
Resveratrol modulates innate and inflammatory responses in fish leucocytes
12
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
-
Upload
independent -
Category
Documents
-
view
6 -
download
0
Transcript of Resveratrol modulates innate and inflammatory responses in fish leucocytes

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Resveratrol modulates innate and inflammatory responses
in fish leucocytes
R. Castro a, J. Lamas a, P. Morais b, M.L. Sanmartın b,F. Orallo c, J. Leiro b,*
a Departamento de Biologıa Celular y Ecologıa, Facultad de Biologıa, Universidad de Santiago de Compostela,
15782 Santiago de Compostela, Spainb Departamento de Microbiologıa y Parasitologıa, Laboratorio de Parasitologıa, Instituto de Investigacion y Analisis Alimentarios,
Universidad de Santiago de Compostela, 15782 Santiago de Compostela, Spainc Departamento de Farmacologıa, Facultad de Farmacia, Universidad de Santiago de Compostela,
15782 Santiago de Compostela, Spain
Received 24 February 2008; received in revised form 23 May 2008; accepted 5 June 2008
Abstract
Resveratrol (RESV; trans-3,5,4’-trihydroxystilbene), a phytoalexin that is produced by some plants, among other effects has
well-known antioxidant, anti-inflammatory and immunomodulatory activities in mammals. In the present study, the effects of
RESVon several functions of turbot, Psetta maxima (L.), kidney leucocytes (KLs) related to the innate and inflammatory responses
were investigated. RESV exerted a dose-dependent inhibitory effect on the migratory response and on the production of reactive
oxygen species in KL, after stimulation of the respiratory burst activity with phorbol myristate acetate (PMA). RESV also
significantly inhibited the generation of the pro-inflammatory mediator prostaglandin E2 (PGE2) in the supernatant of KL cultures
stimulated with acidic sulphated polysaccharides (ASPs) from the seaweed Ulva rigida. The effects of the polyphenol on enzymatic
activity and on myeloperoxidase (MPO) gene expression in neutrophils were also tested. It was found that RESV strongly inhibited
intracellular and extracellular MPO activity, behaving as a noncompetitive and reversible inhibitor, and also induced a decrease in
MPO mRNA levels in turbot neutrophils. These findings indicate that RESV exerts important modulatory effects on inflammatory
responses in fish, and considering the importance of innate immunity in these vertebrates and the similarities with mammals, it may
be possible to use fish for analysis of the effects of different substances on inflammatory responses.
# 2008 Elsevier B.V. All rights reserved.
Keywords: Resveratrol; Turbot leucocytes; Innate immune response; Myeloperoxidase; Inflammation; Respiratory burst
1. Introduction
Polyphenols are a group of phytochemicals that are
increasingly considered as being responsible for the
health benefits provided by fruit and vegetables (Fraga,
2007). Resveratrol (RESV; trans-3,5,4’-trihydroxystil-
bene), a natural polyphenol, was first isolated in 1940 as
a constituent of the roots of white hellebore (Veratrum
grandiflorum O. Loes), but since then it has been found
in various plants, including grapes, berries and peanuts
(Khanna et al., 2007). RESV has a variety of effects on
mammals including protection against ischaemia-
reperfusion injury, as well as antitumour and chemo-
preventative action against malignant tumours (Chen
www.elsevier.com/locate/vetimm
Available online at www.sciencedirect.com
Veterinary Immunology and Immunopathology 126 (2008) 9–19
* Corresponding author at: Laboratorio de Parasitologıa, Instituto de
Investigacion y Analisis Alimentarios, Universidad de Santiago de
Compostela, C/ Constantino Candeira s/n, 15782 Santiago de Com-
postela, Spain. Tel.: +34 981563100; fax: +34 981547171.
E-mail address: [email protected] (J. Leiro).
0165-2427/$ – see front matter # 2008 Elsevier B.V. All rights reserved.
doi:10.1016/j.vetimm.2008.06.001

Author's personal copy
and Tseng, 2007); and it has recently been demonstrated
that RESV extends lifespan in fish (Valenzano and
Cellerino, 2006).
Fish represent the earliest class of vertebrates that
possess elements of both innate and acquired immunity,
and are considered of great interest for study of the
similarities and differences between their defence
mechanisms and those of higher vertebrates (Plouffe
et al., 2005). In fish, the innate immune response has
been considered an essential component in combating
disease due to the constraints placed on the adaptive
immune response in terms of the limited antibody
repertoires, affinity maturation and memory and
relatively slow lymphocyte proliferation (Pedrera
et al., 1992; Collazos et al., 1995; Magnadottir, 2006;
Whyte, 2007). Although inflammation constitutes a
primary response of an organism’s defence mechanism,
inflammation may also underlie various chronic
diseases (Kundu et al., 2004). Both acute and chronic
inflammatory responses occur through cellular reac-
tions mediated by chemical factors such as cytokines,
chemokines, prostaglandins (PGs), nitric oxide (NO)
and other reactive oxygen species (ROS), leukotrienes,
etc., which are produced in response to pro-inflamma-
tory stimuli (Kumar et al., 2004). In animals, including
fish, the inflammatory response to various infectious
agents involves strong stimulation of the polymorpho-
nuclear neutrophils that release ROS and myeloperox-
idase (MPO), and inhibition of the oxidising and MPO
activities of neutrophils offers therapeutic perspectives
in animal pathologies with excessive inflammatory
reactions (Iglesias et al., 2001; Kohnen et al., 2007;
Stakauskas et al., 2007). RESV is an important
inhibitory agent of inflammatory processes, which in
mammals are regulated through the NF-kappa-B
pathway (Leiro et al., 2003, 2004a,b, 2005; Nam,
2006). Because RESV has an inhibitory effect on the
production of ROS by mammalian neutrophils (Caval-
laro et al., 2003; Kohnen et al., 2007), it is possible that
this polyphenol may decrease the inflammatory
response, and that the prolonged lifespan observed in
fish may be at least partly associated with a decrease in
the amount of ROS produced.
The present study was carried out to determine
whether RESV affects the inflammatory response in
turbot, Psetta maxima (L.), by analysis of migration,
production of prostaglandins, respiratory burst activity
and MPO activity and gene expression in fish kidney
leucocytes (KLs). We used turbot because this is one of
the most important fish species in European aqua-
culture, because we have previously studied all these
activities in kidney leucocytes response to different
stimuli and finally because we have characterized turbot
MPO and sequenced a fragment of the gene (Castro
et al., 2008), thereby facilitating the experimental
analyses involved.
2. Materials and methods
2.1. Fish
Male and female turbot (50–100 g; about 3 months
old) were obtained from a local fish farm in Galicia
(northwestern Spain). Prior to experiments, the fish
were acclimatized for at least 15 days in 200 l tanks with
a constant flow of water (14 8C, pH 6.5 � 0.5) and
aeration. The fish were fed daily (between 10:00 and
11:00 a.m.) with a standard, commercially available
semi-dried pelleted food.
2.2. Ethical approval
All experiments were carried out in accordance with
European regulations on animal protection (Directive
86/609), agreed under the Declaration of Helsinki. All
experimental protocols were approved by the Institu-
tional Animal Care and Use Committee of the
University of Santiago de Compostela (Spain).
