
DNA Damage-Sensing Kinases Mediate the Mouse 2-Cell Embryo's Response to Genotoxic Stress
12
BIOLOGY OF REPRODUCTION 85, 524–535 (2011) Published online before print 18 May 2011. DOI 10.1095/biolreprod.110.089334 DNA Damage-Sensing Kinases Mediate the Mouse 2-Cell Embryo’s Response to Genotoxic Stress 1 X.F. Mu, X.L. Jin, M.M.J. Farnham, Y. Li, and C. O’Neill 2 Human Reproduction Unit, Sydney Centre for Developmental and Regenerative Medicine, Kolling Institute of Medical Research, Sydney Medical School, University of Sydney, Sydney, New South Wales, Australia ABSTRACT A critical function of cells is the maintenance of their genomic integrity. A family of phosphoinositide-3-kinase-related protein kinases, which includes ataxia telangiectasia mutated (ATM) and ataxia telangiectasia and Rad3 related (ATR) kinases, play key roles in sensing DNA damage. ATM and ATR were demonstrated in the cleavage stages of mouse embryo development. Genotoxic stress was imposed by exposure to ultraviolet (UV) radiation (causes DNA strand breaks) or cisplatin (causes strand cross- links). UV irradiation or cisplatin treatment of 2-cell embryos in the G 2 phase of the cell cycle caused DNA damage as defined by increased phosphorylation of the H2A histone family, member X (H2AFX; previously H2AX) variant. UV irradiation caused a stable G 2 -M arrest, and cisplatin treatment allowed progression through mitosis followed by activation of a G 1 -S checkpoint. Both checkpoints were transformation-related protein 53- independent. Caffeine (inhibits both ATM and ATR), but not KU55933 (ATM-selective inhibitor), reversed the G 2 -M block induced by UV, inferring a primary role for ATR in sensing this form of DNA damage. Caffeine and KU55933 were equally effective in reversing the cisplatin-induced G 1 -S block, impli- cating ATM as the primary sensing enzyme. Breaching of either checkpoint by treatment with caffeine or KU55933 allowed embryos to progress through several further cell cycles, yet none developed to blastocysts. The results show, to our knowledge for the first time, that the G 2 -M and G 1 -S cell-cycle checkpoints in the early embryo are differentially regulated by ATM and ATR in response to genotoxic stress and that they act as an initial point for containment of genomic damage. Under conditions of extensive or persistent DNA damage, the demise of the embryo is the ultimate method of protecting genomic integrity. apoptosis, ataxia telangiectasia and Rad3 related, ataxia telangiectasia mutated, cell cycle, checkpoint, DNA damage, early development, embryo, H2AFX, transformation-related protein 53, tumor suppressor gene INTRODUCTION A critical function for all cells is monitoring and maintaining the integrity of the genome. Structural damage or alterations to DNA are generally mutagenic, and the propagation of such damaged cells within a lineage may compromise the viability of the organism. If DNA damage occurs within cells of the early embryo, the consequences can be particularly severe, because their perpetuation can poten- tially affect all cells in the developing organism, including the germline. Key to the management of genomic integrity is the detection of DNA damage. A family of phosphoinositide-3- kinase-related protein kinases (PIKKs) plays key roles in sensing DNA damage [1]. Foremost within this family are the tumor suppressor proteins ataxia telangiectasia mutated (ATM) and ataxia telangiectasia and Rad3 related (ATR) kinases. The aim of the present study was to assess the role that ATM and ATR play in sensing DNA damage in the early mouse embryo and the nature of the embryo’s response to this damage. Recruitment of the PIKK family of kinases to sites of DNA damage causes activation of their kinase activity, which in turn activates downstream effectors of cell-cycle regulation or apoptosis. PIKKs are recruited to sites of damage by specific partner proteins. For example, ATR-interacting protein forms a partnership with ATR [2], and the meiotic recombination 11 homolog A complex (formerly known as Mre11) partners with ATM [3]. The minor histone 2A variant, H2A histone family, member X (H2AFX), is an important substrate for the PIKKs. The phosphorylated (serine-129) form of this histone is commonly referred to as gamma-H2AFX. Its phosphorylation in response to DNA damage is very efficient and results in accumulation of H2AFX for many megabase pairs (Mbp) on either side of the lesion, making immunostaining for gamma- H2AFX the most sensitive measure of damage [4, 5]. Gamma- H2AFX serves as a key component in the DNA damage repair response by loosening the conformation of DNA and, thus, facilitating recruitment of the repair enzymes to the lesion. It also recruits molecules, such as cohesion molecules, that provide structural integrity during the repair process [4, 5]. This combination of proteins acts to sense the DNA damage and amplify the DNA damage signal, and this results in the recruitment of the required effectors. A canonical response of cells to DNA damage is the activation of cell-cycle checkpoints [6]. Checkpoints provide a mechanism whereby cells are prevented from progressing past specific points within the cell cycle if conditions such as DNA damage exist. Two important checkpoints are the G 1 -S and G 2 - M checkpoints. ATM is most commonly implicated in the activation of the G 1 -S checkpoint [1], whereas ATR is active in the induction of the G 2 -M checkpoint [7]. It is becoming increasingly recognized, however, that some overlap and redundancy occurs between the action of these two kinases at these checkpoints [1]. The checkpoints also provide an opportunity for repair of DNA before cell division. Where DNA damage is severe or irreparable, cell death, typically via apoptosis, may also result. ATM and ATR can mediate this response by activation of mediators such as transformation- related protein 53 (TRP53; also known as P53) [8, 9], which can induce BCL2-associated X protein (BAX)-mediated apoptosis. 1 Supported by grants from the Australian National Health and Medical Research Council to C.O. 2 Correspondence: FAX: 61 2 9926 6343; e-mail: [email protected] Received: 25 October 2010. First decision: 23 November 2010. Accepted: 13 April 2011. Ó 2011 by the Society for the Study of Reproduction, Inc. eISSN: 1529-7268 http://www.biolreprod.org ISSN: 0006-3363 524 Downloaded from www.biolreprod.org.
Transcript of DNA Damage-Sensing Kinases Mediate the Mouse 2-Cell Embryo's Response to Genotoxic Stress

BIOLOGY OF REPRODUCTION 85, 524–535 (2011)Published online before print 18 May 2011.DOI 10.1095/biolreprod.110.089334
DNA Damage-Sensing Kinases Mediate the Mouse 2-Cell Embryo’s Responseto Genotoxic Stress1
X.F. Mu, X.L. Jin, M.M.J. Farnham, Y. Li, and C. O’Neill2
Human Reproduction Unit, Sydney Centre for Developmental and Regenerative Medicine, Kolling Institute of MedicalResearch, Sydney Medical School, University of Sydney, Sydney, New South Wales, Australia
ABSTRACT
A critical function of cells is the maintenance of their genomicintegrity. A family of phosphoinositide-3-kinase-related proteinkinases, which includes ataxia telangiectasia mutated (ATM) andataxia telangiectasia and Rad3 related (ATR) kinases, play keyroles in sensing DNA damage. ATM and ATR were demonstratedin the cleavage stages of mouse embryo development. Genotoxicstress was imposed by exposure to ultraviolet (UV) radiation(causes DNA strand breaks) or cisplatin (causes strand cross-links). UV irradiation or cisplatin treatment of 2-cell embryos inthe G
2phase of the cell cycle caused DNA damage as defined by
increased phosphorylation of the H2A histone family, member X(H2AFX; previously H2AX) variant. UV irradiation caused astable G
2-M arrest, and cisplatin treatment allowed progression
through mitosis followed by activation of a G1-S checkpoint.
Both checkpoints were transformation-related protein 53-independent. Caffeine (inhibits both ATM and ATR), but notKU55933 (ATM-selective inhibitor), reversed the G
2-M block
induced by UV, inferring a primary role for ATR in sensing thisform of DNA damage. Caffeine and KU55933 were equallyeffective in reversing the cisplatin-induced G
1-S block, impli-
cating ATM as the primary sensing enzyme. Breaching of eithercheckpoint by treatment with caffeine or KU55933 allowedembryos to progress through several further cell cycles, yet nonedeveloped to blastocysts. The results show, to our knowledge forthe first time, that the G
2-M and G
1-S cell-cycle checkpoints in
the early embryo are differentially regulated by ATM and ATR inresponse to genotoxic stress and that they act as an initial pointfor containment of genomic damage. Under conditions ofextensive or persistent DNA damage, the demise of the embryois the ultimate method of protecting genomic integrity.
apoptosis, ataxia telangiectasia and Rad3 related, ataxiatelangiectasia mutated, cell cycle, checkpoint, DNA damage,early development, embryo, H2AFX, transformation-relatedprotein 53, tumor suppressor gene
INTRODUCTION
A critical function for all cells is monitoring andmaintaining the integrity of the genome. Structural damageor alterations to DNA are generally mutagenic, and thepropagation of such damaged cells within a lineage maycompromise the viability of the organism. If DNA damage
occurs within cells of the early embryo, the consequences canbe particularly severe, because their perpetuation can poten-tially affect all cells in the developing organism, including thegermline. Key to the management of genomic integrity is thedetection of DNA damage. A family of phosphoinositide-3-kinase-related protein kinases (PIKKs) plays key roles insensing DNA damage [1]. Foremost within this family are thetumor suppressor proteins ataxia telangiectasia mutated (ATM)and ataxia telangiectasia and Rad3 related (ATR) kinases. Theaim of the present study was to assess the role that ATM andATR play in sensing DNA damage in the early mouse embryoand the nature of the embryo’s response to this damage.
