
Innate immune response to tissue-specific infection: notochord ...
Upload
independentCategory
view
5download
0
www.elsevier.com/locate/aqua-online
Aquaculture 252 (
Seasonal variation in the innate immune parameters
of the Asian catfish Clarias batrachus
Jaya Kumari, P.K. Sahoo *, T. Swain, S.K. Sahoo, A.K. Sahu, B.R. Mohanty
Central Institute of Freshwater Aquaculture, Kausalyaganga, Bhubaneswar 751 002, India
Received 7 March 2005; received in revised form 5 July 2005; accepted 25 July 2005
Abstract
To determine if the innate immunity of Asian catfish (Clarias batrachus) raised in captivity are affected by the rearing
temperature or seasons, various indices of the humoral and cellular innate immune functions were measured in fish reared at a
wide range of water temperatures over a period of 2 years. Measures of humoral immunity included the estimation of serum
myeloperoxidase, lysozyme, haemagglutinin and alternative complement activities. Cellular assay quantified the ability of
blood phagocytes to mount superoxide production. Fish were sampled during different periods of two consecutive years,
maintained at similar prevailing ranges of water temperature. Experiments were performed at mean temperatures of 19, 24, 28,
31, 32.5 8C during the same time of 2 years. The kinetics of the temperature or season mediated immunomodulation in the
innate immune parameters was remarkably fluctuated among the individuals and at different periods or temperatures. Although
a clear seasonal variation was marked in the innate immune parameters of this species, the fluctuations of all the parameters are
not consistent to any of the temperatures except for lysozyme levels that remained significantly lower during summer compared
to other seasons. The probable compensatory mechanism among the innate defence molecules might be playing role to protect
from infections during different water temperatures or seasons.
D 2005 Elsevier B.V. All rights reserved.
Keywords: Innate immunity; Annual variation; Temperature; Season; Clarias batrachus
1. Introduction
Clarias batrachus is a tropical, highly nutritive,
medium-sized catfish of Asian region, which is very
hardy in nature and tolerant to adverse ecosystems. In
0044-8486/$ - see front matter D 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.aquaculture.2005.07.025
* Corresponding author. Tel.: +91 674 2465421; fax: +91 674
2465407.
E-mail addresses: [email protected], [email protected]
(P.K. Sahoo).
intensive culture systems, productions of 100 tonnes/
ha have been reported (Thakur and Das, 1986; Are-
erat, 1987; Zheng et al., 1988). This species experi-
ences large differences in the ambient water
temperature during growout like most aquaculture
species. For instance, during summer, water tempera-
tures rise to around 29–35 8C, whereas in winter the
fish experience temperatures as low as 17–19 8C. Theclose relationship that exists between teleost and their
environment is the basis of a wide variety of studies.
2006) 121–127

J. Kumari et al. / Aquaculture 252 (2006) 121–127122
The fish have a body temperature that is essentially
the temperature of the surrounding water (Fry, 1967),
so that their entire physiology, including immune
functions, is influenced by environmental temperature
(Le Morvan et al., 1998). During different periods of
the seasons or year, C. batrachus show increased
susceptibility to various pathogens. At the onset of
the rainy season during June–July, the early stages of
this fish become highly susceptible to bacterial septi-
caemia (caused by Aeromonas hydrophila, Edward-
siella tarda, etc.) (Sahoo et al., 1998; Swain et al.,
2001) and epizootic ulcerative syndrome (Callinan et
al., 1995). The winter period is also associated with
decreased growth rates and increased susceptibility to
opportunistic pathogens (P.K. Sahoo, unpublished
observations). Little has been explored about the
immune status of this species (Sinha and Chakravarty,
1997; Kumari et al., 2003; Kumari and Sahoo, 2005).
The pathological condition in fish depends both on
temperature-dependent immune system regulation and
pathogenicity of the organism. Numerous studies have
reported the thermosensitivity of specific immune
responses (Avtalion et al., 1973; Stolen et al., 1984;
Bly and Clem, 1992; Le Morvan et al., 1998), whereas
few data are available on the effects of temperature or
season on innate immunity of fish. Lower environ-
mental temperatures adversely affect both cellular and
humoral immune responses in various fish species.
