
The small GTP-binding protein, Rhes, regulates signal transduction from G protein-coupled receptors
10
The small GTP-binding protein, Rhes, regulates signal transduction from G protein-coupled receptors Pierfrancesco Vargiu 1,3 , Ricardo De Abajo 1,3 , Juan Antonio Garcia-Ranea 2,4 , Alfonso Valencia 2 , Pilar Santisteban 1 , Piero Crespo 1 and Juan Bernal* ,1 1 Instituto de Investigaciones Biome´dicas Alberto Sols. Consejo Superior de Investigaciones Cientı´ficas (CSIC) and Universidad Auto´noma de Madrid (UAM), 28029 Madrid, Spain; 2 Centro Nacional de Biotecnologı´a. CSIC-UAM, 28049 Madrid, Spain The Ras homolog enriched in striatum, Rhes, is the product of a thyroid hormone-regulated gene during brain development. Rhes and the dexamethasone-induced Dex- ras1 define a novel distinct subfamily of proteins within the Ras family, characterized by an extended variable domain in the carboxyl terminal region. We have carried this study because there is a complete lack of knowledge on Rhes signaling. We show that in PC12 cells, Rhes is targeted to the plasma membrane by farnesylation. We demonstrate that about 30% of the native Rhes protein is bound to GTP and this proportion is unaltered by typical Ras family nucleotide exchange factors. However, Rhes is not transforming in murine fibroblasts. We have also examined the role of Rhes in cell signaling. Rhes does not stimulate the ERK pathway. By contrast, it binds to and activates PI3K. On the other hand, we demonstrate that Rhes impairs the activation of the cAMP/PKA pathway by thyroid-stimulating hormone, and by an activated b2 adrenergic receptor by a mechanism that suggests uncoup- ling of the receptor to its cognate heterotrimeric complex. Overall, our results provide the initial insights into the role in signal transduction of this novel Ras family member. Oncogene (2004) 23, 559–568. doi:10.1038/sj.onc.1207161 Keywords: Ras; thyroid hormone; striatum; adrenergic receptor; PC12 cells Introduction The Ras family of small GTPases comprises several protein subfamilies (for recent reviews, see Campbell et al., 1998; Reuther and Der, 2000). Among the most recent additions made to the Ras superfamily are two proteins whose expression is regulated by hormones, Dexras1 and Rhes: Dexras1 was originally found in mice, and its name reflects the fact that it was induced by dexamethasone (Kemppainen and Behrend, 1998). A human Dexras homologue was subsequently isolated in a functional screen in yeast designed to identify receptor-independent activators of heterotrimeric G protein signaling, and was named as AGS1 (Cismowski et al., 1999, 2000). AGS1 was proposed to function as a guanine nucleotide exchange factor for Gai/Ga0. Dex- ras1/AGS1 was subsequently found to inhibit receptor- mediated Gai signalling by interfering with receptor-Gai coupling (Graham et al., 2002; Takesono et al., 2002). In other studies, Dexras1 was found to be cytoplasmic and was activated by S-nitrosylation induced by nNOS in response to NMDA receptor activation in brain (Fang et al., 2000; Jaffrey et al., 2002). We have described another Ras family member, which we named Rhes because it is greatly enriched in the striatum (Falk et al., 1999). A human Rhes ortholog was identified as a tumour endothelial marker and was named TEM-2 (St Croix et al., 2000). Rhes shares 62% identity with Dexras1 and is upregulated by thyroid hormone during the postnatal period in the rat (Vargiu et al., 2001). Both proteins contain domains present in prototypical Ras proteins including GTP binding and effector domains, a CAAX box for membrane localiza- tion, and an extended C-terminal variable domain that accounts for their greater molecular mass. However, the specific involvement of Rhes in signal- ling pathways has not been analysed thus far. Herein we present evidence indicating that, under basal conditions, a high proportion of wild-type Rhes exists in the GTP- bound form, but it displays no transforming activity. With regards to effector usage, Rhes does not activate the ERK pathway but is capable of binding to, and activate PI3K. Interestingly, we also found that Rhes inhibited the activation of a cAMP responsive element induced by a constitutively active b2-adrenergic recep- tor, but not when induced by forskolin or by a constitutively active Gas protein. Overall, our results indicate that Rhes displays distinctive characteristics among Ras GTPases, in particular in its role in the coupling between serpentine membrane receptors and heterotrimeric G protein signaling. Results The Rhes/Dexras subfamily On the basis of sequence homology and gene tree calcu- lations, Rhes and Dexras1 define a distinct subfamily of Received 21 May 2003; revised 26 August 2003; accepted 26 August 2003 *Correspondence: J Bernal, Instituto de Investigaciones Biome´dicas, Arturo Duperier 4, 28029 Madrid, Spain; E-mail: [email protected] 3 PV and RDA contributed equally to this work 4 Present Address: Biochemistry and Molecular Biology Department, University College London, Gower Street, London WC1E 6BT, UK Oncogene (2004) 23, 559–568 & 2004 Nature Publishing Group All rights reserved 0950-9232/04 $25.00 www.nature.com/onc
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The small GTP-binding protein, Rhes, regulates signal transduction from G protein-coupled receptors

The small GTP-binding protein, Rhes, regulates signal transduction from G
protein-coupled receptors
Pierfrancesco Vargiu1,3, Ricardo De Abajo1,3, Juan Antonio Garcia-Ranea2,4, Alfonso Valencia2,Pilar Santisteban1, Piero Crespo1 and Juan Bernal*,1
1Instituto de Investigaciones Biomedicas Alberto Sols. Consejo Superior de Investigaciones Cientıficas (CSIC) and UniversidadAutonoma de Madrid (UAM), 28029 Madrid, Spain; 2Centro Nacional de Biotecnologıa. CSIC-UAM, 28049 Madrid, Spain
The Ras homolog enriched in striatum, Rhes, is theproduct of a thyroid hormone-regulated gene during braindevelopment. Rhes and the dexamethasone-induced Dex-ras1 define a novel distinct subfamily of proteins withinthe Ras family, characterized by an extended variabledomain in the carboxyl terminal region. We have carriedthis study because there is a complete lack of knowledgeon Rhes signaling. We show that in PC12 cells, Rhes istargeted to the plasma membrane by farnesylation. Wedemonstrate that about 30% of the native Rhes protein isbound to GTP and this proportion is unaltered by typicalRas family nucleotide exchange factors. However, Rhes isnot transforming in murine fibroblasts. We have alsoexamined the role of Rhes in cell signaling. Rhes does notstimulate the ERK pathway. By contrast, it binds to andactivates PI3K. On the other hand, we demonstrate thatRhes impairs the activation of the cAMP/PKA pathwayby thyroid-stimulating hormone, and by an activated b2adrenergic receptor by a mechanism that suggests uncoup-ling of the receptor to its cognate heterotrimeric complex.Overall, our results provide the initial insights into the rolein signal transduction of this novel Ras family member.Oncogene (2004) 23, 559–568. doi:10.1038/sj.onc.1207161
Keywords: Ras; thyroid hormone; striatum; adrenergicreceptor; PC12 cells
Introduction
The Ras family of small GTPases comprises severalprotein subfamilies (for recent reviews, see Campbellet al., 1998; Reuther and Der, 2000). Among the mostrecent additions made to the Ras superfamily are twoproteins whose expression is regulated by hormones,Dexras1 and Rhes: Dexras1 was originally found inmice, and its name reflects the fact that it was inducedby dexamethasone (Kemppainen and Behrend, 1998).A human Dexras homologue was subsequently isolatedin a functional screen in yeast designed to identify
receptor-independent activators of heterotrimeric Gprotein signaling, and was named as AGS1 (Cismowskiet al., 1999, 2000). AGS1 was proposed to function as aguanine nucleotide exchange factor for Gai/Ga0. Dex-ras1/AGS1 was subsequently found to inhibit receptor-mediated Gai signalling by interfering with receptor-Gaicoupling (Graham et al., 2002; Takesono et al., 2002). Inother studies, Dexras1 was found to be cytoplasmic andwas activated by S-nitrosylation induced by nNOS inresponse to NMDA receptor activation in brain (Fanget al., 2000; Jaffrey et al., 2002).We have described another Ras family member,
which we named Rhes because it is greatly enriched inthe striatum (Falk et al., 1999). A human Rhes orthologwas identified as a tumour endothelial marker and wasnamed TEM-2 (St Croix et al., 2000). Rhes shares 62%identity with Dexras1 and is upregulated by thyroidhormone during the postnatal period in the rat (Vargiuet al., 2001). Both proteins contain domains present inprototypical Ras proteins including GTP binding andeffector domains, a CAAX box for membrane localiza-tion, and an extended C-terminal variable domain thataccounts for their greater molecular mass.However, the specific involvement of Rhes in signal-
ling pathways has not been analysed thus far. Herein wepresent evidence indicating that, under basal conditions,a high proportion of wild-type Rhes exists in the GTP-bound form, but it displays no transforming activity.With regards to effector usage, Rhes does not activatethe ERK pathway but is capable of binding to, andactivate PI3K. Interestingly, we also found that Rhesinhibited the activation of a cAMP responsive elementinduced by a constitutively active b2-adrenergic recep-tor, but not when induced by forskolin or by aconstitutively active Gas protein. Overall, our resultsindicate that Rhes displays distinctive characteristicsamong Ras GTPases, in particular in its role in thecoupling between serpentine membrane receptors andheterotrimeric G protein signaling.
