
A protein transduction domain located at the NH 2-terminus of human translationally controlled tumor...
9
A protein transduction domain located at the NH 2 -terminus of human translationally controlled tumor protein for delivery of active molecules to cells Moonhee Kim, Miyoung Kim, Hyo Young Kim, Sol Kim, Jaehoon Jung, Jeehye Maeng, Jun Chang, Kyunglim Lee * College of Pharmacy, Center for Cell Signaling & Drug Discovery Research, Ewha Womans University, Seoul, Republic of Korea article info Article history: Received 5 August 2010 Accepted 27 August 2010 Available online 22 September 2010 Keywords: Adenovirus Endocytosis Protein transduction domain TCTP abstract Protein transduction domains (PTDs) are small peptides, able to penetrate biological membranes and deliver various types of cargo both in vitro and in vivo. Because use of PTDs originating from viral origins resulted in undesired effects, PTDs originating from non-viral origins are needed. Here, we report that a 10-amino acid peptide (MIIYRDLISH) derived from the NH 2 -terminus of human translationally controlled tumor protein (TCTP) functions as a PTD. This peptide was internalized through lipid raft- dependent endocytosis and partial macropinocytosis, and did not enter lysosome and nucleus. Beta- galactosidase fused to TCTP-PTD, when injected into mice, was efficiently delivered to liver, kidney, spleen, heart, and lungs of the animals. Preincubation of TCTP-PTD with adenovirus increased adenoviral mediated-gene expression in cells and also improved immune response to intranasally administered adenovirus expressing the triple repeat of G glycoprotein of respiratory syncytial virus (RSV), rAd/3G. These findings suggest that TCTP-PTD might overcome the limitations of polycation-mediated trans- duction and serve as an efficient vehicle for drug delivery. Ó 2010 Elsevier Ltd. All rights reserved. 1. Introduction Many protein transduction domains (PTDs), alternatively named cell penetrating peptides (CPPs) have been used to deliver thera- peutic cargo, because of their cell penetration ability. PTDs have been categorized into three groups according to their origin [1]: first group derived from natural proteins such as TAT of the human immunodeficiency virus (HIV), and penetratin of Drosophila Antennapedia (Antp), second group are those modified from natural peptides, such as transportan [2] and synB [3] and the third group are those identified by screening random phage display libraries. Regardless of their origin, all PTDs share some features, including an arginine and lysine rich motif, a hydrophobic core sequence, and an amphipathicity or a-helix. PTDs can transfer a variety of cargos, ranging from small drug molecules to large proteins and nucleic acids, and have therefore been applied as effective vehicles [4,5]. For example, administra- tion of beta-galactosidase fused to TAT to mice resulted in efficient transduction to all tissues, even across the blood-brain-barrier (BBB) [6]. The cross-linking of TAT to siRNA increased cellular uptake of the oligonucleotide without interfering with the peri- nuclear localization required for RNAi activity [7]. However, several studies of transduction of oligonuceotides with PTD-siR- NAs revealed that TAT and penetratin peptide itself affected gene expression [8], underscoring the need for PTDs that do not cause undesirable changes in the organism under study. We identified a PTD derived from the NH 2 terminus of human TCTP (TCTP-PTD) with the sequence, ‘MIIYRDLISH’. TCTP is consid- ered to be a house keeping gene because of its ubiquitous expression, high degree of conservation among species, and occurence in both tumor and normal cells [9]. Human TCTP consists of 172 amino acids and the tertiary structure of fission yeast TCTP shows a possible relationship with Mss4/Dss4 chaperone family [10]. TCTP is known to be involved in human allergic response [11e 13], apoptotic regu- lation [14e16], various cancer-related functions [17e19], and hypertension [20], but its mechanism of the action still remains elusive. We found that this cell penetrating peptide differs from other well-known PTDs in its amino acid sequence and structure, has the ability to internalize into the cells, and translocate cargo in vitro and in vivo. We characterized the mechanism by which TCTP-PTD internalizes into cells and also examined its useful- ness as transduction vehicle using adenoviral gene delivery system. * Corresponding author. Tel.: þ82 2 3277 3024. E-mail address: [email protected] (K. Lee). Contents lists available at ScienceDirect Biomaterials journal homepage: www.elsevier.com/locate/biomaterials 0142-9612/$ e see front matter Ó 2010 Elsevier Ltd. All rights reserved. doi:10.1016/j.biomaterials.2010.08.077 Biomaterials 32 (2011) 222e230
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of A protein transduction domain located at the NH 2-terminus of human translationally controlled tumor...

lable at ScienceDirect
Biomaterials 32 (2011) 222e230
Contents lists avai
Biomaterials
journal homepage: www.elsevier .com/locate/biomateria ls
A protein transduction domain located at the NH2-terminus of humantranslationally controlled tumor protein for delivery of active molecules to cells
Moonhee Kim, Miyoung Kim, Hyo Young Kim, Sol Kim, Jaehoon Jung, Jeehye Maeng, Jun Chang,Kyunglim Lee*
College of Pharmacy, Center for Cell Signaling & Drug Discovery Research, Ewha Womans University, Seoul, Republic of Korea
a r t i c l e i n f o
Article history:Received 5 August 2010Accepted 27 August 2010Available online 22 September 2010
Keywords:AdenovirusEndocytosisProtein transduction domainTCTP
* Corresponding author. Tel.: þ82 2 3277 3024.E-mail address: [email protected] (K. Lee).
0142-9612/$ e see front matter � 2010 Elsevier Ltd.doi:10.1016/j.biomaterials.2010.08.077
a b s t r a c t
Protein transduction domains (PTDs) are small peptides, able to penetrate biological membranes anddeliver various types of cargo both in vitro and in vivo. Because use of PTDs originating from viral originsresulted in undesired effects, PTDs originating from non-viral origins are needed. Here, we report thata 10-amino acid peptide (MIIYRDLISH) derived from the NH2-terminus of human translationallycontrolled tumor protein (TCTP) functions as a PTD. This peptide was internalized through lipid raft-dependent endocytosis and partial macropinocytosis, and did not enter lysosome and nucleus. Beta-galactosidase fused to TCTP-PTD, when injected into mice, was efficiently delivered to liver, kidney,spleen, heart, and lungs of the animals. Preincubation of TCTP-PTD with adenovirus increased adenoviralmediated-gene expression in cells and also improved immune response to intranasally administeredadenovirus expressing the triple repeat of G glycoprotein of respiratory syncytial virus (RSV), rAd/3�G.These findings suggest that TCTP-PTD might overcome the limitations of polycation-mediated trans-duction and serve as an efficient vehicle for drug delivery.
� 2010 Elsevier Ltd. All rights reserved.
1. Introduction
Many protein transduction domains (PTDs), alternatively namedcell penetrating peptides (CPPs) have been used to deliver thera-peutic cargo, because of their cell penetration ability. PTDs havebeen categorized into three groups according to their origin [1]:first group derived from natural proteins such as TAT of the humanimmunodeficiency virus (HIV), and penetratin of DrosophilaAntennapedia (Antp), second group are those modified fromnatural peptides, such as transportan [2] and synB [3] and the thirdgroup are those identified by screening random phage displaylibraries. Regardless of their origin, all PTDs share some features,including an arginine and lysine rich motif, a hydrophobic coresequence, and an amphipathicity or a-helix.
