
Sex-based differences in veterans with pulmonary hypertension
Upload
independentCategory
view
3download
0
Effect of 5-Lipoyxgenase on the Development of Pulmonary Hypertension in Rats
John E. Jones1, Jennifer L. Walker2, Yanli Song2, Norbert Weiss2#, Wellington V. Cardoso3, Rubin M. Tuder4, Joseph Loscalzo2, and Ying-Yi Zhang2*
1Department of Surgery, 2Whitaker Cardiovascular Institute and Evans Department of Medicine, and 3The Pulmonary Center, Boston University School of Medicine, Boston, MA 02118, 4Division of Cardiopulmonary Pathology and Pulmonary and Critical Care Medicine, Department of Pathology and Medicine, The Johns Hopkins University School of Medicine #Current address: Klinikum der Universität Munchen, Medizinische Poliklinik-Innenstadt, D-80336 Münnich, Germany
Running title: 5-Lipoxygenase and pulmonary hypertension
Abbreviations: 5LO-5-lipoxygenase; MCT-monocrotaline; Ad5LO-adenovirous expressing 5LO;
AdGFP-adenovirus expressing green fluorescence protein; PAAT-pulmonary artery acceleration
time; RVSP-right ventricular systolic pressure; PBS-Dulbecco’s phosphate-buffered saline.
Corresponding author:
Ying-Yi Zhang, Ph.D. Boston University School of Medicine Whitaker Cardiovascular Institute 715 Albany St., W-507 Boston, MA 02118 Telephone: (617) 638-4896 Fax: (617) 638-4066 e-mail: [email protected]
Articles in PresS. Am J Physiol Heart Circ Physiol (January 15, 2004). 10.1152/ajpheart.00281.2003
Copyright (c) 2004 by the American Physiological Society.

2
Abstract
5-Lipoxygenase (5LO) and its downstream leukotriene products have been implicated in the
development of pulmonary hypertension. In this study we examined the effects of 5LO
overexpression in rat lungs on pulmonary hypertension, using a recombinant adenovirus
expressing 5LO (Ad5LO). Transthoracic echocardiography and right heart catheterization data
showed that 5LO overexpression in lung did not cause pulmonary hypertension in normal rats;
however, it markedly accelerated the progression of pulmonary hypertension in rats treated with
monocrotaline (MCT). An increase in pulmonary artery pressure occurred earlier in the rats
treated with MCT+Ad5LO (7-10 days) as compared to those treated with control vector,
MCT+AdGFP, or MCT alone (15-18 days). The weight ratio of right ventricle-to-left ventricle plus
septum was higher in the MCT+Ad5LO group than that of MCT+AdGFP or MCT alone group
(0.45±0.08 vs. 0.35±0.03 or 0.33±0.06). Lung tissue histological sections from MCT+Ad5LO
rats exhibited more severe inflammatory cell infiltration and pulmonary vascular muscularization
than those from MCT+AdGFP- or MCT-treated rats. Administration of 5LO inhibitors, zileuton or
MK-886, to either MCT- or MCT+Ad5LO-treated rats prevented the development of pulmonary
hypertension. These data suggest that 5LO plays a critical role in the progression of pulmonary
hypertension in rats, and that the detrimental effect of 5LO is manifest only in the setting of
pulmonary vascular endothelial cell dysfunction.
Key words: 5-lipoxygenase, inflammation, pulmonary hypertension, monocrotaline.

3
Introduction
5-Lipoxygenase (5LO) catalyzes two consecutive reactions that convert arachidonic acid
to leukotriene A4 (6, 32). The latter is a crucial precursor for the formation of leukotriene B4, a
potent chemotactic agent (14), and for the formation of leukotrienes C4, D4, and E4 (collectively
referred to as cysteinyl leukotrienes) (26), substances that cause an increase in vascular
permeability (9, 13) and constriction of bronchi (10, 17) and certain types of blood vessels (4, 5).
The substrate for 5LO is generated by phospholipase A2 (11), and is presented to 5LO by 5LO-
activating protein (FLAP) (12).
5-Lipoxygenase has also been found to participate in other cellular processes, including
cell proliferation (3, 39), tyrosine kinase signaling (21), and F-actin polymerization (21, 27, 29).
In addition, 5LO was found to bind to a TGF-β receptor I-associated protein (TRAP-1) (30),
although the effect of this association is currently unclear.
Increased 5LO expression has been found in the lung tissue of patients with primary
pulmonary hypertension, within infiltrating perivascular alveolar macrophages and in small
pulmonary artery endothelial cells (41). Several studies also suggested that 5LO may play a
role in the development of pulmonary hypertension. For example, cysteinyl leukotrienes were
found in the lung lavage fluid of neonates with persistent pulmonary hypertension (33), patients
with chronic obstructive pulmonary disease (28), and animals subjected to acute hypoxia (25).
Targeted disruption of the 5LO gene (38), or treatment of animals with diethylcarbamazine (24),
MK886 (an inhibitor of FLAP) (38), or leukotriene antagonists (1, 16, 23, 31), reduced hypoxia-
induced pulmonary hypertension. Diethylcarbamazine, an anti-parasitic drug that also inhibits
5LO, was shown to ameliorate the development of pulmonary hypertension caused by
monocrotaline in rats (34).

4
In the present study, we explored two questions: 1) does 5LO overexpression cause
pulmonary hypertension? and 2) does 5LO facilitate the development of pulmonary
hypertension caused by other factors? To address the first question, we expressed an
exogenous 5LO (human 5LO) in rat lungs by adenovirus-mediated gene transfer, and monitored
the development of pulmonary hypertension in the animal. Human 5LO has about 93% protein
sequence identity to rat 5LO (GenBank accession # J03600 and J03960, respectively), and can
be expressed well during transfection in various mammalian cell lines, such as COS-1 and
NIH3T3 cells. We have previously expressed human 5LO in porcine pulmonary artery
endothelial cells and shown the recombinant enzyme was able to utilize endogenous
arachidonic acid to generate leukotrienes (43).
To address the second question, we used monocrotaline-treated rats. Monocrotaline
(MCT) is a plant alkaloid that is dehydrogenated in rat liver to form a highly reactive pyrrole
derivative (8). The latter interacts with the first vascular bed encountered, causing pulmonary
endothelial cell injury and lung inflammation (2, 8, 40). These events lead to progressive
pulmonary vascular remodeling. Elevation of pulmonary arterial pressure occurs in rats in ~2.5
weeks after MCT injection (7, 15, 22). In this study, we examined whether 5LO accelerates the
development of pulmonary hypertension and whether specific 5LO inhibitors prevent the
disease process.

