
Spermatozoon and its relationship with the ovarian lamellae in the internally inseminating catfish...
9
Spermatozoon and Its Relationship With the Ovarian Lamellae in the Internally Inseminating Catfish Trachelyopterus galeatus GLEYDES G. PARREIRA, 1 HE ´ LIO CHIARINI-GARCIA, 1 ROSSANA C.N. MELO, 2 FRANCISCO O. VIEIRA, 1 AND HUGO P. GODINHO 3 * 1 Department of Morphology, Institute of Biological Sciences, Federal University of Minas Gerais, Belo Horizonte, Minas Gerais 31270-901, Brazil 2 Laboratory of Cellular Biology, Department of Biology, Federal University of Juiz de Fora, Juiz de Fora, Minas Gerais 36036-330, Brazil 3 Post-graduate Program of Vertebrate Zoology, Pontifical Catholic University of Minas Gerais, Belo Horizonte, Minas Gerais 30535-610, Brazil KEY WORDS teleost; auchenipterid; introsperm; spermatozeugma; ovarian lamellar epithelium ABSTRACT We examined the spermatozoa and their relationship with the ovarian lamellae in the catfish Trachelyopterus galeatus by classical light microscopy, high-resolution light microscopy, and transmission electron microscopy. Trachelyopterus galeatus is an internally inseminating spe- cies the spermatozoon of which presented an elongated cylindrical head (12.3 6 1.5 lm), elongated midpiece (5.0 6 0.7 lm), and flagellum (23.9 6 2.8 lm). Fertilized eggs or embryos were not found in its ovaries. Spermatozeugmata were demonstrated for the first time in this species. At the ultra- structural level, the anterior region of the head was devoid of chromatin with its shape being rounded with a hyaline tip in frontal sections and flattened in sagittal sections. The proximal cen- triole and most of the distal centriole were contained within a nuclear fossa. Mitochondria with lamellar cristae, as well as glycogen granules, were located just caudal to the nuclear fossa and dis- tally in the midpiece. A single row of accessory microtubules ran peripherally in the midpiece. The flagellar axoneme had the typical 9 1 2 arrangement, having electron-dense and electron-lucent A-tubules at different points along the flagellum; flagellar fins were lacking. The ovarian lamellae were covered by a simple cuboidal epithelium. In maturing/mature females, spermatozoa were free in the ovarian lumen or inserted in pits of the lamellar epithelial cells. Tight junctions and desmo- somes were seen between the epithelial cells. In addition to nourishment of the spermatozoon, the lamellar epithelial cells may play a role in protecting the spermatozoa against the female immune system. Microsc. Res. Tech. 72:889–897, 2009. V V C 2009 Wiley-Liss, Inc. INTRODUCTION The driftwood catfishes (Auchenipteridae, Siluri- formes) are endemic to the Neotropics, comprising at least 20 genera (Buckup et al., 2007; Ferraris, 2003) and up to 94 species (Reis et al., 2003). They are unique among the Siluriformes catfishes because they are thought to present internal insemination (Ferraris, 2003), although the reproductive biology of most mem- bers of this large family has been only cursorily investi- gated. In less than 10% of the auchenipterids, the pres- ence of spermatozoa in the ovaries has been histologi- cally demonstrated, namely, Trachelyopterus striatulus (Trachycorystes striatulus) (Ihering, 1937); Ageneiosus inermis (A. brevifilis), Auchenipterus nuchalis, Tatia brunnea, and Pseudauchenipterus nodosus (Loir et al., 1989); Trachelyopterus galeatus (Parauchenipterus galeatus) (Chacon and Mendes Filho, 1972; Loir et al., 1989; Meisner et al., 2000), and Trachelyopterus luce- nai (Meisner et al., 2000). In this article, we use the term internal insemination (Burns and Weitzman, 2005; Javonillo et al., 2009; Meisner et al., 2000) instead of internal fertilization to indicate the peculiar reproductive condition of T. galeatus, as no documented evidence of the mode of fertilization has been produced for this internally inseminating fish. Trachelyopterus galeatus (Linnaeus, 1766) is a sea- sonal breeder the reproductive period of which coin- cides with the rainy season (Chacon and Mendes Filho, 1972). The male genital system of mature T. galeatus (Chacon and Mendes Filho, 1972; Loir et al., 1989; Meisner et al., 2000) comprises a pair of testes, each organized in two distinct parts: an anterior, composed of dozens of finger-like spermatogenic lobes, and a pos- terior part formed by a variable number of secretory lobes (seminal vesicles); caudally to these lobes, there are sperm store regions. Mature males present a modi- fied anal fin (pseudopenis: Chacon and Mendes Filho, 1972, gonopodium: Loir et al., 1989; Meisner et al., *Correspondence to: Hugo P. Godinho, Post-graduate Program of Vertebrate Zoology, Pontifical Catholic University of Minas Gerais, 500 Dom Cabral Avenue, Belo Horizonte, MG 30535-610, Brazil. E-mail: [email protected] Received 30 June 2009; accepted in revised form 14 August 2009 DOI 10.1002/jemt.20782 Published online 25 September 2009 in Wiley InterScience (www.interscience. wiley.com). V V C 2009 WILEY-LISS, INC. MICROSCOPY RESEARCH AND TECHNIQUE 72:889–897 (2009)
Transcript of Spermatozoon and its relationship with the ovarian lamellae in the internally inseminating catfish...