PTDs can transfer a variety of cargos, ranging from small drugmolecules to large proteins and nucleic acids, and have thereforebeen applied as effective vehicles [4,5]. For example, administra-tion of beta-galactosidase fused to TAT to mice resulted in efficienttransduction to all tissues, even across the blood-brain-barrier(BBB) [6]. The cross-linking of TAT to siRNA increased cellular
All rights reserved.
uptake of the oligonucleotide without interfering with the peri-nuclear localization required for RNAi activity [7]. However,several studies of transduction of oligonuceotides with PTD-siR-NAs revealed that TAT and penetratin peptide itself affected geneexpression [8], underscoring the need for PTDs that do not causeundesirable changes in the organism under study.
We identified a PTD derived from the NH2 terminus of humanTCTP (TCTP-PTD) with the sequence, ‘MIIYRDLISH’. TCTP is consid-ered to be a house keeping gene because of its ubiquitous expression,high degree of conservation among species, and occurence in bothtumor and normal cells [9]. Human TCTP consists of 172 amino acidsand the tertiary structure of fission yeast TCTP shows a possiblerelationship with Mss4/Dss4 chaperone family [10]. TCTP is knownto be involved in human allergic response [11e13], apoptotic regu-lation [14e16], various cancer-related functions [17e19], andhypertension [20], but its mechanism of the action still remainselusive.
We found that this cell penetrating peptide differs from otherwell-known PTDs in its amino acid sequence and structure, hasthe ability to internalize into the cells, and translocate cargo invitro and in vivo. We characterized the mechanism by whichTCTP-PTD internalizes into cells and also examined its useful-ness as transduction vehicle using adenoviral gene deliverysystem.

M. Kim et al. / Biomaterials 32 (2011) 222e230 223
2. Materials and methods
2.1. Cell culture
HeLa, NIH-3T3 and MDCK cell lines were obtained from American Type CultureCollection (ATCC) and maintained in Dulbecco’s minimal essential media (DMEM)supplemented with 10% fetal bovine serum (FBS), 100 units/ml penicillin, and100 units/ml streptomycin. U-937 cell lines also obtained from ATCC were grown inRoswell Park Memorial Institute (RPMI)-1640 Medium supplemented with 10% FBS,100 units/ml penicillin, 100 units/ml streptomycin. Human bronchial epithelial cells,BEAS-2B (ATCC), were maintained in bronchial epithelial growth medium (BEGM,Clonetics, San Diego).
2.2. Peptide synthesis
Highly pure peptides were obtained from Peptron (Daejeon, Korea). Amino-terminals of the peptides were fluorescent labeled with 5-(6)-tetramethylrhod-amine (TAMRA), protecting carboxy-terminals by amidation. Freeze-dried peptideswere reconstituted in high-purity DMSO (10 mM) and stored at �70 �C until use.
2.3. Protein expression
TAT regions of pTAT-HA-LacZ vector (kindly provided by S. F. Dowdy, UCSD, CA,USA) were removed or substituted by TCTP-1-10. These plasmids were expressed inE.coli BL21(DE3) pLysS (Novagen) and purified on a His binding column of nickel-nitrilotriacetic acid (Niþ-NTA) agarose resin (Qiagen). After replacing the buffer withPBS using PD-10 column, proteins were quantitatedwith Bradford Protein Assay Dye(Bio-Rad). Purity of proteins was examined with Coomassie staining by SDS-PAGEand the activity of beta-galactosidase was assessed with 15 mM O-nitrophenyl-b-D-galtopyranoside (ONPG) dissolved in 0.1 M Na.Pi (pH 7.4) buffer. The absorbance wasmeasured at 405 nm with ELISA microplate reader. For preparation of intact andtruncated TCTPs, Escherichia coli BL21(DE3)pLysS cells transformed with the pRSETA/TCTP or pRSET A/deleted TCTPs, were overexpressed and purified as previouslydescribed [13].
2.4. Peptide and protein uptake assay
1.0 � 106 cells/well were seeded onto 6-well plates 24 h before incubation withpeptides or proteins. The cells were then washed with serum-free media andincubated with peptides or proteins in serum-free media. To study the effect of lowtemperature on the cellular internalization of peptides, HeLa cells werewashedwithserum-free DMEM and preincubated in serum-free DMEM for 1 h at 4 �C. The cellswere incubated with 10 mM peptide in serum-free DMEM for 2 h at 4 �C. For energydepletion experiments, HeLa cells were washed with serum free DMEM and pre-incubated in serum- and glucose-free DMEM containing 6 mM 2-deoxy-D-glucose(Sigma) and 10 mM sodium azide (Sigma) for 1 h at 37 �C. The cells were incubatedwith 10 mM peptide in the same DMEM for 2 h at 37 �C. To study the effect of specificendocytosis inhibitors (all were obtained from Sigma), HeLa cells were washed withserum-free DMEM and pretreated for 30 m at 37 �C with 0e15 mM chlorpromazine(CPZ), 0e5mMmethyl-b-cyclodextrin (MbCD), 0e10mM EIPA, 200 mM chloroquine or25 mg/ml heparin. Then the cells were incubated with the 10 mM peptide in serum-free DMEM containing each inhibitor for 2 h at 37 �C. Following the above proce-dures, the cells were washed two times with PBS and immediately trypsinized(1 mg/ml) for 10 min at 37 �C to remove extracellular peptides or proteins and thenwashed two times with PBS. Analysis of peptide internalization was performed byflow cytometry (Becton Dickinson). Flow cytometric analyses were accomplishedusing WinMDI version 2.8 software (free ware).
2.5. Western blotting
When BEAS-2B cells became 80% confluent, they were passaged in 48-wellculture plates, incubated for 20e24 h in bronchial epithelial basal medium (BEBM),washed twice with 1% penicillin-streptomycin/BEGM, and then incubated for 24 hwith 5 mg/ml or 1 M of proteins in 1% penicillin-streptomycin/BEBM. The superna-tants were saved for western blotting, and the pellets were washed twice with PBSand treated with 20 mM of trypsin at 37 �C for 10 min before denaturing in a samplebuffer containing AEBSF (50 mM Tris, pH 6.8, 2.5% SDS, 22% glycerol, 5% b-mercap-toethanol, 1 mM AEBSF). The resulting supernatants and cell extracts were subjectedto western blotting.
2.6. Confocal microscopy
Cells (1.0 � 105/well) were seeded on to glass cover slips 24 h before incubationwith peptides or proteins. After washing with serum-free DMEM, the cells weretreated with 10 mM TAMRA-labeled peptide for 2 h at 37 �C. For subcellular locali-zation of TCTP-PTD peptide in live cells, the cells were coincubated with fluores-cence markers (Mito-Tracker, Lyso-Tracker, Alexa flour 488 labeled trasferrin,Oregon green-labeled neural dxtran-70 kD, Alexa flour 488 labeled cholera toxin B)in serum-free DMEM for 1 h at 37 �C. All fluorescence markers were obtained from
Molecular Probe. The cells were washed and mounted in phenol red-free DMEM(JBI) and immediately visualized by confocal microscopy (Carl Zeiss).
