
Human leukocyte antigen class II-based immune risk model ...
Upload
independentCategory
view
3download
0
ARTICLE IN PRESS
Clinical Nutrition (2006) 25, 923–938
0261-5614/$ - sdoi:10.1016/j.c
AbbreviationEPA, eicosapen3-phosphate demitogen-activaPUFA, polyunsafactor�CorrespondiE-mail addr
http://intl.elsevierhealth.com/journals/clnu
ORIGINAL ARTICLE
Effect of docosahexaenoic acid-rich fish oilsupplementation on human leukocyte function
Renata Gorjaoa,�, Rozangela Verlengiab, Thais Martins de Limaa,Francisco Garcia Sorianoc, Maria Fernanda Cury Boaventuraa,Carla Christine Kanunfred, Carmem Maldonado Peresa,Sandra Concuzzo Sampaioe, Rosemari Ottonf,Alessandra Foladora, Edgair Fernandes Martinsa, Tania CristinaPithon Curif, Erica Paula Portiollia, Philip Newsholmeg, Rui Curia
aDepartment of Physiology and Biophysics, Institute of Biomedical Sciences, University of Sao Paulo, Av.Prof. Lineu Prestes, 1524, 05508-900, Butanta, Sao Paulo, SP, BrazilbMethodist University of Piracicaba, Faculty of Physical Education—FACEF, Sao Paulo, BrazilcDivision of Clinical Emergency, Medical School, University of Sao Paulo, BrazildUniversity of Ponta Grossa, BrazileLaboratory of Pathophysiology, Butantan Institute, BrazilfBiological Sciences and Health Center, Cruzeiro do Sul University, BrazilgDepartment of Biochemistry, Conway Institute of Biomolecular and Biomedical Research, UniversityCollege Dublin, Ireland
Received 7 February 2006; accepted 17 March 2006
KEYWORDSFatty acids;Leukocytes;Phagocytosis;Chemotaxis;Lymphocyte prolif-eration;
ee front matter & 2006lnu.2006.03.004
s: AA, arachidonic acid;taenoic acid (20:5 n-3);hydrogenase; GSTA1, glted protein kinase; PBMturated fatty acids; RO
ng author. Tel.: +55 11ess: [email protected]
SummaryBackground: The effect of a docosahexaenoic acid (DHA)-rich fish oil (FO)supplementation on human leukocyte function was investigated.Methods: Ten male volunteers were supplemented with 3 g/day FO containing 26%eicosapentaenoic acid (EPA, 20:5, n-3) and 54% DHA (22:6, n-3) for 2 months.Results: FO supplementation changed the fatty acid (FA) composition of leukocytesresulting in an increase of n-3/n-6 ratio from 0.18 to 0.62 in lymphocytes and from0.15 to 0.70 in neutrophils. DHA-rich FO stimulated an increase in phagocytic activity
Elsevier Ltd and European Society for Clinical Nutrition and Metabolism. All rights reserved.
BAX, BCL2-associated X protein; ConA, Concanavalin-A; DHA, docosahexaenoic acid (22:6 n-3);FAs, fatty acids; FO, fish oil; GADD45, DNA-damage-inducible protein; GAPDH, glyceraldehyde-utathione S-transferase A1; HSP, heat shock protein; IFN, interferon; IL, interleukin; MAP kinase,Cs, peripheral blood mononuclear cells; PPAR, peroxisome proliferator-activated receptor;S, reactive oxygen species; TBARS, thiobarbituric acid-reactive substances; TNF, tumor necrosis
3091 7245; fax: +55 11 3091 7285.r (R. Gorjao).

ARTICLE IN PRESS
R. Gorjao et al.924
Gene expression;Macroarray
by 62% and 145% in neutrophils and monocytes, respectively. Neutrophil chemotacticresponse was increased by 128%. The rate of production of reactive oxygen speciesby neutrophils was also increased, as it was with lymphocyte proliferation. Thesechanges were partially reversed after a 2-month wash out period. With respect tocytokine production by lymphocytes, interleukin (IL)-4 release was not altered,whereas secretions of IL-10, interferon (IFN)-g and tumor necrosis factor (TNF)-awere raised. These results are in contrast to those described by others using EPA-richFO supplementation. Lymphocyte pleiotropic gene expression was analyzed by amacroarray technique. Of the analyzed genes (588 in total), 77 were modified by thesupplementation. FO supplementation resulted in up-regulation of 6 genes (GATAbinding protein 2, IL-6 signal transducer, transforming growth factor alpha, TNF, heatshock 90 kDa protein 1-alpha and heat shock protein 70 kDa 1A) and a downregulation of 71 genes (92.2% of total genes changed). The largest functional groupof altered genes was that related to signaling pathways (22% of the total modifiedgenes).Conclusions: Therefore, although EPA and DHA are members of n-3 FA family,changes in the proportion of DHA and EPA exert different effects on neutrophil,monocyte and lymphocyte function, which may be a result of specific changes ingene expression.& 2006 Elsevier Ltd and European Society for Clinical Nutrition and Metabolism. Allrights reserved.
Introduction
Over the last 40 years, there has been increasinginterest in the positive effects of fish oil (FO),which is rich in n-3 fatty acids (FAs), on decreasingrisk or severity of cardiovascular1,2 and immune-mediated diseases.3,4 The positive effects of FOsupplementation are related to beneficial effectson cardiovascular diseases,5,6 autoimmune andinflammatory disorders, such as psoriasis,3 rheuma-toid arthritis4 and diabetes.7 In contrast, dietarylipids rich in n-6 FAs, found in vegetable oils, canaffect some inflammatory diseases.8
Eicosapentaenoic (EPA) and docosahexaenoic(DHA) acids are the most important members ofn-3 FA family. However, previous studies on immuneand cardiovascular function were mainly carriedout using FO rich in EPA, generally in a EPA:DHAratio of 2:1.3,9,10
The effects of FO supplementation on cytokineproduction by mononuclear leukocytes have beenfrequently reported.11 The majority of dietarysupplementation studies in animals and humanshave shown that n-3 FAs reduce production oflymphocyte derived pro-inflammatory cytokinesand inhibit mitogen- or antigen-induced lympho-cyte proliferative responses.12,13 FO supplementa-tion is associated with decreased levels ofinterleukin (IL)-1b and tumor necrosis factor(TNF)-a in blood from healthy volunteers.13 Mayeret al.14 reported that infusion of FO suppressesTNF-a, IL-1b, IL-6 and IL-8 release from mono-nuclear leukocytes in patients with sepsis. These
cytokines are overproduced in the progression oftrauma patients towards sepsis.11 The capacity ofn-3 FAs to decrease the levels of inflammatorycytokines suggests that FO might be a useful agentto aid the control of endotoxemia. Meydani et al.15
reported decreased lymphocyte IL-2 productionafter FO supplementation in healthy human volun-teers, whereas Thies et al.16 reported that FOsupplementation (720mg of EPA plus 280mg ofDHA) decreases lymphocyte proliferation. However,administration of 0.2 g/kg/day FO for 4 days inmoderately stressed surgical patients enhanced thelevels of IL-2, interferon (IFN)-g and TNF-a in theblood, indicating that, in this case, FO was notimmunosuppressive.17 Grimble et al.18 hypothe-sized that people with different genotypes thatcontrol TNF-a production would have differentsensitivities to the effects of FO.19 This fact couldexplain the controversial results of leukocytecytokine production observed in the literature.
Many inflammatory processes have been modifiedby n-3 FA supplementation and the mechanismsinclude inducing of changes in signal transductionand gene expression.20 n-3 FA-derived mediatorssuch as resolvins, docosatrienes and neuroprotectinshave been reported to play an important role for theeffect of n-3 FAs.21 The mechanisms involved in theeffects of these FAs seems to be closely correlatedwith the types of eicosanoids produced.22 EPA andAA are competitively metabolized via cyclooxygen-ase and lipoxygenase pathways to specific typesof eicosanoids.23 EPA-derived lipid mediatorspossess markedly reduced inflammatory properties

ARTICLE IN PRESS
Effect of fish oil on human leukocyte function 925
correlated to AA-derived eicosanoids.24 In compar-ison to AA, EPA is the preferential substrate forlipooxygenase, which explains for the higher for-mation of EPA-derived products when both free FAare simultaneously available.20 However, some ofthe effects of FA occur by eicosanoid-independentmechanisms, including actions upon receptor,25
intracellular signaling pathways,26 transcriptionfactor activity27 and gene expression.28
In spite of the information above, the effects ofEPA and DHA are not differentiated in most of thestudies. Some authors have reported differencesbetween these FA when the experiments areperformed with EPA and DHA separately. Verlengiaet al.29,30 have shown that expression of genesrelated to signal transduction, cell survival, apop-tosis and cytokine production was altered by thetreatment with both EPA and DHA in lymphocytes invitro. These FA had different effects on geneexpression analyzed by macroarray technique.The significant increase in expression of 20 selectedgenes and decrease in one gene by EPA and increasein seven selected genes by DHA argues for selectiveeffects of these n-3 PUFA on gene transcription of Blymphocytes.29 In a study with T lymphocytes, DHAraised the expression of 62% of the genes changedby the FAs (39 genes), whereas EPA up-regulated33%.30
Studies showed that EPA is more effective as animmunosupressor when compared with DHA.31,32
Thies et al.16 compared the effects of supplemen-tation with FO (o1 g/d), highly purified DHA (o1 g/day), and a placebo on lymphocyte proliferation inhealthy volunteers and demonstrated that EPA-richFO suppressed lymphocyte proliferation whereasDHA had no effect. Kelley et al.33 demonstratedthat a high intake (6 g/day) of DHA by healthy mendid not lower lymphocyte proliferation and theproduction of IL-2 by these cells and IgG, C3 and IL-2 receptor serum levels.33
The information above led us to investigatechanges in gene expression, cell proliferation,cytokine production, phagocytosis and chemotaxisafter dietary supplementation of a DHA-rich FO inhealthy men.