Spermatozoon and Its Relationship With the OvarianLamellae in the Internally Inseminating CatfishTrachelyopterus galeatusGLEYDES G. PARREIRA,1 HELIO CHIARINI-GARCIA,1 ROSSANA C.N. MELO,2
FRANCISCO O. VIEIRA,1 AND HUGO P. GODINHO3*1Department of Morphology, Institute of Biological Sciences, Federal University of Minas Gerais, Belo Horizonte,Minas Gerais 31270-901, Brazil2Laboratory of Cellular Biology, Department of Biology, Federal University of Juiz de Fora, Juiz de Fora,Minas Gerais 36036-330, Brazil3Post-graduate Program of Vertebrate Zoology, Pontifical Catholic University of Minas Gerais, Belo Horizonte,Minas Gerais 30535-610, Brazil
KEY WORDS teleost; auchenipterid; introsperm; spermatozeugma; ovarian lamellarepithelium
ABSTRACT We examined the spermatozoa and their relationship with the ovarian lamellae inthe catfish Trachelyopterus galeatus by classical light microscopy, high-resolution light microscopy,and transmission electron microscopy. Trachelyopterus galeatus is an internally inseminating spe-cies the spermatozoon of which presented an elongated cylindrical head (12.3 6 1.5 lm), elongatedmidpiece (5.0 6 0.7 lm), and flagellum (23.9 6 2.8 lm). Fertilized eggs or embryos were not foundin its ovaries. Spermatozeugmata were demonstrated for the first time in this species. At the ultra-structural level, the anterior region of the head was devoid of chromatin with its shape beingrounded with a hyaline tip in frontal sections and flattened in sagittal sections. The proximal cen-triole and most of the distal centriole were contained within a nuclear fossa. Mitochondria withlamellar cristae, as well as glycogen granules, were located just caudal to the nuclear fossa and dis-tally in the midpiece. A single row of accessory microtubules ran peripherally in the midpiece. Theflagellar axoneme had the typical 9 1 2 arrangement, having electron-dense and electron-lucentA-tubules at different points along the flagellum; flagellar fins were lacking. The ovarian lamellaewere covered by a simple cuboidal epithelium. In maturing/mature females, spermatozoa were freein the ovarian lumen or inserted in pits of the lamellar epithelial cells. Tight junctions and desmo-somes were seen between the epithelial cells. In addition to nourishment of the spermatozoon, thelamellar epithelial cells may play a role in protecting the spermatozoa against the female immunesystem. Microsc. Res. Tech. 72:889–897, 2009. VVC 2009 Wiley-Liss, Inc.
INTRODUCTION
The driftwood catfishes (Auchenipteridae, Siluri-formes) are endemic to the Neotropics, comprising atleast 20 genera (Buckup et al., 2007; Ferraris, 2003)and up to 94 species (Reis et al., 2003). They are uniqueamong the Siluriformes catfishes because they arethought to present internal insemination (Ferraris,2003), although the reproductive biology of most mem-bers of this large family has been only cursorily investi-gated. In less than 10% of the auchenipterids, the pres-ence of spermatozoa in the ovaries has been histologi-cally demonstrated, namely, Trachelyopterus striatulus(Trachycorystes striatulus) (Ihering, 1937); Ageneiosusinermis (A. brevifilis), Auchenipterus nuchalis, Tatiabrunnea, and Pseudauchenipterus nodosus (Loir et al.,1989); Trachelyopterus galeatus (Parauchenipterusgaleatus) (Chacon and Mendes Filho, 1972; Loir et al.,1989; Meisner et al., 2000), and Trachelyopterus luce-nai (Meisner et al., 2000). In this article, we use theterm internal insemination (Burns and Weitzman,2005; Javonillo et al., 2009; Meisner et al., 2000)instead of internal fertilization to indicate the peculiar
reproductive condition of T. galeatus, as no documentedevidence of the mode of fertilization has been producedfor this internally inseminating fish.
Trachelyopterus galeatus (Linnaeus, 1766) is a sea-sonal breeder the reproductive period of which coin-cides with the rainy season (Chacon and Mendes Filho,1972). The male genital system of mature T. galeatus(Chacon and Mendes Filho, 1972; Loir et al., 1989;Meisner et al., 2000) comprises a pair of testes, eachorganized in two distinct parts: an anterior, composedof dozens of finger-like spermatogenic lobes, and a pos-terior part formed by a variable number of secretorylobes (seminal vesicles); caudally to these lobes, thereare sperm store regions. Mature males present a modi-fied anal fin (pseudopenis: Chacon and Mendes Filho,1972, gonopodium: Loir et al., 1989; Meisner et al.,
*Correspondence to: Hugo P. Godinho, Post-graduate Program of VertebrateZoology, Pontifical Catholic University of Minas Gerais, 500 Dom Cabral Avenue,Belo Horizonte, MG 30535-610, Brazil. E-mail: [email protected]
Received 30 June 2009; accepted in revised form 14 August 2009
DOI 10.1002/jemt.20782
Published online 25 September 2009 in Wiley InterScience (www.interscience.wiley.com).