2.7. Beta-galactosidase staining
4e6 weeks-old Balb/c mice were injected intraperitoreally with 400 mg of b-galcontrol protein and TCTP-PTD/b-gal protein in 0.5ml of PBS. After 2 or 4 h, miceweresacrificed. Tissues were harvested and washed with PBS, fixed in 0.25% glutaralde-hyde for 1 h, and developed overnight in 0.2% X-gal staining solution. All animalprocedures were performed in accordance with institutional guidelines.
2.8. Infection of the cells
For the preparation of adenovirus complex with a TCTP-PTD, adenovirus wasincubated with TCTP-PTD peptide in 100 ml of serum-free DMEM for 30 min at roomtemperature. 6.0 � 104 cells/well were seeded on to 24-well plates 24 h beforeinfection and washed with serum-free DMEM. The complex was added to the cells,left to incubate for 4 h and changed with 10% serum containing DMEM for 24 h.
2.9. Immunization
All animal experiments were performed according to the National Institute ofHealth Publication No. 8523: Guide for Care and Use of Laboratory Animals(Accession No. ELAGC-08-1021). 6- to 8-week-old female BALB/c mice were keptunder specific-pathogen free conditions. For immunization, mice were lightlyanesthetized by ether/chloroform inhalation and intranasally administered5�106 PFU of replication-defective adenovirus complex with TCTP-PTD in a volumeof 70 ml. For measuring total IgG titer, after 2 weeks of infection, blood was obtainedfrom the retro-orbital plexus with a heparinized capillary tube and centrifuged, andserum was stored at �20 �C. For measuring secretory IgA titer, bronchoalveolarlavage (BAL) fluid was obtained from a lung airway. RSV G protein-specific antibodytiters in immunizedmiceweremeasured by a direct enzyme-linked immunosorbentassay (ELISA) using horseradish peroxidase-conjugated affinity-purified rabbit anti-mouse total IgG or IgA secondary antibody (Zymed Laboratoris).
2.10. Toxicity of TCTP-PTD in mice
TCTP-1-10 peptide was intravenously injected into five mice (1.6 mg/kg) everyday for 2 weeks. To assess liver and kidney toxicity, levels of aspartate amino-transferase (AST), alanine aminotrasferase (ALT), blood urea nitrogen (BUN) andcreatinine in the blood were determined and body weight was monitored.
2.11. Statistical analysis
Data are presented as mean � SD. Comparison of differences was conducted byusing an unpaired, two-tailed Student t-test. The difference was considered statis-tically significant when the P value was �0.05.
3. Results
3.1. Identification of a protein transduction domain
We fortuitously observed the translocation of TCTP into the RBL-2H3 cell line while performing confocal microscopic experimentsusing purified full length TCTP. Based on this fortuitous finding, wepostulated that TCTP contains a PTD in its sequence. Whileattempting to locate the PTD present in TCTP protein, we con-structed protein expression vectors of full length TCTP as well theseveral amino terminal truncated proteins, containing only theindicated amino acids residues: TCTP-11-172 and TCTP-35-172.We also purified other truncated derivatives of TCTP includingTCTP-1-38, TCTP-39-110 and TCTP-111-172. In BEAS-2B cells, after24 h incubationwith full length TCTP and the truncated derivatives,TCTP-11-172 and TCTP-35-172, we harvested the supernatants andthen subjected them to western blotting using anti-TCTP antibody.We noted that the full length TCTP disappeared from the super-natant, but the amino terminal truncated proteins, TCTP-11-172and TCTP-35-172, remained in BEAS-2B cell supernatants, sug-gesting that amino-terminus of TCTP is necessary for internalizinginto cells (Fig. 1A). In the next set of experiments, different typesof truncated derivatives, TCTP-1-38, TCTP-39-110 and TCTP-111-172 were treated for 5 min and 30 min in BEAS-2B cells. Asshown in Fig. S1, only TCTP-1-38 disappeared rapidly from the cell

Fig. 1. Identification of a novel PTD derived from human TCTP. (A) Domain mapping of PTD of TCTP by western blotting in supernatant of BEAS-2B cells. (B) Intracellular localizationof TCTP-PTD peptides in HeLa cells. HeLa cells were incubated with various concentrations of TAMRA-labeled TCTP-PTD peptide for 2 h and intracellular fluorescence was analyzedby confocal microscopy. (C) Dose-dependent transduction efficiency in various cell lines including U-937, NIH-3T3 and MDCK cells. After incubation with various amounts of TCTP-PTD for 2 h, intracellular fluorescence of the cells was analyzed by FACS.
M. Kim et al. / Biomaterials 32 (2011) 222e230224
supernatant. In addition, TCTP-1-38 internalized into the cellinterior in a time-dependent manner while the TCTP-39-110 andTCTP-111-172 remained in the supernatants. Taken together, thesedata strongly indicated that the first 10 amino acids of NH2-terminus of TCTP (MIIYRDLISH) enable this protein cross the cellmembranes, acting as a PTD domain. To determine whether TCTP-PTD alone can enter cells, we synthesized 5-(6)-tetramethylrhod-amine (TAMRA)-labeled peptide corresponding TCTP-1-10 andadded various doses of this peptide to the cells. TCTP-PTD alonewasable to translocate into HeLa cells as determined by confocalmicroscopy (Fig. 1B). We also assessed the transduction ability ofTCTP-PTD into other cell lines, U-937, NIH-3T3 and MDCK. Wefound that, TCTP-PTD was transduced in a dose-dependent mannerinto all cell lines studied (Fig. 1C).
3.2. Efficient transduction of b-gal proteins by TCTP-PTD
We then tested the cargo delivering ability of TCTP-PTD, afterfusing beta-galactosidase (b-gal) to the TCTP-PTD. We found thatTCTP-PTD/b-gal fusion complex was transduced into HeLa cells ina dose-dependent manner on 2 h of treatment, but with a lowertransduction efficiency compared to that of TAT-PTD/b-gal (Fig. 2A).We next injected mice intravenously with 200 mg of b-gal protein,and TCTP-PTD/b-gal fusion protein, respectively, and examined thelivers, kidneys, spleens, hearts, lungs and brains at 2 h after injec-tions. All tissues, excluding brain, exhibited strongly increased b-galenzyme activity at 2 h after injection of TCTP-PTD/b-gal but notb-gal protein (Fig. 2B). Interestingly, TCTP-PTD/b-gal was trans-duced into the lung more effectively than into the other tissues ofthemice. Then, to further confirm the internalizing capability of thefusion protein, we isolated and cryo-sectioned tissues from micefollowing intraperitoneal administration of 400 mg of TCTP-PTD/b-gal or b-gal. The cryo-sectioned tissues were then analyzed forthe extent of penetration of the fused protein after staining forb-gal. We found that all tissues other than the brain stronglystained blue, suggesting that their interior regions were penetratedby the TCTP-PTD/b-gal, while b-gal alone showed no penetration(Fig. 2C). The cryo-sectioned brain tissues of TCTP-PTD/b-gal
showed weak response, while the whole brainwas entirely stainedby blue color. This suggests that TCTP-PTD/b-gal can, penetrate theexterior regions of the brain to some extent, and transduction ofTCTP-PTD/b-gal into the interior regions of the brain is relativelylower and/or slower than into the other organs. Thus, we confirmedthat this TCTP-PTD has the ability to carry much larger molecules tonearly all tissues except brain in vivo. This suggests that TCTP-PTDmay be potentially useful in delivering awide variety of biologicallyactive therapeutic cargos.
