
Ectonucleotide Pyrophosphatase/Phosphodiesterase (E-NPP) and Adenosine Deaminase (ADA) activities in...
13
*Departamento de Bioquímica y Biología Molecular, Facultad de Veterinaria, Universidad Complutense de Madrid, Madrid, Spain †Departamento de Bioquímica, Facultad de Optica y Optometría, Universidad Complutense de Madrid, Madrid, Spain Abstract Neuro-2a (N2a) neuroblastoma cells display an ectoenzymatic hydrolytic activity capable of degrading diadenosine polyphos- phates. The Ap n A-cleaving activity has been analysed with the use of the fluorogenic compound BODIPY â FL guanosine 5 0 -O-(3-thiotriphosphate) thioester. Hydrolysis of this dinucle- otide analogue showed a hyperbolic kinetic with a K m value of 4.9 1.3 lM. Diadenosine pentaphosphate, diadenosine tet- raphosphate, diadenosine triphosphate, and the nucleoside monophosphate AMP behaved as an inhibitor of BODIPY â FL guanosine 5 0 -O-(3-thiotriphosphate) thioester extracellular degradation. Ectoenzymatic activity shared the typical charac- teristics of the ectonucleotide pyrophosphatase/phosphodies- terase family, as hydrolysis reached maximal activity at alkaline pH and was dependent on the presence of divalent cations, being strongly inhibited by EDTA and activated by Zn 2+ ions. Both NPP1 and NPP3 isozymes are expressed in N2a cells, their expression levels substantially changing when cells differentiate into a neuronal-like phenotype. In this sense, it is relevant to point the expression pattern of the NPP3 protein, whose levels were drastically reduced in the differentiated cells, being almost completely absent after 24 h of differenti- ation. Enzymatic activity assays carried out with differentiated N2a cells showed that NPP1 is the main isozyme involved in the extracellular degradation of dinucleotides in these cells, this enzyme reducing its activity and changing its subcellular location following neuronal differentiation. Keywords: diadenosine polyphosphates, E-NPP enzymes, N2a cells, NPP1, NPP3. J. Neurochem. (2014) 10.1111/jnc.12794 Diadenosine polyphosphates comprise a group of com- pounds formed by two adenosine nucleosides joined by a phosphate chain of variable length. They are commonly abbreviated as Ap n A, where n represents the number of phosphate residues in the polyphosphate chain. Members of this family of dinucleotides have been identified in the releasable content of storage granules in specialized neuro- secretory cells and platelets and are thought to function as extracellular signalling molecules in the vascular and nervous systems (Flores et al. 1999; Miras-Portugal et al. 1999). In the nervous system, Ap n A(n = 2–6) and other dinucleoside polyphosphates have been found to be costored with mononucleotides, mainly ATP and ADP, in the catecholam- inergic and cholinergic vesicles from adrenomedullary Received February 20, 2014; revised manuscript received June 6, 2014; accepted June 16, 2014. Address correspondence and reprint requests to Dr Javier Gualix, Departamento de Bioqu ımica y Biolog ıa Molecular IV, Facultad de Veterinaria, UCM, Av. Puerta de Hierro s/n, 28040-Madrid, Spain. E-mail: [email protected] Abbreviations used: Ap 3 A, diadenosine triphosphate; Ap 4 A, diadeno- sine tetraphosphate; Ap 5 A, diadenosine pentaphosphate; Ap n A, diadeno- sine polyphosphates; BODIPY â FL, 4,4-difluoro-5,7-dimethyl-4-bora-3a, 4a-diaza-s-indacene-3-propionic acid; BODIPY-FL-GTPcS, BODIPY â FL guanosine 5 0 -O-(3-thiotriphosphate) thioester; BSA, bovine serum albumin; DiBucAMP, dibutyryl-cAMP; E-NPP, ecto-nucleotide pyro- phosphatase/phosphodiesterase (EC 3.1.4.1; EC 3.6.1.9); FBSi, heat- inactivated foetal bovine serum; Gp 4 G, diguanosine tetraphosphate; GTPcS, guanosine 5 0 -O-(3-thiotriphosphate); N2a, neuro-2a neuroblas- toma; PBS, phosphate-buffered saline; Q-PCR, quantitative real-time PCR; RT-PCR, reverse transcription PCR; SDS, sodium dodecyl sulphate. © 2014 International Society for Neurochemistry, J. Neurochem. (2014) 10.1111/jnc.12794 1 JOURNAL OF NEUROCHEMISTRY | 2014 doi: 10.1111/jnc.12794
Transcript of Ectonucleotide Pyrophosphatase/Phosphodiesterase (E-NPP) and Adenosine Deaminase (ADA) activities in...

*Departamento de Bioquímica y Biología Molecular, Facultad de Veterinaria, Universidad
Complutense de Madrid, Madrid, Spain
†Departamento de Bioquímica, Facultad de �Optica y Optometría, Universidad Complutense de Madrid,
Madrid, Spain
AbstractNeuro-2a (N2a) neuroblastoma cells display an ectoenzymatichydrolytic activity capable of degrading diadenosine polyphos-phates. The ApnA-cleaving activity has been analysed with theuse of the fluorogenic compound BODIPY� FL guanosine50-O-(3-thiotriphosphate) thioester. Hydrolysis of this dinucle-otide analogue showed a hyperbolic kinetic with a Km value of4.9 � 1.3 lM. Diadenosine pentaphosphate, diadenosine tet-raphosphate, diadenosine triphosphate, and the nucleosidemonophosphate AMP behaved as an inhibitor of BODIPY� FLguanosine 50-O-(3-thiotriphosphate) thioester extracellulardegradation. Ectoenzymatic activity shared the typical charac-teristics of the ectonucleotide pyrophosphatase/phosphodies-terase family, as hydrolysis reachedmaximal activity at alkalinepH and was dependent on the presence of divalent cations,
being strongly inhibited by EDTA and activated by Zn2+ ions.Both NPP1 and NPP3 isozymes are expressed in N2a cells,their expression levels substantially changing when cellsdifferentiate into a neuronal-like phenotype. In this sense, it isrelevant to point the expression pattern of the NPP3 protein,whose levels were drastically reduced in the differentiatedcells, being almost completely absent after 24 h of differenti-ation. Enzymatic activity assays carried out with differentiatedN2a cells showed that NPP1 is the main isozyme involved inthe extracellular degradation of dinucleotides in these cells, thisenzyme reducing its activity and changing its subcellularlocation following neuronal differentiation.Keywords: diadenosine polyphosphates, E-NPP enzymes,N2a cells, NPP1, NPP3.J. Neurochem. (2014) 10.1111/jnc.12794
Diadenosine polyphosphates comprise a group of com-pounds formed by two adenosine nucleosides joined by aphosphate chain of variable length. They are commonlyabbreviated as ApnA, where n represents the number ofphosphate residues in the polyphosphate chain. Members ofthis family of dinucleotides have been identified in thereleasable content of storage granules in specialized neuro-secretory cells and platelets and are thought to function asextracellular signalling molecules in the vascular and nervoussystems (Flores et al. 1999; Miras-Portugal et al. 1999). Inthe nervous system, ApnA (n = 2–6) and other dinucleosidepolyphosphates have been found to be costored withmononucleotides, mainly ATP and ADP, in the catecholam-inergic and cholinergic vesicles from adrenomedullary
Received February 20, 2014; revised manuscript received June 6, 2014;accepted June 16, 2014.Address correspondence and reprint requests to Dr Javier Gualix,
Departamento de Bioqu�ımica y Biolog�ıa Molecular IV, Facultad deVeterinaria, UCM, Av. Puerta de Hierro s/n, 28040-Madrid, Spain.E-mail: [email protected] used: Ap3A, diadenosine triphosphate; Ap4A, diadeno-
sine tetraphosphate; Ap5A, diadenosine pentaphosphate; ApnA, diadeno-sine polyphosphates; BODIPY� FL, 4,4-difluoro-5,7-dimethyl-4-bora-3a,4a-diaza-s-indacene-3-propionic acid; BODIPY-FL-GTPcS, BODIPY�
FL guanosine 50-O-(3-thiotriphosphate) thioester; BSA, bovine serumalbumin; DiBucAMP, dibutyryl-cAMP; E-NPP, ecto-nucleotide pyro-phosphatase/phosphodiesterase (EC 3.1.4.1; EC 3.6.1.9); FBSi, heat-inactivated foetal bovine serum; Gp4G, diguanosine tetraphosphate;GTPcS, guanosine 50-O-(3-thiotriphosphate); N2a, neuro-2a neuroblas-toma; PBS, phosphate-buffered saline; Q-PCR, quantitative real-timePCR; RT-PCR, reverse transcription PCR; SDS, sodium dodecyl sulphate.
