
Human Hepatocyte Morphology and Functions in a Multibore Fiber Bioreactor
Upload
univ-paris5Category
view
1download
0
Annexin Expressions Are Temporally and SpatiallyRegulated During Rat Hepatocyte DifferentiationBRUNO DELLA GASPERA,1 FRANCOISE BRAUT-BOUCHER,1 MORGANE BOMSEL,2
FRANCOISE CHATELET,3 CHRISTIANE GUGUEN-GUILLOUZO,4 JACQUELINE FONT,1
JACQUELINE WEINMAN,1 AND SERGE WEINMAN1*1UFR Biomedicale des Saints-Peres, Universite Rene Descartes, Paris, France2INSERM U332, ICGM, Paris, France3UFR Saint Antoine, Universite Pierre et Marie Curie, Paris, France4INSERM U522, Rennes, France
ABSTRACT Annexin (Anx) 1, 2, 5, and 6 ex-pressions were determined at the transcriptionaland translational levels in the rat hepatocytesfrom gestational day 15 to postnatal day 17. Dra-matic shifts were observed in Anx 1 and 2 levels,which peaked at day 1 and gestational day 20,respectively, and reached low levels thereafter.However, Anx 5 and 6 rates were more constant.Prenatal administration of dexamethasone (dex)resulted in a decrease of Anx 1 mRNA levels, anda strong increase in Anx 2 mRNA contents. Inadult hepatocytes cultured in the presence ofEGF or HGF, Anx 1 and 2 expressions resumed. Byimmunohistochemistry, Anx 1 was detected only inthe cytoplasm of hepatocytes of 1- to 3-day-old rats,Anx 2 and 6 both exhibited a redistribution fromthe cytoplasm toward the plasma membrane, andAnx 5 was present in the nucleus, cytoplasm, andplasma membrane. Thus, Anx 1, 2, 5, and 6 haveindividual modes of expression and localization inthe differentiating hepatocytes, where they mightplay unique roles at well defined phases of liverontogeny. © 2001 Wiley-Liss, Inc.
Key words: annexins; hepatocytes; development;rat
INTRODUCTION
Annexins (Anx) are proteins composed of four (eightin the case of Anx 6) highly conserved homologousrepeats of approximately 70 amino acids, which allharbour a characteristic Ca21- and phospholipid-bind-ing site within a 17 amino acid consensus sequencecalled the endonexin fold. The N-terminal tails arediverse in length and sequence, and their differencesare seemingly significant for the properties of each Anx(Lieman and Huber, 1997). Annexins are mainly local-ized in the cytosol, but they have been found in theplasma membrane, the vesicular system, the mitochon-dria, and the nucleus. Functions of Anxs are not fullyunderstood. Nevertheless, the common property ofAnxs, i.e., the Ca21-dependent binding to acidic phos-pholipids of the cytosolic leaflet of cellular membranes,strongly suggests that these proteins play a role in
membrane-related events, particularly in membraneorganization, exocytosis, and endocytosis. Anotherproperty of several Anxs is their Ca21 channel activity,which sheds light on some of their functions. As someAnxs are known to be phosphorylated by protein ki-nases involved in the control of cell growth and differ-entiation, it has been suggested that they might par-ticipate in these cellular events. In fact, Anxs seem tobe important regulatory proteins with pluripotent andpleiotropic roles, depending on cell physiological states(Creutz, 1992; Raynald and Pollard, 1994; Gerke andMoss, 1997; Donnelly and Moss, 1997).
During fetal and early postnatal life, the mammalliver is composed of two main cell types: hematopoieticcells and hepatocytes. In the rat, during these periods,the number of hematopoietic cells decreases, whereashepatocytes undergo mitosis and differentiation(Greengard et al., 1972). As Anxs are involved in cellproliferation and differentiation, a description of theexpression patterns of these proteins would be usefulin attempting to understand their roles in the develop-ing liver.
Anxs 1, 2, 5, and 6 have been found and localized inadult liver (Shadle et al., 1985; Mathew et al., 1986;Rainteau et al., 1988; Karasik et al., 1988; Kaetzel etal., 1989; Boustead et al., 1993; Weinman et al., 1994;Masaki et al., 1994; Ortega et al., 1998; Massey-Har-roche et al., 1998). Yet, little is known about the regu-lation of the expressions and cellular localizations ofthese proteins in the developing liver cells.
In this study, we examined the expressions of Anxsin the rat liver during fetal and early postnatal life. Asmechanisms other than changes in mRNA levels mightcontribute to altered protein expression, the rates ofsynthesis of Anxs were determined at the transcrip-
Grant sponsor: Universite Rene Descarte-Paris V; Grant number:plan quadriennal 1998-2001; Grant sponsor: the Institut National dela Sante et de la Recherche Medicale (INSERM); Grant sponsor:European Community; Grant number: BIO4-CT 960052.
*Correspondence to Pr. S Weinman, Laboratoire de Biologie duDeveloppement et de la Differentiation Musculaire (EA2507), UFRBiomedicale des Saints-Peres, 45 rue des Saints-Peres, 75270 Pariscedex 06 France. E-mail: [email protected]
Received 2 January 2001; Accepted 20 June 2001Published online 18 September 2001; DOI 10.1002/dvdy.1183
DEVELOPMENTAL DYNAMICS 222:206–217 (2001)
© 2001 WILEY-LISS, INC.

tional and translational levels. In an attempt to deter-mine the role of the glucocorticoids and growth factorsin the appearance of these proteins, we also investi-gated how dex, epidermal growth factor (EGF), andhepatic growth factor (HGF) could affect their expres-sions. Finally, we carried out the localization of Anxs inthe different cells of the developing liver. These studiesallowed us to obtain data about the temporal and spa-tial controls of Anx expressions during the proliferationand differentiation of hepatocytes.
RESULTSAnx Expressions Are Temporally Regulated inthe Developing Rat Liver
To correlate Anx gene expressions with the develop-ment of the rat liver, total RNA was isolated at differ-ent stages of fetal and postnatal periods: gestationalday (gd) 15, 16, 17, 18, 19, 20, and postnatal day (d) 1,3, 8, 17, respectively. The levels of total mRNA weredetermined by slot blot analysis. Those of the specifictranscripts were determined by Northern blot analysis.The relative amount of each specific mRNA present atthe various stages of the perinatal development wasdetermined as a ratio of the total mRNA and normal-ized to the peak value. Anx expressions were also de-termined at the protein level by Western blot analysis.
A constant expression of the total liver mRNA wasobserved throughout the fetal and early postnatal pe-riods of the rat liver development. In strong contrast,during the same periods, significant changes in Anxexpressions were evident, at the transcriptional as wellas translational levels.