Results
The Rhes/Dexras subfamily
On the basis of sequence homology and gene tree calcu-lations, Rhes and Dexras1 define a distinct subfamily ofReceived 21 May 2003; revised 26 August 2003; accepted 26 August 2003
*Correspondence: J Bernal, Instituto de Investigaciones Biomedicas,Arturo Duperier 4, 28029 Madrid, Spain; E-mail: [email protected] and RDA contributed equally to this work4Present Address: Biochemistry and Molecular Biology Department,University College London, Gower Street, London WC1E 6BT, UK
Oncogene (2004) 23, 559–568& 2004 Nature Publishing Group All rights reserved 0950-9232/04 $25.00
www.nature.com/onc

proteins within the Ras family (Figure 1). Rhes has alsobeen named as Dexras2, but we think that the term isnot appropriate because Rhes is not induced bydexamethasone (Falk et al., 1999). Also, as shown inthis work, there are a number of differences regardingintracellular signaling. Species distribution of Dexras/Rhes proteins suggests an early origin in the evolution ofthis new family, at least previous to the chordatedivergence. Longer branch lengths of the gene treeobserved in Rhes and Dexras sequences point to a moreaccelerated rate of amino-acid replacement and mayreflect reduced functional constrains or adaptation tonovel functions. The gene tree also shows that Dexrasand Rhes sequences appeared in mammals after theirseparation from insects. Distinguishing features ofDexras/Rhes proteins (Falk et al., 1999) include anextended carboxyl terminus variable domain of about 56amino acids in Rhes (residues 210–266) and 70 aminoacids in Dexras1 (residues 210–280), and an insertedregion of eight amino acids preceding the G2 motif. It ispossible that this additional region, shared by Rhes andDexras proteins, could carry out a specific function inthese sequences and it might mediate specific interac-tions with other proteins (Wittinghofer and Valencia,1995). These characteristics prompted us to undertakean analysis of Rhes biochemical properties in furtherdetail.
Rhes expression
Rhes was originally identified as a cDNA cloneencoding an mRNA very enriched in the striatum.As mentioned above, its gene product exhibits remark-able characteristics. Thus, it was of interest to identifyneural pathways that might be influenced by Rhes.
For this purpose, a notion about the areas in the brainwhere Rhes was expressed could provide valuableclues. Thus, we performed in situ hybridization insaggital and coronal slices of adult rat brain usinga radioactively labelled Rhes riboprobe. In Figure 2panel a, representative images from saggital and coronalsections from adult rat brain are displayed. Rheswas prominently expressed in the striatum, but it wasnot restricted to this region and was also detectedin other areas, such as the mitral cell layer of theolfactory bulb, olfactory tubercle, piriform cortex,layers 2/3 and 5 of cerebral cortex, accumbens nucleus,subiculum, pyramidal and granular layers of thehippocampus, anterior thalamic nuclei, inferior collicu-lus, and granular layer of the cerebellum. Interestingly,and although Rhes appeared to be primarily expressedin the brain among other organs, an mRNA of theexpected size was also detected in the thyroid gland(Figure 2, panel c).To gain an insight into what types of cells express
Rhes, we analysed its expression among a panel ofneural cell lines. Rhes mRNA could be detected inundifferentiated PC12 cells (Figure 2, panel b). How-ever, no Rhes transcripts were detected in otherneuronal cells such as GT1-7 (Mellon et al., 1990), orN2a cells, neither under basal conditions nor uponstimulation with thyroid hormones. Rhes mRNA wasalso undetected in the C6 glial cell line in agreement withits neuronal expression in vivo (Vargiu et al., 2001). InPC12 cells, Rhes expression was not altered by T3treatment or after the induction of differentiation byNGF (not shown).
Subcellular localization
We then investigated Rhes subcellular localization.This was undertaken in a physiological environmentsuch as PC12 cells. The presence of a conserved CAAXbox in the Rhes carboxyl terminus (CSIQ in ratRhes and CTIQ in human Rhes) predicts targetingof the protein to the plasma membrane. Thus, toexamine Rhes localization to the plasma membrane,PC12 cells were transfected with vectors encoding Rhesfused to the carboxyl terminus of the green fluorescentprotein. As shown in Figure 3, panel c, confocalmicroscopy revealed that the GFP-–Rhes fusion proteinwas clearly present in the plasma membrane. This wasconfirmed by indirect immunofluorescence utilizing anti-AU5 antibodies, after transfection of PC12 cells withAU5-tagged Rhes (Figure 3, panel a). Conversely, aGFP-–Rhes construct harbouring a Cys to Ala mutationwithin the CAAX box (Rhes C263A) was soluble andcould not be located in the plasma membrane (Figure 3,panel g). We then used specific inhibitors of farnesyla-tion or of geranylgeranylation to check for the nature ofthe lipid moiety attached to the Rhes C-terminus.Treatment of PC12 cells with the farnesylation inhibitorFTI-277 precluded Rhes presence in the cell periphery(Figure 3, panel b), whereas treatment with thegeranylgeranylation inhibitor GGTI-286 had no effectson Rhes membrane distribution (Figure 3, panel f).
