
Distribution and morphology of the ampullary organs of the estuarine long-tailed catfish, Euristhmus...
7
Zoomorphology (2009) 128:111–117 DOI 10.1007/s00435-008-0078-0 123 ORIGINAL PAPER Distribution and morphology of the ampullary organs of the estuarine long-tailed catWsh, Euristhmus lepturus (Plotosidae, Siluriformes) Darryl L. Whitehead · JeVrey T. B. Kwik · Ian R. Tibbetts Received: 29 January 2007 / Accepted: 8 October 2008 / Published online: 1 November 2008 © Springer-Verlag 2008 Abstract Ampullary organs of Euristhmus lepturus occur in high densities along the head and in four parallel path- ways along the trunk of the body. Large ampullary pores (125–130 m) are easily distinguishable from other sensory epithelial pores due to the diVerences in size and the pres- ence of a collar-like structure. Simple, singular ampullary organs of the head region consist of an ampullary pore con- nected to a long canal with a diameter of 115–175 m before terminating as a simple ampulla with an external diameter of 390–480 m. The ampullary canal is composed of 1–2 layers of Xattened squamous epithelial cells, the basement membrane and an interlocking collagen sheath. The innermost cells lining the canal wall are adjoined via tight junctions and numerous desmosomes, as are those of the receptor and supportive cells. Canal wall tissue gives rise to a sensory epithelium containing between 242 and 285 total receptor cells, with an average diameter of 11.7 § 5.3 m, intermixed with medially nucleated support- ive cells. Each receptor cell (21.38 § 4.41 m, height) has an apically positioned nucleus and a luminal surface covered with numerous microvilli. Neural terminals abut the basal region of receptor cells opposite multiple presynaptic bodies and dense mitochondria. Supportive cells extend from the ampullary lumen to the basement membrane, which is adja- cent to the complex system of collagen Wbres. Keywords Ampullary organ · Plasticity · Electroreception · Euristhmus lepturus · Plotosidae · Siluriformes Introduction Siluriformes possess electroreceptive ampullary organs (Kalmijn 1974; Finger 1986; Moller 1995) sensitive to low frequencies DC to 40 Hz (Kalmijn 1974; Zakon 1986) that are used by Wsh in communication, orientation and prey detection (Zakon 1986; Peters et al. 2001). These sensory invaginations of the epidermis exist in many forms, from the small pit organs and microampullae of freshwater silur- ids (Herrick 1901; Whitehead et al. 2003) to the elongated ampullary organs of the marine species, Plotosus anguil- laris Block 1794, (Friedrich-Freska 1930; Obara 1976). The chief morphological diVerences between these forms are the lengths of the canal, the histology of the canal wall, and the morphology and number of receptor cells (Szabo 1974; Zakon 1986; Whitehead et al. 1999, 2000). Freshwa- ter Ameiurus nebulosus Lesueur 1819, has small pit organs that are less than 300 m in total length and have fewer than 20 receptor cells per ampulla (Herrick 1901; Mullinger 1964). In contrast, P. anguillaris (Friedrich-Freska 1930; Obara 1976) has an ampullary organ that is several centi- metres in length and containing hundreds of receptor cells. A freshwater congener, Plotosus tandanus Mitchell 1838, has microampullae much like those documented for Ameiu- rus nebulosus (Herrick 1901; Mullinger 1964; Whitehead et al. 2003). This marked diVerence is believed to result from the conductive properties of the environment (Bennett D. L. Whitehead (&) · I. R. Tibbetts Centre for Marine Studies, The University of Queensland, Brisbane, QLD 4072, Australia e-mail: [email protected] J. T. B. Kwik Ecology and Systematics Laboratory, Department of Biological Sciences, National University of Singapore, 14 Science Drive 4, Singapore 117543, Singapore
Transcript of Distribution and morphology of the ampullary organs of the estuarine long-tailed catfish, Euristhmus...

