
Computational Design of Peptides That Target Transmembrane Helices

doi:10.1016/j.jmb.2007.02.094 J. Mol. Biol. (2007) 368, 1367–1378
The mRNA-binding Site of Annexin A2 Resides inHelices C–D of its Domain IV
Ingvild Aukrust1, Hanne Hollås1, Elin Strand1, Lasse Evensen1
Gilles Travé2, Torgeir Flatmark1 and Anni Vedeler1⁎
1Department of Biomedicine,University of Bergen, Jonas LiesVei 91, N-5009 Bergen, Norway2Équipe Oncoprotéine,UMR-7175-LC1 du CNRS,École Supérieure deBiotechnologie de Strasbourg,67400 Illkirch, FranceAbbreviations used: AnxA2, anneanxA2, annexin A2 cDNA or mRNAIVof annexin A2; MBP, maltose bindresonance units; SA, streptavidin; SPresonance; ss, single-stranded; TEV,Tm, transition temperature; UTR, unwild-type.E-mail address of the correspondi
0022-2836/$ - see front matter © 2007 E
Annexin A2 (AnxA2) is a Ca2+-binding and phospholipid-binding proteininvolved in different intracellular processes including exocytosis, endocy-tosis and membrane-cytoskeleton movements. We have previously identi-fied AnxA2 as an mRNA-binding protein present in cytoskeleton-boundpolysomes, that binds to a specific ∼100 nucleotide region in the 3′-untranslated region of c-myc and its cognate mRNA. In the present study,we show by UV cross-linking assays and surface plasmon resonanceanalyses that the mRNA-binding site of AnxA2 resides in its domain IV.Furthermore, the interaction of full-length AnxA2 with the 3′-untranslatedregion of anxA2 mRNA is Ca2+-dependent. By contrast, the interaction isCa2+-independent for the isolated domain IVof AnxA2, suggesting that themRNA-binding site is masked in Apo-AnxA2 and gains exposure through aCa2+-induced conformational change of AnxA2 generating a favourablemRNA-binding site. The AnxA2-mRNA interaction is specific and involveshelices C and D in domain IVof AnxA2, since point mutagenesis of severalcharged and polar exposed residues of these helices in the full-lengthprotein strongly reduce RNA binding. The interaction appears to besequential involving an initial phase of recognition dominated byelectrostatic interactions, most likely between lysine residues and thephosphate backbone of RNA, followed by a second phase contributing tothe specificity of the interaction.
© 2007 Elsevier Ltd. All rights reserved.
*Corresponding author
Keywords: annexin A2; domain IV; SPR; mRNA; 3′-UTRIntroduction
Annexin A2 (AnxA2) is a member of the multi-gene family of annexin proteins, which are char-acterised by binding to phospholipids in thepresence of Ca2+. On the basis of this property, theannexins have been assigned roles in membraneorganisation, membrane-cytoskeleton interactions,exocytosis, endocytosis and in the regulation of ionchannel activity.1–3 Structurally, the annexins have a
xin A2 protein;; AnxA2-DIV, domaining protein; RU,R, surface plasmontobacco etch virus;translated region; wt,
ng author:
lsevier Ltd. All rights reserve
variable N-terminal tail and a highly conserved C-terminal core structure. The core contains fourhomologous domains (repeats) (I–IV) (except forAnxA6, which has eight repeats), each consisting offive α-helices (helices A through E) generating aright-handed super-helix.4 Helices A and B, andhelices D and E form two parallel helix-loop-helixfolds, whereas helix C connects helices B and D withits axis perpendicular to the other four helices.Generally, the AB-loop and the DE-loop form theCa2+-binding sites in the annexins (type II and typeIII), which differ from the well-characterised EF-hand motif.5–7 Interestingly, the annexins changeconformation upon Ca2+-binding,8–10 and molecularmotion analyses based on available structuralcoordinates (1W7B and 1XJL) have revealed thelargest changes in the backbone conformation atGly100, Thr133, Gly157 and Gly163 in AnxA2†. The
†http://www.molmovdb.org/cgi-bin/phi-angles.cgi?mid=D294633-10718
d.
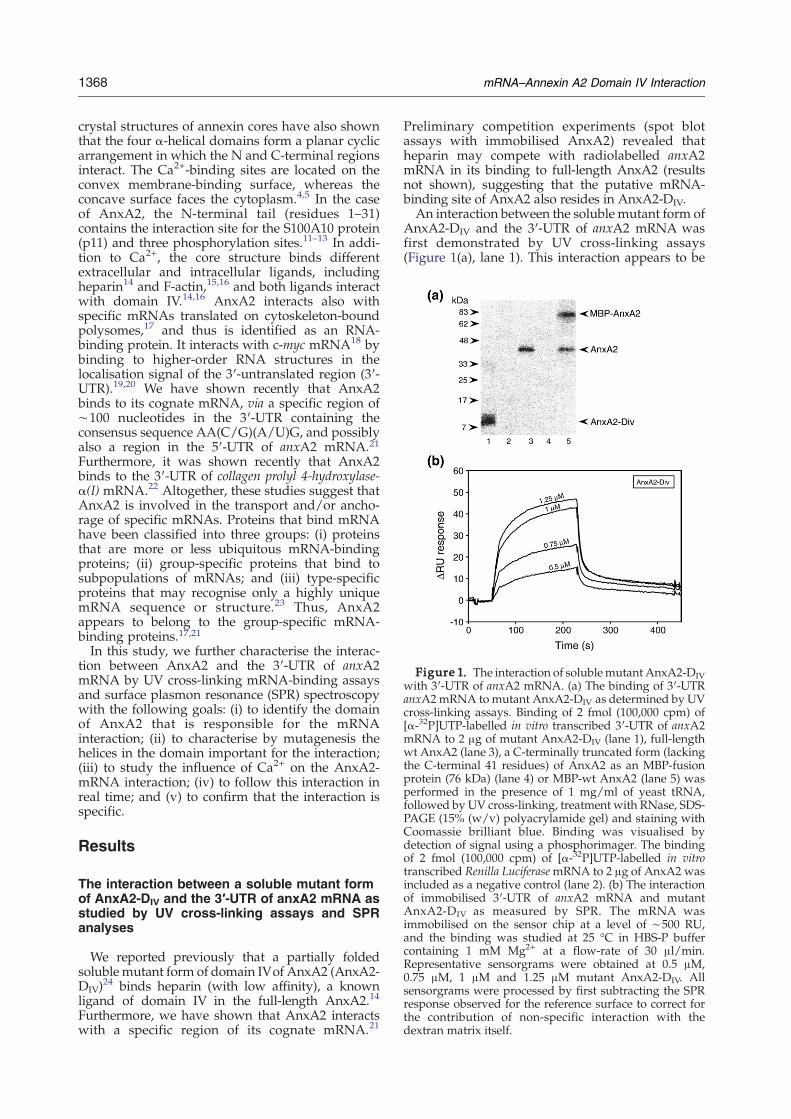
Figure 1. The interaction of solublemutant AnxA2-DIVwith 3′-UTR of anxA2 mRNA. (a) The binding of 3′-UTRanxA2mRNA tomutant AnxA2-DIV as determined by UVcross-linking assays. Binding of 2 fmol (100,000 cpm) of[α-32P]UTP-labelled in vitro transcribed 3′-UTR of anxA2mRNA to 2 μg of mutant AnxA2-DIV (lane 1), full-lengthwt AnxA2 (lane 3), a C-terminally truncated form (lackingthe C-terminal 41 residues) of AnxA2 as an MBP-fusionprotein (76 kDa) (lane 4) or MBP-wt AnxA2 (lane 5) wasperformed in the presence of 1 mg/ml of yeast tRNA,followed by UV cross-linking, treatment with RNase, SDS-PAGE (15% (w/v) polyacrylamide gel) and staining withCoomassie brilliant blue. Binding was visualised bydetection of signal using a phosphorimager. The bindingof 2 fmol (100,000 cpm) of [α-32P]UTP-labelled in vitrotranscribed Renilla LuciferasemRNA to 2 μg of AnxA2 wasincluded as a negative control (lane 2). (b) The interactionof immobilised 3′-UTR of anxA2 mRNA and mutantAnxA2-DIV as measured by SPR. The mRNA wasimmobilised on the sensor chip at a level of ∼500 RU,and the binding was studied at 25 °C in HBS-P buffercontaining 1 mM Mg2+ at a flow-rate of 30 μl/min.Representative sensorgrams were obtained at 0.5 μM,0.75 μM, 1 μM and 1.25 μM mutant AnxA2-DIV. Allsensorgrams were processed by first subtracting the SPRresponse observed for the reference surface to correct forthe contribution of non-specific interaction with thedextran matrix itself.
