
Membrane fusion protein synexin (annexin VII) as a Ca2+/GTP sensor in exocytotic secretion
6
Proc. Natl. Acad. Sci. USA Vol. 93, pp. 10797-10802, October 1996 Cell Biology Membrane fusion protein synexin (annexin VII) as a Ca2+/GTP sensor in exocytotic secretion HUNG CAOHUY, MEERA SRIVASTAVA, AND HARVEY B. POLLARDt Laboratory of Cell Biology and Genetics, National Institute of Diabetes and Digestive and Kidney Diseases, National Institutes of Health, Bethesda, MD 20892 Communicated by Bernard Witkop, National Institutes of Health, Bethesda, MD, June 10, 1996 (received for review April 20, 1996) ABSTRACT Exocytotic membrane fusion and secretion are promoted by the concerted action of GTP and Ca2 , although the precise site(s) of action in the process are not presently known. However, the calcium-dependent membrane fusion reaction driven by synexin (annexin VII) is an in vitro model for this process, which we have now found to be further activated by GTP. The mechanism of fusion activation de- pends on the unique ability of synexin to bind and hydrolyze GTP in a calcium-dependent manner, both in vitro and in vivo in streptolysin 0-permeabilized chromaffin cells. The re- quired [Ca2+] for GTP binding by synexin is in the range of 50-200 ,AM, which is known to occur at exocytotic sites in chromaffin cells, neurons, and other cell types. Previous immunolocalization studies place synexin at exocytotic sites in chromaffin cells, and we conclude that synexin is an atypical G protein that may be responsible for both detecting and mediating the Ca2+/GTP signal for exocytotic membrane fusion. GTP and its nonhydrolyzable analogue guanosine 5'-[y- thio]triphosphate (GTP[,yS]) are known to promote Ca2+- dependent exocytotic secretion from many cell types by a mechanism thought to involve as yet unknown proteins in the GTPase superfamily (1-4). Specific members of this super- family have been considered as mediators of these GTP effects on exocytosis, including heterotrimeric G proteins (5-9) and low molecular weight ras-like proteins such as Rab (10-15) and ARF (16). The current "fusion machine" hypothesis (17-19) envisions a core complex formed between plasma membrane syntaxin and SNAP-25 and the synaptic vesicle protein syn- aptobrevin/VAMP (20), with vesicular synaptotagmin puta- tively identified as a low-affinity calcium sensor that interacts with regulatory syntaxin 1 (21, 22). However, none of the proteins presently identified in the hypothetical fusion ma- chine have actually been shown to be activated by GTP or, indeed, even to fuse membranes (17, 23-25). Therefore, it has been suggested that other GTP-binding proteins, as yet uniden- tified, might control the activity of the fusion complex (19, 25). The site of GTP action in exocytosis has been hypothesized to be closely associated with the site of calcium action in a common pathway (2, 5, 26, 27). The most recent consensus on the nature of this calcium binding site is that it is involved in docking and fusion and that the affinity of the site for Ca2+ may be in the range of 50-200 ,tM (28-36). From this perspective we have considered whether the ubiquitously distributed membrane fusion protein synexin (annexin VII, Anx7; refs. 37-59), with an intrinsic Kd for Ca2+ of '200 ,iM (41, 42), might contribute to the hypothetical GTP/Ca2+- sensitive exocytosis site. Experiments to be described below advance the new concept that purified synexin is also a unique Ca2+-conditional GTP binding protein. We show that synexin fusion activity in vitro is substantially enhanced upon binding to GTP and deactivated by GTP hydrolysis. In addition, studies with intact and streptolysin 0 (SLO)-permeabilized chromaf- fin cells are presented to indicate that synexin behaves in a similar fashion in vivo. MATERIALS AND METHODS Assays for Synexin Using Chromaffin Granules and Phos- phatidylserine (PS) Liposomes. Recombinant human synexin was expressed and isolated as described (60). Isolation and purification of chromaffin granules were performed as de- scribed by Brocklehurst and Pollard (61). Large unilamellar PS liposomes ("LUV") were prepared by the swelling method (62) and fusion analyzed (55, 56). The aggregation assay for synexin activity was performed as described by Brocklehurst and Pollard (61), and nucleotide effects on [Ca2+] were corrected (63) and verified by calcium-selective electrodes. Photoaffinity Labeling of Synexin by 8N3- [y-32P]GTP and Labeling Under Nondenaturing Conditions. The photoaffin- ity-labeling reaction was carried out essentially as described (64, 65), with minor modifications. 32p incorporation was analyzed by SDS/PAGE and quantitated by either autora- diography at -80°C or PhosphorImager counting (Molecular Dynamics). Assays for [35S]GTP[,yS] binding to synexin were also developed using the filtration method described (66). Measurement of Synexin GTPase Activity. Synexin samples (2 ,ug) were incubated for 30 min at 30°C with 0.13 ,uM [a-32P]GTP (Amersham, 3000 Ci/mmol) in a total volume of 10 ,ul of buffer D (25 mM Tris HCl, pH 7.5/250 mM NaCl/1 mM DTT/10 mM ,3-glycerolphosphate) containing 1 mM EGTA (for CaCl2) or 1 mM EDTA (for MgCl2) and various concentrations of CaCl2 or other divalent cations. The reac- tions were stopped by adding an equivalent volume of ice-cold 0.5 M EDTA, and 1-gl aliquots were spotted on cellulose polyethyleneimine plates (PEI, J.T.Baker), which were then developed in 1 M LiCl/1 M HCOOH. For other experiments (see Fig. 4 C-F) buffer E was used (buffer D + 5 mM Ca2 , Mg2+, 1 mM EDTA). The polyclonal anti-synexin antibodies used to inhibit synexin GTPase activity were purified using the ImmunoPure Ag/Ab immobilization kit (Pierce). In Vivo Labeling of Synexin by GTP in SLO-Permeabilized Chromaffin Cells. Chromaffin cells were isolated as described above (61) and cultured at 37°C in DMEM/Ham's F-12 containing 10% fetal bovine serum for 2 days in 12-well tissue culture dish at 5 x 106 cells per well. Cells were then permeabilized with streptolysin 0 (Sigma) at room tempera- ture as described (67). After permeabilization, the cells were washed once in Ca2+-free K+-glutamate buffer and then incubated in 0.5 ml of K+-glutamate buffer containing 0, 100, or 500 ,ztM CaCl2 and 20 ,uCi of either [a-32P]GTP or GTP[9y35S] for 20 min at room temperature. Immunoprecipi- Abbreviations: SLO, streptolysin 0; GTP[,yS], guanosine 5'-[-y- thioltriphosphate; PS, phosphatidylserine; GDP[,BS], guanosine 5'-[,3- thio]triphosphate. tTo whom reprint requests should be addressed at: Building 8/Room 401, National Institutes of Health, Bethesda, MD 20892. e-mail: [email protected]. 10797 The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Transcript of Membrane fusion protein synexin (annexin VII) as a Ca2+/GTP sensor in exocytotic secretion