Recruitment of the PIKK family of kinases to sites of DNAdamage causes activation of their kinase activity, which in turnactivates downstream effectors of cell-cycle regulation orapoptosis. PIKKs are recruited to sites of damage by specificpartner proteins. For example, ATR-interacting protein forms apartnership with ATR [2], and the meiotic recombination 11homolog A complex (formerly known as Mre11) partners withATM [3]. The minor histone 2A variant, H2A histone family,member X (H2AFX), is an important substrate for the PIKKs.The phosphorylated (serine-129) form of this histone iscommonly referred to as gamma-H2AFX. Its phosphorylationin response to DNA damage is very efficient and results inaccumulation of H2AFX for many megabase pairs (Mbp) oneither side of the lesion, making immunostaining for gamma-H2AFX the most sensitive measure of damage [4, 5]. Gamma-H2AFX serves as a key component in the DNA damage repairresponse by loosening the conformation of DNA and, thus,facilitating recruitment of the repair enzymes to the lesion. Italso recruits molecules, such as cohesion molecules, thatprovide structural integrity during the repair process [4, 5].This combination of proteins acts to sense the DNA damageand amplify the DNA damage signal, and this results in therecruitment of the required effectors.
A canonical response of cells to DNA damage is theactivation of cell-cycle checkpoints [6]. Checkpoints provide amechanism whereby cells are prevented from progressing pastspecific points within the cell cycle if conditions such as DNAdamage exist. Two important checkpoints are the G
1-S and G
2-
M checkpoints. ATM is most commonly implicated in theactivation of the G
1-S checkpoint [1], whereas ATR is active in
the induction of the G2-M checkpoint [7]. It is becoming
increasingly recognized, however, that some overlap andredundancy occurs between the action of these two kinases atthese checkpoints [1]. The checkpoints also provide anopportunity for repair of DNA before cell division. WhereDNA damage is severe or irreparable, cell death, typically viaapoptosis, may also result. ATM and ATR can mediate thisresponse by activation of mediators such as transformation-related protein 53 (TRP53; also known as P53) [8, 9], whichcan induce BCL2-associated X protein (BAX)-mediatedapoptosis.
1Supported by grants from the Australian National Health and MedicalResearch Council to C.O.2Correspondence: FAX: 61 2 9926 6343;e-mail: [email protected]
Received: 25 October 2010.First decision: 23 November 2010.Accepted: 13 April 2011.� 2011 by the Society for the Study of Reproduction, Inc.eISSN: 1529-7268 http://www.biolreprod.orgISSN: 0006-3363
524
Dow
nloaded from w
ww
.biolreprod.org.

Damage to DNA can take many forms, including single- ordouble-strand breaks, inter- and intrastrand DNA cross-links,DNA-protein cross-links, and chemical modification, such asgeneration of 8-hydroxydeoxyguanosine residues or polycyclicaromatic hydrocarbon adducts. ATR is thought to be mostimportant for sensing single-strand DNA breaks, whereas ATMis most involved in sensing double-strand breaks [3]. Sensingof interstrand cross-linking defects may involve ATR, but notATM, in somatic cells [10]. A consensus is forming thatimplicates considerable, but not complete, overlap in thefunctions of ATM and ATR in somatic cells [11]. Somenonredundant actions, however, are powerfully demonstratedby the observations that Atr�/� embryos are nonviable by thelate blastocyst stage [12, 13] but that Atm�/� embryos areviable [14]. This indicates fundamental functions of Atr in earlydevelopment that are not compensated for by the presence ofthe Atm gene.
X-irradiation of the zygote or 2-cell mouse embryo inducesa G
2-M arrest [15]. This type of arrest was ameliorated by
caffeine, an inhibitor of both ATR and ATM [16]. X-irradiatedembryos that passed the arrested phase showed increasedmicronuclei formation and reduced developmental viability[17–19]. Drug- and ultraviolet light (UV) irradiation-inducedDNA damage has been less widely investigated; however, UVexposure has been shown to cause increased rates of sisterchromatid exchange [20] and single-strand breaks (hamster)[21]. Exposure of mouse zygotes to UV radiation resulted inunscheduled DNA synthesis, which is interpreted as evidencefor activation of DNA repair processes [22]. Exposure of spermto UV radiation did not prevent fertilization; however, theresulting embryos (bovine) tended to block at the 2-cell stage[23]. X-irradiation of sperm also results in developmental arrestbecause of the activation of a Trp53-dependent, S-phasecheckpoint in the resulting mouse zygotes [24, 25]. Embryostreated in this manner also activated a cyclin-dependent kinase(CDK) inhibitor 1A-dependent cell-cycle arrest at the morula/blastocyst stage.
Caffeine is effective in relieving the cell-cycle blocksinduced by a range of genotoxic stresses in vitro. It achievesthis by inhibiting the phosphorylation and activation ofmammalian PIKKs [26], but it is not a global inhibitor ofother protein kinase activities. The PIKK family of proteinkinases also includes mechanistic target of rapamycin (serine/threonine kinase; MTOR) and protein kinase, DNA-activated,catalytic polypeptide (PRKDC). Caffeine has relatively poorselectivity for PRKDC. It does act on MTOR, but a selectiveMTOR inhibitor (rapamycin) does not influence checkpointblocks induced by irradiation of cells, arguing that MTOR isnot a major caffeine-sensitive effector of these checkpoints[27]. Caffeine is also a well-known inhibitor of cAMP-phosphodiesterase [28], yet specific inhibition of cAMP-phosphodiesterase and increased cellular cAMP concentrationactually increase the radiosensitivity of cells, providing nosupport for a role of phosphodiesterase inhibition in the reliefof checkpoint action in cells with damaged DNA [29]. Thus, itis generally recognized that the ability of caffeine to allow cellswith damaged DNA to bypass checkpoints is primarily theresult of its capacity to inhibit ATR and ATM.
Recently, a highly selective competitive inhibitor of ATMhas been developed (KU55933) [30]. This drug has high ATMspecificity with respect to inhibition of other PIKKs, preventsphosphorylation of selective ATM targets following DNAdamage, and prevents the normal ATM-dependent cell-cyclecheckpoint responses to ionizing radiation [26, 30]. Theselectivity of KU55933 for ATM means that when used inconjunction with caffeine, the relative roles of ATM (caffeine-
and KU55933-sensitive) and ATR (caffeine-sensitive andKU55933-insensitive) in response to defined genotoxic stresscan be assigned.
Most studies of the early embryo to date have used X-irradiation as a source of genotoxic stress; however, exposureof embryos to chemicals or UV radiation seems to be a form ofstress that the embryo is more likely to encounter, particularlywith the advent of assisted reproductive technologies. In thepresent study, 2-cell mouse embryos were subjected to twodifferent forms of genomic stress: UV irradiation, whichtypically induces DNA single-strand breaks and is thereforeexpected to be sensed by ATR, and cisplatin (a chemothera-peutic drug), which induces inter- and intrastrand DNA cross-links and DNA-protein cross-links and is therefore a likelytarget for ATM. We provide evidence for different responsesby the embryo to these distinct forms of DNA damage and thedifferential involvement of ATR and ATM in the embryo’sresponse to each. To our knowledge, the present study providesthe first analysis of the relative roles of key DNA damagesensor kinases in monitoring the integrity of the embryonicgenome and directing the embryo’s response to this damage.
MATERIALS AND METHODS
Animals
The use of animals in the present study was in accordance with theAustralian Code of Practice for the Care and Use of Animals for ScientificPurpose and was approved by the Institutional Animal Care and EthicsCommittee. C57BL6J mice were housed and bred in the Gore Hill ResearchLaboratory (St Leonards, NSW, Australia) under a 12L:12D photoperiod andhad access to food and water ad libitum. Female mice (age, 4–6 wk) weresuperovulated by intraperitoneal injection of 5 IU of equine chorionicgonadotropin (Folligon; Intervet International), followed 48 h later by 5 IUof human chorionic gonadotropin (hCG; Chorulon; Intervet). Females werepaired with males of proven fertility. Pregnancy was confirmed by the presenceof a copulation plug the following morning (Day 1).
Mouse Embryo Collection and Culture
Two-cell embryos were cultured in modified-human tubal fluid medium(mod-HTF) after being flushed from the oviduct with Hepes-buffered mod-HTF [31]. All components of the media were tissue-culture grade (SigmaChemical Co.), and all media contained 3 mg/ml of bovine serum albumin(Sigma) unless otherwise stated. Embryos were cultured in groups of 10 in 10-ll volumes of mod-HTF overlaid by approximately 2 mm of heavy paraffin oil(Sigma) at 378C in 5% CO
2in air for the periods indicated in individual
experiments. The developmental stage and morphology of embryos wereassessed by visualizing the embryos with an inverted phase-contrastmicroscope (Diaphot; Nikon) at 24-h intervals after collection and treatment.