The immunologically dpermissiveT and dnon-permissiveT temperatures depend on fish species,
and the non-permissive temperature has been estab-
lished as 4 8C in salmonids, 14 8C in carp and 22 8Cin channel catfish (Bly and Clem, 1992). The non-
specific immune parameters are useful to determine
the health status of fish, evaluate immunomodulatory
substances, and act as markers of pollution and dis-
ease resistance. Many cells (mostly leucocytes and
macrophages) and their products [myeloperoxidase
(MPO), superoxides, lysozyme, complement, acute-
phase proteins, interferons, agglutinins, properdins,
lysins] contribute to the general immunological
defence mechanism (Sahoo et al., 2005).
Enzymes such as myeloperoxidase, lysins and
lysozyme are microbicidal. Activation of comple-
ment leads to the destruction and elimination of
invading pathogens such as bacteria, viruses and
fungi (Yano, 1992). Out of the three pathways of
the complement system, the alternative complement
pathway (ACP) of fish is many-fold higher than that
of mammals indicating its importance in the innate
response compared to mammals (Holland and Lam-
bris, 2002). The nitroblue tetrazolium (NBT) assay
is indicative of oxidative radical production from
neutrophils and monocytes for use in defence
against pathogens (Anderson and Siwicki, 1995).
Natural fish haemagglutinins/agglutinins bind to car-
bohydrate moieties of microbes causing agglutina-
tion for inactivation and easy immune clearance
(Dalmo et al., 1997).
This work investigated the seasonal fluctuations,
reflecting thermal influences in some of the para-
meters pertaining to non-specific immunity to estab-
lish a physiological normal range in Asian catfish
C. batrachus.
2. Materials and methods
2.1. Experimental design and fish maintenance
C. batrachus (42.23F10.06 g) utilized in this
study were collected from different culture systems
of the Central Institute of Freshwater Aquaculture,
Bhubaneswar farm. Ten to fifty-eight immature fish
were sampled at each time of the year based on the
availability. Apparently healthy fish were collected
and acclimatized in the laboratory for 15 days during
the above periods with the prevailing temperatures in
ferro-cement tanks each with a 500 l water volume.
Each tank was stocked with 10 fish and a similar
stocking density was maintained throughout the
experiment. The fish were fed with a formulated pellet
diet at the rate of 3% of their body weight once daily,
throughout the experimental period. About 10% of
water was removed daily along with the waste feed
and faecal material. The basic physicochemical water
parameters were measured systematically at 3-day
intervals to maintain optimal levels (dissolved oxy-
gen: 5.65F0.72 mg/l; pH: 8.2F0.82; nitrites:
0.015F0.009 mg/l; ammonia: 0.109F0.024 mg/l)
throughout the experiment.
Fish were tested five times a year over a period of
two consecutive years at various mean water tempera-
tures (winter, 19 8C; spring, 24 8C; summer, 32.5 8C;rainy, 31 8C; autumn, 28 8C). The water temperatures
mentioned here represent the mean values over 15

J. Kumari et al. / Aquaculture 252 (2006) 121–127 123
days of acclimatization period in the wet laboratory in
that particular season and that also simulates mean
water temperatures in the culture systems during the
corresponding seasons. The fish were sampled over 2
years in the last week of December, March, May,
August and October, representing winter, spring, sum-
mer, rainy and autumn seasons, respectively.
2.2. Sample collection
A previous health control in these fish indicated the
absence of any parasitic or bacterial infections. In order
to sample the blood, fish were quickly caught, placed in
anaesthetic (0.1 ml/l 2-phenoxyethanol) and sampled
through caudal puncture with a syringe. A part of fresh
blood was placed in heparinized tube for nitroblue
tetrazolium (NBT) test. The rest of the blood was left
to clot at 4 8C for 2 h, the clot removed after centrifuga-
tion, and the serum aliquoted and stored at �70 8C for
lysozyme, complement, haemagglutination and MPO
measurements. The complement activity was studied
on the day following bleeding.
2.3. Non-specific immunological parameters
The total myeloperoxidase content present in
serum was measured according to Quade and Roth
(1997) and a partial modified technique (Sahoo et al.,
2005). Respiratory burst activity was assayed by the
reduction of nitroblue tetrazolium (NBT) to formazan
as a measure of reactive oxygen radical production
(Anderson and Siwicki, 1995).