5
Methods
Animals
Male Sprague-Dawley rats weighing 225-250 grams were obtained from Hilltop Lab
Animals, Inc. (Scottdale, PA). Rats were acclimated for four days before initial treatment.
Drug preparation and treatment
Monocrotaline (Sigma) was prepared at the concentration of 60 mg/ml by dissolving in
HCl-acidified PBS and adjusting the pH to 7.2 with NaOH. The compound was administered to
rats at 60 mg/kg via intraperitoneal injection. MK886 (CalBiochem) was dissolved in
dimethylsulfoxide (DMSO), and administered to rats at 20 mg/kg via subcutaneous injection.
Zileuton (Abbott) was prepared by grinding the tablet into powder and suspending it in water at
50 mg/ml. The drug was administered to rats by oral gavage at 100 mg/kg/day.
Delivery of adenovirus
Replication-deficient adenovirus expressing human 5LO (Ad5LO) was prepared as
previously described (43). Adenovirus expressing green fluorescence protein (AdGFP) was a
kind gift from Dr. John Gray at the Harvard Gene Therapy Initiative. To administer the
recombinant adenovirus to rat lungs, rats were anesthetized with ketamine (100 mg/kg) and
xylazine (7.5 mg/kg). Respiratory tubing was fitted to the snout of the rat, and connected to the
aerosol chamber of a MISTY nebulizer (Allegiance Health Co., McGaw Park, IL) containing 2 x
109 plaque forming units of Ad5LO or AdGFP in 1 ml saline. The chamber was also connected
to a room air source at a flow rate of 8 L/min, causing aerosolization of the virus, and to a tank

6
of 21% oxygen with balanced nitrogen at a flow rate of 1.5 L/min to deliver the adenovirus. The
aerosolized delivery lasted 20 minutes for each rat.
RNA isolation and Northern blotting
Rat lungs were gently perfused with 40 ml of saline through the pulmonary artery, and
~300 mg of lung tissue was homogenized immediately in 10 ml TRIzol Reagent (Invitrogen)
followed by chloroform extraction for total RNA. Northern blotting was carried out by separating
20 µg RNA in 1.2 % agarose gels containing 4% formaldehyde. The RNA was transferred onto
nitrocellulose membranes and hybridized with an [α-32P]-dCTP-labeled 5LO probe that
encompasses nucleotides 45-290 of human 5LO cDNA (GenBank accession #J03600).
Hybridization was carried out at 68°C overnight in MiracleHyb solution (Stratagene) containing
10 µg/ml sheared salmon sperm DNA. The membrane was washed with 2 X SSC/0.1% SDS at
room temperature for 15 min and then in 0.1 X SSC/0.1% SDS at 68°C for 20 min before
exposure to X-ray film. The membrane was then washed in 0.01 X SSC/0.1% SDS at 100°C
twice for 15 min to strip the bound 5LO probe, and hybridized with a [α-32P]-dCTP-labeled
mouse GAPDH probe (Ambion), which crossreacts with rat GAPDH, to estimate total RNA
loading.
Echocardiography
Transthoracic echocardiography was performed on rats according to a previously
described method (18). Rats were anesthetized with a ketamine/xylazine mixture as described
above, and their chests shaved prior to examination. Two-dimensional and Doppler imaging
was performed using an Acuson C256 ultrasonographic system. Pulmonary flow acceleration

7
time (PAAT), pulmonary flow waveform, right ventricular free wall thickness, and tricuspid
regurgitation were examined on all rats on days 3, 7, 10, 15, 18, and 24.
Hemodynamic measurements
Rats underwent right heart catheterization at various time points during the study. Prior
to catheterization, rats were anesthetized with the same ketamine/xylazine mixture as above.
After dissection to expose the right jugular vein, a 1.4F Millar Mikro-Tip pressure catheter (Millar
Instruments) was inserted into the vein and advanced to the right ventricle. The catheter was
connected to a transducer unit (Millar Instruments) and interfaced with a signal amplifier and
recorder (Gould Instrument Systems), and the right ventricular systolic pressure (RVSP) was
recorded. On day 24, the systemic blood pressure of rats from each group was also measured.
The left carotid artery was exposed and cannulated with a 1.4F Millar pressure catheter.
Histology
Rat lungs were inflated with 10% phosphate buffered formalin at a pressure of 20 cm
H2O and fixed for 20 hr at 4°C. The lung tissue was processed and paraffin-embedded using
Hypercenter XP System and Embedding Center (Shandon Inc, Pittsburgh, PA) and cut into 5-
µm sections. The tissue sections were then heat-dried on slides at 56ºC for 1 hr before being
deparaffinized and rehydrated.
For hematoxylin and eosin staining, tissue sections were incubated in Gill-2 hematoxylin
(Thermo Shandon) for 2 min, rinsed with water for 2 min, dipped once in acid alcohol (70%
ethanol and 1% concentrated HCl), rinsed with water for 1 min, dipped in 1% NH4OH for 15 sec,
rinsed with water for 1 min, and dipped twice in 1% Eosin Y (Fisher). The stained sections were

8
dehydrated by incubation in 95% ethanol for 2 x 30 sec, 100% ethanol for 2 x 30 sec, and
xylene for 2 x 2.5 min, and mounted with Cytoseal60 (Richard-Allan Scientific).
For trichrome staining, paraffin-embedded tissue sections were deparaffinized,
rehydrated, and stained with a Masson’s Trichrome staining kit (Poly Scientific R&D Corp)
according to the manufacturer’s instruction. For immunohistochemistry, paraffin embedded lung
tissues were cut into 5-µm sections, heat-dried onto slides, deparaffinized, and rehydrated as
described above. Antigen retrieval was carried out in 10 mM citric acid, pH 6, by heating for 4 x
2 min at 400 watts in a microwave oven and cooling at room temperature for 20 min. The
sections were washed with deionized water twice for 5 min and with PBS for 5 min before
blocking with 10% goat serum in PBS at room temperature for 30 min.
For α-actin staining, the sections were incubated with a mouse monoclonal antibody
against smooth muscle cell α-actin (Sigma, 1:800 dilution in 1% immunohistochemical grade
BSA/PBS) at 4°C overnight. The sections were washed in PBS twice for 5 min and incubated
with the secondary antibody, biotinylated goat anti-mouse IgG (Jackson Immunoresearch), at a
dilution of 1:500 in 1% BSA/PBS for 30 min at room temperature. The sections were washed
with PBS twice for 5 min and incubated with complexes of avidin DH and biotinylated-alkaline
phosphatase H (provided in Vectastain ABC-AP kit, Vector Lab, Burlingame, CA) for 30 min.
The sections were washed with PBS twice for 5 min and incubated with alkaline phosphatase
substrate (provided in Vector Red Substrate kit, Vector Lab) for 20 min. The slides were
washed under running tap water for 5 min before being counterstained with hematoxylin, which
was carried out by dipping the slides in Harris Modified Hematoxylin for 10 sec, washing with
water, dipping in acid alcohol, and rinsing with water.
For monocyte/macrophage staining, the sections were incubated with a mouse
monoclonal antibody against calprotectin (Clone MCA387, Chemicon, Temecula, CA, 1:50