Zoomorphology (2009) 128:111–117
DOI 10.1007/s00435-008-0078-0ORIGINAL PAPER
Distribution and morphology of the ampullary organs of the estuarine long-tailed catWsh, Euristhmus lepturus (Plotosidae, Siluriformes)
Darryl L. Whitehead · JeVrey T. B. Kwik · Ian R. Tibbetts
Received: 29 January 2007 / Accepted: 8 October 2008 / Published online: 1 November 2008© Springer-Verlag 2008
Abstract Ampullary organs of Euristhmus lepturus occurin high densities along the head and in four parallel path-ways along the trunk of the body. Large ampullary pores(125–130 �m) are easily distinguishable from other sensoryepithelial pores due to the diVerences in size and the pres-ence of a collar-like structure. Simple, singular ampullaryorgans of the head region consist of an ampullary pore con-nected to a long canal with a diameter of 115–175 �mbefore terminating as a simple ampulla with an externaldiameter of 390–480 �m. The ampullary canal is composedof 1–2 layers of Xattened squamous epithelial cells, thebasement membrane and an interlocking collagen sheath.The innermost cells lining the canal wall are adjoined viatight junctions and numerous desmosomes, as are those ofthe receptor and supportive cells. Canal wall tissue givesrise to a sensory epithelium containing between 242 and285 total receptor cells, with an average diameter of11.7 § 5.3 �m, intermixed with medially nucleated support-ive cells. Each receptor cell (21.38 § 4.41 �m, height) hasan apically positioned nucleus and a luminal surface coveredwith numerous microvilli. Neural terminals abut the basalregion of receptor cells opposite multiple presynaptic bodiesand dense mitochondria. Supportive cells extend from the
ampullary lumen to the basement membrane, which is adja-cent to the complex system of collagen Wbres.
Keywords Ampullary organ · Plasticity · Electroreception · Euristhmus lepturus · Plotosidae · Siluriformes
Introduction
Siluriformes possess electroreceptive ampullary organs(Kalmijn 1974; Finger 1986; Moller 1995) sensitive to lowfrequencies DC to 40 Hz (Kalmijn 1974; Zakon 1986) thatare used by Wsh in communication, orientation and preydetection (Zakon 1986; Peters et al. 2001). These sensoryinvaginations of the epidermis exist in many forms, fromthe small pit organs and microampullae of freshwater silur-ids (Herrick 1901; Whitehead et al. 2003) to the elongatedampullary organs of the marine species, Plotosus anguil-laris Block 1794, (Friedrich-Freska 1930; Obara 1976).The chief morphological diVerences between these formsare the lengths of the canal, the histology of the canal wall,and the morphology and number of receptor cells (Szabo1974; Zakon 1986; Whitehead et al. 1999, 2000). Freshwa-ter Ameiurus nebulosus Lesueur 1819, has small pit organsthat are less than 300 �m in total length and have fewerthan 20 receptor cells per ampulla (Herrick 1901; Mullinger1964). In contrast, P. anguillaris (Friedrich-Freska 1930;Obara 1976) has an ampullary organ that is several centi-metres in length and containing hundreds of receptor cells.A freshwater congener, Plotosus tandanus Mitchell 1838,has microampullae much like those documented for Ameiu-rus nebulosus (Herrick 1901; Mullinger 1964; Whiteheadet al. 2003). This marked diVerence is believed to resultfrom the conductive properties of the environment (Bennett
D. L. Whitehead (&) · I. R. TibbettsCentre for Marine Studies, The University of Queensland, Brisbane, QLD 4072, Australiae-mail: [email protected]
J. T. B. KwikEcology and Systematics Laboratory, Department of Biological Sciences, National University of Singapore, 14 Science Drive 4, Singapore 117543, Singapore
123

112 Zoomorphology (2009) 128:111–117
1971; Kalmijn 1974; Bennett and Obara 1986; Whiteheadet al. 1999, 2000, 2003).
Arius graeVei Kner and Steindachner 1867, from brack-ish water (24–26 ppt salinity) have an intermediate form ofampullary organ that ranges from 0.2 to 1.75 mm in totallength and contains 85–90 receptor cells (Whitehead et al.1999). The same species from freshwater habitats has mic-roampullary organs that range from 0.2 to 0.5 mm in totallength and have fewer than 60 receptor cells (Whiteheadet al. 2000). The marked polarity in ampullary organ mor-phology between freshwater and marine plotosids and thepresence of an apparent intermediate morphology in anestuarine Ariidae species led us to ask whether estuarineplotosids might possess a similar form of ampullary organ.Here, we describe the distribution of ampullary poresacross the body and the morphology of the ampullaryorgans from the head of Euristhmus lepturus Günther 1864,an estuarine member of the Plotosidae.