Hemolytic assays for the determination of the activ-
ity of the alternative complement pathway were per-
formed following the technique described by
Matsuyama et al. (1988) and Yano (1992) with partial
Table 1
Mean and standard errors for water temperature during a particular season,
corresponding immune parameters analysed over 5 different periods in tw
Variable (period of the year/season)
(mean temperature, 8C)No. of fish sampled/
season/year
Weight
December/Winter (19.00F0.15a) 15 42.80FMarch/Spring (24.00F0.16b) 25 39.65FMay/Summer (32.50F0.36e) 58 39.72FAugust/Rainy (31.00F0.11d) 42 44.18FOctober/Autumn (28.00F0.12c) 10 44.76F
Means bearing different superscript(s) column-wise are significantly ( P b
modifications (Kumari and Sahoo, 2005). Briefly, rab-
bit red blood cells (RaRBC) were washed thrice in
EGTA-Mg-GVB buffer (veronal-buffered saline con-
taining 10 mM EGTA, 10 mM MgCl2, and 0.1%
gelatin) and resuspended to 2�108 cells/ml in the
same buffer. To individual 125 Al aliquots of seriallydiluted test sera in EGTA-Mg-GVB buffer, 50 Al ofRaRBC suspension was added and each mixture was
incubated for 120 min at 20 8C. The hemolytic reaction
was stopped by adding 1.575 ml EDTA-GVB buffer
(10 mM EDTA and 0.1% gelatin) and each reaction
mixture was centrifuged at 1600�g for 5 min. The
supernatant was taken and the absorbance read at 414
nm. Control consists of complete haemolysis (1.7 ml
distilled water and 50 Al RaRBC) and a blank without
serum sample (containing only buffer and RaRBC).
The value y/(1�y) and the reciprocal of the serum
dilution were plotted on log–log graph paper and the
ACH50 (units/ml), the reciprocal dilution giving 50%
haemolysis [ y/(1�y)=1], was read from the graph.
The turbidimetric assay for lysozyme was carried
out according to Sankaran and Shanto (1972) with
partial modifications. A suspension of 150 Al ofMicro-
coccus lysodeikticus (0.2 mg/ml in 0.02 M sodium
acetate buffer, pH 5.5) was added to previously dis-
pensed 15 Al of test serum in a 96-well U-bottom
microtitre plates (Tarson, India). Initial O.D was
taken at 450 nm immediately after adding the substrate
and final O.D was taken after 1 h incubation at 24 8C.Lyophilized hen egg white lysozyme, HEWL (Sigma),
was used to develop a standard curve. Serum lysozyme
values were expressed as Ag/ml equivalent to hen egg
white lysozyme activity.
Haemagglutination assays were performed as in
Kumari and Sahoo (2005). Double serial dilution of
the inactivated sera (56 8C for 20 min) were made in
weight of the fish sampled, number of fish sampled/season/year and
o consecutive years in C. batrachus
(g) Lysozyme
(Ag/ml)
ACH50
(units/ml)
Haemagglutination
titre
0.75 16.79bF0.67 44.84bcF7.43 74.67aF21.43
1.70 17.67bF0.62 57.41cF7.47 145.07abF41.41
1.09 11.21aF0.68 46.54bcF6.39 195.23bF27.95
1.59 15.17bF0.59 28.27abF2.14 45.64aF6.81
1.18 16.97bF0.87 18.19aF7.38 204.8bF96.58
0.05) different.