© 2014 International Society for Neurochemistry, J. Neurochem. (2014) 10.1111/jnc.12794 1
JOURNAL OF NEUROCHEMISTRY | 2014 doi: 10.1111/jnc.12794

chromaffin cells and neurons (Rodriguez del Castillo et al.1988; Pintor et al. 1992a,b,c). All these secretory systemsrespond to depolarizing agents or secretagogues by releasingtheir vesicular content to the extracellular medium (Pintoret al. 1991, 1992a,c). In this regard, push-pull cannulaexperiments performed in living rats showed that afteramphetamine stimulation, rat neostriatum releases diadeno-sine tetraphosphate, Ap4A, and diadenosine pentaphosphate,Ap5A, which can be detected in the perfusion samples atconcentrations in the nanomolar range (Pintor et al. 1995).More recently, nanomolar concentrations of diadenosinepolyphosphates (including diadenosine triphosphate, Ap3A,together with Ap4A and Ap5A) have been also measured inmicrodialysis samples from the cerebellum of conscious ratsunder basal conditions, i.e. in the absence of any exoge-nously added stimulating substance (Gualix et al. 2014).Exocytotically released ApnA interact with specific dinucle-otide receptors but the ionotropic P2X, metabotropic P2Yand P1 adenosine receptors may also function as ApnAtargets (Pintor et al. 2000; Hoyle et al. 2001). Resultsobtained using very different experimental approachessupport the idea that extracellular ApnA, acting on P1, P2or their specific dinucleotide receptors, can effectivelymodulate neural functions (Pereira et al. 2000; Oaknin et al.2001; Jimenez et al. 2002; Delicado et al. 2006). Moreover,neuroprotective effects against injuries induced by ischemiaor 6-hydroxydopamine injection in rat brain have beendescribed for Ap4A (Wang et al. 2003).The inactivation of ApnA by ectoenzymes at the cell
surface provides a necessary mechanism to regulate thereceptor-mediated actions of these dinucleotides (Zimmer-mann et al. 2012). Ectoenzymatic hydrolysis of ApnA hasbeen analysed in adrenomedullary chromaffin cells (Rodri-guez-Pascual et al. 1992; Ramos et al. 1995) and synapticmembranes of the Torpedo electric organ (Mateo et al. 1997).Available data indicate that the membrane-bound enzymescatalyse the hydrolytic cleavage at the polyphosphate chain ofthe dinucleotides to produce the mononucleotidic moietiesAMP + adenosine 50(n � 1) phosphate. The subsequentlygenerated nucleotides are further degraded, yielding adeno-sine as the final product, which may be recovered into thecells through transport systems. Most of these ApnA-hydro-lysing enzymes display biochemical characteristics typical ofmembers of the ectonucleotide pyrophosphatase/phosphodi-esterase (E-NPP) family. This family contains seven mem-bers, which have been numbered NPP1–NPP7 according totheir order of cloning. NPP1–NPP3 have been previouslydescribed as the only members of the family able to hydrolysediadenosine polyphosphates. Each of the three enzymescleaves Ap3A, Ap4A and Ap5A at comparable rates withMichaelis constants (Km) in the low micromolar range(Vollmayer et al. 2003). However, it has been recentlydemonstrated that NPP4 can also hydrolyse Ap3A and Ap4A,although the Km of NPP4 binding to these dinucleotides is
much weaker than previously reported for NPP1–NPP3(Albright et al. 2012, 2014). NPP6 and NPP7 are only knownto hydrolyse phosphodiester bounds in lysophospholipids orother choline phosphodiesters, whereas substrates for NPP5have not yet been identified. Except for NPP2, which onlyexists as a secreted protein, all E-NPPs are single-spantransmembrane proteins (Stefan et al. 2005, 2006; Zimmer-mann et al. 2012). Recent data on the ApnA-hydrolyticactivity in plasma and synaptic membranes isolated from ratbrain point to NPP1 as the main ectoenzyme involved in thecleavage of ApnA by glial cells and neurons (Asensio et al.2007). NPP1 activity is widely distributed through the ratforebrain, with the highest activity in hypothalamus (Oakninet al. 2008). These results add new support for a signallingfunction of diadenosine polyphosphates in brain.Neuro-2a (N2a) is a mouse neural crest-derived tumour
cell line that has been extensively used to study neuronaldifferentiation, axonal growth, and signalling pathways. Thepresence of functional P2X7 nucleotide receptors that areinvolved in the regulation of neuronal differentiation andneurite formation has been described in these cells (Gomez-Villafuertes et al. 2009; Wu et al. 2009). These resultstogether with those obtained in human neuroblastomamodels, showing that P2X7 receptor stimulation resultedinto an enhancement of cell proliferation (Raffaghello et al.2006), support a role for the purinergic signalling in themechanisms that allow a fine control of the balance betweenproliferation and differentiation in neuroblastoma cells. N2acells also constitutively express P2Y2 receptors which aresensitive to dinucleoside tetraphosphates (such as Ap4A) andwhose activation enhances a-secretase activity and the non-amyloidogenic processing of the amyloid precursor protein(Leon-Otegui et al. 2011), thus precluding the formation oftoxic b-amyloid peptides.In this work, we analysed the presence of ectoenzymatic
activities that are capable of hydrolysing extracellulardiadenosine polyphosphates in N2a cells. The ApnA-cleav-ing activity in these cells displays biochemical features thatare typical of the members of the E-NPP family as has beendemonstrated by the use of the fluorogenic dinucleotideanalogue BODIPY� FL guanosine 50-O-(3-thiotriphosphate)thioester (BODIPY-FL-GTPcS) (Invitrogen, San Francisco,CA, USA). Both NPP1 and NPP3 ectoenzymes are expressedin N2a cells and their expression levels dramatically changewhen cells differentiate into a neuronal-like phenotype.Activity assays carried out with differentiated cells showedthat the dinucleotide hydrolytic activity largely depends onthe NPP1 isozyme.
Materials and methods
Materials
Ap5A, Ap4A, Ap3A, ATP, ADP, AMP, dibutyryl-cAMP (DiBu-cAMP) and monoclonal mouse anti-a-tubulin and mouse anti-
© 2014 International Society for Neurochemistry, J. Neurochem. (2014) 10.1111/jnc.12794
2 R. G�omez-Villafuertes et al.

MAP-2 antibodies were purchased from Sigma-Aldrich (St Louis,MO, USA). Micro-Bradford assay, acrylamide electrophoresisreagent and molecular weight protein standards were from Bio-Rad (Munich, Germany). Acetonitrile, HPLC grade, was purchasedfrom Scharlau (Barcelona, Spain). The commercial antibodies usedin this study were raised against NPP1 (also known as PC-1) orNPP3 (also known as CD203c) of human origin but are also suitablefor detection of the corresponding mouse proteins and werepurchased from Santa Cruz Biotechnology (Santa Cruz, CA,USA). Fluorescence immunodetection of NPP1 was carried outusing a different antibody raised against a synthetic peptidecorresponding to C-terminal amino acids 915-925 of human NPP1and was purchased from Abcam (Cambridge, UK). All otherreagents not specified were routinely supplied by Sigma-Aldrich.
Cell culture
N2a cells were cultured in Dulbecco’s modified Eagle’s medium(Sigma-Aldrich) supplemented with glutamax (Invitrogen, SanFrancisco, CA, USA), penicillin⁄streptomycin (Invitrogen) and10% heat-inactivated foetal bovine serum (FBSi, Euro-Clone,Padova, Italy). Cells were grown at 37°C in a humidifiedatmosphere containing 5% CO2.
In differentiation studies, cells were incubated with a lowerpercentage of FBSi (0.5%) and treated with 1 mM DiBucAMP, awell-established inductor of N2a differentiation into a neuronal-likephenotype (Fischer et al. 1986).
HPLC studies on extracellular degradation of ApnA
N2a cells were plated at 750 000 cells/well in six-well plates. After24 h, culture medium was removed and replaced by Locke’ssolution (composition in mM: NaCl, 140; KCl, 4.5; CaCl2, 2.5;KH2PO4, 1.2; MgSO4, 1.2; glucose, 5.5; HEPES, 10; pH 7.4).10-lM Ap5A was added to the wells and cells were maintained at37°C in a humidified atmosphere containing 5% CO2. To analyseAp5A cleavage, samples of the incubation medium were taken atdifferent times (t = 0, 30 min, 1 h, 2 h) and the amount of Ap5Awas measured by HPLC detection. As a control, Ap5A wasincubated in parallel but in the absence of the cells.
The chromatographic system consisted of a Waters (Milford, MA,USA) 1515 isocratic HPLC pump, a 2487 dual absorbance detectorand a Reodyne injector, all managed by the Breeze software fromWaters. Analysis was performed under ion-pair chromatographyconditions by equilibrating the chromatographic system with thefollowing mobile phase: 10 mM KH2PO4, 2 mM tetrabutylammo-nium hydrogen sulphate and 20% acetonitrile, pH 7.5. The columnwas a Nova-Pak C-18 (15-cm length, 0.4-cm diameter) from Waters.Detection was monitored at 260-nm wavelength. Ap5A peaks wereidentified by their retention times. Integrated peak areas of thesamples were normalized with respect to those obtained at t = 0,which were set as a 100%.
Fluorimetric enzyme assays
For the ectonucleotidase activity assays, cells were plated at 50 000cells/well in black with clear bottom 96-well plates and allowed tosettle for 2 h. After that, culture medium was removed and replacedby 50 lL of Locke0s solution. Reaction was started by the additionof the fluorogenic substrate BODIPY-FL-GTPcS dissolved in 50 lLof Locke0s solution for a final reaction volume of 100 lL. Increase
in fluorescence emission owing to BODIPY-FL-GTPcS cleavagewas monitored using a FLUOstar Optima plate reader (BMGLabtech, Ortenberg, Germany). Appropriate wavelengths wereselected with the use of 485-nm excitation and 520-nm emissionfilters. Basal fluorescence was recorded by assaying the fluorescentsubstrate in the presence of 50 mM EDTA (a divalent cationchelator that blocks ectoenzymatic dinucleotide-hydrolysing activityin N2a cells) and was subtracted in all the cases. To establishequivalence between fluorescence units and substrate moles,fluorescence was read again after total substrate hydrolysis, whichwas ensured by the addition of Crotalus adamanteus phosphodies-terase (0.02 U, Sigma-Aldrich). Enzymatic activity values werecalculated by determining the amount of substrate that is hydrolysedafter the initial 30 seconds of reaction in which a linear time-dependent fluorescence increase was obtained and substrate con-sumption was below 10% at any of the substrate concentrationsassayed. For pH and cation dependence studies, reaction mixturewas accordingly modified.
