
A Secretory Golgi Bypass Route to the Apical Surface Domain of Epithelial MDCK Cells

Printed from the CJO service for personal use only by...
New Phytol. (1998), 140, 581–595
Development of the leafy shoot in
Sphagnum (Bryophyta) involves the activity
of both apical and subapical meristems
B ROBERTO LIGRONE" JEFFREY G. DUCKETT#,*
"Facolta[ di Scienze Ambientali, Seconda Universita[ di Napoli, via Arena 22, 81100
Caserta, Italy
#School of Biological Sciences, Queen Mary & Westfield College, Mile End Road,
London E1 4NS, UK
(Received 1 August 1998; accepted 19 August 1998)
This light- and electron-microscope study of four species of Sphagnum reveals that stem elongation involves
meristematic activities unique to the group and hitherto unrecognized. The internal tissue of the mature stem
arises by the concerted activity of an apical (primary) and a subapical (secondary) meristem. The primary
meristem comprises the immediate derivatives of the single apical cell. Following a small number of divisions, the
primary derivatives differentiate into highly vacuolate parenchymatous cells with a storied arrangement.
Subsequently, the large vacuoles are replaced by numerous small vacuoles and the cells then divide repeatedly, by
transverse septa, producing files of about nine short cells. Finally, ninefold elongation of these secondary cells is
responsible for extension growth of the main stem below the mature capitulum. An early step in primary
differentiation is the confinement of pre-existing plasmodesmata to distinct pitted areas. Further enlargement of
the cells during primary and secondary differentiation involves the thickening of non-pitted wall areas, followed
by expansion and thinning out, while the pitted areas remain virtually unchanged. A cortical array of microtubules
is regularly found in association with non-pitted wall areas, while the unexpanded pitted areas are associated with
smooth endoplasmic reticulum showing continuity with desmotubules. Though sharing much the same cytology
as the conducting cells in bryoid mosses, in terms of their development the central stem cells in Sphagnum are not
homologous with those of other mosses. The unique mode of stem development may be an important factor in the
ecological success of Sphagnum.
Key words: Cortical microtubules, differential wall expansion, pitted walls, plasmodesmata, Sphagnum, secondary
meristem.
The genus Sphagnum, the bog mosses, is widespread
in the world, being found on five continents at most
latitudes, with approx. 150–200 species (Gignac,
1993). Sphagnum is a major component of peatland
vegetation and considered in terms of its total mass,
live and dead, it must be regarded as one of the most
important of all plants (Clymo & Hayward, 1982).
Among the reasons for its remarkable biological
success are the ability to colonize oligotrophic wet
habitats and the ability to maintain a low pH that
prevents the growth of most other plants (Clymo &
Hayward, 1982; Clymo, 1984, 1987).
Sphagnum plants consist of leafy shoots of in-
determinate growth with a single apical cell. The
main shoot bears numerous lateral branches of
* To whom correspondence should be addressed.
E-mail : j.g.duckett!qmw.ac.uk
determinate growth arranged in groups (fascicles)
along the stem (Schimper, 1858; Cavers, 1911;
Smith, 1955). The young branches and leaves form
a compact hemispherical head, the capitulum,
around the apex. The growth pattern of lateral
branches varies according to the species and so does
the frequency by which the developing capitulum
forks to form two separate shoots of indeterminate
growth.
As a consequence of their unlimited apical growth
and repeated forking, the older parts of Sphagnum
plants become highly shaded and eventually die. The
only parts remaining alive at some distance from the
apex are groups of cortical cells in the main stem and
the branches, which are able to regenerate new
plants when several-year-old peat cores are exposed
to the light (Clymo & Duckett, 1986). Acropetal
translocation of nutrients from the older parts of the
shoot to the capitulum has been demonstrated with

Printed from the CJO service for personal use only by...
582 R. Ligrone and J. G. Duckett
the use of radioactive tracers and interpreted as
evidence that Sphagnum has the ability to relocate
solutes internally (Rydin & Clymo, 1989).