Figure 1 Schematic phylogenetic tree of the Ras superfamily (Ras,Rho, and Rac, Rab, Arl, and Arf subfamilies). Other main proteingroups related to the Dexras/Rhes subfamily are included(Rr22_human, Rad, Gem, Rheb, Rsr1, Rap, Ral, and Ras-likeproteins: Nras, RasL, K-ras H-ras, Rras2, Rras). Branches lengthsof Ras subfamily cluster are proportional to the rate of amino-acidreplacements
Rhes inhibits GPCR signallingP Vargiu et al
560
Oncogene

Therefore, it may be concluded that Rhes undergoespost-translational farnesylation and is targeted to themembrane.
GTP binding and transforming activity
The Rhes sequence shows the presence of serine andarginine residues in positions 28 and 29, equivalent toRas glycines 12 and 13. In Ras, mutations in thesepositions suppress its GTPase activity and render aconstitutively active, transforming protein. Therefore,we checked whether this was the case for Rhes. First, wedetermined to which guanosine nucleotide Rhes waspreferentially bound under unstimulated conditions.PC12 cells were transfected with expression plasmidsencoding AU5-tagged Rhes or H-Ras, and nucleotidebinding was analysed in anti AU5 immunoprecipitatesfrom serum-starved cells after incubation with32P-orthophosphate in phosphate-free medium. Datafrom different experiments, shown in Figure 4, werepooled and analysed by ANOVA (result: F10,26¼15.497; P¼ 0.000). About 18% of H-Ras was found inthe GTP-bound form. Interestingly, Rhes contained asmuch as 30% in the GTP form, suggesting that wild-type Rhes could be constitutively activated (Po0.05, ascompared to H-Ras). We also tested whether GEFsspecific for the Ras family could catalyse GDP/GTP
Figure 2 Rhes expression. (a) In situ hybridization. The parasagittal (slices 1 and 2) and coronal (slices 3, 4, 5) sections of adult ratbrain are shown. The level of the parasagittal sections is shown in slice 3 by the arrows. Prominent sites of Rhes expression include themitral cell layer of olfactory bulb, olfactory tubercle, piriform cortex, caudate and accumbens nuclei, hippocampus, anterior nuclei ofthe thalamus, inferior colliculus, and granular layer of cerebellum. (b)Northern blotting of RNA from selected cell lines of neuralorigin. Only PC12 cells showed Rhes expression. (c) Northern blotting of RNA from rat tissues. Besides a prominent expression in thebrain, only the thyroid gland was found to express low levels of Rhes mRNA. The blots were stained for rRNA as a control of RNAloading (B: brain; Li: liver; Lu: lung; Ad: adrenals; Mu: muscle; Ki: kidney; Te: testis; Th: thyroid; Thy: thymus)
Figure 3 Rhes is targeted to the plasma membrane. PC12 cellswere analysed by confocal microscopy after transfection with thefollowing constructs: (a) Rhes containing an AU5 epitope anddetected by indirect immunofluorescence using an anti-AU5antibody. (b) GFP-Rhes with the cells treated with the farnesyla-tion inhibitor FTI-277. (c) GFP-Rhes fusion construct. (d) GFP. (e)H-Ras containing an AU5 epitope. (f) GFP-Rhes, with the cellstreated with the geranylgeranylation inhibitor GGTI-286. (g) GFP-Rhes containing a mutation in the CAAX box (Rhes C264A). (h)GFP with a CAAX box from H-Ras in the C-terminus
Rhes inhibits GPCR signallingP Vargiu et al
561
Oncogene
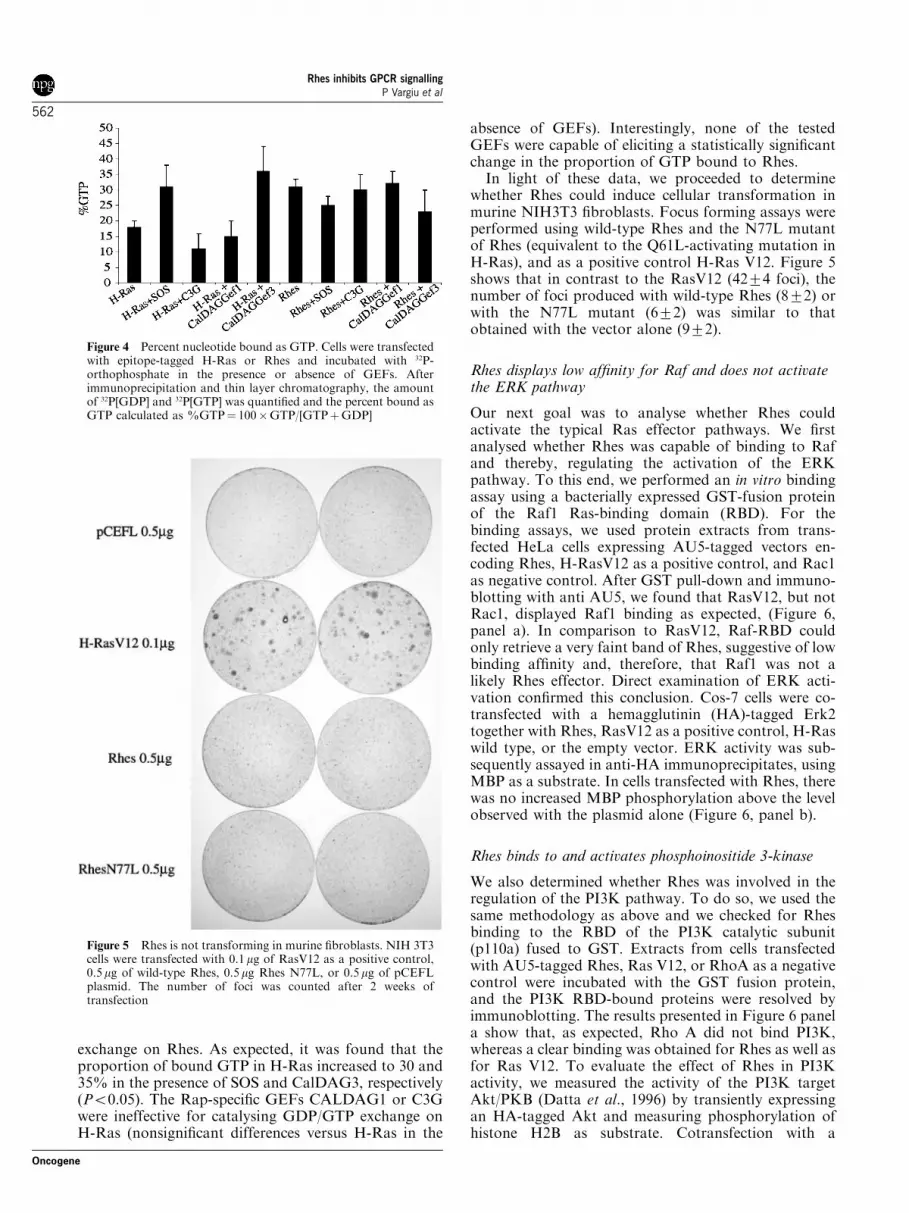
exchange on Rhes. As expected, it was found that theproportion of bound GTP in H-Ras increased to 30 and35% in the presence of SOS and CalDAG3, respectively(Po0.05). The Rap-specific GEFs CALDAG1 or C3Gwere ineffective for catalysing GDP/GTP exchange onH-Ras (nonsignificant differences versus H-Ras in the
absence of GEFs). Interestingly, none of the testedGEFs were capable of eliciting a statistically significantchange in the proportion of GTP bound to Rhes.In light of these data, we proceeded to determine
