
Improved outcome after spinal cord compression injury in mice treated with docosahexaenoic acid
15
Improved outcome after spinal cord compression injury in mice treated with docosahexaenoic acid Siew-Na Lim a, b , Wenlong Huang a, c , Jodie C.E. Hall a, d , Adina T. Michael-Titus a , John V. Priestley a, ⁎ a Centre for Neuroscience and Trauma, Blizard Institute, Barts and The London School of Medicine and Dentistry, Queen Mary University of London, London, UK b Department of Neurology, Chang Gung Memorial Hospital and Chang Gung University College of Medicine, Taipei, Taiwan c Pain Research Group, Department of Surgery and Cancer, Imperial College London, SW10 9NH, UK d Center for Brain and Spinal Cord Repair, The Ohio State University, Columbus, OH 43210, USA abstract article info Article history: Received 17 May 2012 Revised 20 September 2012 Accepted 24 September 2012 Available online 28 September 2012 Keywords: Trauma Spinal cord injury Polyunsaturated fatty acid Docosahexaenoic acid Neuroprotection In this study we have characterised the locomotor recovery, and temporal profile of cell loss, in a novel thoracic com- pression spinal cord injury (SCI) in the mouse. We have also shown that treatment with docosahexaenoic acid (DHA) is neuroprotective in this model of SCI, strengthening the growing literature demonstrating that omega-3 polyunsaturated fatty acids are neuroprotective after SCI. Compression SCI in C57BL/6 mice was produced by placing a 10 g weight for 5 min onto a 2 mm×1.5 mm platform applied to the dura at vertebral level T12. Mice partly recovered from complete hindlimb paralysis and by 28 days post-surgery had plateaued at an average BMS locomotor score of 4.2, equivalent to weight support with plantar stepping. During the same period, neuronal loss at the epicentre increased from 26% of ventral horn neurons by day 1, to 68% by day 28. Delayed loss of oligodendrocytes was also seen (e.g. 84% by day 28 in the dorsal columns) and microglia/macrophage activation was maximal at 7 days. In contrast, axonal damage, judged by a decrease in the non-phosphorylated form of 200 kD neurofilament, was an early event, as the loss was seen by day 1 and did not change markedly over time. Mice that received an intravenous (i.v.) injection of 500 nmol/kg DHA 30 min after SCI, showed improved locomotor recovery and, at 28 day survival, reduced neuronal, oligodendrocyte and neurofilament loss, and reduced microglia/macrophage activation. For some of these indices of SCI, enrichment of the diet with 400 mg/kg/day DHA led to further improvement. However, dietary DHA supplementation, without the initial i.v. injection, was ineffective. © 2012 Elsevier Inc. All rights reserved. Introduction Traumatic spinal cord injury (SCI) has devastating consequences for patients, and current treatments, such as acute decompression (Fehlings and Perrin, 2006) or administration of methylprednisolone (Hurlbert and Hamilton, 2008), are controversial or of limited efficacy. In preclinical studies, major progress has been made in both the fields of neuroprotection and neuroregeneration (Kwon et al., 2010), but translation into the clinic has been disappointing. Neuroprotective strategies are particularly appealing because they aim to stop the spread of injury, and the complex cascade of events that follow the primary insult means there are many possible targets for neuroprotective agents. However, although there are promising new agents under development (Kwon et al., 2011a), many clinical trials have failed, either because of limited efficacy or unexpected toxicity (Tator and Fehlings, 1999). There is, therefore, still a need for the development of new neuroprotective agents, which are safe and effective when delivered after spinal cord injury. Recently, a number of preclinical studies have demonstrated that omega-3 (n−3) polyunsaturated fatty acids (PUFAs) are neuroprotective when administered after SCI. n −3 PUFAs are essential fatty acids that have crucial roles in the development and mature functioning of the nervous system. Particularly important is docosahexaenoic acid (DHA), a long chain (22 carbon) n −3 PUFA that accounts for approximately 50% of the PUFAs in central nervous system (CNS) membranes. We have shown in rat hemisection and compression models of SCI that DHA administered as an intravenous bolus 30 min after injury leads to increased neuronal, oligodendrocyte and axonal survival at the lesion epicentre, and improved locomotor function (Huang et al., 2007b; King et al., 2006; Ward et al., 2010). When the acute bolus is combined with dietary DHA supplementation for several weeks following injury, addi- tional cell and axonal survival is seen, and further improvement of func- tional outcome (Huang et al., 2007b; Ward et al., 2010). Most recently we have shown that a multi-nutrient dietary formulation containing n −3 PUFAs is also neuroprotective (Zbarski-Barquero et al., 2012). Neuroprotection has also been reported in studies involving treatment with DHA prior to SCI (Figueroa et al., 2012) or treatment after SCI with Experimental Neurology 239 (2013) 13–27 ⁎ Corresponding author at: Centre for Neuroscience and Trauma, Blizard Institute, Barts and The London School of Medicine and Dentistry, Queen Mary University of London, 4 Newark Street, London, UK. Fax: +44 20 78822218. E-mail address: [email protected] (J.V. Priestley). 0014-4886/$ – see front matter © 2012 Elsevier Inc. All rights reserved. http://dx.doi.org/10.1016/j.expneurol.2012.09.015 Contents lists available at SciVerse ScienceDirect Experimental Neurology journal homepage: www.elsevier.com/locate/yexnr
Transcript of Improved outcome after spinal cord compression injury in mice treated with docosahexaenoic acid

Experimental Neurology 239 (2013) 13–27
Contents lists available at SciVerse ScienceDirect
Experimental Neurology
j ourna l homepage: www.e lsev ie r .com/ locate /yexnr
Improved outcome after spinal cord compression injury in mice treated withdocosahexaenoic acid
Siew-Na Lim a,b, Wenlong Huang a,c, Jodie C.E. Hall a,d, Adina T. Michael-Titus a, John V. Priestley a,⁎a Centre for Neuroscience and Trauma, Blizard Institute, Barts and The London School of Medicine and Dentistry, Queen Mary University of London, London, UKb Department of Neurology, Chang Gung Memorial Hospital and Chang Gung University College of Medicine, Taipei, Taiwanc Pain Research Group, Department of Surgery and Cancer, Imperial College London, SW10 9NH, UKd Center for Brain and Spinal Cord Repair, The Ohio State University, Columbus, OH 43210, USA
⁎ Corresponding author at: Centre for Neuroscience andand The London School of Medicine and Dentistry, QueeNewark Street, London, UK. Fax: +44 20 78822218.
E-mail address: [email protected] (J.V. Priestl
0014-4886/$ – see front matter © 2012 Elsevier Inc. Allhttp://dx.doi.org/10.1016/j.expneurol.2012.09.015
a b s t r a c t
a r t i c l e i n f oArticle history:Received 17 May 2012Revised 20 September 2012Accepted 24 September 2012Available online 28 September 2012
Keywords:TraumaSpinal cord injuryPolyunsaturated fatty acidDocosahexaenoic acidNeuroprotection
In this studywehave characterised the locomotor recovery, and temporal profile of cell loss, in a novel thoracic com-pression spinal cord injury (SCI) in the mouse. We have also shown that treatment with docosahexaenoic acid(DHA) is neuroprotective in this model of SCI, strengthening the growing literature demonstrating that omega-3polyunsaturated fatty acids are neuroprotective after SCI.Compression SCI in C57BL/6 mice was produced by placing a 10 g weight for 5 min onto a 2 mm×1.5 mmplatform applied to the dura at vertebral level T12. Mice partly recovered from complete hindlimb paralysisand by 28 days post-surgery had plateaued at an average BMS locomotor score of 4.2, equivalent to weightsupport with plantar stepping. During the same period, neuronal loss at the epicentre increased from 26%of ventral horn neurons by day 1, to 68% by day 28. Delayed loss of oligodendrocytes was also seen(e.g. 84% by day 28 in the dorsal columns) and microglia/macrophage activation was maximal at 7 days. Incontrast, axonal damage, judged by a decrease in the non-phosphorylated form of 200 kD neurofilament,was an early event, as the loss was seen by day 1 and did not change markedly over time.Mice that received an intravenous (i.v.) injection of 500 nmol/kg DHA 30 min after SCI, showed improvedlocomotor recovery and, at 28 day survival, reduced neuronal, oligodendrocyte and neurofilament loss, andreduced microglia/macrophage activation. For some of these indices of SCI, enrichment of the diet with400 mg/kg/day DHA led to further improvement. However, dietary DHA supplementation, without the initiali.v. injection, was ineffective.
© 2012 Elsevier Inc. All rights reserved.
Introduction
Traumatic spinal cord injury (SCI) has devastating consequencesfor patients, and current treatments, such as acute decompression(Fehlings and Perrin, 2006) or administration of methylprednisolone(Hurlbert and Hamilton, 2008), are controversial or of limited efficacy.In preclinical studies, major progress has been made in both the fieldsof neuroprotection and neuroregeneration (Kwon et al., 2010), buttranslation into the clinic has been disappointing. Neuroprotectivestrategies are particularly appealing because they aim to stop the spreadof injury, and the complex cascade of events that follow the primaryinsult means there are many possible targets for neuroprotectiveagents. However, although there are promising new agents underdevelopment (Kwon et al., 2011a), many clinical trials have failed,either because of limited efficacy or unexpected toxicity (Tator andFehlings, 1999). There is, therefore, still a need for the development of
Trauma, Blizard Institute, Bartsn Mary University of London, 4
ey).
rights reserved.
new neuroprotective agents, which are safe and effective when deliveredafter spinal cord injury.
Recently, a number of preclinical studies have demonstrated thatomega-3 (n−3) polyunsaturated fatty acids (PUFAs) are neuroprotectivewhen administered after SCI. n−3 PUFAs are essential fatty acids thathave crucial roles in the development and mature functioning of thenervous system. Particularly important is docosahexaenoic acid (DHA),a long chain (22 carbon) n−3 PUFA that accounts for approximately50% of the PUFAs in central nervous system (CNS) membranes. Wehave shown in rat hemisection and compression models of SCI thatDHA administered as an intravenous bolus 30 min after injury leads toincreased neuronal, oligodendrocyte and axonal survival at the lesionepicentre, and improved locomotor function (Huang et al., 2007b; Kinget al., 2006; Ward et al., 2010). When the acute bolus is combined withdietary DHA supplementation for several weeks following injury, addi-tional cell and axonal survival is seen, and further improvement of func-tional outcome (Huang et al., 2007b; Ward et al., 2010). Most recentlywe have shown that a multi-nutrient dietary formulation containingn−3 PUFAs is also neuroprotective (Zbarski-Barquero et al., 2012).Neuroprotection has also been reported in studies involving treatmentwith DHA prior to SCI (Figueroa et al., 2012) or treatment after SCI with

14 S.-N. Lim et al. / Experimental Neurology 239 (2013) 13–27
fenretinide, a synthetic retinoid derivative which increases levels ofendogenous DHA (Lopez-Vales et al., 2010). DHA has also been shownto be neuroprotective if administered after traumatic brain injury inrodents (Bailes and Mills, 2010; Wu et al., 2011). The exact mechanismunderlying these neuroprotective effects is not known, but it is likely toinvolve multiple pathways. Actions of DHA include effects on membraneproperties such as fluidity and permeability (Stillwell andWassall, 2003),effects on membrane proteins including receptors (Lafourcade et al.,2011) and ion channels (Lauritzen et al., 2000), effects on gene transcrip-tion by direct binding of non-esterified DHA to peroxisome proliferatoractivated receptors (PPARs) (Jump, 2002), reduction in the levels ofpro-inflammatory n−6 PUFAs by competition for metabolic andcatabolic pathways, and effects of DHA metabolites, including the anti-inflammatory and pro-homeostatic neuroprotectin D1 (NPD1) (Bazanet al., 2011). These multiple actions, and the established safety profile ofexisting n−3 PUFA formulations (Heller et al., 2006), make DHA a veryattractive candidate for the clinical treatment of acute SCI.