The mature stems of the leafy shoots of Sphagnum
comprise a cortical sheath of one-to-several layers of
dead hyaline cells, with one or more layers of cells
with thick walls (sterome) beneath and an internal
parenchymatous cylinder with no intercellular
spaces. Until very recently, the internal cylinder was
thought to show no cytological differentiation
(Schimper, 1858; Cavers, 1911). However, a recent
study (Ligrone & Duckett, 1998) has revealed that
the internal cells of the mature stem have a highly
specialized cytoplasmic organization including an
axial system of microtubules associated with the
nucleus and a range of organelles.
In the present paper we report a further suite of
features associated with the differentiation of the
leafy stems. These are unique to Sphagnum and have
remained unrecognized hitherto. We describe how
stem development occurs in two distinct phases: an
apical (primary) phase, involving differentiation of
derivatives of the apical cell within the capitulum;
and a wave of subapical (secondary) meristematic
activity. The latter is mainly responsible for stem
elongation below the capitulum. This account details
the cytological changes associated with the primary
and secondary meristems, with particular emphasis
on changes in the cell walls.
Living specimens of Sphagnum cuspidatum Ehrh., S.
palustre L., S. recurvum P. Beauv. and S. magel-
lanicum Bridel were collected from several field sites
in southern England during the spring and early
summer. The capitula were isolated and, after
removing the external leaves and branches, were
fixed in a mixture of 2% glutaraldehyde, 1%
formaldehyde (freshly prepared from paraformal-
dehyde) and 0±5% tannic acid in 0±05 Na-phosphate
buffer at pH 7. After 2 h at room temperature under
gentle vacuum the material was rinsed in 0±1 Na-
phosphate buffer and post-fixed overnight with 1%
osmium tetroxide in 0±1 Na-phosphate buffer,
pH 6±8, at 4 °C. The material was then dehydrated in
ethanol, transferred to propylene oxide and slowly
infiltrated with Spurr’s resin at 4 °C (Ligrone &
Duckett, 1994).
Thin sections, cut with a diamond knife, were
stained with 5% uranyl acetate in methanol for
15 min followed by Reynold’s lead citrate for 10 min
and observed with a Jeol 1200 EX2 or a Philips
CM12 electron microscope. One-µm-thick sections
were cut with glass knives, stained with 1% toluidine
blue in borax and photographed with a Leitz
Ortholux microscope.
A morphometric analysis was performed in S.
cuspidatum at seven developmental stages (Table 1).
Table
1.C
hanges
inth
ein
tern
alce
lls
of
the
leafy
stem
duri
ng
pri
mary
(sta
ges
1–4)
and
seco
ndary
(sta
ges
5–7)
diff
eren
tiation
inSphagnum
cusp
idatu
m
Develo
pm
enta
lst
age
Mic
rogra
phs
(Fig
ure
s)
Dia
gra
mm
atic
repre
senta
tions
(Fig
ure
s)C
ell
length
(µm
)C
ell
wid
th(µ
m)
Wall
thic
kness
inpitte
dare
as
(µm
)
Wall
thic
kness
innon-p
itte
dare
as
(µm
)L
ength
of
pitte
dare
as
(µm
)
Pla
smodesm
al
frequency
inlo
ngitudin
al
walls†
11a,2b
7a,9a
25³
48³
20±1
8³
0±0
3*
0±1
8³
0±0
3*
26±3
³6±4
22c–
e7b,9b
43³
610³
30±2
1³
0±0
50±2
8³
0±0
42±0
8³
0±5
615±6
³4±2
32d,g
7c,
9c
55³
613³
30±5
2³
0±0
61±1
2³
0±2
02±4
0³
0±7
012±5
³4±4
43a,b
7d,9d
75³
816³
20±3
9³
0±0
60±5
3³
0±0
82±3
7³
0±8
18±6
³2±6
54
7e,
8a,9e
78³
12
17³
20±4
3³
0±1
21±0
5³
0±2
02±5
0³
0±5
18±8
³1±9
65
7f–
i,8b–d,9f,
g20³
321³
40±6
7³
0±1
52±0
7³
0±5
02±4
1³
0±7
44±1
³2±2
76
7j,
8e,
9h
180³
45
21³
40±8
5³
0±1
80±8
0³
0±1
22±7
1³
0±8
30±7
³0±3
The
num
bers
are
means³
fr
om
at
least
25
measu
rem
ents
.