Proc. Natl. Acad. Sci. USAVol. 93, pp. 10797-10802, October 1996Cell Biology
Membrane fusion protein synexin (annexin VII) as a Ca2+/GTPsensor in exocytotic secretionHUNG CAOHUY, MEERA SRIVASTAVA, AND HARVEY B. POLLARDtLaboratory of Cell Biology and Genetics, National Institute of Diabetes and Digestive and Kidney Diseases, National Institutes of Health, Bethesda, MD 20892
Communicated by Bernard Witkop, National Institutes of Health, Bethesda, MD, June 10, 1996 (received for review April 20, 1996)
ABSTRACT Exocytotic membrane fusion and secretionare promoted by the concerted action of GTP and Ca2 ,although the precise site(s) of action in the process are notpresently known. However, the calcium-dependent membranefusion reaction driven by synexin (annexin VII) is an in vitromodel for this process, which we have now found to be furtheractivated by GTP. The mechanism of fusion activation de-pends on the unique ability of synexin to bind and hydrolyzeGTP in a calcium-dependent manner, both in vitro and in vivoin streptolysin 0-permeabilized chromaffin cells. The re-quired [Ca2+] for GTP binding by synexin is in the range of50-200 ,AM, which is known to occur at exocytotic sites inchromaffin cells, neurons, and other cell types. Previousimmunolocalization studies place synexin at exocytotic sites inchromaffin cells, and we conclude that synexin is an atypicalG protein that may be responsible for both detecting andmediating the Ca2+/GTP signal for exocytotic membranefusion.
GTP and its nonhydrolyzable analogue guanosine 5'-[y-thio]triphosphate (GTP[,yS]) are known to promote Ca2+-dependent exocytotic secretion from many cell types by amechanism thought to involve as yet unknown proteins in theGTPase superfamily (1-4). Specific members of this super-family have been considered as mediators of these GTP effectson exocytosis, including heterotrimeric G proteins (5-9) andlow molecular weight ras-like proteins such as Rab (10-15) andARF (16). The current "fusion machine" hypothesis (17-19)envisions a core complex formed between plasma membranesyntaxin and SNAP-25 and the synaptic vesicle protein syn-aptobrevin/VAMP (20), with vesicular synaptotagmin puta-tively identified as a low-affinity calcium sensor that interactswith regulatory syntaxin 1 (21, 22). However, none of theproteins presently identified in the hypothetical fusion ma-chine have actually been shown to be activated by GTP or,indeed, even to fuse membranes (17, 23-25). Therefore, it hasbeen suggested that other GTP-binding proteins, as yet uniden-tified, might control the activity of the fusion complex (19, 25).The site of GTP action in exocytosis has been hypothesized
to be closely associated with the site of calcium action in acommon pathway (2, 5, 26, 27). The most recent consensus onthe nature of this calcium binding site is that it is involved indocking and fusion and that the affinity of the site for Ca2+may be in the range of 50-200 ,tM (28-36). From thisperspective we have considered whether the ubiquitouslydistributed membrane fusion protein synexin (annexin VII,Anx7; refs. 37-59), with an intrinsic Kd for Ca2+ of '200 ,iM(41, 42), might contribute to the hypothetical GTP/Ca2+-sensitive exocytosis site. Experiments to be described belowadvance the new concept that purified synexin is also a uniqueCa2+-conditional GTP binding protein. We show that synexinfusion activity in vitro is substantially enhanced upon bindingto GTP and deactivated by GTP hydrolysis. In addition, studies
with intact and streptolysin 0 (SLO)-permeabilized chromaf-fin cells are presented to indicate that synexin behaves in asimilar fashion in vivo.
MATERIALS AND METHODSAssays for Synexin Using Chromaffin Granules and Phos-
phatidylserine (PS) Liposomes. Recombinant human synexinwas expressed and isolated as described (60). Isolation andpurification of chromaffin granules were performed as de-scribed by Brocklehurst and Pollard (61). Large unilamellar PSliposomes ("LUV") were prepared by the swelling method(62) and fusion analyzed (55, 56). The aggregation assay forsynexin activity was performed as described by Brocklehurstand Pollard (61), and nucleotide effects on [Ca2+] werecorrected (63) and verified by calcium-selective electrodes.
Photoaffinity Labeling of Synexin by 8N3- [y-32P]GTP andLabeling Under Nondenaturing Conditions. The photoaffin-ity-labeling reaction was carried out essentially as described(64, 65), with minor modifications. 32p incorporation wasanalyzed by SDS/PAGE and quantitated by either autora-diography at -80°C or PhosphorImager counting (MolecularDynamics). Assays for [35S]GTP[,yS] binding to synexin werealso developed using the filtration method described (66).Measurement of Synexin GTPase Activity. Synexin samples
(2 ,ug) were incubated for 30 min at 30°C with 0.13 ,uM[a-32P]GTP (Amersham, 3000 Ci/mmol) in a total volume of10 ,ul of buffer D (25 mM Tris HCl, pH 7.5/250 mM NaCl/1mM DTT/10 mM ,3-glycerolphosphate) containing 1 mMEGTA (for CaCl2) or 1 mM EDTA (for MgCl2) and variousconcentrations of CaCl2 or other divalent cations. The reac-tions were stopped by adding an equivalent volume of ice-cold0.5 M EDTA, and 1-gl aliquots were spotted on cellulosepolyethyleneimine plates (PEI, J.T.Baker), which were thendeveloped in 1 M LiCl/1 M HCOOH. For other experiments(see Fig. 4 C-F) buffer E was used (buffer D + 5 mM Ca2 ,Mg2+, 1 mM EDTA). The polyclonal anti-synexin antibodiesused to inhibit synexin GTPase activity were purified using theImmunoPure Ag/Ab immobilization kit (Pierce).