As demonstrated in Figure 1, during the fetal andpostnatal periods, dramatic changes occurred in theliver expressions of Anxs 1, 2, 5, and 6, both at themRNA and protein levels. The 1.7-kb Anx 1 mRNAcould be detected at gd 15 and peaked at d 1. It was stillpresent at d 8, but became almost undetectable there-after. The 1.7-kb Anx 2 mRNA was expressed in asimilar time course as Anx 1 mRNA, although itpeaked earlier, at gd 20. Anxs 1 and 2 proteins weremostly detectable during the perinatal period andpeaked at d 1 and gd 21, respectively. A diphasic ex-pression was observed for the 1.7-kb Anx 5 mRNA,with an initial peak at gd 15 followed by a second peakat d 1. A similar pattern was obtained for the expres-sion of the 2.5-kb Anx 6 mRNA, with maxima at gd 16and gd 20. Anx 5 and 6 proteins were detectable at lowlevels in the fetal period and were steadily expressed athigh levels thereafter. To compare the expressions of Anxs 1, 2, 5, and 6
with those of a housekeeping protein, actin expressionwas also determined by Northern and Western blotanalyses (Fig. 2). The 2.1-kb b-actin mRNA, which washighly expressed at the beginning of fetal life, de-creased progressively to reach low levels by d 8. Thesame was true for b-actin protein, although to a lesserextent.
To determine the relationships between Anx expres-sion and cell proliferation in the developing rat liver,
Abbreviations
Anx annexindex dexamethasonegd gestational dayd postnatal dayEGF epidermal growth factorHGF hepatic growth factor
Fig. 1. Anx 1, 2, 5, and 6 expressions in rat liver at different stages ofdevelopment. A: Rat liver RNA analysis on Northern blot by sequentialhybridization to specific cDNA probes to Anx 1, 2, 5, or 6 mRNAs. Thehistograms give a quantitative representation of hybridization with spe-cific cDNA probes obtained by densitometric scanning of the autoradio-grams of the Northern blot of the liver RNA extracts. For each probe, afternormalization to the signal of the corresponding total mRNA, the relativeabundance is normalized to peak level. Results are expressed in arbitraryunits. The expressions of Anx 1 and 2 mRNAs are restricted to theperinatal period and they peak at d 1 and gd 21, respectively. Anxs 5 and6 are diphasically expressed, with an initial peak at gd 15-16 and asecond peak at birth. B: Western blot analysis of Anx 1, 2, 5, or 6 proteinsin extracts from rat liver. Anxs 1 and 2 are strongly expressed only duringthe perinatal period, whereas Anxs 5 and 6 exhibit a slightly increasingexpression throughout the period under consideration. Data shown (6SEM) are the means of three experiments. B, birth; for other abbrevia-tions, see list.
207ANNEXINS IN DIFFERENTIATING HEPATOCYTE

cyclin A mRNAs were assayed in liver RNA (Fig. 3).The two cyclin A mRNA species of 2.7 and 1.8 kb werehighly expressed from gd 15 to gd 20, with a peak at gd
16. On d 1, they were still present, although at muchlower levels. By d 4, they became almost undetectable.
To confirm these findings, cells were isolated fromlivers at different stages of development. Light micros-copy examination, as well as labeling by the antibodiesto vimentin, a hematopoietic cell marker, and to cyto-keratin 8, a hepatocyte marker, showed that the cellpopulation consisted of hematopoietic cells and hepa-tocytes. Anxs 1 and 2 were analyzed for immunofluo-rescence by flow cytometry. A quantitative analysis(Table 1) demonstrated that these proteins were ex-pressed in this cell population essentially during theperinatal period and peaked at d 1 for Anx 1 and gd 21for Anx 2, in accordance with the Northern and West-ern blot data presented in Figure 1. The cycle distribu-tion in the cell population was determined by flowcytometric analysis of DNA content (Fig. 4). During thefetal period, from gd 17 to gd 20, no less than 50% of thecells entered (S1G2/M) phases. By gd 21, this percent-age began to decline and reached the level of 23.5% atd 13. These results were in agreement with the cyclin AmRNA expression presented in Figure 3.
Prenatal Dex Administration Affects AnxmRNA Expressions
During the fetal period, glucocorticoids decrease thenumber and accelerate the involution of hematopoieticcells. In contrast, they increase the volume and accel-erate the differentiation of hepatocytes (Greengard etal., 1972). To examine whether prenatal exposure toglucocorticoids could modify calcium-binding proteinexpressions, livers were removed from the offsprings oftimed pregnant dams given 0.4 mg/kg of dexametha-sone (dex) daily from gd 15 to 21. Total liver RNA wasisolated at gd 17, 18, 19, 20, and d 1. The expressions ofAnx 1, 2, 5, and 6, and cyclin A mRNAs were deter-mined as above. Dex-treatment had significant effectson Anx 1 and 2 mRNA expressions (Fig. 5A). The levelsof Anx 1 mRNA were lowered significantly. In contrast,those of Anx 2 mRNA increased 10-fold at gd 18 andonly 2-fold at gd 20. Anx 5 and 6 mRNA levels (Fig. 5A)were similar in both control and treated animals. A
Fig. 2. Actin expression in rat liver at different stages of development.A: Rat liver RNA analysis on Northern blot by hybridization to a specificcDNA probe to b-actin mRNA. The total RNA was analyzed as describedin Figure 1A. The level of the 2.1-kb transcript decreases progressivelyfrom gd 15 to d 17. B, birth. B: Western blot analysis of b-actin protein inextracts from rat liver. b-actin is expressed at slowly decreasing levelsthroughout the period under consideration. For abbreviations, see list.
Fig. 3. Cyclin A expression in rat liver development at different stagesof development. B, birth. Rat liver RNA analysis on Northern blot byhybridization to a specific cDNA probe to cyclin A mRNAs. The total RNAwas analyzed as described in Figure 1A. The two cyclin mRNAs arestrongly expressed during the fetal period. A dramatic decrease occursafter birth.
TABLE 1. Intensity of Anx 1 and 2 Expressions in LiverCells at Different Stages of Developmenta
Gestational days Postnatal days17 19 21 1 4 13
Anx 1 10 16 230 440 19 14Anx 2 30 40 590 507 73 70aThe liver cell populations, isolated as described in Materialsand Methods section, were labeled either with monoclonalantibodies to vimentin, cytokeratin 8, annexin (Anx) 1, or Anx2, and analyzed by flow cytometry. Results are expressed inarbitrary units. As all the liver cell populations were positivefor vimentin and cytokeratin 8 (not shown), they proved to becomposed of both hematopoietic cells and hepatocytes. Onlythe cell populations isolated at the perinatal period werepositive for Anxs 1 and 2.
208 DELLA GASPERA ET AL.

dramatic shift was observed in the expression of cyclinA mRNAS (Fig. 5B). Their declines occurred earlier inthe prenatal dexamethasone-treated rats than in thecontrol animals, at gestational day 18 instead of day 1.
Anx Expressions in Adult Rat Hepatocytes areDependent on EGF and HGF
EGF and HGF are essential factors for optimal he-patocyte growth and differentiation (Schmidt et al.,1995). To determine the capacity of EGF and HGF,together with glucocorticoids, to induce the synthesis ofAnxs in hepatocytes, primary cultures of adult rathepatocytes, both unstimulated and stimulated by ei-ther EGF/- or HGF/hydrocortisone hemisuccinate,were used. In control cultures, after 48 hr, Anxs 5 and6 could be detected (Fig. 6A, lane 1); after 96 hr, a lowlevel of Anx 2 and a higher level of Anx 5 were ob-served, whereas the rate of Anx 6 remained unchanged(Fig. 6A, lane 2). After 24, 48, 72, or 96 hr, in agreementwith previous results (Loyer et al., 1996), [methyl-3H]thymidine incorporation was extremely low (Fig.6B), demonstrating that, during isolation from ratliver, hepatocytes undergo the G0/G1 transition and, inunstimulated culture, enter into and progress throughthe G1 phase up until a restriction point (R point) inmid-late G1 beyond, which they cannot complete thecell cycle.
