348:565–72.
[14] Flood EC, Hajjar KA. The annexin A2 system and vascularhomeostasis. Vascul Pharmacol 2011;54:59–67.
[15] Thiel C, Osborn M, Gerke V. The tight association of thetyrosine kinase substrate annexin II with the submembranouscytoskeleton depends on intact p11- and Ca(2+)-bindingsites. J Cell Sci 1992;103:733–42.
[16] Zokas L, Glenney JR. The calpactin light chain is tightlylinked to the cytoskeletal form of calpactin I: studies usingmonoclonal antibodies to calpactin subunits. J Cell Biol1987;105:2111–21.
[17] Kumble KD, Vishwanatha JK. Immunoelectron microscopicanalysis of the intracellular distribution of primer recognitionproteins, annexin 2 and phosphoglycerate kinase, in normaland transformed cells. J Cell Sci 1991;99:751–8.
[18] Siever DA, Erickson HP. Extracellular annexin II. Int JBiochem Cell Biol 1997;29:1219–23.
[19] Heyraud S, Jaquinod M, Durmort C, Dambroise E, ConcordE, Schaal JP, et al. Contribution of annexin 2 to the architectureof mature endothelial adherens junctions. Mol Cell Biol2008;28:1657–68.
[20] Radke K, Gilmore T, Martin GS. Transformation by Roussarcoma virus: a cellular substrate for transformation-specificprotein phosphorylation contains phosphotyrosine. Cell1980;21:821–8.
[21] Gould KL, Woodgett JR, Isacke CM, Hunter T. The protein-tyrosine kinase substrate p36 is also a substrate for proteinkinase C in vitro and in vivo. Mol Cell Biol 1986;6:2738–44.
[22] Jost M, Gerke V. Mapping of a regulatory important site forprotein kinase C phosphorylation in the N-terminal domainof annexin II. Biochim Biophys Acta 1996;1313:283–9.
[23] Glenney JR, Tack BF. Amino-terminal sequence of p36 andassociated p10: identification of the site of tyrosine phos-phorylation and homology with S-100. Proc Natl Acad Sci US A 1985;82:7884–8.
[24] He KL, Sui G, Xiong H, Broekman MJ, Huang B, Marcus AJ,et al. Feedback regulation of endothelial cell surface plasmingeneration by PKC-dependent phosphorylation of annexinA2. J Biol Chem 2011;286:15428–39.
[25] Sarafian T, Pradel LA, Henry JP, Aunis D, Bader MF. Theparticipation of annexin II (calpactin I) in calcium-evoked exo-cytosis requires protein kinase C. J Cell Biol 1991;114:1135–47.
[26] Chasserot-Golaz S, Vitale N, Sagot I, Delouche B, Dirrig S,Pradel LA, et al. Annexin II in exocytosis: catecholaminesecretion requires the translocation of p36 to the subplasma-lemmal region in chromaffin cells. J Cell Biol 1996;133:1217–36.
[27] Delouche B, Pradel LA, Henry JP. Phosphorylation by proteinkinaseC of annexin 2 in chromaffin cells stimulated by nicotine.J Neurochem 1997;68:1720–7.
Please cite this article as: Grindheim Ann Kari, et al, Effect of SerinNuclear Localisation and Ligand Interactions of Annexin A2, J Mol
[28] Sagot I, Regnouf F, Henry JP, Pradel LA. Translocation ofcytosolic annexin 2 to a Triton-insoluble membrane sub-domain upon nicotine stimulation of chromaffin cultured cells.FEBS Lett 1997;410:229–34.
[29] Johnsson N, Nguyen Van P, Soling HD,Weber K. Functionallydistinct serine phosphorylation sites of p36, the cellularsubstrate of retroviral protein kinase; differential inhibition ofreassociation with p11. EMBO J 1986;5:3455–60.
[30] Ayala-Sanmartin J, Gouache P, Henry JP. N-terminal domainof annexin 2 regulates Ca(2+)-dependent membrane aggre-gation by the core domain: a site directed mutagenesis study.Biochemistry 2000;39:15190–8.