Before DHA is tested in clinical trials, it should be validated in awiderange of preclinical SCI models, including different types of injury(e.g. contusion and compression) and different species (e.g. ratand mouse). Efficacy in multiple models has been identified as animportant criterion before translation to the clinic (Kwon et al.,2011b). Most of our studies of n−3 PUFAs have been carried outin a rat thoracic compression model of SCI (Hall et al., 2012;Huang et al., 2007b; Lim et al., 2010; Ward et al., 2010), a modelin which cell death extends over a longer period than in contusioninjury models (Huang et al., 2007a). In the present study we havetherefore developed a mouse compression model, which is similarin principle to our rat model (Huang et al., 2007a) and used it to in-vestigate the neuroprotective properties of acute and dietary DHAtreatment. It is well-established that mice exhibit a very differentresponse to SCI, both in terms of functional recovery, and also inthe tissue pathological changes and inflammatory reaction, com-pared to that seen in other mammals. In particular, mice do not ex-hibit the progressive necrosis and larger central cavitation that is sodramatic in rats and other mammals, in which a rim of anatomicallypreserved white matter surrounds a fluid-filled cystic cavity at theinjury epicentre (Grossman et al., 2001; Huang et al., 2007a). In con-trast, the injured mouse spinal cord is completely filled-in with densefibrous connective tissue, and if present, there are only very small cavi-ties (microcysts) in the lesion site (Farooque, 2000; Joshi and Fehlings,2002a,b;Ma et al., 2001). The use of a mouse compressionmodel there-fore allows us to test DHA in a very different pathology, and provides abaseline for future studies in transgenic mice (e.g. Lim et al., 2012).
Material and methods
Compression SCI
C57BL/6 female mice (19–21 g) were deeply anaesthetized with 4%isoflurane (Merial, Essex, UK), as evidenced by lack of response to anociceptive stimulus. Subsequent anaesthesia throughout the proce-dure was maintained using 1.5–2% isoflurane, with oxygen and nitrousoxide at a 1:1 ratio. A laminectomy was performed at vertebral levelT12, leaving the dura undisturbed. The T11 and T13 transverse process-es were clamped in a spinal compression frame, and the compressionwas applied by suspending the base of a compression platform ontothe exposed cord, under microscopic control. Then a weight wasapplied to the platform for a specific period of time. The platform wasthen removed, the muscle layers were sutured and the skin layersclosed with wound clips. In preliminary experiments, platforms of1 mm×1 mm and 2 mm×1.5 mm were tested with a 10 g weightapplied for 10 min. The 2 mm×1.5 mm platform was further testedusing a 10 g weight applied for 5 min, or a 5 g weight applied for10 min. The 10 g weight applied to a 2 mm×1.5 mm platform for5 min, was judged to give the most reproducible results and BMS
recovery profile indicative of a moderate SCI, and so was used in allthe experiments described in this study. After surgery, animals weregiven buprenorphine intramuscularly (0.12 mg/kg; Reckitt Benckiser,UK) and then were placed in warmed cages to recover from anaesthe-sia. Manual bladder expression was performed twice a day until theestablishment of reflex voiding. The mice were given free access tofood and tap water, and were maintained on a 12 hour light/darkcycle at 21 °C±1 °C.
Treatment with DHA
SCI surgery was performed in several groups of animals, whichreceived various types of DHA treatment. To define the optimum in-travenous (i.v.) DHA treatment dose for mice, first we carried out adose–response study. DHA for acute i.v. injection was diluted from1 M stock aliquots (5 μl) of free fatty acid (Sigma, Dorset, UK) madeup in absolute ethanol and stored at −80 °C until use. On the day ofsurgery, the stock aliquot solutions of fatty acids were diluted to therequired concentration in sterilized saline (NaCl, 0.9%) and adjustedto pH 7.4. Mice received the following treatments via a tail vein30 min after SCI: (i) saline vehicle (n=5); or (ii) DHA 100 nmol/kg(n=6); (iii) or DHA 250 nmol/kg (n=6); or (iv) DHA 500 nmol/kg(n=6), in a volume of 5 ml/kg, injected over 10 s. In subsequentstudies, we explored a combined acute and chronic DHA treatmentregime. Mice received the following treatment: (i) an i.v. injectionof saline vehicle 30 min after SCI and a control diet (n=9); or (ii)an i.v. injection of DHA (500 nmol/kg) 30 min after SCI and a controldiet (n=9); or (iii) an i.v. injection of saline vehicle 30 min after SCIand a DHA-enriched diet (400 mg/kg/day) (n=8); or (iv) an i.v. in-jection of DHA (500 nmol/kg) 30 min after injury and a DHA-enriched diet (400 mg/kg/day) (n=8). All animals were maintainedfor 4 weeks until sacrifice. Groups were allocated randomly and theexperimenter was blinded to the treatment groups throughout thesurgery and the behavioural testing period.
Two weeks prior to the start of the DHA dietary enrichment studies,the animals' diet was changed from the standard pelleted form to aground form, in order to allow habituation to the different foodtexture. The control diet consisted of 5KB3 Certified EURodent mainte-nance diet 14% (LabDiet, IPS International Product Supplies Limited,London). The DHA-enriched diet consisted of the 5KB3 diet to whichoil enriched in DHA (Incromega DHA 700E SR; Croda Healthcare, En-gland) was added, resulting in the mice receiving a daily dose of400 mg DHA per kilogramme of animal body mass. The oil contained70–75% DHA, 10% EPA and the remainder contained arachidonic acid(AA) and other fatty acids. Oil-enriched diet was prepared weeklyunder nitrogen by thoroughly mixing the oil with the 5KB3 powderdiet for a minimum of 10 min, ensuring complete and even dispersalof the oil throughout the diet. While still under nitrogen the food wasplaced in air-tight bags and then stored at −20 °C until used. Animalswere found to consume less food for the first week after SCI (20–50%less), but later they resumed normal consumption of the amount ofthe diet provided. The full dose received was recalculated based onthese findings.
Functional assessment
We used the Basso mouse scale (BMS) open-field test (Basso et al.,2006) to assess functional improvement after SCI. The scale consistsof 9 categories which reflect the different stages of recovery. A BMSscore of zero indicates that the animal has complete hindlimb paraly-sis, whereas a BMS score of 9 indicates the animal has no locomotordeficits (uninjured animal). The 11-point BMS subscores, which eval-uate finer aspects of locomotor control, were also assessed. Thebehaviour was assessed and scored in blind, daily for the first weekand either daily, or every 3 days thereafter until sacrifice.

15S.-N. Lim et al. / Experimental Neurology 239 (2013) 13–27
Tissue processing and immunocytochemistry
The tissue sections for immunohistochemical analysis were pre-pared as follows: at specific time points following SCI, the animalswere sacrificed using CO2 inhalation and transcardially perfused withsaline, followed by 4% paraformaldehyde (PFA) in 0.1 M phosphatebuffer (pH 7.4). The 2 mm compression epicentre of the cord wasdissected out, post fixed for 2 h, and cryoprotected in 30% sucrose in0.01 M phosphate buffered saline (PBS) overnight at 4 °C. The tissuewas embedded in OCT medium, frozen and cut into serial transverse10 μm cryostat sections and collected onto Superfrost® slides (VWR,Lutterworth).
Selected slides from the compression epicentre were processedfor immunohistochemistry using the following primary antibodies:mouse neuron-specific nuclear protein (NeuN, 1:1000; Chemicon,USA) to examine neuronal survival, mouse adenomatous polyposiscoli tumour suppressor protein (APC, 1:100; Calbiochem, USA) toexamine the survival of oligodendrocytes, mouse SMI32 (1:1000;Sternberger Monoclonals, UK) to detect non-phosphorylated 200 kDneurofilament, and rabbit Ionized calcium-binding adapter molecule-1(Iba1, 1:1000; Wako, Germany) to label microglia/macrophages.Sections were incubated in 10% normal donkey serum for 1 h followedby an overnight incubation with primary antibodies. Sections werewashed three times (10 min each) in PBS before being incubated for2 h in the appropriate secondary antibodies conjugated to Alexa Fluor488 or 594 (1:1000; Invitrogen, Paisley, UK). After another three10 min washes in PBS, sections labelled with NeuN, and APC werethen counterstained with the fluorescent nuclear dye bis-benzimide(Hoechst 33342; 0.2 mg/100 ml PBS; Sigma, UK) for 2 min to facilitatethe detection of the nucleus of NeuN and APC-labelled cells.
Image analysis
Sections were viewed on a Leica epifluorescence microscope(Wetzlar, Germany) and photographed at 20× magnification usinga Hamamatsu C4742-95 digital camera (Herrsching, Germany) andHiPic software (Hamamatsu). In Photoshop, discrete measuringframes were placed over the areas of interest on each image, andthe number of immunoreactive cells with visible Hoechst-labellednuclei was counted manually in 3 sections per animal. For NeuN anal-ysis, two 144 μm×115 μm measuring frames were placed in the dor-sal horns on both the left and right sides, while two 180 μm×144 μmmeasuring frames were placed on each ventral horn. A similar analy-sis was conducted for APC-labelled cells, with the exception thatareas chosen for APC were the dorsal column (90 μm×72 μm), thecorticospinal tract (90 μm×72 μm), the lateral (180 μm×144 μm),and ventral white matter (180 μm×144 μm) (bilaterally).
Quantification of SMI32 labelled axons was performed in the dorsalcolumn, and lateral and ventral white matter, by using the image anal-ysis programme Image J (Image J 1.43u, National Institute of Health,USA). The size of the measuring frames in each area was the same asthe one used for APC analysis. In the Image J programme, these imageswere converted to a set threshold for all images and then counted auto-matically using the “analyse particles” programme. Iba1 immunoreac-tivity of macrophages/microglia in the epicentre was also quantifiedusing the Image J software. Areas chosen for Iba1 were the dorsal col-umn, dorsal horn, ventral horn and ventral white matter (bilaterally).A measuring frame of 90 μm×72 μm was placed on each dorsal col-umn, while a 180 μm×144 μm measuring frame was placed withinthe dorsal horn, ventral horn and ventral white matter, on both theleft and right sides. In the Image J programme, the appropriate thresh-old value was set manually at a level to detect most of the finestmicroglial processes. The level of immunoreactivity was expressed asthe percentage of the area of the measuring frame that containedimmunoreactivity (field detected area, FDA).
Statistical analysis
The behavioural assessments and histological analysis were perform-ed blind. All statistical analyses were performed using SPSS 18.0. The datawere presented as means and standard errors of the means (S.E.M.).One-way or two-way repeated measures ANOVA was used to comparethe experimental groups, with post-hoc Tukey's test formultiple compar-isons. Differences were considered statistically significant at Pb0.05.
Results
Characterization of mouse compression injury
Locomotor recovery after compression SCICompression SCI at vertebral T12 level produced a long-lasting
deficit in hindlimb function, as assessed using the BMS scale (Fig. 1).Recovering from complete paralysis, all mice demonstrated plantarplacement of the paw (BMS score 3) by 2 weeks after injury, andsome mice started to show occasional weight-supported plantarstepping (BMS score 4). Beyond 2 weeks themice continued to recoververy slowly and by 4 weeks had plateaued at a BMS score of just above 4(4.2). This equates with occasional or frequent plantar stepping, butwithout any coordination. In contrast to the profound locomotor deficitseen in the compression injury mice, laminectomy controls showedonly a small deficit (BMS score 8.5) and had fully recovered by day 3(Fig. 1).
Effect of compression SCI on neuronesIn naïve control mice, NeuN-labelled neurons were present through-
out the grey matter of the spinal cord (Figs. 2A,G). In the laminectomycontrols there appeared to be slightly fewer NeuN-labelled cells(Figs. 2B,H), and this was further confirmed by quantitative analysis,which showed a 8–10% loss of NeuN immunoreactivity between 1 and28 days following the laminectomy. However, this did not reach statisti-cal significance (P>0.05; Fig. 4A). There were no significant effects ofpost-surgery time on the loss of NeuN immunoreactivity following thelaminectomy surgery.
In contrast to the small loss of NeuN immunoreactivity followinglaminectomy, a substantial loss of NeuN immunoreactivity in thecompression epicentre was noted, as compared with naïve controls(Figs. 2, 4). The loss of NeuN immunoreactivity increased withpost-compression time over the 28 day observation period, but wasmost prominent after the first 3 days. Quantitative analysis confirmedthat significantly lower numbers of NeuN-labelled cells were found inthe epicentre overall, at all four post-surgery time points comparedwith the naïve controls (Pb0.05; Fig. 4B). In addition, the loss ofNeuN immunoreactivity was greater in the dorsal horns than in theventral horns (Fig. 4B). The analysis also revealed a significant effectof post-surgery time on the number of NeuN immunoreactive cellsin the compression epicentre. Thus, the losses of NeuN immunoreac-tivity at 7 and 28 days were significantly higher than the effect at1 day after the compression injury (Pb0.05; Fig. 4B). There was nosignificant difference between 3, 7 and 28 days. In contrast to theSCI in rats where large central cavities are seen in the lesion epicentre(Huang et al., 2007a; Liu et al., 1997), there was no central cavitationup to 28 days after SCI in our mouse model (Figs. 2A–F).