In strong contrast, in stimulated cultures EGF/- andHGF/hydrocortisone hemisuccinate have similar,marked effects on the Anx rates (Fig. 6A, lanes 3–6).Anx 1 and 2 expressions resumed, and these proteinswere present at increasing levels, whereas Anx 5 and 6syntheses were significantly enhanced. From 24 to96 h, [methyl-3H]thymidine incorporation increased(Fig. 6B). DNA replication started at 48 hr and reacheda maximum at 96 hr, in agreement with a previousreport showing that most hepatocytes stimulated byEGF or HGF reach the G1/S boundary and completethe cell cycle (Loyer et al., 1996). It is noteworthy thatthe time of increased synthesis of Anxs and the time ofonset of DNA synthesis were identical.
Anx Expressions Are Spatially Regulated in theDeveloping Rat Liver
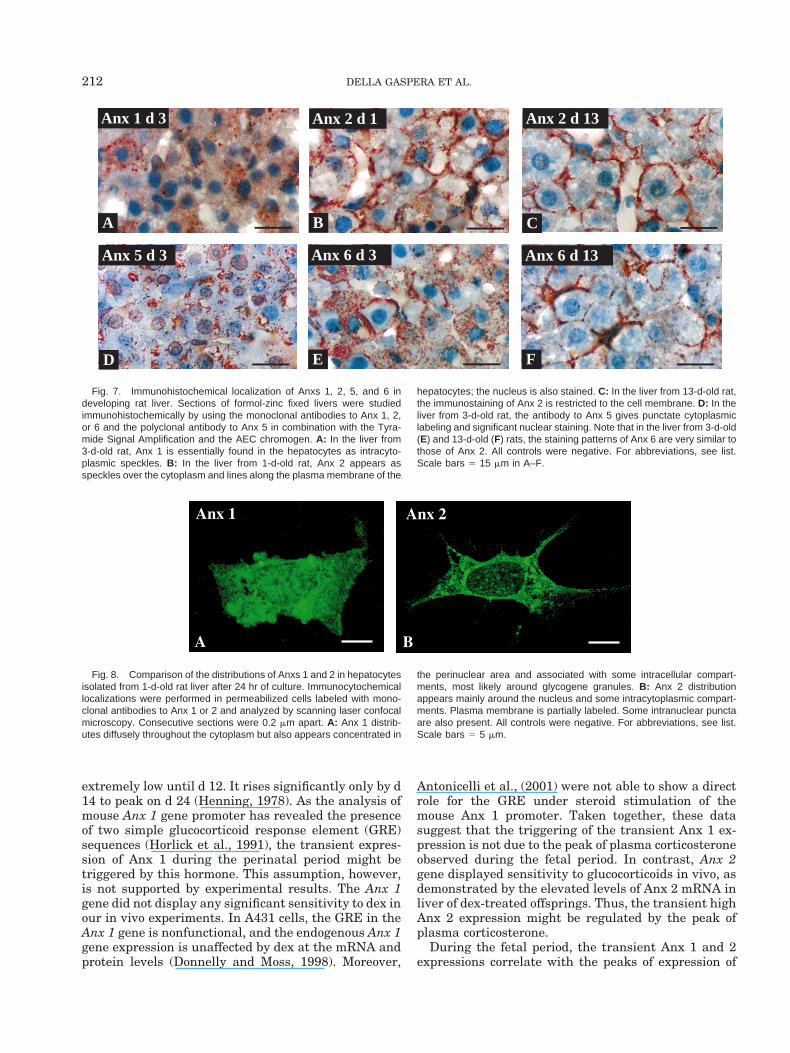
To determine the expressions of Anxs 1, 2, 5, or 6 inthe various cell types of the developing liver and tocompare their specific sites and time of distribution,sections of formol-fixed liver from fetal rats (gd 17) and1-, 3-, and 13-d-old rats were reacted with specific an-tibodies to these proteins and processed for immuno-histochemistry. For Anx 1, slightly positive immunore-activity was detected in the cytoplasm of thehepatocytes of the gd 17 fetal rat liver. In contrast, anintense speckled staining was observed throughout the
Fig. 4. Distribution of the liver cells in G1, S, and G2/M phases atdifferent stages of the perinatal development of the rat. A: Cell popula-tions composed of hematopoietic cells and hepatocytes were isolatedfrom rat liver, and their cell cycle distribution was determined by flowcytometric analysis of DNA content after staining with propidium iodide.
During the fetal period, numerous cells enter S and G2/M phases. Afterbirth, there is an increased percentage of cells blocked in G1, resulting instrongly reduced entry into S and G2/M phases. B: This change ishighlighted by the comparison of the cell cycle distribution of the gesta-tional day (gd) 19 and d 13 cell populations.
209ANNEXINS IN DIFFERENTIATING HEPATOCYTE

cytoplasm of the hepatocytes in the liver from 1-d-oldrat; the same staining was still present at d 3 (Fig. 7A),although at a lower level; no immunoreactivity wasdetected at d 13. In all cases, hematopoietic cells,megacaryoblasts, megacaryocytes, and endothelialcells were not immunoreactive. For Anx 2, immunore-activity was detected in the cytoplasm, and to a lesserextent, over the plasma membrane of the hepatocytesof the gd 17 fetal liver. In sections from 1-d-old ratliver, a strong positive immunoreactivity was evident
(Fig. 7B). The immunostaining appeared both as nu-merous speckles over the cytoplasm and the nucleusand as lines on basolateral and apical plasma mem-branes of the hepatocytes. In the 3-d-old rat liver thepattern was similar, but the labeling of the cytoplasmwas less intense. The immunoreactivity to Anx 2 wasstill present in the hepatocytes of the 13-d-old rat liver(Fig. 7C), although restricted to the basolateral andapical cell membranes. In all cases hematopoietic cells,megacaryoblasts, and megacaryocytes were not immu-
Fig. 5. Anx and cyclin A mRNA expressions in rat liver after prenataldex administration: 0.4 mg/kg per day, from gd 15 to gd 21; B, birth. TotalRNA analysis on Northern blot by sequential hybridization to specificcDNA probes to (A) Anx 1, 2, 5, or 6 mRNAs, and (B) cyclin A mRNAs,in RNA extracts from rat liver. Total RNA was analyzed as described in
Figure 1A. cont, control animals; dex tr, prenatal dex-treated animals.The levels of Anx 1 mRNA are lowered. Those of Anx 2 mRNA stronglyincreased, and those of Anx 5 and 6 mRNAs remained unaffected. CyclinA mRNA expressions decrease at gd 18 instead of d 1. For abbreviations,see list.