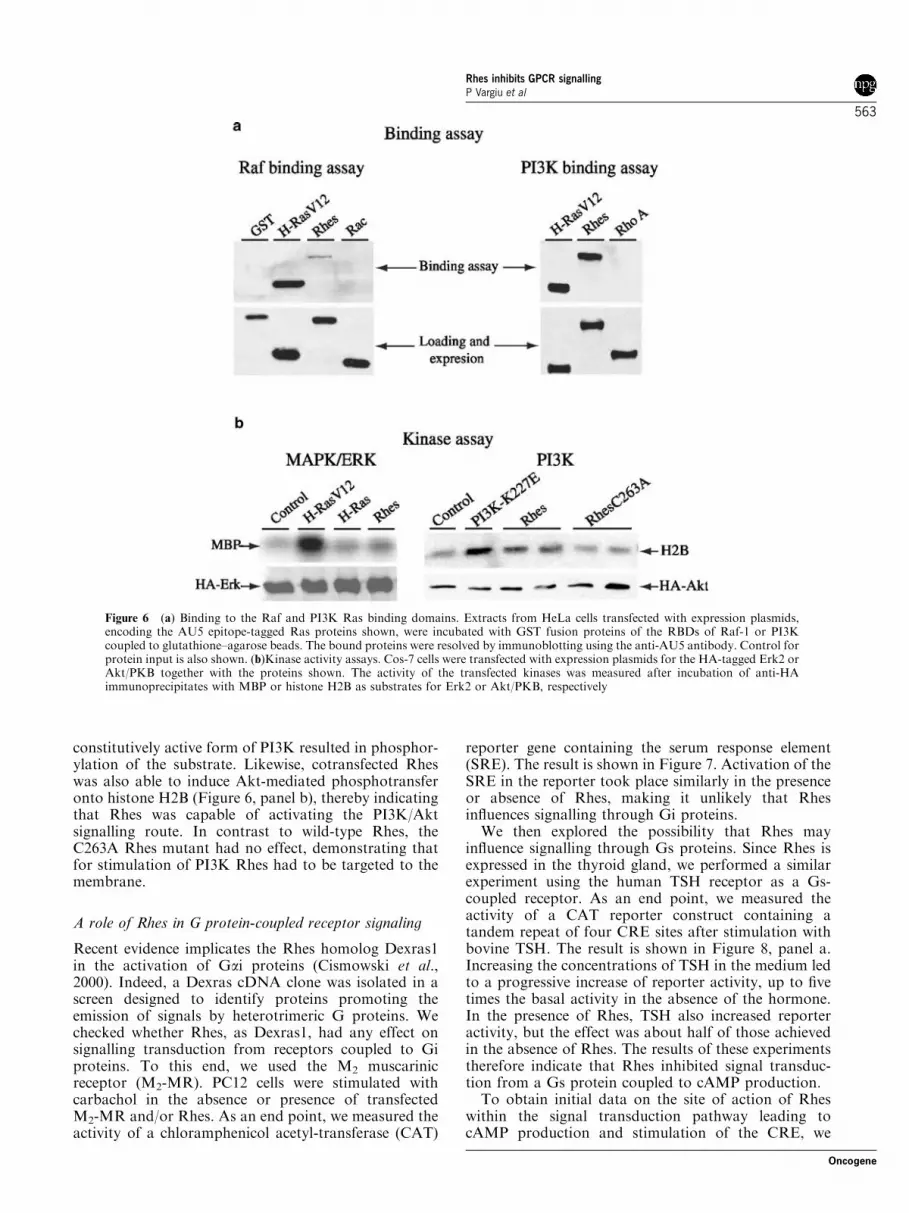
whether Rhes could induce cellular transformation inmurine NIH3T3 fibroblasts. Focus forming assays wereperformed using wild-type Rhes and the N77L mutantof Rhes (equivalent to the Q61L-activating mutation inH-Ras), and as a positive control H-Ras V12. Figure 5shows that in contrast to the RasV12 (4274 foci), thenumber of foci produced with wild-type Rhes (872) orwith the N77L mutant (672) was similar to thatobtained with the vector alone (972).
Rhes displays low affinity for Raf and does not activatethe ERK pathway
Our next goal was to analyse whether Rhes couldactivate the typical Ras effector pathways. We firstanalysed whether Rhes was capable of binding to Rafand thereby, regulating the activation of the ERKpathway. To this end, we performed an in vitro bindingassay using a bacterially expressed GST-fusion proteinof the Raf1 Ras-binding domain (RBD). For thebinding assays, we used protein extracts from trans-fected HeLa cells expressing AU5-tagged vectors en-coding Rhes, H-RasV12 as a positive control, and Rac1as negative control. After GST pull-down and immuno-blotting with anti AU5, we found that RasV12, but notRac1, displayed Raf1 binding as expected, (Figure 6,panel a). In comparison to RasV12, Raf-RBD couldonly retrieve a very faint band of Rhes, suggestive of lowbinding affinity and, therefore, that Raf1 was not alikely Rhes effector. Direct examination of ERK acti-vation confirmed this conclusion. Cos-7 cells were co-transfected with a hemagglutinin (HA)-tagged Erk2together with Rhes, RasV12 as a positive control, H-Raswild type, or the empty vector. ERK activity was sub-sequently assayed in anti-HA immunoprecipitates, usingMBP as a substrate. In cells transfected with Rhes, therewas no increased MBP phosphorylation above the levelobserved with the plasmid alone (Figure 6, panel b).
Rhes binds to and activates phosphoinositide 3-kinase
We also determined whether Rhes was involved in theregulation of the PI3K pathway. To do so, we used thesame methodology as above and we checked for Rhesbinding to the RBD of the PI3K catalytic subunit(p110a) fused to GST. Extracts from cells transfectedwith AU5-tagged Rhes, Ras V12, or RhoA as a negativecontrol were incubated with the GST fusion protein,and the PI3K RBD-bound proteins were resolved byimmunoblotting. The results presented in Figure 6 panela show that, as expected, Rho A did not bind PI3K,whereas a clear binding was obtained for Rhes as well asfor Ras V12. To evaluate the effect of Rhes in PI3Kactivity, we measured the activity of the PI3K targetAkt/PKB (Datta et al., 1996) by transiently expressingan HA-tagged Akt and measuring phosphorylation ofhistone H2B as substrate. Cotransfection with a
Figure 4 Percent nucleotide bound as GTP. Cells were transfectedwith epitope-tagged H-Ras or Rhes and incubated with 32P-orthophosphate in the presence or absence of GEFs. Afterimmunoprecipitation and thin layer chromatography, the amountof 32P[GDP] and 32P[GTP] was quantified and the percent bound asGTP calculated as %GTP¼ 100�GTP/[GTPþGDP]
Figure 5 Rhes is not transforming in murine fibroblasts. NIH 3T3cells were transfected with 0.1mg of RasV12 as a positive control,0.5mg of wild-type Rhes, 0.5 mg Rhes N77L, or 0.5mg of pCEFLplasmid. The number of foci was counted after 2 weeks oftransfection
Rhes inhibits GPCR signallingP Vargiu et al
562
Oncogene
constitutively active form of PI3K resulted in phosphor-ylation of the substrate. Likewise, cotransfected Rheswas also able to induce Akt-mediated phosphotransferonto histone H2B (Figure 6, panel b), thereby indicatingthat Rhes was capable of activating the PI3K/Aktsignalling route. In contrast to wild-type Rhes, theC263A Rhes mutant had no effect, demonstrating thatfor stimulation of PI3K Rhes had to be targeted to themembrane.