*N
ocle
ar
cut
dis
tinction
isvis
ible
at
this
stage
betw
een
pitte
dand
non-p
itte
dare
as.
†N
um
ber
of
pla
smodesm
ata
per
10
µm
wall
pro
file
inm
edia
nlo
ngitudin
alse
ctions
of
cells.

Printed from the CJO service for personal use only by...
Meristems and shoot development in Sphagnum 583
(a)(a)(a) (((ccc))) (((eee)))
(((bbb)))
(((ddd))) (((fff )))
Figure 1. Light microscopy of leafy stem development in Sphagnum. (a) Shoot apex of S. magellanicum,
showing the apical cell (ac) and its immediate derivatives. (b–d ) Longitudinal sections of the shoot apex in S.
magellanicum (b) and S. cuspidatum (c, d ), showing the region of primary differentiation and the onset of
secondary divisions (arrows in b and d ). (e) Higher magnification showing the highly pitted cells that initiate
secondary divisions (arrows) in S. magellanicum. ( f ) Detail of the subapical secondary meristem in S.
cuspidatum. Scale bars: a and f, 100 µm; b, c and d, 200 µm; e, 50 µm.
For each stage, at least 15 low-power electron
micrographs of median longitudinal sections of cells
were utilized. The nucleus was used as a marker of
sections passing through the median longitudinal
plane of cells. The cellular sizes were measured using
light microscopy.
The four species examined showed much the same
morphological organization and developmental pat-
tern. Therefore, except when indicated, the fol-
lowing description applies to all of them.
Printed from the CJO service for personal use only by...
584 R. Ligrone and J. G. Duckett
(a) (d ) (e )
(b) (c)
(g)
(f )
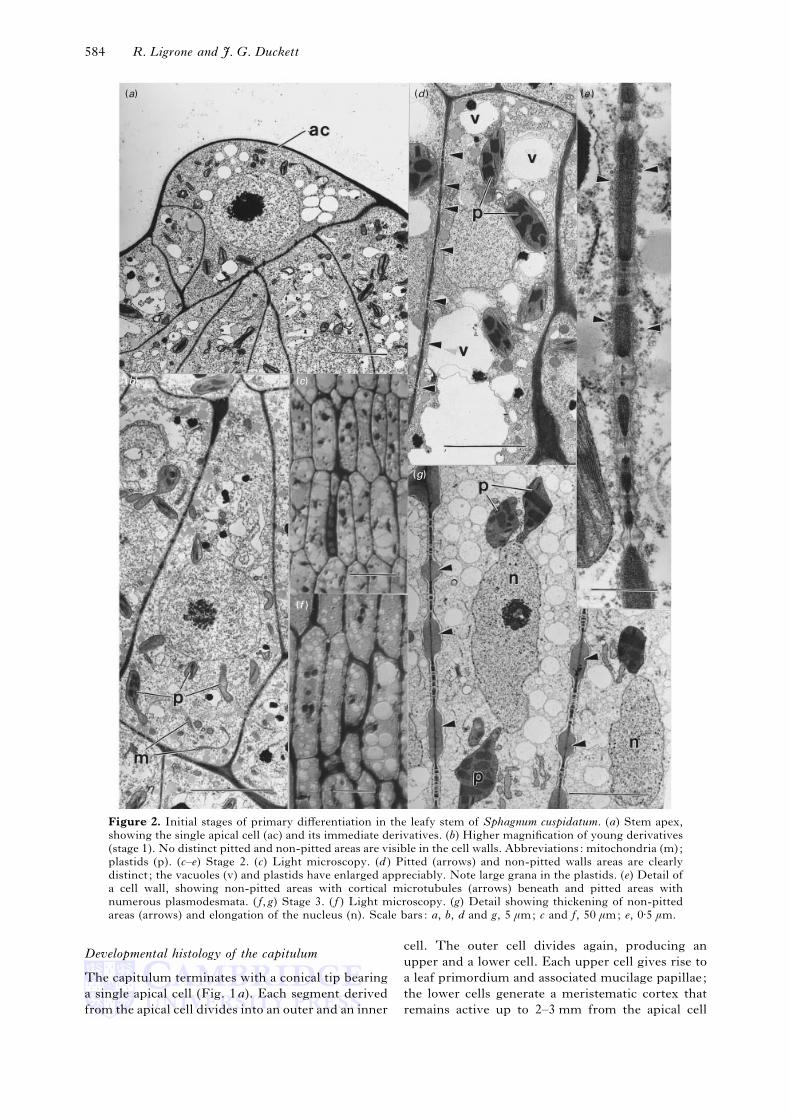
Figure 2. Initial stages of primary differentiation in the leafy stem of Sphagnum cuspidatum. (a) Stem apex,
showing the single apical cell (ac) and its immediate derivatives. (b) Higher magnification of young derivatives
(stage 1). No distinct pitted and non-pitted areas are visible in the cell walls. Abbreviations: mitochondria (m);
plastids (p). (c–e) Stage 2. (c) Light microscopy. (d ) Pitted (arrows) and non-pitted walls areas are clearly
distinct ; the vacuoles (v) and plastids have enlarged appreciably. Note large grana in the plastids. (e) Detail of
a cell wall, showing non-pitted areas with cortical microtubules (arrows) beneath and pitted areas with
numerous plasmodesmata. ( f, g) Stage 3. ( f ) Light microscopy. (g) Detail showing thickening of non-pitted