210 DELLA GASPERA ET AL.

noreactive, whereas endothelial cells were stained. ForAnx 4, no staining could be seen at any time. Theantibody to Anx 5, in contrast, gave faint punctatecytoplasmic labeling and strong nuclear staining inhepatocytes from fetal to 13-day-old rat liver. It wasparticularly intense in the nucleus by d 3 (Fig. 7D). Thenucleolus was not reactive. In dividing hepatocytes,strong immunoreactivity was also detected on the met-aphasic chromosomes (not shown). In erythropoieticcells, only the thin film of cytoplasm was positive. Im-munoreaction was absent when the antibody to Anx 5was preabsorbed with the corresponding immunizingantigen before analysis. For Anx 6, the cytoplasmic andnuclear distribution patterns were similar to those ofAnx 2 (Fig. 7E,F). In particular, a significant redistri-bution from the cytoplasm toward the inner face of theplasma membrane was observed between d 3 and d 13.Nevertheless, some differences must be noted: the bil-iary epithelial cells in the portal tract were strongly
positive, and the labeling of the membrane was mainlybasolateral. In no cases were hematopoietic cells,megacaryoblasts, or megacaryocytes immunoreactive.However, the endothelial cells were stained.
To confirm the immunohistochemical findings con-cerning Anx 1 and 2 localizations in hepatocytes, cellsisolated from 1-day-old rat liver and cultured for 24 hrwere analyzed by fluorescence confocal microscopy. Asillustrated in Figure 8A, Anx 1 labeling appeared par-ticularly intense in the cytoplasm near the nucleuswhich was also stained, although to a lesser extent.Anx 2 labeling (Fig. 8B) presented numerous fluores-cent speckles both on a cytoplasmic network and alongthe plasma membrane. A finely punctuated nucleardistribution was also present. These findings agreewith the immunohistochemical observations in the in-tact tissue presented in Figure 7.
DISCUSSION
In an attempt to better understand the roles of Anxs,correlations of their expressions and localizations withthe developmental events of rat liver organogenesismay provide clues as to the likely functions of theseproteins. In the present study, we have demonstratedthat rat liver perinatal development involves temporaland spatial controls of Anx expressions in the hepato-cytes. The time course of the appearance of the differ-ent Anxs and their characteristic cellular localizationsin the cytoplasm and on the membranes, as well as inthe nucleus, are in accordance with the known pluri-potent and pleiotropic roles of these proteins.
Annexin 1 and Annexin 2
Our biochemical and immunohistochemical studieshave demonstrated the transient high expressions ofAnxs 1 and 2 in hepatocytes, at the transcriptional aswell as translational levels, during the perinatal pe-riod. Anxs 1 and 2 first appear in the cytoplasm, whereprotein synthesis in known to take place; thereafter,they seem to migrate toward the membrane and, to alesser extent, in the nucleus. Indeed, Anx 1 expressionin the fetal rat hepatocytes can be correlated with thepresence of max 3, an Anx related to mammalian Anx1, in the developing liver of the killifish medaka (Os-terloh et al., 1998). In 13-d-old rat and in adult rat, Anx1 disappears entirely, whereas Anx 2 remains only onthe inner face of the plasma membrane (Massey-Har-roche et al., 1998).
The elevated expressions of Anxs 1 and 2 in hepato-cytes during the perinatal period correspond to a periodof differentiation rather than proliferation, as demon-strated by the intermediate levels of cyclin A mRNAs inliver extracts and the decreasing percentages of S 1G2/M phases in the isolated liver cells. They can becompared with the expression of corticosterone, themajor corticosteroid in the rat. In the plasma, the cor-ticosterone level increases from gd 17.5 to gd 19.5,when it reaches a peak value, and then falls to gd 21.5(Martin et al., 1977). During the postnatal period, it is
Fig. 6. A: Western blot analysis of Anx 1, 2, 5, or 6 in extracts fromcultured hepatocytes maintained either under unstimulated conditions orin the presence of either EGF (50 ng/ml) or HGF (25 ng/ml), in 20 mMsodium pyruvate, for 48 or 96 hr. Either EGF or HGF was added just afterseeding and every day thereafter with the renewal of medium. After 48 hr,Anx 1 and 2 expressions resume and the proteins are strongly expressedat increasing levels; Anx 5 and 6 expressions are enhanced. B: Timecourse of [methyl-3H]thymidine incorporation into DNA in unstimulated(black) and EGF- (grey) or HGF- (light grey) stimulated adult rat primarycultures for 24, 48, 72, and 96 hr. For abbreviations, see list.
211ANNEXINS IN DIFFERENTIATING HEPATOCYTE
extremely low until d 12. It rises significantly only by d14 to peak on d 24 (Henning, 1978). As the analysis ofmouse Anx 1 gene promoter has revealed the presenceof two simple glucocorticoid response element (GRE)sequences (Horlick et al., 1991), the transient expres-sion of Anx 1 during the perinatal period might betriggered by this hormone. This assumption, however,is not supported by experimental results. The Anx 1gene did not display any significant sensitivity to dex inour in vivo experiments. In A431 cells, the GRE in theAnx 1 gene is nonfunctional, and the endogenous Anx 1gene expression is unaffected by dex at the mRNA andprotein levels (Donnelly and Moss, 1998). Moreover,
Antonicelli et al., (2001) were not able to show a directrole for the GRE under steroid stimulation of themouse Anx 1 promoter. Taken together, these datasuggest that the triggering of the transient Anx 1 ex-pression is not due to the peak of plasma corticosteroneobserved during the fetal period. In contrast, Anx 2gene displayed sensitivity to glucocorticoids in vivo, asdemonstrated by the elevated levels of Anx 2 mRNA inliver of dex-treated offsprings. Thus, the transient highAnx 2 expression might be regulated by the peak ofplasma corticosterone.
During the fetal period, the transient Anx 1 and 2expressions correlate with the peaks of expression of
Anx 1 d 3
A
Anx 2 d 1
B C
Anx 2 d 13
Anx 5 d 3
D
Anx 6 d 3
E F
Anx 6 d 13
Fig. 7. Immunohistochemical localization of Anxs 1, 2, 5, and 6 indeveloping rat liver. Sections of formol-zinc fixed livers were studiedimmunohistochemically by using the monoclonal antibodies to Anx 1, 2,or 6 and the polyclonal antibody to Anx 5 in combination with the Tyra-mide Signal Amplification and the AEC chromogen. A: In the liver from3-d-old rat, Anx 1 is essentially found in the hepatocytes as intracyto-plasmic speckles. B: In the liver from 1-d-old rat, Anx 2 appears asspeckles over the cytoplasm and lines along the plasma membrane of the
hepatocytes; the nucleus is also stained. C: In the liver from 13-d-old rat,the immunostaining of Anx 2 is restricted to the cell membrane. D: In theliver from 3-d-old rat, the antibody to Anx 5 gives punctate cytoplasmiclabeling and significant nuclear staining. Note that in the liver from 3-d-old(E) and 13-d-old (F) rats, the staining patterns of Anx 6 are very similar tothose of Anx 2. All controls were negative. For abbreviations, see list.Scale bars 5 15 mm in A–F.
Fig. 8. Comparison of the distributions of Anxs 1 and 2 in hepatocytesisolated from 1-d-old rat liver after 24 hr of culture. Immunocytochemicallocalizations were performed in permeabilized cells labeled with mono-clonal antibodies to Anx 1 or 2 and analyzed by scanning laser confocalmicroscopy. Consecutive sections were 0.2 mm apart. A: Anx 1 distrib-utes diffusely throughout the cytoplasm but also appears concentrated in
the perinuclear area and associated with some intracellular compart-ments, most likely around glycogene granules. B: Anx 2 distributionappears mainly around the nucleus and some intracytoplasmic compart-ments. Plasma membrane is partially labeled. Some intranuclear punctaare also present. All controls were negative. For abbreviations, see list.Scale bars 5 5 mm.