Materials and methods
Five specimens (120–250 mm, TL) of E. lepturus Günther1864 (Plotosidae, Siluriformes) were captured by anglingfrom Brisbane River (27°28�, 153°02�; 24–26 ppt, salinity;800 � cm, resistivity) and Pine River (27°18�, 153°04�; 27–29 ppt, salinity; 580 � cm, resistivity), Queensland, Austra-lia. The distribution of ampullary organs was determinedfor two unstained specimens (160 and 142 mm, TL) bytotal body studies with the aid of an illuminated magniWca-tion lamp.
Skin samples for light microscopy were removed fromthe operculum and Wxed in neutral buVered formalin. Afterroutine histological preparation, serial 6-�m sections werestained with Mayer’s haematoxylin and 1% eosin in 70%alcohol. Anatomical measurements were determined fromthe mean dimensions of at least 30 structures (e.g. receptorcells, ampullary pores, supportive cells, and measures oflength of the canal).
Samples for scanning electron microscopy were Wxed in3% glutaraldehyde in 0.1 M phosphate buVer, processedthrough an ascending series of alcohols from 20 to 90%with a Wnal two changes of absolute alcohol, dried in aPolaron Critical Point Dryer, then mounted on 12-mm-diameter stubs with double-sided tape and gold coated in aSPI Sputter Coater. Specimens were examined in either aJEOL 6400F or a JEOL 6300 scanning electron micro-scope. Scanning electron microscopy of skin surface prepa-rations was used to identify ampullary pores, taste buds,and lateral line pores.
For transmission electron microscopy, epidermal tissueblocks (1 mm3) containing ampullary pores were Wxed in3% glutaraldehyde in 0.15 M phosphate buVer. Additional
tissues were processed using 0.05, 0.1, and 0.2 M phos-phate buVer in attempts to reduce the possibly artiWcialstructure of the supportive cells. Additional tissues wereprepared following Karnovsky’s method (Karnovsky 1965)by immersing freshly obtained tissues in a formaldehyde–glutaraldehyde solution (24 h), in 0.067 M cacodylatebuVer, and tissues were washed in cacodylate buVer, pH7.2, overnight before a Wnal three washes (15 min) in 0.1 Mcacodylate buVer. All tissues were further processedthrough a series of ascending alcohols and embedded in anepoxy resin according to routine methods (Whitehead et al.1999), sectioned and viewed with a Hitachi H-300 trans-mission electron microscope at 75 kV.
Results
Distribution of ampullary pores
Whole body surveys revealed high densities of ampullarypores in the head region of both specimens. Total ampul-lary counts for the head regions were 630 and 594 ampul-lary pores for the 160 and 142 mm TL specimens,respectively. A high concentration, the supra-maxilla(sum), of non-linearly arranged ampullary pores surroundsthe base of the Wrst pair of barbels (Fig. 1). Other ampullarypores on the head generally occur in linear tracts. On eachside of the head, a dorsal supraorbital (so) tract originatesaround the nares, passes above the eye before splitting intotwo tracts. One tract continues on the dorsal midtrunkline(dmt) of the body at the base of the dorsal Wn. The secondotic track (o) passes down the posterolateral surface of thehead splitting into a further two tracts immediately poster-ior to the eye. The infra orbital (io) is the anterior-most ofthese tracts and passes beneath the eye to terminate near thebase of the mandibular barbel. The preopercular tract (po)branches from the otic ampullae to terminate on the ventralarea of the preoperculum. Lastly, a submandibular tract(sbm) of ampullary pores occurs along the central mandibu-lar area with high concentrations at the ventral apex alongthe midline of the head. Each of these tracts comprisesmultiple pores in close proximity (Fig. 1). Dissections
Fig. 1 Euristhmus lepturus (Plotosidae). Drawing of the ampullarypore distribution (TL = 160 mm). Each spot represents a single or clus-ters of individual ampullary pores. dmt dorsal midtrunkline, io infraor-bital, o otic, po preopercular, pv posterior ventral, sbl sublateralis, sbmsubmandibular, so supraorbital, sul supralateralis, and sum supraman-dibular
123

Zoomorphology (2009) 128:111–117 113
revealed that the majority of the head pores open to canalsthat end with ampullary organs in the dorso-cranial regionslightly anterior to the eyes and above a slight depression ofthe cranium.