1368 mRNA–Annexin A2 Domain IV Interaction
crystal structures of annexin cores have also shownthat the four α-helical domains form a planar cyclicarrangement in which the N and C-terminal regionsinteract. The Ca2+-binding sites are located on theconvex membrane-binding surface, whereas theconcave surface faces the cytoplasm.4,5 In the caseof AnxA2, the N-terminal tail (residues 1–31)contains the interaction site for the S100A10 protein(p11) and three phosphorylation sites.11–13 In addi-tion to Ca2+, the core structure binds differentextracellular and intracellular ligands, includingheparin14 and F-actin,15,16 and both ligands interactwith domain IV.14,16 AnxA2 interacts also withspecific mRNAs translated on cytoskeleton-boundpolysomes,17 and thus is identified as an RNA-binding protein. It interacts with c-myc mRNA18 bybinding to higher-order RNA structures in thelocalisation signal of the 3′-untranslated region (3′-UTR).19,20 We have shown recently that AnxA2binds to its cognate mRNA, via a specific region of∼100 nucleotides in the 3′-UTR containing theconsensus sequence AA(C/G)(A/U)G, and possiblyalso a region in the 5′-UTR of anxA2 mRNA.21Furthermore, it was shown recently that AnxA2binds to the 3′-UTR of collagen prolyl 4-hydroxylase-α(I) mRNA.22 Altogether, these studies suggest thatAnxA2 is involved in the transport and/or ancho-rage of specific mRNAs. Proteins that bind mRNAhave been classified into three groups: (i) proteinsthat are more or less ubiquitous mRNA-bindingproteins; (ii) group-specific proteins that bind tosubpopulations of mRNAs; and (iii) type-specificproteins that may recognise only a highly uniquemRNA sequence or structure.23 Thus, AnxA2appears to belong to the group-specific mRNA-binding proteins.17,21In this study, we further characterise the interac-
tion between AnxA2 and the 3′-UTR of anxA2mRNA by UV cross-linking mRNA-binding assaysand surface plasmon resonance (SPR) spectroscopywith the following goals: (i) to identify the domainof AnxA2 that is responsible for the mRNAinteraction; (ii) to characterise by mutagenesis thehelices in the domain important for the interaction;(iii) to study the influence of Ca2+ on the AnxA2-mRNA interaction; (iv) to follow this interaction inreal time; and (v) to confirm that the interaction isspecific.
Results
The interaction between a soluble mutant formof AnxA2-DIV and the 3′-UTR of anxA2 mRNA asstudied by UV cross-linking assays and SPRanalyses
We reported previously that a partially foldedsolublemutant form of domain IVof AnxA2 (AnxA2-DIV)
24 binds heparin (with low affinity), a knownligand of domain IV in the full-length AnxA2.14
Furthermore, we have shown that AnxA2 interactswith a specific region of its cognate mRNA.21
Preliminary competition experiments (spot blotassays with immobilised AnxA2) revealed thatheparin may compete with radiolabelled anxA2mRNA in its binding to full-length AnxA2 (resultsnot shown), suggesting that the putative mRNA-binding site of AnxA2 also resides in AnxA2-DIV.An interaction between the soluble mutant form of
AnxA2-DIV and the 3′-UTR of anxA2 mRNA wasfirst demonstrated by UV cross-linking assays(Figure 1(a), lane 1). This interaction appears to be

1369mRNA–Annexin A2 Domain IV Interaction
specific as Renilla Luciferase mRNA does not interactwith AnxA2 (Figure 1(a), lane 2). In addition, wehave shown recently that AnxA2 does not interactwith the coding region or the 3′-end of the 3′-UTR ofits cognate mRNA.21 Furthermore, no interactionbetween AnxA2 and β2-microglobulin mRNA wasdetected in vivo.21 Full-length AnxA2 was includedas a positive control (Figure 1(a), lane 3), while thenegative control protein, a maltose-binding protein(MBP)-AnxA2ΔC41 lacking the putative mRNA-binding site (the three C-terminal helices), does notinteract with the mRNA (Figure 1(a), lane 4). This isnot due to MBP blocking the interaction, since MBP-wtAnxA2 interactswith the 3′-UTR of anxA2mRNA(Figure 1(a), lane 5). The SPR analyses revealed thatmutant AnxA2-DIV interacts with immobilised 3′-UTR of anxA2 mRNA in the absence of Ca2+ (Figure1(b)). The interaction appears to be complex, and thesensorgrams could not be fit to any of the commonbinding models in the BIAevaluation programme.Thus, noKd value could be calculated,most probablydue to the complexity of the interaction.25 Moreover,AnxA2-DIV appeared to self-aggregate at concentra-
tions above 1.25 μM, as suggested by an unexpectedhigh SPR response (results not shown).
The effect of Ca2+ on the interaction of wild-typeAnxA2 with the 3′-UTR of anxA2 mRNA
Ca2+-binding to annexin proteins is generallydescribed as low-affinity binding in the absence ofphospholipids, and for AnxA2 the overall dissocia-tion constant for its multiple Ca2+-binding sites isreported to be ∼0.5 mM.6 Since the binding of Ca2+
results in a conformational change,5,8,9 it was ofinterest to study the effect of Ca2+ on the binding ofwt AnxA2 to the immobilised 3′-UTR of anxA2mRNA as measured by SPR. As seen in Figure 2(a),the binding of AnxA2 was indeed Ca2+-dependent.In contrast to the binding of the soluble mutantAnxA2-DIV to the same transcript (Figure 1(b)), nointeraction was observed at concentrations of Ca2+
<20 μM. Biphasic sensorgrams were observed at50 μM and 70 μM Ca2+. However, at 1 mM Ca2+
(results not shown), the steady-state SPR responsewas much higher than the ΔRUmax response (∼190
Figure 2. The effect of Ca2+ on(a) the interaction of wt AnxA2withimmobilised 3′-UTR of anxA2mRNA as measured by SPR and(b) the solvent accessibility of resi-dues in helix C through helix D ofAnxA2-DIV. (a) The mRNA wasimmobilised on the sensor chip ata level of ∼480 RU, and the bindingwas studied at 25 °C inHBS-P buffercontaining 1 mM Mg2+ at a flow-rate of 30 μl/min. The AnxA2concentration was 2 μM, while theconcentration of Ca2+ was variedfrom 0–70 μM. (b) The static solventaccessibility of residues in helices Cthrough D of AnxA2-DIV in theabsence (□) and in the presence(▪) of Ca2+ was calculated usingCUPSAT28 based on the structuralcoordinates of human AnxA2 ob-tained in the absence (1W7B) andpresence (1XJL) of Ca2+.5 Note thatArg309 in human AnxA2 is equiva-lent to Lys309 in bovine AnxA2. Thecylinders below denote residues inα-helices C and D. The Gly (Gly312)that is indicated by an asterisk (*),changes the backbone dihedral tor-sion angles in the presence of Ca2+
(see the text).