We then examined the toxic, cytotoxic, and immunogeniceffects of long-term transduction of TCTP-PTD in vivo. In HeLa cells,1 mM and 10 mM of TCTP-PTD induced no cytotoxicity and treatmentof TCTP-PTD at 100 mM for 24 h induced weak cytotoxicity, less thanthat caused by TAT-PTD (Fig. S2A). We also found that intravenousinjection of mice with 1.6 mg/kg of TCTP-PTD for 14 days did notinduce any changes in body weight, or liver or kidney toxicity(Fig. S2BeD).
3.3. Characteristics of TCTP-PTD transduction
To investigate the kinetic profiles of the transduction of TCTP-PTD,we treated HeLa cells with TAMRA-labeled TCTP-PTD in a dose- andtime-dependent manner, and compared the mean fluorescenceintensity (MFI) with TAT-PTD using FACS analyzer. We found that thetransduction kinetics of TCTP-PTD and TAT-PTD were significantlydifferent. While TAT-PTD penetrated into cells at low doses (2e8 mM)during the first 1e2 h, TCTP-PTD penetrated into cells at relativelyhigh doses (8e32 mM) (Fig. 3A) in delayed time periods (4e16 h)(Fig. 3B). In parallel experiments using b-gal conjugated protein,TCTP-PTD/b-gal fusion protein was transduced into HeLa cells athigher doses and later times than TAT-PTD/b-gal (Fig. S3). When theincubation was carried out for 16 h, TCTP-PTD as well as TCTP-PTD/b-gal were more efficiently internalized into the cells than those ofTAT-PTD. Longer incubations and higher doses required for trans-duction with TCTP-PTD, in contrast to TAT-PTD, suggests that thesepeptides traverse cell membranes via different mechanisms.
In order to study the mechanism of transduction of TCTP-PTD,we first compared the features of the uptake process such as

Fig. 2. TCTP-PTD facilitates efficient transduction of b-gal in vitro and in vivo. (A) Transduction of TAT-PTD/b-gal and TCTP-PTD/b-gal. After 2 h incubation with various amounts ofTAT-PTD/b-gal and TCTP-PTD/b-gal, intracellular transduction in HeLa cells was analyzed by assay of b-gal activity. (B, C) In vivo delivery of TCTP-PTD/b-gal. After intravenousinjection of 200 mg of TCTP-PTD/b gal in mice, isolated tissues were analyzed for b-gal activity assay. After intraperitoneal injection of 400 mg, cryo-sectioned tissues were analyzedby blue staining of b-gal. Mean � SD.
M. Kim et al. / Biomaterials 32 (2011) 222e230 225
interaction with extracellular heparin sulfate and improvement oftransduction capacity by pretreatment of endolysomotrophicagent, chloroquine employed for TAT-PTD. Before incubation withTAMRA-labeled TAT-PTD or TCTP-PTD, HeLa cells were pretreatedwith soluble heparin or chloroquine, and then the fluorescenceintensity was analyzed by FACS. It has been shown that the TAT-PTDbinds to heparin and that internalization of the TAT protein requiresthe presence of cell-surface heparin sulfate proteoglycans [21,22].Following pretreatment with heparin, transduction of TAT-PTDwasinhibited as expected, but the transduction capability of TCTP-PTDwas not influenced (Fig. 3C). TAT-PTD has been reported to localizeto the nucleus after uptake, suggesting that chloroquine protectsTAT-PTD from proteolytic degradation [23]. Consistent withprevious reports, pretreatment with endolysomotrophic agent,chloroquine facilitated the transfer of TAT-PTD. In contrast, uptakeof TCTP-PTD was not significantly potentiated by pretreatmentwith chloroquine (Fig. 3D). These results imply that TCTP-PTD doesnot require heparin sulfate for transduction and transduced TCTP-PTD itself can efficiently escape from endosome. We then deter-mined whether TCTP-PTD enters into intracellular organelles likenucleus and lysosome. After 2 h incubation of 10 mM TAMRA-labeledTCTP-PTD with Lyso-tracker or DAPI, live HeLa cell images wereachieved by confocal microscope. TCTP-PTD was dispersed widelyin the cytoplasmic region and was not found the interior of lyso-some (Fig. 3E) and nucleus (Fig. 3F). Because TCTP-PTD consists ofhydrophobic amino acids and contains no functional nuclear