Effect of compression SCI on oligodendrocytesIn the spinal cord of naïve control mice, APC-labelled oligodendro-
cytes were present throughout the white matter (Fig. 3A). In thelaminectomy controls there appeared to be slightly fewer APC-labelled cells (Fig. 3B), and this was further confirmed by quantitativeanalysis, which showed a 5–22% loss of APC immunoreactivity be-tween 1 and 28 days following the laminectomy surgery comparedwith the naïve control; however, this did not reach statistical signifi-cance (P>0.05; Fig. 4C). In contrast, within 1 day post-compression

Fig. 1. Behavioural analysis of locomotor recovery after compression SCI. There was only partial recovery of locomotion following SCI. Animals recovered from paralysis to extensiveankle movement (BMS score 2.50±0.2) at 1 week after injury and continued to recover to plantar placement (BMS score 3.55±0.3) at 2 weeks after injury. After that, the animalsshowed very slow recovery, and by 4 weeks after injury, these animals displayed a recovery corresponding to a BMS score of 4.2±0.3. The laminectomy group shows only a tran-sient, non-significant, reduction in BMS score. Results represent mean±S.E.M. (n=9, compression group and n=6, laminectomy group).
16 S.-N. Lim et al. / Experimental Neurology 239 (2013) 13–27
injury, a dramatic loss of APC-immunoreactivity was observed in allfour different white matter areas studied, i.e. the dorsal column,corticospinal tract, lateral and ventral white matter. The loss of APCimmunoreactivity increased with post-compression time over the28 day observation period (Fig. 4D), although there was no statisti-cally significant difference between the 1, 3, 7 and 28 day time points.
Effect of compression SCI on non-phosphorylated neurofilamentAs there were no significant differences in the NeuN and APC im-
munoreactivity after laminectomy surgery, for the next two markers,SMI32 and Iba1, we only performed immunohistochemical analysisfor the laminectomy groups at 28 days post-surgery.
SCI has been reported to lead to a preferential loss of the non-phosphorylated neurofilament (Schumacher et al., 1999). Therefore,the effect of injury on the non-phosphorylated form of the 200 kDneurofilament was studied using the antibody SMI32. In naïve controlmice, SMI32-positive axons were present and arranged in a regularmanner in the white matter of the spinal cord (Fig. 5A). In thelaminectomy controls at 28 days (Fig. 5B), there appeared to be fewerSMI32-positive axons than in naïve mice, but this did not reach statisti-cal significance (P>0.05; Fig. 6A). In contrast, a dramatic loss ofSMI32-positive axons occurred within 1 day post-compression injuryand continued over the 28 day observation period, with many of theremaining axons appearingmisaligned and swollen (Figs. 5C–F). Quan-titative analysis confirmed that compression injury resulted in fewerSMI32-positive axons than naïve and laminectomy controls at all timepoints in the lateral white matter, while in the dorsal column and ven-tral white matter the injury-induced loss of non-phosphorylatedneurofilament only reached statistical significance at 28 days (Fig. 6A).
Effect of compression SCI on microglia/macrophage activationIba1 was used to evaluate the changes in microglia/macrophage
activation following SCI. In the spinal cord of the naïve mice, Iba1immunolabelling showed small resting microglial cells with long, ram-ified thin processes throughout the grey and thewhitematter (Fig. 5G).In the laminectomy controls, the morphology of Iba1 immunolabellingwas very similar to that of the naïve mice (Fig. 5H). One day aftercompression surgery, heavily stained microglia with altered shapesappeared in the spinal cord. At day 3 after compression surgery,microglial/macrophage activation was evident in the spinal cord andwas represented by a reduced number of ramified cells, with processesthat, when present, were thick and less branched, and by an increase inthe size ofmany Iba1-stained nuclei (Fig. 5J). At day 7 after compression
injury, Iba1 immunoreactivity was dramatically increased, with large,rounded cellular Iba1-labelling occupying most of the spinal cordepicentre. By 28 days, the intense microglia/macrophage responsepersisted, but the immunoreactivity of these cells at this time appearedto be weaker than at day 7 (Figs. 5K,L). Detailed quantification con-firmed that a dramatic increase in Iba1 immunoreactivity was seen inthe compression epicentre at day 7 after compression SCI, and declinedat 28 days post-compression, but it remained elevated above that inuninjured controls (Pb0.05 Fig. 6B). There were no significant differ-ences in the Iba1 expression between naïve, laminectomy, post-compression day 1 and day 3 mice (P>0.05; Fig. 6B).
Effect of DHA treatment on mouse compression spinal cord injury
Dose-dependent neuroprotective effects of DHA on motor function aftermouse compression SCI. In the first part of the study, in order to definethe optimum intravenous DHA treatment dose for mice, a preliminarydose–response study was carried out. Compared with saline-treatedmice, mice treated with 500 nmol/kg DHA intravenous injection showedsignificant improvement in locomotor recovery following compressionSCI (Pb0.05; Fig. 7A), and post-hoc analysis revealed significant differ-ences in the BMS score starting at day 4 after injury, The score remainedsignificantly enhanced for the 4 week observation period. DHA at a doseof 250 nmol/kg was less effective. Mice treated with 250 nmol/kg DHAshowed significantly improved functional outcome from 19 to 28 dayspost-injury compared to the saline-treatedmice, but the overall function-al outcome of these mice did not differ significantly from saline-treatedcontrols or the 500 nmol/kg DHA group. At 28 days post-injury, micetreated with 500 nmol/kg and 250 nmol/kg DHA were able to displayfrequent or consistent plantar stepping, with some coordination andmore parallel paw position (BMS 6.6±0.4 and 5.8±0.2, respectively),while mice treated with saline only showed occasional stepping withoutcoordination (BMS 4.1±0.43). In addition, fine locomotor controlassessed by the BMS subscores also showed a significantly improvedresult in mice treated with 500 nmol/kg DHA from 13 to 28 dayspost-injury compared with saline-treated controls. At 28 days post-injury, treatment with 500 nmol/kg DHA resulted in a mean BMSsubscore that was 3.6 points higher than that of saline-treated mice(Pb0.05; Fig. 7B). The functional recovery of the 100 nmol/kg DHAgroup did not differ significantly from the saline-treated controls. Micetreated with 500 nmol/kg DHA showed improved functional outcomeat the 13, 16, 19 and 22 day time points, compared to the 250 nmol/kgDHA group. Thus, the effect of DHA was dose-dependent, with mice
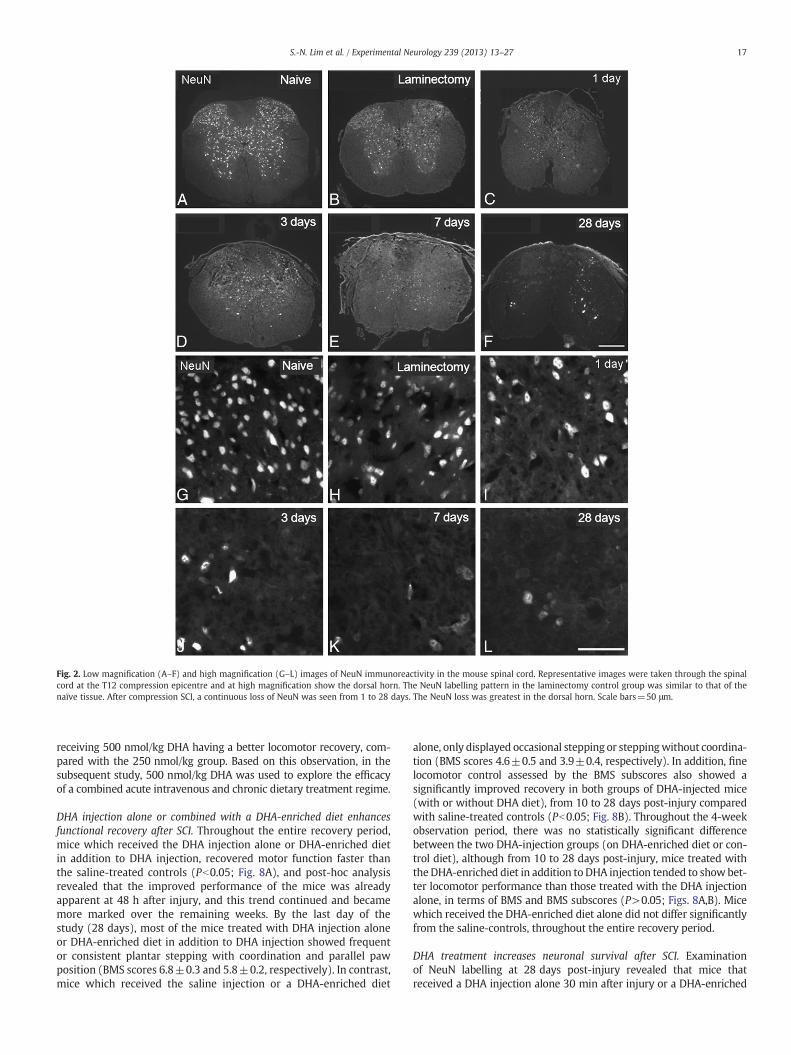
Fig. 2. Low magnification (A–F) and high magnification (G–L) images of NeuN immunoreactivity in the mouse spinal cord. Representative images were taken through the spinalcord at the T12 compression epicentre and at high magnification show the dorsal horn. The NeuN labelling pattern in the laminectomy control group was similar to that of thenaïve tissue. After compression SCI, a continuous loss of NeuN was seen from 1 to 28 days. The NeuN loss was greatest in the dorsal horn. Scale bars=50 μm.
17S.-N. Lim et al. / Experimental Neurology 239 (2013) 13–27
receiving 500 nmol/kg DHA having a better locomotor recovery, com-pared with the 250 nmol/kg group. Based on this observation, in thesubsequent study, 500 nmol/kg DHA was used to explore the efficacyof a combined acute intravenous and chronic dietary treatment regime.
DHA injection alone or combined with a DHA-enriched diet enhancesfunctional recovery after SCI. Throughout the entire recovery period,mice which received the DHA injection alone or DHA-enriched dietin addition to DHA injection, recovered motor function faster thanthe saline-treated controls (Pb0.05; Fig. 8A), and post-hoc analysisrevealed that the improved performance of the mice was alreadyapparent at 48 h after injury, and this trend continued and becamemore marked over the remaining weeks. By the last day of thestudy (28 days), most of the mice treated with DHA injection aloneor DHA-enriched diet in addition to DHA injection showed frequentor consistent plantar stepping with coordination and parallel pawposition (BMS scores 6.8±0.3 and 5.8±0.2, respectively). In contrast,mice which received the saline injection or a DHA-enriched diet
alone, only displayed occasional stepping or steppingwithout coordina-tion (BMS scores 4.6±0.5 and 3.9±0.4, respectively). In addition, finelocomotor control assessed by the BMS subscores also showed asignificantly improved recovery in both groups of DHA-injected mice(with or without DHA diet), from 10 to 28 days post-injury comparedwith saline-treated controls (Pb0.05; Fig. 8B). Throughout the 4-weekobservation period, there was no statistically significant differencebetween the two DHA-injection groups (on DHA-enriched diet or con-trol diet), although from 10 to 28 days post-injury, mice treated withthe DHA-enriched diet in addition to DHA injection tended to show bet-ter locomotor performance than those treated with the DHA injectionalone, in terms of BMS and BMS subscores (P>0.05; Figs. 8A,B). Micewhich received the DHA-enriched diet alone did not differ significantlyfrom the saline-controls, throughout the entire recovery period.
DHA treatment increases neuronal survival after SCI. Examinationof NeuN labelling at 28 days post-injury revealed that mice thatreceived a DHA injection alone 30 min after injury or a DHA-enriched

Fig. 3. APC immunoreactivity in the mouse spinal cord. Representative images were taken from the lateral white matter of the spinal cord at the T12 compression epicentre. The APClabelling in the laminectomy control group was similar to that of the naïve tissue. After compression SCI, a continuous loss of APC immunoreactivity was seen from 1 to 28 days.Scale bar=50 μm.