1370 mRNA–Annexin A2 Domain IV Interaction
RU) expected from the relative molecular masses ofwt AnxA2 and the 3′-UTR of anxA2 mRNA, and theimmobilised level of the transcript. This largeresponse is most likely explained by an aggregationof AnxA2 at high concentrations of Ca2+, a char-acteristic property of several annexin proteins.26,27
In a recent study,24 we have analysed the struc-tural role of the five α-helices of domain IV. Thehelices B and E were shown to be largely involvedin interfacial hydrophobic contacts with the otherdomains of AnxA2. By contrast, helix D as well asmost of helix C (including the CD-loop), does notinteract with the other domains of the protein, andboth helices are largely solvent-exposed. Using theCUPSAT algorithm,28 the static solvent accessibilityof the residues in these helices was calculated(Figure 2(b)), based on the structural coordinates ofAnxA2 in the absence (1W7B) and presence of Ca2+
(1XJL).5 It is seen that all the residues that weremutated, are solvent-exposed in the absence and inthe presence of Ca2+. Importantly, Ca2+ induceschanges of the backbone dihedral torsion angles, i.e.Δ(φ+ψ)=15.6°, at Gly312 (indicated by an asterisk(*) in Figure 2(b)) in the CD loop and thus also of theorientation of helix D relative to helix C (results notshown).
Generation, expression and purification ofAnxA2 mutated in the putative mRNA-bindingsite
The helices C andDof domain IV contain twowell-exposed regions, 308KKK310 and 316YYYIQQD322,
Figure 3. Analysis of surface-exposed basic, aromatic awhich may participate in RNA recognition. (a) Ribbon staromatic and polar residues in helices C, D and in the CD-MOLMOL.52 (b) Substitutions performed in domain IV of AnAnxA2-mutC, AnxA2-mutD and AnxA2-mutCD.
respectively, which togetherwith Gly312 and Lys313in the CD-loop (Figures 2(b) and 3(a)), providepotential sites for interaction with a bulky RNAtarget. To further analyse this possibility, threemutant forms of AnxA2 were prepared (Figure3(b)). AnxA2-mutC contains three multiple Sersubstitutions at Lys308, Lys309 and Lys310 in helixC, while AnxA2-mutD contains three multiple Sersubstitutions of Lys313 in the CD loop and Tyr317and Gln321 in helix D. Finally, AnxA2-mutCDcontains the combined mutations of both AnxA2-mutC and AnxA2-mutD. In all cases Ser was used asthe substituting residue because multiple Ala muta-tions may generate local hydrophobic patches.29
The wt AnxA2 and its mutant forms (Figure 3(b))were cloned into the plasmid pETM-41, allowingtheir expression in Escherichia coli in high yields(Figure 4(a)) as fusion proteins to the C terminus ofHis6-MBP. All proteins were purified on Ni2+-affinity resin and subjected to ultracentrifugation(200,000gav for 16 h) to identify possible tendenciesto aggregate. All fusion proteins were recovered athigh yield as soluble fusion proteins (Figure 4(b),lanes 2, 4, 6 and 8). To ensure that all the mutantswere in a true monodisperse soluble form, and thatthis was not a solubilisation artefact due to thefusion to the soluble MBP partner,30 His6-MBP-wtAnxA2 and the corresponding mutants were sub-jected to ultracentrifugation after cleavage withtobacco etch virus (TEV) protease (Figure 4(c)). Thewt AnxA2 and all the mutants were recoveredmainly in the supernatant (Figure 4(c), lanes 2, 4, 6and 8), indicating that all the passenger proteins
nd polar residues in domain IV of full-length AnxA2,ructure of AnxA2 showing the surface-exposed basic,loop in domain IV. The 3-D image was generated usingxA2 to create mutant forms of full-length AnxA2 termed
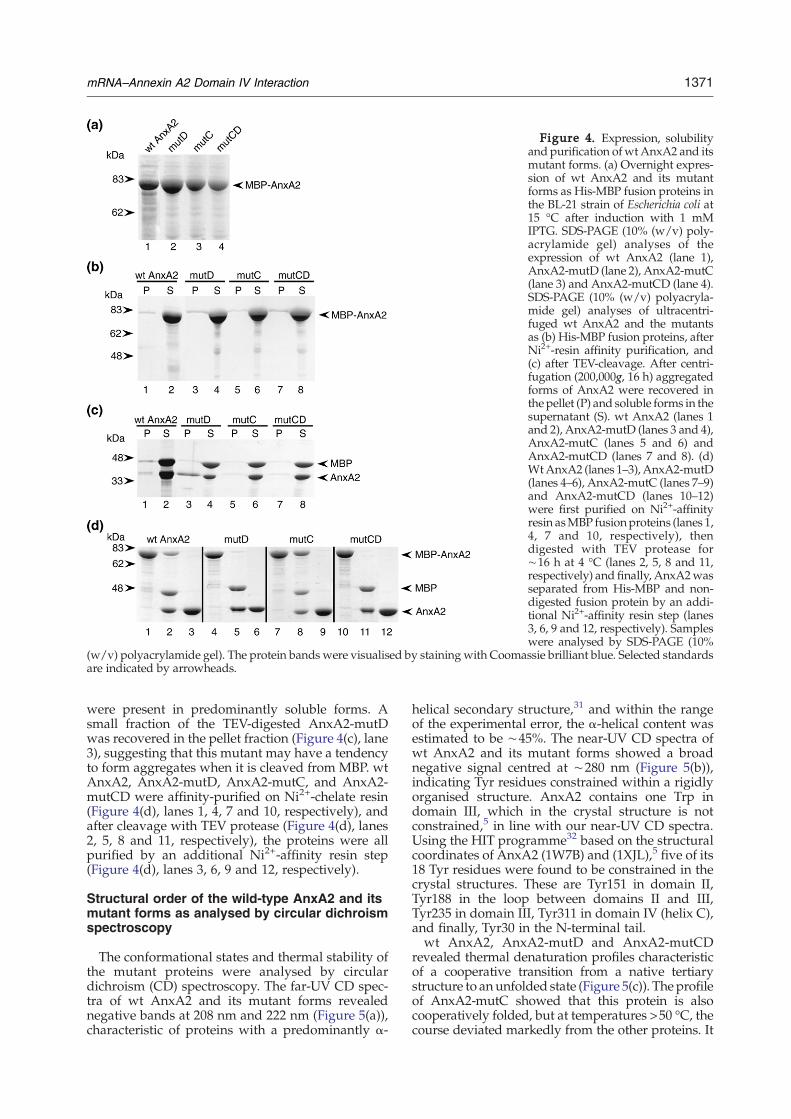
Figure 4. Expression, solubilityand purification ofwtAnxA2 and itsmutant forms. (a) Overnight expres-sion of wt AnxA2 and its mutantforms as His-MBP fusion proteins inthe BL-21 strain of Escherichia coli at15 °C after induction with 1 mMIPTG. SDS-PAGE (10% (w/v) poly-acrylamide gel) analyses of theexpression of wt AnxA2 (lane 1),AnxA2-mutD (lane 2), AnxA2-mutC(lane 3) and AnxA2-mutCD (lane 4).SDS-PAGE (10% (w/v) polyacryla-mide gel) analyses of ultracentri-fuged wt AnxA2 and the mutantsas (b) His-MBP fusion proteins, afterNi2+-resin affinity purification, and(c) after TEV-cleavage. After centri-fugation (200,000g, 16 h) aggregatedforms of AnxA2 were recovered inthe pellet (P) and soluble forms in thesupernatant (S). wt AnxA2 (lanes 1and 2), AnxA2-mutD (lanes 3 and 4),AnxA2-mutC (lanes 5 and 6) andAnxA2-mutCD (lanes 7 and 8). (d)WtAnxA2 (lanes 1–3), AnxA2-mutD(lanes 4–6), AnxA2-mutC (lanes 7–9)and AnxA2-mutCD (lanes 10–12)were first purified on Ni2+-affinityresin asMBP fusionproteins (lanes 1,4, 7 and 10, respectively), thendigested with TEV protease for∼16 h at 4 °C (lanes 2, 5, 8 and 11,respectively) and finally, AnxA2wasseparated from His-MBP and non-digested fusion protein by an addi-tional Ni2+-affinity resin step (lanes3, 6, 9 and 12, respectively). Sampleswere analysed by SDS-PAGE (10%
(w/v) polyacrylamide gel). The protein bands were visualised by staining with Coomassie brilliant blue. Selected standardsare indicated by arrowheads.