Posterior to the head, ampullary organs occur bilaterallyalong the total length of the body in four linear rostro-cau-dally orientated tracts that end at the tip of the tail. The dor-sal midtrunkline (dmt) is an extension of the previouslydescribed dorsal supraorbital tract. The second and thirdtracts are the supralateralis (sul) and sublateralis (sbl), thatoccur slightly dorsal and ventral to the lateral line, respec-tively. This pair of tracts appears to be strongly tied to thepath of the lateral line, although no sharing of neural tissueswas observed. The posterior ventral (pv) tract lies along thepost-anal ventro-lateral surface of the Wsh adjacent to thebase of the pectoral and pelvic Wns. No ampullary poreswere identiWed within the Wn tissues, along the ventral mid-line of the trunk, or along the barbels.
Gross morphology
Ampullary organs are visible on the skin as pores with aninterior diameter of 80–110 �m (Fig. 2a, c). A thick wallsurrounds each pore producing a total diameter of 125–130 �m (Fig. 2a, c). Each ampullary pore lies in a concavityof the skin surface, whereas lateral line pores occur as con-vexities of the skin. Immediately below the ampullary pore,the ampullary canal constricts (Fig. 2b), before widening toform canals ranging from 130 to 190 �m in external diame-ter and approximately 115–175 �m in internal diameter.Each ampullary pores gives rise to an individual canal(Fig. 2c), no bifurcation was observed in the sensory tis-sues. A simple teleost ampulla arises from the distal tip ofeach canal (Fig. 2d), characterised by a single sac consist-ing of receptor and supportive cells attached to an anteriorlateral line nerve (Fig. 2d).
Ampullary canals
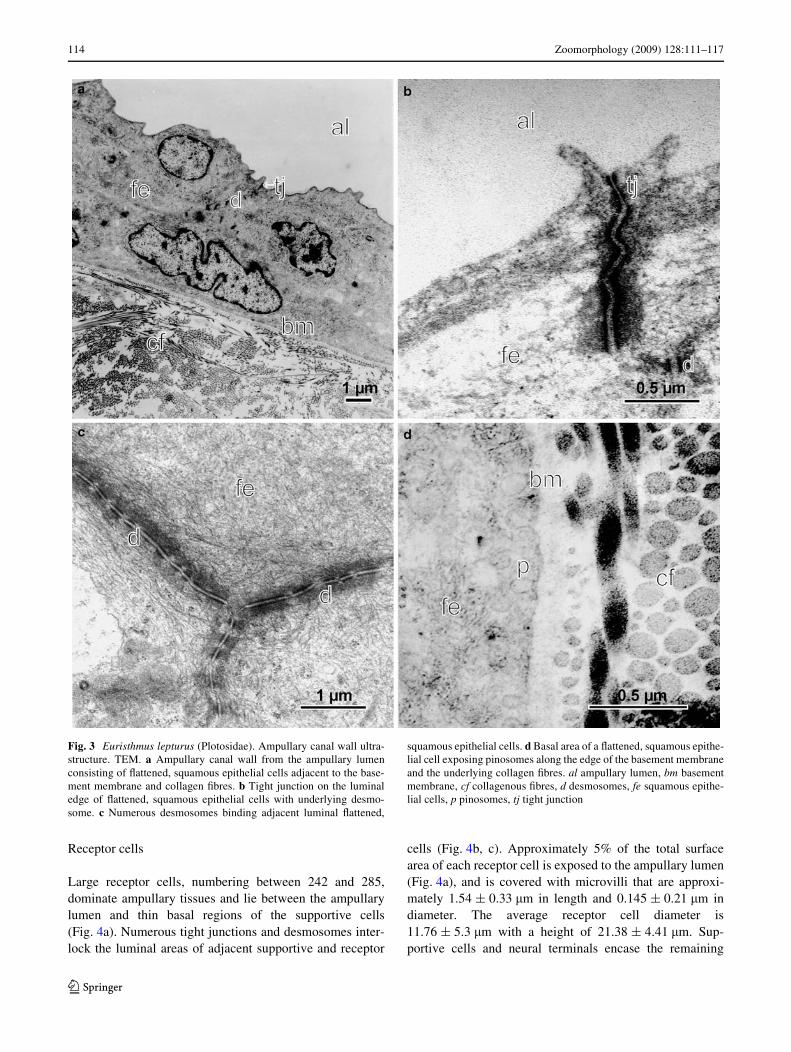
The ampullary canals of E. lepturus comprise two layers ofXattened squamous epithelial cells, a basement membrane,and a collagen sheath (Fig. 3a). The epithelial inner wallthickness ranges from 5 to 12 �m. Canal wall cells areadjoined by tight junctions along the apical cell borders(Fig. 3b) and by numerous desmosomes (Fig. 3c) through-out adjacent membranes of the exterior-most cells that abutthe ampullary lumen. This abundance of connective struc-tures is absent from the underlying layer of squamous, epi-thelial cells in the canal wall, which are directly adjacent tothe basement membrane (Fig. 3a, d) and contain numerousmembrane-bound vesicles or pinosomes (Fig. 3d). Immedi-ately adjacent to the basement membrane are numerous col-lagen Wbres that have no particular orientation, but together
form a complex sheath-like structure around the canal(Fig. 3a, d). This surrounding layer of collagen sheath isapproximately twice the thickness of the squamous celllayers of the ampullary canal wall.