[31] Deora AB, Kreitzer G, Jacovina AT, Hajjar KA. An annexin2 phosphorylation switch mediates p11-dependent trans-location of annexin 2 to the cell surface. J Biol Chem2004;279:43411–8.
[32] Johnstone SA, Hubaishy I, Waisman DM. Phosphorylation ofannexin II tetramer by protein kinase C inhibits aggregation oflipid vesicles by the protein. J Biol Chem 1992;267:25976–81.
[33] Regnouf F, Sagot I, Delouche B, Devilliers G, Cartaud J, HenryJP, et al. "In vitro" phosphorylation of annexin 2 heterotetramerby protein kinase C. Comparative properties of the unpho-sphorylated and phosphorylated annexin 2 on the aggregationand fusion of chromaffin granule membranes. J Biol Chem1995;270:27143–50.
[34] Chiang Y, Davis RG, Vishwanatha JK. Altered expression ofannexin II in human B-cell lymphoma cell lines. BiochimBiophys Acta 1996;1313:295–301.
[35] Liu J, Rothermund CA, Ayala-Sanmartin J, Vishwanatha JK.Nuclear annexin II negatively regulates growth of LNCaP cellsand substitution of Ser 11 and 25 to Glu prevents nucleo-cytoplasmic shuttling of annexin II. BMC Biochem 2003;4:10.
[36] Eberhard DA, Karns LR, VandenBerg SR, Creutz CE.Control of the nuclear-cytoplasmic partitioning of annexin IIby a nuclear export signal and by p11 binding. J Cell Sci2001;114:3155–66.
[37] Yan G, Luo W, Lu Z, Luo X, Li L, Liu S, et al. Epstein-Barrvirus latent membrane protein 1 mediates phosphorylationand nuclear translocation of annexin A2 by activating PKCpathway. Cell Signalling 2007;19:341–8.
[38] Liu J, Vishwanatha JK. Regulation of nucleo-cytoplasmicshuttling of human annexin A2: a proposed mechanism. MolCell Biochem 2007;303:211–20.
[39] LuoW, Yan G, Li L, Wang Z, Liu H, Zhou S, et al. Epstein-Barrvirus latent membrane protein 1 mediates serine 25 phosphor-ylation and nuclear entry of annexin A2 via PI-PLC-PKCalpha/PKCbeta pathway. Mol Carcinog 2008;47:934–46.
[40] Filipenko NR, Waisman DM. The C terminus of annexin IImediates binding to F-actin. J Biol Chem 2001;276:5310–5.
[41] Ozorowski G, Ryan CM, Whitelegge JP, Luecke H. With-aferin A binds covalently to the N-terminal domain of annexinA2. Biol Chem 2012;393:1151–63.
[42] Kelly SM, Jess TJ, Price NC. How to study proteins bycircular dichroism. Biochim Biophys Acta 2005;1751:119–39.
[43] Isacke CM, Trowbridge IS, Hunter T. Modulation of p36phosphorylation in human cells: studies using anti-p36monoclonal antibodies. Mol Cell Biol 1986;6:2745–51.
[44] Hollas H, Aukrust I, Grimmer S, Strand E, Flatmark T,Vedeler A. Annexin A2 recognises a specific region in the 3′-UTR of its cognate messenger RNA. Biochim Biophys Acta2006;1763:1325–34.
[45] Aukrust I, Hollas H, Strand E, Evensen L, Trave G, FlatmarkT, et al. The mRNA-binding site of annexin A2 resides inhelices C-D of its domain IV. J Mol Biol 2007;368:1367–78.
e Phosphorylation and Ser25 Phospho-Mimicking Mutations onBiol (2014), http://dx.doi.org/10.1016/j.jmb.2014.04.019

14 Serine 25 phosphorylation of Annexin A2
[46] Liu L. Calcium-dependent self-association of annexin II: apossible implication in exocytosis. Cell Signalling1999;11:317–24.