18 S.-N. Lim et al. / Experimental Neurology 239 (2013) 13–27
diet in addition toDHA injection, had substantiallymore labelled cells inboth the dorsal and ventral horns at the epicentre compared to thesaline-injected mice (Figs. 9A–O). The quantitative analysis of NeuN la-belling confirmed these differences, with both groups that received aDHA injection resulting in significantly more NeuN labelled cells thansaline controls in the dorsal (average 89% survival in DHA-injectedmice vs. 48% in saline-treated controls, Pb0.05; Fig. 10A) and ventralhorns (average 64% survival in DHA-injected mice vs. 31% in saline-treated controls, Pb0.05; Fig. 10A). There was no statistically significantdifference in NeuN-labelled cells between the two DHA-injectiongroups (on DHA-enriched diet or control diet). Importantly, the NeuNlabelling in the dorsal horns of both DHA-injected groups was not sig-nificantly different from that of naïve mice (Pb0.05; Fig. 10A). The die-tary supplementation with DHA for 4 weeks following injury withoutan acute bolus of DHA did not have any neuroprotective effect inmouse compression SCI. Neuronal survival was similar to that seen inthe saline injected controls, i.e. 56.6±11.9% vs. 48.2±8.0% in the dorsalhorn, and 44.7±5.5% vs. 30.7 vs. 6% in the ventral horn (all compari-sons, P>0.05; Fig. 10A).
DHA treatment increases oligodendrocyte survival after SCI. Both DHA-treatment regimens increased the number of APC labelled oligoden-drocytes in the dorsal column and lateral white matter at 28 dayspost-injury, compared with saline-treated controls (Figs. 9P–Y). De-tailed quantification confirmed these differences, with the numberof surviving spinal cord oligodendrocytes in four different white mat-ter tracts being significantly increased in mice treated with DHAinjection alone (24% to 40% of oligodendrocytes were protected) orDHA-enriched diet in addition to DHA injection (25% to 55% of oligo-dendrocytes were protected) than in the saline-treated controls(Pb0.05; Fig. 10B). However, there were no significant differencesbetween the two DHA-injected groups (on DHA-enriched diet orcontrol diet). The APC labelling in all four white matter tracts of themice receiving DHA injection plus DHA enriched diet, and in thelateral and ventral white matter of mice receiving DHA injection
alone was not significantly different from that in laminectomy mice(P>0.05; Fig. 10B). In addition, there was no significant differencein oligodendrocyte survival between the DHA diet alone and salinegroups, in any of the four regions analysed (Fig. 10B). However, ineach case there was a trend towards more survival in the DHA dietgroup.
DHA treatment ameliorates non-phosphorylated neurofilament loss afterSCI. At 28 days post-injury, mice that received a DHA injection alone30 min after injury or a DHA-enriched diet in addition to DHA injec-tion, had substantially more SMI32-labelled axons in both the lateraland ventral white matter compared to the saline-injected mice(Figs. 11A–J). Quantification confirmed this protection of SMI32-positive axons in both DHA-injected groups compared with thesaline controls, with 30% and 37% more SMI32-positive axonsfound in the ventral and lateral white matter, respectively, in micereceiving DHA injection and DHA diet; 24% and 36% more SMI32-positive axons were found in the ventral and lateral white mater, re-spectively, in mice receiving DHA injection alone (Pb0.05; Fig. 12A).Moreover, in the lateral and ventral white matter, the SMI32 label-ling of axons in both the DHA-injected groups was not significantlydifferent from that of naïve mice. However, there were no signifi-cant differences between the two DHA-injected groups (P>0.05).We did not find any axonal protection effect conferred by the DHAdiet alone (Fig. 12A).
DHA treatment attenuates microglial/macrophage activation. In the spi-nal cord of the naïve mice, Iba1 immunolabelling showed small rest-ing microglial cells with long, ramified thin processes (Figs. 11K,P).The microglia shape in saline-treated controls was larger and irregu-lar. At 28 days after surgery, there appeared to be less Iba1 immuno-reactivity in mice injected with DHA (either on DHA-enriched diet orcontrol diet) compared with those injected with saline (Figs. 11M,R;O,T). Detailed quantification confirmed that the DHA-injected groupshad a significantly lower level of Iba1 expression in the dorsal
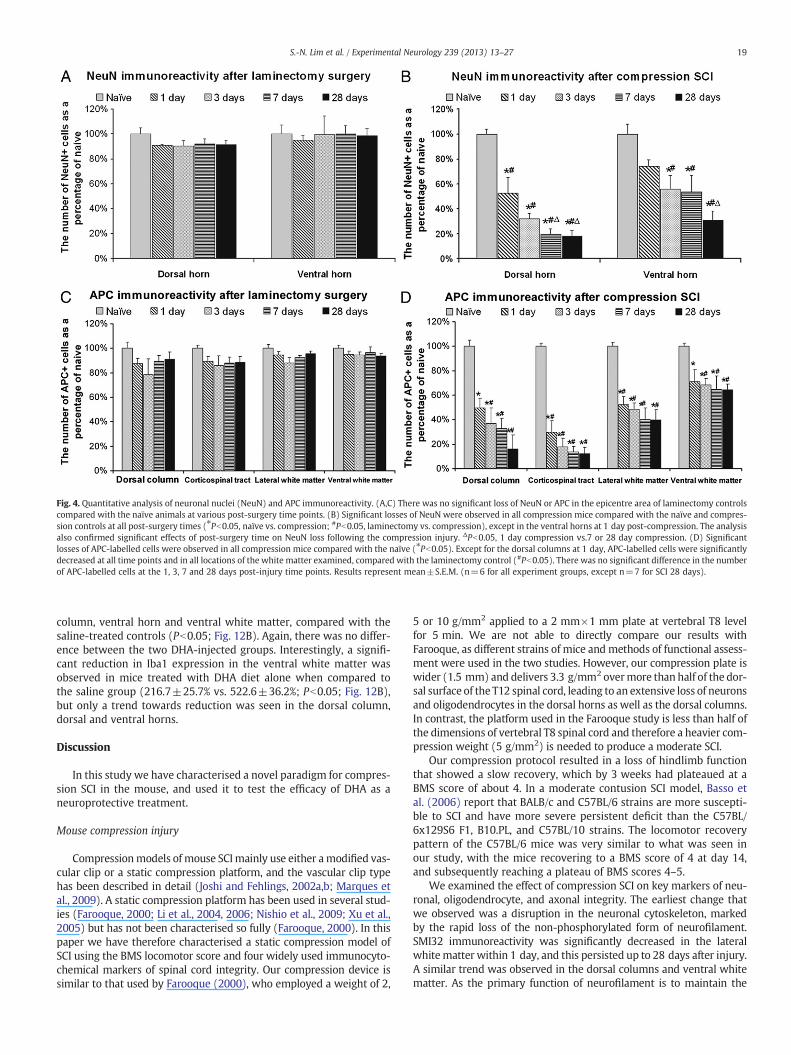
Fig. 4. Quantitative analysis of neuronal nuclei (NeuN) and APC immunoreactivity. (A,C) There was no significant loss of NeuN or APC in the epicentre area of laminectomy controlscompared with the naïve animals at various post-surgery time points. (B) Significant losses of NeuN were observed in all compression mice compared with the naïve and compres-sion controls at all post-surgery times (⁎Pb0.05, naïve vs. compression; #Pb0.05, laminectomy vs. compression), except in the ventral horns at 1 day post-compression. The analysisalso confirmed significant effects of post-surgery time on NeuN loss following the compression injury. ΔPb0.05, 1 day compression vs.7 or 28 day compression. (D) Significantlosses of APC-labelled cells were observed in all compression mice compared with the naïve (⁎Pb0.05). Except for the dorsal columns at 1 day, APC-labelled cells were significantlydecreased at all time points and in all locations of the white matter examined, compared with the laminectomy control (#Pb0.05). There was no significant difference in the numberof APC-labelled cells at the 1, 3, 7 and 28 days post-injury time points. Results represent mean±S.E.M. (n=6 for all experiment groups, except n=7 for SCI 28 days).
19S.-N. Lim et al. / Experimental Neurology 239 (2013) 13–27
column, ventral horn and ventral white matter, compared with thesaline-treated controls (Pb0.05; Fig. 12B). Again, there was no differ-ence between the two DHA-injected groups. Interestingly, a signifi-cant reduction in Iba1 expression in the ventral white matter wasobserved in mice treated with DHA diet alone when compared tothe saline group (216.7±25.7% vs. 522.6±36.2%; Pb0.05; Fig. 12B),but only a trend towards reduction was seen in the dorsal column,dorsal and ventral horns.
Discussion
In this study we have characterised a novel paradigm for compres-sion SCI in the mouse, and used it to test the efficacy of DHA as aneuroprotective treatment.
Mouse compression injury
Compressionmodels of mouse SCImainly use either amodified vas-cular clip or a static compression platform, and the vascular clip typehas been described in detail (Joshi and Fehlings, 2002a,b; Marques etal., 2009). A static compression platform has been used in several stud-ies (Farooque, 2000; Li et al., 2004, 2006; Nishio et al., 2009; Xu et al.,2005) but has not been characterised so fully (Farooque, 2000). In thispaper we have therefore characterised a static compression model ofSCI using the BMS locomotor score and four widely used immunocyto-chemical markers of spinal cord integrity. Our compression device issimilar to that used by Farooque (2000), who employed a weight of 2,
5 or 10 g/mm2 applied to a 2 mm×1 mm plate at vertebral T8 levelfor 5 min. We are not able to directly compare our results withFarooque, as different strains of mice and methods of functional assess-ment were used in the two studies. However, our compression plate iswider (1.5 mm) and delivers 3.3 g/mm2 overmore than half of the dor-sal surface of the T12 spinal cord, leading to an extensive loss of neuronsand oligodendrocytes in the dorsal horns as well as the dorsal columns.In contrast, the platform used in the Farooque study is less than half ofthe dimensions of vertebral T8 spinal cord and therefore a heavier com-pression weight (5 g/mm2) is needed to produce a moderate SCI.
Our compression protocol resulted in a loss of hindlimb functionthat showed a slow recovery, which by 3 weeks had plateaued at aBMS score of about 4. In a moderate contusion SCI model, Basso etal. (2006) report that BALB/c and C57BL/6 strains are more suscepti-ble to SCI and have more severe persistent deficit than the C57BL/6x129S6 F1, B10.PL, and C57BL/10 strains. The locomotor recoverypattern of the C57BL/6 mice was very similar to what was seen inour study, with the mice recovering to a BMS score of 4 at day 14,and subsequently reaching a plateau of BMS scores 4–5.