A role of Rhes in G protein-coupled receptor signaling
Recent evidence implicates the Rhes homolog Dexras1in the activation of Gai proteins (Cismowski et al.,2000). Indeed, a Dexras cDNA clone was isolated in ascreen designed to identify proteins promoting theemission of signals by heterotrimeric G proteins. Wechecked whether Rhes, as Dexras1, had any effect onsignalling transduction from receptors coupled to Giproteins. To this end, we used the M2 muscarinicreceptor (M2-MR). PC12 cells were stimulated withcarbachol in the absence or presence of transfectedM2-MR and/or Rhes. As an end point, we measured theactivity of a chloramphenicol acetyl-transferase (CAT)
reporter gene containing the serum response element(SRE). The result is shown in Figure 7. Activation of theSRE in the reporter took place similarly in the presenceor absence of Rhes, making it unlikely that Rhesinfluences signalling through Gi proteins.We then explored the possibility that Rhes may
influence signalling through Gs proteins. Since Rhes isexpressed in the thyroid gland, we performed a similarexperiment using the human TSH receptor as a Gs-coupled receptor. As an end point, we measured theactivity of a CAT reporter construct containing atandem repeat of four CRE sites after stimulation withbovine TSH. The result is shown in Figure 8, panel a.Increasing the concentrations of TSH in the medium ledto a progressive increase of reporter activity, up to fivetimes the basal activity in the absence of the hormone.In the presence of Rhes, TSH also increased reporteractivity, but the effect was about half of those achievedin the absence of Rhes. The results of these experimentstherefore indicate that Rhes inhibited signal transduc-tion from a Gs protein coupled to cAMP production.To obtain initial data on the site of action of Rhes
within the signal transduction pathway leading tocAMP production and stimulation of the CRE, we
Figure 6 (a) Binding to the Raf and PI3K Ras binding domains. Extracts from HeLa cells transfected with expression plasmids,encoding the AU5 epitope-tagged Ras proteins shown, were incubated with GST fusion proteins of the RBDs of Raf-1 or PI3Kcoupled to glutathione–agarose beads. The bound proteins were resolved by immunoblotting using the anti-AU5 antibody. Control forprotein input is also shown. (b)Kinase activity assays. Cos-7 cells were transfected with expression plasmids for the HA-tagged Erk2 orAkt/PKB together with the proteins shown. The activity of the transfected kinases was measured after incubation of anti-HAimmunoprecipitates with MBP or histone H2B as substrates for Erk2 or Akt/PKB, respectively
Rhes inhibits GPCR signallingP Vargiu et al
563
Oncogene

evaluated the effect of Rhes on the stimulation of thecAMP pathway by different activators acting atdifferent levels of the pathway. For this task, we useda constitutively acting mutant of the b2-adrenergicreceptor (Samama et al., 1993), the activated Gas QL(Landis et al., 1989) or direct stimulation of PKA withforskolin. The effects of these activators were monitoredby measuring the activity of a CAT reporter gene underthe control of a tandem repeat of four CREs. As shownin Figure 8 panel b, expression of the constitutivelyactive b2-adrenergic receptor led to a marked increaseactivity of the CRE-CAT reporter gene. Interestingly,cotransfection with increasing amounts of Rhes led to adose-dependent inhibition of the receptor-induced re-porter activity. This inhibitory effect of Rhes requiredtargeting to the cell membrane, since Rhes C263Aimpaired for binding to the membrane did not affectthe b-adrenergic receptor effect. In an independentexperiment, we compared the effect of Rhes on theb2-adrenergic receptor with that on a constitutivelyacting Gas protein, and with forskolin (Figure 8, panelc). Expression of GasQL, or addition of forskolin, led toa highly increased activity of the reporter. Again, wild-type Rhes, but not Rhes C263A, inhibited the activationinduced by the b2-adrenergic receptor. However, Rheshad no effect on the activation of the CRE-responsivereporter induced by the activated mutant GasQL orby the addition of forskolin. These experiments indi-cate therefore that Rhes regulates the activation of thecAMP/PKA pathway upstream of the activation of theheterotrimeric G protein complex.
Discussion
In this study, we have provided a first insight into Rhesbiochemical properties. Rhes was initially discovered asan mRNA greatly enriched in the striatum. Rhes is oneof the targets of thyroid hormone during the neonatal
Figure 7 Rhes has no effects on stimulation of the muscarinicreceptor by carbachol. PC12 cells were transfected with expressionplasmids encoding the M2-MR with or without Rhes, and thestimulation of a CAT reporter plasmid harbouring a serumresponse element was measured in response to carbachol
Figure 8 (a) Rhes inhibits signalling by the TSH receptor. PC12cells were transfected with expression plasmids encoding the humanTSH receptor with or without Rhes, and the stimulation of a CATreporter plasmid harbouring four tandem copies of the cAMPresponse element was measured in response to different concentra-tions of bovine TSH. (b) Rhes expression inhibits the activation ofa CRE in response to b2-adrenergic receptor stimulation. Cellswere cotransfected with plasmids encoding a constitutively activereceptor and a CAT reporter gene under control of a tandem repeatof four CRE (4� CRE) sites, with or without increasing amountsof wild-type Rhes, or Rhes harbouring a mutation in the CAAXbox that prevents membrane targeting (RhesC263A). (c) Rhes hasno effect on the stimulation of the CRE-CAT reporter geneinduced by the activated GasQL protein or by forskolin
Rhes inhibits GPCR signallingP Vargiu et al
564
Oncogene

period, the critical time for thyroid hormone-dependentbrain maturation. In addition to the striatum, Rhes isalso expressed in other areas of the brain, such as thecerebral cortex, hippocampus, inferior colliculus, andcerebellum. Expression of Dexras1 also occurs mainly inthe brain, with low levels of expression in other tissues(Fang et al., 2000). The distribution of Dexras1 in brainhas not been reported in detail, but the available dataindicate that it is different from Rhes. In addition topredominant expression in the striatum, Rhes is alsoexpressed in the granular cells of the cerebellum,whereas Dexras1 is expressed in the Purkinje cells.Expression in the striatum and the cerebellum suggeststhat Rhes is involved in pathways related to neuromotorcontrol, which would be in agreement with the striato-pallidal syndrome typical of neurological cretinism(Delong et al., 1985). It is interesting that, among theorgans analysed, Rhes was only found in the thyroidgland, suggesting a role in the modulation of thyroidhormone secretion. This idea is further supported by ourfinding that Rhes modulates the action of TSH throughits receptor. Of considerable interest is the recent report(Chan et al., 2002) that Rhes is also expressed in thepancreas where it may control insulin secretion and be atarget of imidazolin secretagogues.Dexras1 is predominantly cytoplasmic (Fang et al.,
2000); this is in sharp contrast to Rhes. Rhes sequencepresents a conserved CAAX box, CSIQ in rat Rhes andCTIQ in human Rhes. A similar C-terminal sequence ispresent in human Rap2 (CNIQ), which undergoesfarnesylation (Farrell et al., 1993). Likewise, our resultsclearly indicate that Rhes is farnesylated and targeted tothe plasma membrane via this post-translational mod-ification.Interestingly our results indicate that, when compared