2.3. Stimuli, drugs, and chemicals
Strain I1 of the ciliate Philasterides dicentrarchi
(Iglesias et al., 2001) was maintained under the culture
conditions described (Iglesias et al., 2003). Turbot was
experimentally infected by intraperitoneal inoculation
of 5 � 104 ciliates and the fish developed fluid in the
abdominal cavity 4 days post-inoculation (Parama et al.,
2004a,b). Ascitic fluid was obtained by abdomenocent-
esis and separation of ciliates by centrifugation at
600 � g for 5 min; the fluid was aliquoted and stored at
�80 8C until use.
Parasite lysate was obtained as described (Parama
et al., 2004a,b). Briefly, the ciliates were washed three
times by centrifugation at 600 � g for 5 min in 0.25 M
sucrose at 4 8C. The cells were then disrupted
ultrasonically at 4 8C in a protease inhibitor cocktail
(Sigma–Aldrich). The homogenate was centrifuged at
15,000 � g for 15 min at 4 8C and aliquots of the
resultant supernatant fraction were stored at �80 8C.
Proteases from P. dicentrarchi trophozoites were
purified on a bacitracin-sepharose affinity column, as
previously described (Parama et al., 2007). Aliquots of
the 2.5 ml fractions collected were assayed for
proteolytic activity, as previously described (Parama
R. Castro et al. / Veterinary Immunology and Immunopathology 126 (2008) 9–1910

Author's personal copy
et al., 2004a,b) and those fractions showing enzyme
activity were pooled and concentrated by ultrafiltration
with Amicon1 Ultra K centrifugal filter devices
(Millipore, USA), dissolved in 0.15 M PBS and stored
at �80 8C until use. The concentration of protein in the
ascitic fluid and in ciliate lysates was determined by the
Bradford method, with a Bio-Rad Protein Assay kit
(Bio-Rad Laboratories, Germany), and with bovine
serum albumin (BSA; Sigma–Aldrich, Spain) as
standard.
Acidic sulphated polysaccharides (ASPs) were
obtained from the seaweed Ulva rigida C. Agardh
and analysed for monosaccharide composition and
structure, as previously described (Castro et al., 2004,
2006; Leiro et al., 2007).
A stock solution (100 mM) of phorbol myristate
acetate (PMA; Sigma–Aldrich), trans-resveratrol
(RESV; Sigma–Aldrich) and indomethacin (INDO;
Sigma–Aldrich) was prepared in dimethyl sulphoxide
(DMSO) and stored in the dark at�80 8C until use. L(+)
Ascorbic acid (Sigma–Aldrich) was prepared in PBS
and stored at �20 8C until use.
2.4. Isolation of turbot kidney leucocytes and
preparation of cell lysates
The kidney is main haematopoietic organ in turbot.
Isolation of kidney leucocytes was carried out as
described by Castro et al. (2004, 2006). Briefly, cells
were suspended in L-15 medium with 0.1% FCS at a
concentration of 2 � 107 viable cells per millilitre
(determined by the trypan blue exclusion test). The
medium was carefully removed before starting the
experiments to obtain a mixed culture of leucocytes,
containing about 50% of peroxidase-positive cells
(neutrophils and neutrophil precursors) and 12% non-
esterase positive cells (monocytes–macrophages).
Before carrying out the experiments, the mean number
of cells per well was estimated by counting the number
of nuclei present after lysing the cells in several wells
with 100 ml lysis buffer containing 0.1 M citric acid, 1%
Tween 20 and 0.05% crystal violet (Castro et al., 2004).
Kidney leucocyte pellets containing 109 cells were
suspended in 8 ml ice-cold disruption buffer [50 mM
Tris–HCl, pH 7.5, containing 0.5 M NaCl, 1 mM
EDTA, and proteinase inhibitors (100 mg/ml phenyl-
methylsulphonyl fluoride (PMSF), 2 mg/ml aprotinin,
2 mg/ml leupeptin)], and were lysed by sonication
(eight bursts each of 1 min duration) on ice. The lysate
was then centrifuged at 12,000 � g for 30 min, and the
supernatant was stored at �80 8C until use. The protein
concentration of the supernatant was determined with
the Bio-Rad Protein Assay kit (Bio-Rad Laboratories,
Germany), as indicated above.
2.5. Purification of myeloperoxidase (MPO)
Turbot neutrophil MPO was purified from KL lysates
on a chromatography column containing acidic
sulphated polysaccharides obtained from U. rigida,
and epoxi-activated Sepharose 6B (GE Healthcare), as
previously described (Castro et al., 2008). Kidney
leucocyte lysates were applied to a column
(1 cm � 8 cm) in 50 mM Tris–HCl, pH 7.5, and eluted
with the same buffer containing different amounts of
NaCl (between 0.5 and 1.0 M NaCl). Eluted fractions
were combined, dialyzed against 20 mM sodium
phosphate buffer pH 7.0, concentrated by Amicon
and lyophilized.
2.6. Peroxidase activity
Enzyme assay peroxidase activity was measured by
spectrophotometry, with the chromogen ortho-pheny-
lenediamine (OPD, Sigma–Aldrich), as previously
described (Castro et al., 2008). Purified MPO and KL
lysates diluted in a solution of OPD (0.4 mg/ml in
0.05 M phosphate–citrate buffer, pH 5.0) were incu-
bated with H2O2 (0.4 ml/ml of commercial solution).
After 30 min incubation at room temperature, the
reaction was stopped by adding 3N sulphuric acid, and
the absorbance was determined at 492 nm. In order to
determine whether the RESV had a reversible or
irreversible inhibitory effect, the purified MPO was
incubated for 1 h with RESVat 100 mM and the enzyme
was then separated from the polyphenol by filtration in a
Microcon YM-10 Centrifugal Filter Unit (Millipore).
Filtration units were centrifuged at 10,000 � g for
10 min, the filtrate was then discarded and the enzyme
retained on the membrane was redissolved in PBS to the
original volume and the enzymatic activity determined
as described above.
2.7. Migration assay
Kidney leucocytes (2 � 105 cells) in L-15 medium
with no FCS, but containing 0.2 mg/ml BSA (Sigma–
Aldrich) were seeded in the upper chamber of a tissue
culture plate well insert with PCF membrane of pore
diameter 3 mm (Tissue culture-treated IsosporeTM
track-etched PVP-free polycarbonate; Millipore,
USA). The lower chamber was filled with 1 ml of L-
15 medium containing 2 mg/ml BSA and the stimulus,
i.e., ascitic fluid or P. dicentrarchi proteases. After
R. Castro et al. / Veterinary Immunology and Immunopathology 126 (2008) 9–19 11

Author's personal copy
incubation of plates for 24 h at 18 8C, migrated cells
were scraped from the lower surface of the membrane
with a cell scraper (Nunc) and then suspended in the
medium in the lower chamber and all migrating cells
(both adherent and those in suspensions) were counted
in a haemocytometer.
2.8. Respiratory burst activity
Extracellular reactive oxygen species produced in
the respiratory burst assays were quantified by the use of
the fluorogenic reagent OxyBURST Green dihydro-
20,4,5,6,7,70-hexafluorofluorescein/BSA (Molecular
Probes), as previously described (Leiro et al.,
2004a,b). Assays were performed in 96-well micro-
culture plates by adding 100 ml of a suspension
containing 106 KL/ml of DMEM medium without
phenol red (Sigma–Aldrich). Cells were then incubated
with 10 mg/ml of fluorescent probe for 2 min at 23 8Cand stimulated by addition of 1 mg/ml of PMA. In some
assays, the medium contained RESV or L(+) ascorbic
acid. Fluorescence emission was then determined in a
microplate fluorescent reader (Bio-Tek instrument;
excitation at 488 nm, detection 530 nm, 55 min). The
results are expressed as increase in fluorescence
(arbitrary units) per minute.