areas (arrows) and elongation of the nucleus (n). Scale bars: a, b, d and g, 5 µm; c and f, 50 µm; e, 0±5 µm.
Developmental histology of the capitulum
The capitulum terminates with a conical tip bearing
a single apical cell (Fig. 1a). Each segment derived
from the apical cell divides into an outer and an inner
cell. The outer cell divides again, producing an
upper and a lower cell. Each upper cell gives rise to
a leaf primordium and associated mucilage papillae;
the lower cells generate a meristematic cortex that
remains active up to 2–3 mm from the apical cell

Printed from the CJO service for personal use only by...
Meristems and shoot development in Sphagnum 585
(a) (c) (d ) (e )
(b )
(f )
Figure 3. Stem development in Sphagnum cuspidatum. (a, b) Final step of primary differentiation (stage 4). (a)
Light microscopy. (b) Detail showing a prominent vacuole displacing the nucleus (n) to the cell periphery; non-
pitted wall areas (arrows) are thinner and longer than in the previous stage. (c–f ) Initial stage in the
development of secondary meristematic cells. (c) Light microscopy. (d ) Low-power electron micrograph of
a cell containing numerous small vacuoles (v) with electron-opaque precipitates and the nucleus (n), which has
moved back to an internal position; pitted and non-pitted areas in the walls are of similar thickness. (e, f ) Cell
wall details ; the wall material in non-pitted areas is less opaque to electrons and shows a more compact fibrillar
texture than in non-pitted areas; arrows indicate cortical microtubules. Scale bars: a and c, 200 µm; b and d,
5 µm; e, 1 µm; f, 0±5 µm.

Printed from the CJO service for personal use only by...
586 R. Ligrone and J. G. Duckett
(a) (b) (c )
(d )
(f )(e )
Figure 4. Fully developed secondary meristematic cells in the leafy stem of Sphagnum (stage 5). (a, b) Light
microscopy in S. recurvum (a) and S. cuspidatum (b) ; note the heavily stained vacuoles (arrows). Abbreviations:
n, nucleus. (c, e) Details from S. palustre (c) and S. recurvum (e) ; the non-pitted wall areas (arrows) are again
thickened; the vacuoles (v) have become smaller and more numerous. (d ) Tangential section of a longitudinal
wall in S. magellanicum, showing a row of six pitted areas with a high concentration of plasmodesmata. ( f ) High
magnification of a pitted area in S. cuspidatum, showing a cortical network of smooth ER continuous with
desmotubules. Scale bars: a and b, 200 µm; c, 5 µm; d and e, 2 µm; f, 0±5 µm.