Immunofluorescence
Embryos were washed three times in PBS with 0.1% (w/v) bovine serumalbumin, 0.1% (w/v) Tween-20 (Sigma), and 0.2% (w/v) sodium azide(washing buffer), then fixed with freshly prepared 2% paraformaldehyde (w/v;Sigma) in PBS (pH 7.4) for 30 min, and then permeabilized with 2% (w/v)paraformaldehyde with 0.3% Tween-20 at room temperature for 30 min.Embryos were washed three times in washing buffer and then blocked in PBScontaining 2% (w/v) bovine serum albumin and 30% (v/v) serum for 3 h. Next,they were stained overnight at 48C with primary antibodies: rabbit anti-ATMantibody (sc-7230; Santa Cruz Biotechnology) at 4 lg/ml, goat polyclonal anti-ATR (sc-1887; Santa Cruz Biotechnology) at 2 lg/ml, and rabbit anti-polyclonal gamma-H2AFX (phosphor-serine 139; ab2893; Abcam) and anti-mouse monoclonal phosphorylated ATM (pATM; phosphorylated serine 1981;ab36810; Abcam) at 4 lg/ml. Each antibody was prepared in PBS with 2%(w/v) bovine serum albumin. Equivalent concentrations of nonimmuneimmunoglobulin (Ig) G were used as negative controls. Detection withfluorescently labeled secondary antibodies was achieved by labeling at roomtemperature for 1 h.
Whole-section imaging was performed with mercury lamp UV illuminationand epifluorescence on a Nikon Optiphot microscope with a DPlan Apo 40 UVobjective. Images were subjected to deconvolution using Image-Pro Plus with
SENSING DNA DAMAGE IN EMBRYOS 525
Dow
nloaded from w
ww
.biolreprod.org.

Sharpstack deconvolution software (Media Cybernetics, Inc.). Opticalsectioning was performed with a Bio-Rad Radiance Confocal microscopeusing a Nikon Plan Apo 60X/1.4 oil-emersion objective as previously described[32]. Images were captured using Lasersharp 2000 software. Microscope andlaser settings were adjusted such that no fluorescence was observed withnonimmune controls. All test specimens were observed with these samesettings. Quantitative analysis of staining was achieved using the area-of-interest marking tool within Image-Pro Plus image analysis software andexpressed as arbitrary units of optical density.
Western Blot Analysis
Western blot analysis was performed as previously described [33]. Freshhybrid 2-cell embryos were collected from reproductive tracts 40 h post-hCG,washed three times in cold PBS, and transferred into 1.5 ll of extraction buffersupplemented with protease and phosphatase inhibitors (23 PBS, 2% [v/v]Triton X-100, 24 mM deoxycholic acid, 0.2% [w/v] SDS, 20 mM NaF, 20 mMNa
4P
2O
7, 2 mM PMSF, 3.08 lM aprotinin, 42 lM leupeptin, and 2.91 lM
Pepstatin A; all from Sigma). The embryos were lysed by three cycles offreezing in liquid nitrogen and thawing (with vortexing). Protein samples werediluted with 1 ll of 53 Laemmli buffer (50 mM Tris-HCl, 5 mMethylenediaminetetra-acetic acid [pH 8.0], 12.5% [w/v] SDS, 0.05% [w/v]bromophenol blue, and 25% beta-mercaptoethanol), incubated for 5 min at958C, and size separated on PhastGel Gradient 4-15 (catalog no. 170678-01;GE Healthcare) using a PhastSystem electrophoresis apparatus (PhastSystemseparation and control unit; Amersham Pharmacia Biotech). The proteins wereblotted onto polyvinylidene fluoride transfer membrane (Hybond-P; GEHealthcare) with transfer buffer containing 12 mM Tris (Sigma), 96 mMglycine (BDH), 0.1% (w/v) SDS, and 10% (v/v) methanol (BDH) using a PhastTransfer system. The membrane was incubated in 10 ml of blocking buffercontaining 2.5% (w/v) skim milk powder (Diploma). The primary antibodieswere the same as used for immunofluorescence staining at a concentration of0.4 lg/ml for each. They were detected with 1:5000 horseradish peroxidase-conjugated secondary antibody (Sigma). The membrane was incubated inSuperSignal West Femto (Pierce) diluted 1:4 and analyzed by chemilumines-cence (Las 4000; Fuji), and HiMark Pre-Stained Standards (Invitrogen) wereused to assess molecular mass. Analyses were performed on groups of 90embryos for each antigen.
Treatments
The ATM inhibitor, 2-morpholin-4-yl-6-thianthren-1-yl-pyran-4-one
(KU55933; gift from Dr. Graeme Smith, KuDOS Pharmaceuticals Ltd.) [30];
the ATR and ATM inhibitor, 1,3,7-trimethylxanthine (caffeine; Sigma); the 1-
o-phosphatidylinositol-3-kinase inhibitor, (2-(4-morpholinyl)-8-phenyl-4H-1-
benzopyran-4-one (LY294002; Calbiochem); the TRP53 inhibitor, 2-(2-imino-
4,5,6,7-tetrahydrobenzothiazol-3-yl)-1-p-tolylethanone (pifithrin-a; Sigma);
and cis-Diammineplatinum(II) dichloride (cisplatin; P4394; Sigma). UV
irradiation was achieved by placing embryos in 1 ml of Hepes-buffered mod-
HTF media in an open, 35-mm, plastic Petri dish. UV irradiation (253.7 nm)
equated to a dose of 1.95 J/cm2. Changes in UV dosage are not readily
calibrated or validated in a biological laboratory; hence, it is common to change
exposure by varying the time of exposure to a standard dose. This strategy has
been adopted in this laboratory. After exposure, embryos were transferred to
mod-HTF, with or without other drugs, in 10-ll drops under mineral oil
(Sigma). Cisplatin treatment was performed as a conventional dose-response
study.
TUNEL Assay
The TUNEL assay was used to detect apoptosis (fluorescein; In Situ Death
Detection Kit; Roche). Embryos were fixed and permeabilized as described
above and incubated in the TUNEL reaction mixture supplied with the kit for 1
h at 378C in the dark. Negative controls lacked terminal deoxynucleotidyl
transferase, and positive controls were embryos incubated in DNase I (50 U/ml;
Sigma) for 10 min at 378C to induce DNA strand breaks. Embryos were
washed three times in PBS, stained with 20-(4-ethoxyphenyl)-5-(4-methyl-1-
piperazinyl)-2,50-bi-1H-benzamidazole trihydrochloride (4 lg/ml; bisBenzi-
mide; Sigma) and then mounted. The total number of nuclei was enumerated by
the bisBenzimide staining, whereas the fluorescein TUNEL staining identified
those nuclei undergoing apoptosis.
DNA Synthesis
Embryos were incubated in media supplemented with 20 lM 5-bromo-20-
deoxyuridine (BrdU; Sigma). They were then fixed and stained using the
FIG. 1. Immunolocalization and Westernblot analysis of ATM and ATR in thepreimplantation mouse embryo. Embryoswere collected from the reproductive tractand immediately fixed and stained. In eachcase, staining with IgG control antisera wasundetectable, and the microscope andimaging settings were the same for eachstage of development for a given antibody.The images shown are representative of atleast five embryos in each of three repli-cates. A-C) Whole-section images of ATMstaining of 2-cell, 8-cell, and blastocyst-stage embryos, respectively. D-F) Whole-section images of ATR staining of 2-cell, 8-cell, and blastocyst-stage embryos, respec-tively. G) A single equatorial confocalsection of ATR stained 2-cell embryo. H) Atwo-dimensional reconstruction of all sec-tions through the embryo shown in G. Bar¼ 10 lm. I) Representative Western blot ofATM and ATR with molecular mass ofmarkers shown with arrows.
526 MU ET AL.
Dow
nloaded from w
ww
.biolreprod.org.

immunolocalization technique described above with a 1:200 dilution of anti-BrdU antibody (Sigma).
Statistical Analysis
Statistical analyses were performed with the SPSS software (Version 11.5;SPSS). The proportion of embryos developing to a given developmentallandmark following culture in vitro or the proportion of embryonic cells wasassessed by binary logistic regression analysis. The proportion developing to agiven landmark or undergoing apoptosis was treated as the dichotomousdependent variable and the treatment and experimental replicate as covariates inthe model.
The effects of treatments on the number of cells and the number of TUNEL-staining cells in blastocysts as well as the intensity of immunostaining wereassessed using univariate regression analysis within the general linear model.When appropriate, the statistical model incorporated the effect of treatmentwith the number of cells as the dependent variable and treatment as fixed-factormain effects. Full factorial analysis was performed with significant factorcontrasts to assess the difference between individual treatments.