Materials and methods
Subjects and study design
The present study was approved by the EthicalCommittees of the University of Sao Paulo Hospitaland Institute of Biomedical Sciences, Sao PauloUniversity. All volunteers were submitted to a
health and lifestyle questionnaire. The nutritionalvalues of the diets were calculated using ‘‘VirtualNutri’’ program that was obtained from theFaculdade de Saude Publica, USP. All subjects hadalimentary habits according to the standards ofBrazilian people and were requested to maintaintheir usual diet. The body mass index (BMI) wasmeasured. The volunteers presented BMI of24.9771.23 kg/m2.
Ten men aged 25–45 years were randomlyassigned to receive a supplement of FO for 2months. Blood was collected before, after 2 monthsof supplementation and after 2 months of thisperiod (wash out period). The capsules adminis-tered to volunteers were obtained from NaturalisAlimentos Naturais Ltdas (Sao Paulo, Brazil). Thevolunteers were supplemented with 3 g/day of oilcontaining 26% EPA (20:5, n-3) and 54% DHA(22:6, n-3). This dose provided 1.62 and0.78 g/day of DHA and EPA, respectively. FOsupplementation increases the unsaturation indexof membrane FA as a result of n-3 FA incorporation.An increase in thiobarbituric acid-reactivesubstance (TBARS) concentration is then expected.Therefore, a-tocopherol (an antioxidant) has to besimultaneously administered during supplementa-tion to prevent lipid peroxidation. The FOwas then given in capsules containing 3.7mg/ga-tocopherol. Plasma TBARS content was deter-mined by the method of Hong et al.34 to evaluatethe occurrence of lipid peroxidation. Therewas no increase of TBARS content in plasmaafter FO supplementation. The values were2.1670.04 nmol/ml of plasma before and 2.1070.03 nmol/ml after supplementation (mean7SEMof eight volunteers).
Cell isolation
Blood venous was collected from the volunteers intubes containing ethylenediamine tetra acetic acid(EDTA; 1mg/ml). Blood was diluted in PBS (1:1) andthis suspension was layered on Histopaque-1077(Sigma Chemical Co., St. Louis, MO, EUA) andcentrifuged for 30min at 400g and room tempera-ture. Peripheral blood mononuclear cells (PBMCs; amixture of monocytes and lymphocytes) werecollected from the interphase and washed oncewith PBS. Neutrophils were prepared from theinferior sediment after erythrocytes being lysedwith 150mM NH4Cl, 10mM NaHCO3, 0.1mM EDTA,pH 7.4. The PBMC were maintained in RPMI 1640medium for 2 h to allow the adherence ofmonocytes to the plates and separation fromlymphocytes.

ARTICLE IN PRESS
R. Gorjao et al.926
Lipid extraction and determination of fattyacids by HPLC
The lipids were extracted from plasma and lym-phocytes as previously described.35 The lipids weresaponified using 2ml of an alkaline methanolsolution (1mol/ml NaOH in 90% methanol) at37 1C, for 2 h, in a shaking water bath. Afterwards,the alkaline solution was acidified to pH 3 with HClsolution (1mol/l). FA were then extracted threetimes with 2ml hexane. After the extractionprocedure and saponification,36–38 the FA werederivatized with 4-bromomethyl-7 coumarim39
and the analysis performed in a liquid chromato-grapher Shimadzu model LC-10A. The samples wereeluted using a C8 column (25 cm� 4.6 id, 5 mm ofparticles) with pre-column C8 (2.5 cm 4.6 id, 5 mmof particles), 1ml/min of acetonitrile/water(77:23) flow and fluorescence detector (325 nmexcitation and 395 emission). The standard FA(C17:0) were obtained from Sigma Chemical Co.Margaric acid was used to calculate recovery. Thecapacity factor (K0), elution sequence, linearity,recovery, precision, interference and limit ofdetection were determined. The minimum limit ofquantification of the FA ranged from 1 to 10 ng. Onecurve of calibration for each standard, determiningcoefficients of correlation and regression, wasprepared.
Phagocytic activity of monocytes andneutrophils
The zymosan particles (Zymosan A, an insolublelarge polysaccharide fraction of yeast cell walls)were suspended in PBS providing a concentration ofparticles of 5.7mg/ml. For opsonization, 2mlzymosan particles were mixed with 2ml normalhuman serum and incubated for 30min at 37 1C.40
The opsonized zymosan particles were then cen-trifuged at 180g for 10min in PBS and suspended inRPMI 1640 medium for the phagocytosis assay. Thenumber of particles per cover slip was of approxi-mately 2� 106.41
The coverslips containing adherent monocyteswere incubated in 1ml RPMI 1640 medium contain-ing opsonized zymosan for 40min, at 37 1C, in anatmosphere containing 5% CO2. Monocyte phagocy-tosis was performed in smears stained with apanchromatic dye.41
An aliquot of 2� 106 neutrophils were incubatedunder the same conditions described above. Neu-trophils were diluted (1:10) in a solution of Crystalviolet (in acetic acid) and counted in a Neubauerchamber.
The phagocytosis is expressed as the percentageof cells that had phagocytosed more than threeparticles (as determined by using a light micro-scope� 1000).
Chemotaxis assay
The neutrophil chemotactic response was studiedby using a modification of a Boyden chambertechnique as previously described.42 The assaywas carried out in transwell plates (CorningIncorporated, NY) using a filter with 8 mm poresize. Briefly, the upper compartment was filled with1� 106 cell suspension. The lower compartmentwas filled with activated plasma (containing com-ponent C5a of the complement system) or withminimal essential medium as control. The assayswere performed in triplicate. After incubation at371C for 180min, the filters were removed. Theresults are expressed in number of cells thatmigrated to the inferior compartment of thechamber. The assay containing RPMI medium with-out activated plasma in the inferior compartmentwas used for correction of background migration.
Production of reactive oxygen metabolitesby neutrophils
Hydroethidine was used for the flow cytometricmeasurement of reactive oxygen metabolites pro-duced by neutrophils. Hydroethidine is rapidlyoxidized to ethidium bromide (a red fluorescentcompound) by hydrogen peroxide (in the presenceof peroxidase) and KO2, source of superoxide.Samples containing 1� 106 neutrophils stimulatedwith phorbol myristate acetate (PMA-20 nM) andnon-stimulated were washed with PBS and thenincubated with hydroethidine (Polysciences, Niles,IL; 10 mM in PBS), for 30min, at 37 1C. Afterwashing, fluorescence was then evaluated by flowcytometry. The data are presented as the ratiowith the respective control (before treatment),which received an arbitrary value of 1 in eachexperiment.
Proliferation assay
Lymphocytes (3.3� 105 cells/ml) were plated in 96-well microtiter plates and cultured for a period of24 h in the presence of 5 mg/ml concanavalin A (ConA)—a T-lymphocyte mitogen.43 The plates wereincubated in a humidified atmosphere of 5% CO2
and 95% air at 37 1C. After 48 h, [2-14C] thymidine(1 mCi/ml) was added to the medium and the cellswere incubated for a further period of 18 h. The

ARTICLE IN PRESS
Effect of fish oil on human leukocyte function 927
radioactivity was analyzed by using a liquidscintillation counter (Packard TRI-CARB 2100 TRCounters; Downers Grove, IL).
Measurement of cytokines
Lymphocytes (2� 105 cells/ml) were plated in 24-well plates and cultured for a period of 24 h in thepresence of 5 mg/ml Con A. Afterwards, cell culturesupernatant was harvested to carry out thedetermination of secreted cytokines.
The production of IL-2, IL-10, IL-4, TNF-a andIFN-g was evaluated by enzyme-linked immunosor-bent assay (ELISA) using Kit OptEIATM from Pharmin-gen (San Diego, CA). The detection limit of IL-2, IL-10, IL-4 and TNF-a was 7.8 pg/ml and of INF-g was4.7 pg/ml according to the manufacturer.
Values are presented as the difference betweencytokine production before and after 2 months ofsupplementation with DHA-rich FO.
Analysis of pleitropic gene expression inlymphocytes
Gene expression in lymphocytes isolated beforeand after 2 months of supplementation with FO wasanalyzed by macroarray technique. For this analy-sis, a single pool of 20 mg total RNA was prepared,consisting of equal amounts (2 mg) of lymphocyteRNA from each of the ten volunteers.
Total RNA extraction
Total RNA was obtained from 1� 107 cells usingTrizol reagentTM (Invitogen, Carlsbad, CA). Briefly,the cells were lysed using 1ml Trizol reagent(Invitrogen). After 5min incubation at room tem-perature, 200 ml chloroform were added to thetubes and centrifuged at 12,000g. The aqueousphase was transferred to another tube and the RNAwas pelleted by centrifugation (12,000g) with coldethanol and dried in air. RNA pellets were eluatedin RNase-free water and treated with DNase I. TheRNA preparation was then stored at �70 1C until tothe time of the experiment. The RNA was quanti-fied by measuring absorbance at 260 nm. The purityof the RNA preparation was assessed by the 260/280 nm ratio and on a 1% agarose gel stained withethidium bromide at 5 mg/ml.44 These sampleswere used for macroarray and RT-PCR analysis.
Synthesis of cDNA probes
The cDNA probes were synthesized using the puretotal RNA labeling system Atlas KitTM according to
manufacturer’s recommendations (ClontechLaboratories, California, USA). Briefly, 15 mgof a pool of total RNA of the eight volunteers and2 ml primers mix ‘‘CDs’’ (a mixture of primersrelative to the genes spotted in macroarraymembrane) were heated at 70 1C for 5min in aTechne-Genius Thermal cycler (Oxford, UK).The temperature was decreased to 50 1C and13.5 ml of the mix of the following reagents wereadded: 4 ml reaction buffer 5� , 0.5 ml 100mM DTT,2 ml 10� dNTP mix (dCTP, dGTP, dTTP), 5 ml [a-33P]ATP (at 10 mCi/ml) and 2 ml reverse superscriptenzyme (Invitrogen). The reaction was incubatedfor 25min at 50 1C and stopped by using 2 mlTermination Mix. The 33P-labeled probe was pur-ified from unincorporated nucleotides by passingthe reaction mixture through a push column(NucleoSpin Extraction Spin Column—ClontechLaboratories).