In Vivo Labeling of Synexin by GTP in SLO-PermeabilizedChromaffin Cells. Chromaffin cells were isolated as describedabove (61) and cultured at 37°C in DMEM/Ham's F-12containing 10% fetal bovine serum for 2 days in 12-well tissueculture dish at 5 x 106 cells per well. Cells were thenpermeabilized with streptolysin 0 (Sigma) at room tempera-ture as described (67). After permeabilization, the cells werewashed once in Ca2+-free K+-glutamate buffer and thenincubated in 0.5 ml of K+-glutamate buffer containing 0, 100,or 500 ,ztM CaCl2 and 20 ,uCi of either [a-32P]GTP orGTP[9y35S] for 20 min at room temperature. Immunoprecipi-
Abbreviations: SLO, streptolysin 0; GTP[,yS], guanosine 5'-[-y-thioltriphosphate; PS, phosphatidylserine; GDP[,BS], guanosine 5'-[,3-thio]triphosphate.tTo whom reprint requests should be addressed at: Building 8/Room401, National Institutes of Health, Bethesda, MD 20892. e-mail:[email protected].
10797
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement" inaccordance with 18 U.S.C. §1734 solely to indicate this fact.
Proc. Natl. Acad. Sci. USA 93 (1996)
tated synexin was then assayed for bound nucleotides, asdescribed (68).
RESULTSEffect of GTP and GTP Analogues on Synexin Activity.
Synexin drives chromaffin granule aggregation in a calcium-dependent manner, and as shown in Fig. IA, both GTP andGTP[,yS] were able to increase the initial rates of this reaction.GTP[,yS] was substantially more potent that GTP, exhibitinga k/,5app of 100 ,uM compared with the ki/2,app of -500 ,uMfor GTP. By comparison, the optimal GTP[,yS] concentrationsin most permeabilized cell studies has been 50-100 ,tM, whilethe average cytosolic GTP concentrations are -200 ,tM (69).Furthermore, we found that at GTP[,yS] concentrationsgreater than 200 ,LM, this nonhydrolyzable analogue becameprogressively less potent at activating baseline synexin activity.By contrast, we did not note inhibition at higher GTP con-centrations. Finally, we found that the nonhydrolyzable ana-logue guanosine 5'-[,3-thio]dephosphate (GDP[,BS]) was en-tirely inactive. As shown in Fig. 1B, we observed virtuallyidentical relative activities of GTP[,yS], GTP, and GDP[13S]with synexin-driven fusion of pure PS liposomes. By contrast,ATP, CTP, UTP, GDP, or GMP, at 1 mM concentrations, hadno effects on synexin activity, regardless of whether the targetmembranes were chromaffin granules or PS liposomes (datanot shown).
o 170
100C*' 1 50
r 130.2
110
< 90-,0.0 0.2 0.4 0.6 0.8
[Nucleotide], mM
a
E 0.010-
a 0.008 -p0< 0.006-8)
0.004 -C.2, 0.002-U.
1.0
0.0 0.2 0.4 0.6 0.8 1.0[Synexin], gg/mL
1.2 0.0 0.2
[Nucleotide], mM
0.4 0.6 0.8 1.0 1.5 2.0[Ca2+], mM
FIG. 1. Effect of GTP and GTP analogues on synexin activity. (A)Synexin driven aggregation of chromaffin granules. Various concen-trations of GTP (0), GTP[-yS] (a), or GDP[,BS] (A) were added, andthe aggregation reactions were then initiated by the addition ofrecombinant human synexin to a final concentration of 0.5 ,ug/ml.Data are the mean ± SD (n = 2). The P values indicating significantdifferences from control are as follows: *, P < 0.004; *, P < 0.013 (forGTP[,yS]); and 4, P < 0.03; *, P < 0.003 (for GTP). (B) Synexin-drivenPS liposome fusion. The aggregation reactions were performed andanalyzed as described in A, except that PS liposomes were suspendedat a final A540 of 0.15 at 22°C. Results are the the mean + SD (n = 2or 3). The P values are shown as follows: *, P < 0.001; 4, P < 0.024 (forGTP[-yS]); and t, P < 0.005; *, P < 0.0032 (for GTP). (C) Kineticanalysis of the effect of GTP on the synexin-driven PS liposome fusionreaction. Various concentrations of synexin were used in the presenceor absence of 1 mM GTP under the same conditions as inA and B. Theinitial rates of aggregation were then measured and plotted as shown.Data are the mean ± SD (n = 2) and are fitted by theoretical curvesbased on Hill plots. The Hill coefficient for each curve is shown withinthe parentheses. (D) Kinetic analysis of the effect of GTP and [Ca2+]on the synexin-driven PS liposome fusion reaction. The GTP concen-tration was set at 1 mM and the synexin concentration was set at 0.5,tg/ml. Data are mean ± SD (n = 2).