We examined the effect of compression SCI on key markers of neu-ronal, oligodendrocyte, and axonal integrity. The earliest change thatwe observed was a disruption in the neuronal cytoskeleton, markedby the rapid loss of the non-phosphorylated form of neurofilament.SMI32 immunoreactivity was significantly decreased in the lateralwhitematter within 1 day, and this persisted up to 28 days after injury.A similar trend was observed in the dorsal columns and ventral whitematter. As the primary function of neurofilament is to maintain the
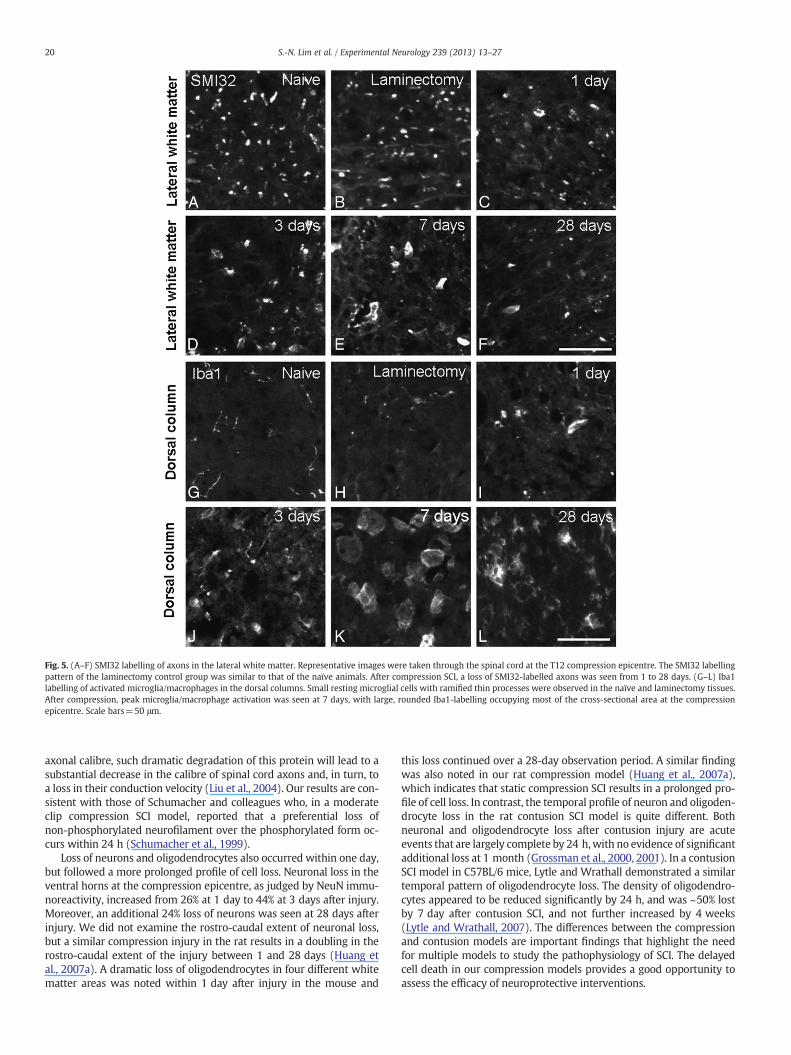
Fig. 5. (A–F) SMI32 labelling of axons in the lateral white matter. Representative images were taken through the spinal cord at the T12 compression epicentre. The SMI32 labellingpattern of the laminectomy control group was similar to that of the naïve animals. After compression SCI, a loss of SMI32-labelled axons was seen from 1 to 28 days. (G–L) Iba1labelling of activated microglia/macrophages in the dorsal columns. Small resting microglial cells with ramified thin processes were observed in the naïve and laminectomy tissues.After compression, peak microglia/macrophage activation was seen at 7 days, with large, rounded Iba1-labelling occupying most of the cross-sectional area at the compressionepicentre. Scale bars=50 μm.
20 S.-N. Lim et al. / Experimental Neurology 239 (2013) 13–27
axonal calibre, such dramatic degradation of this protein will lead to asubstantial decrease in the calibre of spinal cord axons and, in turn, toa loss in their conduction velocity (Liu et al., 2004). Our results are con-sistent with those of Schumacher and colleagues who, in a moderateclip compression SCI model, reported that a preferential loss ofnon-phosphorylated neurofilament over the phosphorylated form oc-curs within 24 h (Schumacher et al., 1999).
Loss of neurons and oligodendrocytes also occurred within one day,but followed a more prolonged profile of cell loss. Neuronal loss in theventral horns at the compression epicentre, as judged by NeuN immu-noreactivity, increased from 26% at 1 day to 44% at 3 days after injury.Moreover, an additional 24% loss of neurons was seen at 28 days afterinjury. We did not examine the rostro-caudal extent of neuronal loss,but a similar compression injury in the rat results in a doubling in therostro-caudal extent of the injury between 1 and 28 days (Huang etal., 2007a). A dramatic loss of oligodendrocytes in four different whitematter areas was noted within 1 day after injury in the mouse and
this loss continued over a 28-day observation period. A similar findingwas also noted in our rat compression model (Huang et al., 2007a),which indicates that static compression SCI results in a prolonged pro-file of cell loss. In contrast, the temporal profile of neuron and oligoden-drocyte loss in the rat contusion SCI model is quite different. Bothneuronal and oligodendrocyte loss after contusion injury are acuteevents that are largely complete by 24 h,with no evidence of significantadditional loss at 1 month (Grossman et al., 2000, 2001). In a contusionSCI model in C57BL/6 mice, Lytle and Wrathall demonstrated a similartemporal pattern of oligodendrocyte loss. The density of oligodendro-cytes appeared to be reduced significantly by 24 h, and was ~50% lostby 7 day after contusion SCI, and not further increased by 4 weeks(Lytle and Wrathall, 2007). The differences between the compressionand contusion models are important findings that highlight the needfor multiple models to study the pathophysiology of SCI. The delayedcell death in our compression models provides a good opportunity toassess the efficacy of neuroprotective interventions.

Fig. 6. Quantitative analysis of SMI32 and Iba1 immunoreactivity in naïve mice, 28 days laminectomy or 1, 3, 7 or 28 days following compression SCI. (A) There was no significantloss of SMI32-labelled axons in the epicentre of laminectomy controls compared with the naïve animals. A significant loss of SMI32 immunoreactivity in the lateral white matteroccurred within 1 day post-injury, and was sustained up to 28 days, compared with both the naïve and laminectomy controls. The loss of SMI32 immunoreactivity in the dorsalcolumns and ventral white matter was also noted within 1 day after compression, but this only reached statistical significance at 28 days post-injury ( Pb0.05 vs. naïvecontrols; #Pb0.05 vs. laminectomy controls). (B) Quantification confirmed that Iba1 was dramatically and significantly increased at days 7 and 28 after compression injury in allfour regions analysed. ⁎Pb0.05, vs. naïve; #Pb0.05, vs. laminectomy; ΔPb0.05, vs. SCI 1 day; §Pb0.05, vs. SCI 3 days; ¶Pb0.05, vs. SCI 28 days. Results represent mean±S.E.M.(n=6 for all experimental groups, except n=7 for SCI 28 days).
21S.-N. Lim et al. / Experimental Neurology 239 (2013) 13–27
The difference in the temporal profile of neuronal and oligodendro-cyte loss between contusion and compression SCI may reflect differ-ences in the pathogenesis of the secondary insults such as ischaemia,inflammation and free radical production. Iba1 immunostaining re-vealed that microglial/macrophage activation was evident by 1 day,with a dramatic peak at 7 days after compression injury and declinedat 28 days, but remained elevated above the naïve controls. A similartemporal pattern ofmicroglia/macrophage activation has been reportedin C57BL/6mice receiving contusion SCI (Kigerl et al., 2006), whichmayindicate that other mechanisms linked to cell death account for thedifferent temporal profiles of cell loss in the contusion and compressionmodels. The delayed phase of the microglia/macrophage response maybe related to a reopening of the blood–spinal cord barrier (Popovich etal., 1996) and/or secondary axonal degeneration in ascending and de-scending tracts (Popovich et al., 1997; Sroga et al., 2003).
DHA treatment
Previous studies in our laboratory have shown that the acute i.v.administration of 250 nmol/kg DHA has significant neuroprotectiveeffects after compression SCI in rats, and that neuroprotection and func-tional recovery are greater if the acute injection is combined with
chronic administration of DHA in the diet (Huang et al., 2007b). In thepresent study, the neuroprotective effect of a single bolus DHA30 min after injury was clearly confirmed in the mouse compressionSCI model. However, although there was a trend towards furtherneuroprotection by combination with chronic dietary DHA supple-mentation, this effect was not statistically significant. Furthermore,dietary DHA alone for 4 weeks following compression injury didnot protect against either the neurological deficit or the histologicaldamage.
Our preliminary dose–response study showed that the maximumlocomotor recovery after SCI in mice could be obtained by an acuteadministration of 500 nmol/kg DHA. The neuroprotective effect of500 nmol/kg DHA was further confirmed in the subsequent study,and was clearly demonstrated by a number of end points, includingincreased neuronal and oligodendrocyte survival, non-phosphorylatedneurofilament protection, and decreased microglial/macrophage acti-vation. These parameters were all studied at 28 days post-injury. Anal-ysis at other time points would be informative, andwould allow amoredetailed correlation with the locomotor recovery.
Even though the dose we injected in mice (500 nmol/kg,i.e. 164 μg/kg) is double than what we used in rats (250 nmol/kg),it is still very small compared with the oral DHA dose which is
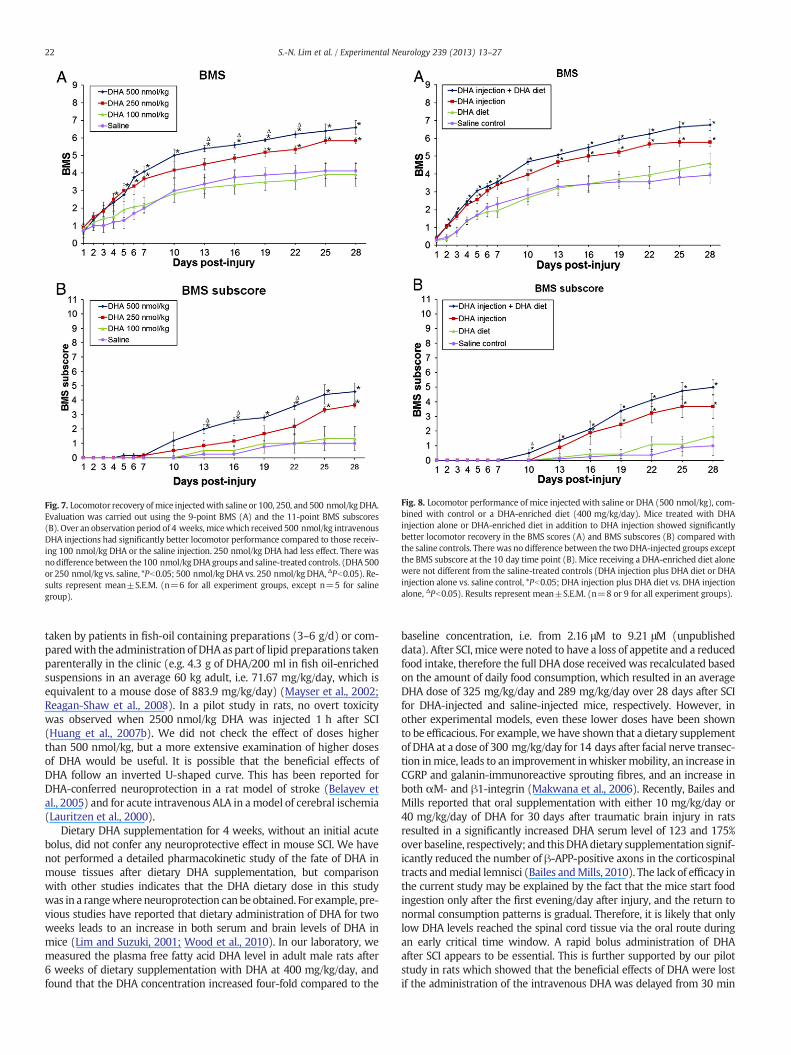
Fig. 7. Locomotor recovery ofmice injectedwith saline or 100, 250, and 500 nmol/kg DHA.Evaluation was carried out using the 9-point BMS (A) and the 11-point BMS subscores(B). Over an observation period of 4 weeks,micewhich received 500 nmol/kg intravenousDHA injections had significantly better locomotor performance compared to those receiv-ing 100 nmol/kg DHA or the saline injection. 250 nmol/kg DHA had less effect. There wasnodifference between the 100 nmol/kgDHAgroups and saline-treated controls. (DHA500or 250 nmol/kg vs. saline, *Pb0.05; 500 nmol/kg DHA vs. 250 nmol/kg DHA, ΔPb0.05). Re-sults represent mean±S.E.M. (n=6 for all experiment groups, except n=5 for salinegroup).
Fig. 8. Locomotor performance of mice injected with saline or DHA (500 nmol/kg), com-bined with control or a DHA-enriched diet (400 mg/kg/day). Mice treated with DHAinjection alone or DHA-enriched diet in addition to DHA injection showed significantlybetter locomotor recovery in the BMS scores (A) and BMS subscores (B) compared withthe saline controls. There was no difference between the two DHA-injected groups exceptthe BMS subscore at the 10 day time point (B). Mice receiving a DHA-enriched diet alonewere not different from the saline-treated controls (DHA injection plus DHA diet or DHAinjection alone vs. saline control, *Pb0.05; DHA injection plus DHA diet vs. DHA injectionalone, ΔPb0.05). Results represent mean±S.E.M. (n=8 or 9 for all experiment groups).