9
dilution in 1% immunohistochemical grade BSA/PBS) at 4ºC overnight. Calprotectin is
expressed in monocytes, macrophages, and granulocytes. The sections were washed and
stained with the secondary antibody as described above. The sections were developed by
washing with PBS twice for 5 min, incubated with avidin and biotinylated horseradish peroxidase
complex (Vectastatin ABC kit) for 30 min, washed with PBS twice again for 5 min, and incubated
with peroxidase substrate (DAB substrate kit, Vector Lab) for 7 min. The sections were washed
with tap water and counterstained with methyl green for 4 min at 60ºC, followed by rinsing in
water for 1 min and in acetone containing 0.05% acetic acid for 6 min. The stained sections
were dehydrated, and the slides were mounted as described above.
For 5LO staining, the sections were incubated with rabbit anti-human 5LO antiserum
(1551-4b) at 1:1600 dilution or pre-bleed serum at 1:1600 dilution (43) at 4°C overnight and
developed with the same procedure as described above for α-actin staining.
Statistical analysis
Data are presented as the mean ± standard deviation of the mean. Statistical analysis
was performed by two-way ANOVA, with the Tukey test used for post-hoc analysis. P<0.05
indicates statistical significance.

10
Results
Overexpression of 5LO in rat lungs
To examine the role of 5LO in the development of pulmonary hypertension, recombinant
human 5LO was expressed in rat lungs via adenovirus-mediated gene transfer. Approximately
2 x 109 plaque forming units of adenovirus expressing human 5LO (Ad5LO) were aerosolized
and delivered to the rat lung through inhalation. The time course of the transgene expression in
the lungs was analyzed by Northern blotting. Three groups of rats were examined: rats given
vehicle - Dulbecco’s phosphate-buffered saline (PBS), MCT, or MCT+Ad5LO. As shown in
Figure 1, rats in the last group expressed human 5LO mRNA; the expression peaked between
7-10 days and decreased by day 15 after Ad5LO delivery.
The localization of the expressed 5LO protein in rat lungs was examined by
immunohistochemical staining. As shown in Figure 2, the expression occurred in a wide range
of cell types, including alveolar pneumocytes, bronchial epithelial cells, alveolar macrophages,
and small pulmonary vessels. The anti-human 5LO antiserum used for the study crossreacts
with rat 5LO, which is normally expressed in leukocytes and macrophages.
.
Effect of 5LO overexpression on the development of pulmonary hypertension
To determine the effect of 5LO on the development of pulmonary hypertension in rats,
five groups of rats were studied: rats given PBS, Ad5LO, MCT, MCT+AdGFP, and MCT+Ad5LO.
All the treatments were performed on day 0.
On days 3, 7, 10, 15, 18, and 24, rats were examined by echocardiography to assess
pulmonary artery acceleration time (PAAT), pulmonary flow waveform, and right ventricular free
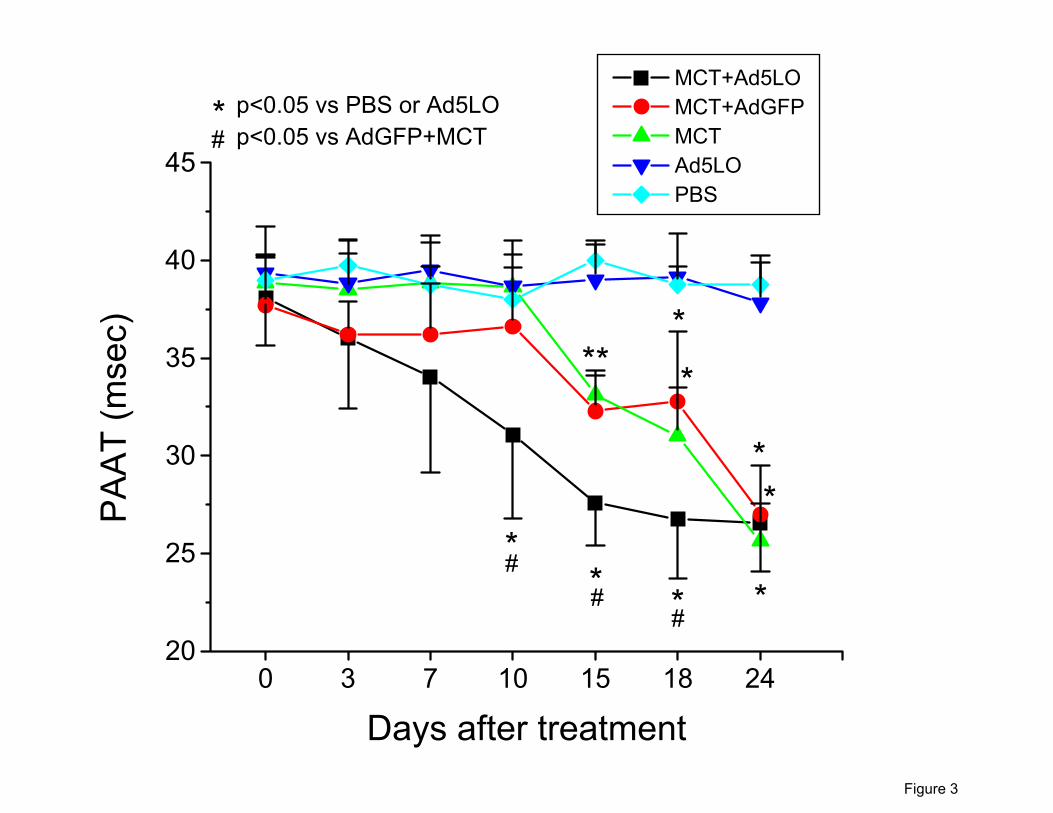
wall thickness. As shown in Figure 3, the decrease in PAAT occurred significantly earlier in the