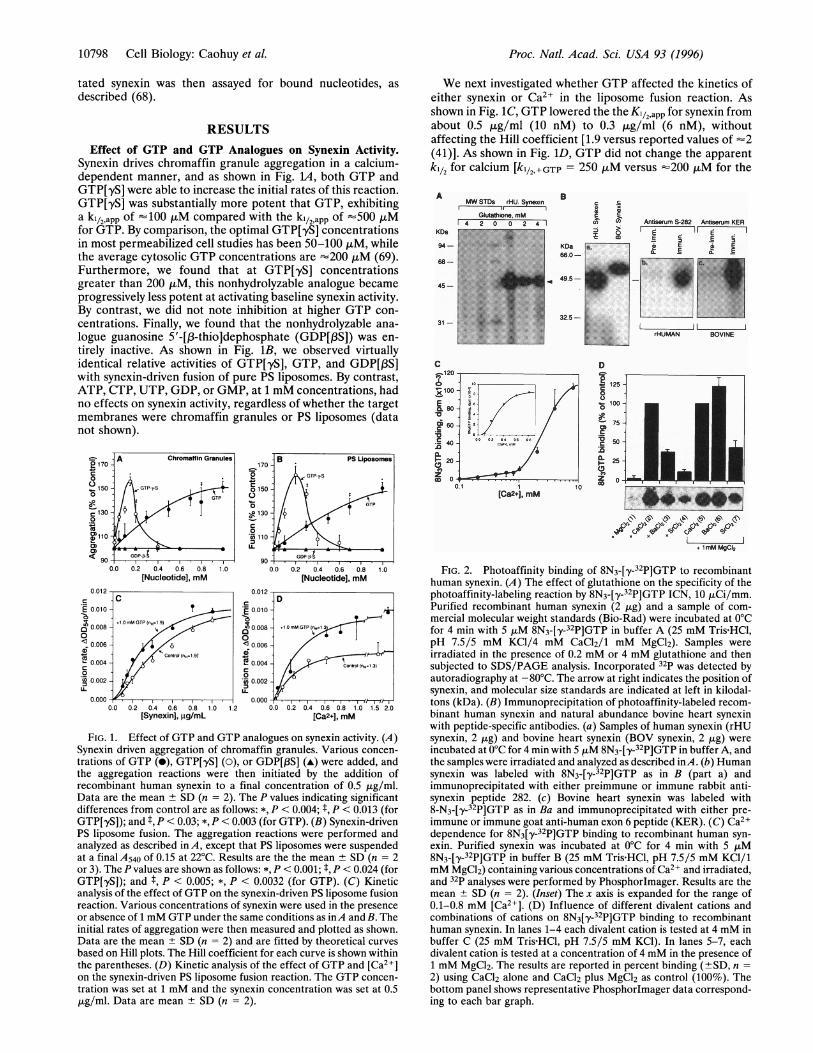
We next investigated whether GTP affected the kinetics ofeither synexin or Ca2 in the liposome fusion reaction. Asshown in Fig. 1C, GTP lowered the the K1/2,app for synexin fromabout 0.5 p,g/ml (10 nM) to 0.3 ,tg/ml (6 nM), withoutaffecting the Hill coefficient [1.9 versus reported values of -2(41)]. As shown in Fig. 1D, GTP did not change the apparentkl/2 for calcium [k1/2,+GTP = 250 ,tM versus =200 ,uM for the
AMWSTDs rHU.Synexin B
GIGlutalhone, mM
r, . . . .
KDa
94-
68-
45-
31-
C.120
-ioo -
_0-X
, 60-
:140-
20)z o0
0. 1C[Ca2+], mM
D
9 125-
p 00
75a .so
25C,Z 0
I, /X+ 1mM MgC1
FIG. 2. Photoaffinity binding of 8N3-[y-32P]GTP to recombinanthuman synexin. (A) The effect of glutathione on the specificity of thephotoaffinity-labeling reaction by 8N3-[,y-32P]GTP ICN, 10 ,LCi/mm.Purified recombinant human synexin (2 ,ug) and a sample of com-mercial molecular weight standards (Bio-Rad) were incubated at 0°Cfor 4 min with 5 ,lM 8N3-[Yy-32P]GTP in buffer A (25 mM Tris HCl,pH 7.5/5 mM KCI/4 mM CaCl2/1 mM MgCl2). Samples wereirradiated in the presence of 0.2 mM or 4 mM glutathione and thensubjected to SDS/PAGE analysis. Incorporated 32p was detected byautoradiography at -80°C. The arrow at right indicates the position ofsynexin, and molecular size standards are indicated at left in kilodal-tons (kDa). (B) Immunoprecipitation of photoaffinity-labeled recom-binant human synexin and natural abundance bovine heart synexinwith peptide-specific antibodies. (a) Samples of human synexin (rHUsynexin, 2 ,ug) and bovine heart synexin (BOV synexin, 2 ,ug) wereincubated at 0°C for 4 min with 5 FM 8N3-[,y-32P]GTP in buffer A, andthe samples were irradiated and analyzed as described inA. (b) Humansynexin was labeled with 8N3-[,y-32P]GTP as in B (part a) andimmunoprecipitated with either preimmune or immune rabbit anti-synexin peptide 282. (c) Bovine heart synexin was labeled with8-N3-[-y-32P]GTP as in Ba and immunoprecipitated with either pre-immune or immune goat anti-human exon 6 peptide (KER). (C) Ca2+dependence for 8N3[,y-32P]GTP binding to recombinant human syn-exin. Purified synexin was incubated at 0°C for 4 min with 5 utM8N3-[,y-32P]GTP in buffer B (25 mM Tris HCl, pH 7.5/5 mM KCl/1mM MgCl2) containing various concentrations of Ca2+ and irradiated,and 32P analyses were performed by Phosphorlmager. Results are themean + SD (n = 2). (Inset) The x axis is expanded for the range of0.1-0.8 mM [Ca2+]. (D) Influence of different divalent cations andcombinations of cations on 8N3[4y-32P]GTP binding to recombinanthuman synexin. In lanes 1-4 each divalent cation is tested at 4 mM inbuffer C (25 mM Tris HCl, pH 7.5/5 mM KCl). In lanes 5-7, eachdivalent cation is tested at a concentration of 4 mM in the presence of1 mM MgCl2. The results are reported in percent binding (-+SD, n =2) using CaCl2 alone and CaCl2 plus MgCl2 as control (100%). Thebottom panel shows representative Phosphorlmager data correspond-ing to each bar graph.
A Chromaffin Granules
GDP--tS
1O0 2 4 06 -eI9 C-,o
a v -i *
C
+1.0 M GTP (nl9)
,(Conlrol (n,,=1.9y
I36F
DE°°r°L'E0.010 +Z 0.008- 1O0 mM GTP(3.3
0< 0.0066-~~~~4'
0.004D - Coto/=. 3)C0iii 0.002LL :I
10798 Cell Biology: Caohuy et al.
.1

Proc. Natl. Acad. Sci. USA 93 (1996) 10799
-4 -3.5 -3
log [NucleotideJ, M
GDP-P-S (A), mM
........ .......
GTP (0), mM
c,°c~,&o0 0 o.........~
GTP-)'S (9), mMM
8N3 GTP (0), mM
oo vi
FIG. 3. Inhibition of 8N3[,y-32P]GTP binding to recombinant human synexin by guanine nucleotide analogues. Samples of purified synexin wereincubated with various concentrations of 8N3-GTP (-), GTP[,yS] (-), GTP (0), or GDP[,BS] (A) for 4 min at 0°C. The reaction mixtures were thenmixed with 5 ,uM 8N3-[Yy-32P]GTP and incubated for an additional 4 min at 0°C. Irradiation and 32P analyses were performed as described in Fig.2, and the inhibitions are reported as percent of control. The control in each case is a sample not preincubated with unlabeled nucleotides. Panelsat right shows the representative Phosphorlmager data. Data are the mean SD (n = 2-4).
control (41, 43)]. The Hill plots (results not shown) were alsovirtually identical.