Macroarray hibridization
All solutions for hybridization were obtained fromthe Clontech Laboratories. The list of genes in themacroarray membranes is available on the ClontechWebsite (www.bdbiosciences.com/clontech/atlas/genelists/7733-1_HuTox2.pdf). The membrane waspre-hybridized for 30min at 68 1C in Express Hybcontaining 50 mg freshly denaturated salmon spermDNA. Subsequently, the membrane was hybridizedduring 18 h at 68 1C with 33P-labeled denaturatedprobe (2� 106 cpm/ml). The membrane waswashed four times at 68 1C with 1� SSC, 0.1% SDS;followed by one washing in 1� SSC, 1% SDS andthen exposed to phosphor screen for 48 h andscanned in the Storm 840 (Molecular Dynamics,Sunnyvale, CA).
Analysis of macroarray results
Changes in lymphocyte gene expression induced bysupplementation with FO for 2 months wereanalyzed by comparison with the expressionobserved before the supplementation using thesoftware Array-ProTM Analyzer, version 4 (MediaCyberneticss, Silver Spring, MD). Normalizationwas performed by housekeeping genes presentin the membrane. Triplicate hybridizationsusing separate sets of nylon membranes wereperformed for all conditions. Only signalsthat differed from the control by at least 2-fold inthe four independent experiments were consideredas significant. Yamazak et al.45 used a similarprocedure.

ARTICLE IN PRESS
R. Gorjao et al.928
RT-PCR
RT-PCR using specific primers was performed toconfirm the differential expression of the mRNAsdetected with the macroarray analysis. The se-quences of the primers were designed usinginformation of the public database in the GeneBankof the National Center for Biotechnology Informa-tion (NCBI).
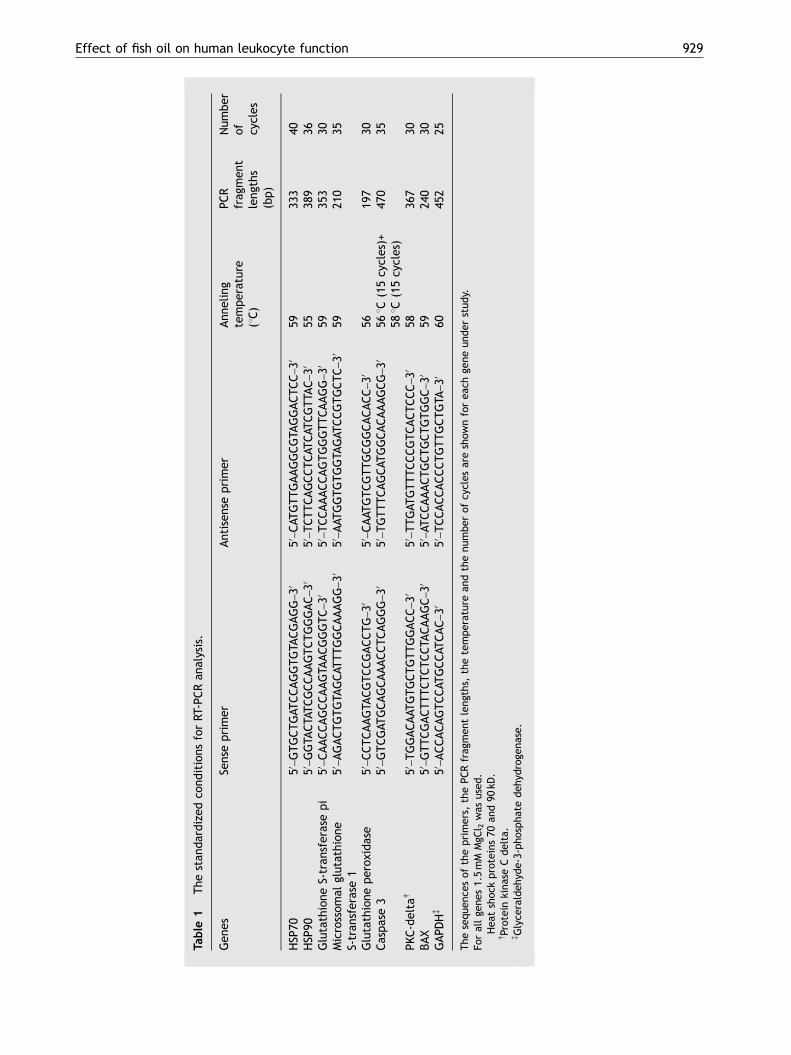
The RT-PCR was performed using parametersdescribed by Innis and Gelfand.46 The number ofcycles allowed linear amplification of the cDNAunder study. For semi-quantitative PCR analysis,the housekeeping glyceraldehyde-3-phosphate de-hydrogenase (GAPDH) gene was used as reference.The primer sequences and their respective PCRfragment lengths are shown in the Table 1.Published guidelines were followed to guard againstbacterial and nucleic acid contamination.47
Analysis of the PCR products
The analysis of PCR amplification products wasperformed in 1.5% agarose gels containing 0.5 mg/ml ethidium bromide and electrophoresed for 1 h at100 V. The gels were photographed using a DC120Zoom Digital Camera System from Kodak (Invitro-gen). The images were processed and analyzed inthe software Kodak Digital Science 1D ImageAnalysis (Invitrogen).
PCR band intensities were expressed as ODnormalized for GAPDH expression. The data arepresented as the ratio with the respective controls,which received an arbitrary value of 1 in eachexperiment.
Statistical analysis
The results of FA composition, lymphocyte prolif-eration, monocyte and neutrophil phagocytosis,neutrophil chemiotaxis, production of reactiveoxygen metabolites and RT-PCR analysis are ex-pressed as mean7SEM. Comparisons betweengroups were analyzed using the paired t-test(INStat—Graph Pad Software Inc., San Diego, CA).Results were considered statistically significant forPo0:05.
Results
Fatty acid composition
The FA composition of plasma and leukocyte lipidswas clearly changed after 2 months of DHA-rich FO
supplementation. The AA content was decreasedsignificantly in neutrophils. This decrease wasaccompanied by an increase in n-3 FA content(data not shown). The n-3/n-6 ratio increasedqfrom 0.15 to 0.70. A similar effect was observedin FA composition of lymphocytes where then-3/n-6 ratio increased from 0.18 to 0.62 aftersupplementation.
Phagocytic activity of monocytes andneutrophils
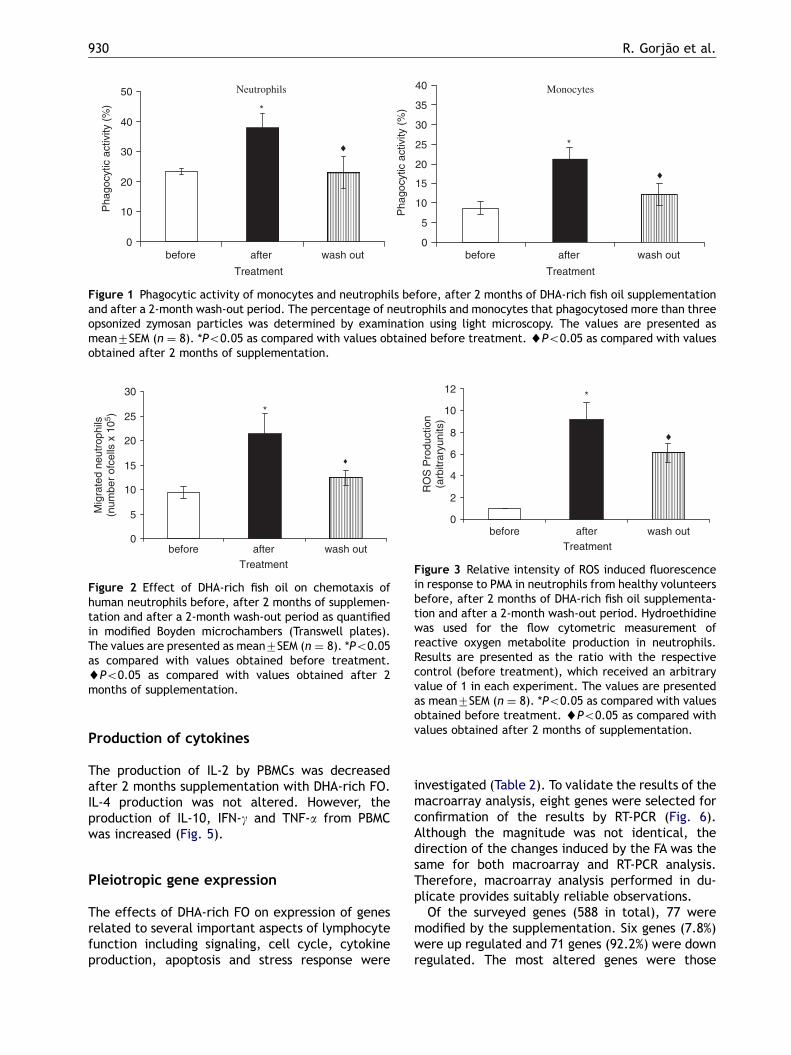
DHA-rich FO supplementation significantly stimu-lated phagocytic activity of neutrophils and mono-cytes (Fig. 1). The percentage of cell engaging inphagocytosis was increased by 62% and 145% inneutrophils and monocytes, respectively (Fig. 1).Both neutrophil and monocyte phagocytosis wasreturned towards normal after a 2-month wash-outperiod.
Chemotaxis
Neutrophil chemotactic responsiveness to comple-ment C5a was significantly increased (by 128%)after 2 months of FO supplementation (Fig. 2); from9.4� 105 migrated cells to 2.1� 106. Neutrophilmigration was returned towards normal after a2-month wash-out period (11.4� 105).