22 S.-N. Lim et al. / Experimental Neurology 239 (2013) 13–27
taken by patients in fish-oil containing preparations (3–6 g/d) or com-paredwith the administration of DHA as part of lipid preparations takenparenterally in the clinic (e.g. 4.3 g of DHA/200 ml in fish oil-enrichedsuspensions in an average 60 kg adult, i.e. 71.67 mg/kg/day, which isequivalent to a mouse dose of 883.9 mg/kg/day) (Mayser et al., 2002;Reagan-Shaw et al., 2008). In a pilot study in rats, no overt toxicitywas observed when 2500 nmol/kg DHA was injected 1 h after SCI(Huang et al., 2007b). We did not check the effect of doses higherthan 500 nmol/kg, but a more extensive examination of higher dosesof DHA would be useful. It is possible that the beneficial effects ofDHA follow an inverted U-shaped curve. This has been reported forDHA-conferred neuroprotection in a rat model of stroke (Belayev etal., 2005) and for acute intravenous ALA in amodel of cerebral ischemia(Lauritzen et al., 2000).
Dietary DHA supplementation for 4 weeks, without an initial acutebolus, did not confer any neuroprotective effect in mouse SCI. We havenot performed a detailed pharmacokinetic study of the fate of DHA inmouse tissues after dietary DHA supplementation, but comparisonwith other studies indicates that the DHA dietary dose in this studywas in a rangewhere neuroprotection can be obtained. For example, pre-vious studies have reported that dietary administration of DHA for twoweeks leads to an increase in both serum and brain levels of DHA inmice (Lim and Suzuki, 2001; Wood et al., 2010). In our laboratory, wemeasured the plasma free fatty acid DHA level in adult male rats after6 weeks of dietary supplementation with DHA at 400 mg/kg/day, andfound that the DHA concentration increased four-fold compared to the
baseline concentration, i.e. from 2.16 μM to 9.21 μM (unpublisheddata). After SCI, micewere noted to have a loss of appetite and a reducedfood intake, therefore the full DHA dose received was recalculated basedon the amount of daily food consumption, which resulted in an averageDHA dose of 325 mg/kg/day and 289 mg/kg/day over 28 days after SCIfor DHA-injected and saline-injected mice, respectively. However, inother experimental models, even these lower doses have been shownto be efficacious. For example, we have shown that a dietary supplementof DHA at a dose of 300 mg/kg/day for 14 days after facial nerve transec-tion inmice, leads to an improvement inwhiskermobility, an increase inCGRP and galanin-immunoreactive sprouting fibres, and an increase inboth αM- and β1-integrin (Makwana et al., 2006). Recently, Bailes andMills reported that oral supplementation with either 10 mg/kg/day or40 mg/kg/day of DHA for 30 days after traumatic brain injury in ratsresulted in a significantly increased DHA serum level of 123 and 175%over baseline, respectively; and thisDHAdietary supplementation signif-icantly reduced the number of β-APP-positive axons in the corticospinaltracts andmedial lemnisci (Bailes andMills, 2010). The lack of efficacy inthe current study may be explained by the fact that the mice start foodingestion only after the first evening/day after injury, and the return tonormal consumption patterns is gradual. Therefore, it is likely that onlylow DHA levels reached the spinal cord tissue via the oral route duringan early critical time window. A rapid bolus administration of DHAafter SCI appears to be essential. This is further supported by our pilotstudy in rats which showed that the beneficial effects of DHA were lostif the administration of the intravenous DHA was delayed from 30 min
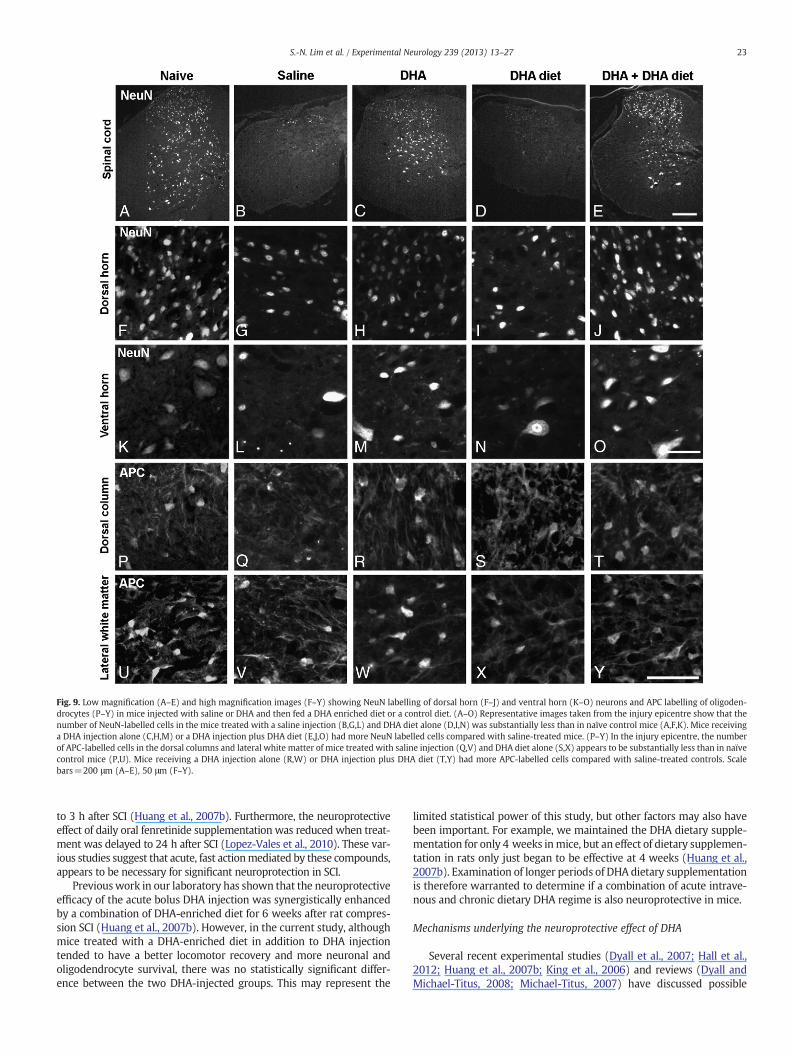
Fig. 9. Low magnification (A–E) and high magnification images (F–Y) showing NeuN labelling of dorsal horn (F–J) and ventral horn (K–O) neurons and APC labelling of oligoden-drocytes (P–Y) in mice injected with saline or DHA and then fed a DHA enriched diet or a control diet. (A–O) Representative images taken from the injury epicentre show that thenumber of NeuN-labelled cells in the mice treated with a saline injection (B,G,L) and DHA diet alone (D,I,N) was substantially less than in naïve control mice (A,F,K). Mice receivinga DHA injection alone (C,H,M) or a DHA injection plus DHA diet (E,J,O) had more NeuN labelled cells compared with saline-treated mice. (P–Y) In the injury epicentre, the numberof APC-labelled cells in the dorsal columns and lateral white matter of mice treated with saline injection (Q,V) and DHA diet alone (S,X) appears to be substantially less than in naïvecontrol mice (P,U). Mice receiving a DHA injection alone (R,W) or DHA injection plus DHA diet (T,Y) had more APC-labelled cells compared with saline-treated controls. Scalebars=200 μm (A–E), 50 μm (F–Y).
23S.-N. Lim et al. / Experimental Neurology 239 (2013) 13–27
to 3 h after SCI (Huang et al., 2007b). Furthermore, the neuroprotectiveeffect of daily oral fenretinide supplementation was reduced when treat-ment was delayed to 24 h after SCI (Lopez-Vales et al., 2010). These var-ious studies suggest that acute, fast actionmediated by these compounds,appears to be necessary for significant neuroprotection in SCI.
Previouswork in our laboratory has shown that the neuroprotectiveefficacy of the acute bolus DHA injection was synergistically enhancedby a combination of DHA-enriched diet for 6 weeks after rat compres-sion SCI (Huang et al., 2007b). However, in the current study, althoughmice treated with a DHA-enriched diet in addition to DHA injectiontended to have a better locomotor recovery and more neuronal andoligodendrocyte survival, there was no statistically significant differ-ence between the two DHA-injected groups. This may represent the
limited statistical power of this study, but other factors may also havebeen important. For example, we maintained the DHA dietary supple-mentation for only 4 weeks inmice, but an effect of dietary supplemen-tation in rats only just began to be effective at 4 weeks (Huang et al.,2007b). Examination of longer periods of DHA dietary supplementationis therefore warranted to determine if a combination of acute intrave-nous and chronic dietary DHA regime is also neuroprotective in mice.
Mechanisms underlying the neuroprotective effect of DHA
Several recent experimental studies (Dyall et al., 2007; Hall et al.,2012; Huang et al., 2007b; King et al., 2006) and reviews (Dyall andMichael-Titus, 2008; Michael-Titus, 2007) have discussed possible

Fig. 10. Quantitative analysis of NeuN and APC immunoreactivity. (A) Significant losses of NeuN-labelled cells were observed in all injured mice compared with the naive controls(⁎Pb0.05), except in the dorsal horns of both DHA-injected groups. There were significantly more NeuN-labelled cells in the dorsal and ventral horns in the two DHA-injectedgroups compared with saline-treated controls (#Pb0.05). The percentage of surviving dorsal horn neurons was also significantly increased in mice treated with DHA injectionbolus (either on control or DHA diet) compared to the mice treated with DHA-enriched diet alone (ΔPb0.05). (B) Significant losses in APC-labelled cells were observed in all injuredmice compared with the naïve controls (⁎Pb0.05), except in the lateral and ventral white matter of the two DHA-injected groups, and the dorsal column and corticospinal tract ofthe DHA injection plus DHA-enriched diet mice. There were significantly more APC-labelled cells in the four different white matter tracts in DHA-injected mice compared withsaline-treated controls (#Pb0.05). Results represent mean±S.E.M. (n=6 per group).
24 S.-N. Lim et al. / Experimental Neurology 239 (2013) 13–27
mechanisms underlying the neuroprotective effects of DHA, so only keyelements related to the current study will be discussed.
Following mouse compression injury, we have found that DHAsuppressed macrophage/microglial activation in both the grey andwhite matter. Although the role of the inflammatory response in SCIremains controversial, the general consensus is that functional out-come after SCI is improved by therapies that reduce inflammation(Kwon et al., 2004; Popovich and Jones, 2003). After SCI, activatedmicroglia express the retinoic acid receptor (Mey et al., 2005), andit has been shown that DHA is an endogenous ligand for retinoid X re-ceptor (RXRs) (de Urquiza et al., 2000), and can activate these recep-tors at low micromolar concentrations (Lengqvist et al., 2004). An invitro study has found that 9-cis-retinoic acid, a RXR agonist, sup-presses the lipopolysaccharide-triggered inflammatory response ofmicroglia and astrocytes (Xu and Drew, 2006). Thus, DHAmay reduceinflammation by modulating the response of reactive microglia.
Finally, the anti-inflammatory and anti-oxidative properties of DHAcan be mediated partly via its metabolites, the D-series of resolvinsand docosatrienes (Serhan, 2004).
Although we did not see the major effects of DHA dietary supple-mentation, chronic changes mediated by DHA treatment may play animportant role. For example, the neuroprotective effect of DHA mayinvolve the activation of the GluR2 receptor. The GluR2 subunit isdecreased following SCI and leads to an increase in AMPA-mediatedCa2+ influx following injury (Wu et al., 2005). DHA has been shownto reduce AMPA-mediated excitotoxicity in the hippocampus via amechanism that includes the modulation of both the GluR1 and theGluR2 subunit (Menard et al., 2009). Work in our laboratory hasshown that DHA can increase the expression of the GluR2 glutamatereceptor subunit in aged rats (Dyall et al., 2007). The chronic expo-sure to DHA also affects membrane properties, and leads to anincreased accumulation of phosphatidylserine in membranes, which

Fig. 11. SMI32 labelling of axons (A–J) and Iba1 labelling of activated microglia/macrophages (K–T) in mice injected with saline or DHA and then fed a DHA enriched diet or controldiet. (A–J) Representative images taken from the injury epicentre show that the number of SMI32-labelled axons in the mice treated with saline injection (B,G,) and a DHA dietalone (D,I) was substantially less than in naïve control mice (A,F). Mice receiving a DHA injection alone (C,H) or DHA injection and DHA diet (E,J) had more SMI32-labelledaxons compared with saline-treated controls. (K–T) In the epicentre, small resting microglia cells with ramified thin processes were noted in the naïve tissue (K,P). The microgliashape in saline-treated controls (L,Q) and DHA diet alone (N,S) was larger and irregular. At 28 days post-injury, there appeared to be less Iba1 immunoreactivity in mice receiving aDHA injection alone (M,R) or a DHA injection and a DHA-enriched diet (O,T), compared with saline-treated controls. Scale bars=50 μm.