1371mRNA–Annexin A2 Domain IV Interaction
were present in predominantly soluble forms. Asmall fraction of the TEV-digested AnxA2-mutDwas recovered in the pellet fraction (Figure 4(c), lane3), suggesting that this mutant may have a tendencyto form aggregates when it is cleaved from MBP. wtAnxA2, AnxA2-mutD, AnxA2-mutC, and AnxA2-mutCD were affinity-purified on Ni2+-chelate resin(Figure 4(d), lanes 1, 4, 7 and 10, respectively), andafter cleavage with TEV protease (Figure 4(d), lanes2, 5, 8 and 11, respectively), the proteins were allpurified by an additional Ni2+-affinity resin step(Figure 4(d), lanes 3, 6, 9 and 12, respectively).
Structural order of the wild-type AnxA2 and itsmutant forms as analysed by circular dichroismspectroscopy
The conformational states and thermal stability ofthe mutant proteins were analysed by circulardichroism (CD) spectroscopy. The far-UV CD spec-tra of wt AnxA2 and its mutant forms revealednegative bands at 208 nm and 222 nm (Figure 5(a)),characteristic of proteins with a predominantly α-
helical secondary structure,31 and within the rangeof the experimental error, the α-helical content wasestimated to be ∼45%. The near-UV CD spectra ofwt AnxA2 and its mutant forms showed a broadnegative signal centred at ∼280 nm (Figure 5(b)),indicating Tyr residues constrained within a rigidlyorganised structure. AnxA2 contains one Trp indomain III, which in the crystal structure is notconstrained,5 in line with our near-UV CD spectra.Using the HIT programme32 based on the structuralcoordinates of AnxA2 (1W7B) and (1XJL),5 five of its18 Tyr residues were found to be constrained in thecrystal structures. These are Tyr151 in domain II,Tyr188 in the loop between domains II and III,Tyr235 in domain III, Tyr311 in domain IV (helix C),and finally, Tyr30 in the N-terminal tail.wt AnxA2, AnxA2-mutD and AnxA2-mutCD
revealed thermal denaturation profiles characteristicof a cooperative transition from a native tertiarystructure to anunfolded state (Figure 5(c)). The profileof AnxA2-mutC showed that this protein is alsocooperatively folded, but at temperatures >50 °C, thecourse deviated markedly from the other proteins. It

Figure 5. CD spectra of wt AnxA2 and its mutantforms. (a) Far-UV and (b) near-UV CD spectra wereobtained at a protein concentration of 18 μM and 15 μM,respectively, in 20 mM Tris–HCl (pH 8) at 20 °C. (c) CD-monitored thermal disruption of the α-helicity at 18 μM ofthe respective proteins. The change in ellipticity at 222 nmwasmeasured at pH 8 at a heating rate of 40 deg.C/h from20 °C to 90 °C. The apparent transition temperature (Tm)was determined from the first derivative of the curve. Allthe spectra represented in this Figure are background-corrected and smoothed, and the observed optical activityis expressed as the mean residue molar ellipticity [θ]MRW(deg cm2 dmol−1). Figure 6. The interaction of wt AnxA2 and its mutant
forms with 3′-UTR anxA2 mRNA as measured by UVcross-linking assays. (a) The binding of 4 fmol(∼200,000 cpm) of the radiolabelled 3′-UTR of anxA2mRNA to 2 μg of wt AnxA2 (lane 1), AnxA2-mutD (lane2), AnxA2-mutC (lane 3) or AnxA2-mutCD (lane 4). (b)Coomassie brilliant blue staining of AnxA2 after SDS-PAGE before phosphorimaging shown in (a). Binding wasperformed in the presence of 0.5 mg/ml of yeast tRNA,followed by UV cross-linking, treatment with RNase, SDS-PAGE (15% (w/v) polyacrylamide gel) and staining withCoomassie brilliant blue. Binding was visualised bydetection of signal using a phosphorimager.
1372 mRNA–Annexin A2 Domain IV Interaction
was noticed that the photomultiplier voltage of theinstrument in this case continued to increase attemperatures above 50 °C, in contrast to the otherAnxA2 forms (results not shown), compatiblewith anincreasing aggregation of AnxA2-mutC above thistemperature. wt AnxA2 and AnxA2-mutD havesimilar transition temperatures (Tm) of ∼55 °C,whereas AnxA2-mutC and AnxA2-mutCD show a
lower thermal stability with Tm-values at∼49 °C and∼48 °C, respectively. Altogether, these results revealthat all the mutants adopt an overall cooperativelyfolded structure with a content of helical secondaryand tertiary structure comparable to wt AnxA2, butwith some structural perturbations.
The interaction of wild-type AnxA2 and itsmutant forms with the 3′-UTR of anxA2 mRNA
The UV cross-linking experiments (Figure 6)revealed that the interaction of the 3′-UTR ofanxA2 mRNA with AnxA2-mutD (lane 2), AnxA2-mutC (lane 3) and AnxA2-mutCD (lane 4) is reducedby ∼80%, ∼90% and ∼98%, respectively, whencompared to wt AnxA2 (Figure 6, lane 1). Thesenumbers were obtained bymeasuring the binding ofradioactive nucleotides (cpm) to AnxA2 (Figure6(a)) relative to the amount of protein loaded on theSDS/polyacrylamide gel (Figure 6(b)). Furtherinformation on these interactions was obtained byreal-time SPR analyses. The interaction between theimmobilised 3′-UTR of anxA2 mRNA and wtAnxA2 (Figure 7(a)) resulted in sensorgrams thatwere biphasic both during the association and thedissociation phase. The association phase revealed arapid burst (∼2 s), followed by a slower increase ofsignal. As for the sensorgrams shown in Figure 1(b),the data could not be explained by any of thecommon binding models in the BIAevaluationprogramme, and therefore no Kd could be calculatedfrom the observed association and dissociation rateconstants. Since electrostatic interactions betweenthe negatively charged RNA and basic amino acidsare often observed in RNA–protein interactions,33
the effect of ionic strength was studied. The sen-sorgrams shown in Figure 7(a) were obtained at150 mM NaCl, but when the concentration wasincreased to 200 mM NaCl, the interaction between

Figure 7. The binding of wt AnxA2 and its mutant forms to immobilised 3′-UTR anxA2 mRNA as measured by SPR.Representative SPR sensorgrams for the interaction of (a) wt AnxA2, (b) AnxA2-mutC, (c) AnxA2-mutCD and (e) AnxA2-mutD with immobilised 3′-UTR of anxA2 mRNA. The protein was injected in random order at a flow-rate of 30 μl/minand in HBS-P buffer with 70 μM Ca2+ and 1 mM Mg2+. The level of immobilised mRNA was ∼530 RU in all theexperiments, except for those presented in (e), where it was∼380 RU. All sensorgrams were processed by first subtractingthe SPR response observed for the reference surface to correct for the contribution of non-specific interaction with thedextranmatrix itself. (d) Comparison of the interaction of 3 μMwtAnxA2, AnxA2-mutC andAnxA2-mutCDwith 3′-UTRof anxA2 mRNA as studied by SPR analysis; the sensorgrams are taken from (a), (b) and (c). AnxA2-mutD is omitted dueto its tendency to aggregate (see the text).