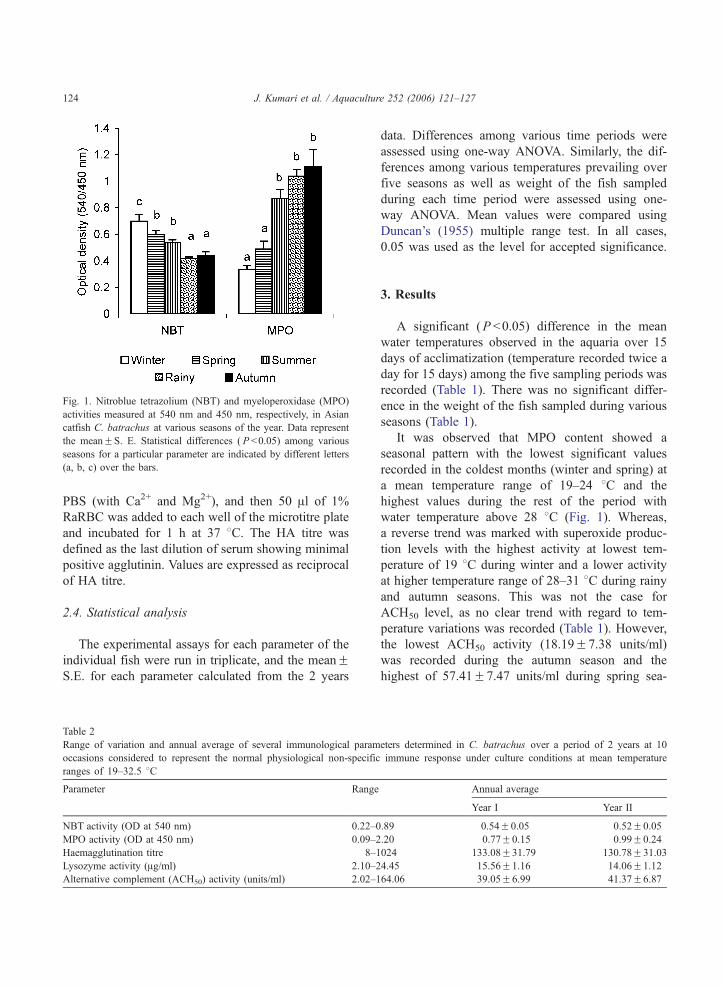
Fig. 1. Nitroblue tetrazolium (NBT) and myeloperoxidase (MPO)
activities measured at 540 nm and 450 nm, respectively, in Asian
catfish C. batrachus at various seasons of the year. Data represent
the meanFS. E. Statistical differences ( P b0.05) among various
seasons for a particular parameter are indicated by different letters
(a, b, c) over the bars.
J. Kumari et al. / Aquaculture 252 (2006) 121–127124
PBS (with Ca2+ and Mg2+), and then 50 Al of 1%
RaRBC was added to each well of the microtitre plate
and incubated for 1 h at 37 8C. The HA titre was
defined as the last dilution of serum showing minimal
positive agglutinin. Values are expressed as reciprocal
of HA titre.
2.4. Statistical analysis
The experimental assays for each parameter of the
individual fish were run in triplicate, and the meanFS.E. for each parameter calculated from the 2 years
Table 2
Range of variation and annual average of several immunological param
occasions considered to represent the normal physiological non-specific
ranges of 19–32.5 8C
Parameter Range
NBT activity (OD at 540 nm) 0.22–0
MPO activity (OD at 450 nm) 0.09–2
Haemagglutination titre 8–1
Lysozyme activity (Ag/ml) 2.10–2
Alternative complement (ACH50) activity (units/ml) 2.02–1
data. Differences among various time periods were
assessed using one-way ANOVA. Similarly, the dif-
ferences among various temperatures prevailing over
five seasons as well as weight of the fish sampled
during each time period were assessed using one-
way ANOVA. Mean values were compared using
Duncan’s (1955) multiple range test. In all cases,
0.05 was used as the level for accepted significance.
3. Results
A significant (P b0.05) difference in the mean
water temperatures observed in the aquaria over 15
days of acclimatization (temperature recorded twice a
day for 15 days) among the five sampling periods was
recorded (Table 1). There was no significant differ-
ence in the weight of the fish sampled during various
seasons (Table 1).
It was observed that MPO content showed a
seasonal pattern with the lowest significant values
recorded in the coldest months (winter and spring) at
a mean temperature range of 19–24 8C and the
highest values during the rest of the period with
water temperature above 28 8C (Fig. 1). Whereas,
a reverse trend was marked with superoxide produc-
tion levels with the highest activity at lowest tem-
perature of 19 8C during winter and a lower activity
at higher temperature range of 28–31 8C during rainy
and autumn seasons. This was not the case for
ACH50 level, as no clear trend with regard to tem-
perature variations was recorded (Table 1). However,
the lowest ACH50 activity (18.19F7.38 units/ml)
was recorded during the autumn season and the
highest of 57.41F7.47 units/ml during spring sea-
eters determined in C. batrachus over a period of 2 years at 10
immune response under culture conditions at mean temperature
Annual average
Year I Year II
.89 0.54F0.05 0.52F0.05
.20 0.77F0.15 0.99F0.24
024 133.08F31.79 130.78F31.03
4.45 15.56F1.16 14.06F1.12
64.06 39.05F6.99 41.37F6.87

J. Kumari et al. / Aquaculture 252 (2006) 121–127 125
son. Significantly lower lysozyme activity was
noticed during the peak summer (mean temperature
of 32.5 8C) (Table 1) compared to other seasonal
temperatures. On the other hand, there was a
decrease in haemagglutination titre in the coldest
winter period at 19 8C and rainy season at a tem-
perature of 31 8C compared to high titre during
summer and autumn seasons (Table 1). Wide varia-
tions in all the parameters were noticed throughout
the year among the individuals (Table 2). The mean
values of each of the immune parameters obtained
over two consecutive years did not differ signifi-
cantly (Table 2).