[47] Wolff B, Sanglier JJ, Wang Y. Leptomycin B is an inhibitor ofnuclear export: inhibition of nucleo-cytoplasmic translocation ofthe human immunodeficiency virus type 1 (HIV-1) Rev proteinand Rev-dependent mRNA. Chem Biol 1997;4:139–47.
[48] Eichholtz T, de Bont DB, de Widt J, Liskamp RM, Ploegh HL.A myristoylated pseudosubstrate peptide, a novel proteinkinase C inhibitor. J Biol Chem 1993;268:1982–6.
[49] Hoque M, Rentero C, Cairns R, Tebar F, Enrich C, Grewal T.Annexins—scaffolds modulating PKC localization and sig-naling. Cell Signalling 2014;26:1213–25.
[50] Parthiban V, Gromiha MM, Abhinandan M, Schomburg D.Computational modeling of protein mutant stability: analysisand optimization of statistical potentials and structuralfeatures reveal insights into prediction model development.BMC Struct Biol 2007;7:54.
[51] Pollastri G, McLysaght A. Porter: a new, accurate server forprotein secondary structure prediction. Bioinformatics2005;21:1719–20.
[52] Adamczak R, Porollo A, Meller J. Combining prediction ofsecondary structure and solvent accessibility in proteins.Proteins 2005;59:467–75.
[53] Kelley LA, Sternberg MJE. Protein structure prediction onthe Web: a case study using the Phyre server. Nat Protoc2009;4:363–71.
[54] Porte F, de Santa Barbara P, Phalipou S, Liautard JP,Widada JS. Change in the N-terminal domain conformationof annexin I that correlates with liposome aggregation isimpaired by Ser-27 to Glu mutation that mimics phosphory-lation. Biochim Biophys Acta 1996;1293:177–84.
[55] Mickleburgh I, Burtle B, Hollas H, Campbell G, Chrzanowska-Lightowlers Z, Vedeler A, et al. Annexin A2 binds to thelocalization signal in the 3′ untranslated region of c-mycmRNA. FEBS J 2005;272:413–21.
[56] Filipenko NR, MacLeod TJ, Yoon CS, Waisman DM.Annexin A2 is a novel RNA-binding protein. J Biol Chem2004;279:8723–31.
[57] Anji A, Kumari M. A cis-acting region in the N-methyl-D-aspartate R1 3′-untranslated region interacts with the novelRNA-binding proteins beta subunit of alpha glucosidase IIand annexin A2—effect of chronic ethanol exposure in vivo.Eur J Neurosci 2011;34:1200–11.
[58] Kwak H, Park MW, Jeong S. Annexin A2 binds RNA andreduces the frameshifting efficiency of infectious bronchitisvirus. PLoS One 2011;6:e24067.
[59] Gerke V, Weber K. Calcium-dependent conformationalchanges in the 36-kDa subunit of intestinal protein I relatedto the cellular 36-kDa target of Rous sarcoma virus tyrosinekinase. J Biol Chem 1985;260:1688–95.
[60] Hayes MJ, Rescher U, Gerke V, Moss SE. Annexin–actininteractions. Traffic 2004;5:571–6.
[61] Ikebuchi NW, Waisman DM. Calcium-dependent regulationof actin filament bundling by lipocortin-85. J Biol Chem1990;265:3392–400.
[62] Harder T, Kellner R, Parton RG, Gruenberg J. Specificrelease of membrane-bound annexin II and cortical cytoskel-etal elements by sequestration of membrane cholesterol. MolBiol Cell 1997;8:533–45.
[63] Hubaishy I, Jones PG, Bjorge J, Bellagamba C, Fitzpatrick S,Fujita DJ, et al. Modulation of annexin II tetramer by tyrosinephosphorylation. Biochemistry 1995;34:14527–34.
Please cite this article as: Grindheim Ann Kari, et al, Effect of SerinNuclear Localisation and Ligand Interactions of Annexin A2, J Mol
[64] Hayes MJ, Moss SE. Annexin 2 has a dual role as regulatorand effector of v-Src in cell transformation. J Biol Chem2009;284:10202–10.