2.9. Assay for measuring extracellular MPO
activity
The activity of extracellular MPO released by
incubation of turbot KL with PMA (100 ng/ml) was
determined in 96-well flat bottom microtitre plates. Test
wells received 100 ml of 106 KL incubated with 0.1 mg/
ml of PMA and RESV (100, 50 and 25 mM) in HBSS.
Control wells received only PMA and DMSO (10 mM)
and plates were incubated for 1 h at 21 8C. After
incubation, the plates were centrifuged at 600 � g for
5 min, and 89 ml of supernatant from each well was
transferred to another plate and mixed with 10 ml of a
solution of OPD (4 mg/ml in 0.05 M phosphate–citrate
buffer, pH 5.0) and 1 ml of H2O2 at 0.5%. After 30 min
incubation at room temperature, the reaction was
stopped by adding 25 ml of 3N sulphuric acid, and
the absorbance was determined at 492 nm.
2.10. Intracellular MPO activity
The intracellular MPO activity was visualized
microscopically as previously described (Leiro et al.,
2000). Kidney leucocyte cells were incubated with
RESV (100, 50 or 25 mM) for 1 h at 21 8C. After
incubation, cell suspensions were smeared on micro-
scope slides, fixed for 1 min in PBS containing 1%
formaldehyde and 2% glutaraldehyde, and the MPO
activity was determined by incubating the cells for
15 min with 0.02% of 3,30-diaminobenzidine tetrahy-
drochloride (DAB) and 0.006% H2O2 in 0.15 M Tris
buffer pH 7.6. The slides were then washed three times
with Tris buffer and cells were stained with haemotoxylin
for 4 min. After dehydration with ethanol, permanent
mounts were prepared with Eukitt. Stained cells (dark
brown cytoplasm) were counted as peroxidase-positive.
2.11. Determination of PGE2 in culture
supernatants
Aliquots (100 ml) of 107 KL/ml were incubated for
24 h at 18 8C with 100 mg/ml of ASP in the presence or
absence of indomethacin (10 mg/ml), a non-selective
inhibitor of cyclooxygenase-2-enzymatic activity, in 96-
well microculture plates (Leiro et al., 2007). The plates
were centrifuged at 400� g for 5 min, and prostaglandin
E2 (PGE2) levels were determined in the supernatant by a
competitive binding immunoassay (R&D Systems), by
following the manufacturer’s instructions. Briefly,
100 ml of PGE2 standard (dilutions from 1000 to
7.8 pg/ml in assay buffer containing blocking proteins
and preservative, buffer ED1) or samples (in assay buffer)
were incubated for 18 h at 4 8C with 50 ml of PGE2
conjugated to alkaline phosphatase (PGE2HS) and 50 ml
of mouse monoclonal antibody against PGE2. The wells
were washed three times with wash buffer, then 200 ml of
p-nitrophenylphosphate ( pNPP) in buffered solution
were added to each well, the plates were incubated for 1 h
at 37 8C and then the reaction stopped with 50 ml of
trisodium phosphate (TSP) solution. The optical density
(OD) was determined in a microplate reader (Titertek
Multiscan, Flow Laboratories) at 405 nm, with wave-
length correction between 570 and 590 nm. To estimate
the concentrations of PGE2 in samples, a calibration
curve of PGE2 concentration against 100B/B0 was
constructed (B: average OD for that concentration minus
non-specific binding; B0: average OD in wells without
PGE2HS minus non-specific binding; non-specific
binding: average OD in wells without either PGE2HS
or anti-PGE2 antibody).
2.12. Isolation of RNA, production of cDNA and
reverse transcriptase-polymerase chain reaction
(RT-PCR)
Total RNA without any contaminating DNA from
KL samples (107 cells/ml) was isolated with an mRNA
R. Castro et al. / Veterinary Immunology and Immunopathology 126 (2008) 9–1912

Author's personal copy
isolation kit (NucleoSpin1; Macherey-Nagel), in
accordance with the manufacturer’s instructions. The
resulting RNA was dried and dissolved in diethylpyr-
ocarbonate (DEPC)-treated RNase-free water at a
concentration of 1 mg/ml. cDNA synthesis (25 ml/
reaction) was achieved with 1.25 mM random hexamer
primers (Roche) as described previously (Leiro et al.,
2003). One microlitre of cDNA was amplified by PCR
with the MPO forward/reverse primer pair, 50-CAA-
GAGAGAGCAGATCAACGC-30/50-GGCTGGATGG-
CCAAGTGG-30 (GenBank of NCBI accession number
EF112175) and turbot b-actin forward/reverse primer
pair, 50-AACTGGGATGACATGGAGA-30/50-GTCAG-
GATCTTCATGAGGTA-30 (NCBI, accession number
AY0083055) as a control for gene expression (Castro
et al., 2008). These primers amplified fragments of
567 bp (MPO) and 330 bp (b-actin). Thermal cycling
conditions were as follows: initial denaturating at 94 8Cfor 5 min; 30 cycles at 94 8C for 30 s, 65 8C (MPO) or
52 8C (b-actin) for 45 s, and 72 8C for 1 min and finally,
a 7-min extension phase at 72 8C. In all experiments,
RT-PCR controls (without RNA or without RT) were
included; no amplification products were obtained in
any of the experiments. Aliquots (20 ml) of PCR
products were separated on a 2% agarose gel, stained
with ethidium bromide and photographed with a digital
camera. Bands were quantified in tagged image file
format (TIFF) with densitometry analysis software
(ImageMaster Total Laboratory, version 2.00, GE
Healthcare).
2.13. Statistical analysis
Results shown are expressed as means � standard
error of the mean (S.E.M.). Statistical significances
(a = 0.05) were determined by one-way analysis of
variance (ANOVA) followed by Tukey–Kramer test for
multiple comparisons.
3. Results
3.1. Effect of RESV on migration of turbot kidney
leucocytes
The effect of RESV on migration of turbot KL
stimulated with ascitic fluid obtained from turbot
infected experimentally with the pathogen ciliate P.
dicentrarchi was evaluated, and the results obtained
after adding RESV or parasite proteases to the fluid are
shown in Fig. 1. At concentrations higher than 25 mM,
RESV had a significant inhibitory effect on migration of
KL stimulated with ascitic fluid. This inhibition was
equivalent to that obtained with cysteine proteases
obtained from P. dicentrarchi (Fig. 1).