4. Discussion
A number of reports have shown the changes in
non-specific immune parameters with relation to
infection, toxicity, diet, stressors, pollution or tem-
perature fluctuations (Ingram, 1980; Studnicka et al.,
1986; Anderson, 1990; Anderson et al., 1992; Dalmo
et al., 1997; Ellis, 2001). Nevertheless, just a few of
these studies are related to C. batrachus (Saha et al.,
1993; Sinha and Chakravarty, 1997; Kumari and
Sahoo, 2005). No report on the effect of temperature
fluctuations or normal range in the innate immune
parameters in C. batrachus is available.
Temperature has an effect on a number of phy-
siological and immune parameters (Sinha and Chak-
ravarty, 1997; Hernandez and Tort, 2003; Tort et al.,
2004). On the other hand, the temperature fluctua-
tions have shown variable effects in the immune
parameters among different species. For example,
alternative complement activity did not reveal any
change with relation to variable temperatures (12 or
24 8C) in snapper (Pagrus auratus) (Cook et al.,
2003); whereas a clear reduction in ACH50 activity
was marked in Sparus aurata during winter period
(temperature of b10 8C). In this study, although a
significant seasonal variation was noticed in ACH50
activity of C. batrachus, the variation was not
temperature-dependent. The lowest activity was
marked during autumn at a mean temperature of
28 8C. Collectively, our results are in agreement
with recent studies in cyprinid fish on the alternative
complement pathway activities which were found to
be predominant at lower temperatures (Yano et al.,
1984; Collazos et al., 1994; Le Morvan et al., 1998;
Alcorn et al., 2002).
Similarly, one study revealed that lysozyme level
in S. aurata was less sensitive to seasonal or tempera-
ture changes (Hernandez and Tort, 2003); whereas a
clear reduction in lysozyme level was marked in S.
aurata at lower temperature (Tort et al., 2004). In our
study, the lowest lysozyme level was noticed at the
highest temperature (32.5 8C) in C. batrachus. The
wide variations in all the immune parameters noticed
in this species were also marked in other tropical
Indian major carps (Sahoo et al., 2005). Earlier studies
have shown that the adaptation to lower temperature
did lead to an increase in the respiratory burst activity
in fish (Dexiang and Ainsworth, 1991; Hardie et al.,
1994; Le Morvan et al., 1998). Similarly, the highest
superoxide production level was marked in winter at
19 8C in C. batrachus. On the contrary, the lowest
MPO activities were evident during winter or spring at
temperature of 19–24 8C. Natural haemagglutinin
level appeared to be insensitive to low temperatures
although seasonal fluctuations in this parameter was
noticed in this study.
A clear seasonal variation in all the immune para-
meters is marked in this species. However, the fluctua-
tions noticed in all the parameters are not consistent in
any one of the temperatures. This indicated that C.
batrachus might be adaptable to this temperature
changes and the compensatory mechanism among
the defence molecules might be playing a role to
protect from major pathogen attacks at various periods
of the year. The specific pathogen problem at a parti-
cular period needs a thorough elucidation into the
pathogenic mechanism of the etiological agent to say
which part of the immune system is particularly
responsible for the failure of defence. The baseline
data obtained here might be of help to work as health
indicators in pollution, stress, nutrition or infection-
related studies.
Acknowledgements
The authors wish to thank the Director, Central
Institute of Freshwater Aquaculture, Kausalyaganga,
Bhubaneswar, India and Catfish Unit for providing
the necessary facilities and fish, respectively, during
this study.