11
rats treated with MCT+Ad5LO than those treated with MCT alone (7-10 days vs. 15-18 days).
Administration of MCT+AdGFP had the same effect as that of MCT alone. Rats that received
Ad5LO alone did not have decreased PAAT over the entire study period, which was similar to
that observed in rats given a PBS injection. Pulmonary flow waveform change (development of
a midsystolic notch) occurred also earlier in the rats treated with MCT+Ad5LO than those
treated with MCT only or MCT+AdGFP (data not shown).
Right ventricular systolic pressures, measured by right heart catheterization, are shown in
Figure 4 for all groups of rats on days 0, 10, 15, 18, and 24. Consistent with the
echocardiographic findings, rats treated with MCT+Ad5LO had a significantly higher right
ventricular systolic pressure on day 10 compared to that of rats treated with MCT. Rats treated
with MCT+AdGFP behaved similarly to rats treated with MCT alone. Administration of Ad5LO
alone or PBS did not cause increases in right ventricular systolic pressure. By day 24, no
significant difference in right ventricular systolic pressure was found among the MCT+Ad5LO,
MCT+AdGFP, and MCT alone treatment groups.
Changes in right ventricle to left ventricle plus septum weight ratio
Rats in the above treatment groups were sacrificed at day 26. Their systolic pressures
and body weights were measured, and hearts dissected. The weight ratio of right ventricle-to-left
ventricle plus septum (RV/LV+S) was calculated. As shown in Figure 5, the ratio was
significantly higher in the group treated with MCT+Ad5LO than in the groups treated with
MCT+AdGFP or MCT alone (0.45±0.08 vs. 0.35±0.03 or 0.33±0.06, P<0.02). In addition, the
ratios in the groups treated with MCT were significantly higher than those without MCT. As the
MCT+Ad5LO-treated group were exposed to a significantly longer period of increased
pulmonary artery pressure, it may explain the higher RV/LV+S found in this group, even though

12
its right ventricular systolic pressure was no longer higher than that of the MCT and
MCT+AdGFP treatment groups by day 24. There was no significant difference in systolic blood
pressure or heart rate among the 5 groups at day 26 (data not shown).
Histological changes
Lungs from the treated rats were harvested at various time points and were fixed in
formalin. After paraffin embedding and sectioning, the tissues were stained with hematoxylin
and eosin. Three major histological changes were found in the MCT-treated groups:
inflammatory cell infiltration, muscularization of the distal pulmonary vessels, and fibrosis of the
interstitium. These changes in the MCT+Ad5LO group were more severe than those in the
MCT+AdGFP or MCT alone groups. The sections also show scattered post-inflammatory
extension of bronchiolar epithelium to thickened septa, which appeared to be similar in all MCT-
treated groups (data not shown). To compare the degree of inflammation and muscularization of
distal vessels, lung tissue sections were stained with specific antibodies against
monocytes/macrophages or smooth muscle cell α-actin, and the numbers of the antibody-
stained cells or vessels were counted. In addition, Masson’s trichrome staining was performed
on tissue sections collected at day 26 after treatment to monitor the fibrosis in these sections.
Figure 6A shows the photomicrographs of hematoxylin and eosin-stained lung sections
prepared from rats treated with PBS, MCT, MCT+AdGFP, and MCT+Ad5LO at day 15. The
staining revealed the presence of inflammatory cells, predominantly monocytes/macrophages
(include enlarged foam cells) and lymphocytes. The distribution of the infiltrated inflammatory
cells in the lung is patchy, many being in the proximity of blood vessels. To estimate the degree
of inflammation, lung sections from MCT, MCT+AdGFP, and MCT+Ad5LO treatment groups
collected at days 7, 15, and 22 were stained with anti-monocyte/macrophage antibody and the

13
number of stained cells was counted. As shown in Figure 6B, the number of macrophages in
the MCT+AdGFP and MCT+Ad5LO groups was the highest at day 15 and was reduced by day
22. The number of macrophages in the MCT group was increased by day 22. The
MCT+Ad5LO group had significantly more macrophage infiltration than the MCT+AdGFP group
at day 15. The MCT only group had noticeably less macrophage infiltration than the
MCT+Ad5LO and MCT+AdGFP groups, indicating that part of the inflammation was caused by
adenovirus infection.
Shown in Figure 7A are photomicrographs of an anti-smooth muscle cell α-actin stained
lung sections prepared from rats treated with PBS, MCT+AdGFP, and MCT+Ad5LO at day 22.
The muscularization and thickening of distal pulmonary vessels is more visible in the
MCT+Ad5LO group than the MCT+AdGFP group. The numbers of the muscularized distal
vessels were counted in groups treated with MCT+Ad5LO and MCT+AdGFP, at days 7, 15, 22,
and 26. As shown in Figure 7B, the number of muscularized distal pulmonary vessels
increased significantly at 15 day and reached a plateau by day 22. The MCT+Ad5LO treatment
group displayed a trend toward higher numbers of muscularized vessels than those of the
MCT+AdGFP treatment group on days 22 and 26, but the difference did not reach statistical
significance.
Figure 8 shows representative photomicrographs of lung sections stained with Masson’s
trichrome stain from rats treated with PBS, MCT+AdGFP, or MCT+Ad5LO at day 26. As
demonstrated, the lung sections had extensive collagen deposition, consistent with fibrosis. The
MCT+Ad5LO treatment group appeared to have more alveolar thickening and collagen
deposition than the MCT+AdGFP treatment group.

14
Effect of 5LO inhibition on the development of pulmonary hypertension
To examine whether these effects of 5LO is dependent on its activity, two specific
inhibitors, one for 5LO (zileuton) and one for FLAP (MK886), were utilized. Rats were treated
with MCT+Ad5LO on day 0, and DMSO (solvent for MK886), MK886, or zileuton was
administered on day 1 and daily thereafter. The one-day delay of the administration of inhibitors
was designed to avoid potential interference of the activation of MCT in the liver. Right heart
catheterization was performed on all groups at days 10, 15, and 26. As shown in Figure 9A,
both inhibitors prevented the increase of right ventricular pressure in the MCT+Ad5LO-treated
rats at day 10 (15±2 mmHg and 16±4 mmHg for MCT+Ad5LO+ MK886 and
MCT+Ad5LO+zileuton, respectively, vs. 25±2 mmHg for MCT+Ad5LO; p<0.05). Interestingly,
MK886 and zileuton also prevented the late development of pulmonary hypertension in the rats
(day 26: MCT+Ad5LO = 42±4 mmHg vs. 25±5 mmHg and 22±8 mmHg for
MCT+Ad5LO+MK886 and MCT+Ad5LO+zileuton rats, respectively; p<0.05 vs. MCT+Ad5LO).
To examine the possibility that 5LO plays a role in MCT-induced pulmonary hypertension
in rats, rats were treated with MCT only and received daily administration of vehicle (DMSO),
MK886, or zileuton as described above. Right heart catheterization was performed on all
groups at day 35. As shown in Figure 9B, both MK886 and zileuton prevented the development
of pulmonary hypertension in the MCT-treated rats, which suggests that endogenous 5LO
played an important role in mediating MCT-induced pulmonary hypertension in rats.