3.2. Effect of RESV on respiratory burst of turbot
kidney leucocytes stimulated with phorbol myristate
acetate (PMA)
The KL stimulated with PMA displayed a highly
significant increase in the extracellular production of
ROS. However, the levels of extracellular ROS
decreased significantly when cells were incubated with
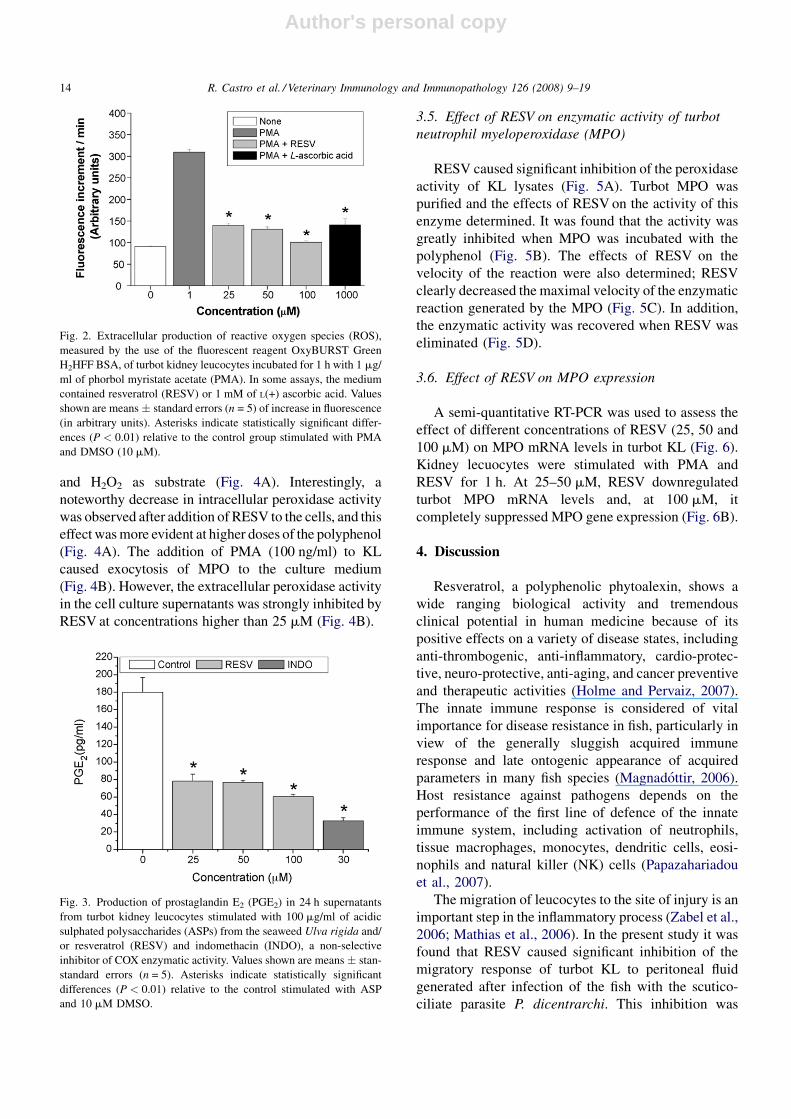
RESV at concentrations higher than 25 mM. This
decrease was equivalent to that induced by L(+)
ascorbic acid, a scavenger of ROS, at 1 mM (Fig. 2).
3.3. Effect of RESV on PGE2 production by kidney
leucocytes stimulated with acidic sulphated
polysaccharides (ASPs)
The addition of ASP from the seaweed U. rigida
(50 mg/ml) caused an increase in the levels of PGE2 in
the supernatants of cultures of turbot KL (Fig. 3). At
concentrations higher than 25 mM, RESV significantly
inhibited the production of PGE2 (Fig. 3).
3.4. Effect of RESV on the intracellular and
extracellular activity of turbot kidney leucocyte
MPO
Only the neutrophil precursors and the neutrophils
showed intracellular peroxidase activity as determined
in kidney leucocytes by the use of DAB as chromogen
R. Castro et al. / Veterinary Immunology and Immunopathology 126 (2008) 9–19 13
Fig. 1. Migration activity of turbot kidney leucocytes (KLs) in
response to ascitic fluid from turbot infected experimentally with
Philasterides dicentrarchi and DMSO (10 mM) (control group), and to
ascitic fluid containing resveratrol (RESV) or P. dicentrarchi cysteine
proteases. Values are means � standard errors. Asterisks indicate
statistically significant differences (P < 0.01; n = 5) relative to the
control.
Author's personal copy
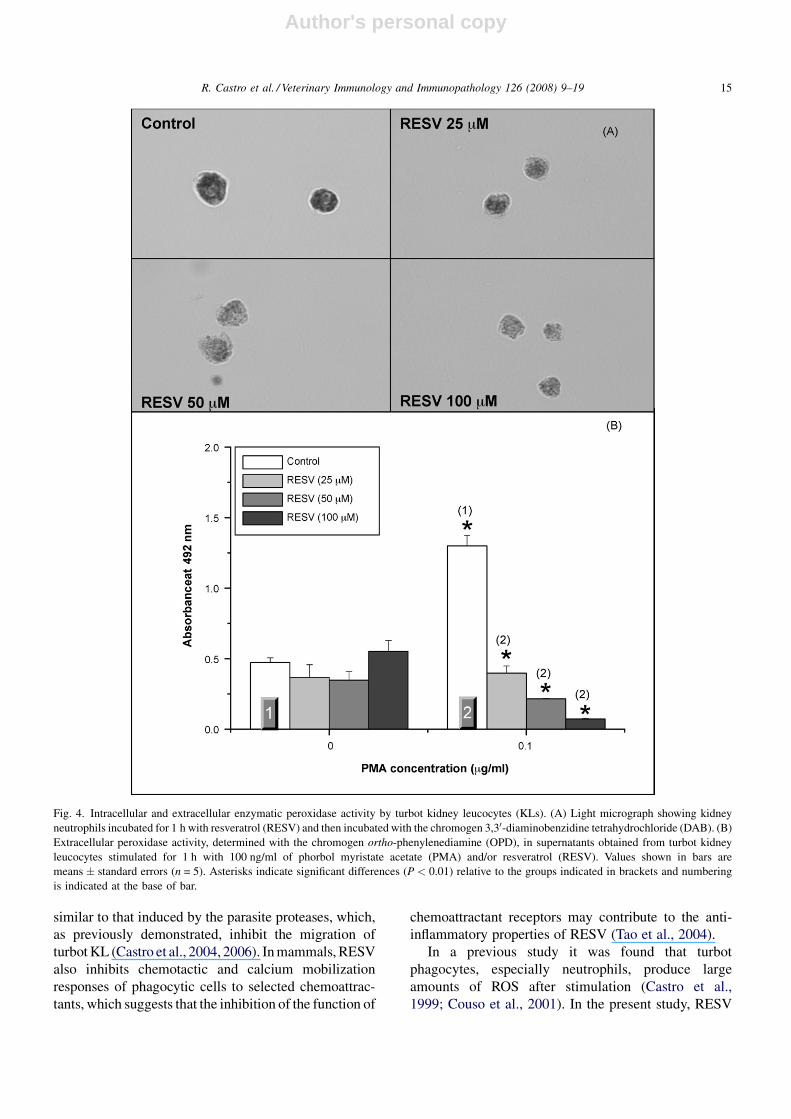
and H2O2 as substrate (Fig. 4A). Interestingly, a
noteworthy decrease in intracellular peroxidase activity
was observed after addition of RESV to the cells, and this
effect was more evident at higher doses of the polyphenol
(Fig. 4A). The addition of PMA (100 ng/ml) to KL
caused exocytosis of MPO to the culture medium
(Fig. 4B). However, the extracellular peroxidase activity
in the cell culture supernatants was strongly inhibited by
RESV at concentrations higher than 25 mM (Fig. 4B).