to H-Ras, wild-type Rhes contained a high proportionof bound nucleotide in the form of GTP underunstimulated conditions. It is known that singleamino-acid substitutions within Ras residues 116–119,a region that interacts with the guanine ring, result in aconstitutive activation as a result of reduced nucleotideaffinity, resulting in GEF-independent nucleotide ex-change (Clanton et al., 1986; Walter et al., 1986). It isnoteworthy that Rhes maintains the consensus sequenceN-K-X-D (NKND in Rhes) within this region. Thus, areduced affinity for binding to the guanine ring shouldnot account for the high levels of GTP-bound Rhes.Alternatively, this phenomenon could be explained by areduced intrinsic GTPase activity. This may find aprecedent in RhoE, a constitutively active protein thatdisplays no GTPase activity and contains all ofnucleotide bound in the form of GTP (Foster et al.,1996). This property of RhoE is due to amino-acidsubstitutions in positions corresponding to H-Ras 12,59, and 61. Most notably, the corresponding positionsof Rhes and Dexras proteins show similar amino-acidsubstitutions, that is, a serine residue is present inpositions corresponding to residues 12 and 59 of H-Ras,although residue 61 is conservatively replaced byasparagine. These and other amino-acid substitutionsin the corresponding positions 13 (arginine instead of
glycine) and 63 (proline instead of glutamic acid) ofH-Ras, which are also important for GTPase activity,predict that Rhes intrinsic GTPase activity is low.Recent reports suggest that Dexras1 is activated by
nNOS-induced S-nitrosylation in Cys 11 (Jaffrey et al.,2002), a reaction facilitated by Dexras1 binding to theadaptor protein CAPON. Interestingly, H-Ras can beinduced to undergo nucleotide exchange by NO,through a GEF-independent mechanism that involvesthe destabilization of the interaction between residuesin the GDP-binding pocket and the nucleotide, ratherthan the NO-induced modification of cysteine 118 (Mottet al., 1997). Whether Rhes is also subject toS-nitrosylation is not known, but it is possible thatRhes and Dexras1 are subject to the same type ofregulation by nitrogen metabolites. Such a regulatorymechanism may also help to explain the high proportionof Rhes bound to GTP under what we consider basalconditions in our experimental setting. Furthermore,our results demonstrate that Rhes is insensitive to Ras-and Rap-specific GEFs. Although at this stage wecannot rule out that others as yet unidentified GEFscatalyse Rhes GDP/GTP transit, a GEF-independentactivation such as the one proposed is an attractivealternative. In addition, how Rhes responds to GAPproteins GTPase-potentiating activity is another poten-tial level of regulation that has not been explored thusfar, which may shed some light into its regulation.The multiple sequence alignment of Rhes and Dexras
sequences (Graham et al., 2001) shows the conservationof all GTP-binding motives of Ras-like proteins. Theyalso share conserved important residues in the effectorregions compared with the Ras subfamily, like Iso-leucine 52 in rat Rhes (Ile 36 in Human H-Ras) andthe glutamic acid residue in position 53 (37 in humanH-Ras). These last residues are known to be critical forRBD-specific recognition by Ras proteins (Bauer et al.,1999). However, it is important to note that within itseffector domain Rhes exhibits an important substitutionin Thr 49 corresponding to Asp 33 in H-Ras. Thisposition is very important in the Ras family for theinteraction with Raf (Chuang et al., 1994). This agreeswith our findings demonstrating that Rhes had lowaffinity for Raf1 and did not activate the ERK pathway.By contrast, Rhes was able to associate with andactivate PI3K, an interaction which is unaffected bythe substitution at position 49.Finally, we have observed that Rhes has a remarkable
role in GPCR signalling. Rhes interfered with stimula-tion of the cAMP/PKA pathway mediated by the TSHreceptor and by a constitutively active form of theb2-adrenergic receptor. In contrast, it was without effecton a constitutively active Gas or to the direct activationof PKA. These results strongly suggest that Rhes isacting somewhere upstream of the activation of theheterotrimeric complex by the receptor. This would bein agreement with data from Cismowski et al. (1999,2000), Takesono et al. (2002), and Graham et al. (2002)in which a Ras family protein would directly modulatethe activation of a heterotrimeric G protein. Themechanism is not clear. Concerning Dexras1/AGS1, it
Rhes inhibits GPCR signallingP Vargiu et al
565
Oncogene

has been proposed that this protein alters the pool of Gprotein available for receptor interaction, therebyuncoupling the receptor from its effector (Grahamet al., 2002). In our case, we contemplate three hypo-theses. (1) Rhes competes with Gas for binding to thereceptor, and therefore an overexpression of Rhes woulduncouple the receptor from the heterotrimeric complexand preclude its activation. (2) Rhes heterodimerizeswith Gas when GDP-bound precluding it from loadingwith GTP. Rhes would not affect signalling by Gas QLas it is bound to GTP constitutively and would not beable to interact with it. (3) The inhibitory effect wouldnot be directly mediated by Rhes, but by a hithertounidentified kinase activated by the GTPase that wouldphosphorylate and thereby uncouple the b2-adrenergicreceptor from the heterotrimeric complex. Uncouplingby phosphorylation is a common mechanism and hasbeen previously demonstrated in the case of the b2-adrenergic receptor (Daaka et al., 1997). These threepossibilities warrant further investigation.
Materials and methods
Sequence analysis
Sequences belonging to Dexras/Rhes protein group wereobtained by BLAST searches on Swissprot, TREMBL, andTREMBLNEW databases (Bairoch and Apweiler, 2000) usingRhes1 sequence from Rat (EMBLAccession number: Q9WVD3).Ras-related sequences in Swissprot belonging to main subfamilies(Ras, Rho, and Rab) were exhaustively collected using ISS-BLAST (Intermediate Sequence Searches) (Park et al., 1997).Multiple sequences alignment were built with CLUSTALWusing BLOSUM62 matrix (Henikoff and Henikoff, 1992) forRhes and the other Ras sequence subfamilies. Phylogeneticclassification of Rhes sequences into the whole Ras sequenceswere carried out with CLUSTALW using gene tree boot-strapping calculation method (Thompson et al., 1994).
Cell cultures
NIH 3T3 cells were grown in Dulbecco’s modified Eagle’smedium (DMEM) supplemented with 10% calf serum. COS-7and HeLa cells were grown in the same medium. The ratpheochromocytoma cell line, PC12, was cultured in DMEMsupplemented with 5% foetal calf serum (FCS) and 10% horseserum (HS). For focus forming assays, 1.5� 105 NIH3T3 cellswere plated the day before transfection onto a 100mm culturedish. DNA transfection was performed using Lipofectamine Plusreagent (Life Technologies, Rockville, MD, USA), as recom-mended by the manufacturer. After transfection, the mediumwas changed twice a week, and the number and quality of fociwere scored every week. The plates were fixed in 100% coldmethanol and stained with 10% Giemsa (Merk) in phosphate-buffered saline (PBS). Other cells used for studies of Rhesexpression included N2a neuroblastoma cells (Americal TypeCell Culture Collection, CCL 131), hypothalamic GT1-7 cells(Mellon et al., 1990), C6 glioma cells, and ‘P’ cells, immortalizedneurons from E18 embryonic rat brain (Munoz et al., 1993).