Production of reactive oxygen metabolitesby neutrophils
The fluorescence intensity (proportional to ROSproduction) in response to PMA was increasedin neutrophils isolated from volunteers after 2months of DHA-rich FO supplementation (Fig. 3).Therefore, DHA-rich FO promoted an increase inthe responsiveness of neutrophils to PMA stimula-tion. This enhanced responsiveness was alsoreduced towards normal after a 2-month wash-outperiod.
Lymphocyte proliferation
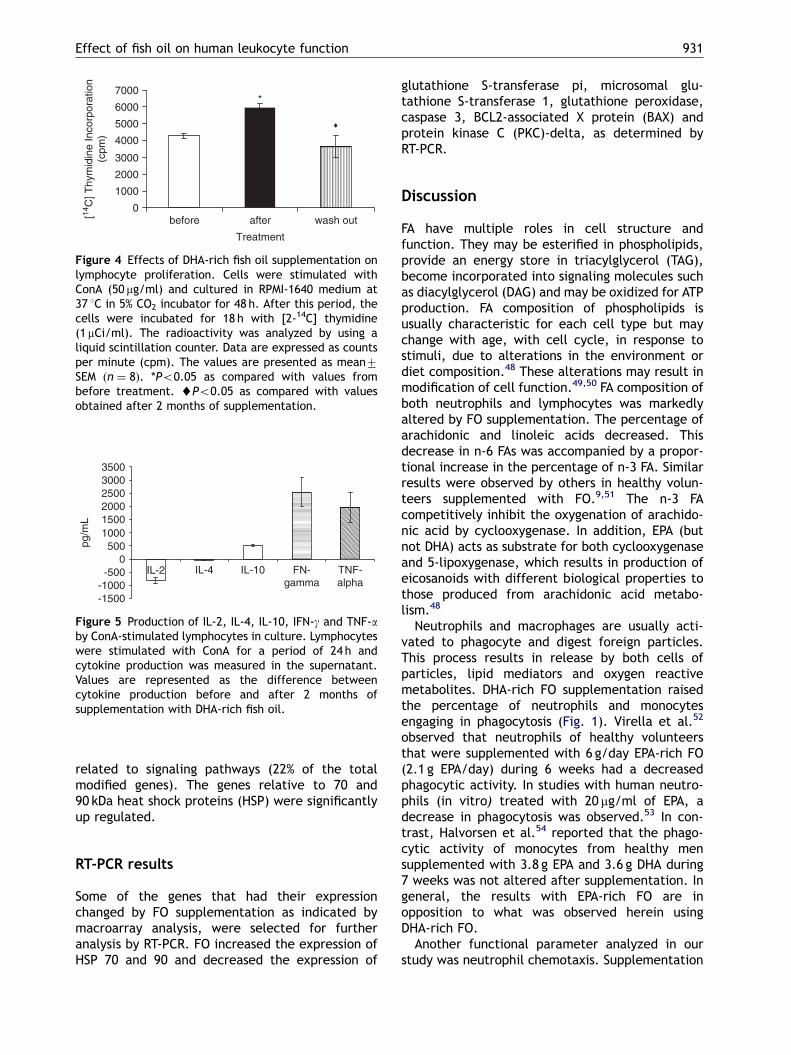
The supplementation with DHA-rich FO promotedan increase of 40% in ConA-dependent lymphocyteproliferative capacity (Fig. 4). These results sug-gest that DHA-rich FO stimulates the proliferativeresponse of lymphocytes to ConA. This enhancedresponse was reduced to normal levels after a2-month wash-out period.
ARTICLE IN PRESS
Table
1The
stan
dardized
cond
itions
forRT
-PCRan
alysis.
Gen
esSe
nseprimer
Antisen
seprimer
Ann
eling
temperature
(1C)
PCR
frag
men
tleng
ths
(bp)
Num
ber
of cycles
HSP
70�
50–GTGCTGAT
CCAGGTGTA
CGAGG
–30
50–CAT
GTTGAAGGCGTA
GGACTCC
–30
5933
340
HSP
90�
50–GGTA
CTATCGCCAAGTCTGGGAC
–30
50–TCTTCAGCCTCAT
CAT
CGTTA
C–30
5538
936
Glutathione
S-tran
sferasepi
50–CAACCAGCCAAGTA
ACGGGTC
–30
50–TCCAAACCAGTGGGTTCAAGG
–30
5935
330
Microssom
alglutathion
eS-tran
sferase1
50–AGACTGTGTA
GCAT
TTGGCAAAGG
–30
50–AAT
GGTGTGGTA
GAT
CCGTGCTC
–30
5921
035
Glutathione
perox
idase
50–CCTCAAGTA
CGTCCGACCTG
–30
50–CAAT
GTCGTTGCGGCACACC
–30
5619
730
Caspase3
50–GTCGAT
GCAGCAAACCTCAGGG
–30
50–TGTTTCAGCAT
GGCACAAAGCG
–30
561C(15cycles)+
581C(15cycles)
470
35
PKC-deltay
50–TGGACAAT
GTGCTGTTGGACC
–30
50–TTGAT
GTTTCCCGTCACTCCC
–30
5836
730
BAX
50–GTTCGACTTTCTCTCCTA
CAAGC
–30
50–AT
CCAAACTGCTGCTGTGGC
–30
5924
030
GAPD
Hz
50–ACCACAGTCCAT
GCCAT
CAC
–30
50–TCCACCACCCTGTTGCTGTA
–30
6045
225
The
sequ
ence
sof
theprimers,
thePCRfrag
men
tleng
ths,
thetempe
rature
andthenu
mbe
rof
cycles
areshow
nforea
chge
neun
der
stud
y.Fo
rallge
nes1.5mM
MgC
l 2was
used
.�Hea
tshoc
kproteins70
and90
kD.
yProteinkina
seCdelta.
zGlyce
raldeh
yde-3-ph
ospha
tedeh
ydroge
nase.
Effect of fish oil on human leukocyte function 929
ARTICLE IN PRESS
0
10
20
30
40
50
before after wash out
Pha
gocy
tic a
ctiv
ity (
%) *
♦
Neutrophils
0
5
10
15
20
25
30
35
40
before after wash out
Pha
gocy
tic a
ctiv
ity (
%)
*
♦
Monocytes
Treatment Treatment
Figure 1 Phagocytic activity of monocytes and neutrophils before, after 2 months of DHA-rich fish oil supplementationand after a 2-month wash-out period. The percentage of neutrophils and monocytes that phagocytosed more than threeopsonized zymosan particles was determined by examination using light microscopy. The values are presented asmean7SEM (n ¼ 8). *Po0.05 as compared with values obtained before treatment. ~Po0.05 as compared with valuesobtained after 2 months of supplementation.
0
5
10
15
20
25
30
before after wash out
Mig
rate
d ne
utro
phils
(num
ber
ofce
lls x
105 )
*
♦
Treatment
Figure 2 Effect of DHA-rich fish oil on chemotaxis ofhuman neutrophils before, after 2 months of supplemen-tation and after a 2-month wash-out period as quantifiedin modified Boyden microchambers (Transwell plates).The values are presented as mean7SEM (n ¼ 8). *Po0.05as compared with values obtained before treatment.~Po0.05 as compared with values obtained after 2months of supplementation.
0
2
4
6
8
10
12
before after wash out
RO
S P
rodu
ctio
n(a
rbitr
aryu
nits
)
*
♦
Treatment
Figure 3 Relative intensity of ROS induced fluorescencein response to PMA in neutrophils from healthy volunteersbefore, after 2 months of DHA-rich fish oil supplementa-tion and after a 2-month wash-out period. Hydroethidinewas used for the flow cytometric measurement ofreactive oxygen metabolite production in neutrophils.Results are presented as the ratio with the respectivecontrol (before treatment), which received an arbitraryvalue of 1 in each experiment. The values are presentedas mean7SEM (n ¼ 8). *Po0.05 as compared with valuesobtained before treatment. ~Po0.05 as compared withvalues obtained after 2 months of supplementation.
R. Gorjao et al.930
Production of cytokines
The production of IL-2 by PBMCs was decreasedafter 2 months supplementation with DHA-rich FO.IL-4 production was not altered. However, theproduction of IL-10, IFN-g and TNF-a from PBMCwas increased (Fig. 5).