Specificity of GTP Binding to Synexin. To determine un-
ambiguously whether synexin indeed bound GTP, we chose touse the photoactivatable GTP analogue 8-N3-[ y-32P]GTP (Fig.24). There was no labeling when either Ca>2 or the irradiationstep were omitted. Glutathione (2 mM) was optimal forblocking nonspecific labeling. As shown in Fig. 2Ba, bothhuman and bovine proteins could be labeled by 8-N3-[y-32P]GTP. Synexin from bovine heart muscle expresses theadditional sequence defined by cassette exon 6 and is, there-fore, -2.2 kDa larger than the nonmuscle human synexin. Asshown in Fig. 2Bb, only the immune antiserum precipitated thelabeled human synexin. We also immunoprecipitated labeledbovine heart synexin with a specific anti-exon 6 peptide goatantiserum (Fig. 2Bc).Dependence of Labeling on Calcium and Other Divalent
Cations. Since synexin labeling is so dependent upon thepresence of calcium, we performed a detailed examination ofthe calcium dependence for 8-N3-[,y-32P]GTP binding. Asshown in Fig. 2C, we found that the effect of calcium was
biphasic. The high-affinity site has a ki12,app. for [Ca2+] of 250,uM, while the lower-affinity system has a kil2app. for [Ca]2+of 2.5 mM. In both cases the Hill coefficients (nH) were -2.Different individual divalent cations were active in the follow-ing order: Ca>2 > Ba2+ > Sr2+ > Mg2+. By contrast, in thesimultaneous presence of 1 mM Mg2+, the following activityseries was obtained: Ba2+ > Ca2+ > Sr2+. Thus while bindingof GTP is crucially dependent upon the presence of Ca>,barium and strontium can also function A4-fold more effica-ciously if Mg2+ were also included in the reaction mixture.Dependence of Labeling on Guanine Nucleotides. As a
further test of the specificity of the binding of the labeled GTPanalogue, we investigated the ability of different guaninenucleotides to displace 8-N3-[ y-32P]GTP from synexin. Asshown in Fig. 3, the more hydrophobic 8N3-GTP displaced thelabel with an apparent Kd of 15 ,u M, followed by GTP[,yS] withan apparent Kd of =50 ,uM, and GTP with an apparent Kd of150 ,tM. By contrast, GDP[,BS] was relatively inactive, achiev-ing 50% displacement at -1.50 mM. At 1 mM concentrations,the following activity series was obtained: 8N3GTP > GTP[,yS]> GTP > GMP-PNP > GDP[,BS] >> ATP, CTP, and UTP(data not shown).
Synexin GTPase Activity. We next turned our attention tothe question of whether the binding of GTP was accompaniedby the ability to hydrolyze the bound nucleotide. As shown inFig. 4A, synexin exhibited readily detectable GTPase activitywhich, at the optimal temperature of 30°C, was dependentupon the presence of calcium (kl/2,app. of 50 ,uM). By contrast,a much higher concentration of Mg2> alone was needed toactivate the enzyme (k1/2app = 400 ,uM). As further summa-
rized in Fig. 4B, we found that barium and strontium alonewere much less potent than either Ca>2 or Mg2+. However, inthe presence of Mg2+, barium and strontium were againpotentiated. Finally, we found that the synexin GTPase activ-ity, in the presence of 4 mM Mg2> and 4 mM Ca2+, could beblocked by affinity purified goat anti-synexin antibody (Fig.4F). Thus, synexin GTPase activity is blocked by anti-synexinantibody regardless of whether activation is due to Mg2> orc2+.Ca.
In the presence of calcium, synexin GTPase activity is linearfor at least 30 min (Fig. 4C). The kinetics revealed saturationof the initial rate of hydrolysis as a function of GTP concen-tration with a Km of 1 ,uM, and a Hill coefficient (nH) of 1.0(see Fig. 4D). The mass curve (Fig. 4E) shows an initial lag inactivity as a function of synexin concentration, allowing us tocalculate a minimum value of the steady state kcat for Ca2+_activated synexin of 4 x 10-4 mol of GDP per mol of synexinper min. The kcat value for synexin is likely to be a minimumnumber since mastoparan activates the hydrolysis rate by atleast 10-fold (data not shown). MAS-7, an active analogue ofmastoparan, activates 8-N3-[-y-32P]GTP labeling of synexin at4°C by 15-fold and GTP['y35S] labeling at 30°C by =20-fold(data not shown). The maximal molar ratios of 8-N3-GTP/synexin range between 3 and 7%.GTP Binding and Hydrolysis by Synexin in Permeabilized
Chromaffin Cells. Permeabilized bovine chromaffin cells wereprepared with SLO in the presence of either [a-32P]GTP or
GTP[y35S], and challenged with Ca2+, and immunoprecipi-tated endogenous synexin was assayed for bound nucleotides.As shown in Fig. SA, Phosphorlmager analysis indicated thatapproximately twice as much guanine nucleotides were boundin the presence of 100 ,tM free Ca> than in - 1 nM free Ca>.As shown in Fig. SB, clear signals indicating bound GTP[935S]were seen at the two higher Ca>2 concentrations. However,only very poor signals could be obtained at -1 nM Ca2+. Thusboth types of experiments support the concept that synexin
ct
00a
IOCl
0)cvz5
Cell Biology: Caohuy et al.