CAT activity assays
Cells were transfected with CAT reporter constructs usingLipofectamine. As an internal control for transfection
efficiency, we used the plasmid pCH110 (Pharmacia LKB)encoding b-galactosidase under control of SV-40 earlypromoter. At 36 h after transfection, the cell extracts wereincubated with 25 nCi of [14C]chloramphenicol and 800 mMacetyl-CoA, in a final volume of 100ml of 0.25M Tris HCl, pH7.5 at 371C for 2 h. The reaction products were extracted with550 ml of ethyl acetate and fractionated by thin-layer chroma-tography on silicagel plates followed by autoradiography.Quantifications were performed after exposure of the plates inan Instant Imager electronic detector (Packard).
GTP/GDP binding assay
Cells were transfected with 1 mg each of H-RasV12, Rhes andH-Ras, in the PCEFL expression vector containing the AU5epitope for immunoprecipitation. The cells were kept for 48 hin DMEM supplemented with 10% FCS, and then incubatedovernight with phosphate-free DMEM (Sigma-Aldrich, StLouis, MO, USA). The cells were incubated with 0.05mCi/mlof 32P-orthophosphate for 6 h, and the GTP-binding proteinwas immunoprecipitated with anti-AU5 antibody at a final1 : 500 dilution (BabCo) and incubated with 20 ml of g-bindG-Sepharose (Pharmacia) for 15min on ice. The immunopre-cipitates were washed three times with cold buffer (50mM
TrisHCl, pH 7.5, 20mM MgCl2, 500mM NaCl, 0.5% NonidetP-40), and the bound nucleotides were released by heating at681C for 2min. Eluted nucleotides were analysed by PEIcellulose thin-layer chromatography (Merck) and the propor-tion of GTP-bound over total GTPþGDP was calculated.The data from different experiments were pooled to calculatestatistical significance by ANOVA. The calculations wereperformed using the SPSS statistical package (SPSS Inc.,Chicago, IL, USA).
Intracellular localization
PC12 cells were plated at low density on poly-L-lysine-coatedcoverslips. They were then transfected with either GFP-expression vectors containing the constructs of interest, orpCEFL expression vectors containing the AU5 epitope, usingLipofectamine in serum-free medium (Optimem 1, GibcoBRL) for 5 h. After washing, the cells were maintained ingrowth medium for 24 h. Cells were rinsed in PBS, fixed in3.7% formaldehyde for 15min, washed with PBS, andmounted in Mowiol for direct fluorescence of GFP. Forindirect immunofluorescence, the cells were fixed in 4%paraformaldehyde, washed with 0.1M glycine in PBS for15min and then permeabilized with 0.5% Triton X-100 in0.1M glycine in PBS for 15min and blocked with 3% BSA inPBS for 30min. Incubation with the mouse anti-AU5 antibodyat a 1 : 100 dilution (BAbCo) was for 1 h at room temperature.After rinsing three times with PBS, the cells were stained withFITC-conjugated goat anti-mouse IgG (Jackson ImmunoResearch Laboratories, 1 : 100 dilution) at room temperaturein the dark. After washing, the coverslips were mounted withMowiol and viewed in a confocal microscope. The effect offarnesylation or geranylgeranylation inhibitors was assessed bytreatment of the cells with FTI-277 or GGTI-298 (Calbio-chem), respectively, as described (Lerner et al., 1995a, b).
In vitro binding assays
The Raf(amino acids 1–149) and p110 subunit of PI3K (aminoacids 127–314) RBDs, in the expression vector pGEX, wereprepared from 400ml of overnight Escherichia coli culture andcoupled to 300 ml of glutathione–agarose beads (Pharmacia–Biotech). HeLa cells were transfected with expression vectorsencoding Ras proteins, and 400ml of cell lysate (lysis buffer
Rhes inhibits GPCR signallingP Vargiu et al
566
Oncogene

was 25mM HEPES pH 7.3, 150mM NaCl, 10mM MgCl2 10,0.5mM EGTA, 0.5% Triton X-100, 4% glycerol, 100 mg/mlPMSF, 10mM NaF, 20mM b-glycerol phosphate, 10mM
Na3VO4, and 2mg/ml each of leupeptin and aprotinin) wasincubated with 30mg of GST fusion proteins for 3 h at 41C in arotating wheel. After pelleting the beads, the proteins weresolubilized and immunoblotted using the anti-AU5 antibody.
Kinase activity assays
To evaluate the effect of Rhes on the MAP kinase pathway,approximately 1� 106 COS-7 cells were transfected bystandard calcium phosphate precipitation with 1.0 mg of anHA-tagged MAPK/Erk2 expression plasmid together witheither 3 mg of wild-type Rhes, 3 mg of H-Ras, 1mg of H-RasV12, or 3 mg of H-Ras N17 in the expression plasmid pCEFL.At 48 h after transfection, the cells were starved in serum-free medium for 16 h and subsequently lysed in 400 ml of20mM HEPES (pH 7.5), 10mM EGTA, 40mM b-glycerolphosphate, 1% NP-40, 2.5mM MgCl2, 2mM Na3VO4, 1mM
DTT, 100mg/ml PMSF, 2mg/ml leupeptin, 2mg/ml aprotinin.Exogenously expressed Erk2 was immunoprecipitated fromthe total cell lysate using an anti-HA antibody for 1 h at 41C,followed by coupling 15 ml of g-bind G-Sepharose (Pharmacia)for 20min at 41C. Immune complexes were washed three timeswith 1% NP-40 and 2mM Na3VO4 in PBS, followed by onewash in 5mM LiCl, 100mM Tris (pH 7.5), and one wash inkinase reaction buffer (12.5mM b-glycerol phosphate, 7.5mM
MgCl2, 0.5mM EGTA, 0.5mM NaF, 0.5mM Na3VO4, 1mMDTT, 12.5mM MOPS pH 7.5). Kinase activity assay wasperformed in 30ml of reaction buffer, containing 1.0 mCi of[g32P]ATP, 20 mM unlabelled ATP, 1mM DTT, and 40 mg ofmyelin basic protein (Sigma, MO, USA), and incubated at301C for 30min. Aliquots (30 ml) of the reaction mixture weresubjected to 14% acrylamide SDS gel electrophoresis.To evaluate the effect of Rhes on the PI3 kinase pathway,
3.0mg of expression plasmid was cotransfected with 1.0 mg ofHA–Akt expression plasmid in COS–7 cells. Approximately48 h after transfection, cells were starved in serum-depletedmedium for 16 h and subsequent cell lysis and kinase reactionwere performed. For Akt protein kinase activity assay, cellswere lysed in NP-40 lysis buffer (1% NP-40, 10% glycerol,150mM NaCl, 20mM Tris pH 7.4, containing 2 mg/mlaprotinin, 2 mg/ml leupeptin, 100mg/ml PMSF, 20mM NaFand 1mM Na3VO4). Immunoprecipitation was carried out for
2 h at 41C with rotation, using an anti-HA antibody followedby coupling 20 ml of g-bind G-Sepharose (Pharmacia) for 1 h at41C with rotation. The immunoprecipitates were washed threetimes with lysis buffer, once with water, and once with the Aktkinase buffer (20mM HEPES, pH 7.4, 10mM MgCl2, 10mM
MnCl2. Kinase assay was performed in 30ml of Akt kinasebuffer, containing 3mCi of g32P-ATP, 20M unlabelled ATP,and 3mg of histone H2B), and incubated at 301C for 30min.Equal aliquots (30 ml) of the reaction mixture were subjected to16% polyacrylamide–SDS gel electrophoresis.