25S.-N. Lim et al. / Experimental Neurology 239 (2013) 13–27
has profound consequences for cell survival mechanisms. The changein these acidic phospholipids facilitates the translocation and activa-tion of the phosphatidylinositol 3-kinase/Akt pathway (Akbar et al.,2005; Kim et al., 2000, 2001).
The neuroprotective effect of DHA may also involve specific freefatty acid receptors, such as the recently identified G-protein-coupledreceptor (GPR)40,which haswidespread expression in themonkey spi-nal cord (Ma et al., 2007). It has been suggested that GPR40 is activatedby PUFAs (Briscoe et al., 2003; Ma et al., 2007), and a role for the DHA/GPR40 signalling pathway in adult neurogenesis has been proposed byYamashima (2008). Finally, the fatty acid binding proteins (FABPs), afamily of carrier proteins for fatty acids, eicosanoids and retinoids,which can facilitate the transfer of fatty acids between extra- and intra-cellularmembranes (Chmurzynska, 2006) and transport lipophilicmol-ecules from outer cellular membranes to certain intracellular receptorssuch as PPARs (Tan et al., 2002), may also play an important role in theneuroprotective effect of DHA. B-FABP may be especially important,since DHA has a high affinity for it (Xu et al., 1996).
Conclusions
In this study we have characterised a compression model of SCI inthe mouse, and shown that it results in a prolonged period of neuro-nal and oligodendrocyte cell death. In the same mouse model, we
have shown that an i.v. bolus of 500 nmol/kg DHA promotes functionalrecovery and reduces neurofilament loss, neuronal and oligodendrocytecell death, and microglial/macrophage activation. This study thus pro-vides a detailed validation in a second species (mouse) of the significantneuroprotective potential of DHA after SCI.
Acknowledgments
We acknowledge the generous support of Chang Gung MemorialHospital, Taiwan (CMRPG360622). We thank CRODA Healthcare forthe donation of DHA oil, and Dr Julian Taylor (Toledo) for assistancewith design of the compression platform.
References
Akbar, M., Calderon, F., Wen, Z., Kim, H.Y., 2005. Docosahexaenoic acid: a positive mod-ulator of Akt signaling in neuronal survival. Proc. Natl. Acad. Sci. U. S. A. 102,10858–10863.
Bailes, J.E., Mills, J.D., 2010. Docosahexaenoic acid reduces traumatic axonal injury in arodent head injury model. J. Neurotrauma 27, 1617–1624.
Basso, D.M., Fisher, L.C., Anderson, A.J., Jakeman, L.B., McTigue, D.M., Popovich, P.G.,2006. Basso Mouse Scale for locomotion detects differences in recovery after spinalcord injury in five common mouse strains. J. Neurotrauma 23, 635–659.
Bazan, N.G., Molina, M.F., Gordon, W.C., 2011. Docosahexaenoic acid signalolipidomicsin nutrition: significance in aging, neuroinflammation, macular degeneration,Alzheimer's, and other neurodegenerative diseases. Annu. Rev. Nutr. 31, 321–351.
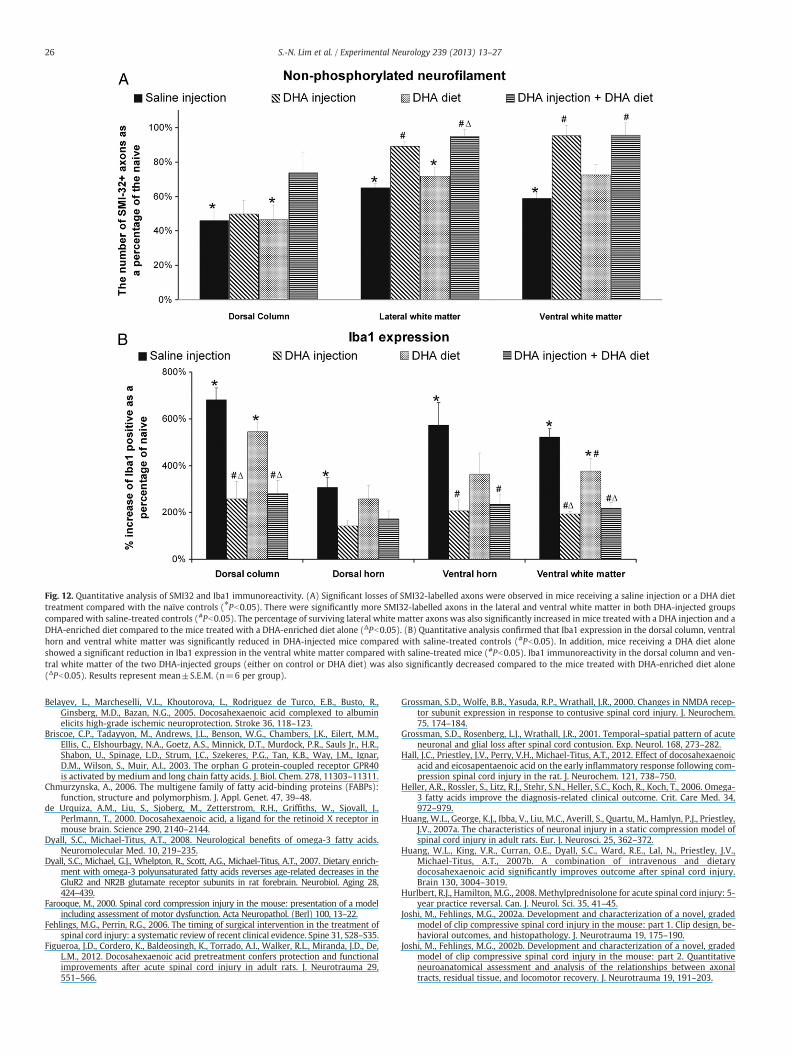
Fig. 12. Quantitative analysis of SMI32 and Iba1 immunoreactivity. (A) Significant losses of SMI32-labelled axons were observed in mice receiving a saline injection or a DHA diettreatment compared with the naïve controls (⁎Pb0.05). There were significantly more SMI32-labelled axons in the lateral and ventral white matter in both DHA-injected groupscompared with saline-treated controls (#Pb0.05). The percentage of surviving lateral white matter axons was also significantly increased in mice treated with a DHA injection and aDHA-enriched diet compared to the mice treated with a DHA-enriched diet alone (ΔPb0.05). (B) Quantitative analysis confirmed that Iba1 expression in the dorsal column, ventralhorn and ventral white matter was significantly reduced in DHA-injected mice compared with saline-treated controls (#Pb0.05). In addition, mice receiving a DHA diet aloneshowed a significant reduction in Iba1 expression in the ventral white matter compared with saline-treated mice (#Pb0.05). Iba1 immunoreactivity in the dorsal column and ven-tral white matter of the two DHA-injected groups (either on control or DHA diet) was also significantly decreased compared to the mice treated with DHA-enriched diet alone(ΔPb0.05). Results represent mean±S.E.M. (n=6 per group).
26 S.-N. Lim et al. / Experimental Neurology 239 (2013) 13–27
Belayev, L., Marcheselli, V.L., Khoutorova, L., Rodriguez de Turco, E.B., Busto, R.,Ginsberg, M.D., Bazan, N.G., 2005. Docosahexaenoic acid complexed to albuminelicits high-grade ischemic neuroprotection. Stroke 36, 118–123.
Briscoe, C.P., Tadayyon, M., Andrews, J.L., Benson, W.G., Chambers, J.K., Eilert, M.M.,Ellis, C., Elshourbagy, N.A., Goetz, A.S., Minnick, D.T., Murdock, P.R., Sauls Jr., H.R.,Shabon, U., Spinage, L.D., Strum, J.C., Szekeres, P.G., Tan, K.B., Way, J.M., Ignar,D.M., Wilson, S., Muir, A.I., 2003. The orphan G protein-coupled receptor GPR40is activated by medium and long chain fatty acids. J. Biol. Chem. 278, 11303–11311.
Chmurzynska, A., 2006. The multigene family of fatty acid-binding proteins (FABPs):function, structure and polymorphism. J. Appl. Genet. 47, 39–48.
de Urquiza, A.M., Liu, S., Sjoberg, M., Zetterstrom, R.H., Griffiths, W., Sjovall, J.,Perlmann, T., 2000. Docosahexaenoic acid, a ligand for the retinoid X receptor inmouse brain. Science 290, 2140–2144.
Dyall, S.C., Michael-Titus, A.T., 2008. Neurological benefits of omega-3 fatty acids.Neuromolecular Med. 10, 219–235.
Dyall, S.C., Michael, G.J., Whelpton, R., Scott, A.G., Michael-Titus, A.T., 2007. Dietary enrich-ment with omega-3 polyunsaturated fatty acids reverses age-related decreases in theGluR2 and NR2B glutamate receptor subunits in rat forebrain. Neurobiol. Aging 28,424–439.
Farooque, M., 2000. Spinal cord compression injury in the mouse: presentation of a modelincluding assessment of motor dysfunction. Acta Neuropathol. (Berl) 100, 13–22.
Fehlings, M.G., Perrin, R.G., 2006. The timing of surgical intervention in the treatment ofspinal cord injury: a systematic review of recent clinical evidence. Spine 31, S28–S35.
Figueroa, J.D., Cordero, K., Baldeosingh, K., Torrado, A.I., Walker, R.L., Miranda, J.D., De,L.M., 2012. Docosahexaenoic acid pretreatment confers protection and functionalimprovements after acute spinal cord injury in adult rats. J. Neurotrauma 29,551–566.
Grossman, S.D., Wolfe, B.B., Yasuda, R.P., Wrathall, J.R., 2000. Changes in NMDA recep-tor subunit expression in response to contusive spinal cord injury. J. Neurochem.75, 174–184.
Grossman, S.D., Rosenberg, L.J., Wrathall, J.R., 2001. Temporal–spatial pattern of acuteneuronal and glial loss after spinal cord contusion. Exp. Neurol. 168, 273–282.
Hall, J.C., Priestley, J.V., Perry, V.H., Michael-Titus, A.T., 2012. Effect of docosahexaenoicacid and eicosapentaenoic acid on the early inflammatory response following com-pression spinal cord injury in the rat. J. Neurochem. 121, 738–750.
Heller, A.R., Rossler, S., Litz, R.J., Stehr, S.N., Heller, S.C., Koch, R., Koch, T., 2006. Omega-3 fatty acids improve the diagnosis-related clinical outcome. Crit. Care Med. 34,972–979.
Huang, W.L., George, K.J., Ibba, V., Liu, M.C., Averill, S., Quartu, M., Hamlyn, P.J., Priestley,J.V., 2007a. The characteristics of neuronal injury in a static compression model ofspinal cord injury in adult rats. Eur. J. Neurosci. 25, 362–372.
Huang, W.L., King, V.R., Curran, O.E., Dyall, S.C., Ward, R.E., Lal, N., Priestley, J.V.,Michael-Titus, A.T., 2007b. A combination of intravenous and dietarydocosahexaenoic acid significantly improves outcome after spinal cord injury.Brain 130, 3004–3019.
Hurlbert, R.J., Hamilton, M.G., 2008. Methylprednisolone for acute spinal cord injury: 5-year practice reversal. Can. J. Neurol. Sci. 35, 41–45.
Joshi, M., Fehlings, M.G., 2002a. Development and characterization of a novel, gradedmodel of clip compressive spinal cord injury in the mouse: part 1. Clip design, be-havioral outcomes, and histopathology. J. Neurotrauma 19, 175–190.
Joshi, M., Fehlings, M.G., 2002b. Development and characterization of a novel, gradedmodel of clip compressive spinal cord injury in the mouse: part 2. Quantitativeneuroanatomical assessment and analysis of the relationships between axonaltracts, residual tissue, and locomotor recovery. J. Neurotrauma 19, 191–203.

27S.-N. Lim et al. / Experimental Neurology 239 (2013) 13–27
Jump, D.B., 2002. Dietary polyunsaturated fatty acids and regulation of gene transcrip-tion. Curr. Opin. Lipidol. 13, 155–164.
Kigerl, K.A., McGaughy, V.M., Popovich, P.G., 2006. Comparative analysis of lesiondevelopment and intraspinal inflammation in four strains of mice following spinalcontusion injury. J. Comp. Neurol. 494, 578–594.
Kim, H.Y., Akbar, M., Lau, A., Edsall, L., 2000. Inhibition of neuronal apoptosis bydocosahexaenoic acid (22:6n−3). Role of phosphatidylserine in antiapoptoticeffect. J. Biol. Chem. 275, 35215–35223.