VVC 2009 WILEY-LISS, INC.
MICROSCOPY RESEARCH AND TECHNIQUE 72:889–897 (2009)

2000), which serves in the transfer of sperm to thefemale. Upon ejaculation, a gelatinous component ofthe sperm secretion plugs the female genital papilla(Chacon and Mendes Filho, 1972). The sperm head ofT. galeatus is greatly elongated (Chacon and MendesFilho, 1972). Meisner et al. (2000) reported findingno spermatozeugmata in this fish, contrary to T. luce-nai in which they were readily observed in lightmicroscopy.
The female genital system of T. galeatus consists,briefly, of a pair of ovaries, which merge caudally intothe gonoduct before reaching the genital pore. Eachovary is externally limited by the ovarian capsule theinternal surface of which supports the ovarian lamel-lae. The gonoduct of mature females is sac-like toaccommodate the sperm introduced by the males. Sper-matozoa are found lining the ovarian lamellae (Chaconand Mendes Filho, 1972; Loir et al., 1989; Meisneret al., 2000) but only in maturing and mature females(Chacon and Mendes Filho, 1972; Loir et al., 1989).
The relationship between lined up spermatozoa andthe ovarian lamellar epithelium in internally insemi-nating fishes has been so far obtained mainly in histo-logical studies. Thus, knowledge in this area is limitedmainly due to restrictions imposed by the resolution of
light microscopy (Meisner et al., 2000). In this article,in addition to classical light microscopy, we examinedspermatozoa and their relationship with the ovarianlamellae in T. galeatus by means of high-resolutionlight microscopy (HRLM) and transmission electronmicroscopy (TEM). Spermatozeugmata were demon-strated for the first time in this species.
MATERIALS ANDMETHODSAnimals
Adult males and females of T. galeatus, in differentstages of the reproductive cycle, were collected in theSao Francisco River, Brazil. They were killed with anoverdose of Quinaldine (Merck, Germany) in 0.25 mL/Lof water, and then fragments of testes and ovaries werecollected.
Tissue Preparation
Semen from two adults was collected from the tip ofthe gonopodium after gentle coelomic hand pressure.Care was taken to avoid contaminations to the spermduring handling. Smears were immediately preparedfrom fresh, undiluted semen, stained with Giemsa-May-Grunwald (Carvalho, 2008) and examined with alight microscope.
Fig. 1. Light microscopy of T. galeatus spermatozoa. A: Bundles ofspermatozoa (black arrows) in the lumen of a seminiferous tubule isshown; the seminiferous tubule wall is indicated by a white arrow. B:
Various spermatozeugmata in semen smear in which the cells arepacked together through the heads. C: The components of the T. galea-tus spermatozoon: head, midpiece, and tail. Bars: A, B: 10 lm; C: 5 lm.
Microscopy Research and Technique
890 G.G. PARREIRA ET AL.
Testis fragments were fixed in 5% buffered glutaralde-hyde (0.1 M, pH 7.2 phosphate buffer), for 12 h, at 48C,and postfixed in reduced osmium tetroxide for 2 h. After-wards, the fragments were washed several times in thebuffer solution, dehydrated in a graded series of ethanol,and finally embedded in araldite resin (Chiarini-Garciaand Meistrich, 2008). Semithin (1 lm) and ultrathin (70–100 nm) sections were obtained, respectively, for HRLMand TEM studies. The HRLM studies were made in tis-sue sections stained with toluidine blue-sodium boratewith a BX-41 microscope connected to a Q-Color 3 digitalcamera (Olympus, Tokyo, Japan). The TEM studies wereperformed on tissue sections stained with uranyl acetateand lead citrate, and examined in a Tecnai G2-20 trans-mission electron microscope (FEI Company, The Nether-lands) and photographed with a digital camera.