RESULTS
Immunolocalization of ATM (Fig. 1, A–C) and ATR (Fig.1, D–F) in embryos collected directly from the reproductivetract showed that both antigens were present in the 2-cell and 8-cell embryos. Blastocysts stained intensely for ATM butshowed negligible ATR staining (Fig. 1F). In whole-embryosections, nuclear accumulation of ATM was evident, althoughit was less clear whether this was also the case for ATR. Thus,single equatorial confocal sections of ATR staining wereimaged in fresh 2-cell embryos. These showed some nuclearaccumulation of ATM staining (Fig. 1G), which had arelatively uniform distribution, except that it was reducedwithin the presumptive nucleoli. Cytoplasmic staining showeda heterogeneous granular pattern, which was more intense inthe cortical regions. A two-dimensional reconstruction ofconfocal sections through the depth of the embryo showedextensive distribution of these ATR-staining granules (Fig.1H). The nuclear localization of ATM and ATR in embryosthat were assumed to be normal and not subject to genotoxicstress was surprising. To ensure that it was not an acuteresponse to the process of collecting and preparing embryos, 2-cell embryos were collected again but flushed from the oviductwith fixative (paraformaldehyde) to minimize any stressresponse before fixation. These embryos showed the samepattern of staining as those subjected to conventional collectionprocedures (not shown). Western blot analyses (Fig. 1I) ofextracts of 2-cell embryos (using the same antibodies used forimmunolocalization) showed a single dominant high-molecu-lar-weight band for each of the ATM and ATR antibodies,confirming their suitability for immunolocalization studies.
The next component of the study was to assess the responseof embryos to two different forms of genotoxic stress. Thedesign of the experiment was to expose 2-cell embryos to thegenotoxic stressors during the G
2phase of the cell cycle. If the
stress caused a G2-M cell-cycle checkpoint, then a 2-cell block
was expected. If the block occurred at the G1-S checkpoint,
then progression of embryos through the 2-cell stage with asubsequent block in the 4-cell stage was expected. Todetermine the time that 2-cell embryos completed the S phaseand had thus entered the G
2phase, cell-cycle analysis was
performed by BrdU incorporation. Zygotes were collected at29 h post-hCG and cultured to 50 h post-hCG in mediasupplemented with BrdU at progressive times after culture wasinitiated. Addition of BrdU commenced 37 h post-hCG and at1.5-h intervals thereafter (at least 25 embryos were analyzed ateach time point). Immunolocalization of BrdU was performedin the resulting 2-cell embryos, and nuclei were categorized aseither with or without BrdU incorporation. When incubation
with BrdU commenced before 40 h post-hCG, many cells(.50%) incorporated BrdU, but by 43 h post-hCG, no cellsincorporated BrdU. Based on this analysis, we concluded that2-cell embryos collected 44 h post-hCG were within the G
2phase, and this was the time point used for further analyses.
We next investigated the capacity of UV irradiation orcisplatin treatment to induce cell-cycle blocks in the earlyembryo. Two-cell embryos (44 h post-hCG) were treated withincreasing exposure times to UV radiation or increasingcisplatin concentration. Increasing the time of exposure of 2-cell mouse embryos to UV radiation caused a progressiveincrease in the proportion of embryos that failed to progressthrough the cell cycle (2-cell block) 24 h after treatment (P ,0.001) (Fig. 2A). UV exposure for 4 min prevented all embryosfrom undergoing mitosis. Treatment of embryos with anincreasing concentration of cisplatin for 4 h caused only a smallproportion to block at the 2-cell stage, and this proportion
FIG. 2. Development of 2-cell embryos in culture after exposure togenotoxic stress. Two-cell embryos were collected from the reproductivetract and immediately exposed to different levels of two genotoxic stress,UV irradiation and cisplatin treatment. After exposure, the embryos werewashed and placed in culture for (A) 24 h, after which the proportion ofembryos that had developed past the 2-cell stage was recorded, or (B) 48h, after which the proportion of embryos that developed past the 4-cellstage was recorded. The results represent three independent replicates,with at least 30 embryos per treatment per replicate.
SENSING DNA DAMAGE IN EMBRYOS 527
Dow
nloaded from w
ww
.biolreprod.org.

plateaued as the concentration increased (P , 0.001 vs. UVtreatment) (Fig. 2A).
Examination of the proportion of embryos that progressedpast the 4-cell stage after 48 h of culture showed the UV-treated embryos that bypassed the 2-cell block were mainlyblocked at the 4-cell stage (Fig. 2B). A strong cisplatin dose-dependent block to progression past the 4-cell stage wasobserved. More than 80% of 2-cell embryos did not progresspast the 4-cell stage after treatment with 12 lM cisplatin (Fig.2B).
Phosphorylation of H2AFX (gamma-H2AFX) is a highlysensitive marker of damage induced by a range of genotoxicstressors. Two-cell embryos were immunostained for this
marker at various times after UV irradiation (Fig. 3A) orcisplatin treatment (Fig. 3B). Untreated 2-cell embryos showedlittle detectable gamma-H2AFX. The amount of nuclearstaining increased with the time that embryos were exposedto UV at each of four assessed time points after treatment (1–4h). Exposure of embryos to cisplatin also caused an increase ingamma-H2AFX staining, but this was only observed consis-tently after 4 h of exposure. The results show that both UV andcisplatin treatment caused DNA damage in the 2-cell embryo.
Severe genotoxic stress might conceivably cause the acuteonset of cell death by apoptosis. Cohorts of embryos treatedwith the highest UV exposure times and the highestconcentration of cisplatin were subjected to TUNEL staining
FIG. 3. The phosphorylation of H2AFX inresponse to genotoxic stress in the 2-cellembryo. Immunofluorescence images of 2-cell embryos stained for gamma-H2AFXafter their treatment with (A) UV radiationor (B) cisplatin. Embryos were exposed toUV radiation for 0, 1, 2, or 4 min andstained after 1, 2, or 4 h in culture. Cisplatintreatment was at a concentration of 0, 3, 6,or 12 lM. The images are representative ofresponse for each treatment. Nonimmunesera resulted in no detectable staining. Bar¼ 10 lm.
528 MU ET AL.
Dow
nloaded from w
ww
.biolreprod.org.

24 h after treatment. No UV-treated or control embryos wereTUNEL positive (n ¼ 25 for each). Cisplatin-treated embryosthat were blocked at the 4-cell stage showed only a smallnumber of apoptotic cells (5% 6 2% of cells, mean 6 SEM, n¼ 15 embryos). During TUNEL staining, the nucleus was alsostained with bisBenzimide; this showed that the blocks inducedby both UV irradiation and cisplatin treatment were notassociated with the entrapment of cells at the metaphase stage.To assess whether the 4-cell block induced by cisplatinoccurred before cells entering the next round of DNA synthesis(S phase), control or cisplatin (12 lM)-treated embryos werecultured in the presence of BrdU and then tested forincorporation by immunolocalization. All cells in controlembryos showed levels of BrdU incorporation consistent withprogression through the S phase, but this was only the case for7% 6 4% (mean 6 SEM) of cells in embryos treated withcisplatin (n¼15 embryos in each case). These results show thatG
2-stage embryos exposed to UV radiation were blocked
before entering metaphase, and we therefore conclude that UVirradiation activated a G
2-M checkpoint. Cisplatin treatment
did not block cells from progressing through metaphase but,rather, induced a block before the next S phase; therefore, it canbe concluded to have induced a G
1-S checkpoint.
To determine whether the embryos at the 2-cell (UVtreatment) and 4-cell (cisplatin treatment) stage were blockedrather than transiently delayed, their fate was followed over 72h in culture. None of embryos blocked at the 2-cell stage afterUV exposure progressed past this stage after 72 h of culture.Most of the embryos exposed to cisplatin were at the 4-cellstage 24 h after treatment (Fig. 4A). At 48 h (Fig. 4B) or 72 h(Fig. 4C), this block persisted, with only a small increase in thenumber of embryos progressing to the 8-cell stage (P , 0.01)and none progressing past this stage. Over this time, anincrease (P , 0.01) was observed in the proportion of embryosshowing morphological degeneration (fragmentation). It isconcluded that the G
1-S checkpoint activated by cisplatin and
the G2-M checkpoint activated by UV radiation were relatively
stable over at least 72 h.
FIG. 4. The rate of development of embryos treated with cisplatin over72 h in culture. Two-cell embryos recovered from the reproductive tractwere immediately exposed to control media or media supplemented with12 lM cisplatin. After washing, they were cultured in media for 72 h. Theproportion of embryos reaching each developmental landmark after (A) 24h, (B) 48 h, and (C) 72 h is shown. The results represent a minimum of 90embryos in each treatment group.
FIG. 5. The effect of inhibitors of ATM and ATR on the developmentalblocks induced by UV irradiation and cisplatin treatment of 2-cellembryos. Embryos were treated with (A) UV radiation (4 min) or (B)cisplatin (12 lM) in the presence of increasing concentrations of caffeineor KU55933. After extensive washing, embryos were cultured in mediawith the same concentration of caffeine or KU55933 for 24 h (UVirradiation) or 48 h (cisplatin treatment), and the proportion achievingdevelopmental landmarks was recorded. The results represent a minimumof 30 embryos per treatment in each of three replicates.