Kim, H.Y., Akbar, M., Kim, K.Y., 2001. Inhibition of neuronal apoptosis by polyunsatu-rated fatty acids. J. Mol. Neurosci. 16, 223–227.
King, V.R., Huang, W.L., Dyall, S.C., Curran, O.E., Priestley, J.V., Michael-Titus, A.T., 2006.Omega-3 fatty acids improve recovery, whereas omega-6 fatty acids worsen out-come, after spinal cord injury in the adult rat. J. Neurosci. 26, 4672–4680.
Kwon, B.K., Tetzlaff, W., Grauer, J.N., Beiner, J., Vaccaro, A.R., 2004. Pathophysiology andpharmacologic treatment of acute spinal cord injury. Spine J. 4, 451–464.
Kwon, B.K., Sekhon, L.H., Fehlings, M.G., 2010. Emerging repair, regeneration, and transla-tional research advances for spinal cord injury. Spine (Phila Pa 1976) 35, S263–S270.
Kwon, B.K., Okon, E., Hillyer, J., Mann, C., Baptiste, D., Weaver, L.C., Fehlings, M.G., Tetzlaff,W., 2011a. A systematic review of non-invasive pharmacologic neuroprotectivetreatments for acute spinal cord injury. J. Neurotrauma 28, 1545–1588.
Kwon, B.K., Okon, E.B., Tsai, E., Beattie, M.S., Bresnahan, J.C., Magnuson, D.K., Reier, P.J.,McTigue, D.M., Popovich, P.G., Blight, A.R., Oudega, M., Guest, J.D., Weaver, L.C.,Fehlings, M.G., Tetzlaff, W., 2011b. A grading system to evaluate objectively thestrength of pre-clinical data of acute neuroprotective therapies for clinical transla-tion in spinal cord injury. J. Neurotrauma 28, 1525–1543.
Lafourcade, M., Larrieu, T., Mato, S., Duffaud, A., Sepers, M., Matias, I., Smedt-Peyrusse,V., Labrousse, V.F., Bretillon, L., Matute, C., Rodriguez-Puertas, R., Laye, S., Manzoni,O.J., 2011. Nutritional omega-3 deficiency abolishes endocannabinoid-mediatedneuronal functions. Nat. Neurosci. 14, 345–350.
Lauritzen, I., Blondeau, N., Heurteaux, C., Widmann, C., Romey, G., Lazdunski, M., 2000.Polyunsaturated fatty acids are potent neuroprotectors. EMBO J. 19, 1784–1793.
Lengqvist, J., Mata, D.U., Bergman, A.C., Willson, T.M., Sjovall, J., Perlmann, T., Griffiths,W.J., 2004. Polyunsaturated fatty acids including docosahexaenoic and arachidonicacid bind to the retinoid X receptor alpha ligand-binding domain. Mol. Cell. Prote-omics 3, 692–703.
Li, G.L., Farooque, M., Isaksson, J., Olsson, Y., 2004. Changes in synapses and axons dem-onstrated by synaptophysin immunohistochemistry following spinal cord com-pression trauma in the rat and mouse. Biomed. Environ. Sci. 17, 281–290.
Li, Y., Oskouian, R.J., Day, Y.J., Rieger, J.M., Liu, L., Kern, J.A., Linden, J., 2006. Mouse spinalcord compression injury is reduced by either activation of the adenosine A2A re-ceptor on bone marrow-derived cells or deletion of the A2A receptor on non-bone marrow-derived cells. Neuroscience 141, 2029–2039.
Lim, S., Suzuki, H., 2001. Changes in maze behavior of mice occur after sufficient accu-mulation of docosahexaenoic acid in brain. J. Nutr. 131, 319–324.
Lim, S.N., Huang, W., Hall, J.C., Ward, R.E., Priestley, J.V., Michael-Titus, A.T., 2010. Theacute administration of eicosapentaenoic acid is neuroprotective after spinal cordcompression injury in rats. Prostaglandins Leukot. Essent. Fatty Acids 83, 193–201.
Lim, S.N., Gladman, S.J., Dyall, S.C., Platel, U., Virani, N., Kang, J.X., Priestley, J.V., Michael-Titus, A.T., 2012. Improved outcome after spinal cord injury in transgenic micewith high levels of endogenous omega-3 polyunsaturated fatty acids. ISSFAL Annu-al Meeting Abstract.
Liu, X.Z., Xu, X.M., Hu, R., Du, C., Zhang, S.X., McDonald, J.W., Dong, H.X., Wu, Y.J., Fan,G.S., Jacquin, M.F., Hsu, C.Y., Choi, D.W., 1997. Neuronal and glial apoptosis aftertraumatic spinal cord injury. J. Neurosci. 17, 5395–5406.
Liu, Q., Xie, F., Siedlak, S.L., Nunomura, A., Honda, K., Moreira, P.I., Zhua, X., Smith, M.A.,Perry, G., 2004. Neurofilament proteins in neurodegenerative diseases. Cell. Mol.Life Sci. 61, 3057–3075.
Lopez-Vales, R., Redensek, A., Skinner, T.A., Rathore, K.I., Ghasemlou, N., Wojewodka,G., DeSanctis, J., Radzioch, D., David, S., 2010. Fenretinide promotes functionalrecovery and tissue protection after spinal cord contusion injury in mice. J. Neurosci.30, 3220–3226.
Lytle, J.M., Wrathall, J.R., 2007. Glial cell loss, proliferation and replacement in thecontused murine spinal cord. Eur. J. Neurosci. 25, 1711–1724.
Ma, M., Basso, D.M., Walters, P., Stokes, B.T., Jakeman, L.B., 2001. Behavioral and histo-logical outcomes following graded spinal cord contusion injury in the C57Bl/6mouse. Exp. Neurol. 169, 239–254.
Ma, D., Tao, B., Warashina, S., Kotani, S., Lu, L., Kaplamadzhiev, D.B., Mori, Y., Tonchev,A.B., Yamashima, T., 2007. Expression of free fatty acid receptor GPR40 in the cen-tral nervous system of adult monkeys. Neurosci. Res. 58, 394–401.
Makwana, M., Dyall, S., Raivich, G., Michael-Titus, A.T., 2006. The effect ofdocosahexaenoic acid, an omega-3 polyunsaturated fatty acid, in a mouse facialnerve injury model. BPS 75th Anniversary Meeting, 181P.
Marques, S.A., Garcez, V.F., Del Bel, E.A., Martinez, A.M., 2009. A simple, inexpensiveand easily reproducible model of spinal cord injury in mice: morphological andfunctional assessment. J. Neurosci. Methods 177, 183–193.
Mayser, P., Mayer, K., Mahloudjian, M., Benzing, S., Kramer, H.J., Schill, W.B., Seeger, W.,Grimminger, F., 2002. A double-blind, randomized, placebo-controlled trial of n-3versus n-6 fatty acid-based lipid infusion in atopic dermatitis. JPEN J. Parenter. En-teral Nutr. 26, 151–158.
Menard, C., Patenaude, C., Gagne, A.M., Massicotte, G., 2009. AMPA receptor-mediatedcell death is reduced by docosahexaenoic acid but not by eicosapentaenoic acid inarea CA1 of hippocampal slice cultures. J. Neurosci. Res. 87, 876–886.
Mey, J., Morassutti, J., Brook, G., Liu, R.H., Zhang, Y.P., Koopmans, G., McCaffery, P., 2005.Retinoic acid synthesis by a population of NG2-positive cells in the injured spinalcord. Eur. J. Neurosci. 21, 1555–1568.
Michael-Titus, A.T., 2007. Omega-3 fatty acids and neurological injury. ProstaglandinsLeukot. Essent. Fatty Acids 77, 295–300.
Nishio, Y., Koda, M., Hashimoto, M., Kamada, T., Koshizuka, S., Yoshinaga, K., Onodera,S., Nishihira, J., Okawa, A., Yamazaki, M., 2009. Deletion of macrophage migrationinhibitory factor attenuates neuronal death and promotes functional recoveryafter compression-induced spinal cord injury in mice. Acta Neuropathol. 117,321–328.
Popovich, P.G., Jones, T.B., 2003. Manipulating neuroinflammatory reactions in the in-jured spinal cord: back to basics. Trends Pharmacol. Sci. 24, 13–17.
Popovich, P.G., Horner, P.J., Mullin, B.B., Stokes, B.T., 1996. A quantitative spatial analy-sis of the blood-spinal cord barrier. I. Permeability changes after experimental spi-nal contusion injury. Exp. Neurol. 142, 258–275.
Popovich, P.G., Wei, P., Stokes, B.T., 1997. Cellular inflammatory response afterspinal cord injury in Sprague–Dawley and Lewis rats. J. Comp. Neurol. 377,443–464.
Reagan-Shaw, S., Nihal, M., Ahmad, N., 2008. Dose translation from animal to humanstudies revisited. FASEB J. 22, 659–661.
Schumacher, P.A., Eubanks, J.H., Fehlings, M.G., 1999. Increased calpain I-mediated pro-teolysis, and preferential loss of dephosphorylated NF200, following traumatic spi-nal cord injury. Neuroscience 91, 733–744.
Serhan, C.N., 2004. A search for endogenous mechanisms of anti-inflammationuncovers novel chemical mediators: missing links to resolution. Histochem. CellBiol. 122, 305–321.
Sroga, J.M., Jones, T.B., Kigerl, K.A., McGaughy, V.M., Popovich, P.G., 2003. Rats and miceexhibit distinct inflammatory reactions after spinal cord injury. J. Comp. Neurol.462, 223–240.
Stillwell, W., Wassall, S.R., 2003. Docosahexaenoic acid: membrane properties of aunique fatty acid. Chem. Phys. Lipids 126, 1–27.
Tan, N.S., Shaw, N.S., Vinckenbosch, N., Liu, P., Yasmin, R., Desvergne, B., Wahli, W., Noy,N., 2002. Selective cooperation between fatty acid binding proteins and peroxi-some proliferator-activated receptors in regulating transcription. Mol. Cell. Biol.22, 5114–5127.
Tator, C.H., Fehlings, M.G., 1999. Review of clinical trials of neuroprotection in acutespinal cord injury. Neurosurg. Focus 6, e8.
Ward, R.E., Huang, W., Curran, O.E., Priestley, J.V., Michael-Titus, A.T., 2010.Docosahexaenoic acid prevents white matter damage after spinal cord injury.J. Neurotrauma 27, 1769–1780.
Wood, J.T., Williams, J.S., Pandarinathan, L., Janero, D.R., Lammi-Keefe, C.J., Makriyannis,A., 2010. Dietary docosahexaenoic acid supplementation alters select physiologicalendocannabinoid-system metabolites in brain and plasma. J. Lipid Res. 51,1416–1423.
Wu, X., Yoo, S., Wrathall, J.R., 2005. Real-time quantitative PCR analysis of temporal–spatial alterations in gene expression after spinal cord contusion. J. Neurochem.93, 943–952.
Wu, A., Ying, Z., Gomez-Pinilla, F., 2011. The salutary effects of DHA dietary supplemen-tation on cognition, neuroplasticity, and membrane homeostasis after brain trau-ma. J. Neurotrauma 28, 2113–2122.
Xu, J., Drew, P.D., 2006. 9-Cis-retinoic acid suppresses inflammatory responses ofmicroglia and astrocytes. J. Neuroimmunol. 171, 135–144.
Xu, L.Z., Sanchez, R., Sali, A., Heintz, N., 1996. Ligand specificity of brain lipid-bindingprotein. J. Biol. Chem. 271, 24711–24719.
Xu, W., Chi, L., Xu, R., Ke, Y., Luo, C., Cai, J., Qiu, M., Gozal, D., Liu, R., 2005. Increasedproduction of reactive oxygen species contributes to motor neuron death in a com-pression mouse model of spinal cord injury. Spinal Cord 43, 204–213.
Yamashima, T., 2008. A putative link of PUFA, GPR40 and adult-born hippocampalneurons for memory. Prog. Neurobiol. 84, 105–115.
Zbarski-Barquero, V., Sijben, J., De Wilde, M., Priestley, J.V., Michael-Titus, A.T., 2012. Amulti-nutrient composition with neuroprotective properties in spinal cord injury.ISSFAL Annual Meeting Abstract.




