Reverse transcription PCR (RT-PCR) and quantitative real-time PCR
(Q-PCR)
Total RNA from N2a cells was extracted using RNeasy� plus minikit (Qiagen, Hilden, Germany), following the manufacturer’sinstructions. After digestion with TURBO DNase (Ambion, Austin,TX, USA), total RNA was quantified and reverse transcribed usingM-MLV reverse transcriptase, 6 lg of random primers and 350 lMdNTPs (Invitrogen, San Francisco, CA, USA). PCR reactions werecarried out using AmpliTools Master Mix (Biotools), 5 lL of theRT product and specific oligonucleotide primers for mouse ectonu-cleotide pyrophosphatases/phosphodiesterases NPP1 and NPP3(Sigma-Aldrich). For NPP1 (NM_008813.3), the primers usedwere: forward 50-CGGACGCTATGATTCCTTAGA-30 and reverse50-AGCACAATGAAGAAGTGAGTCG-30. For NPP3 (NM_134005.2), the primers used were: forward 50-GATGCACAGGACGAGGAGAC-30 and reverse 50-TGCACGTCCATATTTGAGTTG-30. Amplicon lengths were 93 bp and 75 bp for NPP1and NPP3 respectively. PCR reactions involved an initial denatur-ation step at 94�C for 5 min, followed by 40 cycles of amplification(94°C for 30 s; 60°C for 30 s and 72°C for 30s) conducted with a2720 Thermal Cycler (Applied Biosystems). Control reactions werecarried out in the absence of RT product (template) to avoid crosscontamination. Amplified PCR products were electrophoresed on a2.5% agarose gel and visualized by SYBR� Safe DNA gel stain(Invitrogen). GeneRuler 1 kb Plus DNA Ladder (Thermo Scientific,Rockford, IL, USA) was used as DNA size marker.
Q-PCR reactions were performed using the previous gene-specific primers in combination with specific Taqman 30-minorgroove binder (MGB)-DNA probes (Roche, Basel, Switzerland).The probes designed were FAM-50-GCCAGGAA-30-MGB andFAM-50-CTGCTGGG-30-MGB for NPP1 and NPP3 respectively.Fast thermal cycling was performed using a StepOnePlus� Real-Time System (Applied Biosystems, Foster City, CA, USA) asfollows: denaturation, one cycle of 95�C for 20 s, followed by 40cycles each of 95�C for 1 s and 60�C for 20 s. The results werenormalized as indicated by parallel amplification of glyceralde-hyde-3-phosphate dehydrogenase housekeeping gene. For glycer-aldehyde-3-phosphate dehydrogenase, commercial primers andTaqMan MGB probe were supplied by Applied Biosystems.
© 2014 International Society for Neurochemistry, J. Neurochem. (2014) 10.1111/jnc.12794
Analysis of E-NPP activity in N2a cells 3

Western blot detection of E-NPP isozymes
N2a cells were lysed and homogenized for 1 h at 4°C in lysis buffercontaining 50 mm Tris⁄HCl, 150 mm NaCl, 1% Nonidet P-40 andComplete� Protease Inhibitor Cocktail Tablets (Roche), pH 7.4.Protein content in the cell extracts was determined by the Bradfordmethod and then samples were mixed with loading buffer 5X (50%glycerol, 250 mM Tris, 10% sodium dodecyl sulphate, 0.5%bromophenol blue, 7.7 % dithiothreitol, pH = 6.8) and heated at99°C for 5 min. Aliquots of 30-lg protein were subjected to sodiumdodecyl sulphate gel electrophoresis using 8% acrylamide gels andtransferred to nitrocellulose membranes (Amersham GE Healthcare,Madrid, Spain). Membranes were blocked for 1 h at 25�C with 5%non-fat dried milk or 10% donkey serum in phosphate-bufferedsaline containing 0.1% Tween 20 [phosphate-buffered saline (PBS)-Tween] and incubated overnight at 4°C with the following antiseraat the dilutions specified in parentheses: mouse anti-NPP1 (1 : 100),goat anti-NPP3 (1 : 200) and mouse anti-a-tubulin (1 : 10 000).Blots were then washed in PBS-Tween and incubated for 1 h at25�C with goat anti-mouse or rabbit anti-goat IgGs coupled tohorseradish peroxidase (Dako Cytomation, Glostrup, Denmark),both used at 1 : 1000 dilution. Protein bands were detected byenhanced chemiluminescence detection (Amersham GE Health-care). Chemiluminescence images were acquired using the Image-Quant LAS 500 imager (GE Healthcare Bio-Sciences, Uppsala,Sweden) and quantified by means of the ImageQuant TL 8.1software (GE Healthcare Bio-Sciences).
Immunocytochemistry
N2a cells cultured on coverslips placed in 35-mm dishes (350 000cells/well) were washed with PBS and fixed with 4% paraformal-dehyde for 15 min. After washing with PBS, cells were permea-bilized with 0.1% Triton X-100 and blocked with 5% donkeyserum and 3% bovine serum albumin in PBS for 1 h at 25�C.After washing with 3% bovine serum albumin in PBS, the cellswere incubated for 1 h with primary antibodies: goat anti-NPP1(1 : 250), goat anti-NPP3 (1 : 50) and mouse anti-MAP-2(1 : 100). Positive immunostaining was revealed using AlexaFluor 647 donkey anti-goat and Alexa Fluor 488 goat anti-mouseIgGs (Invitrogen). Nuclei were counter-stained with 40,6-diami-dino-2-phenylindole (Invitrogen). Coverslips were mounted onglass slides using ProLong� Gold Antifade Reagent (Life Tech-nologies, Carlsbad, CA, USA). Images were acquired using aLeica CTR 6500 confocal microscope (Leica Microsystems,Wetzlar, Germany) with an ACS APO 409 immersion oilobjective (NA 1.15).
Statistical analysis
Data were analysed using one-way ANOVA followed by Dunnett’s orBonferroni’s post hoc comparisons test (Graph Pad Prism 5, GraphPad Software Inc., San Diego, CA, USA). Data are expressed as themean � SEM. Differences were considered significant at p ≤ 0.05.
Results
Presence of an ectoezymatic ApnA-cleaving activity in N2acells
To analyse the presence of ectoenzymatic activities able tohydrolyse diadenosine polyphosphates, N2a cells were
incubated with 10 lM Ap5A and aliquots of the incubationmedium were taken at different times (t = 0, 30 min, 1 h and2 h) to follow the extracellular dinucleotide degradation byHPLC detection. As it is shown in Fig. 1a and b, extracel-lular levels of Ap5A were significantly reduced after 30 minand 1 h of incubation with the N2a cells and this dinucle-otide could not be detected at measurable quantities in theextracellular medium after a 2-h incubation period. Thepresence of mononucleotide moieties that are generated asintermediate products of Ap5A hydrolysis could not beaccurately analysed as these compounds do not accumulatein the extracellular medium. Rather, they are rapidlydegraded by nucleotidases and converted to adenosine whichappeared at the chromatogram front (Fig 1a). No significanthydrolysis of Ap5A was observed in the absence of cells(Fig. 1b).
0 2 4 6 8 10 12 14 16 18 20
0,0002 AUFS
Time
Ap5A
Retention time (min)
t = 0
t = 30
'
t = 1
ht =
2 h
0
20
40
60
80
100
120
Ap 5
A (%
)
(a)
(b)
Fig. 1 Presence of an ApnA-hydrolytic activity in neuro-2a (N2a) cells.(a) Representative HPLC profiles showing extracellular degradation of
diadenosine pentaphosphate (Ap5A) by N2a cells. Cells were incu-bated at 37°C with 10 lM Ap5A for 0, 30, 60 and 120 min. Aliquots of15 lL of the extracellular medium were injected into the chromato-
graph and peaks were detected by absorbance detection at 260-nmwavelength. Successive chromatograms in the figure (from top tobottom) show the results obtained at increased incubation times. Ap5Apeaks in the samples were identified by their retention time. (b)
Integrated areas under Ap5A peaks were normalized with respect tothose obtained at t = 0 which were set as a 100% (white bars). As acontrol, Ap5A was incubated under the same conditions but in the
absence of cells (black bars). Results are the mean � SEM (n = 3).