SENSING DNA DAMAGE IN EMBRYOS 529
Dow
nloaded from w
ww
.biolreprod.org.

H2AFX is a substrate for ATM and ATR, and theobservation of the induction of gamma-H2AFX staining inthe nuclei of 2-cell embryos exposed to UV radiation andcisplatin (Fig. 3) implicates the activation of these PIKKs as acomponent of the embryo’s response to these stressors. Therole of ATM and ATR in the developmental blocks induced byUV irradiation or cisplatin treatment was assessed by the use oftheir inhibitors, caffeine and/or KU55933. Caffeine causeddose-dependent (P , 0.001) amelioration of the 2-cellcheckpoint induced by UV irradiation (Fig. 5A) and the 2-cell and 4-cell blocks caused by cisplatin (Fig. 5B) (P ,0.001). KU55933 had no dose-dependent effect on the UVirradiation-induced 2-cell checkpoint (P . 0.05) but partiallyameliorated the cisplatin-induced 4-cell checkpoint (P ,0.001). The amelioration of a UV irradiation-induced 2-cellcheckpoint by caffeine, but not by KU55933, implicates aprimary role for ATR in this response. The capacity ofKU55933 to ameliorate the 4-cell checkpoint induced bycisplatin implicates ATM in this response. The increasedstaining of gamma-H2AFX in response to UV irradiation wasameliorated by treatment with caffeine but not by KU55933
(Fig. 6A). No additive interaction between caffeine andKU55933 was observed. The accumulation of gamma-H2AFX in 2-cell nuclei in response to cisplatin was equallyblocked by caffeine and KU55933, and no further additiveeffect of these two inhibitors was found (Fig. 6B). Phosphor-ylation of H2AFX is a canonical response to DNA damagesensed by ATR and ATM. The inhibition of this response byinhibitors of ATR and ATM confirm the role of these PIKKs inthe embryo’s response to DNA damage.
The activation of ATM is associated with its phosphoryla-tion (at serine 1981; pATM) [34]. To date, an equivalentactivation state has not been identified for ATR. Immunolo-calization of pATM showed little staining in untreated 2-cellembryos, but increased staining was found following exposureto UV radiation or cisplatin (Fig. 7). This increased stainingwas characterized by an accumulation of pATM staining withinthe nucleus. The increased pATM staining in response to UVirradiation was largely blocked by treatment with caffeine butnot with KU55933 (Fig. 7). Caffeine and KU55933 reduced thelevel of pATM staining after cisplatin treatment (Fig. 7).
FIG. 6. The effect of ATR and ATMinhibitors on the activation of H2AFX inresponse genotoxic stress. A) Embryos wereexposed to UV radiation for 4 min, treatedwith cisplatin (12 lM), or untreated (Con-trol). During exposure, embryos were eitheruntreated (Vehicle) or cotreated with caf-feine (10 mM), KU55933 (20 lM), or acombination of the two (caffeine þKU55933). They were cultured for 4 h andthen stained for gamma-H2AFX. B) Theintensity of nuclear staining was measured,and the graph shows the mean 6 SEM ofthis staining in arbitrary units. **P , 0.001vs. respective control treatment, *P , 0.01vs. respective vehicle treatment group. Bar¼ 10 lm.
530 MU ET AL.
Dow
nloaded from w
ww
.biolreprod.org.

Both ATM and ATR act in some circumstances via theactivation of TRP53. To test for a role of TRP53, embryoswere subjected to both forms of genotoxic stress in thepresence of a TRP53 inhibitor, pifithrin-a, over a concentrationrange known to block TRP53-mediated loss of viability inembryos [35]. This treatment had no beneficial effect on thecapacity of embryos to bypass the checkpoints induced byeither UV irradiation (Fig. 8A) or cisplatin treatment (Fig. 8B).The results show that the stable checkpoints induced by UVirradiation and cisplatin treatment acted in a TRP53-indepen-dent manner.
The inhibitor LY294002 is a potent inhibitor of phospha-tidylinositol-3-kinase activity, but it is not an effective inhibitorof the protein kinase activities of ATM and ATR. Two-cellembryos subjected to UV irradiation or cisplatin treatment werecotreated with the phosphoinositide-3-kinase inhibitor,LY294002, at a concentration that blocks phosphatidylinosi-tol-3-kinase activity [36]. This treatment caused a dose-dependent augmentation (P , 0.01) of the adverse responseof embryos to cisplatin treatment but had no effect (P . 0.05)on the block caused by UV irradiation (Fig. 8C). The resultsshow that a phosphatidylinositol-3-kinase activity was notresponsible for the developmental blocks induced by UVirradiation or cisplatin treatment. It further showed that aLY294002-sensitive phosphatidylinositol-3-kinase activitymay play a role in mitigating the effects of some forms ofgenotoxic stress.
It was expected that bypassing the cell-cycle checkpointsinduced by UV irradiation or cisplatin treatment would meanthat these embryos would develop further, with marked DNAdamage. The developmental fate of these embryos wastherefore of interest and was studied by monitoring develop-ment over 72 h of culture. Caffeine treatment allowed most UVradiation-treated embryos to escape the 2-cell G
2-M block (P ,
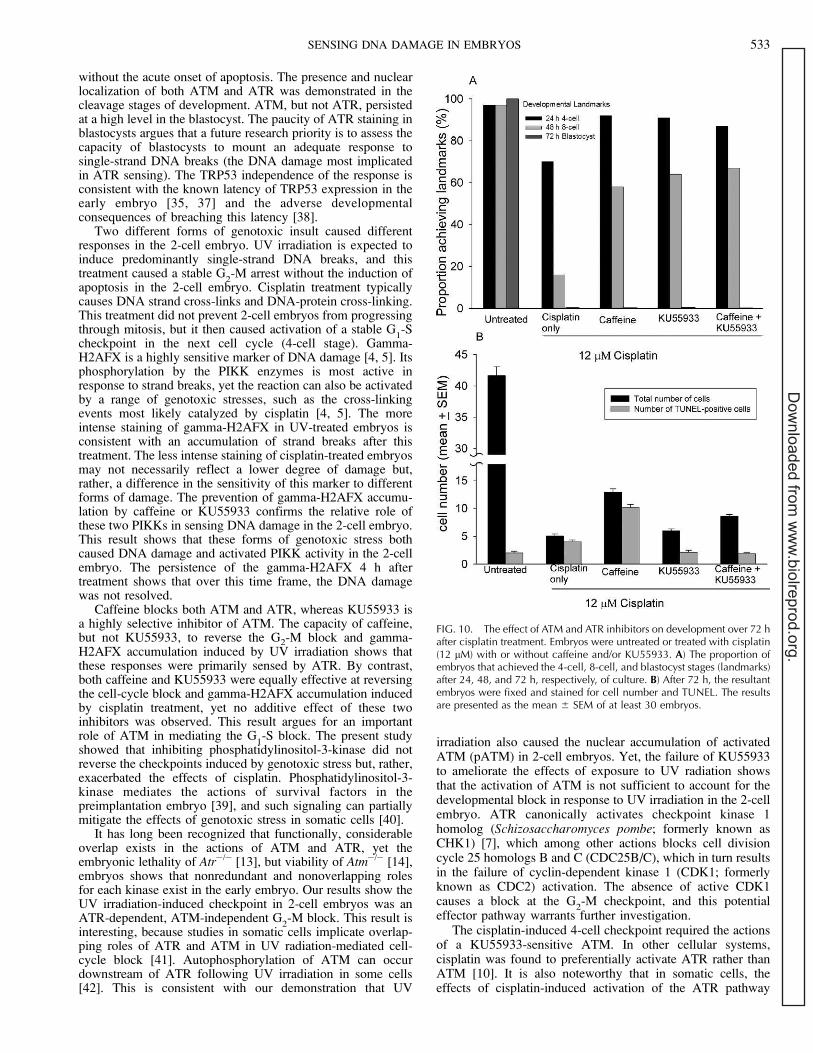
0.001), although fewer achieved the 4-cell landmark after 24 hcompared to control embryos (P , 0.01), indicating embryoswere delayed in the 2-cell stage even after caffeine treatment(Fig. 9A). Many UV-exposed embryos reached the 8-celllandmark after 48 h culture with caffeine treatment; however,this proportion was less than that in untreated controls (P ,
0.01) and less than the proportion of caffeine-treated embryosthat reached the 4-cell landmark after 24 h of culture (P ,0.05). No caffeine-treated, UV-exposed embryos achieved theblastocyst landmark after 72 h of culture, with about half at themorula stage at this time. Over this culture period, KU55933was without any beneficial effect on embryo development, andno additive effect of combined treatment with caffeine andKU55933 was observed (P . 0.05) (Fig. 9A).