DAPI (40, 6-diamidino-2-phenylindole dihydrochlor-ide; Sigma-Aldrich, St Louis, MO) staining was per-formed on spermatozoa kept moist in an agarosematrix that combines cell labeling with optimal preser-vation of cell morphology (Melo et al., 2009). Semenwas collected and gently mixed with a low-gelling tem-perature 1.25% agarose (Pierce, Rockford, IL) in a 1:3ratio (cell:agarose) and carefully spread onto slides(� 20 lL/slide). A perfusion chamber (CoverWell, GraceBio-Labs, Bend, OR) was affixed over the cells and cal-cium magnesium free HBSS (HBSS2/2) added tomaintain moisture. Slides were then incubated for 15min with a permeabilization solution (0.5% bovine se-rum albumin (BSA), 2 mM azide, 1% normal goat se-rum, 5% saponin in phosphate buffer saline (PBS)).Permeabilized cells were washed with HBSS2/2, incu-
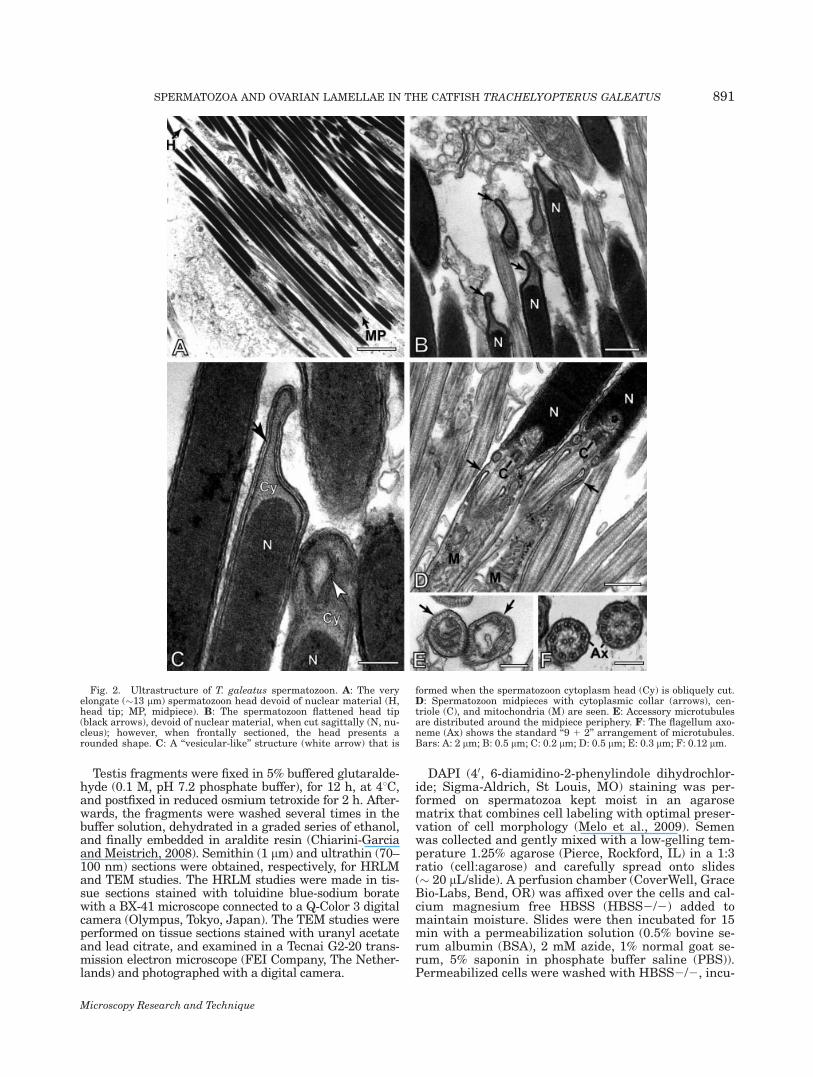
Fig. 2. Ultrastructure of T. galeatus spermatozoon. A: The veryelongate (�13 lm) spermatozoon head devoid of nuclear material (H,head tip; MP, midpiece). B: The spermatozoon flattened head tip(black arrows), devoid of nuclear material, when cut sagittally (N, nu-cleus); however, when frontally sectioned, the head presents arounded shape. C: A ‘‘vesicular-like’’ structure (white arrow) that is
formed when the spermatozoon cytoplasm head (Cy) is obliquely cut.D: Spermatozoon midpieces with cytoplasmic collar (arrows), cen-triole (C), and mitochondria (M) are seen. E: Accessory microtubulesare distributed around the midpiece periphery. F: The flagellum axo-neme (Ax) shows the standard ‘‘9 1 2’’ arrangement of microtubules.Bars: A: 2 lm; B: 0.5 lm; C: 0.2 lm; D: 0.5 lm; E: 0.3 lm; F: 0.12 lm.
Microscopy Research and Technique
891SPERMATOZOA AND OVARIAN LAMELLAE IN THE CATFISH TRACHELYOPTERUS GALEATUS

bated with DAPI (1:500 in PBS plus 2% BSA), washedin HBSS2/2, and mounted with an aqueous mountingmedium. Analyses were performed with fluorescencemicroscopy under an Olympus microscope (Olympus,Melville, NY).
Cell Measurements
Length measurements of the head, midpiece, andflagellum (lm 6 standard deviation) were made on 15loose spermatozoa in Giemsa-May-Grunwald stainedsemen smear of two males. We also measured, in elec-tron micrograph prints, the diameter (lm 6 standarddeviation) of 30 transversely cut spermatozoon headsthat were inserted into pits of the ovarian lamellar epi-thelial cells of one female.
Image Treatment
Digital HRLM and TEM images received equal digi-tal adjustment concerning focus, sharpness, contrast,and brightness, using the software Photoshop (AdobeSystem, USA).
RESULTSMorphology of the Spermatozoon
Histologically, the spermatozoa were arranged in theseminiferous tubule lumen in bundles tightly packedalong the tip of their heads with no cellular or acellularcoat. Apparently no loose spermatozoa occurred withinthe testis (Fig. 1A). A relatively large amount of a thickmucous secretion accompanied the hand-strippedsemen. The semen smears revealed discrete spermato-zeugmata the components of which were maintained
Fig. 3. Light microscopy of the lamellar arrangement in T. galea-tus ovary. A: The ovary is externally enveloped by the tunica albugi-nea (A), the lamellae are covered by the lamellar epithelium (LE) andsupport the oocytes (previtellogenic oocytes, Ov); a distinct lumen(Lu) is seen between lamellae. B: No spermatozoon is seen either in
the lumen (Lu) or in the simple cubical lamellar epithelium (LE), theoocyte (Ov) is in previtellogenic stage. C: Spermatozoa in the lumenand lying on the LE which is also simple and cubical (arrow); anoocyte (Ov) in early previtellogenic stage is seen. Bars: A: 150 lm; B,C: 30 lm.