localization signal (NLS) like TAT-PTD, TCTP-PTD differs from TAT-PTD in intracellular localization and characteristics of transductionmechanisms. Consequently, the fact that TCTP-PTD does nottranslocate to the nucleus, suggests that it may not disturb thenormal expression of genes.
3.4. Internalization of TCTP-PTD
We tested whether internalization of TCTP-PTD is an energy-dependent process. After HeLa cells were cooled to 4 �C orpretreated in energy depletion media at 37 �C, the uptake ofTAMRA-labeled TCTP-PTD was completely inhibited at 4 �C, andalso dramatically reduced under circumstances of energy depletion(Fig. 4A). These data indicate that TCTP-PTD is internalized viaa mechanism involving energy-dependent endocytosis. Endocy-tosis, can occur through four pathways including macropinocytosis,or clathrin-mediated, lipid raft-mediated, or clathrin-, lipid raft-independent pathways [24]. To investigatewhich of these endocyticpathways is involved, we first examined the role of clathrin-coatedpits in the process of TCTP-PTD uptake. HeLa cells were pretreatedwith chlorpromazine (CPZ), a specific inhibitor of clathrin-mediatedendocytosis, in a dose-dependent manner. Pretreatment with 15 mMCPZ had little effect on internalization of TCTP-PTD (Fig. 4B). Also,after 1 h incubation of both TCTP-PTD and Alexa Fluor 488-labeledtransferrin, fluorescence images were achieved in live HeLa cells byconfocal microscopy. We found that TCTP-PTD peptide did not
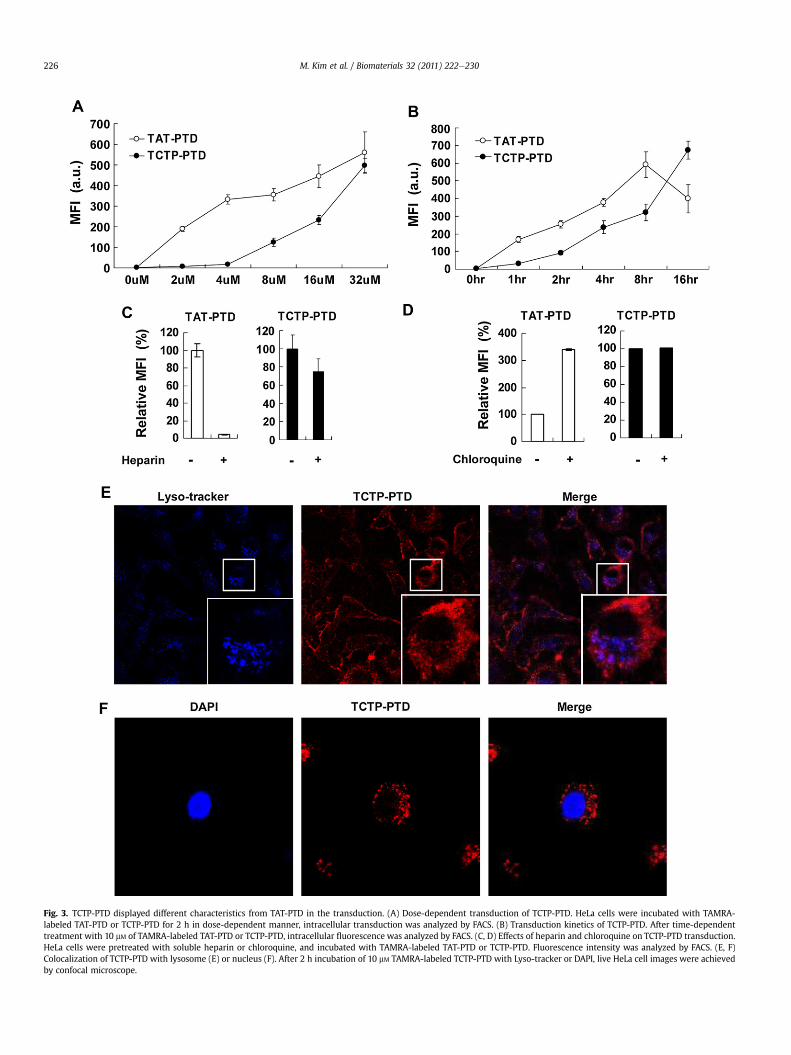
Fig. 3. TCTP-PTD displayed different characteristics from TAT-PTD in the transduction. (A) Dose-dependent transduction of TCTP-PTD. HeLa cells were incubated with TAMRA-labeled TAT-PTD or TCTP-PTD for 2 h in dose-dependent manner, intracellular transduction was analyzed by FACS. (B) Transduction kinetics of TCTP-PTD. After time-dependenttreatment with 10 mM of TAMRA-labeled TAT-PTD or TCTP-PTD, intracellular fluorescence was analyzed by FACS. (C, D) Effects of heparin and chloroquine on TCTP-PTD transduction.HeLa cells were pretreated with soluble heparin or chloroquine, and incubated with TAMRA-labeled TAT-PTD or TCTP-PTD. Fluorescence intensity was analyzed by FACS. (E, F)Colocalization of TCTP-PTD with lysosome (E) or nucleus (F). After 2 h incubation of 10 mM TAMRA-labeled TCTP-PTD with Lyso-tracker or DAPI, live HeLa cell images were achievedby confocal microscope.
M. Kim et al. / Biomaterials 32 (2011) 222e230226
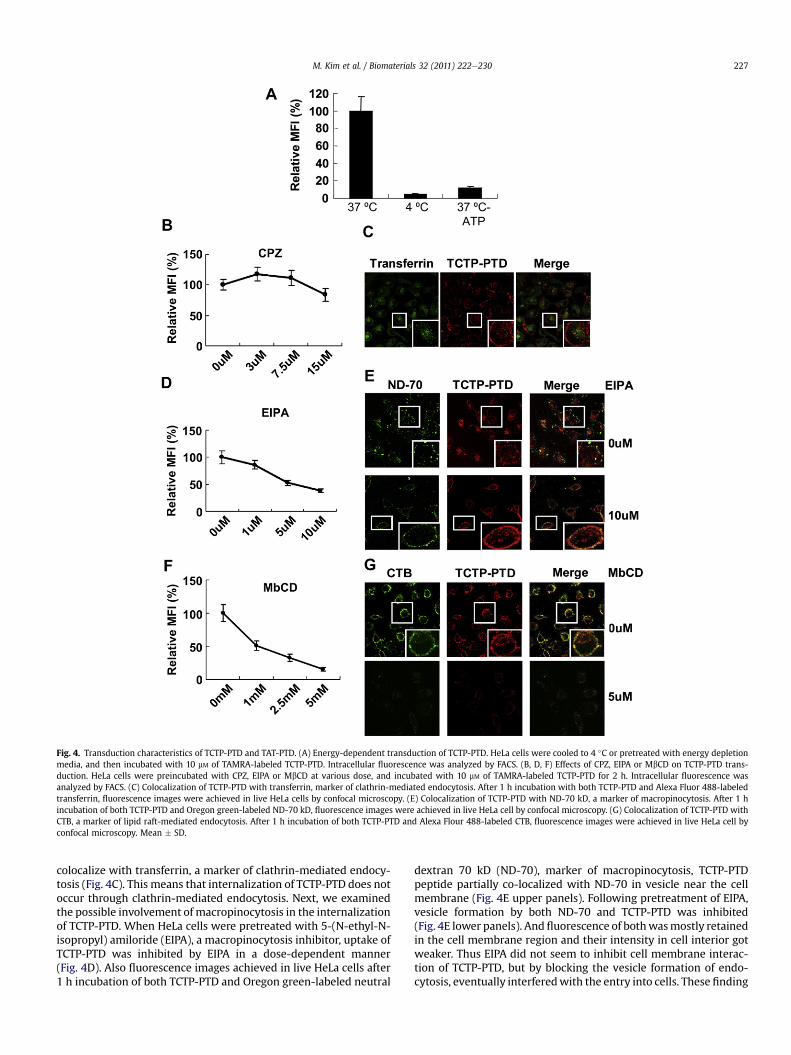
Fig. 4. Transduction characteristics of TCTP-PTD and TAT-PTD. (A) Energy-dependent transduction of TCTP-PTD. HeLa cells were cooled to 4 �C or pretreated with energy depletionmedia, and then incubated with 10 mM of TAMRA-labeled TCTP-PTD. Intracellular fluorescence was analyzed by FACS. (B, D, F) Effects of CPZ, EIPA or MbCD on TCTP-PTD trans-duction. HeLa cells were preincubated with CPZ, EIPA or MbCD at various dose, and incubated with 10 mM of TAMRA-labeled TCTP-PTD for 2 h. Intracellular fluorescence wasanalyzed by FACS. (C) Colocalization of TCTP-PTD with transferrin, marker of clathrin-mediated endocytosis. After 1 h incubation with both TCTP-PTD and Alexa Fluor 488-labeledtransferrin, fluorescence images were achieved in live HeLa cells by confocal microscopy. (E) Colocalization of TCTP-PTD with ND-70 kD, a marker of macropinocytosis. After 1 hincubation of both TCTP-PTD and Oregon green-labeled ND-70 kD, fluorescence images were achieved in live HeLa cell by confocal microscopy. (G) Colocalization of TCTP-PTD withCTB, a marker of lipid raft-mediated endocytosis. After 1 h incubation of both TCTP-PTD and Alexa Flour 488-labeled CTB, fluorescence images were achieved in live HeLa cell byconfocal microscopy. Mean � SD.