Analysis of the cellular composition of the embryos after 72h of culture presented a slightly more complex picture (Fig.9B). It showed that whereas caffeine partially ameliorated theeffects of UV treatment, the resulting embryos, on average, hadmany fewer cells (P , 0.01) than control embryos, and manyof these were apoptotic, as evident by TUNEL-staining (P ,0.01). The cellular analysis confirmed the lack of effect ofKU55933 but showed that combining KU55933 with caffeineactually slightly reduced the beneficial effect of caffeine on thenumber of cells within embryos (P , 0.01). The relativeproportion of cells undergoing apoptosis was similar (P .0.05) after treatment with both inhibitors, as it was aftercaffeine treatment alone, with a large proportion of embryoniccells undergoing apoptosis.
Cisplatin treatment at the 2-cell stage caused progressivelyworsening development over the 72 h of culture after treatment.No cisplatin-treated embryos developed to morphologicalblastocysts (Fig. 10A). Caffeine treatment prevented theactivation of the 2-cell block (P , 0.001) and allowed manyembryos to escape the 4-cell block and reach the 8-celllandmark after 48 h (P , 0.001), yet none of these embryosdeveloped to blastocysts by 72 h. Treatment with KU55933 ora combination of caffeine and KU55933 resulted in develop-ment rates that were not different (P . 0.05) from the rate fortreatment with caffeine alone.
After 72 h of culture, cisplatin-treated embryos containedfewer cells than untreated controls (P , 0.001), and almost allof these were TUNEL-positive (Fig. 10B). Whereas apoptosiswas not an acute response to the 4-cell checkpoint, itspersistence did result in extensive apoptosis after 72 h.Caffeine treatment partially relieved the 4-cell block. However,the total number of cells in the resulting embryos was still
FIG. 7. The phosphorylation of ATM inresponse to genotoxic stress in the 2-cellembryo. Immunofluorescence images of 2-cell embryos stained for pATM after theirtreatment with UV irradiation or cisplatin inthe presence or absence of ATM and ATRinhibitors. Embryos were exposed to UVradiation for 4 min, treated with cisplatin(12 lM), or untreated (Control). Duringexposure, embryos were either untreated(vehicle) or cotreated with caffeine (10mM), KU55933 (20 lM), or a combinationof the two (caffeine þ KU55933). Theywere cultured for 4 h and then stained forpATM. Nonimmune control sera resulted inno detectable staining. Bar ¼ 10 lm.
SENSING DNA DAMAGE IN EMBRYOS 531
Dow
nloaded from w
ww
.biolreprod.org.
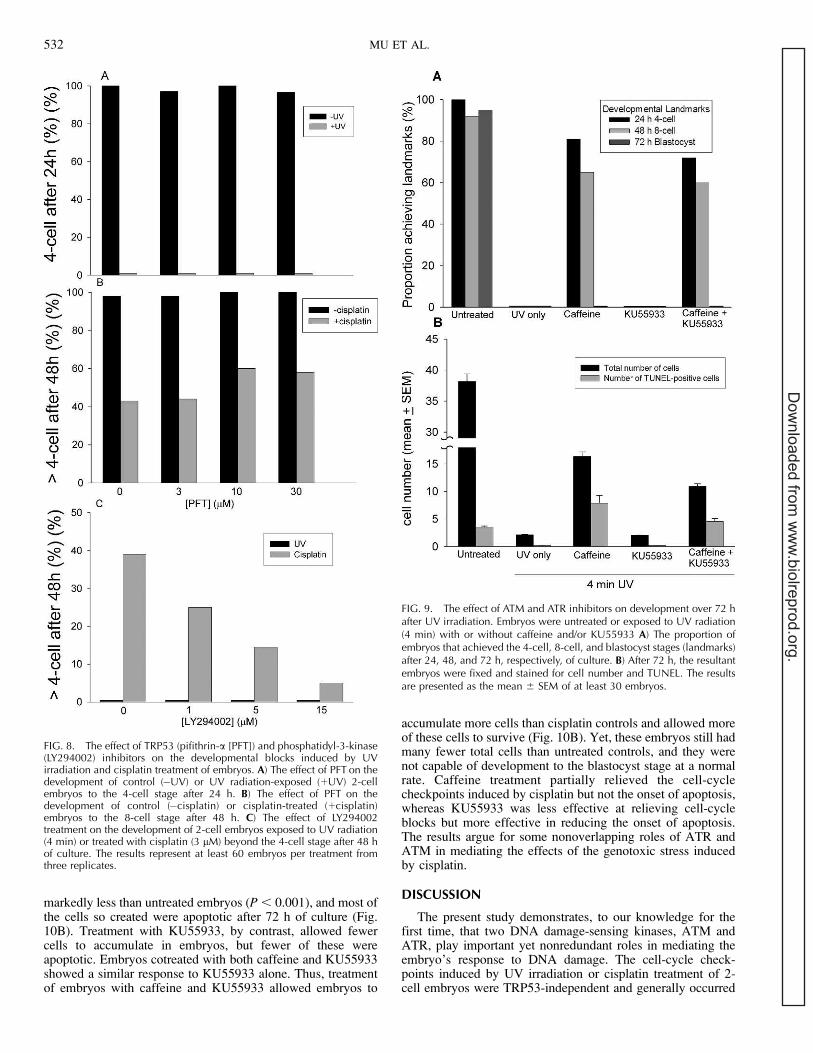
markedly less than untreated embryos (P , 0.001), and most ofthe cells so created were apoptotic after 72 h of culture (Fig.10B). Treatment with KU55933, by contrast, allowed fewercells to accumulate in embryos, but fewer of these wereapoptotic. Embryos cotreated with both caffeine and KU55933showed a similar response to KU55933 alone. Thus, treatmentof embryos with caffeine and KU55933 allowed embryos to
accumulate more cells than cisplatin controls and allowed moreof these cells to survive (Fig. 10B). Yet, these embryos still hadmany fewer total cells than untreated controls, and they werenot capable of development to the blastocyst stage at a normalrate. Caffeine treatment partially relieved the cell-cyclecheckpoints induced by cisplatin but not the onset of apoptosis,whereas KU55933 was less effective at relieving cell-cycleblocks but more effective in reducing the onset of apoptosis.The results argue for some nonoverlapping roles of ATR andATM in mediating the effects of the genotoxic stress inducedby cisplatin.
DISCUSSION
The present study demonstrates, to our knowledge for thefirst time, that two DNA damage-sensing kinases, ATM andATR, play important yet nonredundant roles in mediating theembryo’s response to DNA damage. The cell-cycle check-points induced by UV irradiation or cisplatin treatment of 2-cell embryos were TRP53-independent and generally occurred
FIG. 8. The effect of TRP53 (pifithrin-a [PFT]) and phosphatidyl-3-kinase(LY294002) inhibitors on the developmental blocks induced by UVirradiation and cisplatin treatment of embryos. A) The effect of PFT on thedevelopment of control (�UV) or UV radiation-exposed (þUV) 2-cellembryos to the 4-cell stage after 24 h. B) The effect of PFT on thedevelopment of control (�cisplatin) or cisplatin-treated (þcisplatin)embryos to the 8-cell stage after 48 h. C) The effect of LY294002treatment on the development of 2-cell embryos exposed to UV radiation(4 min) or treated with cisplatin (3 lM) beyond the 4-cell stage after 48 hof culture. The results represent at least 60 embryos per treatment fromthree replicates.
FIG. 9. The effect of ATM and ATR inhibitors on development over 72 hafter UV irradiation. Embryos were untreated or exposed to UV radiation(4 min) with or without caffeine and/or KU55933 A) The proportion ofembryos that achieved the 4-cell, 8-cell, and blastocyst stages (landmarks)after 24, 48, and 72 h, respectively, of culture. B) After 72 h, the resultantembryos were fixed and stained for cell number and TUNEL. The resultsare presented as the mean 6 SEM of at least 30 embryos.
532 MU ET AL.
Dow
nloaded from w
ww
.biolreprod.org.
without the acute onset of apoptosis. The presence and nuclearlocalization of both ATM and ATR was demonstrated in thecleavage stages of development. ATM, but not ATR, persistedat a high level in the blastocyst. The paucity of ATR staining inblastocysts argues that a future research priority is to assess thecapacity of blastocysts to mount an adequate response tosingle-strand DNA breaks (the DNA damage most implicatedin ATR sensing). The TRP53 independence of the response isconsistent with the known latency of TRP53 expression in theearly embryo [35, 37] and the adverse developmentalconsequences of breaching this latency [38].