Proc. Natl. Acad. Sci. USA 93 (1996)
0 10 20Time, min
30
D FIG. 4. GTPase activity of recombinant human synexin. (A) Ca2+ andMg2+ dependence of synexin GTPase activity. GTPase activity wasmeasured by incubating purified recombinant human synexin with[a-32P]GTP at 30°C in GTPase buffer containing various concentrations of
/-4 __- CaCl2 (a) or MgCl2 (A). [a-32P]GDP products were separated by chro-I-/;t / matography on a cellulose PEI plate and quantitated by Phosphorlmager
analysis. The data are corrected to show the total product in each reaction.-. b X[Me2+] represents either Ca2+ or Mg2+. (Inset) Representative
Phosphorlmager data for GTPase activity are shown. Numbers given areconcentrations of either Ca> or Mg> in each reaction. Individual data
GTP[00 2P]04 M points are mean ± SD (n = 2). (B) Effect of different divalent cations onGTP[a32P]_M synexin GTPase activity. Except as shown the final concentrations ofE divalent cations are 4 mM. Conditions are as follows: Lanes: 1, no synexin;
2, EGTA (no CaCl2 or MgCl2 added); 3, 1.8 mM Mg2+; 4, 2 mM Ca2+; 5,3 mM Ba2+; 6, 3 mM Sr2+; 7, 4 mM Ca2+/1 mM Mg2+; 8, 4 mM Ba2+/1mM Mg2+; 9, 4 mM Sr2+/1 mM Mg2+. (C) Time course for synexinGTPase activity. Purified synexin (2 ,ug in 40 ,ul) was incubated with
* / -,S.Kn [xa-32P]GTP, and aliquots (0.2 jig/10 jil) were taken every 10 min forGD91 measurement of the [a-32p]GDP product as described in A. Statistics and
0 +"tmi<°lP conditions are also as for A. (D) Determination ofKm for synexin GTPase0.2- tT''''-- activity. Various concentrations of [a-32P]GTP were incubated with 1 jig
000.2 0.4 0.6 O.B 1.0 1.2 1 4 of purified synexin for 30 min at 30°C. (Inset) Lineweaver-Burke plot of[Synexinl, AM synexin GTPase activity, from which kinetic parameters (Km and Kcat) are
derived. Statistics are as forA. (E) GTPase activity as a function of synexinconcentration. Various concentrations of synexin were incubated for 30
F o ^l min at 30°C with [a-32P]GTP. (Inset) A representative autoradiogram ofthe GTPase activity is shown as a function of synexin concentration.Statistics are as forA. (F) Inhibition of synexin GTPase activity by affinitypurified anti-synexin antibody. Synexin was preincubated with or without
_ _ 12356 affinity-purified goat polyclonal antibody ("Ram6n") for 30 min on ice inbuffer E. Immediately thereafter, 1 ,uCi of [a-33P]GTP (DuPont/NEN2000) was added and the incubation was extended for an additional 30 minat 30°C. Data are the mean ± SD (n = 3). (Inset) RepresentativePhosphorlmager data are given for GTPase activities. Numbers indicate
(4. t+ conditions as follows: Lanes: 1, blank; 2, synexin control (1 jig); 3, synexin'^ <i>vKx* ',i6 + 10 jig of antibody; 4, synexin + 20 jig of antibody; 5, 10 jig of antibody
OP V011 alone; 6, 20 jig of antibody alone.
inside SLO-permeabilized chromaffin cells can be furtheractivated to bind and hydrolyze GTP in the presence ofelevated free [Ca21].
DISCUSSIONThese data show that Ca2+-dependent synexin-driven mem-brane aggregation and fusion activity is substantially poten-tiated by GTP and by the nonhydrolyzable analogueGTP[,yS], but not by GDP[,3S]. The mechanism of GTP
A.
Pre-Imm. Antiserum S-282
GDP [a-32p]
GTP [a-32P]
A.
_iI
ORIGIN -
[Ca2+],jM 100 0.001 100 500
potentiation involves Ca2+-dependent binding of GTP to thesynexin molecule, and this correlation is manifest qualita-tively by close parallels between the potencies of differenttypes of nucleotides and divalent cations in both processes.The sensitivity of synexin to these other divalent cations is ofparticular importance in the field of secretion since it iswidely found that Ba>2 can sometimes be a more effectivesecretagogue than Ca>2 and that Sr2+ also has activity (70,71). Synexin is thus, to our knowledge, the first example ofa GTP binding protein for which nucleotide binding can be
B.
Antiserum S-282.--.-.=I
GTP--S [35S]
ORIGIN0.001 100 500
FIG. 5. In vivo labeling of synexin by GTP in SLO-permeabilized chromaffin cells. Thin layer chromatogram of the nucleotides eluted fromimmunoprecipitates of synexin from SLO-permeabilized chromaffin cells that had been incubated in different concentrations of Ca2+ with either[a-32P]GTP (A) or GTP[9?5S] (B). After a 20-min incubation, cells were lysed and the lysate immunoprecipitated with either preimmune rabbitserum or immune serum S-282 for 4 h at 4°C. Migration positions of nucleotide standards are shown at the left or right, respectively.
CO 0.40 -
x
E 0.30 -
CL
Io 0.200
- 0.20 -0-
C)1 .
10800 Cell Biology: Caohuy et aL

Proc. Natl. Acad. Sci. USA 93 (1996) 10801
REGULATION OF SYNEXIN-DRIVEN MEMBRANEFUSION BY Ca2+ AND GTP
[S -Mg2+-GTrP
FIG. 6. Model for synexin-driven membrane fusion regulated byCa2+ and GTP. Under a resting low-calcium condition, synexin existsin an "off' state (SO-Mg2+GDP) that is formed by a process ofconstitutive Mg2+-dependent hydrolysis of GTP. Upon elevation ofCa2 , synexin binds GTP and assumes a stable "on" state that candrive membrane fusion (S**-Ca2+/Mg2+GTP). This transition can beactivated by mastoparan. Upon hydrolysis of the GTP to GDP, atransient (S*-Ca2+/Mg2+GDP) complex is formed. Then, with thereduction in the free Ca2+ concentration, the "off' state (SO-Mg2+GDP) complex is reformed, and the cycle can start once again.AlF3 can activate the GDP-bound form of synexin, as long as elevatedCa2+ is also present. The different number of * symbols representdifferent activated states and have no quantitative significance. Thesubscripted numbers on the bracketed complexes, [ I1 and [ 12, refer tothe presumed physiological order of the reaction. Broken arrow isnonphysiological activation by A1F3.