1373mRNA–Annexin A2 Domain IV Interaction
AnxA2 and the 3′-UTR of anxA2 mRNA wasabolished completely (results not shown).The two mutant forms, AnxA2-mutC (Figure 7(b))
and AnxA2-mutCD (Figure 7(c)), revealed a markeddecrease in the apparent affinity of the interactionwith the 3′-UTR of anxA2 mRNA, confirming theresults obtained by UV cross-linking (Figure 6). Acomparison of the sensorgrams for the interaction ofwt AnxA2, AnxA2-mutC and AnxA2-mutCD, all at3 μM, with the 3′-UTR of anxA2 mRNA is presented
in Figure 7(d), which demonstrates that AnxA2-mutC and AnxA2-mutCD interact poorly withmRNA.The SPR signal observed for AnxA2-mutD (Figure
7(e)) was about fourfold higher than for wt AnxA2(Figure 7(a)). The ΔRUmax was far above thetheoretically estimated value (∼190 RU), indicatingaggregation of this mutant protein, as also observedduring its purification in the absence of Ca2+ (Figure4(c), lane 3). The signal in the sensorgrams was

1374 mRNA–Annexin A2 Domain IV Interaction
expected to be low, since the monomers are bindingwith low affinity to the 3′-UTR of anxA2 mRNA asshown by the UV cross-linking experiments (Figure6) but, due to aggregation of the protein, the signal isenhanced dramatically.
Discussion
RNA transcripts interact with different RNA-binding proteins, forming mRNP complexes. Someproteins are associated with the mRNA throughouttheir lifetime, while others participate in specificsteps of mRNA processing, such as transport,anchorage and/or translation.34 This interactionwith trans-acting proteins is mediated by cis-actingelements located mainly in the 5′ and 3′-UTRs ofmRNAs.35–37
In general, it has been proposed that protein–mRNA interactions may be either strictly single-stranded (ss) RNA sequence-dependent or relymostly on specific higher-order RNA structures,including stem–loop structures.38 In the first case,β-sheet domains in theRNA-bindingproteins appearto be involved, while in the latter case α-helical do-mains seem to be important.38 AnxA2 is composed offour compact domains, each containing five α-he-lices.4 Interestingly, the structure of another RNA-binding protein, S15, also contains five α-helices,39
not normally encountered in RNA-binding proteins.AnxA2 does not contain any of the most abundantRNA-binding motifs such as the RNA-recognitionmotif (RRM), the K-homology (KH) domain and thedouble-stranded RNA-binding motif (dsRBM).Neither does it contain any zinc finger, the oligonu-cleotide/oligosaccharide binding (OB) fold, RGGboxes nor the Pumilio homology domain found inPUF proteins.34,40,41 The closest resemblance appearsto be with the helix-turn-helix motif found in Ffh/SRP54, a component of the signal recognition particle(SRP),42 and the Rev protein of HIV-1.43
Protein–RNA interactions have generally beeninvestigated by assays describing RNA bindingunder non-equilibrium conditions, such as spotblots, UV cross-linking and electrophoretic mobi-lity-shift assays, which are inadequate to demon-strate transient and low-affinity interactions. Bycontrast, SPR spectroscopy is often a more reliablemethod to study such interactions in real-time,providing information about the kinetics as well asthe affinity of the interaction.44 However, care shouldbe taken in using SPR data to calculate Kd valuesbased on the kon and koff, since each of these phasesmay consist of several events, such as binding,conformational changes and aggregation (possiblyupon binding) as observed in the present study.
Surface-exposed basic, polar and aromaticresidues in helices C and D of domain IV areimportant for the AnxA2-mRNA interaction
Here, we demonstrate by two independent meth-ods (Figure 1) that the mRNA-binding site of AnxA2
resides in its domain IV. The interaction of themutantAnxA2-DIVwith the 3′-UTR of anxA2mRNAis specific (Figure 1(a)) and Ca2+-independent asmeasured by SPR (Figure 1(b)). By contrast, theinteraction of full-length wt AnxA2 with the 3′-UTRof anxA2mRNA is Ca2+-dependent (Figure 2(a)). Wealso observed by SPR that the binding of AnxA2 toimmobilised heparin was Ca2+-dependent (resultsnot shown), in agreement with previous studies.14
This Ca2+-dependent interaction has been linked to aCa2+-induced conformational change of the protein,as shown crystallographically,14 generating afavourable binding site. A similar mechanism mayexplain the effect of Ca2+ on the interaction of full-length AnxA2 with the 3′-UTR of anxA2 mRNA. Inline with this suggestion, the binding of Ca2+ toAnxA2 results in changes of the backbone dihedraltorsion angles at Gly312 in the CD-loop and thus alsoin the orientation of helix D relative to helix C.UV cross-linking (Figure 6) and SPR analyses
(Figure 7) have revealed that an array of basicresidues in helix C (Lys308, Lys309 and Lys310) ofAnxA2 is important for its interaction with 3′-UTRanxA2 mRNA. Furthermore, multiple mutations ofLys313 in the CD-loop together with Tyr317 andGln321 in helix D also affect the interaction. Thestructurally related AnxA6 is known to bind thenucleotides ATP and GTP in vitro, and a consensussequence, (F/Y)XXX(F/Y)XKSL, of the putativenucleotide-binding site was proposed.45 Interest-ingly, the four Lys residues in the sequenceFKKKYGKSL of bovine AnxA2 that seem to beinvolved in the recognition of the 3′-UTR of anxA2mRNA, are located at a position equivalent to thenucleotide-binding site in AnxA6.The loss of RNA binding for the AnxA2 mutant
proteins could be provoked indirectly by thestructural perturbations that were detected in bothAnxA2-mutC and AnxA2-mutCD. However, bio-physical characterisation of the full-length multiplemutants revealed preservation of the structuralintegrity of the protein at the working temperatureused for the interaction assays. The individualcontributions of constrained Tyr residues to thenear-UV signal (Figure 5(b)) are known to bedifficult to quantify. However, the presence of thissignal and its conservation in all the mutants aswell as wt AnxA2 support the far-UV CD data(Figure 5(a)) by indicating that all the full-lengthAnxA2 forms present a cooperatively foldedtertiary structure. Since the folding process ofAnxA2 has been shown to be a fully cooperativeprocess in which the individual folding of domainsis regarded as critical for folding of the entiremolecule,46 our data suggest strongly that, attemperatures below 45 °C, all the mutants arefully folded in their overall structure and at thelevel of domain IV.A two-step model for protein–RNA interactions
has been proposed by Katsamba et al.47 for thebinding of U1A to U1hpII RNA, where the firstrapid step occurs via electrostatic interactionsbetween the negatively charged phosphate back-

1375mRNA–Annexin A2 Domain IV Interaction
bone of the RNA and the positively charged aminoacid residues of the protein. In the second step,short-range hydrogen bonding and aromatic stack-ing interactions take place between the protein andRNA, which are subsequently locked into a morestable complex.33,47 The present SPR results supportsuch a “lure and lock” model to describe theinteraction of AnxA2 with the 3′-UTR of its cognatemRNA. Increasing concentrations of salt (>200 mM)inhibit the wt AnxA2-mRNA interaction, most likelyby interfering with the initial recognition phaseinvolving electrostatic interactions between theRNA and the protein. It is likely that Lys313 in theCD-loop together with the array of Lys residues inhelix C form a patch ideal for electrostatic interac-tions with the phosphate backbone of RNA. Suchpatches are often involved in RNA binding.33,48 Theinitial phase is, by nature, rather unspecific. How-ever, AnxA2 binds to a specific region of the 3′-UTRof its cognate mRNA21 and c-myc mRNA,19 while itdoes not interact with the Renilla Luciferase mRNA(Figure 1(a)). This indicates the existence of a secondmore specific binding phase in which AnxA2-mRNA complexes are formed, possibly accompa-nied by some degree of conformational change ofthe complex. By binding to the 3′-UTRs of c-myc,19
anxA221 and collagen prolyl 4-hydroxylase-α(I)22
mRNAs, it is likely that AnxA2 is involved in thetransport and/or anchorage of specific mRNAs.This conclusion is further supported by datashowing that AnxA2 binds to the stem–looplocalisation signal present in the 3′-UTR of c-mycmRNA19 and to F-actin.1
In principle, the interaction of wt AnxA2 with themRNA could be mediated by a direct contact (e.g.electrostatic interactions) or indirectly via Ca2+.Since high concentrations of salt inhibit the AnxA2-mRNA interaction,21 we favour a direct bindingmechanism. By contrast, the interaction of AnxA2with another negatively charged molecule, heparin,is of a different mode. The latter interaction in-volves main-chain and side-chain interactions, andCa2+ bound to the AB and DE-loops of domainIV.14
In conclusion, our results provide a more preciseunderstanding of the molecular mechanismwhereby AnxA2 interacts with its cognate mRNA.We show by two independent methods that themRNA-binding site resides in its domain IV andinvolves helices C and D. The AnxA2-mRNAinteraction is Ca2+-dependent and specific, interms of both the mRNA19,21 and the protein. Bymutating only three surface-exposed residues inhelices C or D (including the CD-loop) of domain IVof AnxA2, the interaction with mRNA is stronglyinhibited, indicating that the interaction is highlyspecific also at the protein level. Furthermore,solvent-exposed residues, most likely Lys, in thisregion appear to be involved in electrostatic inter-actions with the mRNA. Work is in progress toproduce and analyse crystals of AnxA2-mRNAcomplexes in order to obtain more detailed informa-tion on the molecular basis of the interaction.