212 DELLA GASPERA ET AL.

EGF, HGF, and their receptors. EGF receptor expres-sion and EGF binding to the plasma membrane aredetected in the rat liver as early as gd 17, and bothincrease more than twofold within 24 hr. By the end ofd 1, there is a precipitous drop in the number of recep-tors to less than 20% of the adult level (De et al., 1991).HGF, a mesenchymal-derived factor, acts on neighbor-ing epithelia by coupling with its receptor c-Met, amembrane-spanning tyrosine kinase. HGF and c-MetmRNA expressions have been demonstrated in the rat,from gd 14 to d 19 (Defrances et al., 1992). This findingis in close accordance with our results showing thatAnx 1 and 2 expressions resume in primary cultures ofadult rat hepatocytes that reach the G1/S boundaryand complete the cell cycle after stimulation by eitherEGF/ or HGF/hydrocortisone hemisuccinate. This find-ing is also in agreement with the results published byMasaki et al. (1994) who reported elevated expressionsof Anxs 1 and 2 in adult rat liver after damage orpartial hepatectomy. Moreover, Anx 1, which is notfound in normal or chronic hepatitis human liver, isoverexpressed in tumorous and nontumorous regions ofhuman hepatocellular carcinoma (Masaki et al., 1996).Anx 2 is also a growth-regulated gene whose expres-sion is significantly increased in various human can-cers. Taken together, these findings strongly suggestthat Anx 1 and 2 synthesis in the developing or regen-erating hepatocytes are up-regulated by EGF andHGF. Moreover, the changes in Anx 1 and 2 levels inrat hepatocytes during the perinatal period can be com-pared with those observed in rat developing pancreaticislets in which Anx 1 and 2 expressions gradually in-crease during postnatal development, simultaneouslyto those of EGF receptor, pp60src, and protein kinase C(Ohnishi et al., 1994). Thus, the time courses of appear-ance of Anxs 1 and 2, although quite different in bothorgans, seem to be solely regulated by growth factors.
It has been proposed that Anx 1 is implicated inphospholipase inhibition and secretion. Anx 1 can bephosphorylated on Tyr21 by the protein tyrosine kinaseactivity of two receptors, EGF receptor (Huang et al.,1986) and c-Met (Skouteris and Schroder, 1996); it canalso be phosphorylated by protein kinase C on Ser andThr belonging to the N-terminal tail. A role for Tyr21
phosphorylated Anx 1 in proliferation is strongly sug-gested by studies that have demonstrated that thelevel of phosphorylated Anx 1 is significantly increasedin cultures of A549 cells 48–72 hr after coaddition ofHGF/dex, in comparison with the level observed up to24 hr (Skouteris and Schroder, 1996). Moreover Anx 1expression and phosphorylation are up-regulated dur-ing liver regeneration and transformation (De Coupadeet al., 2000). Phosphorylated Anx 1 may be implicatedin the intracellular translation of the mitogenic signalsin A 549 cells stimulated with HGF. The phosphoryla-tion of Anx 1 results in the release of cPLA2 activityand the stimulated production of prostaglandin E2(PGE2) which, in turn, induces cell proliferation. Asprostaglandins E2 and F2a trigger hepatocyte prolifer-
ation (Adachi et al., 1995), it may be postulated thatgrowth factor mitogenic action on hepatocytes is alsotranslated by phosphorylated Anx 1 and prostaglan-dins. Another mechanism involving activation of PLA2by a mitogen-activated protein kinase (MAP kinase),however, has also been reported (Adachi et al., 1995).Nevertheless, it can be suggested that both mecha-nisms which release cPLA2 activity, Anx 1 phosphory-lation and mitogen-activated MAP kinase, cooperate,perhaps depending on physiological conditions. Thepossible role of Anx 1 in the intracellular translation ofmitogenic signals correlates well with its cytoplasmiclocalization. In addition, participation in nuclear-re-lated events is also suggested by its partial movementtoward the nucleus.
Anx 2 is implicated in several membrane transportevents along the Ca21-regulated secretory pathway(Creutz, 1992). Anx 2 is also involved in the endocytoticprocesses in which it affects fusion properties and/orthe organization and intracellular location of endoso-mal membranes in a Ca21-independent manner (Jostet al., 1997; Turpin et al., 1998). Biochemical propertiesand intracellular distribution of Anx 2 are regulated bycomplex formation with p11, a member of the S100protein family. Anx 2, which is essentially localized onthe plasma membrane of differentiated hepatocytes(Massey-Harroche et al., 1998), may play a role in theintensive intracellular trafficking along with otherAnxs, particularly Anx 6 (see below). Anx 2 can also bephosphorylated by the activity of pp60src, a cytosolicnonreceptor protein tyrosine kinase (Huang et al.,1986), and by protein kinase C. However, the mecha-nism by which phosphorylated Anx 2 may play a role incell processes remains largely unknown.
Annexin 5 and Annexin 6
In strong contrast to Anx 1 and 2 expressions, Anx 5and 6 expressions can be detected in hepatocytes atevery stage of perinatal development, at both mRNAand protein levels. After birth, the levels of Anx 5 and6 mRNAs decline, whereas Anx 5 and 6 protein con-tents increase, suggesting that up-regulation of bothproteins in the postnatal period may occur through aposttranscriptional process. Moreover, Anx 5 and 6mRNA expressions are slightly increased by prenataldex administration, and Anx 5 and 6 synthesis is mod-erately enhanced in primary cultures of adult rat hepa-tocytes that enter S phase after stimulation by EGF orHGF. The presence of Anx 5 in the rat hepatocytes inthe early fetal period parallels the expression of max 1,a member of the Anx family related to mammalian Anx5, in the developing liver of the killifish medaka (Os-terloh et al., 1998).
A striking feature is the wide distribution of Anx 5 inthe nucleus and the cytoplasm, as well as on theplasma membrane of the fetal and neonatal hepato-cytes. This finding is in agreement with our previousultrastructural studies of adult rat hepatocytes (Wein-man et al., 1994), which show the presence of Anx 5 in
213ANNEXINS IN DIFFERENTIATING HEPATOCYTE

epithelial cells lining the bile ducts (Boustead et al.,1993) and Anx 5 association with the internal nu-clear matrix fraction in chicken liver (Altieri et al.,1996). Anx 5 is known to bind to phosphatidylserine.It has been demonstrated that in the fetal rat brainhelicase 2 and DNA (cytosine-5) methyltransferaseare candidates for Anx 5-binding proteins (Ohsawaet al., 1996). Anx 5 also exhibits ion channel, an-tiphospholipase, and antiprotein kinase C activities.Taken together, these findings strongly suggest thathepatocytic Anx 5 has a multifunctional and pleio-tropic role and regulates many Ca21-dependent pro-cesses in the nucleus and the cytoplasm and also onthe plasma membrane.
Anx 6 concentrates on the hepatocyte plasma mem-brane during the postnatal period, as already shown inhepatocytes of adult rat (Massey-Harroche et al.,1998). This finding is also consistent with the presenceof Anx 6 on apical endosomes in rat liver hepatocytes(Ortega et al., 1998). Altogether, these findings rein-force the assumption that Anx 6, together with otherAnxs and proteins, is involved in vesicular traffickingin hepatocytes and other cells (Jost et al., 1997; Turpinet al., 1998).