Printed from the CJO service for personal use only by...
Meristems and shoot development in Sphagnum 587
(a) (b) (c) (d )
(e) (f )
Figure 5. Details of the subapical secondary meristem in the leafy stem of Sphagnum. (a) Light microscopy
showing the first division (arrow) in a secondary meristem cell of S. cuspidatum. (b) Dividing secondary
meristem cells in S. recurvum. (c) Files of cells at the beginning of secondary differentiation in S. palustre.Primary septa between non-sister cells are arrowed. (d ) Dividing cell showing a well-developed phragmoplast
(arrows) between the two daughter nuclei (n) ; plastids (p) are associated with each nucleus. (e) Newly formed
secondary cell at the end of a cell file in S. cuspidatum, showing both a primary (1) and a secondary (2) septum.
( f ) Detail of a cell wall in S. magellanicum, showing cortical microtubules (arrows) along a non-pitted area but
lacking at the level of the pitted area nearby. Scale bar: a, 20 µm; b, 100 µm; c, 200 µm; d and e, 5 µm; f, 0±5 µm.
(Fig. 1b, c) and produces the lateral branches and the
hyaline tissue ensheathing the mature stem.
The inner derivatives of the apical cell generate
the internal cylinder of the stem through a two-step
process. During the first step, which we refer to as
primary differentiation, the cells stop dividing after
a few divisions, become highly vacuolate and enlarge
(Fig. 1b, c). At a distance of 0±7–1±2 mm from the
apical cell the cells form thick, pitted walls and their
cytoplasm becomes denser (Fig. 1b,d, e). Following
these changes, the cells divide repeatedly by trans-
verse septa (Fig. 1e), thus forming a subapical
meristem about 0±8–1±5 mm in length (Fig. 1 f ), the
derivatives of which will generate the internal
cylinder of the mature stem by secondary differen-
tiation (Figs 2–6).
The following description of cytological events
accompanying primary and secondary differentiation
focusses on the internal cells of the cylinder, which
form the bulk of the mature stem. Morphological
and cytological differences between these and the
thick-walled peripheral cells of the cylinder arise
mostly during secondary differentiation and have
been described in a previous study (Ligrone &

Printed from the CJO service for personal use only by...
588 R. Ligrone and J. G. Duckett
(a) (b) (d ) (f )
(c) (g) (h)
Figure 6. Secondary differentiation of leafy stem cells in Sphagnum. (a) Light microscopy of the mature stem
of S. palustre. The external sheath of hyaline cells (arrows) arise from the cortical meristem; all the internal
tissue, including the stereids, is of secondary origin. (b) Detail from a showing fully elongate internal cells. (c)Detail of an internal cell in the course of elongation in S. cuspidatum. The non-pitted wall areas are very thin
and long, but the pitted area (arrow) retains much the same length and thickness as in former stages; n, nucleus.
(d–f ) Details of the cell wall in the course of (d, e) and at the end of ( f ), secondary expansion in S. cuspidatum.
Associated with the expanding wall are numerous cortical microtubules (arrowed) perpendicular to the cell

Printed from the CJO service for personal use only by...
Meristems and shoot development in Sphagnum 589
Duckett, 1998). Quantitative parameters of cellular
differentiation in S. cuspidatum are summarized in
Table 1 and the developmental changes illustrated
diagrammatically in Figures 7–9. For clarity we have
divided development into 7 stages: stages 1–4 refer
to primary differentiation of the inner derivatives of
the apical cell ; stages 5–7 refer to secondary
differentiation of the inner cells of the stem.
Primary differentiation
The apical cell has dense cytoplasm with a prominent
spheroidal nucleus, numerous small plastids and
vacuoles and, frequently, dumbell-shaped mito-
chondria (Fig. 2a). The immediate derivatives have
a similar ultrastructural morphology, except for the
vacuoles, which are more irregular in shape and
often accumulate electron-opaque material (Fig. 2b).