M. Kim et al. / Biomaterials 32 (2011) 222e230 227
colocalize with transferrin, a marker of clathrin-mediated endocy-tosis (Fig. 4C). This means that internalization of TCTP-PTD does notoccur through clathrin-mediated endocytosis. Next, we examinedthe possible involvement of macropinocytosis in the internalizationof TCTP-PTD. When HeLa cells were pretreated with 5-(N-ethyl-N-isopropyl) amiloride (EIPA), a macropinocytosis inhibitor, uptake ofTCTP-PTD was inhibited by EIPA in a dose-dependent manner(Fig. 4D). Also fluorescence images achieved in live HeLa cells after1 h incubation of both TCTP-PTD and Oregon green-labeled neutral
dextran 70 kD (ND-70), marker of macropinocytosis, TCTP-PTDpeptide partially co-localized with ND-70 in vesicle near the cellmembrane (Fig. 4E upper panels). Following pretreatment of EIPA,vesicle formation by both ND-70 and TCTP-PTD was inhibited(Fig. 4E lower panels). And fluorescence of bothwasmostly retainedin the cell membrane region and their intensity in cell interior gotweaker. Thus EIPA did not seem to inhibit cell membrane interac-tion of TCTP-PTD, but by blocking the vesicle formation of endo-cytosis, eventually interferedwith the entry into cells. These finding

Fig. 5. TCTP-PTD improves adenoviral gene expression in vitro and in vivo. (A, B, C) Adenoviral gene expression of Ad-GFP is enhanced in HeLa and NIH-3T3 cells by TCTP-PTD. HeLacells were infected with Ad-GFP pre-complexed with increasing concentrations of TCTP-PTD. GFP expression was analyzed by FACS or fluorescence microscope. (D, E) TCTP-PTDEnhanced adenoviral gene delivery in vivo. Humoral immune responses induced by complex formation of rAd/3�G with TCTP-PTD. BALB/c mice were intranasally immunized oncewith rAd/3�G precomplexed with TCTP-PTD and systemic anti-RSV IgG antibody titers were measured by serum ELISA 2 weeks after immunization. The results represent log2 endpoint values from four or five individual mice (D). Mucosal immune responses induced by complex formation of rAd/3�G with TCTP-PTD. Mucosal IgA titers in the BAL fluid weremeasured 2 weeks after immunization (E). Mean � SD (n � 4) *P � 0.05.
M. Kim et al. / Biomaterials 32 (2011) 222e230228
imply that macropinocytosis is a partial component of the inter-nalization process of TCTP-PTD. Lastly, to elucidate whetherTCTP-PTD enters cell via the lipid raft-mediated endocytosis, wepretreated the cells with various doses of methyl-b-cyclodextrin(MbCD), an inhibitor of lipid raft-mediated endocytosis. After the2 h incubation of TAMRA-labeled TCTP-PTD, internalization ofTCTP-PTD was completely inhibited by 5 mM of MbCD (Fig. 4F). Asubsequent experiment using fluorescence imaging with TCTP-PTDand Alexa Flour 488-labeled cholera toxin B (CTB), revealed thatTCTP-PTD was co-localized with CTB, a marker of lipid-raft medi-ated endocytosis (Fig. 4G upper panel). But when pretreatedwith 5 mM MbCD, cell membrane binding as well as intracellularinternalization of TCTP-PTD were almost completely inhibited(Fig. 4G lower panel). These results indicate that internalization of
TCTP-PTD occurs through lipid raft-mediated endocytosis. Takentogether, our data provide evidence that TCTP-PTD is internalizedmainly via lipid-raft mediated endocytosis and partly by adoptingmacropinocytosis.
3.5. Enhanced adenoviral gene delivery by TCTP-PTD
Although there have been many different approaches to theintroduction of DNA and proteins into target cells, low efficiency ofthe non-viral vector systems, as well as the immunogenicity andtransiency of the expression of many of the viral vectors [25,26],have limited their applications in vivo. Viral infection requires theinteraction of coxackivirus and adenovirus receptor (CAR) at cellmembrane before viral entry into host cells [27]. As many viral

M. Kim et al. / Biomaterials 32 (2011) 222e230 229
vectors are unable to infect cell types which have low levels ofspecific receptors or co-receptors, cationic PTDs, such as TAT, Antpand HP-4 were used to enhance viral entry and infection efficiency[28,29]. In this work, we examined whether TCTP-PTD can alsopotentiate the infection efficiency of adenovirus. We comparedvirus-delivered gene expression both in CAR positive HeLa cells(Fig. 5A) and CAR deficient NIH-3T3 cells (Fig. 5B) using anadenovirus encoding green fluorescent protein (Ad-GFP). Twentyfour h after viral infection, we analyzed GFP expression by FACS.TCTP-PTD showed enhanced expression in a dose-dependentmanner from 1 mM to 30 mM. But in high concentrations above100 mM, TCTP-PTD showed did not cause any increase in infectionefficacy, unlike TAT-PTD, suggesting that its hydrophobic propertiesformed self aggregates and blocked the internalization. Ata concentration of 30 mM, TCTP-PTD increased GFP expression by4.87 fold (in HeLa) and 7.68 fold (in NIH-3T3) compared with virusalone.We also noted that GFP expression by TCTP-PTD at 30 mM wasgreater than by TAT-PTD at 300 mM. This can be an advantagebecause viral delivery by TCTP-PTD can be accomplished withlower viral levels, thereby avoiding any potential cytotoxicity. Inaddition, examination of GFP expression in HeLa cells by fluores-cence microscopy, confirmed that TCTP-PTD enhanced adenoviralGFP expression in a dose-dependent manner up to 30 mM (Fig. 5C).Finally, we examined whether TCTP-PTD improves the antigen-specific immune responses in vivo, using low doses of adenovirusencoding three copies of RSV G protein fragment (rAd/3�G)(5.0 � 106 PFU). RSV G glycoprotein was reported as a majorattachment protein of the virus [30] and a single administration ofrAd/3�G induced immunity against RSV infection, a dangerous toinfants [31,32]. Before initiating the in vivo study, we firstconfirmed that pre-incubation of rAd/3�G with TCTP-PTDincreased RSV G protein expression in HeLa cells (data not shown).Next, we immunized mice once with rAd/3�G pre-complex withTCTP-PTD and measured serum anti-RSV IgG antibody titers byELISA 2 wks after immunization. In control groups, a complex ofrAd/3�G and TAT peptide induced strong RSV-specific total IgGresponses in serum, compared with those elicited by rAd/3�G only.Also administration of a complex of rAd/3�G and TCTP-PTD led toserum IgG response comparable to that induced by TAT complex(Fig. 5D). Then, we examined whether immunization with thecomplex of rAd/3�G and TCTP-PTD peptide elicits a RSV-specificsecretory IgA response. Mucosal IgA titers in the bronchoalveolarlavage fluid (BALF) were measured 2 weeks after immunization.Notably, the level of mucosal IgA was higher in BALFs of the groupadministrated the complex of rAd/3�G and TCTP-PTD peptide, thanin the group administrated rAd/3�G and TAT-PTD peptide (Fig. 5E).Based on these results, we conclude that TCTP-PTD significantlyenhanced the delivery and expression of adenoviral gene, withdramatic improvements in mucosal and humoral immunogenicity.