Each canal terminates in a large, single ampulla that con-sists of receptor and supportive cells, which together pro-duce a wall, 17–25-�m thick, around the central ampullarylumen that is 350–450 �m in diameter. A nerve passesthrough the surrounding collagen Wbres and upon enteringthe ampulla divide into individual neurons that upon con-tact with the ampullary tissues lose their myelin sheath. Theindividual neurons appear to terminate in the form of axonboutons adjacent to receptor cells.
Fig. 2 Euristhmus lepturus (Plotosidae). Ampullary organ micro-structure. SEM. a Dermal surface containing ampullary pores showingtheir large size in comparison to the lateral line pores, and in back-ground the large ampullary canals passing leading from the epidermis.b Vertical section through dermis showing an ampullary canal passingfrom the ampullary pore through underlying connective tissues. Dou-ble head arrow details the point of canal constriction before the swell-ing of the canal. Secondarily two pointing arrows detail the canal wallthickness. c Dermal surface preparation demonstrating the concentricepidermal thickening (collar) around the pore and a single ampullarycanal arising from it. Scale bar 100 �m. d Lateral view of a dissectedampullary canal and its single ampulla, with an aVerent nerve arisingfrom the ampulla. a ampulla, ac ampullary canals, an aVerent nerve, apampullary pores, ct connective tissues, e epidermis, llp lateral linepores
123
114 Zoomorphology (2009) 128:111–117
Receptor cells
Large receptor cells, numbering between 242 and 285,dominate ampullary tissues and lie between the ampullarylumen and thin basal regions of the supportive cells(Fig. 4a). Numerous tight junctions and desmosomes inter-lock the luminal areas of adjacent supportive and receptor
cells (Fig. 4b, c). Approximately 5% of the total surfacearea of each receptor cell is exposed to the ampullary lumen(Fig. 4a), and is covered with microvilli that are approxi-mately 1.54 § 0.33 �m in length and 0.145 § 0.21 �m indiameter. The average receptor cell diameter is11.76 § 5.3 �m with a height of 21.38 § 4.41 �m. Sup-portive cells and neural terminals encase the remaining
Fig. 3 Euristhmus lepturus (Plotosidae). Ampullary canal wall ultra-structure. TEM. a Ampullary canal wall from the ampullary lumenconsisting of Xattened, squamous epithelial cells adjacent to the base-ment membrane and collagen Wbres. b Tight junction on the luminaledge of Xattened, squamous epithelial cells with underlying desmo-some. c Numerous desmosomes binding adjacent luminal Xattened,
squamous epithelial cells. d Basal area of a Xattened, squamous epithe-lial cell exposing pinosomes along the edge of the basement membraneand the underlying collagen Wbres. al ampullary lumen, bm basementmembrane, cf collagenous Wbres, d desmosomes, fe squamous epithe-lial cells, p pinosomes, tj tight junction
123

Zoomorphology (2009) 128:111–117 115
receptor cell surface (Fig. 4a). Each receptor cell contains alarge nucleus that may occur anywhere between the centraland apical regions, and is surrounded by numerousmitochondria. The nucleus contains a pronounced, darklystaining nucleolus. Rough endoplasmic reticulum is con-centrated along the basal region of each receptor cell.
Unmyelinated neural terminal boutons (Fig. 4a, d),which lie adjacent to the basal region of each receptor cell,are replete with mitochondria and vesicles. Multiple pre-synaptic bodies and numerous mitochondria also lie oppo-site these boutons within the basal areas of the receptorcells (Fig. 4d). These presynaptic bodies are of variousshapes with approximately 20 ribbon synapses with eachindividual neural bouton.