potently activated by Ca2+, Ba2+, or Sr2+. Binding of GTPinduced by calcium leads to formation of a synexin-GTPcomplex that is sufficiently stable and long-lived to be readilydetectable by the photolabeling reaction. This calcium-dependent complex is thus operationally different from theGTP complex with synexin formed in the presence of Mg2+,for which the photolabeling reaction is much less efficient.These in vitro data are likely to be relevant to membranefusion processes occurring in vivo, since calcium also acti-vates synexin to bind and hydrolyze GTP in SLO-permeabilized chromaffin cells. The average cytosolic GTPconcentration is '200 ,uM (69), which we interpret toindicate that both GTP binding and GTP hydrolysis bysynexin can be constitutively saturated processes in cells.We have therefore envisioned the mechanism of this
process explicitly in terms of a G-protein-like molecularswitch (72-75) in which calcium-conditional activation leadssynexin into a GTP-bound fusogenic "on" state. Subse-quently, inactivation to the "off" state occurs by GTPhydrolysis. This concept is summarized in Fig. 6. In the "off'state (S), synexin slowly hydrolyzes GTP to GDP by a
Mg2+-dependent reaction that, for example, occurs in thelow [Ca2+] condition in cells in Fig. 5. Upon elevation of thefree [Ca2+] into the range of 50-200 ,AM, synexin convertsto the "on" state (S*). Alternatively, synexin binds Ca2+ andGTP to form the S**-Ca2+/Mg2+-GTP complex. This pro-cess can be further potentiated by mastoparan, which hasbeen reported to activate secretion from SLO-permeabilizedchromaffin cells (76). Fusion then ensues until GTP ishydrolyzed and the calcium is reduced, leaving synexin againin the "off" state. This model is generally characteristic ofthe molecular switch mechanism identified for different
members of the GTPase superfamily in which a conditionalactivator induces an active GTP-bound state and for whichinactivation occurs by GTP hydrolysis. In the case of synexin,the conditional activator seems to be calcium. The model inFig. 6 also describes activation of calcium-dependent mem-brane fusion activity by A1F3, which we have observed withliposomes (data not shown)., A1F3 also activates heterotri-meric G proteins by forming a GTP-like structure withbound GDP, and a similar mechanism may be obtained withrecombinant synexin isolated from Escherichia coli in aGDP-bound state (data not shown).
In conclusion, these studies suggest that the function ofsynexin in cells may be to transduce the calcium signal forexocytosis by binding GTP, polymerizing, and directly medi-ating a membrane fusion process. It is our opinion that thesedata are entirely compatible with activities of "fusion ma-chines" or "fusion scaffolds," which are widely thought to beassociated with initial membrane docking events, but whosecurrently defined components lack "fusion" activity.
We thank Drs. Eduardo Rojas, Nelson Arispe, Eleazar B. Cohen,Allan Minton, Shiv Srivastava, Constantine Londos, Yossi Raviv, andJan Wolfe for helpful discussions, preliminary reading of the manu-script, and the sharing of preliminary data.
1. Neher, E. (1988) J. Physiol. (London) 395, 193-214.2. Monck, J. R. & Fernandez, J. M. (1994) Neuron 12, 707-716.3. Ahnert-Hilger, G., Wegenhorst, U., Stecher, B., Spicher, K.,
Rosenthal, W. & Gratzl, M. (1992) Biochem. J. 284, 321-326.4. Aridor, M., Rajmilevich, G., Beaven, M. A. & Sagi-Eisenberg, R.
(1993) Science 262, 1569-1572.5. Gomperts, B. D., Cockroft, S., Howell, T. W., Niisse, 0. &
Tatham, P. E. R. (1987) Biosci. Report 7, 369-381.6. Gomperts, B. D. (1990) Annu. Rev. Physiol. 52, 591-606.7. Brocklehurst, K. W. & Pollard, H. B. (1988) FEBS Lett. 234,
439-445.8. Cefia, V., Brocklehurst, K. W., Pollard, H. B. & Rojas, E. (1991)
J. Membr. Biol. 122, 23-31.9. Ohara-Imaizumi, M., Kameyama, K., Kawae, N., Takeda, K.,
Muramatsu, S. & Kumakura, K. (1992) J. Neurochem. 58, 2275-2284.
10. Oberhauser, A. F., Monck, J. R., Balch, W. E. & Fernandez, J.M.(1992) Nature (London) 360, 270-273.
11. Lledo, P.-M. Vernier, P., Vincent, J.-D., Mason, W. T. & Zorec,R. (1993) Nature (London) 364, 540-544.
12. Johannes, L., Liedo, P. M., Roa, M., Vincent, J.-D., Henry, J.-P.& Darchen, F. (1994) EMBO J. 13, 2029-2037.
13. Darchen, F., Zahraoui, A., Hammel, F., Monteils, M.-P., Tavi-tian, A. & Scherman, D. (1990) Proc. Natl. Acad. Sci. USA 87,5692-5696.
14. Fischer von Mollard, G., Mignery, G. A., Baumert, M., Perin,M. S., Hanson, T. J., Burger, P. M., Jahn, R. & Sudhof, T. C.(1990) Proc. Natl. Acad. Sci. USA 87, 1988-1992.
15. Geppert, M., Bolshakov, V. Y., Siegelbaum, S. A., Takel, K., DeCamilli, P., Hammer, R. E. & Sudhof, T. C. (1994) Nature(London) 369, 493-497.
16. Morgan, A. & Burgoyne, R. D. (1993) FEBS Lett. 329, 121-124.17. Rothman, J. E. & Orci, L. (1992) Nature (London) 355,409-415.18. Bennett, M. K. & Scheller, R. H. (1993) Proc. Natl. Acad. Sci.
USA 90, 2559-2563.19. Ferro-Novick, S. & Jahn, R. (1994) Nature (London) 370, 191-
193.20. Sudhof, T. C. (1995) Nature (London) 375, 645-653.21. Chapman, E. R., Hansen, P. I. & Jahn, R. (1995) J. Biol. Chem.
270, 23667-23671.22. Morgan, A. & Burgoyne, R. D. (1995) EMBO J. 14, 232-239.23. White, J. (1992) Science 258, 917-923.24. Morgan, A. (1995) Essays Biochem. 30, 77-95.25. O'Connor, V., Augustine, G. J. & Betz, H. (1994) Cell 76,
785-787.26. Howell, T. W., Cockroft, S. & Gomperts, B. D. (1987)J. Cell Biol.
105, 191-197.27. Okano, K., Monck, J. R. & Fernandez, J. M. (1993) Neuron 11,
165-172.
Cell Biology: Caohuy et al.

Proc. Natl. Acad. Sci. USA 93 (1996)
28. Augustine, G. J. & Neher, E. (1992) J. Physiol. (London) 450,247-271.
29. Neher, E. & Augustine, G. J. (1992) J. Physiol. (London) 450,273-301.
30. Neher, E. & Zucker, R. S. (1993) Neuron 10, 21-30.31. Heinemann, C., Chow, R. H., Neher, E. & Zucker, R. S. (1994)
Biophys. J. 67, 2546-2557.32. von Ruden, L. & Neher, E. (1993) Science 262, 1061-1065.33. Rosenboom, H. & Lindau, M. (1994) Proc. Natl. Acad. Sci. USA
91, 5267-5271.34. Heldelberger, R., Heinemann, C., Neher, E. & Mathews, G.