4. Discussion
The tertiary structure of TCTP is predicted to contain threehelixes: a very short first helix (H1) with the second (H2) and thirdhelices (H3) exposed to the outside. In the predicted helical wheelmodel of the H2 and H3 structures of TCTP in Schizosaccharomycespombe [10], basic amino acids are distributed toward outer surfaceof helix, and are assumed to be involved in protein transductionactivity. However, our preliminary experiments revealed that thesehelixes have nothing to do with PTD’s functional abilities (data notshown). In follow-up experiments, we found that the TCTP-PTD,MIIYRDLISH, at the NH2-terminus of TCTP, rather than H2 and H3regions was responsible for the transduction function. TCTP-PTDhas hydrophobic cores of 1MII and 7LI and 5 polar or charged aminoacids, 4Y, 5R, 6D, 9S and 10H. Notably, when 6th aspartate residue
(6D) is substituted by alanine, the cell penetrating activity increased(data not shown). This finding suggests that hydrophobicity ofTCTP-PTD contributes to its cell penetrating ability.
The mechanism by which PTDs internalize into cells is notcompletely understood. Several kinds of mechanisms have beensuggested for the internalization of PTDs into the cells, but it isobvious that PTDs use at least two independent multi-step mech-anisms. The first step in the internalization process is the interac-tion of PTD with the cell surface. It has been shown that cationicPTDs bind electrostatically to the exposed charged parts of phos-pholipids and that loosely bound PTD can be removed from the cellsurface by ice-cold acidic solutions [33]. The interaction of PTDwithlipid bilayer was also studied by molecular modeling [2] and theaccumulation of labeled PTDs in or at the cell membrane wasreadily observed by confocal microscopy. Recently, heparan sulfate(HS) a proteoglycan in ECM, was reported as molecular portal ofbasic PTDs in many studies. But, as our data indicated, the trans-location mechanism of TCTP-PTD is different from those of otherPTDs such as TAT and penetratin. TCTP-PTD did not require heparansulfate for the initial interaction with cell membrane and accu-mulation of rhodamine-labeled TCTP-PTD in HeLa cell membranewas observed by confocal microscopy image, suggesting that TCTP-PTD may interact with other component(s) in ECM besides HS. Themechanism of action of MbCD, used in the present study, to blocklipid raft-mediated endocytosis, is achieved via depletion ofcholesterols. Pretreatment of MbCD dramatically inhibited thetransduction of TCTP-PTD in FACS study and blocked the accumu-lation of this peptide at the membrane, as shown in confocalmicroscopy studies, which implies that cholesterol may be a majorcomponent in ECM of membrane portal of TCTP-PTD.
Although different PTDs are internalized through different kindsof endocytosis, their common destiny is the lysosome becausepretreatment of the cells with chloroquine enhances the trans-duction efficiency through lysosomal escape. Treatment withchloroquine has been shown to prevent degradation of the fusedprotein and aid its release from lysosomes. However, TCTP-PTDwastransduced in a chloroquine-independent manner and was not co-localized with Lyso-Tracker, indicating that TCTP-PTD itself has thecapacity to escape from lysosomes. Thus it appears that TCTP-PTD isstable in the cells, enabling it to be more effective transducer atlater time period, than TAT-PTD (Fig. 3B, Fig. S3). Although TCTP-PTD/b-gal exhibited better transduction efficiency at higher dosesand later time points than TAT-PTD/b-gal, in vivo experimentsconducted under equivalent conditions showed that TCTP-PTD/b-gal exhibited comparable b-gal activity with TAT-PTD/b-gal. Apossible explanation for this might be that positively charged PTDslike TAT-PTDmight transduce more efficiently into single cell layersvia the interaction with the negatively charged heparan sulfatelocated at the plasma membrane than hydrophobic PTDs such asTCTP-PTD. While TCTP-PTD cannot transduce efficiently into singlecell layers because of its hydrophobicity, it can transduce better intotissues comprising multiple cell layers, as the in vivo studies haveshown.
Antp and TAT were shown to improve cellular uptake andtherapeutic gene delivery of replication-deficient viruses in cellsand in vivo. These studies provide a simple approach which is justto pre-incubate the adenovirus with PTD at room temperature. Inthis study, we tried this approach to improve adenoviral delivery tocells and tissues, particularly the lung, using TCTP-PTD. Wepostulated that TCTP-PTD can interact with another component ofadenovirus coat, different from cationic PTDs. Although pre-incu-bation with amounts less than 1 mM of TCTP-PTD improved theexpression of adenovirus expressing GFP, higher doses of TCTP-PTD(>5 mM) did not enhance the viral expression in cells (data notshown). This might be because the remaining unbound peptides

M. Kim et al. / Biomaterials 32 (2011) 222e230230
compete with viral particle at the site of infection withoutincreasing the effective concentration of viral particles on thesurfaces. Therefore this method of incubation with PTDs, will beeffective only when appropriate doses of PTD and virus are used.Still systemic non-viral therapies to treat human disease will be oflimited success because of plasma membrane’s impermeability tomacromolecules and polar molecules. We have shown here thata small hydrophobic peptide, derived from amino terminus of TCTP,is a useful vehicle for efficient adenoviral gene delivery.
5. Conclusions
We identified and characterized a PTD, 10 amino acids long, atthe NH2-terminus of human TCTP. We demonstrated that TCTP-PTDhas a good transduction ability; it efficiently mediates the transferof cargos across biological membranes both in cell culture and inliving animals without provoking toxicity. We have also shown thatTCTP-PTD is internalized in cells mainly by lipid-raft mediatedendocytosis and partially by macropinocytosis. Using adenoviraldelivery systems, we further demonstrated that TCTP-PTD alsoenhances the efficacy of viral infection in cells and boosts immuneresponses in vivo. Our results suggest that because of its hydro-phobic nature, TCTP-PTD might overcome the limitations ofpolycationic- transducers such as undesirable ionic interactionswith nucleic acids. This study should help in the development ofTCTP-PTD variants with different degrees of hydrophobicity andimproved ability to deliver cargos such as siRNA and cancer killingagents. Further studies in this regard are currently underway in ourlaboratory.
Acknowledgments
This study was supported by a grant of the Korea Healthcaretechnology R&D Project, Ministry for Health,Welfare & Family Affairs(A090030), NRF of Korea Grant funded by the Korean Government(2009-0064401), Mid-career Research Program through NRF grantfunded by the MEST (R01-2007-000-20263-0), Seoul R&BD Program(ST090801), and the NCRC programofMOST/KOSEF (R15-2006-020).
Appendix. Supplementary information
The supplementary data associated with this article can be foundin the on-line version at doi:10.1016/j.biomaterials.2010.08.077.
Appendix
Figures with essential color discrimination. Figs. 1e5 in thisarticle are difficult to interpret in black and white. The full colorimages can be found in the on-line version, at doi:10.1016/j.biomaterials.2010.08.077.
References
[1] Jarver P, Langel U. The use of cell-penetrating peptides as a tool for generegulation. Drug Discov Today 2004;9(9):395e402.
[2] Soomets U, Lindgren M, Gallet X, Hallbrink M, Elmquist A, Balaspiri L, et al.Deletion analogues of transportan. Biochim Biophys Acta 2000;1467(1):165e76.