[65] Rescher U, Ludwig C, Konietzko V, Kharitonenkov A, GerkeV. Tyrosine phosphorylation of annexin A2 regulates Rho-mediated actin rearrangement and cell adhesion. J Cell Sci2008;121:2177–85.
[66] de Graauw M, Tijdens I, Smeets MB, Hensbergen PJ,Deelder AM, van de Water B. Annexin A2 phosphorylationmediates cell scattering and branching morphogenesis viacofilin activation. Mol Cell Biol 2008;28:1029–40.
[67] Morel E, Gruenberg J. Annexin A2 binding to endosomes andfunctions in endosomal transport are regulated by tyrosine 23phosphorylation. J Biol Chem 2009;284:1604–11.
[68] Illien F, Piao HR, Coue M, di Marco C, Ayala-Sanmartin J.Lipid organization regulates annexin A2 Ca(2+)-sensitivity formembrane bridging and its modulator effects on membranefluidity. Biochim Biophys Acta 2012;1818:2892–900.
[69] Hayes MJ, Merrifield CJ, Shao D, Ayala-Sanmartin J,Schorey CD, Levine TP, et al. Annexin 2 binding tophosphatidylinositol 4,5-bisphosphate on endocytic vesiclesis regulated by the stress response pathway. J Biol Chem2004;279:14157–64.
[70] Montaville P, Neumann JM, Russo-Marie F, Ochsenbein F,Sanson A. A new consensus sequence for phosphatidylserinerecognition by annexins. J Biol Chem 2002;277:24684–93.
[71] Gokhale NA, Abraham A, Digman MA, Gratton E, Cho W.Phosphoinositide specificity of and mechanism of lipiddomain formation by annexin A2-p11 heterotetramer. J BiolChem 2005;280:42831–40.
[72] Kristensen T, Saris CJ, Hunter T, Hicks LJ, Noonan DJ,Glenney JR, et al. Primary structure of bovine calpactin Iheavy chain (p36), a major cellular substrate for retroviralprotein-tyrosine kinases: homology with the human phos-phol ipase A2 inh ib i tor l ipocor t in . B iochemist ry1986;25:4497–503.
[73] Zanier K, Nomine Y, Charbonnier S, Ruhlmann C, Schultz P,Schweizer J, et al. Formation of well-defined solubleaggregates upon fusion to MBP is a generic property of E6proteins from various human papillomavirus species. ProteinExpression Purif 2007;51:59–70.
[74] Aukrust I, Evensen L, Hollas H, Berven F, Atkinson RA,Trave G, et al. Engineering, biophysical characterisation andbinding properties of a soluble mutant form of annexin A2domain IV that adopts a partially folded conformation. J MolBiol 2006;363:469–81.
[75] BradfordMM. A rapid and sensitive method for the quantitationof microgram quantities of protein utilizing the principle ofprotein-dye binding. Anal Biochem 1976;72:248–54.
[76] Chen MH, Sheng J, Hind G, Handa AK, Citovsky V.Interaction between the tobacco mosaic virus movementprotein and host cell pectin methylesterases is required forviral cell-to-cell movement. EMBO J 2000;19:913–20.
[77] Trave G, Cregut D, Lionne C, Quignard JF, Chiche L, SriWidada J, et al. Site-directed mutagenesis of a calciumbinding site modifies specifically the different biochemicalproperties of annexin I. Protein Eng 1994;7:689–96.
[78] Sannerud R, Marie M, Nizak C, Dale HA, Pernet-Gallay K,Perez F, et al. Rab1 defines a novel pathway connecting thepre-Golgi intermediate compartment with the cell periphery.Mol Biol Cell 2006;17:1514–26.
e Phosphorylation and Ser25 Phospho-Mimicking Mutations onBiol (2014), http://dx.doi.org/10.1016/j.jmb.2014.04.019
Copyright © 2022 FDOKUMEN




