15
Discussion
This study examined the role of 5LO in the development of pulmonary hypertension by
overexpressing the enzyme in rat lungs through adenovirus-mediated gene transfer, and by
administrering specific 5LO inhibitors to MCT-treated rats. The results show that
overexpression of 5LO in normal lungs did not cause pulmonary hypertension; however, it
markedly accelerated the development of pulmonary hypertension in rats treated with MCT. 5LO
inhibitors inhibited the effect of 5LO, and also prevented MCT-induced pulmonary hypertension.
Two questions arise from the results of this study: 1) why did the overexpression of 5LO
have an adverse effect on the pulmonary vasculature in MCT-treated rats but not in normal rats?
and 2) why did the 5LO inhibitors prevent the pulmonary hypertension caused by MCT?
The first question can be explained by two possibilities. One is that the normal
pulmonary vasculature is somewhat protected from leukotriene-dependent vasoconstriction, but
in the MCT-injured pulmonary vessels, this protection is weakened. The source of the
protection has been implicated by Bäck and colleagues who reported that cysteinyl leukotrienes
stimulate prostacyclin production in the isolated human pulmonary artery, and that endothelium-
denuded or indomethacin-treated human pulmonary artery is significantly more sensitive to
cysteinyl leukotriene-induced vasoconstriction (in an organ chamber) than endothelialized
pulmonary arteries (4). The second possibility is that MCT-treated rats had more pulmonary
inflammation than normal rats, and the inflammation provided a supportive environment for 5LO-
catalyzed leukotriene biosynthesis. Leukotriene biosynthesis consists of three major steps:
release of arachidonic acid from cell membranes by phospholipase A2, conversion of
arachidonic acid to leukotriene A4 by 5LO, and transformation of leukotriene A4 to leukotrienes
B4 or cysteinyl leukotrienes. Both phospholipase A2 and 5LO require calcium for their activity,
and intracellular calcium release is related to the state of cell activation. Inflammatory stimuli

16
can cause activation of a variety of cells. Further analysis of prostacyclin and leukotriene
release in normal and dysfunctional pulmonary vasculature in animals expressing 5LO in the
lung would help to confirm these hypotheses.
The finding that 5LO inhibitors prevented the MCT-induced pulmonary hypertension
suggests endogenous 5LO is essential for MCT-induced pulmonary hypertension. MCT-
induced pulmonary endothelial injury is known to be followed by significant pulmonary vascular
and alveolar inflammation (34, 40), and the latter is thought to be important in subsequent
pulmonary vascular remodeling and pulmonary hypertension. Previous studies have shown that
inhibition of inflammation by interleukin-1 receptor antagonists (37) or by an antibody that
neutralizes monocyte chemotactic and activating factor/monocyte chemoattractant protein-1
(19) alleviate MCT-induced pulmonary hypertension. As 5LO is predominantly expressed in
inflammatory cells, infiltration of these cells in the lung results in an increase in 5LO in the
tissue. Downstream leukotriene products of 5LO could markedly amplify the inflammatory
process by attracting more inflammatory cells to the lung (leukotriene B4 ) and by increasing
vascular permeability to facilitate cell infiltration. It is possible that 5LO inhibitors prevented the
MCT-induced pulmonary hypertension via a similar mechanism as that by other inflammatory
inhibitors.
In addition to the facilitation of inflammatory processes, 5LO could exert its effect through
several other possible mechanisms. These include: 1) causing pulmonary vasoconstriction by
increasing cysteinyl leukotriene production; 2) promoting vascular cell proliferation by its nuclear
localization or through interaction with cytoskeletal proteins (21, 27, 29); and 3) altering TGF-β
superfamily signaling through binding to a TGF-β receptor I-associated protein (TRAP-1). To
address these mechanisms, further studies need to explore 1) the types of cysteinyl leukotriene
receptors on pulmonary arteries or arterioles and the potential influence of endothelial

17
dysfunction on the function of these receptors; 2) the role of the cytoskeleton, actin, or tyrosine
kinase signaling in vascular remodeling during the development of pulmonary arterial
hypertension; and 3) the effect of the 5LO-TRAP-1 interaction on TGF-β signaling. This latter
interaction may be of particular relevance to familial and sporadic forms of primary pulmonary
hypertension in which imbalanced TGF-β superfamily signaling is believed to play a role in the
disease development (20, 35, 36, 42). Since 5LO overexpression has been demonstrated in
primary pulmonary hypertension, examining these possibilities in the future would help clarify
the specific role of 5LO in pulmonary hypertension, as well as the mechanism of the
development of pulmonary hypertension, in general.

18
Acknowledgements:
The authors thank Stephanie Tribuna for expert secretarial assistance, Antoinette Hayes
for invaluable contribution with animal work, and Ray Qian for excellent assistance in
morphometric analysis of histology slides.
This work is supported by NIH Grants P01 HL66254 project III (RMT), P50 HL55993,
HL58976, HL61795 (JL), and AHA National Grant-in-Aid 0256282N (YYZ).

19
References
1. Ahmed, T., and W. Oliver, Jr. Does slow-reacting substance of anaphylaxis mediate
hypoxic pulmonary vasoconstriction? Am Rev Respir Dis 127: 566-71., 1983.
2. Allen, J. R., and L. A. Carstens. Pulmonary vascular occlusions initiated by endothelial
lysis in monocrotaline-intoxicated rats. Exp Mol Pathol 13: 159-71., 1970.
3. Avis, I. M., M. Jett, T. Boyle, M. D. Vos, T. Moody, A. M. Treston, A. Martinez, and J. L.
Mulshine. Growth control of lung cancer by interruption of 5-lipoxygenase- mediated growth
factor signaling. J Clin Invest 97: 806-13, 1996.
4. Back, M., X. Norel, L. Walch, J. Gascard, V. de Montpreville, S. Dahlen, and C. Brink.
Prostacyclin modulation of contractions of the human pulmonary artery by cysteinyl-
leukotrienes. Eur J Pharmacol 401: 389-95., 2000.
5. Berkowitz, B. A., B. Zabko-Potapovich, R. Valocik, and J. G. Gleason. Effects of the
leukotrienes on the vasculature and blood pressure of different species. J Pharmacol Exp Ther
229: 105-12., 1984.
6. Borgeat, P., and B. Samuelsson. Arachidonic acid metabolism in polymorphonuclear
leukocytes: unstable intermediate in formation of dihydroxy acids. Proceedings of the National
Academy of Sciences of the United States of America 76: 3213-7, 1979.
7. Bruner, L. H., K. S. Hilliker, and R. A. Roth. Pulmonary hypertension and ECG changes
from monocrotaline pyrrole in the rat. Am J Physiol 245: H300-6., 1983.
8. Butler, W. H., A. R. Mattocks, and J. M. Barnes. Lesions in the liver and lungs of rats
given pyrrole derivatives of pyrrolizidine alkaloids. J Pathol 100: 169-75., 1970.
9. Dahlen, S. E., J. Bjork, P. Hedqvist, K. E. Arfors, S. Hammarstrom, J. A. Lindgren, and B.
Samuelsson. Leukotrienes promote plasma leakage and leukocyte adhesion in postcapillary