At this stage (stage 1) the cells present slightly
elongate hexagonal longitudinal profiles. They
usually have very thin walls (about 0±15–0±20 µm)
with numerous evenly scattered plasmodesmata. The
transition to stage 2 (i.e. the onset of primary
differentiation) is characterized by cell elongation
(Table 1) with the cells developing a clearly storied
(tiered) arrangement (Fig. 1c, d ) that persists until
the onset of secondary differentiation. The vacuoles
enlarge (Fig. 2c, d ) as do the plastids, which also
develop an extensive inner-membrane system (Fig.
2d ). The plasmodesmata in longitudinal walls now
become confined to distinct pitted regions separated
from each other by non-pitted areas (Fig. 2d ). The
transverse walls usually contain a single, central,
pitted area and a relatively small margin with no
plasmodesmata. From this stage, a cortical array of
microtubules with a transverse orientation is regu-
larly found along the non-pitted wall areas, while no
microtubules are visible in the cytoplasm adjoining
pitted regions (Fig. 2e). A collar of electron-
transparent material surrounds each plasmodesma;
moreover, the wall material between plasmodesmata
in the same pitted area is more electron-opaque than
in adjacent non-pitted areas (Fig. 2e).
The subsequent stage in primary differentiation
(stage 3) comprises a change in nuclear shape from
spheroidal to elongate, and a marked thickening of
non-pitted wall areas along with a moderate increase
in cell size (Table 1; Fig. 2 f, g). In stage 4, the cells
elongate considerably, and the numerous vacuoles
present in stage 3 are replaced with one or a few large
vacuoles that displace the nucleus and most
organelles to the periphery (Fig. 3a, b). Cellular
elongation only affects the non-pitted wall areas,
which thus become longer and thinner, whereas the
axis ; the last-deposited wall layers exhibit an axial orientation of cellulose microfibrils ( f, arrows). (g) Mature
internal cell showing secondary wall layering (arrows) in S. recurvum ; pw, primary wall. (h) Detail from g ;
secondary wall layering is conspicuous in the longitudinal walls but it is very scarce on the transverse septa.
Scale bars: a, 200 µm; b, 50 µm; c, d, f and g, 5 µm; e, 0±5 µm; h, 1±0 µm.
pitted areas retain much the same dimensions (Fig. 3
b, Table 1). The same is observed in transverse walls,
where expansion only affects the non-pitted area.
Electron-opaque finely granular material is now
visible in the vacuoles (Fig. 3b). At the end of
primary differentiation, the cells appear as highly
vacuolate parenchyma cells. An approximate evalu-
ation of changes in cell wall volumes indicates that a
very limited deposition of new wall material occurs
during the transition from stage 3 to stage 4.
Although quite variable, the region of primary
differentiation is markedly longer in S. cuspidatum
than in the other three species (cf. Fig. 1b, c). In
S. palustre, cellular vacuolization is more pro-
nounced throughout development and the different
stages in primary differentiation are not so distinct as
in the other species. By contrast, the pattern of
secondary development is remarkably uniform in all
the species examined.
The development of secondary meristematic activity
The first signs of the development of a secondary
meristem from fully differentiated primary cells are
the replacement of the large vacuole(s) by numerous
smaller vacuoles containing electron-opaque coarse
deposits, and the concomitant migration of the nucleus
from the periphery to the interior of the cells (Fig.
3c, d ). Initially, the non-pitted areas are only slightly
thicker than pitted areas, although morphological
differences between the two regions are still evident
(Fig. 3e, f ). Subsequently, the non-pitted wall areas
become strongly thickened while retaining a homo-
geneous fibrillar structure with no visible layering.