© 2014 International Society for Neurochemistry, J. Neurochem. (2014) 10.1111/jnc.12794
4 R. G�omez-Villafuertes et al.

BODIPY-FL-GTPcS: a fluorogenic substrate for the
characterization of the ApnA-cleaving activity in N2a cells
ApnA-hydrolysing enzymatic activity was analysed with theuse of the fluorogenic compound BODIPY-FL-GTPcS,formed by the BODIPY FL fluorophore linked through thec-thiol of guanosine 50-O-(3-thiotriphosphate) (GTPcS).BODIPY-FL-GTPcS cleavage is accompanied by a 5-foldincrease in fluorescence emission because BODIPY fluores-cence is quenched by stacking with the guanosine base in theintact molecule (Draganescu et al. 2000). Time-dependentincrease in the fluorescence emission because of BODIPY-FL-GTPcS cleavage can be used to calculate the hydrolysisrate.Extracellular BODIPY-FL-GTPcS was degraded by intact
N2a cells. Cleavage of the fluorogenic dinucleotide analoguewas concentration dependent, showing a typical hyperbolickinetic with a Km value of 4.9 � 1.3 lM (Fig 2a). SimilarKm values in the low micromolar range have been observedfor the extracellular hydrolysis of dinucleotides by chromaf-fin cells and synaptic membranes from Torpedo electricorgan or rat brain (Rodriguez-Pascual et al. 1992; Ramoset al. 1995; Mateo et al. 1997; Asensio et al. 2007).Moreover, heterologously expressed NPP1, NPP2 andNPP3 also hydrolyse the diadenosine polyphosphates withKm values in the low micromolar range (Vollmayer et al.2003). The extracellular hydrolysis of BODIPY-FL-GTPcS
was inhibited by Ap5A in a dose-dependent manner, thisdinucleotide reducing the cleavage of the fluorogenicsubstrate by a 78% at 200 lM concentration (Fig. 2b). Asimilar behaviour was observed when Ap4A was assayed asan inhibitor of BODIPY-FL-GTPcS degradation, this com-pound being able to reduce the hydrolysis of the fluorogenicsubstrate by a 61% at 100-lM concentration (Fig. 2c). Ap3A,at 100 lM concentration, also acted as an inhibitor byreducing extracellular BODIPY-FL-GTPcS cleavage by a65% (Fig. 2c). These results indicate that a common
0
20
40
60
80
100
** *****
******
(b)
(c)
0 50 100 150 2000
20
40
60
80
100
[Ap5A] (μM)
Act
ivity
(%)
Control
250 μ
M Ap 4A
100 μ
M Ap 3A
100 μ
M ATP
100 μ
M ADP
100 μ
M AMP
Act
ivity
(%)
0 5 10 15 200
10
20
30
40
[BODIPY-FL-GTPγS] (μM)
Act
ivity
(pm
ol/m
in)
(a)
100 μ
M Ap 4A
500 μ
M Ap 4A
Fig. 2 Use of the fluorogenic BODIPY� FL guanosine 50-O-(3-thiotri-phosphate) thioester (BODIPY-FL-GTPcS) substrate to characterizethe ApnA-cleaving activity in neuro-2a (N2a) cells. (a) Kinetic of thehydrolysis of BODIPY-FL-GTPcS by N2a cells. Cells were incubated at
37°C with BODIPY-FL-GTPcS concentrations ranging from 1 to 20 lMand time-dependent increase in fluorescence emission because ofsubstrate cleavage was monitored by the use of a plate reader. To
establish equivalence between fluorescence units and substrate molesfluorescence was read again after total substrate hydrolysis by theaddition ofC. adamanteus phosphodiesterase. Initial slope of the traces
was used to calculate the reaction rates. Graph represents themean � SEM of three experiments performed in triplicate. (b)Inhibition of BODIPY-FL-GTPcS cleaving activity by diadenosine
pentaphosphate (Ap5A). N2a cells were incubated with 10 lMBODIPY-FL-GTPcS at varying concentrations of Ap5A (10–200 lM). Activities are expressed as percentage of linear fluores-cence trace slopes in the presence of Ap5A relative to control slope
without the inhibitor. Graph represents the mean � SEM of twoexperiments performed in triplicate. (c) Inhibition of BODIPY-FL-GTPcS cleaving activity by mono- and dinucleotides. Cells were
incubated with 10 lM BODIPY-FL-GTPcS in the absence (control) orin the presence of the indicated concentrations of diadenosinetetraphosphate (Ap4A), diadenosine triphosphate (Ap3A), ATP, ADP
or AMP. Activities are expressed as percentage of those measuredin control situation. Bars represent the mean � SEM of threeexperiments performed in triplicate. Statistical significance versus
control, **p < 0.01, ***p < 0.001 (ANOVA with the Dunnett’s post hoctest).
© 2014 International Society for Neurochemistry, J. Neurochem. (2014) 10.1111/jnc.12794
Analysis of E-NPP activity in N2a cells 5

enzymatic activity showing broad substrate specificity is theresponsible for both the extracellular hydrolysis of BODIPY-FL-GTPcS and diadenosine polyphosphates, ApnA thenacting as competitors on BODIPY-FL-GTPcS degradation.Similar broad substrate specificity has been described for theectoenzymatic activity able to hydrolyse diadenosinepolyphosphates in chromaffin cells and Torpedo electricorgan or rat brain membranes (Rodriguez-Pascual et al. 1992;Ramos et al. 1995; Mateo et al. 1997; Asensio et al. 2007).Also, heterologously expressed NPP1, NPP2 and NPP3 havebeen shown to be able to hydrolyse several different ApnAalong with the diguanosine polyphosphate, diguanosinetetraphosphate (Gp4G) (Vollmayer et al. 2003). Thus,BODIPY-FL-GTPcS can be used as a useful substrate withwhich to analyse and characterize the ectoenzymatic activityresponsible for the extracellular degradation of dinucleotidesin N2a cells. A similar strategy, using etheno-derivatives ofApnA as fluorogenic substrates, has been previously used tocharacterize the dinucleotide-hydrolysing activities in chro-maffin cells and Torpedo electric organ and brain membranes(Ramos et al. 1995; Mateo et al. 1997; Asensio et al. 2007).Regarding mononucleotides, only AMP was effective byreducing the extracellular cleavage of BODIPY-FL-GTPcS.This nucleotide inhibited BODIPY-FL-GTPcS-hydrolysingactivity by a 55% at 100-lM concentration (Fig. 2c). It isrelevant to note that E-NPP enzymes asymmetrically cleavethe polyphosphate chain of ApnA to produce the mononu-cleotidic moieties AMP + adenosine 50(n � 1) phosphate asreaction products (Bollen et al. 2000; Vollmayer et al. 2003;Stefan et al. 2005; Zimmermann et al. 2012). AMP is notfurther hydrolysed by the E-NPP, as expected from theabsence of phosphodiester or pyrophosphate bonds, but exertscompetitive product inhibition on the ectoenzyme activity(Bollen et al. 2000; Vollmayer et al. 2003; Zimmermannet al. 2012). In this sense, strong inhibition by the reactionproduct AMP has been reported for the NPP1 activity presentin brain membranes (Asensio et al. 2007).
Biochemical features of the ApnA-cleaving activity in N2a
cells
The effect of divalent cations and pH on the ectoenzymaticApnA-hydrolysing activity was analysed by the use offluorogenic substrate BODIPY-FL-GTPcS. Dinucleotide-cleaving activity in the N2a cells was dependent on thepresence of divalent cations in the extracellular medium, asdemonstrated by the effect of EDTA which reduced thehydrolytic activity in a dose-dependent manner, BODIPY-FL-GTPcS cleavage being almost completely abolished inthe presence of 50 mM of the cation chelator (Fig 3a). Whenthe enzymatic activity was assayed in a Mg2+-free Locke’ssolution (containing only 2.5 mM Ca2+ as divalent cation),activity was 63.4 � 6.5 % of that in the standard medium(containing both 1.2 mM Mg2+ and 2.5 mM Ca2+). Simi-larly, hydrolysing activity was reduced to a 33.9 � 1.6 % in
a virtually Ca2+-free medium (obtained by the addition of6 mM of the selective Ca2+ chelator EGTA to the Locke’ssolution). This indicated an additive effect of both Mg2+ andCa2+ stimulating ectoenzymatic dinucleotide-cleaving activ-ity. When assays were carried out in the absence of thesedivalent cations (in a Mg2+-free Locke’s solution supple-mented with 6 mM EGTA), BODIPY-FL-GTPcS-cleavingactivity was reduced to a 19.5 � 4.0 %, this residual activitybeing increased to a 36.2 � 2.5 % when 200 lM Zn2+ wasadded to the medium (Fig. 3b), thus demonstrating theactivatory effect exerted by Zn2+ ions.Regarding the influence of pH, dinucleotide hydrolase
activity was stimulated in alkaline conditions. In the pHrange 6.5–8.5, maximal activity was observed at pH 8.5, as itis shown in Fig. 3c. There were no changes in thefluorescence emission of BODIPY-FL-GTPcS within thisinterval.