20
venules: in vivo effects with relevance to the acute inflammatory response. Proc Natl Acad Sci U
S A 78: 3887-91., 1981.
10. Dahlen, S. E., P. Hedqvist, S. Hammarstrom, and B. Samuelsson. Leukotrienes are
potent constrictors of human bronchi. Nature 288: 484-6., 1980.
11. Dennis, E. A. Phospholipase A2 in eicosanoid generation. American Journal of
Respiratory & Critical Care Medicine 161: S32-5, 2000.
12. Dixon, R. A., R. E. Diehl, E. Opas, E. Rands, P. J. Vickers, J. F. Evans, J. W. Gillard, and
D. K. Miller. Requirement of a 5-lipoxygenase-activating protein for leukotriene synthesis.
Nature 343: 282-4, 1990.
13. Drazen, J. M., K. F. Austen, R. A. Lewis, D. A. Clark, G. Goto, A. Marfat, and E. J. Corey.
Comparative airway and vascular activities of leukotrienes C-1 and D in vivo and in vitro. Proc
Natl Acad Sci U S A 77: 4354-8., 1980.
14. Ford-Hutchinson, A. W., M. A. Bray, M. V. Doig, M. E. Shipley, and M. J. Smith.
Leukotriene B, a potent chemokinetic and aggregating substance released from
polymorphonuclear leukocytes. Nature 286: 264-5, 1980.
15. Ghodsi, F., and J. A. Will. Changes in pulmonary structure and function induced by
monocrotaline intoxication. Am J Physiol 240: H149-55., 1981.
16. Goldberg, R. N., C. Suguihara, T. Ahmed, B. D. de Cudemus, P. Barrios, E. S. Setzer,
and E. Bancalari. Influence of an antagonist of slow-reacting substance of anaphylaxis on the
cardiovascular manifestations of hypoxia in piglets. Pediatr Res 19: 1201-5., 1985.
17. Hanna, C. J., M. K. Bach, P. D. Pare, and R. R. Schellenberg. Slow-reacting substances
(leukotrienes) contract human airway and pulmonary vascular smooth muscle in vitro. Nature
290: 343-4., 1981.

21
18. Jones, J. E., L. Mendes, M. A. Rudd, G. Russo, J. Loscalzo, and Y. Y. Zhang. Serial
noninvasive assessment of progressive pulmonary hypertension in a rat model. Am J Physiol
Heart Circ Physiol 283: H364-71., 2002.
19. Kimura, H., Y. Kasahara, K. Kurosu, K. Sugito, Y. Takiguchi, M. Terai, A. Mikata, M.
Natsume, N. Mukaida, K. Matsushima, and T. Kuriyama. Alleviation of monocrotaline-induced
pulmonary hypertension by antibodies to monocyte chemotactic and activating factor/monocyte
chemoattractant protein-1. Lab Invest 78: 571-81., 1998.
20. Lane, K. B., R. D. Machado, M. W. Pauciulo, J. R. Thomson, J. A. Phillips, 3rd, J. E.
Loyd, W. C. Nichols, and R. C. Trembath. Heterozygous germline mutations in BMPR2,
encoding a TGF-beta receptor, cause familial primary pulmonary hypertension. The
International PPH Consortium. Nat Genet 26: 81-4., 2000.
21. Lepley, R. A., and F. A. Fitzpatrick. 5-Lipoxygenase contains a functional Src homology
3-binding motif that interacts with the Src homology 3 domain of Grb2 and cytoskeletal proteins.
J Biol Chem 269: 24163-8, 1994.
22. Meyrick, B., and L. Reid. Development of pulmonary arterial changes in rats fed
Crotalaria spectabilis. Am J Pathol 94: 37-51., 1979.
23. Morganroth, M. L., J. T. Reeves, R. C. Murphy, and N. F. Voelkel. Leukotriene synthesis
and receptor blockers block hypoxic pulmonary vasoconstriction. J Appl Physiol 56: 1340-6.,
1984.
24. Morganroth, M. L., K. R. Stenmark, K. G. Morris, R. C. Murphy, M. Mathias, J. T. Reeves,
and N. F. Voelkel. Diethylcarbamazine inhibits acute and chronic hypoxic pulmonary
hypertension in awake rats. Am Rev Respir Dis 131: 488-92., 1985.

22
25. Morganroth, M. L., K. R. Stenmark, J. A. Zirrolli, R. Mauldin, M. Mathias, J. T. Reeves, R.
C. Murphy, and N. F. Voelkel. Leukotriene C4 production during hypoxic pulmonary
vasoconstriction in isolated rat lungs. Prostaglandins 28: 867-75., 1984.
26. Murphy, R. C., S. Hammarstrom, and B. Samuelsson. Leukotriene C: a slow-reacting
substance from murine mastocytoma cells. Proceedings of the National Academy of Sciences of
the United States of America 76: 4275-9, 1979.
27. Peppelenbosch, M. P., L. G. Tertoolen, W. J. Hage, and S. W. de Laat. Epidermal growth
factor-induced actin remodeling is regulated by 5- lipoxygenase and cyclooxygenase products.
Cell 74: 565-75., 1993.
28. Piperno, D., Y. Pacheco, R. Hosni, P. Moliere, C. Gharib, M. Lagarde, and M. Perrin-
Fayolle. Increased plasma levels of atrial natriuretic factor, renin activity, and leukotriene C4 in
chronic obstructive pulmonary disease. Chest 104: 454-9., 1993.
29. Provost, P., J. Doucet, T. Hammarberg, G. Gerisch, B. Samuelsson, and O. Radmark. 5-
Lipoxygenase interacts with coactosin-like protein. J Biol Chem 276: 16520-7., 2001.
30. Provost, P., B. Samuelsson, and O. Radmark. Interaction of 5-lipoxygenase with cellular
proteins. Proc Natl Acad Sci U S A 96: 1881-5., 1999.
31. Schreiber, M. D., M. A. Heymann, and S. J. Soifer. Leukotriene inhibition prevents and
reverses hypoxic pulmonary vasoconstriction in newborn lambs. Pediatr Res 19: 437-41., 1985.
32. Shimizu, T., O. Radmark, and B. Samuelsson. Enzyme with dual lipoxygenase activities
catalyzes leukotriene A4 synthesis from arachidonic acid. Proceedings of the National Academy
of Sciences of the United States of America 81: 689-93, 1984.
33. Stenmark, K. R., S. L. James, N. F. Voelkel, W. H. Toews, J. T. Reeves, and R. C.
Murphy. Leukotriene C4 and D4 in neonates with hypoxemia and pulmonary hypertension. N
Engl J Med 309: 77-80., 1983.