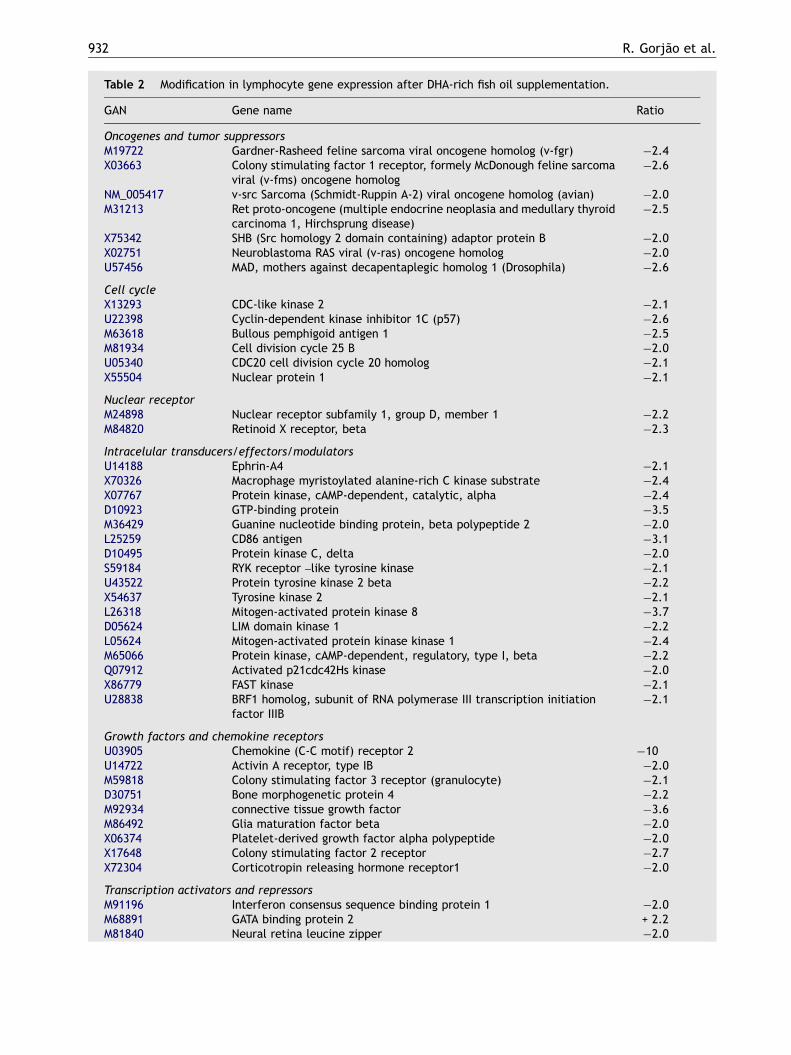
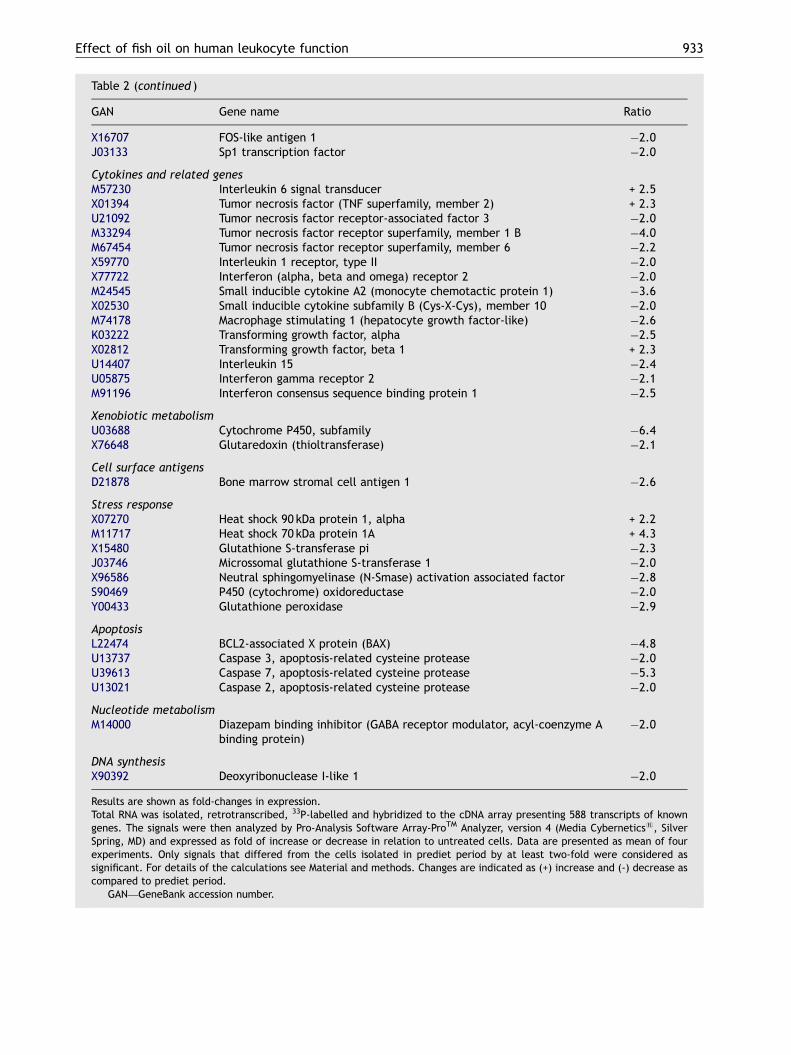
Pleiotropic gene expression
The effects of DHA-rich FO on expression of genesrelated to several important aspects of lymphocytefunction including signaling, cell cycle, cytokineproduction, apoptosis and stress response were
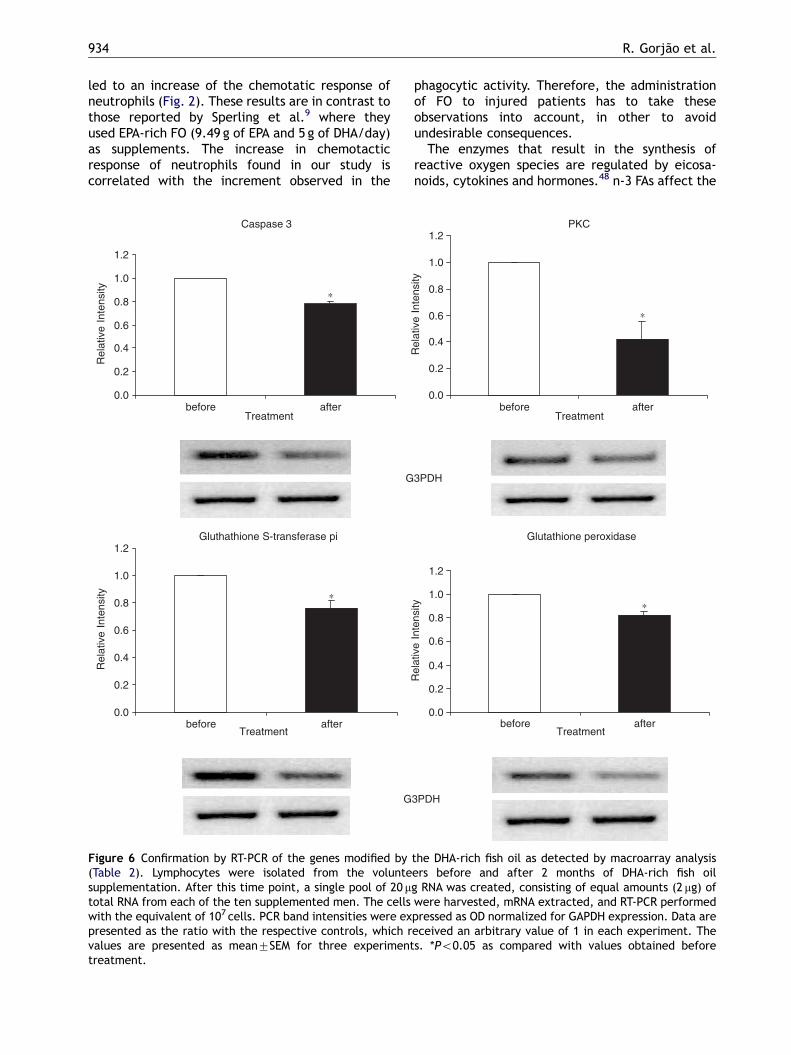
investigated (Table 2). To validate the results of themacroarray analysis, eight genes were selected forconfirmation of the results by RT-PCR (Fig. 6).Although the magnitude was not identical, thedirection of the changes induced by the FA was thesame for both macroarray and RT-PCR analysis.Therefore, macroarray analysis performed in du-plicate provides suitably reliable observations.
Of the surveyed genes (588 in total), 77 weremodified by the supplementation. Six genes (7.8%)were up regulated and 71 genes (92.2%) were downregulated. The most altered genes were those
ARTICLE IN PRESS
-1500-1000-500
0500
100015002000250030003500
IL-2 IL-4 IL-10 FN-gamma
TNF-alpha
pg/m
L
Figure 5 Production of IL-2, IL-4, IL-10, IFN-g and TNF-aby ConA-stimulated lymphocytes in culture. Lymphocyteswere stimulated with ConA for a period of 24 h andcytokine production was measured in the supernatant.Values are represented as the difference betweencytokine production before and after 2 months ofsupplementation with DHA-rich fish oil.
0
1000
2000
3000
4000
5000
6000
7000
before after wash out
Treatment
[14C
] Thy
mid
ine
Inco
rpor
atio
n(c
pm)
*
♦
Figure 4 Effects of DHA-rich fish oil supplementation onlymphocyte proliferation. Cells were stimulated withConA (50 mg/ml) and cultured in RPMI-1640 medium at37 1C in 5% CO2 incubator for 48 h. After this period, thecells were incubated for 18 h with [2-14C] thymidine(1mCi/ml). The radioactivity was analyzed by using aliquid scintillation counter. Data are expressed as countsper minute (cpm). The values are presented as mean7SEM ðn ¼ 8Þ. *Po0.05 as compared with values frombefore treatment. ~Po0.05 as compared with valuesobtained after 2 months of supplementation.
Effect of fish oil on human leukocyte function 931
related to signaling pathways (22% of the totalmodified genes). The genes relative to 70 and90 kDa heat shock proteins (HSP) were significantlyup regulated.
RT-PCR results
Some of the genes that had their expressionchanged by FO supplementation as indicated bymacroarray analysis, were selected for furtheranalysis by RT-PCR. FO increased the expression ofHSP 70 and 90 and decreased the expression of
glutathione S-transferase pi, microsomal glu-tathione S-transferase 1, glutathione peroxidase,caspase 3, BCL2-associated X protein (BAX) andprotein kinase C (PKC)-delta, as determined byRT-PCR.
Discussion
FA have multiple roles in cell structure andfunction. They may be esterified in phospholipids,provide an energy store in triacylglycerol (TAG),become incorporated into signaling molecules suchas diacylglycerol (DAG) and may be oxidized for ATPproduction. FA composition of phospholipids isusually characteristic for each cell type but maychange with age, with cell cycle, in response tostimuli, due to alterations in the environment ordiet composition.48 These alterations may result inmodification of cell function.49,50 FA composition ofboth neutrophils and lymphocytes was markedlyaltered by FO supplementation. The percentage ofarachidonic and linoleic acids decreased. Thisdecrease in n-6 FAs was accompanied by a propor-tional increase in the percentage of n-3 FA. Similarresults were observed by others in healthy volun-teers supplemented with FO.9,51 The n-3 FAcompetitively inhibit the oxygenation of arachido-nic acid by cyclooxygenase. In addition, EPA (butnot DHA) acts as substrate for both cyclooxygenaseand 5-lipoxygenase, which results in production ofeicosanoids with different biological properties tothose produced from arachidonic acid metabo-lism.48
Neutrophils and macrophages are usually acti-vated to phagocyte and digest foreign particles.This process results in release by both cells ofparticles, lipid mediators and oxygen reactivemetabolites. DHA-rich FO supplementation raisedthe percentage of neutrophils and monocytesengaging in phagocytosis (Fig. 1). Virella et al.52
observed that neutrophils of healthy volunteersthat were supplemented with 6 g/day EPA-rich FO(2.1 g EPA/day) during 6 weeks had a decreasedphagocytic activity. In studies with human neutro-phils (in vitro) treated with 20 mg/ml of EPA, adecrease in phagocytosis was observed.53 In con-trast, Halvorsen et al.54 reported that the phago-cytic activity of monocytes from healthy mensupplemented with 3.8 g EPA and 3.6 g DHA during7 weeks was not altered after supplementation. Ingeneral, the results with EPA-rich FO are inopposition to what was observed herein usingDHA-rich FO.