N2a cells express NPP1 and NPP3 isozymes
General characteristics of ectoenzymatic dinucleotide-cleav-ing activity in N2a cells, such as alkaline optimum pH,activation by Ca2+, Mg2+ and Zn2+ ions and inhibition byEDTA fit well with those of the E-NPP family members thathydrolyse ApnA (NPP1-NPP4). NPP4 has shown a loweraffinity for the diadenosine polyphosphates than thosereported here for the dinucleotide-hydrolysing activity inN2a cells. NPP4 present on the surface of vascular endothe-lium hydrolyses Ap3A and Ap4A with Km values of 843 lMand 210 lM respectively (Albright et al. 2012), whereas asoluble form of the enzyme, obtained via C-terminaltruncation to eliminate the cytoplasmic and transmembranedomains of human NPP4, cleaved Ap3A with a Km value of685 lM (Albright et al. 2014). However, the affinity valuesobserved here for the dinucleotide hydrolysis by N2a cells,which are in the low lM range, fit well with those describedfor NPP1–NPP3 (Bollen et al. 2000; Vollmayer et al. 2003;Zimmermann et al. 2012). From these, NPP2 only exists as asecreted protein (Stefan et al. 2005, 2006; Zimmermannet al. 2012). As culture medium was replaced by Locke’ssolution immediately before the enzyme activity measure-ments, it seems unlikely that a secreted protein could be theresponsible of the dinucleotide-cleaving activity observed inour assays. Thus, we analysed the expression of NPP1 andNPP3 to discriminate the isozyme involved in the extracel-lular degradation of dinucleotides by N2a cells.RT-PCR analysis demonstrated that both NPP1 and
NPP3�are simultaneously expressed in N2a cells (Fig. 4a).To characterize the relative amount of each NPP transcript,Q-PCR assays were performed. As it is shown in Fig. 4bcomparable levels were obtained for both NPP1 and NPP3mRNAs. Western blot analyses were performed on totalprotein extracts obtained from N2a cells using commercialisozyme-specific antibodies. As expected, the expression ofNPP proteins correlated well with the results obtained by
© 2014 International Society for Neurochemistry, J. Neurochem. (2014) 10.1111/jnc.12794
6 R. G�omez-Villafuertes et al.

quantitative PCR. Thus, bands corresponding to NPP1 andNPP3 proteins were immunodetected (Fig. 4c). These pro-teins contain several consensus sequences for N-linked
glycosylation. The occurrence of N-glycosylation in vivoaffects the electrophoretic mobility of the proteins (Bollenet al. 2000; Zimmermann et al. 2012) that can varydepending on the cell type and species. The bands obtainedin this study were consistent with the range of molecularsizes reported for NPP1 and NPP3 in the literature (Donget al. 2005; Jankowski et al. 2011).
Changes in E-NPP expression in differentiated N2a cellsIt is well known that purinergic signalling may regulateneuroblastoma cells growth (Raffaghello et al. 2006) andchanges in the expression and function of nucleotidereceptors occurs when N2a cells differentiate into neuronal-like cells (Gomez-Villafuertes et al. 2009; Wu et al. 2009).Ectonucleotidases play a pivotal role in the regulation of theextracellular levels of the nucleotidic effectors and hencevariations in their activity can have a deep influence in thecell proliferation and differentiation processes (Stefan et al.2005, 2006; Zimmermann et al. 2012). Here, we analyse theexpression of the E-NPP proteins when N2a cells differen-tiate. Variations in the expression of these enzymes, both atthe transcriptional and protein level, compared to non-differentiated cells, were measured.Q-PCR studies revealed that the endogenous levels of the
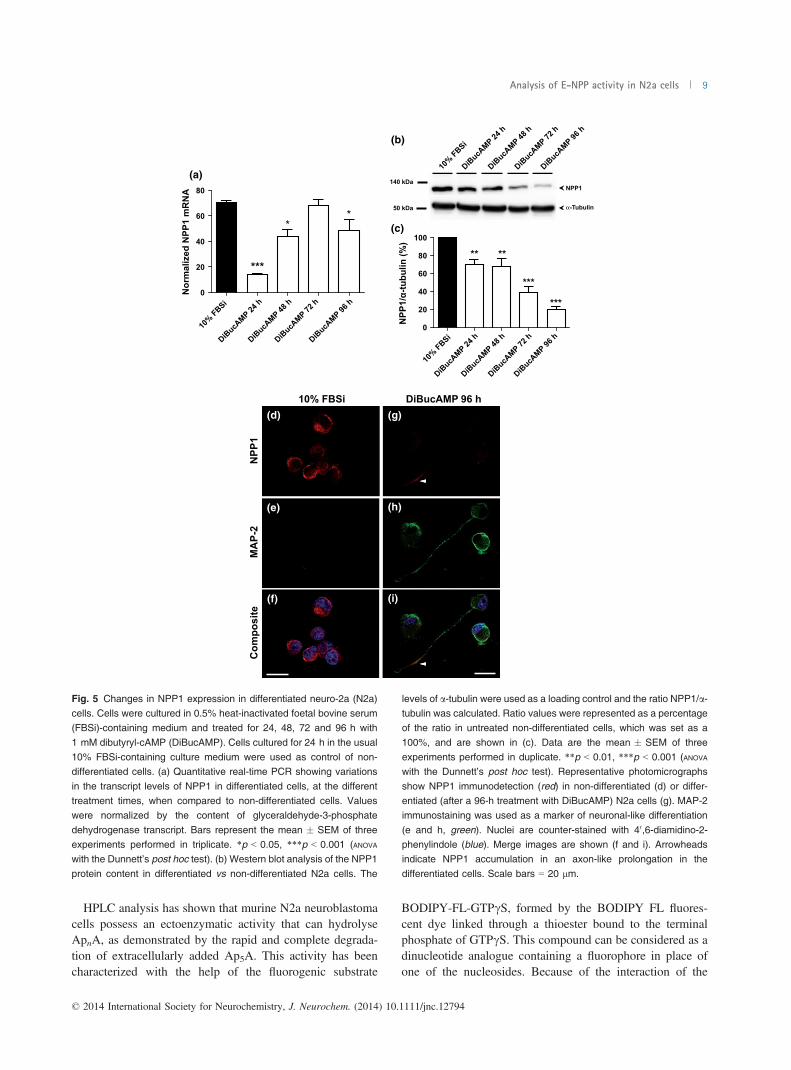
NPP1 transcript were very significantly reduced after 24 h ofdifferentiation treatment, NPP1 mRNA levels being recov-ered at longer times, 48 h and 72 h (Fig. 5a). Curiously,western blot analysis showed that NPP1 protein levels onlyexperimented a substantial decrease after 72 h and 96 h ofdifferentiation (Fig. 5b and c). Immunofluorescence studiesconfirmed that NPP1 protein is present in non-differentiatedcells (Fig. 5d–f), whereas their levels are significantlyreduced after 96 h of differentiation treatment (Fig. 5g–i).
(a)
(b)
+ 1 m
M EDTA
+ 3 m
M EDTA
+ 5 m
M EDTA
+ 10 m
M EDTA
+ 25 m
M EDTA
+ 50 m
M EDTA0
20
40
60
80
100
Act
ivity
(%)
– Mg2
+
+ EGTA
(–Ca2
+ )
–Mg2
+ + EGTA
(–Ca2
+ )
–Mg2
+ + EGTA
(–Ca2
+ ) + Zn2
+
0
20
40
60
80
100
**
******
***
#
Act
ivity
(%)
pH = 6.5
pH = 7.4
pH = 8.5
0
10
20
30
40
50
Act
ivity
(pm
ol/m
in)
(c)
Control
Control
Fig. 3 Effect of divalent cations and pH on ectoenzymatic dinucleo-tide-cleaving activity. (a) Cells were incubated with 10 lM BODIPY�
FL guanosine 50-O-(3-thiotriphosphate) thioester (BODIPY-FL-GTPcS)in the absence (control) and in the presence of the indicatedconcentrations of the cation chelator EDTA. Activities are expressed
as percentage of those measured in control situation. Bars representthe mean � SEM of three experiments performed in triplicate. (b) Cellswere incubated with 10 lM BODIPY-FL-GTPcS in the usual Locke0ssolution (control) or in a Mg2+-free Locke’s solution (obtained byreplacing MgSO4 by 7.9 mM glucose, a concentration that conservedthe solution osmolarity), a virtually Ca2+-free medium (obtained byadding 6 mM of the selective Ca2+ chelator EGTA to the Locke0ssolution), a medium in which both divalent cations were absent (Mg2+-free Locke’s solution plus 6 mM EGTA) or the latest medium butsupplemented with 200 lM ZnSO4. Bars represent the mean � SEM
of three experiments performed in triplicate. **p < 0.01, ***p < 0.001versus control; #p < 0.05 (ANOVA with the Bonferroni’s post hoc test). (c)neuro-2a (N2a) cells were incubated with 10 lM BODIPY-FL-GTPcS
and hydrolysis rates at different pH values were measured. Bars arethe mean � SEM of three experiments performed in triplicate.
© 2014 International Society for Neurochemistry, J. Neurochem. (2014) 10.1111/jnc.12794
Analysis of E-NPP activity in N2a cells 7

It is relevant to note that the remaining NPP1 protein seemsto change its intracellular distribution from a somatic to anaxon-like location in the differentiated cells (Fig. 5g and i).A similar pattern to that obtained for NPP1 was observed
when the levels of the NPP3 transcript were analysed. NPP3mRNA was substantially reduced in the differentiated cellsafter 24 h of treatment, and transcript levels were recoveredat longer times (Fig. 6a). However, a completely differentbehaviour was observed when the amounts of protein wereanalysed by western blot. NPP3 protein was dramaticallyreduced in the differentiated N2a cells, being almostcompletely absent after 24 and 48 h of treatment andshowing only a very slight recovery at longer times oftreatment (Fig. 6b and c). Fluorescence immunodetectionexperiments confirmed that NPP3 protein is present in non-differentiated cells (Fig. 6d–f), whereas their levels areundetectable in neurite-bearing N2a cells that express theneuronal marker MAP-2 after 96 h of differentiation(Fig. 6g–i).