J. Kumari et al. / Aquaculture 252 (2006) 121–127126
References
Alcorn, S.W., Murra, L., Pascho, R.J., 2002. Effects of rearing
temperature on immune functions in sockeye salmon (Oncor-
hynchus nerka). Fish Shellfish Immunol. 12, 303–334.
Anderson, D.P., 1990. Immunological indicators: effects of envir-
onmental stress on immune protection and disease outbreaks.
Am. Fish. Soc. Symp. 8, 38–50.
Anderson, D.P., Moritomo, T., de Grooth, R., 1992. Neutrophil,
glass adherent, nitroblue tetrazolium assay gives early indication
of immunization effectiveness in rainbow trout. Vet. Immunol.
Immunopathol. 30, 419–429.
Anderson, D.P., Siwicki, A.K., 1995. Basic haematology and serol-
ogy for fish health programs. In: Shariff, M., Arthur, J.R.,
Subasinghe, R.P. (Eds.), Diseases in Asian Aquaculture II.
Manila, Philippines. Fish Health Section. Asian Fisheries
Society, p. 185.
Areerat, S., 1987. Clarias culture in Thailand. Aquaculture 63,
355–362.
Avtalion, R.R., Wojdani, A., Malik, Z., Shahrabani, R., Ducsyminer,
M., 1973. Influence of environmental temperature on the immune
response in fish. Curr. Top. Microbiol. Immunol. 61, 1–35.
Bly, J.E., Clem, L.W., 1992. Temperature and teleost immune
functions. Fish Shellfish Immunol. 2, 159–171.
Callinan, R.B., Paclibare, J.O., Reantaso, M.B., Lumanlan-Mayo,
S.C., Fraser, G.C., Sammut, J., 1995. EUS outbreaks in
estuarine fish in Australia and the Philippines: associations
with acid sulfate soils, rainfall and Aphanomyces. In: Shariff,
M., Arthur, J.R., Subasinghe, R.P. (Eds.), Diseases in Asian
Aquaculture II. Fish Health Section. Asian Fisheries Society,
Manila, pp. 291–298.
Collazos, M.E., Barriga, C., Ortega, E., 1994. Optimum conditions
for the activation of the alternative complement pathway of a
cyprinid fish (Tinca tinca, L.). Seasonal variations in the titres.
Fish Shellfish Immunol. 4, 499–506.
Cook, M.T., Hayball, P.J., Hutchinson, W., Nowak, B.F., Hayball,
J.D., 2003. Administration of a commercial immunostimulant
preparation, EcoActivak as a feed supplement enhances macro-
phage respiratory burst and the growth rate of snapper (Pagrus
auratus, Sparidae (Bloch and Schneider)) in winter. Fish Shell-
fish Immunol. 14, 333–345.
Dalmo, R.A., Ingebrightsen, K., Bogwald, J., 1997. Non-specific
defense mechanisms in fish, with particular reference to the
reticuloendothelial system (RES). J. Fish Dis. 20, 241–273.
Dexiang, C., Ainsworth, A.J., 1991. Effect of temperature on the
immune system of channel catfish (Ictalurus punctatus). II.
Adaptation of anterior kidney phagocytes to 10 8C. Comp.
Biochem. Physiol. 100A, 913–918.
Duncan, D.B., 1955. Multiple range and multiple dFT tests. Bio-
metrics 11, 1–42.
Ellis, A.E., 2001. Innate host defense mechanisms of fish against
viruses and bacteria. Dev. Comp. Immunol. 25, 827–839.
Fry, F.E.J., 1967. Responses of vertebrate poikilotherms to tempera-
ture. In: Rose, A.H. (Ed.), Thermobiology. Academic Press,
London, pp. 375–409.
Hardie, L.J., Fletcher, T.C., Secombes, C.J., 1994. Effect of tem-
perature on macrophage activation and the production of macro-
phage activating factor by rainbow trout (Oncorhynchus mykiss)
leucocytes. Dev. Comp. Immunol. 18, 57–66.
Hernandez, A., Tort, L., 2003. Annual variation of complement,
lysozyme and haemagglutinin levels in serum of the gilt-
head seabream Sparus aurata. Fish Shellfish Immunol. 15,
479–481.
Holland, M.C.H., Lambris, J.D., 2002. The complement system in
teleosts. Fish Shellfish Immunol. 12, 399–420.