Microscopy Research and Technique
892 G.G. PARREIRA ET AL.

together with a deeply stained core, among loose sper-matozoa (Fig. 1B). Trachelyopterus galeatus spermato-zoon (Fig. 1C) possessed a very elongated head (12.3 61.5 lm), elongate midpiece (5.0 6 0.7 lm), and flagel-lum (23.9 6 2.8 lm).
At the ultrastructural level, the uniformly cylindrical,needlelike head (0.46 6 0.05 lm in diameter) containeda nucleus with dense chromatin showing denser irregu-lar areas along its length (Figs. 2A–2C). A large areadevoid of chromatin was located at the rostral tip of thehead. The remaining surface of the nucleus was coveredby a slim rim of cytoplasm. No acrosome was present.The head tip showed a peculiar morphology. When cutfrontally, the tip of the sperm head was round andbecause of its hyaline appearance resembled the eye of aneedle (Fig. 2A), and when cut sagittally it was flat-tened anteriorly (Figs. 2B and 2C). Within the ovary,only ‘‘rounded" type was found (Figs. 6A and 6B).
The elongated midpiece was well defined (Figs. 1Cand 2D). The proximal centriole (always poorlyresolved) and part of the distal centriole (Fig. 2D) were
contained in a deep nuclear fossa and oriented approxi-mately perpendicular to each other. The distal centriolegave rise to the flagellum the initial segment of whichwas encompassed by the cytoplasmic collar and limitedby the cytoplasmic canal (Fig. 2D). The remaining seg-ment of the midpiece was a discrete projection not pene-trated by the flagellum. Mitochondria with lamellarcristae were located just caudal to the nuclear fossaand within the cytoplasmic collar where they wereaccompanied by glycogen granules (Fig. 2D). A singlerow of accessory microtubules ran peripherally in thecytoplasmic collar paralleled to its longitudinal axis(Fig. 2E).
The flagellar axoneme (Fig. 2F) had the typical 9 1 2arrangement and lacked fins. It exhibited electron-dense and electron-lucent A-tubules at different pointsalong the flagellum.
Ovarian Lamellae and RelationshipsWith Spermatozoa
Histologically, the ovarian lamellae were attached tothe entire inner surface of the ovarian capsule and iso-lated from each other by an irregular lumen. In matur-ing/mature fish, the ovarian lumen was reduced to avery irregular and narrow space. Previtellogenic oocyteswere predominantly arranged close to the lamellaperiphery, whereas those in more advanced stages ofdevelopment were randomly arranged in the lamellae.The lamellae were covered by a simple cuboidal epithe-lium (Figs. 3A–3C). Spermatozoa were observed freelyin the ovarian lumen and conspicuously inserted in thelamellar epithelium (Figs. 3C and 4A–4C) where theywere arranged in bundles probably reflecting introducedspermatozeugmata (Figs. 4A and 4B).
At the ultrastructural level, the epithelial cells fac-ing the ovarian lumen rested on a well-developed basalmembrane. Their highly constricted nucleus, basallypositioned (Figs. 5A, 5B, and 6A), contained nucleoliand smooth endoplasmic reticulum. Tight junctionsand desmosomes were seen between the epithelial cells(Figs. 5C and 5D). The cell surface was marked bynumerous pits containing multiple discrete spermheads (Figs. 5C and 5D). Epithelial cell microtubulesand intermediate filaments were seen well organizedclose to the spermatozoa (Fig. 6B).
Fertilized eggs or embryos were not found in theT. galeatus ovaries.
DISCUSSION
We characterized Trachelyopterus galeatus as aninternally inseminating species based on traits recog-nized elsewhere (Chacon and Mendes Filho, 1972;Meisner et al., 2000) and in the present work: (i) intro-mittent organ resulting from a modification in themale anal fin, (ii) striking elongation of the spermato-zoon head and midpiece, (iii) presence of unencapsu-lated sperm packets (spermatozeugmata), (iv) abun-dant secretion from nonspermatogenic testicular lobes,including the formation of a genital plug, and (v) pres-ence of spermatozoa in the ovarian lumen or embeddedwithin the ovarian lamellar epithelium. Spermatozoaare present in the ovaries of maturing/mature femalesbut not in resting individuals (Chacon and MendesFilho, 1972; Godinho, unpublished observations; Loiret al., 1989). Although a study of the genital tract anat-
Fig. 4. Light microscopy of lamellar epithelium in T. galeatusovary. A: the arrows indicate spermatozoa arranged in groups inthe LE or freely distributed in the Lu; oocyte, Ov. B: An enlargementof Figure 4A, showing spermatozoa in the lumen and embedded inlamellar cells; LE, lamellar epithelium; Lu, lumen. Bars: A: 30 lm;B: 6 lm.