3.5. Effect of RESV on enzymatic activity of turbot
neutrophil myeloperoxidase (MPO)
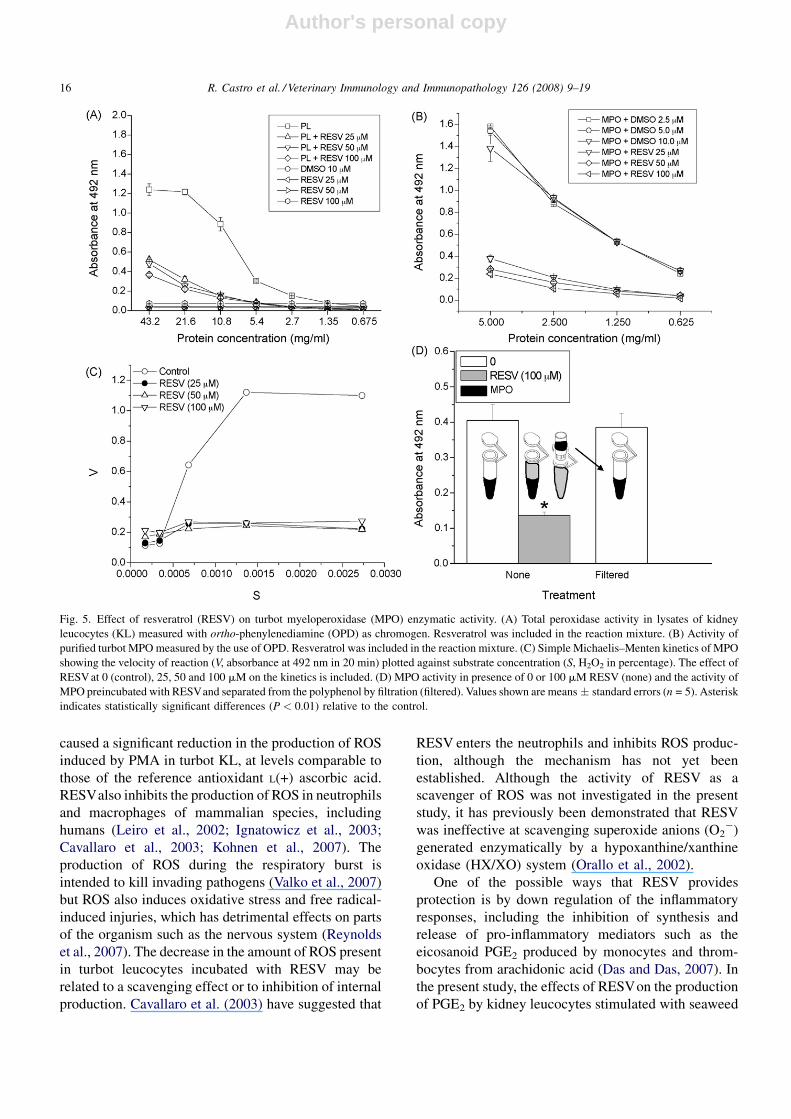
RESV caused significant inhibition of the peroxidase
activity of KL lysates (Fig. 5A). Turbot MPO was
purified and the effects of RESV on the activity of this
enzyme determined. It was found that the activity was
greatly inhibited when MPO was incubated with the
polyphenol (Fig. 5B). The effects of RESV on the
velocity of the reaction were also determined; RESV
clearly decreased the maximal velocity of the enzymatic
reaction generated by the MPO (Fig. 5C). In addition,
the enzymatic activity was recovered when RESV was
eliminated (Fig. 5D).
3.6. Effect of RESV on MPO expression
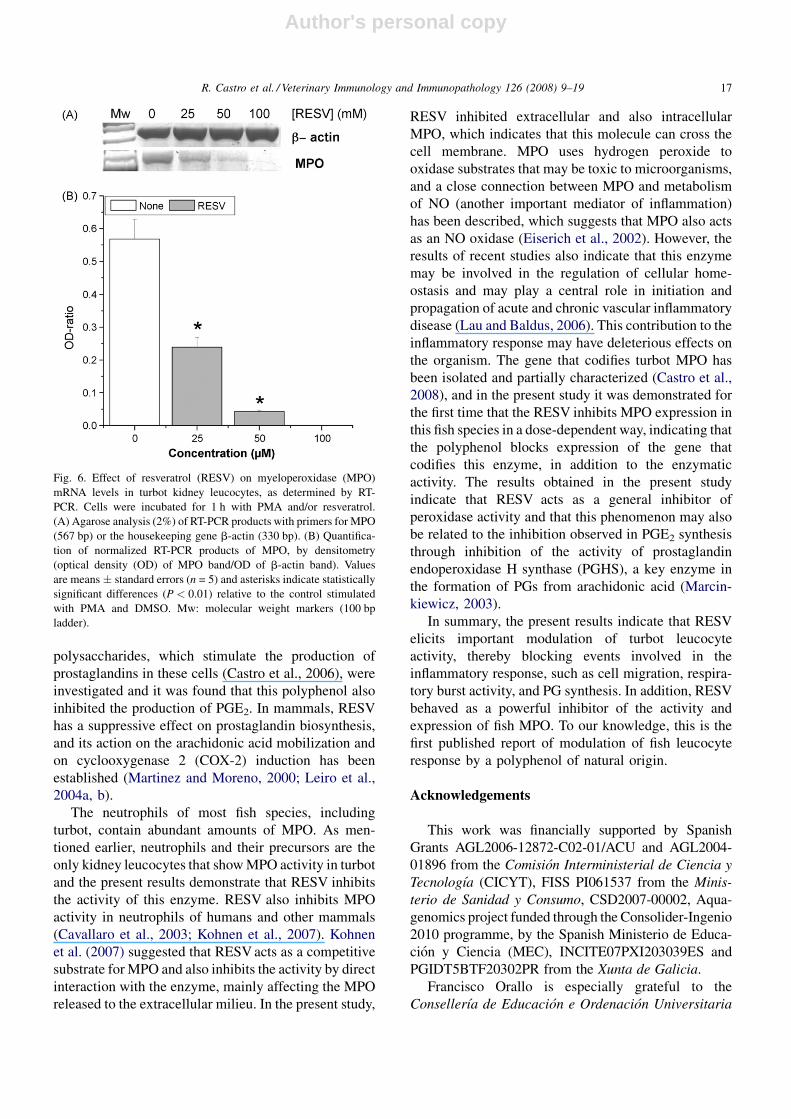
A semi-quantitative RT-PCR was used to assess the
effect of different concentrations of RESV (25, 50 and
100 mM) on MPO mRNA levels in turbot KL (Fig. 6).
Kidney lecuocytes were stimulated with PMA and
RESV for 1 h. At 25–50 mM, RESV downregulated
turbot MPO mRNA levels and, at 100 mM, it
completely suppressed MPO gene expression (Fig. 6B).
4. Discussion
Resveratrol, a polyphenolic phytoalexin, shows a
wide ranging biological activity and tremendous
clinical potential in human medicine because of its
positive effects on a variety of disease states, including
anti-thrombogenic, anti-inflammatory, cardio-protec-
tive, neuro-protective, anti-aging, and cancer preventive
and therapeutic activities (Holme and Pervaiz, 2007).
The innate immune response is considered of vital
importance for disease resistance in fish, particularly in
view of the generally sluggish acquired immune
response and late ontogenic appearance of acquired
parameters in many fish species (Magnadottir, 2006).
Host resistance against pathogens depends on the
performance of the first line of defence of the innate
immune system, including activation of neutrophils,
tissue macrophages, monocytes, dendritic cells, eosi-
nophils and natural killer (NK) cells (Papazahariadou
et al., 2007).