Another functional parameter analyzed in ourstudy was neutrophil chemotaxis. Supplementation
ARTICLE IN PRESS
Table 2 Modification in lymphocyte gene expression after DHA-rich fish oil supplementation.
GAN� Gene name Ratio
Oncogenes and tumor suppressorsM19722 Gardner-Rasheed feline sarcoma viral oncogene homolog (v-fgr) �2.4X03663 Colony stimulating factor 1 receptor, formely McDonough feline sarcoma
viral (v-fms) oncogene homolog�2.6
NM_005417 v-src Sarcoma (Schmidt-Ruppin A-2) viral oncogene homolog (avian) �2.0M31213 Ret proto-oncogene (multiple endocrine neoplasia and medullary thyroid
carcinoma 1, Hirchsprung disease)�2.5
X75342 SHB (Src homology 2 domain containing) adaptor protein B �2.0X02751 Neuroblastoma RAS viral (v-ras) oncogene homolog �2.0U57456 MAD, mothers against decapentaplegic homolog 1 (Drosophila) �2.6
Cell cycleX13293 CDC-like kinase 2 �2.1U22398 Cyclin-dependent kinase inhibitor 1C (p57) �2.6M63618 Bullous pemphigoid antigen 1 �2.5M81934 Cell division cycle 25 B �2.0U05340 CDC20 cell division cycle 20 homolog �2.1X55504 Nuclear protein 1 �2.1
Nuclear receptorM24898 Nuclear receptor subfamily 1, group D, member 1 �2.2M84820 Retinoid X receptor, beta �2.3
Intracelular transducers/effectors/modulatorsU14188 Ephrin-A4 �2.1X70326 Macrophage myristoylated alanine-rich C kinase substrate �2.4X07767 Protein kinase, cAMP-dependent, catalytic, alpha �2.4D10923 GTP-binding protein �3.5M36429 Guanine nucleotide binding protein, beta polypeptide 2 �2.0L25259 CD86 antigen �3.1D10495 Protein kinase C, delta �2.0S59184 RYK receptor –like tyrosine kinase �2.1U43522 Protein tyrosine kinase 2 beta �2.2X54637 Tyrosine kinase 2 �2.1L26318 Mitogen-activated protein kinase 8 �3.7D05624 LIM domain kinase 1 �2.2L05624 Mitogen-activated protein kinase kinase 1 �2.4M65066 Protein kinase, cAMP-dependent, regulatory, type I, beta �2.2Q07912 Activated p21cdc42Hs kinase �2.0X86779 FAST kinase �2.1U28838 BRF1 homolog, subunit of RNA polymerase III transcription initiation
factor IIIB�2.1
Growth factors and chemokine receptorsU03905 Chemokine (C-C motif) receptor 2 �10U14722 Activin A receptor, type IB �2.0M59818 Colony stimulating factor 3 receptor (granulocyte) �2.1D30751 Bone morphogenetic protein 4 �2.2M92934 connective tissue growth factor �3.6M86492 Glia maturation factor beta �2.0X06374 Platelet-derived growth factor alpha polypeptide �2.0X17648 Colony stimulating factor 2 receptor �2.7X72304 Corticotropin releasing hormone receptor1 �2.0
Transcription activators and repressorsM91196 Interferon consensus sequence binding protein 1 �2.0M68891 GATA binding protein 2 + 2.2M81840 Neural retina leucine zipper �2.0
R. Gorjao et al.932
ARTICLE IN PRESS
Table 2 (continued )
GAN� Gene name Ratio
X16707 FOS-like antigen 1 �2.0J03133 Sp1 transcription factor �2.0
Cytokines and related genesM57230 Interleukin 6 signal transducer + 2.5X01394 Tumor necrosis factor (TNF superfamily, member 2) + 2.3U21092 Tumor necrosis factor receptor-associated factor 3 �2.0M33294 Tumor necrosis factor receptor superfamily, member 1 B �4.0M67454 Tumor necrosis factor receptor superfamily, member 6 �2.2X59770 Interleukin 1 receptor, type II �2.0X77722 Interferon (alpha, beta and omega) receptor 2 �2.0M24545 Small inducible cytokine A2 (monocyte chemotactic protein 1) �3.6X02530 Small inducible cytokine subfamily B (Cys-X-Cys), member 10 �2.0M74178 Macrophage stimulating 1 (hepatocyte growth factor-like) �2.6K03222 Transforming growth factor, alpha �2.5X02812 Transforming growth factor, beta 1 + 2.3U14407 Interleukin 15 �2.4U05875 Interferon gamma receptor 2 �2.1M91196 Interferon consensus sequence binding protein 1 �2.5
Xenobiotic metabolismU03688 Cytochrome P450, subfamily �6.4X76648 Glutaredoxin (thioltransferase) �2.1
Cell surface antigensD21878 Bone marrow stromal cell antigen 1 �2.6
Stress responseX07270 Heat shock 90 kDa protein 1, alpha + 2.2M11717 Heat shock 70 kDa protein 1A + 4.3X15480 Glutathione S-transferase pi �2.3J03746 Microssomal glutathione S-transferase 1 �2.0X96586 Neutral sphingomyelinase (N-Smase) activation associated factor �2.8S90469 P450 (cytochrome) oxidoreductase �2.0Y00433 Glutathione peroxidase �2.9
ApoptosisL22474 BCL2-associated X protein (BAX) �4.8U13737 Caspase 3, apoptosis-related cysteine protease �2.0U39613 Caspase 7, apoptosis-related cysteine protease �5.3U13021 Caspase 2, apoptosis-related cysteine protease �2.0
Nucleotide metabolismM14000 Diazepam binding inhibitor (GABA receptor modulator, acyl-coenzyme A
binding protein)�2.0
DNA synthesisX90392 Deoxyribonuclease I-like 1 �2.0
Results are shown as fold-changes in expression.Total RNA was isolated, retrotranscribed, 33P-labelled and hybridized to the cDNA array presenting 588 transcripts of knowngenes. The signals were then analyzed by Pro-Analysis Software Array-ProTM Analyzer, version 4 (Media Cyberneticss, SilverSpring, MD) and expressed as fold of increase or decrease in relation to untreated cells. Data are presented as mean of fourexperiments. Only signals that differed from the cells isolated in prediet period by at least two-fold were considered assignificant. For details of the calculations see Material and methods. Changes are indicated as (+) increase and (-) decrease ascompared to prediet period.�GAN—GeneBank accession number.
Effect of fish oil on human leukocyte function 933
ARTICLE IN PRESS
R. Gorjao et al.934
led to an increase of the chemotatic response ofneutrophils (Fig. 2). These results are in contrast tothose reported by Sperling et al.9 where theyused EPA-rich FO (9.49 g of EPA and 5 g of DHA/day)as supplements. The increase in chemotacticresponse of neutrophils found in our study iscorrelated with the increment observed in the
Caspase 3
0.0
0.2
0.4
0.6
0.8
1.0
1.2
before afterTreatment
Rel
ativ
e In
tens
ity
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Rel
ativ
e In
tens
ity
∗
G
Gluthathione S-transferase pi
before afterTreatment
∗
G
Figure 6 Confirmation by RT-PCR of the genes modified by(Table 2). Lymphocytes were isolated from the voluntesupplementation. After this time point, a single pool of 20 mtotal RNA from each of the ten supplemented men. The cellswith the equivalent of 107 cells. PCR band intensities were expresented as the ratio with the respective controls, which rvalues are presented as mean7SEM for three experimenttreatment.
phagocytic activity. Therefore, the administrationof FO to injured patients has to take theseobservations into account, in other to avoidundesirable consequences.
The enzymes that result in the synthesis ofreactive oxygen species are regulated by eicosa-noids, cytokines and hormones.48 n-3 FAs affect the
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Rel
ativ
e In
tens
ity
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Rel
ativ
e In
tens
ity
PKC
before afterTreatment
∗
3PDH
Glutathione peroxidase
before afterTreatment
∗
3PDH
the DHA-rich fish oil as detected by macroarray analysisers before and after 2 months of DHA-rich fish oilg RNA was created, consisting of equal amounts (2 mg) ofwere harvested, mRNA extracted, and RT-PCR performedpressed as OD normalized for GAPDH expression. Data areeceived an arbitrary value of 1 in each experiment. Thes. *Po0.05 as compared with values obtained before

ARTICLE IN PRESS
HSP70
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
before afterTreatment
Rel
ativ
e In
tens
ity
0.0
0.2
0.4
0.6
0.8
1.0
1.6
1.4
1.2
Rel
ativ
e In
tens
ity
∗HSP90
before afterTreatment
∗
G3PDH
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Rel
ativ
e In
tens
ity
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Rel
ativ
e In
tens
ity
Gluthathione S-transferase pi
before afterTreatment
∗
Bax
before afterTreatment
∗
G3PDH
Figure 6 (Continued)
Effect of fish oil on human leukocyte function 935
production of eicosanoids and cytokines and thusmight modulate PKC activity. Therefore, these FAmay regulate the production of reactive oxygenspecies by phagocytes via PKC and NADPH oxidase.In our study, the production of reactive oxygenspecies by neutrophils stimulated with PMA wasincreased after supplementation with DHA-rich FO.Poulos et al.55 observed that EPA and DHA potenti-ate PMA-stimulated ROS production by humanneutrophils. On the other hand, Chen et al.56
observed a decrease in the production of ROS by
human neutrophils incubated in the presence ofEPA and DHA. Therefore, supplementationcarried out with FOs containing different composi-tion of DHA and EPA may result in different effects.The results observed in our study suggest thatDHA-rich FO may increase the cytolytic activity ofneutrophils.