Materials and Methods
Construction of domain IV of AnxA2, wild-type AnxA2and its mutant forms
The bovine anxA2 cDNA,49 cloned into the pGEM-3Zf(+)vector, was used as a template for the construction of wtAnxA2 and the three AnxA2 mutants (AnxA2-mutC,AnxA2-mutD and AnxA2-mutCD, for details see Figure 3)by PCR. wt AnxA2 was generated as a single fragment atthe DNA level, while AnxA2-mutC, AnxA2-mutD andAnxA2-mutCD were generated from two fragments,using the PCR/BbsI seamless cloning method.50 ThePCR fragments were ligated into the TOPO vector(Invitrogen) using TA cloning before digestion withrestriction enzymes (BbsI, NcoI and Acc651) and ligationof the PCR products into the NcoI and Acc651 sites in thepETM-41 expression vector. The pETM-41 vector waskindly provided by Gunter Stier (EMBL, Heidelberg). Thevector is derived from the pET24d expression vector(Novagen) and contains a His6-MBP tag followed by aTEV protease-sensitive linker (ENLYFQG). The TEVcleavage site results in four additional residues (GAMG)at the N terminus of the constructs. Oligonucleotideprimers were obtained from Eurogentec and TIB MOL-BIOL. DNA sequencing verified the identities of the finalconstructs. The construction of the mutant AnxA2-DIV hasbeen described.24
Protein expression and purification
The expression and purification of the His-MBP-AnxA2fusion proteins have been explained in detail.24 Briefly, thefusion proteins were purified on His-Select Ni2+-affinityresin (Sigma) and subsequently treated with His-TEVprotease to separate AnxA2 from its fusion partner.Digested AnxA2 was purified from His-MBP, His-TEVand the undigested form, by loading it to His-Select Ni2+-affinity resin once again.
Determination of the aggregation state byultracentrifugation
Protein samples (after Ni2+-affinity purification or TEVcleavage) of 55–143 μg in 20 mM Tris–HCl (pH 8) weresubjected to ultracentrifugation (200,000gav, 16 h at 4 °C)before SDS-PAGE analysis. Equal amounts of pellet andsupernatant of each protein sample were loaded indifferent lanes.
Circular dichroism measurements
Far-UV CD spectra (185–260 nm, light path 1 mm) andnear-UV CD spectra (260–300 nm, light path 5 mm) wererecorded at 20 °C in a Jasco J-810 spectropolarimeterequipped with a Peltier temperature control unit. Theprotein was diluted in 20 mM Tris–HCl (pH 8) to aconcentration of 18 μM (far-UV) or 15 μM (near-UV). Thespectra obtained were the average of four (far-UV) or 16(near-UV) scans. Buffer scans were subtracted from theprotein spectra. Thermal transition curves (20–90 °C)were determined by monitoring the decrease in ellipticityat 222 nm at a scanning rate of 40 degC/h. Back-calculations of the helical secondary structure contentwere performed using the VARiable SELECtion method(VARSELEC).51

1376 mRNA–Annexin A2 Domain IV Interaction
In vitro transcription
The 3′-UTR of full-length bovine anxA2 cDNA49 wassubcloned into the pGEM3Zf+vector. The construct wassequenced in both directions to verify the sequence of theinsert. The mRNA was transcribed either unlabelled or[α-32P]UTP-labelled as described for the RibomaxTMLarge Scale RNA Production System-SP6 (Promega). Uni-formly [α-32P]UTP-labelled in vitro transcribed 3′-UTR ofanxA2 mRNA achieved a specific activity of 1.0×108–2.5×108 cpm/μg. The purity and the integrity of the RNAsample were verified by agarose gel electrophoresis.
UV cross-linking assays
The mRNA-AnxA2 binding assays were performedessentially as described21 but with a few modifications.RNase T1 and RNase Awere added for 60 min at 37 °C to afinal concentration of 0.01 U/μl and 0.1 μg/μl, respec-tively, to digest unprotected RNA. The RNA bindingsolution was 10 mM Hepes (pH 7.4), 150 mM NaCl and0.005% surfactant P20 (HBS-P from Biacore), containing70 μM Ca2+, 0.5 mg/ml of tRNA and 1 mM Mg2+.