23
34. Stenmark, K. R., M. L. Morganroth, L. K. Remigio, N. F. Voelkel, R. C. Murphy, P. M.
Henson, M. M. Mathias, and J. T. Reeves. Alveolar inflammation and arachidonate metabolism
in monocrotaline- induced pulmonary hypertension. Am J Physiol 248: H859-66., 1985.
35. Thomson, J. R., R. D. Machado, M. W. Pauciulo, N. V. Morgan, M. Humbert, G. C. Elliott,
K. Ward, M. Yacoub, G. Mikhail, P. Rogers, J. Newman, L. Wheeler, T. Higenbottam, J. S.
Gibbs, J. Egan, A. Crozier, A. Peacock, R. Allcock, P. Corris, J. E. Loyd, R. C. Trembath, and
W. C. Nichols. Sporadic primary pulmonary hypertension is associated with germline mutations
of the gene encoding BMPR-II, a receptor member of the TGF- beta family. J Med Genet 37:
741-5., 2000.
36. Trembath, R. C., J. R. Thomson, R. D. Machado, N. V. Morgan, C. Atkinson, I. Winship,
G. Simonneau, N. Galie, J. E. Loyd, M. Humbert, W. C. Nichols, N. W. Morrell, J. Berg, A.
Manes, J. McGaughran, M. Pauciulo, and L. Wheeler. Clinical and molecular genetic features of
pulmonary hypertension in patients with hereditary hemorrhagic telangiectasia. N Engl J Med
345: 325-34., 2001.
37. Voelkel, N. F., R. M. Tuder, J. Bridges, and W. P. Arend. Interleukin-1 receptor
antagonist treatment reduces pulmonary hypertension generated in rats by monocrotaline. Am J
Respir Cell Mol Biol 11: 664-75., 1994.
38. Voelkel, N. F., R. M. Tuder, K. Wade, M. Hoper, R. A. Lepley, J. L. Goulet, B. H. Koller,
and F. Fitzpatrick. Inhibition of 5-lipoxygenase-activating protein (FLAP) reduces pulmonary
vascular reactivity and pulmonary hypertension in hypoxic rats. J Clin Invest 97: 2491-8., 1996.
39. Walker, J. L., J. Loscalzo, and Y. Y. Zhang. 5-Lipoxygenase and human pulmonary artery
endothelial cell proliferation. Am J Physiol Heart Circ Physiol 282: H585-93., 2002.

24
40. Wilson, D. W., H. J. Segall, L. C. Pan, and S. K. Dunston. Progressive inflammatory and
structural changes in the pulmonary vasculature of monocrotaline-treated rats. Microvasc Res
38: 57-80., 1989.
41. Wright, L., R. M. Tuder, J. Wang, C. D. Cool, R. A. Lepley, and N. F. Voelkel. 5-
Lipoxygenase and 5-lipoxygenase activating protein (FLAP) immunoreactivity in lungs from
patients with primary pulmonary hypertension. Am J Respir Crit Care Med 157: 219-29., 1998.
42. Yeager, M. E., G. R. Halley, H. A. Golpon, N. F. Voelkel, and R. M. Tuder. Microsatellite
instability of endothelial cell growth and apoptosis genes within plexiform lesions in primary
pulmonary hypertension. Circ Res 88: E2-E11., 2001.
43. Zhang, Y. Y., J. L. Walker, A. Huang, J. F. Keaney, C. B. Clish, C. N. Serhan, and J.
Loscalzo. Expression of 5-lipoxygenase in pulmonary artery endothelial cells. Biochem J 361:
267-76., 2002.

25
Figure Legends
Figure 1. Northern blotting analysis of the time course of 5LO transgene expression. Rats were
treated with PBS, MCT, or MCT+Ad5LO, and total RNA was isolated from the lung tissues of
the rats at days 3, 7, 10, and 15. Northern blotting analysis was carried out using an [α-32P]-
dCTP-labeled human 5LO cDNA probe (upper panel) and a probe against rat GAPDH (lower
panel). RNA samples from three individual rats were combined and examined at each time
point.
Figure 2. Distribution of 5LO transgene protein in rat lungs delivered by intratracheal
aerosolization. Rats were treated with PBS (panel A) or MCT+Ad5LO (panels B and C), and the
lung tissues were collected at day 10 after treatment. Paraffin embedded lung sections were
stained with rabbit anti-human 5LO antiserum (1:1600 dilution) (panels A and C) and rabbit pre-
bleed serum (1:1600 dilution) (panel B), and developed with biotinylated-alkaline phosphatase
and Vector Red Substrate. The slides were counterstained with hemotoxylin.
Figure 3. Pulmonary artery acceleration time (PAAT). Rats were treated with MCT+Ad5LO ( ),
MCT+AdGFP ( ), MCT only ( ), Ad5LO only ( ), or PBS ( ) and followed by transthoracic
echocardiography over time. The PAAT was significantly shorter in the MCT+Ad5LO group
from day 10 to day 18 as compared to all other groups. No difference existed between the
MCT+AdGFP and the MCT only groups. The Ad5LO alone did not decrease the PAAT as
compared to the PBS treatment group. *p<0.05 vs. PBS treatment and Ad5LO only groups.
#p<0.05 vs. MCT+AdGFP and MCT only groups. All values expressed as mean ± standard
deviation; n=3-10 per group.