Microscopy Research and Technique
893SPERMATOZOA AND OVARIAN LAMELLAE IN THE CATFISH TRACHELYOPTERUS GALEATUS

omy in T. galeatus was not within our objectives, obser-vations made in the course of our work corresponded tothose previously reported for this species (Chaconand Mendes Filho, 1972; Loir et al., 1998, Meisneret al., 2000).
Some inseminating teleosts exhibit internal gameticassociation characterized by a contact between sperma-tozoa and oocytes within the female genital tract priorto spawning. However, the spermatozoa apparently ini-tiate fertilization only when shed in the external envi-
Fig. 5. Ultrastructure of T. galeatus ovary. In A: The narrow Lucontains some spermatozoa (arrows); in both sides of the lumen, thestructural components are similar to each other, showing lamellarepithelial cells (E), theca cells (T), basal membrane (BM), follicularcells (F), zona pellucida (PZ), and oocyte (Ov). B: The following struc-
tures are seen: lamellar epithelium, from Lu to BM, and epithelialcells with convoluted nucleus (N). The electron-dense round struc-tures are transverse spermatozoon head sections (S) within epithelialcell pits. C, D: Enlarged electron micrographies squared in (B); TJ,tight junctions and D, desmosomes. Bars: A, B: 3 lm; C, D: 10 lm.
Microscopy Research and Technique
894 G.G. PARREIRA ET AL.

ronment (Javonillo et al., 2009). Although it seems notunlikely that this is the case, our data are insufficientto support whether or no this reproduction mode ispresent in T. galeatus. Among auchenipterids, fertiliza-tion biology has been examined only in T. striatulus(Ihering, 1937) in which the eggs obtained from previ-ously inseminated females were all fertilized anddeveloped normally.
In Table 1, we compare morphological characteristicsof the spermatozoon and spermatozeugmata of T. luce-nai and T. galeatus. Sperm heads and midpieces of T.galeatus and T. lucenai are strikingly elongated whencompared with the spherical to ovoid sperm heads andshort midpieces commonly seen in externally fertilizing
teleosts (Burns and Weitzman, 2005; Jamieson, 2009),although sperm heads containing spherical nuclei havebeen reported in some inseminating teleosts (Burnset al., 1997; Grier et al., 2005). Possible selectiveadvantages of elongated sperm nucleus and midpiecehave been reviewed by Burns and Weitzman (2005); insummary, a greater number of thinner cells may beable to pass through the female genital papilla at onetime, elongated shaped cells may move more easilywithin the ovarian lumen, formation of sperm packetsmay reduce cell losses to the environment during tran-sition from male to female genitalia, an elongated mid-piece may provide abundant energy for spermatozoondispersal within the ovaries and allow for an increasedlife-span of the spermatozoon during storage withinthe ovary.
The sperm head and midpiece of T galeatus are mor-phologically similar to those of T. lucenai (Burns et al.,2002, 2009). The anterior tip of T. galeatus nucleus isdevoid of chromatin (present work), similar to that ofT. lucenai and T. striatulus (Burns et al., 2009) and itsshape resembles that of T. lucenai (Figs. 11.44A and11.44B in Burns et al., 2009). The possibility that the
Fig. 6. Spermatozoa embedded in lamellar epithelial cells. A:Spermatozoa (black arrow) are deeply inserted in the lamellar epithe-lial cell pit. B: A close contact between spermatozoon head and themembrane of the epithelia cells is seen only at the spermatozoon headtip (white arrow). Microtubules (Mt) and intermediate filaments (IF)are seen well organized in epithelial cell (E) close to spermatozoonhead. BM, basal membrane; Su, supporting cell (epithelial cell); Lu,lumen; Cy, spermatozoon cytoplasm; N, nucleus and T, thecal cell.Bars: A: 1 lm and B: 0.2 lm.
TABLE 1. Comparative morphological characteristics ofspermatozoon and spermatozeugma of Trachelyopterus lucenai and
Trachelyopterus galeatus
Component T. lucenai T. galeatus
HEADShape Elongateda,b Elongatedc,d
Conicalb Cylindricald
Length (lm) 10.1a,b 12.7c
12.3d
Width/diameter (lm) 0.34b 0.46d
Tip deprivedof chromatin
Presentb Presentc,d
Tip shape Not available Flattenedd
Tip vesicles Presentb Presentc
Absentd
Acrosome Absenta,b Acrosome-likevesiclesc
Absentd
MidpieceShape Elongateda,b Elongatedc,d
Length (lm) Not available 5.0c
Nuclear fossa Presenta Presentc,d
Centrioles (relativemutual position)
Perpendiculara Approximatelyperpendiculard
Cytoplasmic collarlength (lm)
1.4a,b Not available
Accessorymicrotubules
Presenta,b Presentd
Glycogenrosettes/granules
Presenta,b Presentd
Mitochondria Presenta,b Presentd
FlagellumLength (lm) �60a 23.9d
Axonemal fins Absenta,b Absentd
Axonemalmicrotubules
9 1 2a,b 9 1 2c,d
A-tubules Electron-densea,b Electron-dense1electron-lucentd
SpermatozeugmaLength (lm) 80–84a Not availableNumber of cells/
spermatozeugmaUp to 130a,b Not available
Specialized junctions Presenta Absentd
Stainable core Not available Presentd
Presence in ovary Absenta Presentd
Source of data: aBurns et al., 2002; bBurns et al., 2009; cParreira and Godinho;dpresent study.