In conclusion, our studies demonstrate that Anxs 1,2, 5, and 6 have individual modes of transcriptionaland translational regulation and specific locations dur-ing hepatocyte differentiation. These proteins, withpluripotent and pleiotropic physiological functions,might play unique roles at well defined periods of liverontogenesis and participate in many membrane-, cyto-plasmic-, or nuclear-related events.
EXPERIMENTAL PROCEDURESChemicals, Reagents, and Cell Culture Media
Chemicals and reagents of the highest purity availablewere purchased from Merck (Darmstadt, Germany),Sigma Chemical (St. Louis, MO), or Boehringer (Mann-heim, Germany), cell culture media from Life Technolo-gies (Paisley, UK), the 18-mer oligo-dT and the 24-meroligonucleotide [ACGGTATCTGATCGCTTCGAACC, 59to 39] from the Pasteur Institute (Paris, France),[g-32P]dCTP (111 TBq/mmol), [g-32P]ATP (259 TBq/mmol) and [methyl-3H]thymidine (248 GBq/mmol) fromICN Pharmaceuticals (Costa Mesa, CA), the MegaprimeDNA labeling system, Hybond-N membranes, Hyper-films MP, the enhanced chemiluminescence (ECL) West-ern blotting analysis system, and the glutathione-S-transferase (GST) gene fusion system from AmershamPharmacia Biotech (Orsay, France), reagents for poly-acrylamide gel electrophoresis, and 0.2-mm polyvinyli-dene difluoride (PVDF) membrane from Bio-Rad Labora-tories (Hercules, CA).
Antibodies
Monoclonal antibodies (mAb) to Anxs 1, 2, 4, and 6were purchased from Zymed Laboratories (SouthSan Francisco, CA). Purified recombinant Anx 5 wasbiosynthesized from a 1.6-kb NotI fragment frompNN 08 (Pepinsky et al., 1988) by using the GST
gene fusion system, and a polyclonal antibody waselicited in rabbit and purified by affinity chromatog-raphy. All antibodies reacted specifically with theirrespective antigens in Western blotting experiments.Monoclonal antibodies to vimentin and cytokeratin 8were purchased from Sigma Immunochemicals (St.Louis, MO). Purified mouse IgG1 and fluorescein iso-thiocyanate- (FITC) or phycoerythrin-conjugatedgoat anti-mouse IgGs were obtained from CaltagLaboratories (Burlingame, CA).
Animals
Sprague-Dawley timed pregnant rats (CD, CharlesRiver, France) were the source of all prenatal and post-natal livers. In some experiments, dams were injectedsubcutaneously from gd 15 to gd 21 with 0.4 mg/kg ofdex 21-phosphate dissolved in water.
Isolation of Cells from Fetal and PostnatalRat Liver
Cells were isolated from fetal and postnatal rat livers(Braut-Boucher et al., 1988). Briefly, livers wereminced and incubated in Ca21-Mg21 free Dulbecco’sPBS, enriched with glucose (1 g/l) and containing tryp-sin (0.25%) for 15 min at 37°C. The tissue fragmentswere suspended in MEM and dissociated by shaking inthe presence of glass microspheres. Isolated cell viabil-ity was not less than 96%.
Cultured Hepatocytes from Adult Rat
Hepatocytes from adult male Sprague-Dawley ratswere isolated by a two-step collagenase perfusion pro-cedure (Guguen et al., 1975), seeded, and cultured in amixture of 75% MEM and 25% medium 199 supple-mented with bovine insuline (5 mg/ml) and hydrocorti-sone hemisuccinate (7 3 1027 M). The hepatocyteswere stimulated by either epidermal growth factor(EGF) (50 ng/ml) or hepatic growth factor (HGF) (25ng/ml) in combination with 20 mM of sodium pyruvate(Loyer et al., 1996). DNA replication was estimatedby measuring [methyl-3H]thymidine incorporation (2mCi/ml, final concentration).
Quantification of Total mRNA
Liver RNA was prepared according to Chirgwin etal. (1979). To compare total cellular mRNA and spe-cific mRNA expressions, slot blots of liver RNA wereperformed as explained in detail elsewhere (Mer-cadier and Dubus, 1991). Briefly, serial dilutions ofdenatured RNA were fixed to a Hybond-N nylonmembrane. Total mRNA for each liver RNA samplewas determined by hybridization of the membrane tothe g-32P end-labeled 18-mer oligo-dT. The mem-brane was exposed to Hyperfilms MP to obtain den-sitometric scans by using a Schimadzu CS 9000 den-sitometer (Tokyo, Japan). To control the RNAamount bound to the filter, the stripped membranewas rehybridized to the g-32P end-labeled 24-meroligonucleotide complementary to a sequence of therat 18S ribosomal RNA sequences (Boheler et al.,
214 DELLA GASPERA ET AL.

1991). All values of total mRNA were normalized to18S ribosomal RNA and expressed as arbitrary ODunits.
Quantification of Specific mRNAs
The methods used here, including probe preparation,have been published in detail (Weinman et al., 1991).For Northern blot analysis, Hybond-N membraneswere successively hybridized with 32P labeled probes. A1.3-kb EcoRI fragment from pUC13 (Wallner et al.,1986), a 0.8-kb EcoRI fragment from pUC18 (Huang etal., 1986), a 1.6-kb NotI fragment from pNN 08 (Pep-insky et al., 1988), and a 2-kb EcoRI fragment frompAT 153 (Crompton et al., 1988), were used as Anxs1,2,5, and 6 probes, respectively. For the detection ofb-actin mRNA, a 1.2-kb PstI fragment from pBR322(Alonso et al., 1986), which hybridizes essentially withb-cytoplasmic actin mRNA, was used. A 1.7-kb EcoR1fragment of a Bluescript vector was used as cyclin Aprobe (Wang et al., 1990). The probes were labeled with[a-32P]dCTP by random primer extension. To controlthe amount of RNA actually bound in each lane, themembranes were also hybridized with the g-32P end-labeled 24-mer oligonucleotide complementary to 18Sribosomal RNA sequences. Hybridization signals werequantitated by scanning densitometry. All values ofspecific mRNAs were normalized to 18S ribosomalRNA and expressed as arbitrary OD units.
Western Blot Analysis
The protein content of the samples was estimatedaccording to Bradford (1976). Immunoblot analysis wasperformed according to Laemmli (1970) and Towbin etal. (1979). Samples of rat liver lysed in Laemmli sam-ple buffer were analysed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) on13% gels and electroblotted to 0.2-mm PVDF mem-branes. The proteins were detected by using the pri-mary antibodies to Anxs, or actin, in combination withthe appropriate peroxidase-conjugated secondary anti-body and the ECL system.