The migration of leucocytes to the site of injury is an
important step in the inflammatory process (Zabel et al.,
2006; Mathias et al., 2006). In the present study it was
found that RESV caused significant inhibition of the
migratory response of turbot KL to peritoneal fluid
generated after infection of the fish with the scutico-
ciliate parasite P. dicentrarchi. This inhibition was
R. Castro et al. / Veterinary Immunology and Immunopathology 126 (2008) 9–1914
Fig. 2. Extracellular production of reactive oxygen species (ROS),
measured by the use of the fluorescent reagent OxyBURST Green
H2HFF BSA, of turbot kidney leucocytes incubated for 1 h with 1 mg/
ml of phorbol myristate acetate (PMA). In some assays, the medium
contained resveratrol (RESV) or 1 mM of L(+) ascorbic acid. Values
shown are means � standard errors (n = 5) of increase in fluorescence
(in arbitrary units). Asterisks indicate statistically significant differ-
ences (P < 0.01) relative to the control group stimulated with PMA
and DMSO (10 mM).
Fig. 3. Production of prostaglandin E2 (PGE2) in 24 h supernatants
from turbot kidney leucocytes stimulated with 100 mg/ml of acidic
sulphated polysaccharides (ASPs) from the seaweed Ulva rigida and/
or resveratrol (RESV) and indomethacin (INDO), a non-selective
inhibitor of COX enzymatic activity. Values shown are means � stan-
standard errors (n = 5). Asterisks indicate statistically significant
differences (P < 0.01) relative to the control stimulated with ASP
and 10 mM DMSO.
Author's personal copy
similar to that induced by the parasite proteases, which,
as previously demonstrated, inhibit the migration of
turbot KL (Castro et al., 2004, 2006). In mammals, RESV
also inhibits chemotactic and calcium mobilization
responses of phagocytic cells to selected chemoattrac-
tants, which suggests that the inhibition of the function of
chemoattractant receptors may contribute to the anti-
inflammatory properties of RESV (Tao et al., 2004).
In a previous study it was found that turbot
phagocytes, especially neutrophils, produce large
amounts of ROS after stimulation (Castro et al.,
1999; Couso et al., 2001). In the present study, RESV
R. Castro et al. / Veterinary Immunology and Immunopathology 126 (2008) 9–19 15
Fig. 4. Intracellular and extracellular enzymatic peroxidase activity by turbot kidney leucocytes (KLs). (A) Light micrograph showing kidney
neutrophils incubated for 1 h with resveratrol (RESV) and then incubated with the chromogen 3,30-diaminobenzidine tetrahydrochloride (DAB). (B)
Extracellular peroxidase activity, determined with the chromogen ortho-phenylenediamine (OPD), in supernatants obtained from turbot kidney
leucocytes stimulated for 1 h with 100 ng/ml of phorbol myristate acetate (PMA) and/or resveratrol (RESV). Values shown in bars are
means � standard errors (n = 5). Asterisks indicate significant differences (P < 0.01) relative to the groups indicated in brackets and numbering
is indicated at the base of bar.
Author's personal copy
caused a significant reduction in the production of ROS
induced by PMA in turbot KL, at levels comparable to
those of the reference antioxidant L(+) ascorbic acid.
RESValso inhibits the production of ROS in neutrophils
and macrophages of mammalian species, including
humans (Leiro et al., 2002; Ignatowicz et al., 2003;
Cavallaro et al., 2003; Kohnen et al., 2007). The
production of ROS during the respiratory burst is
intended to kill invading pathogens (Valko et al., 2007)
but ROS also induces oxidative stress and free radical-
induced injuries, which has detrimental effects on parts
of the organism such as the nervous system (Reynolds
et al., 2007). The decrease in the amount of ROS present
in turbot leucocytes incubated with RESV may be
related to a scavenging effect or to inhibition of internal
production. Cavallaro et al. (2003) have suggested that
RESV enters the neutrophils and inhibits ROS produc-
tion, although the mechanism has not yet been
established. Although the activity of RESV as a
scavenger of ROS was not investigated in the present
study, it has previously been demonstrated that RESV
was ineffective at scavenging superoxide anions (O2�)
generated enzymatically by a hypoxanthine/xanthine
oxidase (HX/XO) system (Orallo et al., 2002).
One of the possible ways that RESV provides
protection is by down regulation of the inflammatory
responses, including the inhibition of synthesis and
release of pro-inflammatory mediators such as the
eicosanoid PGE2 produced by monocytes and throm-
bocytes from arachidonic acid (Das and Das, 2007). In
the present study, the effects of RESVon the production
of PGE2 by kidney leucocytes stimulated with seaweed
R. Castro et al. / Veterinary Immunology and Immunopathology 126 (2008) 9–1916
Fig. 5. Effect of resveratrol (RESV) on turbot myeloperoxidase (MPO) enzymatic activity. (A) Total peroxidase activity in lysates of kidney
leucocytes (KL) measured with ortho-phenylenediamine (OPD) as chromogen. Resveratrol was included in the reaction mixture. (B) Activity of
purified turbot MPO measured by the use of OPD. Resveratrol was included in the reaction mixture. (C) Simple Michaelis–Menten kinetics of MPO
showing the velocity of reaction (V, absorbance at 492 nm in 20 min) plotted against substrate concentration (S, H2O2 in percentage). The effect of
RESV at 0 (control), 25, 50 and 100 mM on the kinetics is included. (D) MPO activity in presence of 0 or 100 mM RESV (none) and the activity of
MPO preincubated with RESVand separated from the polyphenol by filtration (filtered). Values shown are means � standard errors (n = 5). Asterisk
indicates statistically significant differences (P < 0.01) relative to the control.
Author's personal copy
polysaccharides, which stimulate the production of
prostaglandins in these cells (Castro et al., 2006), were
investigated and it was found that this polyphenol also
inhibited the production of PGE2. In mammals, RESV
has a suppressive effect on prostaglandin biosynthesis,
and its action on the arachidonic acid mobilization and
on cyclooxygenase 2 (COX-2) induction has been
established (Martinez and Moreno, 2000; Leiro et al.,
2004a, b).
The neutrophils of most fish species, including
turbot, contain abundant amounts of MPO. As men-
tioned earlier, neutrophils and their precursors are the
only kidney leucocytes that show MPO activity in turbot
and the present results demonstrate that RESV inhibits
the activity of this enzyme. RESV also inhibits MPO
activity in neutrophils of humans and other mammals
(Cavallaro et al., 2003; Kohnen et al., 2007). Kohnen
et al. (2007) suggested that RESV acts as a competitive
substrate for MPO and also inhibits the activity by direct
interaction with the enzyme, mainly affecting the MPO
released to the extracellular milieu. In the present study,
RESV inhibited extracellular and also intracellular
MPO, which indicates that this molecule can cross the
cell membrane. MPO uses hydrogen peroxide to
oxidase substrates that may be toxic to microorganisms,
and a close connection between MPO and metabolism
of NO (another important mediator of inflammation)
has been described, which suggests that MPO also acts
as an NO oxidase (Eiserich et al., 2002). However, the
results of recent studies also indicate that this enzyme
may be involved in the regulation of cellular home-
ostasis and may play a central role in initiation and
propagation of acute and chronic vascular inflammatory
disease (Lau and Baldus, 2006). This contribution to the
inflammatory response may have deleterious effects on
the organism. The gene that codifies turbot MPO has
been isolated and partially characterized (Castro et al.,
2008), and in the present study it was demonstrated for
the first time that the RESV inhibits MPO expression in
this fish species in a dose-dependent way, indicating that
the polyphenol blocks expression of the gene that
codifies this enzyme, in addition to the enzymatic
activity. The results obtained in the present study
indicate that RESV acts as a general inhibitor of
peroxidase activity and that this phenomenon may also
be related to the inhibition observed in PGE2 synthesis
through inhibition of the activity of prostaglandin
endoperoxidase H synthase (PGHS), a key enzyme in
the formation of PGs from arachidonic acid (Marcin-
kiewicz, 2003).