Surface plasmon resonance analyses
SPR analyses were performed with a Biacore 3000biosensor system (Biacore AB, Uppsala, Sweden) at 25 °C.Biotinylated RNA molecules were immobilised to astreptavidin (SA) sensor chip, which is the method usedmost often in SPR studies of RNA–protein interactions.The commercial SA chip from Biacore, a CM5 sensor chippre-immobilised with SA, was found to be unsuitable inthis study due to the high negatively charged dextransurface at pH 7, which caused a substantial non-specificbinding of the basic AnxA2 protein. The matrix of thesensor chip CM4 is similar to that of CM5, but the formerhas a lower degree of carboxymethylation and therefore areduced net negative charge at pH 7. Thus, non-specificbinding of AnxA2 to the dextran matrix was eliminatedusing these CM4 sensor chips.SA (a generous gift from Biacore AB, Uppsala, Sweden)
was first immobilised to the CM4 sensor chip using thestandard primary amine coupling reaction described bythe manufacturer. Before immobilisation, SA was dilutedin 10 mM sodium acetate buffer (pH 4.5) to a finalconcentration of 200 μg/ml. A surface concentration of2500–4300 RU SA was immobilised, and the runningbuffer was HBS-P (10 mM Hepes (pH 7.4), 150 mM NaCland 0.005% surfactant P20) (Biacore) at a flow rate of 5 μl/min. SA was also immobilised to the reference surface.After SAwas immobilised, the sensorgramwas run until
a stable baseline was obtained. Both flow-cells weresubsequently coated with 100–300 RU biotinylatedssDNA oligonucleotide (5′-GCATGCAAGCTTGAG-TATTC-3′biotin) using a solution of 10–100 nM oligonu-cleotide in HBS-P buffer at a flow-rate of 2 μl/min. Theoligonucleotide is complementary to a linker region of 20nucleotides at the 5′end of 3′-UTR anxA2 mRNA, leavingthe entire 3′-UTR of anxA2mRNA available for interactionwith the protein. It should be noted that the 307nt 3′-UTRof anxA2 mRNA was used as a ligand in all the SPRexperiments, rather than the 1381nt full-length mRNAcontaining more than one AnxA2-binding site,21 whichcould further complicate the interpretation of the SPR data.The 3′-UTR of anxA2 mRNA was diluted in 10 mM
Hepes (pH 7.4), 1 M NaCl and 0.005% surfactant P20,
heated at 72 °C for 3 min and subsequently cooled on icefor 5 min before injection, to ensure that the linker regionwould be available for hybridisation to the biotinylatedssDNA oligonucleotide. The mRNA was injected step-wise at a concentration of 500 nM using a flow-rate of 2 μl/min, until the desired immobilisation level of ∼500 RUwas achieved. The amount of RNAwas minimised to limitmass transport and rebinding effects. No anxA2 mRNAwas captured on the reference surface, which contained anantisense oligonucleotide hybridised to the immobilisedbiotinylated ssDNA oligonucleotide. A limiting factor inthe indirect immobilisation strategy in which RNA isimmobilised via biotinylated ssDNA oligonucleotides, isthat these RNA surfaces are less stable than those made bydirect coupling.44 We also observed such instability and,therefore, after each series of analyte injection, new RNAwas immobilised to the same level as in the previous cycle.The AnxA2-mRNA binding assays were performed in
HBS-P buffer in the presence of 1 mM Mg2+ and theindicated concentrations of Ca2+; Mg2+ was included tostabilise the mRNA secondary structure. The protein wasultracentrifuged at 360,000gav for 10min in the same bufferto remove possible aggregates before injection. Differentprotein concentrations (0.5–4 μM) were injected over bothflow-cells at 25 °C using a flow-rate of 30 μl/min. Allsensorgrams were processed by first subtracting the SPRresponse observed for the reference surface to correct forany contribution of non-specific interaction with thedextran matrix itself.
Acknowledgements
The study was supported by grants from the EWSand Oddrun Mjåland′s foundation. Ali Sepulvedaand JeffreyMcKinney are greatly appreciated for thepurification of TEV protease. Gunther Stier at EMBL(Heidelberg) is thanked for the pETM-41 vector. Weare grateful to Anita Larsson and Åsa Rosengren atBiacore AB (Uppsala, Sweden) for their assistance inthe SPR analyses and for the donation of streptavi-din. Finally, we are indebted to Volker Gerke andRobert H. Kretsinger for valuable comments on themanuscript.
References
1. Hayes, M. J., Shao, D., Bailly, M. & Moss, S. E. (2006).Regulation of actin dynamics by annexin 2. EMBO J.25, 1816–1826.
2. Gerke, V., Creutz, C. E. &Moss, S. E. (2005). Annexins:linking Ca2+ signalling to membrane dynamics.NatureRev. Mol. Cell Biol. 6, 449–461.
3. Hayes, M. J., Rescher, U., Gerke, V. & Moss, S. E.(2004). Annexin-actin interactions. Traffic, 5, 571–576.
4. Gerke, V. & Moss, S. E. (2002). Annexins: fromstructure to function. Physiol. Rev. 82, 331–371.
5. Rosengarth, A. & Luecke, H. (2004). Annexin A2. Doesit induce membrane aggregation by a new multimericstate of the protein? Annexins, 1, 129–136.
6. Thiel, C., Weber, K. & Gerke, V. (1991). Character-ization of a Ca(2+)-binding site in human annexin IIby site-directed mutagenesis. J. Biol. Chem. 266,14732–14739.

1377mRNA–Annexin A2 Domain IV Interaction
7. Rosengarth, A., Rösgen, J., Hinz, H. J. & Gerke, V.(2001). Folding energetics of ligand binding proteins.II. Cooperative binding of Ca2+ to annexin I. J. Mol.Biol. 306, 825–835.
8. Follenius-Wund, A., Pigault, C. & Gerard, D. (1993).Ca(2+)- and Mg(2+)-dependent binding of annexin IIand its complex to phospholipid vesicles; a compara-tive spectroscopic study. Biochem. Mol. Biol. Int. 29,653–660.
9. Ayala-Sanmartin, J., Vincent, M., Sopkova, J. & Gallay,J. (2000). Modulation by Ca(2+) and by membranebinding of the dynamics of domain III of annexin 2(p36) and the annexin 2-p11 complex (p90): implica-tions for their biochemical properties. Biochemistry, 39,15179–15189.
10. Rosengarth, A. & Luecke, H. (2003). A calcium-drivenconformational switch of the N-terminal and coredomains of annexin A1. J. Mol. Biol. 326, 1317–1325.
11. Glenney, J. R. & Tack, B. F. (1985). Amino-terminalsequence of p36 and associated p11: identification ofthe site of tyrosine phopshorylation and homologywith S-100. Proc. Natl Acad. Sci. USA 82, 7884–7888.
12. Jost, M. & Gerke, V. (1996). Mapping of a regulatoryimportant site for protein kinase C phosphorylation inthe N-terminal domain of annexin II. Biochim. Biophys.Acta, 3, 283–289.
13. Johnsson, N., Nguyen, V. P., Soling, H. D. & Weber, K.(1986). Functionally distinct serine phosphorylationsites of p36, the cellular substrate of retroviral proteinkinase; differential inhibition of reassociation withp11. EMBO J. 5, 3455–3460.
14. Shao, C., Zhang, F., Kemp, M. M., Linhardt, R. J.,Waisman, D. M., Head, J. F. & Seaton, B. A. (2006).Crystallographic analysis of calcium-dependent hepa-rin binding to annexin A2. J. Biol. Chem. 281,31689–31695.
15. Gerke, V. & Weber, K. (1984). Identity of p36Kphosphorylated upon Rous sarcoma virus transfor-mation with a protein purified from brush borders:calcium-dependent binding to non-erythroid spectrinand F-actin. EMBO J. 3, 227–233.
16. Filipenko, N. R. & Waisman, D. M. (2001). The Cterminus of annexin II mediates binding to F-actin.J. Biol. Chem. 276, 5310–5315.
17. Vedeler, A.&Hollås, H. (2000). Annexin II is associatedwith mRNAs which may constitute a distinct sub-population. Biochem. J. 348, 565–572.
18. Filipenko, N. R., MacLeod, T. J., Yoon, C. S. &Waisman, D. M. (2004). Annexin A2 is a novel RNA-binding protein. J. Biol. Chem. 279, 8723–8731.
19. Mickleburgh, I., Burtle, B., Hollås, H., Campbell, G.,Chrzanowska-Lightowlers, Z., Vedeler, A. & Hesketh,J. (2005). Annexin A2 binds to the localization signal inthe 3′ untranslated region of c-myc mRNA. FEBS J.272, 413–421.
20. Chabanon, H., Mickleburgh, I., Burtle, B., Pedder, C. &Hesketh, J. (2005). An AU-rich stem-loop structure is acritical feature of the perinuclear localization signal ofc-myc mRNA. Biochem. J. 392, 475–483.