Immunofluorescence and Cell Cycle Analyses ofIsolated Liver Cells by Flow Cytometry
For immunofluorescence studies, 106 liver cells werefixed and permeated, by using the Fix and Perm kitfrom Caltag Laboratories (Burlingame, CA) and incu-bated with the antibodies to vimentin, cytokeratin 8,Anx 1 or Anx 2 (1 mg/ml) which were then revealed withFITC- or phycoerythrin-conjugated goat anti-mouseIgG1. Purified mouse IgG1 was used as a negativecontrol. Cells were analysed by flow cytometry by usingan Ortho Cytofluorograph 30 L (Ortho Diagnostic Sys-tems). Calibration and standardization were per-formed with fluorescent microspheres. Fluorescence in-tensity was given by the ratio (A-B)/C , where A, B, andC were the mean channel values for the specific anti-body, purified isotype IgG1, and microspheres, respec-tively. Cell cycle analysis was performed according toVindelov et al. (1983).
Immunohistochemistry of Rat Liver
For immunohistochemistry, pieces of rat liver werefixed in formol-zinc and embedded in paraffin wax.Sections of 3 mm were de-waxed, and nonspecific pro-tein-binding sites were blocked by incubation in Trisbuffer pH 7.4 containing serum from the same animalin which the secondary antibody was raised (5%, v/v).To detect Anxs 1, 2, 4, 5, and 6, the sections wereincubated with the corresponding primary antibody ata final concentration of 5 mg/ml in PBS. Primary anti-body labeling was visualized by Tyramide Signal Am-plification (TSA) technology (Adams, 1992) by usingthe TSA-Indirect kit for chromogenic immunohisto-chemistry from NEN Life Science Products (Boston,MA) and amino-ethylcarbazole (AEC) as chromogenicsubstrate. The sections were counterstained with Har-ris hematoxylin. Controls used were a purified IgGfraction from the same animal in which the primaryantibody was raised at a final concentration of 5 mg/mlin PBS. Moreover, the specificity of the Anx 5 immu-nolabeling was assessed by comparison with labelingpatterns obtained with anti-Anx 5 preadsorbed with a10 M excess of the recombinant Anx 5 (3 mg Anx 5 permg IgG). In no case was staining above backgroundobserved.
Fluorescence Confocal Microscopy Analysis ofIsolated Liver Cells
For fluorescence confocal microscopy analysis, iso-lated cells from 1-d-old rat liver were seeded on 8-wellLabTek (Nunc Inc., Naperville, IL) in MEM supple-mented with 10% heat inactivated fetal calf serum.After cell attachment (24 hr later), cells were processedfor indirect immunofluorescence and observed by con-focal microscopy (Bio-Rad 1024 confocal microscope)with a 360 optique (Nikon) as described in detail else-where (Turpin et al., 1998). In short, mAb anti-Anx 1 or2 (5 mm/ml) was used as primary antibody and FITC-conjugated goat anti-mouse IgGs (5 mm/ml) as second-ary antibody. Control stainings were carried out byreplacing the first antibody by nonspecific purified IgGsor by omitting the antibody. No staining above back-ground was observed.
ACKNOWLEDGMENTS
The authors thank R.B. Pepinsky and S.E. Moss forkindly providing the Anx cDNA probes used in thiswork and Michel Kornprobst for the cell sorting fluo-rescence analysis, Denise Glaise for the primary cul-tures of rat hepatocytes, Isabelle Carriere and GinetteBoissonot for help in immunocytochemistry and Isa-belle Bouchaert for expert assistance with confocal mi-croscopy. They also thank Pierre Mehul for preparingthe Figures and James Mac Cabe (Centre Universitairede Langues) for preparing the manuscript.
REFERENCES
Adachi T, Nakashima S, Saji S, Nakamura T, Nozawa Y. 1995. Rolesof prostaglandin production and mitogen-activated protein kinase
215ANNEXINS IN DIFFERENTIATING HEPATOCYTE

activation in hepatocytes growth factor-mediated rat hepatocyteproliferation. Hepatology 21:1668–1674.
Adams JC. 1992. Biotin amplification of biotin and horseradish per-oxidase signals in histochemical stains. J Histochem Cytochem40:1457–1463.
Alonso S, Minty A, Bourlet Y, Buckingham M. 1986. Comparison ofthree actin-coding sequences in the mouse evolutionary relation-ships between the actin genes of warm-blooded vertebrates. J MolEvol 23:11–22.
Altieri F, Maras H, Turano C. 1996. Nuclear matrix localization ofannexin V in chicken liver. Biochem Biophys Res Commun 225:448–454.
Antonicelli F, De Coupade C, Russo-Marie F, Le Garrec Y. 2001.CREB is involved in mousse annexin A1 regulation by cAMP andglucocorticoids. Eur J Biochem 268:62–69.
Boheler KR, Carrier L, de la Bastie D, Allen PD, Komadja M, Mer-cadier JJ, Schwartz K. 1991. Skeletal actin mRNA increases in thehuman heart during ontogenic development and is the major iso-form of control and failing adult heart. J Clin Invest 88:323–330.
Boustead CM, Brown R, Walker JH. 1993. Isolation, characterizationand localization of annexin V from chicken liver. Biochem J 291:601–608.
Bradford MMW. 1976. A rapid and sensitive method for the quanti-tation of microgram quantities of protein. Anal Biochem 72:248–254.
Braut-Boucher F, Achard-Ellouk S, Hoellinger H, Pauthe-Daide D ,Henry M. 1988. Isolated and cultured rat hepatocytes as models forstudying the cytotoxicity of two triterpenoid saponins. In: GuillouzoA, editor. Liver cells and drugs, Colloque INSERM: John LibbeyEurotext Ltd. p 379–383.
Chirgwin JM, Przybyla AP, MacDonal RJ, Rutter WJ. 1979. Isolationof biologically active ribonucleic acid from sources enriched in ribo-nucleases. Biochemistry 18:5294–5299.
Creutz CE. 1992. The annexins and exocytosis. Science 258:924–931.Crompton MR, Owens RJ, Totty NF, Moss SE, Waterfield MD,
Crumpton MJ. 1988. Primary structure of the human, membrane-associated Ca21-binding protein p68: a novel member of a proteinfamily. EMBO J 7:21–27.
De BK, Brown TL, Suchy FJ. 1991. Ontogeny of epidermal growthfactor receptor tyrosine kinase in rat liver. Am J Physiol 260:G290–G298.
De Coupade C, Gillet R, Bennoun M, Briand P, Russo-Marie F, SolitoE. 2000. Annexin 1 expression and phosphorylation are upregulatedduring liver regeneration and transformation in antithrombin IIISV40 T large antigen transgenic mice. Hepatology 31:371–380.
Defrances MC, Wolf HK, Michalopoulos GK, Zarnegar R. 1992. Thepresence of hepatocyte growth factor in the developing rat. Devel-opment 116:387–395.
Donnelly SR, Moss SE. 1997. Annexins in the secretory pathway. CellMol Life Sci 53:533–538.
Donnelly SR, Moss SE. 1998. Functional analysis of the human an-nexin I and VI gene promoters. Biochem J 332:681–687.
Gerke V, Moss SE. 1997. Annexins and membrane dynamics. BiochimBiophys Acta 1357:129–154.
Greengard O, Federman M, Enox WE. 1972. Cytomorphometry ofdeveloping rat liver and its application to enzymic differentiation.J Cell Biol 52:261–272.
Guguen C, Guillouzo A, Boisnard M, Le Cam A, Bourel M. 1975.Etude ultrastructurale des hepatocytes de rat cultives en mono-couches en presence d’hemisuccinate d’hydrocortisone. Biol Gastro-enterol 8:223–231.