Microscopy Research and Technique
895SPERMATOZOA AND OVARIAN LAMELLAE IN THE CATFISH TRACHELYOPTERUS GALEATUS

cytoplasmic vesicles may represent an acrosome inT. galeatus was suggested by Parreira and Godinho(2004). However, detailed study in T. striatulusrevealed that these vesicles probably develop duringfinal chromatin condensation at the end of spermiogen-esis and represent expansions of the nuclear envelope(Ortiz in Burns et al., 2009). Moreover, this work hasshown that these ‘‘vesicles,’’ in T. galeatus, appearedonly when the head was sectioned in specific planesand they were bounded by membranes similar to thecell membrane. Thus, we conclude that T. galeatuslacks an acrosome as previously demonstrated inT. lucenai (Burns et al., 2002, 2009) and T. striatulus(Burns et al., 2009). The midpiece arrangement andcomponents of T. galeatus (this study) were similar tothose found in T. lucenai (Burns et al., 2002, 2009).Accessory microtubules, as seen in Trachelyopterus,may increase the rigidity of the cell thus facilitatingmovement within the ovary (Burns and Weitzman,2005).
The structure of the flagellum is similar in T. galea-tus and T. lucenai, except the length which is muchlarger in T. lucenai (Burns et al., 2002). In T. elongatus,besides electron-lucent A-tubules, we found electron-dense A-tubules along most of the flagellum similar towhat is observed in T. lucenai (Burns et al., 2002,2009).
Thus far, spermatozeugmata have been documentedin at least four auchenipterid species: Tatia brunnea(Loir et al., 1989), T. lucenai (Meisner et al., 2000;Burns et al., 2002, 2009), Auchenipterus nuchalis(Mazzoldi et al., 2007), and T. galeatus (this study).Regarding the spermatozeugmata of T. galeatus, Meis-ner et al. (2000) did not observed them in any of thespecimens examined. The site and mechanism of sper-matozeugma formation and its transfer to the femalegenital tract were not addressed in this study. Thepresence of spermatozeugmata in fresh T. galeatussemen (this study) indicated that their transfer to thefemale genital system would likely occur during nor-mal mating conditions. In the auchenipterids so farstudied, spermatozoa within a given spermatozeugmamay be adhered to one another by means of specializedcell junctions (T. lucenai: Burns et al., 2002) or a stain-able core structure (this study).
Free inseminated spermatozoa in the ovarian lumen,as well cells embedded in the lamellar epithelial cells,have been reported in T. lucenai (Meisner et al., 2000)and T. galeatus (Meisner et al., 2000; this study).Although no spermatozeugmata were observed withinthe ovaries in these species (Burns et al., 2002; Meisneret al., 2000), in this study Figure 4B (arrows) indicatesthat intact spermatozeugmata may indeed be embed-ded within the pits of the epithelial cells lining theovarian lumen.
This article evaluated for the first time the ultra-structural relationships between spermatozoa and theovarian lamellar epithelium in an auchenipterid cat-fish. Bundles of sperm heads were partially embeddedin pits of the cells lining the ovarian cavity. However,all cells remained discrete as evidenced by the lack ofmembrane fusion. More complex relationships aredescribed in nonauchenipterid fishes (Vila et al., 2007).
The ovarian lamellar epithelium supports somaticand germ cells (the germinal epithelium: Grier et al.,
2005), but only the somatic component was in the scopeof our study. In inseminating teleosts, these epithelialcells function as nurse cells to help keep the insemi-nated spermatozoa alive inside the ovary for anextended period of time prior to fertilization (see Uribeet al., 2005). Being functionally active, as indicated bya number of morphological characteristics (i.e., pro-nounced nuclear indentation, nucleoli and smoothendoplasmic reticulum), the epithelial cells in thisstudy may participate in the nourishment of theinseminated spermatozoon. Adjacent lamellar epithe-lial cells are joined together by junctional complexes(this study) the function of which has been regarded asforming a barrier to protect the spermatozoa from thefemale immune system (Koya et al., 1997; Vila et al.,2007).
ACKNOWLEDGMENTS
The authors thank M. L. Silva for the tissue process-ing. The ultrastructural analyses were conducted inthe Electron Microscopy Center, Federal University ofMinas Gerais, Belo Horizonte, Brazil.
REFERENCES
Buckup PA, Menezes NA, Ghazzi MSA. 2007. Introducao. In: BuckupPA, Menezes NA, Ghazzi MAS, editors. Serie Livros 23: Catalogodas especies de peixes de agua doce do Brasil. Rio de Janeiro:Museu Nacional/Universidade Federal do Rio de Janeiro. pp. 11–16.
Burns JR, Weitzman SH. 2005. Insemination in ostariophysan fishes.In: Grier HJ, Uribe MC, editors. Viviparous fishes. Homestead, FL:New Life Publications. pp. 105–132.
Burns JR, Weitzman SH, Malabarba LR. 1997. Insemination in eightspecies of Cheirodontinae fishes (Teleostei: Characidae: Cheirodon-tinae). Copeia 2:433–438.