Ingram, G.A., 1980. Substances involved in the natural resistance of
fish to infection—a review. J. Fish Biol. 16, 23–60.
Kumari, Jaya, Sahoo, P.K., 2005. Effects of cyclophosphamide on
the immune system and disease resistance of Asian catfish,
Clarias batrachus. Fish Shellfish Immunol. 19, 307–316.
Kumari, Jaya, Swain, T., Sahoo, P.K., 2003. Dietary bovine lacto-
ferrin induces changes in immunity level and disease resistance
in Asian catfish Clarias batrachus. Vet. Immunol. Immuno-
pathol. 94, 1–9.
Le Morvan, C., Troutand, D., Deschaux, P., 1998. Differential
effects of temperature on specific and non-specific immune
defences in fish. J. Exp. Biol. 201, 165–168.
Matsuyama, H., Tanaka, K., Nakao, M., Yano, T., 1988. Character-
ization of the alternative complement pathway of carp. Dev.
Comp. Immunol. 12, 403–408.
Quade, M.J., Roth, J.A., 1997. A rapid, direct assay to measure
degranulation of bovine neutrophil primary granules. Vet.
Immunol. Immunopathol. 58, 239–248.
Saha, K., Dash, K., Sahu, A., 1993. Antibody dependent haemo-
lysin, complement and opsonin in sera of a major carp, Cir-
rhina mrigala and catfish, Clarias batrachus and
Heteropneustes fossilis. Comp. Immunol. Microbiol. Infect.
Dis. 16, 323–330.
Sahoo, P.K., Mukherjee, S.C., Sahoo, S.K., 1998. Aeromonas
hydrophila versus Edwardsiella tarda: a pathoanatomical
study in Clarias batrachus. J. Aquac. 6, 57–66.
Sahoo, P.K., Kumari, Jaya, Mishra, B.K., 2005. Non-specific
immune responses in juveniles of Indian major carps. J. Appl.
Ichthyol. 21, 151–155.
Sankaran, K., Shanto, G., 1972. On the variation in catalytic
activity of lysozyme in fishes. Indian J. Biochem. Biophys.
91, 162–165.
Sinha, A., Chakravarty, A.K., 1997. Immune responses in an air-
breathing teleost Clarias batrachus. Fish Shellfish Immunol. 7,
105–114.
Stolen, J.S., Draxler, S., Nagle, J.J., 1984. A comparison of tem-
perature-mediated immunomodulation between two species of
flounder. Immunol. Commun. 13, 245–253.
Studnicka, M., Siwicki, A., Ryka, B., 1986. Lysozyme level in carp
(Cyprinus carpio L.). Bamidgeh 38, 22–25.
Swain, P., Mukherjee, S.C., Das, B.K., Sahoo, P.K., Pattnaik, P.,
Murjani, G., Nayak, S.K., 2001. Dot-enzyme linked immuno-
sorbent assay (Dot-ELISA) for diagnosis of Edwardsiella tarda
infection in fish. Asian Fish. Sci. 14, 89–95.
Thakur, N.K, Das, P., 1986. Synopsis of biological data on magur,
Clarias batrachus (Linnaeus, 1758). CIFRI, Bull. 41, 1–42.
Tort, L., Rotllant, J., Liarte, C., Acerete, L., Hernandez, A., Ceule-
mans, S., Coutteau, P., Padros, F., 2004. Effects of temperature
decrease on feeding rates, immune indicators and histopatholo-

J. Kumari et al. / Aquaculture 252 (2006) 121–127 127
gical changes of gilthead seabream Sparus aurata fed with an
experimental diet. Aquaculture 229, 55–65.
Yano, T., 1992. Assays of haemolytic complement activity. In:
Stolen, J.S., Fletcher, T.C., Anderson, D.P., Kaatari, S.L., Row-
ley, A.F. (Eds.), Techniques in Fish Immunology, vol. 2. SOS,
USA, pp. 131–141.
Yano, T., Ando, H., Nakao, M., 1984. Optimum conditions for the
assay of haemolytic titer of carp and seasonal variation of the
titers. J. Fac. Agric., Kyushu Univ. 29, 91–101.
Zheng, W.B., Pan, J.H., Liu, W.S., 1988. Culture of catfish in China.
Aquaculture 75, 35–44.
Copyright © 2022 FDOKUMEN




