NPP1 is the main isozyme involved in the extracellularcleavage of ApnA by N2a cells
To determine the NPP isozyme that is involved in theextracellular degradation of dinucleotides in N2a cells, weanalysed the changes in the ectoenzymatic hydrolytic activityin differentiated cells after 24 h or 96 h of treatment (inwhich one or both of the E-NPP proteins significantlyreduced their expression when compared to untreated cells).As it is shown in Fig. 7, hydrolytic activity was only partially
reduced in differentiated cells after 24 h of treatment, inwhich NPP3 protein was almost absent and NPP1 onlypartially reduced it expression. Moreover, in differentiatedcells after 96 h of treatment, in which NPP3 still had a verylow expression rate and expression of NPP1 was substan-tially reduced, ectoenzymatic activity was also significantlydecreased (when compared to both untreated cells ordifferentiated cells after 24 h of treatment). These resultsseem to indicate that ectoenzymatic activity able to hydrolyseApnA in N2a cells largely resides in the NPP1 isozyme, aschanges in ectoenzymatic hydrolytic activity appeared tomatch changes in NPP1 protein expression. Reduction inNPP3 expression, on the contrary, seems to have little or noeffect on ectoenzymatic dinucleotide-cleaving activity.
Discussion
Diadenosine polyphosphates (ApnA) have emerged as a newfamily of extracellular signalling molecules that can interactwith several membrane protein targets (Miras-Portugal et al.1999; Pintor et al. 2000; Hoyle et al. 2001), being involved inneurotransmitter/neuromodulatory functions in the nervoussystem (Pereira et al. 2000; Oaknin et al. 2001; Jimenez et al.2002; Delicado et al. 2006). Ectonucleotidases readily hydro-lysing ApnA into AMP and adenosine 50(n�1) phosphatecould provide a well-suited mechanism to enzymaticallyregulate signalling by these dinucleotides (Rodriguez-Pascualet al. 1992; Ramos et al. 1995; Mateo et al. 1997; Asensioet al. 2007).
NPP1 NPP3
140 kDa
-RT
-RT
NPP1NPP3
0
50 000
100 000
150 000
200 000
E-N
PP m
RN
A
NPP1NPP3
(a) (b)
(c)
Fig. 4 Ectonucleotide pyrophosphatases/phosphodiesterases NPP1 and NPP3
are simultaneously expressed in non-differentiated murine neuro-2a (N2a) neuro-blastoma cells. (a) RT-PCR expression of
NPP1 and NPP3. No amplification productswere observed in parallel assays carried outwithout template (-RT). DNA sizemarkerwas
GeneRuler 1 kb Plus DNA Ladder (ThermoScientific). (b) Quantitative real-time PCRshowing the transcript levels of NPP1 and
NPP3 in N2a cells. (c) Total protein isolatedfrom N2a cells was subjected to sodiumdodecyl sulphate (SDS)–polyacrylamide gelelectrophoresis, transferred onto nitrocell-
ulose membrane and incubated withspecific antibodies against NPP1 and NPP3.
© 2014 International Society for Neurochemistry, J. Neurochem. (2014) 10.1111/jnc.12794
8 R. G�omez-Villafuertes et al.
HPLC analysis has shown that murine N2a neuroblastomacells possess an ectoenzymatic activity that can hydrolyseApnA, as demonstrated by the rapid and complete degrada-tion of extracellularly added Ap5A. This activity has beencharacterized with the help of the fluorogenic substrate
BODIPY-FL-GTPcS, formed by the BODIPY FL fluores-cent dye linked through a thioester bound to the terminalphosphate of GTPcS. This compound can be considered as adinucleotide analogue containing a fluorophore in place ofone of the nucleosides. Because of the interaction of the
10% FBSi
(a)
(b)
(c)
0
20
40
60
80
**
Nor
mal
ized
NPP
1 m
RN
A
***
140 kDa
10% FBSi
DiBucA
MP 24 h
DiBucA
MP 48 h
DiBucA
MP 72 h
DiBucA
MP 96 h
10% FBSi
DiBucA
MP 24 h
DiBucA
MP 48 h
DiBucA
MP 72 h
DiBucA
MP 96 h
10% FBSi
DiBucA
MP 24 h
DiBucA
MP 48 h
DiBucA
MP 72 h
DiBucA
MP 96 h
NPP1
α-Tubulin50 kDa
0
20
40
60
80
100
** **
***
***
NPP
1/α-
tubu
lin (%
)
NPP
1M
AP-
2C
ompo
site
DiBucAMP 96 h(d) (g)
(e) (h)
(f) (i)
Fig. 5 Changes in NPP1 expression in differentiated neuro-2a (N2a)cells. Cells were cultured in 0.5% heat-inactivated foetal bovine serum(FBSi)-containing medium and treated for 24, 48, 72 and 96 h with
1 mM dibutyryl-cAMP (DiBucAMP). Cells cultured for 24 h in the usual10% FBSi-containing culture medium were used as control of non-differentiated cells. (a) Quantitative real-time PCR showing variations
in the transcript levels of NPP1 in differentiated cells, at the differenttreatment times, when compared to non-differentiated cells. Valueswere normalized by the content of glyceraldehyde-3-phosphate
dehydrogenase transcript. Bars represent the mean � SEM of threeexperiments performed in triplicate. *p < 0.05, ***p < 0.001 (ANOVA
with the Dunnett’s post hoc test). (b) Western blot analysis of the NPP1
protein content in differentiated vs non-differentiated N2a cells. The
levels of a-tubulin were used as a loading control and the ratio NPP1/a-tubulin was calculated. Ratio values were represented as a percentageof the ratio in untreated non-differentiated cells, which was set as a
100%, and are shown in (c). Data are the mean � SEM of threeexperiments performed in duplicate. **p < 0.01, ***p < 0.001 (ANOVA
with the Dunnett’s post hoc test). Representative photomicrographs
show NPP1 immunodetection (red) in non-differentiated (d) or differ-entiated (after a 96-h treatment with DiBucAMP) N2a cells (g). MAP-2immunostaining was used as a marker of neuronal-like differentiation
(e and h, green). Nuclei are counter-stained with 40,6-diamidino-2-phenylindole (blue). Merge images are shown (f and i). Arrowheadsindicate NPP1 accumulation in an axon-like prolongation in the
differentiated cells. Scale bars = 20 lm.
© 2014 International Society for Neurochemistry, J. Neurochem. (2014) 10.1111/jnc.12794
Analysis of E-NPP activity in N2a cells 9

BODIPY FL dye with the guanine base of GTP, BODIPY-FL-GTPcS shows significant fluorescence quenching that isrelieved when the phosphate chain of the molecule becomescleaved (Draganescu et al. 2000). This fluorogenic com-pound has been used to analyse the diadenosine triphosphatehydrolase activity of Fhit, one of the most frequently
inactivated proteins in lung cancer that functions as a tumoursuppressor (Draganescu et al. 2000). BODIPY-FL-GTPcS iscleaved by the N2a cells in a dose-dependent manner with aKm value in the low micromolar range. Inhibition studiesshowed that BODIPY-FL-GTPcS cleavage becomes reducedby Ap5A, Ap4A and Ap3A, thus indicating that a common
0
20
40
60
80
100
*** *** ******
NPP
3/α-
tubu
lin (%
)
140 kDa
10% FBSi
DiBucA
MP 24 h
DiBucA
MP 48 h
DiBucA
MP 72 h
DiBucA
MP 96 h
10% FBSi
DiBucA
MP 24 h
DiBucA
MP 48 h
DiBucA
MP 72 h
DiBucA
MP 96 h
10% FBSi
DiBucA
MP 24 h
DiBucA
MP 48 h
DiBucA
MP 72 h
DiBucA
MP 96 h
NPP3
α-Tubulin 50 kDa
(b)
(c)N
PP3
MA
P-2
Com
posi
te
10% FBSi DiBucAMP 96 h
0
20
40
60
80
***
** **
Nor
mal
ized
NPP
3 m
RN
A
(a)
(d) (g)
(e) (h)
(f) (i)
Fig. 6 Changes in NPP3 expression in differentiated neuro-2a (N2a)
cells. Cells were cultured in 0.5% heat-inactivated foetal bovine serum(FBSi)-containing medium and treated for 24, 48, 72 and 96 h with1 mM dibutyryl-cAMP (DiBucAMP). Cells cultured for 24 h in the usual
10% FBSi-containing culture medium were used as control of non-differentiated cells. (a) Quantitative real-time PCR showing variationsin the levels of the NPP3 transcript in differentiated cells, at different
treatment times. Values were normalized by the content of glycer-aldehyde-3-phosphate dehydrogenase transcript. Bars represent themean � SEM of three experiments performed in triplicate. **p < 0.01,***p < 0.001 (ANOVA with the Dunnett’s post hoc test). (b) Western blot
analysis of the NPP3 protein content in differentiated vs non-differen-
tiated N2a cells. The levels of a-tubulin were used as a loading control
and the ratio NPP3/a-tubulin was calculated. Ratio values wererepresented as a percentage of the ratio in non-differentiated cells,which was set as a 100%, and are shown in (c). Data are the
mean � SEM of three experiments in duplicate. ***p < 0.001 (ANOVA
with the Dunnett’s post hoc test). Representative photomicrographsshow NPP3 immunodetection (red) in non-differentiated (d) or differ-
entiated (after a 96-h treatment with DiBucAMP) N2a cells (g). MAP-2immunostaining was used as a marker of neuronal-like differentiation(e and h, green). Nuclei are counter-stained with 40,6-diamidino-2-phenylindole (blue). Merged images are shown (f and i). Scale
bars = 20 lm.