(1994) Nature (London) 371, 513-515.35. Thomas. P., Wong, J. G., Lee, A. K. & Almers, W. (1993) Neuron
11, 93-104.36. Bittner, M. A. & Holz, R. W. (1992) J. Biol. Chem. 267, 16219-
16225.37. Pollard, H. B., Rojas, E., Pastor, R. W., Rojas, E. M., Guy, H. R.
& Burns, A. L. (1991) Ann. N.Y Acad. Sci. 635, 328-351.38. Pollard, H. B., Burns, A. L. & Rojas, E. (1990) J. Membr. Biol.
117, 101-112.39. Shirvan, A., Srivastava, M., Wang, M. A., Cultraro, C., Ma-
gendzo, K., McBride, 0. W., Pollard, H. B. & Burns, A. L. (1994)Biochemistry 33, 6888-6901.
40. Zhang-Keck, Z.-Y., Burns, A. L. & Pollard, H. B. (1993) Bio-chem. J. 289, 735-741.
41. Creutz, C. E., Pazoles, C. J. & Pollard, H. B. (1978)J. Biol. Chem.253, 2858-2866.
42. Raynal, P. & Pollard, H. B. (1994) Biochim. Biophys. ActaBiomembranes 1197, 63-93.
43. Creutz, E. C., Pazoles, C. J. & Pollard, H. B. (1979)J. Biol. Chem.254, 553-558.
44. Kuijpers, G. A. J., Lee, G. & Pollard, H. B. (1992) Cell Tissue Res.269, 323-330.
45. Cardenas, A. M., Kuijpers, G.A. J. & Pollard, H. B. (1994)Biochim. Biophys. Acta 21234, 255-260.
46. Gierke, V. (1991) J. Biol. Chem. 266, 1697-1700.47. Doring, V., Schleicher, M. & Noegel, A. A. (1991) J. Biol. Chem.
266, 17509-17515.48. Srivastava, M., Zhang-Keck, Z.-Y., Caohuy, H., McPhie, P. &
Pollard, H. B. (1996) Biochem. J. 316, 729-735.49. Zhang-Keck, Z.-Y., Srivastava, M.,. Kozak, C. A., Caohuy, H.,
Shirvan, A., Burns, A. L. & Pollard, H. B. (1994) Biochem. J. 301,835-845.
50. Burns, A. Lee, Magendzo, K., Shirvan, A., Srivastava, M., Rojas,E., Alijani, M. & Pollard, H. B. (1989) Proc. Natl. Acad. Sci. USA86, 3798-3802.
51. Creutz, C. E. (1981) J. Cell Biol. 91, 247-256.
52. Stutzin, A. (1986) FEBS Lett. 197, 274-280.53. Stutzin, A., Cabantchik, I., Lelkes, P. I. & Pollard, H. B. (1987)
Biochim. Biophys. Acta 905, 205-212.54. Nir, S., Stutzin, A. & Pollard, H. B. (1987) Biochim. Biophys. Acta.
903, 309-318.55. Hong, K., Duzgunes, N. & Papahadjopulos, D. (1981) J. Biol.
Chem. 256, 3641-3644.56. Hong, K., Duzgunes, N., 'Ekerdt, R. & Papahadjopoulos, D.
(1982) Proc. Natl. Acad. Sci. USA 79, 4642-4644.57. Pollard, H. B., Rojas, E., Pastor, R. W., Rojas, E. M., Guy, H. R.
& Burns, A. L. (1991) Ann. N. Y Acad. Sci. 635, 328-351.58. Rojas, E. & Pollard, H. B. (1987) FEBS Lett. 217, 25-31.59. Pollard, H. B. & Rojas, E. (1988) Proc. Natl. Acad. Sci. USA 85,
2974-2978.60. Burns, A. L., Magendzo, K., Srivastava, M., Rojas, E., Parra, C.,
de la Fuente, M., Cultraro, C., Shirvan, A., Vogel, T., Heldman,J., Caohuy, H., Tombaccini, D. & Pollard, H. B. (1990) Biochem.Soc. Trans. 18, 1118-1131.
61. Brocklehurst, K. W. & Pollard, H. B. (1990) in Peptide Hormones:A PracticalApproach, eds. Siddle, K. & Hutton, J. (IRL, Oxford),pp. 233-255.
62. Reeves, J. & Dowben, R. (1968) Cell. Physiol. 78, 49-60.63. Fabiato, A. & Fabiato, F. (1979) J. Physiol. (Paris) 75, 463-465.64. Pak, C., Krumbiegel, M., Blumenthal, R. & Raviv, Y. (1994)
J. Biol. Chem. 269, 14614-14619.65. Thomas, R. & Pfeuffer, T. (1991) Methods Enzymol. 195, 280-
287.66. Ferguson, K. M., Higashima, T., Smigel, M. D. & Gilman, A. G.
(1986) J. Biol. Chem. 261, 7393-7399.67. Ahnert-Hilger, G. (1994) Cell Biology: A Laboratory Manual
(Academic, London), Vol. 3, pp. 77-82.68. Downward, J., Graves, J. D., Warne, P. H., Rayter, S. & Cantrell,
D. A. (1990) Nature (London) 346, 719-723.69. Peveri, P., Heyworth, P. G. & Curnutte, J. T. (1992) Proc. Natl.
Acad. Sci. USA 89, 2494-2498.70. Forsberg, E. J. & Pollard, H. B. (1988) Neuroscience 27, 711-715.71. Heldman, E., Levine, M., Raveh, L. & Pollard, H. B. (1989)
J. Biol. Chem. 264, 7914-7920.72. Gilman, A. G. (1987) Annu. Rev. Biochem. 56, 615-649.73. Bourne, H. R., Sanders, D. A. & McCormick, F. (1990) Nature
(London) 348, 125-132.74. Bourne, H. R., Sanders, D. A. & McCormick, F. (1991) Nature
(London) 349, 117-127.75. Ross, E. M. (1988) Concepts Biochem. 65, 937-942.76. Vitale, N., Mukai, H., Rouot, B., Thierse, D., Aunis, D. &
France-Bader, M. F. (1995) J. Biol. Chem. 268, 14715-14723.
10802 Cell Biology: Caohuy et al.




