26
Figure 4. Right ventricular systolic pressure (RVSP). Rats were treated with MCT+Ad5LO ( ),
MCT+AdGFP ( ), MCT only ( ), Ad5LO only ( ), or PBS ( ) and right heart catheterization
was performed on rats from each group on days 0, 10, 15, 18, and 24. The RVSP was
significantly higher in the MCT+Ad5LO group from day 10 to day 18 as compared to all other
groups. No difference existed between the MCT+AdGFP and the MCT only groups. Treatment
with Ad5LO alone did not increase the RVSP as compared to controls. *indicates p<0.05 vs.
PBS treatment and Ad5LO only groups. #indicates p<0.05 vs. the MCT+AdGFP and MCT only
groups. All values expressed as mean ± standard deviation. n=3-8 per group.
Figure 5. Right ventricle to left ventricle plus septum (RV/LV+S) weight ratio. Rats were treated
with PBS, Ad5LO, MCT, MCT+AdGFP, or MCT+Ad5LO, and sacrificed at day 26. The
RV/LV+S weight ratio was measured immediately after sacrifice. * indicates p<0.01 vs. the PBS
groups and # indicates p<0.02 vs. MCT+AdGFP group. All values are expressed as mean ±
standard deviation. n=3-8 per group.
Figure 6. Inflammatory cell infiltration in lungs. Rats were treated with PBS, MCT,
MCT+AdGFP, or MCT+Ad5LO. Paraffin-embedded lung tissue sections were prepared at days
7, 15, and 22 after treatment. Figure 6A shows hematoxylin and eosin-stained lung sections
from rats treated with PBS (A), MCT (B), MCT+AdGFP (C), and MCT+Ad5LO (D) harvested on
day 15. Figure 6B shows the number of monocytes/macrophages in lung sections prepared
from rats treated with MCT (white), MCT+AdGFP (gray), and MCT+Ad5LO (cross hatch), and
prepared at days 7, 15, and 22. The lung sections were stained with anti-monocyte/
macrophage antibody, and the cells in each section were counted in twenty consecutive fields
(200x). Data represent mean ± standard deviation of the number of cells in the lung tissue

27
sections obtained from 3-5 rats at each time point. * indicates p<0.05 vs. MCT+AdGFP group
and # indicates p<0.05 vs. MCT group.
Figure 7. Muscularized distal pulmonary vessels. Rats were treated with PBS, MCT+AdGFP,
and MCT+Ad5LO on day 0, and the lung tissues were collected on days 7, 15, 22, and 26 after
treatment. Tissue sections were stained with anti-smooth muscle cell α-actin antibody and
developed with biotinylated-alkaline phosphatase and Vector Red Substrate. The numbers of
muscularized vessels with diameter of 10-50 µm, and located distal to terminal bronchioles were
counted in twenty consecutive fields (200x) per section. Figure 7A shows representative
photomicrographs (400x) of stained lung section from rats treated with PBS (A), MCT+AdGFP
(B), and MCT+Ad5LO (C) at day 22. Figure 7B shows the average number of the muscularized
distal vessels in the lung tissue sections obtained from 3-5 rats at each time point. Open bars
represent treatment with MCT+AdGFP, and solid bars represent treatment with MCT+Ad5LO.
All values are expressed as mean ± standard deviation.
Figure 8. Fibrosis in lung tissues of rats treated with MCT+Ad5LO and with MCT+AdGFP.
Lung sections from day 26 were stained with Masson’s trichrome stain in order to visualize
collagen deposition. Panels A-C show representative photomicrographs of lung sections from a
rat treated with PBS (A), a rat treated with MCT+AdGFP (B), and a rat treated with MCT+Ad5LO
(C).
Figure 9. Inhibition of 5-lipoxygenase reduces the development of pulmonary hypertension in
MCT+Ad5LO- and MCT-treated rats. A) Rats were treated with MCT+Ad5LO on day 0 and
received vehicle (DMSO) (gray bar), MK886 (black bar), or zileuton (white bar) on day 1 and

28
daily thereafter. Right heart catheterization was performed on days 10, 15, and 26. B) Rats
were treated with MCT on day 0 and received DMSO, MK886, or zileuton on day 1 and daily
thereafter. Right heart catheterization was performed on day 35. Data represent mean ±
standard deviation of values obtained from 4 (panel A) and 5 rats (panel B), *indicates p<0.05
vs. MCT+DMSO (MCT) group.

Human 5LO
3 7 10 153 7 10 153 7 10 15Day
PBS MCT MCT+Ad5LO
GAPDH
Figure 1

Figure 2
A
B
C
0 3 7 10 15 18 2420
25
30
35
40
45#*
Figure 3
* *
*
*
p<0.05 vs PBS or Ad5LOp<0.05 vs AdGFP+MCT
*
**
##
#
***
MCT+Ad5LO MCT+AdGFP MCT Ad5LO PBS
PA
AT
(mse
c)
Days after treatment

0 5 10 15 20 2510
20
30
40
50
60#*
Figure 4
p<0.05 vs. PBS or Ad5LOp<0.05 vs. AdGFP+MCT or MCT
**
*
****
#
##
*
**
MCT+Ad5LO MCT+AdGFP MCT Ad5LO PBS
RV
SP
(mm
Hg)
Days after treatment

PBS
Ad5LO MCT
MCT+AdG
FPMCT+A
d5LO
0.0
0.1
0.2
0.3
0.4
0.5
0.6
Figure 5
p< 0.02 vs. MCT+AdGFP group#p<0.01 vs. PBS group*
**
#
*
RV
/LV
+S w
eigh
t rat
io

A
C D
B
Figure 6A

7 15 220
100
200
300
400
500
600
MCT MCT+AdGFP MCT+Ad5LO
Figure 6B
*# p<0.05 vs. MCT
p<0.05 vs. MCT+AdGFP
#
#*
# m
onoc
ytes
/Mac
roph
ages
Day

A
B
C
Figure 7A

7 15 22 260
20
40
60
80
100
Figure 7B
# M
uscu
lariz
ed d
ista
l ves
sels
Day
MCT+AdGFP MCT+Ad5LO

Figure 8
A
B
C

Day 10 Day 15 Day 260
10
20
30
40
50
* p<0.05 vs. MCT+Ad5LO
**
**
**
Figure 9A
RV
SP
(mm
Hg)
MCT+Ad5LO MCT+Ad5LO+MK886 MCT+Ad5LO+zileuton

MCT MCT+MK886 MCT+zileuton0
10
20
30
40
50
60
Figure 9B
*
p<0.05 vs. MCT*
*
RV
SP
(mm
Hg)
Copyright © 2022 FDOKUMEN




