Supportive cells
Thin, elongate supportive cells extend from the lumen ofthe ampulla to the basement membrane (Fig. 4a). Support-ive cells separate and lie between all receptor cells. Theirsmooth apical surface is exposed to the lumen of the canaland bear only short protrusions, but no diVerentiatedmicrovilli (Fig. 4b). Basally, the supportive cells lie indirect contact with the basement membrane under whichlies complex arrays of collagen Wbres. The nucleus of thesupportive cells appears to be ovoid, located typically mid-level of the ampullary wall and contains a dense nucleolus(Fig. 4a).
Discussion
As in other members of the Siluridae, the ampullary organsof E. lepturus are most dense on the head (Friedrich-Freska1930; Mullinger 1964; Obara 1976; Jakubowski 1987;Whitehead et al. 1999, 2000). Ampullary pores are readilydistinguishable from lateral line pores by their larger size,location in a concavity in the skin surface, the external sur-rounding wall position, and the morphology of underlyingcanal structures. The arrangement of the ampullary poresresembles that of P. anguillaris (Friedrich-Freska 1930);Lekander 1949; Obara 1976). The majority of ampullarypores lead to sensory units in the mid-dorso-cranial regionand resemble arrangements described for P. anguillaris(Lekander 1949; Obara 1976). These cranial ampullae areelongate and are diVerentiated from the main dermal tissuesto lie in connective tissues of the head as described for P.anguillaris (Friedrich-Freska 1930; Lekander 1949; Obara1976). Another similarity between these two species wasthat ampullary organs are in linear arrays along the lengthof the body (Friedrich-Freska 1930; Lekander 1949; Bauerand Denizot 1972). There appears to be a correlation of thecentral tracts of ampullary pores to the lateral line, which
suggests that studies into their respective development maybe fruitful.
The collagen sheath that surrounds the canal resembledthat of the ampullary organs of P. anguillaris and marineelasmobranchs (Friedrich-Freska 1930; Zakon 1986;Andres and von Düring 1988; Collin and Whitehead 2004).This sheath was present along the entire length of the canaland possibly provides a mechanically stable structure thatprotects that epithelial wall from mechanical deformation(Whitehead et al. 1999). The complex arrangement of theseWbres in the sheath is reminiscent of those that surround theampulla of Plotosus tandanus (Whitehead et al. 2003).
Flattened epithelial cells form the smooth internal wallof the ampullary canal. This wall was similar to thatdescribed in other electroreceptive species (Friedrich-Freska 1930; Obara 1976; Whitehead et al. 1999). Waltman(1966) suggested that the apical borders of these cells,being bound together by tight junctions and desmosomes,provides a reXective wall for the passage of electrical sig-nals along the length of the canal, reducing electrical leak-age (Waltman 1966; Bennett and Clusin 1979).
The basal region of the ampullary canals of E. lepturusswells to form a single ampulla that resembles those ofother siluroids and freshwater elasmobranchs (Szabo et al.1972; Obara 1976; Szamier and Bennett 1980; Raschi et al.1997; Whitehead et al. 1999, 2000, 2003). The receptorcells are similar to those of other siluroids (Herrick 1901;Sato 1956; Mullinger 1964; Szabo 1974; Obara 1976; Fin-ger 1986; Zakon 1986; Whitehead et al. 2000). One unify-ing characteristic of these receptor cells is that they possessmicrovilli on the part of the apical region that is exposed tothe lumen (Zakon 1986; Collin and Whitehead 2004). Thereceptor cells of E. lepturus closely resemble thosedescribed for P. anguillaris (Friedrich-Freska 1930; Bauerand Denizot 1972; Obara 1976), and are similar in positionand size to those of other siluroids. The principal diVerenceoccurs in the density of receptor cells within a singleampulla. The high numbers observed in E. lepturus placethem into an intermediate position between the freshwaterand marine siluroids (Friedrich-Freska 1930; Mullinger1964; Whitehead et al. 1999, 2000, 2003).
Tight junctions and desmosomes are present at the con-nections between adjacent receptor and supportive cells ofE. lepturus. These are likely to provide a stable connectionsite between adjacent cells. We support suggestions thatthis structural union probably also provides electrical insu-lation, and results in the charges being directed through thereceptor cells (Waltman 1966; Whitehead et al. 1999).