© 2014 International Society for Neurochemistry, J. Neurochem. (2014) 10.1111/jnc.12794
10 R. G�omez-Villafuertes et al.

activity with a broad substrate specificity is responsible forthe extracellular degradation of both ApnA and the fluoro-genic substrate. BODIPY-FL-GTPcS cleavage was alsoinhibited by the nucleoside monophosphate AMP. Theseresults point to a member of the E-NPP family as beingresponsible for the extracellular dinucleotide degradation inthe N2a cells. NPP1-NPP3 enzymes have been found to beable to hydrolyse pyrophosphate and phosphodiester bondsin a wide range of nucleotidic substrates, including ApnAwhich exhibited affinity values in the low micromolar range(Bollen et al. 2000; Vollmayer et al. 2003; Asensio et al.2007; Zimmermann et al. 2012). Nucleoside monophos-phates such as AMP are not hydrolysed but exert competitiveproduct inhibition on NPP reaction (Bollen et al. 2000;Vollmayer et al. 2003; Zimmermann et al. 2012). In addi-tion, the ectoenzymatic dinucleotide cleaving activity in N2acells exhibited biochemical features that are characteristic ofthe members of the E-NPP family (Bollen et al. 2000;Vollmayer et al. 2003; Zimmermann et al. 2012): (i) cata-lytic activity was dependent on divalent cations, beingstrongly inhibited by EDTA, (ii) activity was drasticallyreduced in the absence of both Ca2+ and Mg2+ ions, butenzyme reactivation was observed by the addition on Zn2+,(iii) finally, pH dependence studies revealed an alkalineoptimum pH for the enzyme activity.Fluorimetric enzyme assays were performed by replacing
the incubation medium by fresh Locke0s solution
immediately before the start of the experiments, whichavoids the participation of a secreted enzyme such as NPP2(Stefan et al. 2005, 2006; Zimmermann et al. 2012).Regarding NPP1 and NPP3, both could be detected in theN2a cells at the transcriptional and protein levels. Q-PCRanalysis showed that these two isozymes are expressed to asimilar extent in the neuroblastoma cells.E-NPP expression levels vary when N2a differentiate into
neuronal-like cells, which was achieved by incubating themin a medium with a low (0.5%) percentage of FBSi,supplemented with DiBucAMP. Both NPP1 and NPP3mRNA become significantly reduced at the early stages ofdifferentiation (after 24 h of treatment with DiBucAMP).However, there are differences in the time required for thesechanges to be translated into changes in the protein levels.NPP3 protein was dramatically reduced in the differentiatedN2a cells being almost completely absent after 24 h oftreatment with DiBucAMP. However, NPP1 protein levelshave only been partially reduced in the differentiated cellsafter 24 or 48 h of treatment with DiBucAMP, requiringlonger treatment times to be substantially decreased. Inaddition, the remaining NPP1 protein in the differentiatedcells seems to change its intracellular distribution from asomatic to a ‘neurite-like’ location. These differences mayreflect differences in the turnover of both NPP1 and NPP3proteins. Rapid fall in the levels of NPP3 protein when N2acells differentiate could make this protein a suitable surfacemarker for undifferentiated N2a cells. NPP1 (also known asPC-1) is expressed at limited stages of antibody-producingB-cell differentiation (Anderson et al. 1984), whereas NPP3seems to have a role in the differentiation and invasiveproperties of glial cells (Deissler et al. 1999). Regardingtumour cells, NPP3 has been suggested to be an early markerof cholangiocarcinoma, an adenocarcinoma derived frombiliary cells (Meerson et al. 1998). Moreover, NPP1 hasbeen shown to be expressed in astrocytic brain tumours, anda correlation was found between the up-regulated expressionof this ectoenzyme and the histological grade of theastrocytoma (Aerts et al. 2011b). Besides, cyclic AMP-dependent induction of differentiation inhibits NPP1 expres-sion in the rat C6 glioma cells (Aerts et al. 2011a). However,the physiological implications of changes in the expressionof E-NPP in the differentiated N2a cells remain to beelucidated.Changes in ectoenzymatic dinucleotide-hydrolysing activ-
ity seem to reflect variations in the levels of NPP1 protein, asthe activity is only partially reduced in differentiated cellsafter 24 h of treatment with DiBucAMP, being substantiallydecreased after 96 h: a pattern that quite exactly matcheschanges in NPP1 protein expression. However, N2a cells stillretain a significant capacity to hydrolyse dinucleotides inconditions, in which NPP3 protein is almost completely loss,i.e. after 24 h of treatment with DiBucAMP. These resultsindicate that the dinucleotide cleaving activity of N2a cells
10% FBSi
DiBucA
MP 24 h
DiBucA
MP 96 h
0102030405060708090
100
*
***
#A
ctiv
ity %
Fig. 7 Changes in the ectoenzymatic dinucleotide-cleaving activity indifferentiated neuro-2a (N2a) cells. Cells were cultured in 0.5% heat-inactivated foetal bovine serum (FBSi)-containing medium and treated
for 24 h or 96 h with 1 mM dibutyryl-cAMP (DiBucAMP). After that,hydrolysis of 10 lM BODIPY� FL guanosine 50-O-(3-thiotriphosphate)thioester (BODIPY-FL-GTPcS) was assayed in the differentiated cells.
Activities were represented as a percentage of those measured in cellsthat were incubated for 24 h in the usual 10% FBSi-containing culturemedium, which were used as control of non-differentiated cells. Data
are the mean � SEM of three experiments performed in triplicate.*p < 0.05, ***p < 0.001 versus non-differentiated cells; #p < 0.05differentiated cells after 24-h treatment versus differentiated cells after96-h treatment (ANOVA with the Bonferroni’s post hoc test).
© 2014 International Society for Neurochemistry, J. Neurochem. (2014) 10.1111/jnc.12794
Analysis of E-NPP activity in N2a cells 11

largely resides in NPP1, NPP3 contributing in a minor, ifany, extent to the extracellular degradation of dinucleotidesin these cells. However, it is necessary to take into accountthat both NPP1 and NPP3 can hydrolyse several mononu-cleotides in addition to dinucleotides. Moreover, theirspectrum of substrates is not restricted to nucleotidiccompounds, these enzymes being able to hydrolyse pyro-phosphate and phosphodiester bonds in a wide variety ofsubstrates (Bollen et al. 2000; Vollmayer et al. 2003;Zimmermann et al. 2012). Thus, actions of NPP3 onextracellular compounds, other than dinucleotides, cannotbe discarded. On the other hand, it is necessary to bear inmind that although the major functional role of ectonucle-otidases is the extracellular hydrolysis of mono- anddinucleotides and the production of extracellular nucleo-sides, thus regulating the ligand availability at nucleotideand adenosine receptors, some ectonucleotidases are mul-tifunctional proteins that can interact with the extracellularmatrix or even signal into the cell (Zimmermann et al.2012).
Acknowledgements and conflict of interestdisclosure
This work has been supported by research grants from Ministerio deCiencia e Innovaci�on (BFU2011-24743), the Spanish Ion ChannelInitiative (CSD2008-00005) and Fundaci�on Marcelino Bot�ın.
All experiments were conducted in compliance with the ARRIVEguidelines. The authors have no conflict of interest to declare.
References
Aerts I., Grobben B., Van Ostade X. and Slegers H. (2011a) Cyclic AMP-dependent down regulation of ecto-nucleotide pyrophosphatase/phosphodiesterase 1 (NPP1) in rat C6 glioma. Eur. J. Pharmacol.654, 1–9.
Aerts I., Martin J. J., De Deyn P. P., Van Ginniken C., Van Ostade X.,Kockx M., Dua G. and Slegers H. (2011b) The expression of ecto-nucleotide pyrophosphatase/phosphodiesterase 1 (E-NPP1) iscorrelated with astrocytic tumor grade. Clin. Neurol. Neurosurg.113, 224–229.
Albright R. A., Chang W. C., Robert D. et al. (2012) NPP4 is aprocoagulant enzyme on the surface of vascular endothelium.Blood 120, 4432–4440.
Albright R. A., Ornstein D. L., Cao W. et al. (2014) Molecular basis ofpurinergic signal metabolism by ectonucleotide pyrophosphatase/phosphodiesterases 4 and 1 and implications in stroke. J. Biol.Chem. 289, 3294–3306.
Anderson K. C., Bates M. P., Slaughenhoupt B., Schlossman S. F. andNadler L. M. (1984) A monoclonal antibody with reactivityrestricted to normal and neoplastic plasma cells. J. Immunol. 132,3172–3179.
Asensio A. C., Rodriguez-Ferrer C. R., Castaneyra-Perdomo A., OakninS. and Rotllan P. (2007) Biochemical analysis of ecto-nucleotidepyrophosphatase phosphodiesterase activity in brain membranesindicates involvement of NPP1 isoenzyme in extracellularhydrolysis of diadenosine polyphosphates in central nervoussystem. Neurochem. Int. 50, 581–590.
Bollen M., Gijsbers R., Ceulemans H., Stalmans W. and Stefan C.(2000) Nucleotide pyrophosphatases/phosphodiesterases on themove. Crit. Rev. Biochem. Mol. Biol. 35, 393–432.
Deissler H., Blass-Kampmann S., Bruyneel E., Mareel M. and RajewskyM. F. (1999) Neural cell surface differentiation antigen gp130(RB13-6) induces fibroblasts and glioma cells to express astroglialproteins and invasive properties. FASEB J. 13, 657–666.
Delicado E. G., Miras-Portugal M. T., Carrasquero L. M., Leon D.,Perez-Sen R. and Gualix J. (2006) Dinucleoside polyphosphatesand their interaction with other nucleotide signaling pathways.Pflugers Arch. 452, 563–572.