In situ hybridization
In situ hybridization was performed as described previously(Vargiu et al., 2001). Briefly, 25 mm thick coronal and saggitalslices were obtained in a cryostat from paraformaldehyde-fixedadult rat brains. The slices were hybridized using a free floatingtechnique with a Rhes riboprobe synthesized from nucleotides1431–1869 of Rhes cDNA in the presence of [35S]UTP. Afterhybridization and further processing of the slices as described,the slices were mounted on glass slides for autoradiography.
Activation of the cAMP pathway
The cAMP pathway was activated in PC12 cells by transfec-tion with expression vectors encoding a constitutively activatedb2-adrenergic receptor (Samama et al., 1993), the activatedGas mutant, GasQL (Landis et al., 1989; Crespo et al., 1995;Vargiu et al., 2001), or by 10mM forskolin. In anotherexperiment, the cells were stimulated with bovine TSH(Sigma-Aldrich Co., St Louis, MO, USA) after transfectionwith the human TSH receptor (kindly supplied by Dr GilbertVassart, Free University of Brussels, Belgium). The cells werecotransfected with a CAT reporter under control of a tandemrepeat of four cAMP responsive elements (CRE) and CATassays were performed as described above.
Acknowledgements
We acknowledge the technical help provided by EulaliaMoreno. Supported by Grants 08.5/0044/2000 1 from theCommunity of Madrid, BFI2002-00489 from the Ministry ofScience and Technology, European Union Q663-2000-00930and FIS, Instituto de Salud Carlos III, Red de Centros RCMN(C03/08), Madrid, Spain.
References
Bairoch A and Apweiler R. (2000). Nucleic Acids Res., 28,45–48.
Bauer B, Mirey G, Vetter IR, Garcia-Ranea JA, Valencia A,Wittinghofer A, Camonis JH and Cool RH. (1999). J. Biol.Chem., 274, 17763–17770.
Campbell SL, Khosravi-Far R, Rossman KL, Clark GJ andDer CJ. (1998). Oncogene, 17, 1395–1413.
Chan SL, Monks LK, Gao H, Deaville P and Morgan NG.(2002). Br. J. Pharmacol., 136, 31–36.
Chuang E, Barnard D, Hettich L, Zhang XF, Avruch J andMarshall MS. (1994). Mol. Cell. Biol., 14, 5318–5325.
Cismowski MJ, Ma C, Ribas C, Xie X, Spruyt M, Lizano JS,Lanier SM and Duzic E. (2000). J. Biol. Chem., 275,23421–23424.
Cismowski MJ, Takesono A, Ma C, Lizano JS, Xie X,Fuernkranz H, Lanier SM and Duzic E. (1999). Nat.Biotechnol., 17, 878–883.
Clanton DJ, Hattori S and Shih TY. (1986). Proc. Natl. Acad.Sci. USA, 83, 5076–5080.
Crespo P, Cachero TG, Xu N and Gutkind JS. (1995). J. Biol.Chem., 270, 25259–25265.
Daaka Y, Luttrell LM and Lefkowitz RJ. (1997). Nature, 390,88–91.
Datta K, Bellacosa A, Chan TO and Tsichlis PN. (1996).J. Biol. Chem., 271, 30835–30839.
Delong GR, Stanbury JB and Fierro-Benıtez R. (1985).Develop. Med. Child Neurol., 27, 317–324.
Falk JD, Vargiu P, Foye PE, Usui H, Perez J, Danielson PE,Lerner DL, Bernal J and Sutcliffe JG. (1999). J. Neurosci.Res., 57, 782–788.
Fang M, Jaffrey SR, Sawa A, Ye K, Luo X and Snyder SH.(2000). Neuron, 28, 183–193.
Farrell FX, Yamamoto K and Lapetina EG. (1993). Biochem.J., 289 (Part 2), 349–355.
Rhes inhibits GPCR signallingP Vargiu et al
567
Oncogene

Foster R, Hu KQ, Lu Y, Nolan KM, Thissen J and SettlemanJ. (1996). Mol. Cell. Biol., 16, 2689–2699.
Graham TE, Key TA, Kilpatrick K and Dorin RI. (2001).Endocrinology, 142, 2631–2640.
Graham TE, Prossnitz ER and Dorin RI. (2002). J. Biol.Chem., 277, 10876–10882.
Henikoff S and Henikoff JG. (1992). Proc. Natl. Acad. Sci.USA, 89, 10915–10919.
Jaffrey SR, Fang M and Snyder SH. (2002). Chem. Biol., 9,1329–1335.
Kemppainen RJ and Behrend EN. (1998). J. Biol. Chem., 273,3129–3131.
Landis CA, Masters SB, Spada A, Pace AM, Bourne HR andVallar L. (1989). Nature, 340, 692–696.
Lerner EC, Qian Y, Blaskovich MA, Fossum RD, Vogt A,Sun J, Cox AD, Der CJ, Hamilton AD and Sebti SM.(1995a). J. Biol. Chem., 270, 26802–26806.
Lerner EC, Qian Y, Hamilton AD and Sebti SM. (1995b).J. Biol. Chem., 270, 26770–26773.
Mellon PL, Windle JJ, Goldsmith PC, Padula CA, Roberts JLand Weiner RI. (1990). Neuron, 5, 1–10.
Mott HR, Carpenter JW and Campbell SL. (1997). Biochem-istry, 36, 3640–3644.
Munoz A, Wrighton C, Seliger B, Bernal J and Beug H. (1993).J. Cell Biol., 121, 423–438.
Park J, Teichmann SA, Hubbard T and Chothia C. (1997).J. Mol. Biol., 273, 349–354.
Reuther GW and Der CJ. (2000). Curr. Opin. Cell. Biol., 12,157–165.
Samama P, Cotecchia S, Costa T and Lefkowitz RJ. (1993).J. Biol. Chem., 268, 4625–4636.
St Croix B, Rago C, Velculescu V, Traverso G, Romans KE,Montgomery E, Lal A, Riggins GJ, Lengauer C, VogelsteinB and Kinzler KW. (2000). Science, 289, 1197–1202.
Takesono A, Nowak MW, Cismowski M, Duzic E and LanierSM. (2002). J. Biol. Chem., 277, 13827–13830.
Thompson JD, Higgins DG and Gibson TJ. (1994). NucleicAcids Res., 22, 4673–4680.
Vargiu P, Morte B, Manzano J, Perez J, de Abajo R, GregorSutcliffe J and Bernal J. (2001). Brain Res. Mol. Brain Res.,94, 1–8.
Walter M, Clark SG and Levinson AD. (1986). Science, 233,649–652.
Wittinghofer A and Valencia A. (1995). Guidebook to the SmallGTPases, Zerial M, Huber LA, Tooze J (eds). OxfordUniversity Press: Oxford, New York, Tokyo, pp. 20–29.
Rhes inhibits GPCR signallingP Vargiu et al
568
Oncogene




