The structure of the ampullary organs, from the headregions, described for this estuarine plotosid reveals simi-larities to the ampullae of Lorenzini described for themarine P. anguillaris (Friedrich-Freska 1930; Lekander1949; Nishihara and Yamamoto 1971; Bauer and Denizot
123

116 Zoomorphology (2009) 128:111–117
1972; Obara 1976), and marked diVerences from theampullary organs of the freshwater P. tandanus (Whiteheadet al. 2003). The ampullary organs from the trunk and tailregions of the species remain to be studied. The relatively
high salinities, ranging from 24 to 29 ppt., and the resistiv-ity (580 and 800 � cm) of the areas from which E. lepturuswere captured, could result in the similarity of their ampul-lary organs to those of marine species.
123

Zoomorphology (2009) 128:111–117 117
�
Variations among genera are more likely to be con-founded by phylogenetic history than diVerences within agenus or species (e.g. Arius graeVei; Whitehead et al. 1999,2000). The lack of a truly transhaline plotosid for analysisprecluded a convincing corollary in a second genus of Silu-ridae. However, our results agree with general principlethat electroreceptor morphology is inXuenced by environ-mental conductivity.
Acknowledgments The authors thank Maurizio Bigazzi, AndrewNorris, Ben Kay, Stacey Robinson for specimens; Lina Daddow forassistance with electron microscopy; Tom Gorringe for graphics. Ourappreciation is extended to the Australian Defence Science and Tech-nology Organisation for their support and direct involvement with thisresearch. This research complies with the University of Queenslandanimal experimentation ethics committee, approval numbers SMS/ZOO/068/98/URG and ZOO/489/98/ADCS/DSTO/URG/PHD.
References
Andres KH, von Düring M (1988) Comparative anatomy of vertebrateelectroreceptors. Prog Brain Res 74:113–131. doi:10.1016/S0079-6123(08)63006-X
Bauer R, Denizot JP (1972) Sur la présence et la répartition des organesampullaires chez Plotosus anguillaris. Arch Anat Microsc 61:85–90
Bennett MVL (1971) Electroreception. In: Hoar WS, Randall DJ (eds)Fish physiology, vol 5. Academic Press, New York, pp 493–574
Bennett MVL, Clusin WT (1979) Transduction at electroreceptors:origins of sensitivity. In: Cone RA, Dowling JE (eds) Membranetransduction mechanisms. Raven Press, New York, pp 91–116
Bennett MVL, Obara S (1986) Ionic mechanisms and pharmacology ofelectroreceptors. In: Bullock TH, Heiligenberg W (eds) Electro-reception. Wiley, New York, pp 157–181
Collin SP, Whitehead DL (2004) The functional roles of passive elec-troreception in non electric Wshes. Anim Biol 54:1–25.doi:10.1163/157075604323010024
Finger TE (1986) Electroreception in catWsh. Behavior, anatomy, andelectrophysiology. In: Bullock TH, Heiligenberg W (eds) Electro-reception. Wiley, New York, pp 287–318
Friedrich-Freska VH (1930) Lorenzinische Ampullen bei dem Siluro-iden Plotosus anguillaris Bloch. Zool Anz 87:49–66
Günther A (1864) Catalogue of the Wshes in the British Museum.Catalogue of the Physostomi, containing the families Siluridae,
Characinidae, Haplochitonidae, Sternoptychidae, Scopelidae,Stomiatidae in the collection of the British Museum. V 5 i–xxii,pp 1–455
Herrick C (1901) The cranial nerves and cutaneous sense organs of theNorth American siluroid Wshes. J Comp Neurol XI:177–254.doi:10.1002/cne.910110302
Jakubowski M (1987) Ultrastructure of the ampullary electroreceptiveorgans in Silurus glanis (Pisces). Z Mikrosk Anat Forsch101:461–472
Kalmijn AJ (1974) The detection of electric Welds from inanimate andanimate sources other than electric organs. In: Fessard A (ed)Handbook of sensory physiology. Springer-Verlag, New York, pp147–200
Karnovsky MJ (1965) A formaldehyde–glutaraldehyde Wxative of highosmolality for use in electron microscopy. J Cell Biol 27(2):A137