Two different forms of genotoxic insult caused differentresponses in the 2-cell embryo. UV irradiation is expected toinduce predominantly single-strand DNA breaks, and thistreatment caused a stable G
2-M arrest without the induction of
apoptosis in the 2-cell embryo. Cisplatin treatment typicallycauses DNA strand cross-links and DNA-protein cross-linking.This treatment did not prevent 2-cell embryos from progressingthrough mitosis, but it then caused activation of a stable G
1-S
checkpoint in the next cell cycle (4-cell stage). Gamma-H2AFX is a highly sensitive marker of DNA damage [4, 5]. Itsphosphorylation by the PIKK enzymes is most active inresponse to strand breaks, yet the reaction can also be activatedby a range of genotoxic stresses, such as the cross-linkingevents most likely catalyzed by cisplatin [4, 5]. The moreintense staining of gamma-H2AFX in UV-treated embryos isconsistent with an accumulation of strand breaks after thistreatment. The less intense staining of cisplatin-treated embryosmay not necessarily reflect a lower degree of damage but,rather, a difference in the sensitivity of this marker to differentforms of damage. The prevention of gamma-H2AFX accumu-lation by caffeine or KU55933 confirms the relative role ofthese two PIKKs in sensing DNA damage in the 2-cell embryo.This result shows that these forms of genotoxic stress bothcaused DNA damage and activated PIKK activity in the 2-cellembryo. The persistence of the gamma-H2AFX 4 h aftertreatment shows that over this time frame, the DNA damagewas not resolved.
Caffeine blocks both ATM and ATR, whereas KU55933 isa highly selective inhibitor of ATM. The capacity of caffeine,but not KU55933, to reverse the G2-M block and gamma-H2AFX accumulation induced by UV irradiation shows thatthese responses were primarily sensed by ATR. By contrast,both caffeine and KU55933 were equally effective at reversingthe cell-cycle block and gamma-H2AFX accumulation inducedby cisplatin treatment, yet no additive effect of these twoinhibitors was observed. This result argues for an importantrole of ATM in mediating the G
1-S block. The present study
showed that inhibiting phosphatidylinositol-3-kinase did notreverse the checkpoints induced by genotoxic stress but, rather,exacerbated the effects of cisplatin. Phosphatidylinositol-3-kinase mediates the actions of survival factors in thepreimplantation embryo [39], and such signaling can partiallymitigate the effects of genotoxic stress in somatic cells [40].
It has long been recognized that functionally, considerableoverlap exists in the actions of ATM and ATR, yet theembryonic lethality of Atr�/� [13], but viability of Atm�/� [14],embryos shows that nonredundant and nonoverlapping rolesfor each kinase exist in the early embryo. Our results show theUV irradiation-induced checkpoint in 2-cell embryos was anATR-dependent, ATM-independent G
2-M block. This result is
interesting, because studies in somatic cells implicate overlap-ping roles of ATR and ATM in UV radiation-mediated cell-cycle block [41]. Autophosphorylation of ATM can occurdownstream of ATR following UV irradiation in some cells[42]. This is consistent with our demonstration that UV
irradiation also caused the nuclear accumulation of activatedATM (pATM) in 2-cell embryos. Yet, the failure of KU55933to ameliorate the effects of exposure to UV radiation showsthat the activation of ATM is not sufficient to account for thedevelopmental block in response to UV irradiation in the 2-cellembryo. ATR canonically activates checkpoint kinase 1homolog (Schizosaccharomyces pombe; formerly known asCHK1) [7], which among other actions blocks cell divisioncycle 25 homologs B and C (CDC25B/C), which in turn resultsin the failure of cyclin-dependent kinase 1 (CDK1; formerlyknown as CDC2) activation. The absence of active CDK1causes a block at the G
2-M checkpoint, and this potential
effector pathway warrants further investigation.The cisplatin-induced 4-cell checkpoint required the actions
of a KU55933-sensitive ATM. In other cellular systems,cisplatin was found to preferentially activate ATR rather thanATM [10]. It is also noteworthy that in somatic cells, theeffects of cisplatin-induced activation of the ATR pathway
FIG. 10. The effect of ATM and ATR inhibitors on development over 72 hafter cisplatin treatment. Embryos were untreated or treated with cisplatin(12 lM) with or without caffeine and/or KU55933. A) The proportion ofembryos that achieved the 4-cell, 8-cell, and blastocyst stages (landmarks)after 24, 48, and 72 h, respectively, of culture. B) After 72 h, the resultantembryos were fixed and stained for cell number and TUNEL. The resultsare presented as the mean 6 SEM of at least 30 embryos.
SENSING DNA DAMAGE IN EMBRYOS 533
Dow
nloaded from w
ww
.biolreprod.org.

commonly are mediated largely by the TRP53 pathway viaTRP53 phosphorylation [10]. The TRP53 independence andthe absence of any additivity between caffeine and KU55933 inthe 2-cell embryo’s response to cisplatin do not support acritical role for ATR in this response and provide anotherdifference in the embryo’s response to genotoxic stress fromthat commonly observed in somatic cells. A potential effectorpathway is the ATM activation of checkpoint kinase 2homolog (Schizosaccharomyces pombe) protein (CHEK2;formerly known as CHK2) [43], which has a number ofdownstream targets capable of inducing a G
1-S phase
checkpoint. For example, ATM and CHEK2 cause inhibitionof cell division cycle 25 homolog A (CDC25A), and this leadsto the failed activation of CDK2, resulting in checkpointactivation.
More detailed analysis of the fate of embryos following theirrelease from the cisplatin-induced G1-S checkpoint revealed apotential role for both ATR and ATM in the embryo’s responseto their stress. Caffeine unblocked the 4-cell checkpoint. After72 h of culture, however, the resulting embryos had fewer cellsthan untreated controls, and most of these cells were apoptotic.KU55933 was equally effective at unblocking the 4-cellcheckpoint. However, the resulting embryos had fewer cellsthan those treated with caffeine, but fewer of these cells wereapoptotic. The combined treatment with caffeine and KU55933resulted in embryos that had a total cell number (and rate ofapoptosis) that was intermediate between those of caffeine orKU55933 treatment alone. If caffeine and KU55933 were bothacting to inhibit ATM, then a similar outcome was expected.The difference in outcome indicates that caffeine acts, in part,on a different target, presumably ATR. Whereas an action ofcaffeine against phosphodiesterase can not be unequivocallyexcluded, other studies do not implicate the cAMP pathways inthe DNA damage response [29]. The difference in response ofcisplatin-treated embryos to caffeine and KU55933 points todual roles: a caffeine-sensitive ATR component of theembryo’s response to cisplatin, which is more important inregulating cell-cycle progression after the G
1-S checkpoint is
bypassed but is a less important mediator of apoptosis, and aKU55933-sensitive ATM component, which may be mostimportant as a mediator of apoptosis. By contrast, caffeine-treatment caused the UV-induced G
2-M checkpoint in 2-cell
embryos to be breached and allowed several further rounds ofcell division in embryos to occur. Of the cells resulting, aroundhalf were apoptotic by 72 h of culture. This checkpoint wasunaffected by KU55933 treatment, showing no overlap in theaction of ATR and ATM in this response. The results show, toour knowledge for the first time, that the G
2-M and G
1-S cell-
cycle checkpoints in the early embryo are differentiallyregulated by ATM and ATR and act as an initial point ofcontainment of different forms of genomic damage induced byUV irradiation or exposure to cisplatin.
The actions of ATM and ATR in sensing DNA damageindicate that these pathways are important for maintaining theintegrity of the genome during early development. Anoteworthy finding of the present study was that followinginhibition of these sensing activities, delayed yet effectivealternative protections against DNA damage existed in theembryo. An important future direction for investigation will beto define whether the mechanisms of DNA sensing andresponses remain the same through early embryo developmentor whether maturation and elaboration of the range of potentialresponses occur. The differences observed between ATR andATM responses observed in the embryos in the present study,compared to responses in somatic cells, indicates that someelaboration of responses likely occurs throughout development.
The results show that under the conditions of extensive orpersistent DNA damage, the demise of the embryo is theultimate method of protecting genomic integrity.
ACKNOWLEDGMENT
We thank Dr. Graeme Smith, KuDOS Pharmaceuticals Ltd., for thekind gift of KU55933.
REFERENCES
1. Abraham RT. Cell cycle checkpoint signaling through the ATM and ATRkinases. Genes Dev 2001; 15:2177–2196.
2. Zou L, Elledge SJ. Sensing DNA damage through ATRIP recognition ofRPA-ssDNA complexes. Science 2003; 300:1542–1548.
3. Falck J, Coates J, Jackson SP. Conserved modes of recruitment of ATM,ATR and DNA-PKcs to sites of DNA damage. Nature 2005; 434:605–611.
4. Dickey J, Redon C, Nakamura A, Baird B, Sedelnikova O, Bonner W.H2AX: functional roles and potential applications. Chromosoma 2009;118:683–692.
5. Mah LJ, El-Osta A, Karagiannis TC. [gamma]H2AX: a sensitivemolecular marker of DNA damage and repair. Leukemia 2010; 24:679–686.
6. Niida H, Nakanishi M. DNA damage checkpoints in mammals.Mutagenesis 2006; 21:3–9.
7. Liu QH, Guntuku S, Cui XS, Matsuoka S, Cortez D, Tamai K, Luo GB,Carattini-Rivera S, DeMayo FJ, Bradley A, Donehower LA, Elledge SJ.Chk1 is an essential kinase that is regulated by Atr and required for theG(2)/M DNA damage checkpoint. Genes Dev 2000; 14:1448–1459.