The effects of FO on lymphocyte function are stillcontroversial. Supplementation with EPA-rich FOinhibited lymphocyte proliferation in elderlyhealthy women (age between 51 and 68 years)

ARTICLE IN PRESS
R. Gorjao et al.936
but this effect was not observed in young women(age between 21 and 33 years).15 In other studies, adecrease in ConA stimulated human lymphocyteproliferation occurred after supplementation with1.2 g/day of EPA-rich FO.16,57 However, enhancedproliferation was not observed in lymphocytes ofhuman volunteers supplemented with high concen-trations of EPA and DHA.58 In our study, DHA-rich FOsupplementation promoted a marked increase inConA stimulated lymphocyte proliferation. Theenhanced functional responses described in thisstudy were reduced significantly after a 2-monthwash-out period, demonstrating a FA-induced ef-fect which was reversible.
A stimulatory effect of DHA-rich FO on TNF-a,IFN-g and IL-10 production by ConA stimulatedlymphocytes is reported herein. It is noteworthythat the expression of TNF-a was increased asevaluated by macroarray analysis. In spite of this,the expression of TNF receptor was decreased. Thisfinding may limit the effect of TNF-a under theseconditions. As reported by others, EPA-rich FOsupplementation does not show consistent changesin TNF-a production.10,18 An increase of IFN-gproduction after DHA-rich FO supplementation wasalso observed by Xi et al.59 that studied the effects ofFO supplementation in mice with AIDS. On the otherhand, other author reported that FO supplementa-tion did not alter IFN-g production by humanlymphocytes.16 The inconsistent results from thesestudies could be due to healthy versus infected state,as the immune function of subjects is different inthese two states, or the level of FO (proportion ofEPA and DHA) or vitamin E ingested is not the same
We observed an overall suppression of geneexpression (92.2%), in contrast to an increase of7.8% in expression of detected genes. To validatethe results of the macroarray analysis, eight geneswere selected for confirmation by RT-PCR (Fig. 6).Although the magnitude was not the same, thedirection of changes induced by the FA wasconsistent in both macroarray and RT-PCR analysis.
The DHA-rich FO supplementation responsivegenes were clustered according to main putativebiological functions of their encoded proteins. Thegenes most altered by the FO supplementationwere related to signal transduction. In studies byothers, n-3 FAs regulated specific aspects of cellsignaling, as enzyme activity such as protein kinaseC.60 Denys et al.61 reported that EPA and DHAdecrease MAP kinase activation in T lymphocytes.In our study, DHA-rich FO supplementation inhib-ited the expression of MAP kinase kinase I, proteinkinase C delta, MAP kinase 8, cAMP-dependentprotein kinase at the transcriptional level. It ispossible that polyunsaturated FA may exert effects
on cell signaling by activation of transcriptionfactors such as peroxisome proliferator activatedreceptors (PPARs).62,63
Our study has shown that n-3 FAs are involved inthe regulation of genes related to cellular defense.The expression of HSP 70 and 90 was increased byFO supplementation. In the unstressed cell, theseproteins have constitutive functions that areessential in various aspects of protein metabo-lism.64 HSP 90 is active in supporting variouscomponents of cytoskeleton and steroid hormonereceptors. HSP 70 is known to mediate repair anddegradation of altered or denaturated proteins.65
The HSPs are known to be involved with immuneresponse in mammals.65 These proteins may also beimportant in providing maintenance and protectionto phagocytic cells66 by repairing damage orprotecting against autolysis or apoptosis due toauto-oxidation brought about by the cells owninternal defense systems.66 DHA accelerated theenhancement of HSP 70 mRNA expression in rain-bow trout leukocytes when these animals weresubmitted to a heat shock.67
DHA-rich FO supplementation suppressed expres-sion of pro-apoptotic genes such as caspases 3, 7and 2. However, the expression of BAX (an anti-apoptotic gene) was also suppressed. In otherstudies, Heimli et al.68 analyzed the direct effectof EPA on apoptosis of cultured leukemia/lympho-ma cells. EPA increased the activity of caspases 3and 9, but not of caspase 8. Further work isrequired to elucidate the mechanism by which FAmodulate leukocyte apoptosis.
The findings of our study have demonstrated thatFOs containing different composition of n-3 FAs canexert divergent effects on immune response. EPA-rich FO exhibits immunossupressor effects, whereasevidence is presented herein that DHA-rich FOstimulate several aspects of immune function.Thus, the selection of DHA or EPA-rich FOs mustbe considered carefully before being used tomodulate inflammatory diseases.
Acknowledgments
The authors are indebted to J.R. Mendonc-a and G.de Souza for technical assistance. This research issupported by FAPESP, CAPES and CNPq.
References
1. Bang H O, Dyerberg J. Plasma lipids and lipoproteins inGreenlandic west coast Eskimos. Acta Med Scand1972;192:85–94.

ARTICLE IN PRESS
Effect of fish oil on human leukocyte function 937
2. De Caterina R, Madonna R, Zucchi R, La Rovere MT.Antiarrhythmic effects of omega-3 fatty acids: from epide-miology to bedside. Am Heart J 2003;146:420–30.
3. Mayser P, Grimm H, Grimminger F. n-3 fatty acids inpsoriasis. Br J Nutr 2002;87:S77–82.
4. Kremer J M. n-3 Fatty acid supplements in rheumatoidarthritis. Am J Clin Nutr 2000;71:S349–51.
5. Calder P C. n-3 Fatty acids and cardiovascular disease:evidence explained and mechanisms explored. Clin Sci2004;107:1–11.
6. Colussi G L, Baroselli S, Sechi L. Omega-3 polyunsaturatedfatty acids decrease plasma lipoprotein(a) levels in hyper-tensive subjects. Clin Nutr 2004;23(5):1246–7.
7. Woodman RJ, Mori TA, Burke V, et al. Effects of purifiedeicosapentaenoic acid and docosahexaenoic acid on plate-let, fibrinolytic and vascular function in hypertensive type 2diabetic patients. Atherosclerosis 2003;166:85–93.
8. Simopoulos AP. Omega-3 fatty acids in inflammationand autoimmune diseases. J Am Coll Nutr 2002;21:495–505.
9. Sperling RI, Benincaso AI, Knoell CT, Larkin JK. Dietary $-3polyunsaturated fatty acids inhibit phosphoinositide forma-tion and chemotaxis in neutrophils. J Clin Invest 1993;91:651–60.
10. Yaqoob P, Pala HS, Cortina-Borja M, Newsholme EA, CalderPC. Encapsulated fish oil enriched in a-tocopherol altersplasma phospholipid and mononuclear cell fatty acidcompositions but not mononuclear cell functions. Eur J ClinInvest 2000;30:260–74.
11. Calder PC. n-3 Fatty acids, inflammation, and immunity-relevance to postsurgical and critically ill patients. Lipids2004;39(12):1147–61.
12. Terada S, Takizawa M, Yamamoto S, Ezaki O, Itakura H,Akagawa KS. Suppressive mechanisms of EPA on human Tcellproliferation. Microbiol Immunol 2001;45:473–81.
13. Endres S, Ghorbani R, Kelley VE, et al. The effect of dietarysupplementation with n-3 polyunsaturated fatty acids on thesynthesis of interleukin-1 and tumor necrosis factor bymononuclear cells. N Engl J Med 1989;320:265–71.
14. Mayer K, Gokorsch S, Fegbeutel C, et al. Parenteral nutritionwith fish oil modulates cytokine response in patients withsepsis. Am J Respir Crit Care Med 2003;167:1321–8.
15. Meydani SN, Endres S, Woods MM, et al. Oral (n-3) fatty acidsupplementation suppresses cytokine production and lym-phocyte proliferation: comparison between young and olderwomen. J Nutr 1991;121:547–55.
16. Thies F, Nebe-von-Caron G, Powell JR, Yaqoob P, NewsholmeEA, Calder PC. Dietary supplementation with gamma-linolenic acid or fish oil decreases T lymphocyte prolifera-tion in healthy older humans. J Nutr 2001;131:1918–27.
17. Schauder P, Rohn U, Schafer G, Korff G, Schenk HD. Impactof fish oil enriched total parenteral nutrition on DNAsynthesis, cytokine release and receptor expression bylymphocytes in the postoperative period. Br J Nutr2002;87:S103–10.
18. Grimble RF, Howell WM, O’Reilly G, et al. The ability of fishoil to suppress tumor necrosis factor alpha production byperipheral blood mononuclear cells in healthy men isassociated with polymorphisms in genes that influencetumor necrosis factor alpha production. Am J Clin Nutr2002;76:454–9.
19. Markovic O, O’Reilly G, Fussell HM, et al. Role of singlenucleotide polymorphisms of pro-inflammatory cytokinegenes in the relationship between serum lipids andinflammatory parameters, and the lipid-lowering effect offish oil in healthy males. Clin Nutr 2004;23(5):1084–95.
20. Grimm H, Mayer K, Mayser P, Eigenbrodt E. Regulatorypotential of n-3 fatty acids in immunological and inflamma-tory processes. Br J Nutr 2002;87:S59–67.
21. Serhan CN, Arita M, Hong S, Gotlinger K. Resolvins,docosatrienes, and neuroprotectins, novel omega-3-derivedmediators, and their endogenous aspirin-triggered epimers.Lipids 2004;39(11):1125–32.
22. Lee TH, Hoover RL, Williams JD, et al. Effect of dietaryenrichment with eicosapentaenoic and docosahexaenoicacids on in vitro neutrophil and monocyte leukotrienegeneration and neutrophil function. N Engl J Med1985;312:1217–24.