Lekander B (1949) The sensory line system and the canal bones in thehead of some Ostariophysi. Acta Zool 30:1–131
Moller P (1995) Electric Wshes: history and behavior. Chapman andHall, London
Mullinger AM (1964) The Wne structure of ampullary electric receptorsin Amiurus. Proc R Soc Lond Biol 160:345–359
Nishihara H, Yamamoto T (1971) Fine structure of ampullary organsof Lorenzini in marine catWsh, Plotosus anguillaris. J Elecron Mi-crosc 20:231
Obara S (1976) Mechanisms of electroreception in ampullae of Lo-renzini of the marine catWsh Plotosus. In: Reuben JP, Purpura DP(eds) Electrobiology of nerve synapse and muscle. Raven Press,New York, pp 128–147
Peters RC, Struik ML, Bretscheider F (2001) Electroreception in fresh-water catWsh: the biologically adequate stimulus. In: Kapoor BG,Hara TJ (eds) Sensory biology of jawed Wshes: new insights. Sci-ence Publishers Inc, EnWeld, pp 275–296
Raschi W, Keithan ED, Rhee WCH (1997) Anatomy of the ampullaryelectroreceptor in the freshwater stingray, Himantura signifer.Copeia 1997:101–107. doi:10.2307/1447844
Sato M (1956) Studies on the pit organs of Wshes IV. The distribution,histological structure and development of the small pit organs.Annot Zool Jpn 29:207–212
Szabo T (1974) Anatomy of the specialized lateral line organs of elec-troreception. In: Fessard A (ed) Handbook of sensory physiology.Springer-Verlag, New York, pp 13–58
Szabo T, Kalmijn AJ, Enger PS, Bullock TH (1972) Microampullaryorgans and a submandibular sense organ in the freshwater ray,Potamotrygon. J Comp Physiol 79:15–27. doi:10.1007/BF00693615
Szamier RB, Bennett MVL (1980) Ampullary electroreceptors in thefreshwater ray, Potamotrygon. J Comp Physiol [A] 138:225–230.doi:10.1007/BF00657040
Waltman B (1966) Electrical properties and Wne structure of theampullary canals of Lorenzini. Acta Physiol Scand 66(Suppl.264):1–60
Whitehead DL, Tibbetts IR, Daddow LYM (1999) Distribution andmorphology of the ampullary organs of the salmontail catWsh,Arius graeVei. J Morphol 239:97–105. doi:10.1002/(SICI)1097-4687(199901)239:1<;97::AID-JMOR7>;3.0.CO;2-#
Whitehead DL, Tibbetts IR, Daddow LYM (2000) Ampullary organmorphology of freshwater salmontail catWsh, Arius graeVei. JMorphol 246:142–149. doi:10.1002/1097-4687(200011)246:2<;142::AID-JMOR8>;3.0.CO;2-D
Whitehead DL, Tibbetts IR, Daddow LYM (2003) Microampullary or-gans of a freshwater eel-tailed catWsh, Plotosus (tandanus) tand-anus. J Morphol 246:142–149. doi:10.1002/1097-4687(200011)246:2<;142::AID-JMOR8>;3.0.CO;2-D
Zakon HH (1986) The electroreceptive periphery. In: Bullock TH, Hei-ligenberg W (eds) Electroreception. Wiley, New York, pp 103–156
Fig. 4 Euristhmus lepturus (Plotosidae) ampullary organ ultrastruc-ture. TEM. a Receptor and supportive cells lie between the ampullarylumen and the basement membrane, which underlain by collagenWbres. The luminal surface of the receptor cell bears microvilli, has alarge proximal nucleus and basally communicates with a neural termi-nal by a series of presynaptic bodies. Supportive cells are relativelythin and have central, polymorphic nuclei. b Exposed to the ampullarylumen, the apical areas of a receptor cell and adjacent supportive cellare connected by tight junctions and desmosomes. c Tight junction andmany desmosomes bind adjacent supportive cells (sc). d Basal regionof a receptor cell showing presynaptic bodies lying adjacent to the cellmembrane and opposite a neural terminal. al ampullary lumen, bmbasement membrane, cf collagen Wbres, d desmosome, mv microvilli,n nucleus, nt neural terminal, pb presynaptic bodies, rc receptor cell,rer rough endoplasmic reticulum, sc supportive cell, scn supportivecell nucleus, tj tight junction
123




