In summary, the present results indicate that RESV
elicits important modulation of turbot leucocyte
activity, thereby blocking events involved in the
inflammatory response, such as cell migration, respira-
tory burst activity, and PG synthesis. In addition, RESV
behaved as a powerful inhibitor of the activity and
expression of fish MPO. To our knowledge, this is the
first published report of modulation of fish leucocyte
response by a polyphenol of natural origin.
Acknowledgements
This work was financially supported by Spanish
Grants AGL2006-12872-C02-01/ACU and AGL2004-
01896 from the Comision Interministerial de Ciencia y
Tecnologıa (CICYT), FISS PI061537 from the Minis-
terio de Sanidad y Consumo, CSD2007-00002, Aqua-
genomics project funded through the Consolider-Ingenio
2010 programme, by the Spanish Ministerio de Educa-
cion y Ciencia (MEC), INCITE07PXI203039ES and
PGIDT5BTF20302PR from the Xunta de Galicia.
Francisco Orallo is especially grateful to the
Consellerıa de Educacion e Ordenacion Universitaria
R. Castro et al. / Veterinary Immunology and Immunopathology 126 (2008) 9–19 17
Fig. 6. Effect of resveratrol (RESV) on myeloperoxidase (MPO)
mRNA levels in turbot kidney leucocytes, as determined by RT-
PCR. Cells were incubated for 1 h with PMA and/or resveratrol.
(A) Agarose analysis (2%) of RT-PCR products with primers for MPO
(567 bp) or the housekeeping gene b-actin (330 bp). (B) Quantifica-
tion of normalized RT-PCR products of MPO, by densitometry
(optical density (OD) of MPO band/OD of b-actin band). Values
are means � standard errors (n = 5) and asterisks indicate statistically
significant differences (P < 0.01) relative to the control stimulated
with PMA and DMSO. Mw: molecular weight markers (100 bp
ladder).

Author's personal copy
de la Xunta de Galicia (Spain) for granting him
financial support for intensification of research activity
and reduction of teaching activity during the academic
year 2007–2008 [Programa de promocion de intensi-
ficacion de la actividad investigadora en el sistema
Universitario de Galicia (SUG)].
References
Castro, R., Couso, N., Obach, A., Lamas, J., 1999. Effect of different
b-glucans on the respiratory burst of turbot (Psetta maxima) and
gilthead seabream (Sparus aurata) phagocytes. Fish Shellfish
Immunol. 9, 529–541.
Castro, R., Piazzon, M.C., Noya, M., Leiro, J.M., Lamas, J., 2008.
Isolation and molecular cloning of a fish myeloperoxidase. Mol.
Immunol. 45, 428–437.
Castro, R., Piazzon, M.C., Zarra, I., Leiro, J., Noya, M., Lamas, J.,
2004. Stimulation of turbot phagocytes by Ulva rigida C. Agardh
polysaccharides. Aquaculture 254, 9–20.
Castro, R., Zarra, I., Lamas, J., 2006. Water-soluble seaweed extracts
modulate the respiratory burst activity of turbot phagocytes.
Aquaculture 229, 67–78.
Cavallaro, A., Ainis, T., Bottari, C., Fimiani, V., 2003. Effect of
resveratrol on some activities of isolated and in whole blood
human neutrophils. Physiol. Res. 52, 555–562.
Chen, Y., Tseng, S.H., 2007. Review. Pro- and anti-angiogenesis
effects of resveratrol. In Vivo 21, 365–370.
Collazos, M.E., Barriga, C., Ortega, E., 1995. Effect of high summer
temperatures upon granulocyte phagocytic function of the tench
(Tinca tinca L.). Comp. Immunol. Microbiol. Infect. Dis. 18,
115–121.
Couso, N., Castro, R., Noya, M., Lamas, J., 2001. Location of
superoxide production sites in turbot neutrophils and gilthead
seabream acidophilic granulocytes during phagocytosis of glucan
particles. Dev. Comp. Immunol. 25, 607–618.
Das, S., Das, D.K., 2007. Anti-inflammatory responses of resveratrol.
Inflamm. Allergy Drug Targets 6, 168–173.
Eiserich, J.P., Baldus, S., Brennan, M.L., Ma, W., Zhang, C., Tousson,
A., Castro, L., Lusis, A.J., Nauseef, W.M., White, C.R., Freeman,
B.A., 2002. Myeloperoxidase, a leukocyte-derived vascular NO
oxidase. Science 296, 2391–2394.
Fraga, C.G., 2007. Plant polyphenols: how to translate their in vitro
antioxidant actions to in vivo conditions. IUBMB Life 59,
308–315.
Holme, A.L., Pervaiz, S., 2007. Resveratrol in cell fate decisions. J.
Bioenerg. Biomembr. 39, 59–63.
Iglesias, R., Parama, A., Alvarez, M.F., Leiro, J., Fernandez, J.,
Sanmartın, M.L., 2001. Philasterides dicentrarchi (Ciliophora,
Scuticociliatida) as the causative agent of scuticociliatosis in
farmed turbot Scophthalmus maximus in Galicia (NW Spain).
Dis. Aquat. Org. 46, 47–55.
Iglesias, R., Parama, A., Alvarez, M.F., Leiro, J., Ubeira, F.M.,
Sanmartın, M.L., 2003. Philasterides dicentrarchi (Ciliophora:S-
cuticociliatida) expresses surface immobilization antigens that
probably induce protective immune responses in turbot. Parasitol-
ogy 126, 125–134.
Ignatowicz, E., Balana, B., Vulimiri, S.V., Szaefer, H., Baer-Dubowska,
W., 2003. The effect of plant phenolics on the formation of 7,12-
dimethylbenz[a]anthracene–DNA adducts and TPA-stimulated
polymorphonuclear neutrophils chemiluminescence in vitro. Tox-
icology 189, 199–209.
Khanna, D., Sethi, G., Ahn, K.S., Pandey, M.K., Kunnumakkara, A.B.,
Sung, B., Aggarwal, A., Aggarwal, B.B., 2007. Natural products
as a gold mine for arthritis treatment. Curr. Opin. Pharmacol. 7,
344–351.
Kohnen, S., Franck, T., Van Antwerpen, P., Boudjeltia, K.Z.,
Mouithys-Mickalad, A., Deby, C., Moguilevsky, N., Deby-
Dupont, G., Lamy, M., Serteyn, D., 2007. Resveratrol inhibits
the activity of equine neutrophil myeloperoxidase by a direct
interaction with the enzyme. J. Agric. Food Chem. 55, 8080–8087.
Kumar, V., Abbas, A.K., Fausto, N., 2004. Robbins and Cotran
Pathologic Basis of Disease. Elsevier Saunders, Philadelphia, PA.