23. Pompeia C, Lopes LR, Miyasaka CK, Procopio J, Sannomiya P,Curi R. Effect of fatty acids on leukocyte function. Braz JMed Biol Res 2000;33:1–14.
24. Calder PC. Immunoregulatory and anti-inflammatory effectsof n-3 polyunsaturated fatty acids. Braz J Med Biol Res1998;31:467–90.
25. Yagaloff KA, Franco L, Simko B, Burghardt B. Essential fattyacids are antagonists of the leukotriene B4 receptor.Prostaglandins Leukot Essent Fatty Acids 1995;52:293–7.
26. Graber R, Sumida C, Nunez EA. Fatty acids and cell signaltransduction. J Lipid Mediat Cell Signal 1994;9:91–116.
27. Camandola S, Leonarduzzi G, Musso T, et al. Nuclear factorkB is activated by arachidonic acid but not by eicosapen-taenoic acid. Biochem Biophys Res Commun 1996;229:643–7.
28. Sellmayer A, Danesch U, Weber PC. Effects of differentpolyunsaturated fatty acids on growth-related early geneexpression and cell growth. Lipids 1996;31:S37–40.
29. Verlengia R, Gorjao R, Kanunfre CC, et al. Effects of EPA andDHA on proliferation, cytokine production, and geneexpression in Raji cells. Lipids 2004;39:857–64.
30. Verlengia R, Gorjao R, Kanunfre CC, et al. Comparativeeffects of eicosapentaenoic acid and docosahexaenoicacid on proliferation, cytokine production, and pleiotropicgene expression in Jurkat cells. Nutr Biochem 2004;15:657–65.
31. Sasaki T, Kake Y, Kudoh K, Misawa Y, Shimizu J, Takita T.Effects of dietary docosahexaenoic acid on surface mole-cules involved in T cell proliferation. Biochem Biophys Acta1999;1436:519–30.
32. Volker DH, Fitzgerald PE, Garg ML. The eicosapentaenoic todocosahexaenoic acid ratio of diets affects the pathogenesisof arthritis in Lew/SSN rats. J Nutr 2000;130:559–65.
33. Kelley DS, Taylor PC, Nelson GJ, Mackey BE. Dietarydocosahexaenoic acid and immunocompetence in younghealthy men. Lipids 1998;33(6):559–66.
34. Hong YL, Yeh SL, Chang CY, Hu ML. Total plasma malondial-dehyde levels in 16 Taiwanese college students determinedby various thiobarbituric acid tests and an improved high-performance liquid chromatography-based method. ClinBiochem 2000;33:619–25.
35. Folch J, Lee M, Stanley GHA. A simple method for isolationpurification of total lipid from animal tissue. J Biol Chem1957;226:497–503.
36. Beyer ES, Jensen LN. Overestimation of the cholesterolcontent of egg. Agric Food Chem 1989;37:917.
37. Hamilton S, Hamilton RJ, Sewell PA. Extraction of lipids andderivate formation. In: Hamilton RJ, Hamilton S, editors.Lipid analysis—a practical approach. 1st ed. Oxford: IRLPress at Oxford University Press; 1992 [chapter 2].
38. Nishiyama-Naruke A, de Sousa JAA, Carnelos Filho M, Curi R.HPLC determination of underivatized fatty acid saponified atlow temperature analysis of fatty acids in oils and tissues.Analy Lett 1998;31:2565–76.

ARTICLE IN PRESS
R. Gorjao et al.938
39. Abushufa R, Reed P, Weinkove C. Fatty acids in erythrocytesmeasured by isocratic HPLC. Clin Chem 1994;40:1707–12.
40. Guimaraes AR, Costa Rosa LF, Safi DA, Curi R. Metabolic andfunctional changes in macrophages of rats fed polyunsatu-rated or saturated fatty acid rich-diets during ageing.Biochem Int 1992;27:9–16.
41. Sampaio SC, Brigatte P, Sousa-e-Silva MC, et al. Contributionof crotoxin for the inhibitory effect of Crotalus durissusterrificus snake venom on macrophage function. Toxicon2003;41(7):899–907.
42. Bignold LP. Use of sparse-pore polycarbonate (Nuclepore)membrane for the measurement of chemotaxis of poly-morphonuclear leucocytes. J Immunol Methods 1989;118:217–25.
43. Boyum A. Isolation of mononuclear cells and granulocytesfrom human blood. Isolation of monuclear cells by onecentrifugation, and of granulocytes by combining centrifu-gation and sedimentation at 1 g. Scand J Clin Lab Invest1968;97:77–89.
44. Sambrook J, Russell DW. Molecular cloning: a laboratorymanual, 3rd ed. New York: Cold Spring Harbor LaboratoryPress; 2001 A8.21.
45. Yamazaki K, Kuromitsu J, Tanaka I. Microarray analysis ofgene expression changes in mouse liver induced by peroxi-some proliferator activated receptor alpha agonists. Bio-chem Biophys Res Commun 2002;290:1114–22.
46. Innis MA, Gelfand DH. Optimization of PCRs. In: Innis MA,Gelfand DH, Sninsky JJ, White TJ, editors. PCR protocols: aguide to methods and applications. 1st ed. San Diego, CA:Academic Press; 1990. p. 3–12.
47. Kwok S, Higuchi R. Avoiding false positives with PCR. Nature1989;339:237–8.
48. Calder P C. n-3 Polyunsaturated fatty acids and immune cellfunction. Adv. Enzyme Regul 1997;37:197–237.
49. Goodwin JS, Ceuppens J. Regulation of the immune responseby prostaglandins. J Clin Immunol 1983;3:295–315.
50. Switzer KC, McMurray DN, Chapkin R S. Effects of dietary n-3polyunsaturated fatty acids on T-cell membrane compositionand function. Lipids 2004;39:1163–70.
51. Kew S, Mesa MD, Tricon S, Buckley R, Minihane AM, Yaqoob P.Effects of oils rich in eicosapentaenoic and docosahexaenoicacids on immune cell composition and function in healthyhumans. Am J Clin Nutr 2004;79(4):674–81.
52. Virella G, Kilpatrick JM, Rugeles MT, Hyman B, Russell R.Depression of humoral responses and phagocytic functions invivo and in vitro by fish oil and eicosapentanoic acid. ClinImmunol Immunopathol 1989;52:257–70.
53. Sipka S, Dey I, Buda C, Csongor J, Szeged G, Farkas T. Themechanism of inhibitory effect of eicosapentaenoic acid onphagocytic activity and chemotaxis of human neutrophilgranulocytes. Clin Immunol Immunopathol 1996;79:224–8.
54. Halvorsen DS, Hansen JB, Grimsgaard S, Bonaa KH, Kierulf P,Nordoy A. The effect of highly purified eicosapentaenoic and
docosahexaenoic acids on monocyte phagocytosis in man.Lipids 1997;32:935–42.
55. Poulos A, Robinson BS, Ferrante A, Harvey DP, Hardy SJ,Murray AW. Effect of 22–32 carbon n-3 polyunsaturated fattyacids on superoxide production in human neutrophils:synergism of docosahexaenoic acid with f-met-leu-phe andphorbol ester. Immunol 1991;73:102–8.
56. Chen LY, Lawson DL, Mehta JL. Reduction in humanneutrophil superoxide anion generation by n-3 polyunsatu-rated fatty acids: role of cyclooxygenase products andendothelium-derived relaxing factor. Thromb Res 1994;76:317–22.
57. Molvig J, Pociot F, Worssaae H, et al. Dietary supplementa-tion with $-3 polyunsatured fatty acids decreases mono-nuclear cell proliferation and interleukin-1b content but notmonokine secretion in healthy and insulin-dependent dia-betic individuals. Scand J Immunol 1991;34:399–410.
58. Wallace FA, Miles EA, Calder PC. Comparison of the effectsof linseed oil and different doses of fish oil on mononuclearcell function in healthy human subjects. Br J Nutr2003;89(5):679–89.
59. Xi S, Cohen D, Chen LH. Effects of fish oil on cytokines andimmune functions of mice with murine AIDS. J Lipid Res1998;39:1677–87.
60. Fan YY, Ly LH, Barhoumi R, McMurray DN, Chapkin RS.Dietary docosahexaenoic acid suppresses T cell proteinkinase C theta lipid raft recruitment and IL-2 production. JImmunol 2004;173:6151–60.
61. Denys A, Hichami A, Khan NA. Eicosapentaenoic acid anddocosahexaenoic acid modulate MAP kinase (ERK1/ERK2)signaling in human T cells. J Lipid Res 2001;42:2015–20.
62. Nakamura MT, Cheon Y, Li Y, Nara TY. Mechanisms ofregulation of gene expression by fatty acids. Lipids2004;39(11):1077–83.
63. Stulnig TM, Zeyda M. Immunomodulation by polyunsaturatedfatty acids: impact on T-cell signalling. Lipids2004;39(12):1171–5.
64. Kiang JG, Tsokos GC. Heat shock protein 70 kDa: molecularbiology, biochemistry, and physiology. Pharmacol Ther1998;80:183–201.
65. Young RA. Stress proteins and immunology. Annu RevImmunol 1990;8:401–20.
66. Polla BS, Stubbe H, Kantengwa S, Maridonneau-Parini I,Jacquier-Sarlin MR. Differential induction of stress proteinsand functional effects of heat shock in human phagocytes.Inflammation 1995;19:363–78.
67. Samples BL, Pool GL, Lumb RH. Polyunsaturated fatty acidsenhance the heat induced stress response in rainbow trout(Oncorhynchus mykiss) leukocytes. Comp Biochem Physiol BBiochem Mol Biol 1999;123:389–97.
68. Heimli H, Giske C, Naderi S, Drevon CA, Hollung K.Eicosapentaenoic acid promotes apoptosis in Ramos cellsvia activation of caspase-3 and -9. Lipids 2002;37:797–802.
Copyright © 2022 FDOKUMEN



