The vacuoles further increase in number and reduce
in size (stage 5; Fig. 4a–c, e). Each cell is usually in
contact laterally with five or six other cells and there
are rows of six to eight pitted areas on each cellular
face (Fig. 8). The pitted areas are elliptical or
roughly circular in shape, with a diameter of
about 2–6 µm, and contain numerous plasmodesmata
(Fig. 4d ) with a density ranging from 22 µm−#
(S. magellanicum and S. recurvum) to 30 µm−# (S.
palustre and S. cuspidatum). Plasmodesma numbers
per pit thus range from 60 to 800 with most contain-
ing 200–500. A network of tubular endoplasmic
reticulum (ER), showing continuity with the desmo-
tubules, is regularly found in the cortical cytoplasm
adjoining pitted areas (Fig. 4 f ).
The cells reach their maximum width just before
the onset of the secondary divisions. The first of
these separates two cells of either similar or different
sizes (cf. Figs 1d, e and 5a). Irrespective of whether
the first division is symmetrical or asymmetrical, this

Printed from the CJO service for personal use only by...
590 R. Ligrone and J. G. Duckett
(a)
n
vn
(b ) (c )
v
(d )
n
v
(e )
n
mt
v
(f )
n
n
mt
v
n
(g ) (h ) ( j )
(i )
v
mt
n
Figure 7. For legend see opposite.

Printed from the CJO service for personal use only by...
Meristems and shoot development in Sphagnum 591
is followed during stage 6 by other, both terminal
and intercalary, divisions (Fig. 5b) ; these produce
files of 6–11 (occasionally more but on average nine)
short cells (Fig. 5c). A tightly controlled balance of
cellular elongation and cellular division keeps the
initial length of secondary derivatives rather constant
(about 20 µm in S. cuspidatum, slightly more in the
other species). The files of newly formed daughter
cells are conspicuously longer than the mother cells
from which they arise (about 180 µm as compared
with about 80 µm in S. cuspidatum). In spite of this
considerable expansion, the longitudinal walls are
further thickened in the course of the divisions (Fig
5a, b, e). As in former stages, the thickening only
affects non-pitted areas, which exhibit a compact
fibrillar texture with a predominately transverse
orientation of microfibrils (Fig. 5 f ). Cortical micro-
tubules in the adjoining cytoplasm also show trans-
verse orientation (Fig. 5 f ). Cellular division is
preceded by plastid migration to the nuclear poles,
probably ensuring that the daughter cells receive
equivalent plastid numbers, and terminates with a
phragmoplast depositing a new septum between the
two daughter nuclei (Fig. 5d ). The new transverse
walls are slightly thinner and less electron-opaque
than the old ones; moreover, they lack the non-
pitted, thickened margin present in the transverse
septa of primary origin (Fig. 5e).
Secondary differentiation
During subsequent differentiation, the secondarily
formed cells elongate about ninefold while remain-
ing almost unchanged in width (Fig. 6a, b). The
longitudinal walls of the elongating cells are very
thin (Fig. 6c), show a loose fibrillar texture with a
predominately transverse or oblique orientation and
have numerous cortical microtubules beneath (Fig.
6d, e). In the fully elongated cells, the longitudinal
walls are slightly thicker because of the late de-
position of additional layers with a lengthwise
orientation of microfibrils (Fig. 6 f ). Once again, the
elongation does not affect the pitted areas; these
retain much the same length and morphology that
was established at stage 2 (Fig. 6c and Table 1). An
evaluation of cell wall volumes in unexpanded and
Figure 7. Diagrammatic representation of the major subcellular changes in the ontogeny of the central cells
of Sphagnum stems. Abbreviations; mt, microtubules; n, nuclei ; v, vacuoles. Because of economies of scale,
plastids, mitochondria, ER, dictyosomes and plasmodesmata are not illustrated. Figure numbers in parentheses
refer to micrographs of the various stages. All drawings are to the same scale. (a) Stage 1: thin-walled
rectangular cells (Fig. 2b). (b) Stage 2: elongation of the initially hexagonal apical cell derivatives has produced
a storied arrangement of elongate cells with large vacuoles (Fig. 2c, d ). (c) Stage 3: further cell enlargement is
associated with wall thickening, nuclear elongation and a reduction in the size of the vacuoles (Fig. 2 f, g). (d )
Stage 4: note the now peripheral nucleus, thinner walls and large central vacuole (Fig. 3a, b). (e) Stage 5:
Secondary meristematic cell with thick pitted walls, an elongate central nucleus and small vacuoles with dense
contents (Fig. 4). ( f–i) Stage 6: the secondary meristematic cells undergo several divisions to produce files of
6-9-11 cells (Fig. 5) concomitant with cell elongation and thinning of the walls. These processes continue after
the cessation of meristematic activity (i), when axial endoplasmic strands begin to appear. ( j) Stage 7: mature
central cell with endoplasmic strands, polarized cytoplasm and thin walls (Fig. 6).