Burns JR, Meisner AD, Weitzman SH, Malabarba LR. 2002. Spermand spermatozeugma ultrastructure in the inseminating catfish,Trachelyopterus lucenai (Ostariophysi: Siluriformes: Auchenipteri-dae). Copeia 1:173–179.
Burns JR, Quagio-Grassiotto I, Jamieson BGM. 2009. Ultrastructureof spermatozoa: Ostariophysi. In: Jamieson BGM, editor. Reproduc-tive biology and phylogeny of fishes (Agnathans and bony fishes),Part 8A. Enfield, NH: Science Publishers. pp. 287–388.
Carvalho HF. 2008. Tecnicas medicas de hematologia e imuno-hema-tologia, 8th ed. Belo Horizonte: Coopmed. 341 p.
Chacon JO, Mendes Filho A. 1972. Estudo morfologico do aparelhogenital de cangati. Trachycorystes galeatus, Linnaeus, 1756. CienCult 24:531–536.
Chiarini-Garcia H, Meistrich ML. 2008. High-resolution light micro-scopic characterization of spermatogonia. Methods Mol Biol450:95–107.
Ferraris CJ, Jr. 2003. Family Auchenipteridae (Driftwood catfishes).In: Reis RE, Kullander SO, Ferraris CJ, Jr, editors. Check list ofthe freshwater fishes of South and Central America. Porto Alegre:EDIPUCRS. pp. 470–486.
Grier HJ, Uribe MC, Parenti LR, Rosa-Cruz G. 2005. Fecundity, thegerminal epithelium, and folliculogenesis in viviparous fishes. In:Grier HJ, Uribe MC, editors. Viviparous fishes. Homestead, FL:New Life Publications. pp. 191–216.
Ihering R. 1937. Oviducal fertilization in the South American catfish,Trachycorystes. Copeia 4:2001–2005.
Jamieson BGM. 2009. Ultrastructure of spermatozoa: Euteleostei:Argentiformes, Esociformes, Salmoniformes and Osmeriformes. In:Jamieson BGM, editor. Reproductive biology and phylogeny offishes (Agnathans and bony fishes), Part 8A. Enfield, NH: SciencePublishers. pp. 389–413.
Javonillo R, Burns JR, Weitzman SH. 2009. Sperm modificationsrelated to insemination, with examples from the Ostariophisi. In:Jamieson BGM, editor. Reproductive biology and phylogeny offishes (Agnathans and bony fishes), Part 8A. Enfield, NH: SciencePublishers. pp. 723–763.
Koya Y, Munehara H, Takano K. 1997. Sperm storage and degrada-tion in the ovary of a marine copulating sculpin, Alcichthys alcicor-nis (Teleostei: Scorpaeniformes): Role of intercellular junctionsbetween inner ovarian epithelial cells. J Morphol 233:153–163.
Microscopy Research and Technique
896 G.G. PARREIRA ET AL.

Loir M, Cauty C, Planquette P, Le Bail PY. 1989. Comparative studyof the male reproductive tract in seven families of South-Americancatfishes. Aquat Living Resour 2:45–56.
Mazzoldi C, Lorenzi V, Rasotto MB. 2007. Variation of male reproduc-tive apparatus in relation to fertilization modalities in the catfishfamilies Auchenipteridae and Callichthyidae (Teleostei: Siluri-formes). J Fish Biol 70:243–256.
Meisner AD, Burns JR, Weitzman SH, Malabarba LR. 2000. Morphol-ogy and histology of the male reproductive system in two species oninternally inseminating South American catfishes. Trachelyopteruslucenai and T. galeatus (Teleostei: Auchenipteridae). J Morphol246:131–141.
Melo RCN, Spencer LA, Perez SA, Neves JS, Bafford SP, Morgan ES,Dvorak AM, Weller PF. 2009. Vesicle-mediated secretion of humaneosinophil granule-derived major basic protein. Lab Invest 89:769–781.
Parreira GG, Godinho HP. 2004. Sperm ultrastructure of thecangati Trachelyopterus galeatus (Linnaeus, 1766) (Siluri-formes, Auchenipteridae), an internal fertilizer fish. Mol Biol Cell15:91A.
Reis RE, Kullander SO, Ferraris CJ, Jr. 2003. Introduction. In: ReisRE, Kullander SO, Ferraris CJ, Jr, editors. Check list of the fresh-water fishes of South and Central America. Porto Alegre: EDI-PUCRS. pp 1–3.
Uribe MC, Rosa-Cruz G, Garcıa-Alarcon A. 2005. The ovary of vivipa-rous teleosts; morphological differences between the ovaries ofGoodea atripinnis and Ilyodon whitei (Goodeidae). In: Grier HJ,Uribe MC, editors. Viviparous fishes. Homestead, FL: New LifePublications. pp. 217–235.
Vila S, Munoz M, Sabat M, Casadevall M. 2007. Annual cycle of storedspermatozoawithin the ovaries of Helicolenus dactylopterus dacty-lopterus (Teleostei. Scorpaenidae). J Fish Biol 71:596–609.
Microscopy Research and Technique
897SPERMATOZOA AND OVARIAN LAMELLAE IN THE CATFISH TRACHELYOPTERUS GALEATUS




