Kundu, J.K., Shin, Y.K., Surh, Y.J., 2004. Resveratrol modulates
phorbol ester-induced pro-inflammatory signal transduction path-
ways in mouse skin in vivo, NF-kappaB and AP-1 as prime targets.
Biochem. Pharmacol. 72, 1506–1515.
Lau, D., Baldus, S., 2006. Myeloperoxidase and its contributory role
in inflammatory vascular disease. Pharmacol. Ther. 111, 16–26.
Leiro, J., Alvarez, E., Arranz, J.A., Laguna, R., Uriarte, E., Orallo, F.,
2004a. Effects of cis-resveratrol on inflammatory murine macro-
phages: antioxidant activity and down-regulation of inflammatory
genes. J. Leukoc. Biol. 75, 1156–1165.
Leiro, J., Garcıa, D., Arranz, J.A., Delgado, R., Sanmartın, M.L.,
Orallo, F., 2004b. An Anacardiaceae preparation reduces the
expression of inflammation-related genes in murine macrophages.
Int. Immunopharmacol. 4, 991–1003.
Leiro, J., Alvarez, E., Garcıa, D., Orallo, F., 2002. Resveratrol
modulates rat macrophage functions. Int. Immunopharmacol. 2,
767–774.
Leiro, J., Arranz, J.A., Fraiz, N., Sanmartın, M.L., Quezada, E., Orallo,
F., 2005. Effect of cis-resveratrol on genes involved in nuclear
factor kappa B signalling. Int. Immunopharmacol. 5, 393–406.
Leiro, J., Castro, R., Arranz, J.A., Lamas, J., 2007. Immunomodulat-
ing activities of acidic sulphated polysaccharides obtained from
the seaweed Ulva rigida C. Agardh. Int. Immunopharmacol. 7,
879–888.
Leiro, J., Ortega, M., Sanmartın, M.L., Ubeira, F.M., 2000. Non-
specific responses of turbot (Scophthalmus maximus L.) adherent
cells to microsporidian spores. Vet. Immunol. Immunopathol. 75,
81–95.
Leiro, J.M., Alvarez, E., Arranz, J.A., Siso, I.G., Orallo, F., 2003. In
vitro effects of mangiferin on superoxide concentrations and
expression of the inducible nitric oxide synthase, tumour necrosis
factor-alpha and transforming growth factor-beta genes. Biochem.
Pharmacol. 65, 1361–1371.
Magnadottir, B., 2006. Innate immunity of fish (overview). Fish
Shellfish Immunol. 20, 137–151.
Marcinkiewicz, J., 2003. Prostanoids and MPO–halide system pro-
ducts as a link between innate and adaptive immunity. Immunol.
Lett. 89, 187–191.
Martinez, J., Moreno, J.J., 2000. Effect of resveratrol, a natural
polyphenolic compound, on reactive oxygen species and prosta-
glandin production. Biochem. Pharmacol. 59, 865–870.
Mathias, J.R., Perrin, B.J., Liu, T.X., Kanki, J., Look, A.T., Hutten-
locher, A., 2006. Resolution of inflammation by retrograde che-
motaxis of neutrophils in transgenic zebrafish. J. Leukoc. Biol. 80,
1281–1288.
Nam, N.H., 2006. Naturally occurring NF-kappaB inhibitors. Mini
Rev. Med. Chem. 6, 945–951.
Orallo, F., Alvarez, E., Camina, M., Leiro, J.M., Gomez, E., Fernan-
dez, P., 2002. The possible implication of trans-resveratrol in the
R. Castro et al. / Veterinary Immunology and Immunopathology 126 (2008) 9–1918

Author's personal copy
cardioprotective effects of long-term moderate wine consumption.
Mol. Pharmacol. 61, 294–302.
Papazahariadou, M., Athanasiadis, G.I., Papadopoulos, E., Symeoni-
dou, I., Hatzistilianou, M., Castellani, M.L., Bhattacharya, K.,
Shanmugham, L.N., Conti, P., Frydas, S., 2007. Involvement of
NK cells against tumors and parasites. Int. J. Biol. Markers 22,
144–153.
Parama, A., Castro, R., Arranz, J.A., Sanmartın, M.L., Lamas, J.,
Leiro, J., 2007. Scuticociliate cysteine proteinases modulate turbot
leucocyte functions. Fish Shellfish Immunol. 23, 945–956.
Parama, A., Iglesias, R., Alvarez, M.F., Leiro, J., Ubeira, F.M.,
Sanmartın, M.L., 2004a. Cysteine proteinase activities in the fish
pathogen Philasterides dicentrarchi (Ciliophora: Scuticocilia-
tida). Parasitology 128, 541–548.
Parama, A., Iglesias, R., Alvarez, M.F., Sanmartın, M.L., Leiro, J.,
2004b. Chemotactic responses of the fish-parasitic scuticociliate
Philasterides dicentrarchi to blood and blood components of the
turbot Scophthalmus maximus, evaluated using a new microplate
multiassay. J. Microbiol. Methods 58, 361–366.
Pedrera, I.M., Collazos, M.E., Ortega, E., Barriga, C., 1992. In vitro
study of the phagocytic processes in splenic granulocytes of the
tench (Tinca tinca L.). Dev. Comp. Immunol. 16, 431–439.
Plouffe, D.A., Hanington, P.C., Walsh, J.G., Wilson, E.C., Belosevic,
M., 2005. Comparison of select innate immune mechanisms of fish
and mammals. Xenotransplantation 12, 266–277.
Reynolds, A., Laurie, C., Mosley, R.L., Gendelman, H.E., 2007.
Oxidative stress and the pathogenesis of neurodegenerative dis-
orders. Int. Rev. Neurobiol. 82, 297–325.
Stakauskas, R., Schuberth, H.J., Leibold, W., Steinhagen, D., 2007.
Modulation of carp (Cyprinus carpio) neutrophil functions during
an infection with the haemoparasite Trypanoplasma borreli. Fish
Shellfish Immunol. 23, 446–458.
Tao, H.Y., Wu, C.F., Zhou, Y., Gong, W.H., Zhang, X., Iribarren, P.,
Zhao, Y.Q., Le, Y.Y., Wang, J.M., 2004. The grape component
resveratrol interferes with the function of chemoattractant
receptors on phagocytic leukocytes. Cell. Mol. Immunol. 1,
50–56.
Valenzano, D.R., Cellerino, A., 2006. Resveratrol and the pharmacol-
ogy of aging: a new vertebrate model to validate an old molecule.
Cell Cycle 5, 1027–1032.
Valko, M., Leibfritz, D., Moncol, J., Cronin, M.T., Mazur, M.,
Telser, J., 2007. Free radicals and antioxidants in normal phy-
siological functions and human disease. Int. J. Biochem. Cell
Biol. 39, 44–84.
Whyte, S.K., 2007. The innate immune response of finfish—a review
of current knowledge. Fish Shellfish Immunol. 23, 1127–1151.
Zabel, B.A., Zuniga, L., Ohyama, T., Allen, S.J., Cichy, J., Handel,
T.M., Butcher, E.C., 2006. Chemoattractants, extracellular pro-
teases, and the integrated host defense response. Exp. Hematol. 34,
1021–1032.
R. Castro et al. / Veterinary Immunology and Immunopathology 126 (2008) 9–19 19




