Dong H., Maddux B. A., Altomonte J., Meseck M., Accili D.,Terkeltaub R., Johnson K., Youngren J. F. and Goldfine I. D.(2005) Increased hepatic levels of the insulin receptor inhibitor,PC-1/NPP1, induce insulin resistance and glucose intolerance.Diabetes 54, 367–372.
Draganescu A., Hodawadekar S. C., Gee K. R. and Brenner C. (2000)Fhit-nucleotide specificity probed with novel fluorescent andfluorogenic substrates. J. Biol. Chem. 275, 4555–4560.
Fischer I., Shea T. B., Sapirstein V. S. and Kosik K. S. (1986)Expression and distribution of microtubule-associated protein 2(MAP2) in neuroblastoma and primary neuronal cells. Brain Res.390, 99–109.
Flores N. A., Stavrou B. M. and Sheridan D. J. (1999) The effects ofdiadenosine polyphosphates on the cardiovascular system.Cardiovasc. Res. 42, 15–26.
Gomez-Villafuertes R., del Puerto A., Diaz-Hernandez M., Bustillo D.,Diaz-Hernandez J. I., Huerta P. G., Artalejo A. R., Garrido J. J. andMiras-Portugal M. T. (2009) Ca2+/calmodulin-dependent kinase IIsignalling cascade mediates P2X7 receptor-dependent inhibition ofneuritogenesis in neuroblastoma cells. FEBS J. 276, 5307–5325.
Gualix J., Gomez-Villafuertes R., Pintor J., Llansola M., Felipo V. andMiras-Portugal M. T. (2014) Presence of diadenosinepolyphosphates in microdialysis samples from rat cerebellumin vivo: effect of mild hyperammonemia on their receptors.Purinergic Signal. 10, 349–356.
Hoyle C. H. V., Hilderman R. H., Pintor J. J., Schluter H. and King B. F.(2001) Diadenosine polyphosphates as extracellular signalmolecules. Drug Dev. Res. 52, 260–273.
Jankowski M., Piwkowska A., Rogacka D., Audzeyenka I., Janaszak-Jasiecka A. and Angielski S. (2011) Expression of membrane-bound NPP-type ecto-phosphodiesterases in rat podocytes culturedat normal and high glucose concentrations. Biochem. Biophys. Res.Commun. 416, 64–69.
Jimenez A. I., Castro E., Delicado E. G. and Miras-Portugal M. T.(2002) Specific diadenosine pentaphosphate receptor coupledto extracellular regulated kinases in cerebellar astrocytes.J. Neurochem. 83, 299–308.
Leon-Otegui M., Gomez-Villafuertes R., Diaz-Hernandez J. I., Diaz-Hernandez M., Miras-Portugal M. T. and Gualix J. (2011)Opposite effects of P2X7 and P2Y2 nucleotide receptors ona-secretase-dependent APP processing in Neuro-2a cells. FEBSLett. 585, 2255–2262.
Mateo J., Rotllan P., Marti E., Gomez De Aranda I., Solsona C. andMiras-Portugal M. T. (1997) Diadenosine polyphosphate hydrolasefrom presynaptic plasma membranes of Torpedo electric organ.Biochem. J. 323, 677–684.
Meerson N. R., Delautier D., Durand-Schneider A. M., Moreau A.,Schilsky M. L., Sternlieb I., Feldmann G. and Maurice M. (1998)Identification of B10, an alkaline phosphodiesterase of theapical plasma membrane of hepatocytes and biliary cells, in ratserum: Increased levels following bile duct ligation and duringthe development of cholangiocarcinoma. Hepatology 27,563–568.
© 2014 International Society for Neurochemistry, J. Neurochem. (2014) 10.1111/jnc.12794
12 R. G�omez-Villafuertes et al.

Miras-Portugal M. T., Gualix J., Mateo J., Diaz-Hernandez M., Gomez-Villafuertes R., Castro E. and Pintor J. (1999) Diadenosinepolyphosphates, extracellular function and catabolism. Prog.Brain Res. 120, 397–409.
Oaknin S., Rodriguez-Ferrer C. R., Aguilar J. S., Ramos A. and RotllanP. (2001) Receptor binding properties of di (1, N6-ethenoadenosine) 50, 50’’-P1, P4-tetraphosphate and itsmodulatory effect on extracellular glutamate levels in ratstriatum. Neurosci. Lett. 309, 177–180.
Oaknin S., Rodriguez-Ferrer C. R., Ramos A., Aguilar J. S. and Rotllan P.(2008)Bindingof50-O-(2-thiodiphosphate) to rat brainmembranes isprevented by diadenosine tetraphosphate and correlates with ecto-nucleotide pyrophosphatase phosphodiesterase 1 (NPP1) activity.Neurosci. Lett. 432, 25–29.
Pereira M. F., Hernandez M. D., Pintor J., Miras-Portugal M. T., CunhaR. A. and Ribeiro J. A. (2000) Diadenosine polyphosphatesfacilitate the evoked release of acetylcholine from rat hippocampalnerve terminals. Brain Res. 879, 50–54.
Pintor J., Torres M. and Miras-Portugal M. T. (1991) Carbachol inducedrelease of diadenosine polyphosphates–Ap4A and Ap5A–fromperfused bovine adrenal medulla and isolated chromaffin cells. LifeSci. 48, 2317–2324.
Pintor J., Diaz-Rey M. A., Torres M. and Miras-Portugal M. T. (1992a)Presence of diadenosine polyphosphates–Ap4A and Ap5A–in ratbrain synaptic terminals. Ca2+ dependent release evoked by 4-aminopyridine and veratridine. Neurosci. Lett. 136, 141–144.
Pintor J., Kowalewski H. J., Torres M., Miras-Portugal M. T. andZimmermann H. (1992b) Synaptic vesicle storage of diadenosinepolyphosphates in the Torpedo electric organ. Neurosci. Res.Commun. 10, 9–15.
Pintor J., Rotllan P., Torres M. and Miras-Portugal M. T. (1992c)Characterization and quantification of diadenosine hexaphosphatein chromaffin cells: granular storage and secretagogue-inducedrelease. Anal. Biochem. 200, 296–300.
Pintor J., Porras A., Mora F. and Miras-Portugal M. T. (1995) Dopaminereceptor blockade inhibits the amphetamine-induced release ofdiadenosine polyphosphates, diadenosine tetraphosphate anddiadenosine pentaphosphate, from neostriatum of the consciousrat. J. Neurochem. 64, 670–676.
Pintor J., Diaz-Hernandez M., Gualix J., Gomez-Villafuertes R.,Hernando F. and Miras-Portugal M. T. (2000) Diadenosine
polyphosphate receptors. from rat and guinea-pig brain to humannervous system. Pharmacol. Ther. 87, 103–115.
Raffaghello L., Chiozzi P., Falzoni S., Di Virgilio F. and Pistoia V.(2006) The P2X7 receptor sustains the growth of humanneuroblastoma cells through a substance P-dependentmechanism. Cancer Res. 66, 907–914.
Ramos A., Pintor J., Miras-Portugal M. T. and Rotllan P. (1995) Useof fluorogenic substrates for detection and investigation ofectoenzymatic hydrolysis of diadenosine polyphosphates: afluorometric study on chromaffin cells. Anal. Biochem. 228,74–82.
Rodriguez del Castillo A., Torres M., Delicado E. G. and Miras-PortugalM. T. (1988) Subcellular distribution studies of diadenosinepolyphosphates–Ap4A and Ap5A–in bovine adrenal medulla:presence in chromaffin granules. J. Neurochem. 51, 1696–1703.
Rodriguez-Pascual F., Torres M., Rotllan P. and Miras-Portugal M. T.(1992) Extracellular hydrolysis of diadenosine polyphosphates,ApnA, by bovine chromaffin cells in culture. Arch. Biochem.Biophys. 297, 176–183.
Stefan C., Jansen S. and Bollen M. (2005) NPP-typeectophosphodiesterases: unity in diversity. Trends Biochem. Sci.30, 542–550.
Stefan C., Jansen S. and Bollen M. (2006) Modulation of purinergicsignaling by NPP-type ectophosphodiesterases. Purinergic Signal.2, 361–370.
Vollmayer P., Clair T., Goding J.W., SanoK., Servos J. and ZimmermannH. (2003) Hydrolysis of diadenosine polyphosphates by nucleotidepyrophosphatases/phosphodiesterases. Eur. J. Biochem. 270, 2971–2978.
Wang Y., Chang C. F., Morales M., Chiang Y. H., Harvey B. K., Su T.P., Tsao L. I., Chen S. and Thiemermann C. (2003) Diadenosinetetraphosphate protects against injuries induced by ischemia and6-hydroxydopamine in rat brain. J. Neurosci. 23, 7958–7965.
Wu P. Y., Lin Y. C., Chang C. L., Lu H. T., Chin C. H., Hsu T. T.,Chu D. and Sun S. H. (2009) Functional decreases in P2X7receptors are associated with retinoic acid-induced neuronaldifferentiation of Neuro-2a neuroblastoma cells. Cell. Signal. 21,881–891.
Zimmermann H., Zebisch M. and Strater N. (2012) Cellular function andmolecular structure of ecto-nucleotidases. Purinergic Signal. 8,437–502.
© 2014 International Society for Neurochemistry, J. Neurochem. (2014) 10.1111/jnc.12794
Analysis of E-NPP activity in N2a cells 13




