Henning SJ. 1978. Plasma concentration of total and free corticoste-rone during development in the rat. Am J Physiol 325:E451–E456.
Horlick KR, Cheng IC, Wong W, Wakeland EK, Nick HS 1991. Mouselipocortin 1 gene structure and chromosomal assignment: gene du-plication and the origins of a gene family. Genomics 10:365–374.
Huang KS, Wallner BP, Mattaliano RJ, Tizard R. 1986. Two human35 kd inhibitors of phospholipase A2 are related to substrates ofpp60v-src and the epidermal growth factor receptor kinase. Cell46:191–199.
Jost M, Zeuschner D, Seeman J, Weber K, Gerke V. 1997. Identifica-tion and characterization of a novel annexin-membrane interaction:
Ca21 is not required for the association of Annexin II with earlyendosomes. J Cell Sci 110:221–228.
Kaetzel MA, Hazarika P, Dedman JR. 1989. Differential tissue ex-pression of three 35-kDa annexin calcium-dependent phospholipid-binding proteins. J Biol Chem 264:14463–14470.
Karasik A, Pepinsky RB, Shoelson SE, Kahn CR. 1988. Lipocortins 1and 2 as substrates for the insulin receptor kinase in rat liver. J BiolChem 263:11862–11867.
Laemmli UK. 1970. Cleavage of structural proteins during the assem-bly of the head of bacteriophage T4. Nature (Lond) 227:680–685.
Lieman S, Huber R. 1997. Three-dimensional structure of annexins.Cell Mol Life Sci 53:516–521.
Loeb JN, Borek C, Yeung LL. 1973. Suppression of DNA synthesis inhepatoma cells exposed to glucocorticoids. Proc Natl Acad Sci USA70:3852–3856.
Loyer P, Cariou S, Glaise D, Bilodeau M, Baffet G, Guguen-GuillouzoC. 1996. Growth factor dependence of progression through G1 ans Sphases of adult rat hepatocytes in vitro. J Biol Chem 271:11484–11492.
Martin CE, Cake MH, Hartmann PE, Cook IF. 1977. Relationshipbetween foetal corticosteroids, maternal progesterone and parturi-tion in the rat. Acta Endocr 84:167–176.
Masaki T, Tokuda M, Fujimura T, Ohnishi M, Tai Y. 1994. Involve-ment of annexin I and annexin II in hepatocyte proliferation: canannexins I and II be markers for proliferative hepatocytes. Hepa-tology 20:425–435.
Masaki T, Tokuda M, Ohnishi M, Watanabe S, Fujimura T. 1996.Enhanced expression of the protein kinase substrate Ann in humanhepatocellular carcinoma. Hepatology 24:72–81.
Massey-Harroche D, Mayran N, Maroux S. 1998. Polarized localiza-tions of annexins I, II, VI, and XIII in epithelial cells of intestinal,hepatic, and pancreatic tissues. J Cell Sci 111:3007–3015.
Mathew JK, Krolak JM, Dedman JR 1986. Calcimedins: purificationand characterization from chicken gizzard and rat and bovine liv-ers. J Cell Biochem 32:223–234.
Mercadier JJ, Dubus I. 1991. Assay of natriuretic factor messengerribonucleic acid. Methods Neurosci 5:22–34.
Ohnishi M, Tokuda Masaki MT, Fujimura T, Tai Y. 1994. Changes inannexin I and II levels during the postnatal development of ratpancreatic islets. J Cell Sci 107:2117–2125.
Ohsawa K, Imai Y, Ito D, Kohsaka S. 1996. Molecular cloning andcharacterization of annexin V-binding proteins with highly hydro-philic peptide structure. J Neurochem 67:89–97.
Ortega D, Pol A, Biermer M, Jackle M, Henrich C. 1998. Annexin VIdefines an apical endocytic compartment in rat liver hepatocyte.J Cell Sci 111:261–269.
Osterloh D, Wittbrodt J, Gerke V. 1998. Characterization and devel-opmentally regulated expression of four annexins in the killifishmedaka. DNA Cell Biol 17:835–847.
Pepinsky RB, Tizard R, Mattaliano RJ, Sinclair JK, Miller GT. 1988.Five distinct calcium and phospholipid binding proteins share ho-mology with lipocortin I. J Biol Chem 263:10799–10811.
Rainteau DP, Weinman SJ, Kabaktchis CAM, Smith VL, Kaetzel MA,Dedman JR, Weinman SJ. 1988. The expression of the 35- and67-kDa calcimedins is dependent on thyroid hormones. J Biol Chem263:1244–1248.
Raynal P, Pollard HB. 1994. Annexins: the problem of assessing thebiological role for a gene family of multifunctional calcium- andphospholipid-binding proteins. Biochim Biophys Acta 1197:63–93.
Schmidt C, Bladt F, Goedecke S, Brinkmann V, Zschieche W, SharpeM, Gherardi E, Birchmeier C. 1995. Scatter factor/hepatocytegrowth factor is essential for liver development. Nature 373:699–702.
Shadle PJ, Gerke V, Weber K. 1985. Three Ca21-binding proteinsfrom porcine liver and intestine differ immunologically and physi-cochemically and are distinct in Ca21 affinities. J Biol Chem 260:16354–16360.
Skouteris GG, Schroder CH. 1996. The hepatocyte growth factor re-ceptor kinase-mediated phosphorylation of lipocortin-1 transducesthe proliferating signal of the hepatocyte growth factor. J Biol Chem271:27266–27273.
216 DELLA GASPERA ET AL.

Towbin H, Staehelin T, Gordon J. 1979. Electrophoretic transfer ofproteins from polyacrylamide gels to nitrocellulose sheets: proce-dure and some applications. Proc Natl Acad Sci USA 76:4350–4354.
Turpin E, Russo-Marie F, Dubois T, de Paillerets C, Alfsen A, BomselM. 1998. In adrenocortical tissue, annexins II and VI are attachedto clathrin coated vesicles in a calcium-independent manner. Bio-chim Biophys Acta 1402:115–130.
Vindelov LL, Christensen IJ, Nissen NI. 1983. Standardization ofhigh resolution flow cytometry DNA analysis by simultaneous useof chicken and trout red blood cells as internal reference standard.Cytometry 3:328–331.
Wallner BP, Mattaliano RJ, Hession C, Cate RL, Tizard R. 1986.Cloning and expression of human lipocortin, a phospholipase A2
inhibitor with potential anti-inflammatory activity. Nature (Lond)320:77–81.
Wang J, Chenivesse X, Henglein B, Brechot C. 1990. Hepatitis B virusintegration in a cyclin A gene in a hepatocellular carcinoma. Nature(Lond) 343:555–557.
Weinman J, Della Gaspera B, Dautigny A, Pham Dinh D, Wang J,Nojima H, Weinman SJ. 1991. Developmental regulation of calmod-ulin gene expression in rat brain and skeletal muscle. Cell Regul2:819–826.
Weinman J, Feinberg JM, Rainteau DP, Della Gaspera B, Wein-man SJ. 1994. Annexins in rat enterocytes and hepatocytes:an immunogold electron-microscope study. Cell Tissue Res 278:389 –397.
217ANNEXINS IN DIFFERENTIATING HEPATOCYTE
Copyright © 2022 FDOKUMEN




