8. Geyer RK, Nagasawa H, Little JB, Maki CG. Role and regulation of p53during an ultraviolet radiation-induced G
1cell cycle arrest. Cell Growth
Differ 2000; 11:149–156.9. Banin S, Moyal L, Shieh S-Y, Taya Y, Anderson CW, Chessa L,
Smorodinsky NI, Prives C, Reiss Y, Shiloh Y, Ziv Y. Enhancedphosphorylation of p53 by ATM in response to DNA damage. Science1998; 281:1674–1677.
10. Siddik ZH. Cisplatin: mode of cytotoxic action and molecular basis ofresistance. Oncogene 2003; 22:7265–7279.
11. Jazayeri A, Falck J, Lukas C, Bartek J, Smith GCM, Lukas J, Jackson SP.ATM- and cell cycle-dependent regulation of ATR in response to DNAdouble-strand breaks. Nature Cell Biol 2005; 8:37–45.
12. de Klein A, Muijtjens M, van Os R, Verhoeven Y, Smit B, Carr AM,Lehmann AR, Hoeijmakers JHJ. Targeted disruption of the cell-cyclecheckpoint gene ATR leads to early embryonic lethality in mice. Curr Biol2000; 10:479–482.
13. Brown EJ, Baltimore D. ATR disruption leads to chromosomalfragmentation and early embryonic lethality. Genes Dev 2000; 14:397–402.
14. Xu Y, Ashley T, Brainerd E, Bronson R, Meyn M, Baltimore D. Targeteddisruption of ATM leads to growth retardation, chromosomal fragmen-tation during meiosis, immune defects, and thymic lymphoma. Genes Dev1996; 10:2411–2422.
15. Baatout S, Muller W, Michaux A, Buset J, Schoonjans W, Jacquet P.Histone H1 and Cdk1 kinase activities in early embryos of four mousestrains after X-irradiation. In Vivo 2007; 21:571–582.
16. Jacquet P, de Saint-Georges L, Barrio S, Baugnet-Mahieu L. Morpholog-ical effects of caffeine, okadaic acid and genistein in one-cell mouseembryos blocked in G
2by X-irradiation. Int J Radiat Biol 1995; 67:347–
358.17. Molls M, Streffer C, Zamboglou N. Micronucleus formation in
preimplanted mouse embryos cultured in vitro after irradiation with x-rays and neutrons. Int J Radiat Biol Relat Stud Phys Chem Med 1981;39:307–314.
18. Muller WU, Schlusen I, Streffer C. Direct evidence that radiation inducedmicronuclei of early embryos require a mitosis for expression. RadiatEnviron Biophys 1991; 30:117–122.
19. Molls M, Streffer C. The influence of G2
progression on X-ray sensitivityof two-cell mouse embryos. Int J Radiat Biol Relat Stud Phys Chem Med1984; 46:355–365.
20. Muller W, Spindle A. Induction of sister chromatid exchange inpreimplantation mouse embryos in vitro by 3H-thymidine or ultravioletlight in combination with caffeine. Teratog Carcinog Mutagen 1986;6:107–114.
21. Takahashi M, Saka N, Takahashi H, Kanai Y, Schultz RM, Okano A.Assessment of DNA damage in individual hamster embryos by cometassay. Mol Reprod Dev 1999; 54:1–7.
534 MU ET AL.
Dow
nloaded from w
ww
.biolreprod.org.

22. Brandriff B, Pedersen RA. Repair of the ultraviolet-irradiated malegenome in fertilized mouse eggs. Science 1981; 211:1431–1433.
23. Bordignon V, Smith LC. Ultraviolet-irradiated spermatozoa activateoocytes but arrest preimplantation development after fertilization andnuclear transplantation in cattle. Biol Reprod 1999; 61:1513–1520.
24. Shimura T, Inoue M, Taga M, Shiraishi K, Uematsu N, Takei N, Yuan Z-M, Shinohara T, Niwa O. p53-Dependent S-phase damage checkpoint andpronuclear cross talk in mouse zygotes with X-irradiated sperm. Mol CellBiol 2002; 22:2220–2228.
25. Toyoshima M, Shimura T, Adiga SK, Taga M, Shiraishi K, Inoue M,Yuan Z, Niwa O. Transcription-independent suppression of DNAsynthesis by p53 in sperm-irradiated mouse zygotes. Oncogene 2005;24:3229–3235.
26. Sarkaria JN, Busby EC, Tibbetts RS, Roos P, Taya Y, Karnitz LM,Abraham RT. Inhibition of ATM and ATR kinase activities by theradiosensitizing agent, caffeine. Cancer Res 1999; 59:4375–4382.
27. Rosenzweig KE, Youmell MB, Palayoor ST, Price BD. Radiosensitizationof human tumor cells by the phosphatidylinositol 3-kinase inhibitorswortmannin and LY294002 correlates with inhibition of DNA-dependentprotein kinase and prolonged G
2-M delay. Clin Cancer Res 1997; 3:1149–
1156.28. Butcher RW, Sutherland EW. Adenosine 30,50-phosphate in biological
materials. I. Purification and properties of cyclic 3 0,5 0-nucleotidephosphodiesterase and use of this enzyme to characterize adenosine30,50-phosphate in human urine. J Biol Chem 1962; 237:1244–1250.
29. Chirkov YY, Malatsidze MA, Kazarov AR, Sobolev AS. Depression ofthe radioprotective effect of isoproterenol on mammalian cells in vitroafter desensitization of the cAMP system. Int J Radiat Biol 1985; 48:753–760.
30. Hickson I, Zhao Y, Richardson CJ, Green SJ, Martin NMB, Orr AI,Reaper PM, Jackson SP, Curtin NJ, Smith GCM. Identification andcharacterization of a novel and specific inhibitor of the ataxia-telangiectasia mutated kinase ATM. Cancer Res 2004; 64:9152–9159.
31. O’Neill C. Evidence for the requirement of autocrine growth factors fordevelopment of mouse preimplantation embryos in vitro. Biol Reprod1997; 56:229–237.
32. Lu DP, Li Y, Bathgate R, Day M, O’Neill C. Ligand-activated signaltransduction in the 2-cell embryo. Biol Reprod 2003; 69:106–116.
33. Cahana A, Jin XL, Reiner O, Wynshaw-Boris A, O’Neill C. A study of thenature of embryonic lethality in LIS1�/� mice. Mol Reprod Dev 2003;66:134–142.
34. So S, Davis AJ, Chen DJ. Autophosphorylation at serine 1981 stabilizesATM at DNA damage sites. J Cell Biol 2009; 187:977–990.
35. Jin XL, Chandrakanthan V, Morgan HD, O’Neill C. Preimplantationembryo development in the mouse requires the latency of TRP53expression, which is induced by a ligand-activated PI3 kinase/AKT/MDM2-mediated signaling pathway. Biol Reprod 2009; 80:286–294.
36. Lu DP, Chandrakanthan V, Cahana A, Ishii S, O’Neill C. Trophic signalsacting via phosphatidylinositol-3 kinase are required for normalpreimplantation mouse embryo development. J Cell Sci 2004;117:1567–1576.
37. de Rozieres S, Maya R, Oren M, Lozano G. The loss of mdm2 inducesp53 mediated apoptosis. Oncogene 2000; 19:1691–1697.
38. Li A, Chandrakanthan V, Chami O, O’Neill C. Culture of zygotesincreases TRP53 expression in B6 mouse embryos which reduces embryoviability. Biol Reprod 2007; 76:362–367.
39. Li Y, Chandrakanthan V, Day ML, O’Neill C. Direct evidence for theaction of phosphatidylinositol (3,4,5)-trisphosphate-mediated signal trans-duction in the 2-cell mouse embryo. Biol Reprod 2007; 77:813–821.
40. Kao GD, Jiang Z, Fernandes AM, Gupta AK, Maity A. Inhibition ofphosphatidylinositol-3-OH kinase/Akt signaling impairs DNA repair inglioblastoma cells following ionizing radiation. J Biol Chem 2007;282:21206–21212.
41. Yajima H, Lee K-J, Zhang S, Kobayashi J, Chen BPC. DNA double-strand break formation upon UV-induced replication stress activates ATMand DNA-PKcs kinases. J Mol Biol 2009; 385:800–810.
42. Dodson GE, Tibbetts RS. DNA replication stress-induced phosphorylationof cyclic AMP response element-binding protein mediated by ATM. J BiolChem 2006; 281:1692–1697.
43. Chaturvedi P, Eng WK, Zhu Y, Mattern MR, Mishra R, Hurle MR, ZhangX, Annan RS, Lu Q, Faucette LF, Scott GF, Li X, et al. Mammalian Chk2is a downstream effector of the ATM-dependent DNA damage checkpointpathway. Oncogene 1999; 18:4047–4054.
SENSING DNA DAMAGE IN EMBRYOS 535
Dow
nloaded from w
ww
.biolreprod.org.




