[3] Drin G, Rousselle C, Scherrmann JM, Rees AR, Temsamani J. Peptide delivery tothe brain via adsorptive-mediated endocytosis: advances with SynB vectors.AAPS PharmSci 2002;4(4):E26.
[4] Moghimi SM, Symonds P, Murray JC, Hunter AC, Debska G, Szewczyk A. A two-stage poly(ethylenimine)-mediated cytotoxicity: implications for gene trans-fer/therapy. Mol Ther 2005;11(6):990e5.
[5] Veldhoen S, Laufer SD, Trampe A, Restle T. Cellular delivery of small inter-fering RNA by a non-covalently attached cell-penetrating peptide: quantita-tive analysis of uptake and biological effect. Nucleic Acids Res 2006;34(22):6561e73.
[6] Schwarze SR, Ho A, Vocero-Akbani A, Dowdy SF. In vivo protein transduction:delivery of a biologically active protein into the mouse. Science 1999;285(5433):1569e72.
[7] Chiu YL, Ali A, Chu CY, Cao H, Rana TM. Visualizing a correlation betweensiRNA localization, cellular uptake, and RNAi in living cells. Chem Biol 2004;11(8):1165e75.
[8] Waldeck W, Pipkorn R, Korn B, Mueller G, Schick M, Toth K, et al. Transportermolecules influence the gene expression in HeLa cells. Int J Med Sci 2009;6(1):18e27.
[9] Bommer UA, Thiele BJ. The translationally controlled tumour protein (TCTP).Int J Biochem Cell Biol 2004;36(3):379e85.
[10] Thaw P, Baxter NJ, Hounslow AM, Price C, Waltho JP, Craven CJ. Structure ofTCTP reveals unexpected relationship with guanine nucleotide-free chaper-ones. Nat Struct Biol 2001;8(8):701e4.
[11] Langdon JM, Schroeder JT, Vonakis BM, Bieneman AP, Chichester K,Macdonald SM. Histamine-releasing factor/translationally controlled tumorprotein (HRF/TCTP)-induced histamine release is enhanced with SHIP-1knockdown in cultured human mast cell and basophil models. J Leukoc Biol2008;84(4):1151e8.
[12] Kim M, Min HJ, Won HY, Park H, Lee JC, Park HW, et al. Dimerization oftranslationally controlled tumor protein is essential for its cytokine-likeactivity. PLoS One 2009;4(7):e6464.
[13] MacDonald SM, Rafnar T, Langdon J, Lichtenstein LM. Molecular identificationof an IgE-dependent histamine-releasing factor. Science 1995;269(5224):688e90.
[14] Susini L, Besse S, Duflaut D, Lespagnol A, Beekman C, Fiucci G, et al. TCTPprotects from apoptotic cell death by antagonizing bax function. Cell DeathDiffer 2008;15(8):1211e20.
[15] Liu H, Peng HW, Cheng YS, Yuan HS, Yang-Yen HF. Stabilization andenhancement of the antiapoptotic activity of mcl-1 by TCTP. Mol Cell Biol2005;25(8):3117e26.
[16] Yang Y, Yang F, Xiong Z, Yan Y, Wang X, Nishino M, et al. An N-terminal regionof translationally controlled tumor protein is required for its antiapoptoticactivity. Oncogene 2005;24(30):4778e88.
[17] Hsu YC, Chern JJ, Cai Y, Liu M, Choi KW. Drosophila TCTP is essential forgrowth and proliferation through regulation of dRheb GTPase. Nature2007;445(7129):785e8.
[18] Tuynder M, Fiucci G, Prieur S, Lespagnol A, Geant A, Beaucourt S, et al.Translationally controlled tumor protein is a target of tumor reversion. ProcNatl Acad Sci U S A 2004;101(43):15364e9.
[19] Arcuri F, Papa S, Carducci A, Romagnoli R, Liberatori S, Riparbelli MG, et al.Translationally controlled tumor protein (TCTP) in the human prostate andprostate cancer cells: expression, distribution, and calcium binding activity.Prostate 2004;60(2):130e40.
[20] Kim MJ, Kwon JS, Suh SH, Suh JK, Jung J, Lee SN, et al. Transgenic over-expression of translationally controlled tumor protein induces systemichypertension via repression of Naþ, Kþ-ATPase. J Mol Cell Cardiol 2008;44(1):151e9.
[21] Richard JP, Melikov K, Brooks H, Prevot P, Lebleu B, Chernomordik LV. Cellularuptake of unconjugated TAT peptide involves clathrin-dependent endocytosisand heparan sulfate receptors. J Biol Chem 2005;280(15):15300e6.
[22] Argyris EG, Kulkosky J, Meyer ME, Xu Y, Mukhtar M, Pomerantz RJ, et al. Theperlecan heparan sulfate proteoglycan mediates cellular uptake of HIV-1 Tatthrough a pathway responsible for biological activity. Virology 2004;330(2):481e6.
[23] Wadia JS, Stan RV, Dowdy SF. Transducible TAT-HA fusogenic peptideenhances escape of TAT-fusion proteins after lipid raft macropinocytosis. NatMed 2004;10(3):310e5.
[24] Conner SD, Schmid SL. Regulated portals of entry into the cell. Nature2003;422(6927):37e44.
[25] Frankel AD, Pabo CO. Cellular uptake of the tat protein from human immu-nodeficiency virus. Cell 1988;55(6):1189e93.
[26] Green M, Loewenstein PM. Autonomous functional domains of chemicallysynthesized human immunodeficiency virus tat trans-activator protein. Cell1988;55(6):1179e88.
[27] Bergelson JM, Cunningham JA, Droguett G, Kurt-Jones EA, Krithivas A, Hong JS,et al. Isolation of a common receptor for Coxsackie B viruses and adenoviruses2 and 5. Science 1997;275(5304):1320e3.
[28] Youn JI, Park SH, Jin HT, Lee CG, Seo SH, Song MY, et al. Enhanced deliveryefficiency of recombinant adenovirus into tumor and mesenchymal stem cellsby a novel PTD. Cancer Gene Ther 2008;15(11):703e12.
[29] Gratton JP, Yu J, Griffith JW, Babbitt RW, Scotland RS, Hickey R, et al. Cell-permeable peptides improve cellular uptake and therapeutic gene delivery ofreplication-deficient viruses in cells and in vivo. Nat Med 2003;9(3):357e62.
[30] Levine S, Klaiber-Franco R, Paradiso PR. Demonstration that glycoprotein G isthe attachment protein of respiratory syncytial virus. J Gen Virol 1987;68(Pt9):2521e4.
[31] Yu JR, Kim S, Lee JB, Chang J. Single intranasal immunization with recombi-nant adenovirus-based vaccine induces protective immunity against respi-ratory syncytial virus infection. J Virol 2008;82(5):2350e7.
[32] Glezen WP, Greenberg SB, Atmar RL, Piedra PA, Couch RB. Impact of respi-ratory virus infections on persons with chronic underlying conditions. JAMA2000;283(4):499e505.
[33] Pooga M, Hallbrink M, Zorko M, Langel U. Cell penetration by transportan.FASEB J 1998;12(1):67e77.




