21. Hollås, H., Aukrust, I., Grimmer, S., Strand, E.,Flatmark, T. & Vedeler, A. (2006). Annexin A2 rec-ognises a specific region in the 3′-UTR of its cognatemessenger RNA. Biochim. Biophys. Acta, 1763,1325–1334.
22. Fähling, M., Mrowka, R., Steege, A., Nebrich, G.,Perlewitz, A., Persson, P. B. & Thiele, B. J. (2006).Translational control of collagen prolyl-4-hydroxy-lase-alpha(I) gene expression under hypoxia. J. Biol.Chem. 281, 26089–26101.
23. Keene, J. D. (2001). Ribonucleoprotein infrastructureregulating the flow of genetic information between thegenome and the proteome. Proc. Natl Acad. Sci. 98,7018–7024.
24. Aukrust, I., Evensen, L., Hollås, H., Berven, F.,Atkinson, R. A., Travé, G. et al. (2006). Engineering,biophysical characterisation and binding properties ofa soluble mutant form of annexin A2 domain IV thatadopts a partially folded conformation. J. Mol. Biol.363, 469–481.
25. Geitmann, M. & Danielson, U. H. (2004). Studies ofsubstrate-induced conformational changes in humancytomegalovirus protease using optical biosensortechnology. Anal. Biochem. 332, 203–214.
26. Zaks, W. J. & Creutz, C. E. (1991). Ca(2+)-dependentannexin self-association on membrane surfaces. Bio-chemistry, 30, 9607–9615.
27. Travé, G., Cregut, D., Lionne, C., Quignard, J. F.,Chiche, L., Sri, J. & Widada, J. P. (1994). Site-directedmutagenesis of a calcium binding site modifies speci-fically the different biochemical properties of annexinI. Protein Eng. 7, 689–696.
28. Parthiban, V., Gromiha, M. M. & Schomburg, D.(2006). CUPSAT: prediction of protein stability uponpoint mutations. Nucl. Acids Res. 34, W239–W242.
29. Ristriani, T., Nominé, Y., Laurent, C., Weiss, E. &Travé, G. (2002). Protein mutagenesis with monodis-persity-based quality probing: selective inactivation ofp53 degradation and DNA-binding properties of HPVE6 oncoprotein. Protein Expr. Purif. 26, 357–367.
30. Nominé, Y., Ristriani, T., Laurent, C., Lefèvre, J. F.,Weiss, E. & Travé, G. (2001). Formation of solubleinclusion bodies by hpv E6 oncoprotein fused tomaltose-binding protein. Protein Expr. Purif. 23, 22–32.
31. Kelly, S. M., Jess, T. J. & Price, N. C. (2005). How tostudy proteins by circular dichroism. Biochim. Biophys.Acta, 10, 119–139.
32. Burba, A. E., Lehnert, U., Yu, E. Z. & Gerstein, M.(2006). Helix Interaction Tool (HIT): a web-based toolfor analysis of helix-helix interactions in proteins.Bioinformatics, 22, 2735–2738.
33. Law, M. J., Linde, M. E., Chambers, E. J., Oubridge, C.,Katsamba, P. S., Nilsson, L. et al. (2006). The role ofpositively charged amino acids and electrostaticinteractions in the complex of U1A protein and U1hairpin II RNA. Nucl. Acids Res. 34, 275–285.
34. Moore, M. J. (2005). From birth to death: the complexlives of eukaryotic mRNAs. Science, 309, 1514–1518.
35. De Moor, C. H., Meijer, H. & Lissenden, S. (2005).Mechanisms of translational control by the 3′ UTR indevelopment and differentiation. Semin. Cell Dev. Biol.16, 49–58.
36. Pickering, B. M. & Willis, A. E. (2005). The implica-tions of structured 5′ untranslated regions on transla-tion and disease. Semin. Cell Dev. Biol. 16, 39–47.
37. Wilkie, G. S., Dickson, K. S. & Gray, N. K. (2003).Regulation of mRNA translation by 5′ and 3′UTR-binding factors. Trends Biochem. Sci. 28, 182–188.
38. Draper, D. E. (1999). Themes in RNA-protein recogni-tion. J. Mol. Biol. 293, 255–270.
39. Berglund, H., Rak, A., Serganov, A., Garber, M. &Hard, T. (1997). Solution structure of the ribosomalRNA binding protein S15 from Thermus thermophilus.Nature Struct. Biol. 4, 20–23.
40. Stefl, R., Skrisovska, L. & Allain, F. H. (2005). RNAsequence- and shape-dependent recognition by pro-teins in the ribonucleoprotein particle. EMBO Rep. 6,33–38.
41. Singh, R. & Valcárcel, J. (2005). Building specificity

1378 mRNA–Annexin A2 Domain IV Interaction
with nonspecific RNA-binding proteins.Nature Struct.Mol. Biol. 12, 645–653.
42. Rupert, P. B. & Ferré-D'Amaré, A. R. (2000). SRPrisesin RNA-protein recognition. Structure, 8, R99–R104.
43. Battiste, J. L., Mao, H., Rao, N. S., Tan, R., Muhan-diram, D. R., Kay, L. E. et al. (1996). Alpha helix-RNAmajor groove recognition in anHIV-1 rev peptide-RRERNA complex. Science, 273, 1547–1551.
44. Katsamba, P. S., Park, S. & Laird-Offringa, I. A. (2002).Kinetic studies of RNA-protein interactions usingsurface plasmon resonance. Methods, 26, 95–104.
45. Bandorowicz-Pikula, J., Kirilenko, A., vanDeursen, R.,Golczak, M., Kuhnel, M., Lancelin, J. M. et al. (2003). Aputative consensus sequence for the nucleotide-bind-ing site of annexin A6. Biochemistry, 42, 9137–9146.
46. Cordier-Ochsenbein, F., Guerois, R., Russo-Marie, F.,Neumann, J. M. & Sanson, A. (1998). Exploring thefolding pathways of annexin I, a multidomain protein.II. Hierarchy in domain folding propensities maygovern the folding process. J. Mol. Biol. 279, 1177–1185.
47. Katsamba, P. S., Myszka, D. G. & Laird-Offringa, I. A.(2001). Two functionally distinct steps mediate highaffinity binding of U1A protein to U1 hairpin II RNA.J. Biol. Chem. 276, 21476–21481.
48. Cheng, S. & Gallie, D. R. (2006). Wheat eukaryoticinitiation factor 4B organizes assembly of RNA andeIFiso4G, eIF4a, and poly(A)-binding protein. J. Biol.Chem. 281, 24351–24364.
49. Kristensen, T., Saris, C. J., Hunter, T., Hicks, L. J.,Noonan, D. J., Glenney, J. R., Jr. et al. (1986). Primarystructure of bovine calpactin I heavy chain (p36), amajor cellular substrate for retroviral protein-tyro-sine kinases: homology with the human phospho-lipase A2 inhibitor lipocortin. Biochemistry, 25,4497–4503.
50. Zanier, K., Nominé, Y., Charbonnier, S., Ruhlmann, C.,Schultz, P., Schweizer, J. & Travé, G. (2006). Formationof well-defined soluble aggregates upon fusion toMBP is a generic property of E6 proteins from varioushuman papillomavirus species. Protein Expr. Purif. 51,59–70.
51. Johnson, W. C. (1999). Analyzing protein circulardichroism spectra for accurate secondary structures.Proteins: Struct. Funct. Genet. 35, 307–312.
52. Koradi, R., Billeter, M. & Wuthrich, K. (1996).MOLMOL: a program for display and analysis ofmacromolecular structures. J. Mol. Graph. 14, 51–55,29–32.
Edited by D. E. Draper
(Received 29 January 2007; received in revised form 26 February 2007; accepted 27 February 2007)Available online 7 March 2007
Copyright © 2022 FDOKUMEN



