(a ) (b ) (d ) (e )
(c )
Figure 8. Diagrammatic representation showing the
apportioning of pits in the secondary meristematic cells
during stages 6 and 7. Numbers in parentheses refer to
micrographs and to the drawings of the same stages in
Fig. 7. (a) Secondary meristematic cell with a row of pits
on each longitudinal face (Figs 7e ; 4c–e). (b) The first
division of the meristematic cells halves the number of pits
on each face (Figs 7 f, 5a). (c) Subsequent divisions again
halve the number of pits per face (Fig. 7g). (d ) Following
the final division each face contains but one pit (Figs 7h,
6c). (e) The single pit in each face remains unchanged
during maturation of the central cells (Fig. 7 j).
fully elongated cells of S. cuspidatum indicates that
cellular elongation during secondary differentiation
must involve the synthesis of approx. 200% new
wall material. Further thickening of the longitudinal
walls follows the completion of cellular expansion in
S. palustre, S. recurvum and S. magellanicum (Fig.
6g,h) ; no secondary layering is usually visible in S.
cuspidatum (Fig. 6c). A full description of the
cytology of the mature cells is given in Ligrone &
Duckett (1998).

Printed from the CJO service for personal use only by...
592 R. Ligrone and J. G. Duckett
(h)(g)(f )(e )(d )(c )(b )(a )
2 lm(a )
2 lm(b ) – (h )
Figure 9. Diagrammatic representation of the changes in wall thickness, pit spacing and cortical microtubule
distribution during central cell ontogeny in Sphagnum. Numbers in parentheses refer to micrographs and to the
drawings of the same stages in Figure 7. (a) Apical cell derivative; thin wall with uniformly distributed
plasmodesmata and microtubules (Figs 7a, 2a, b). (b) Stage 2: distinct pitted and non-pitted areas (Figs 7b, 2d,
e). Shading with arrows indicates the wall segments that have expanded between stages a and b. (c) Stage 3:
thicker walls between the pitted areas (Figs 7c, 2g). (d ). Stage 4: thinner walls and wider areas between the pits
(Figs 7d, 3b). (e) Stage 5: secondary meristematic cell with thicker walls (Figs 7e, 4). ( f ) Onset of stage 6; the
walls reach their maximum thickness and the distance between the pits remains the same as in stage 4 (Figs 7 f,5e, f ). (g) Stage 6: midway; elongation of the derivatives of the secondary meristematic cells accompanied by
an increased spacing of the pits (Fig. 7 i). (h) Stage 7: mature central cell ; the thin walls now generally contain
no more than one pit per lateral face (Figs 7 j, 6c).
This study shows that stem development in
Sphagnum is a two-step process. Primary meris-
tematic activity in derivatives of the apical cell
produces tiers of differentiated, highly vacuolate
cells in the centre of the capitulum. In the secondary,
subapical phase, redifferentiation and resumption of
meristematic activity in these cells, producing files of
elongate derivatives, is responsible for stem
elongation underlying the expansion of the mature
capitulum.
Though the presence of subapical meristematic
cells, dividing transversely, can be surmised from
previously published drawings of Sphagnum capitula
by Schimper (1858), a subapical secondary meristem
has not previously been reported. Moreover, this
appears to be a feature unique to Sphagnum not only
among the mosses (cf. Crosby, 1980) but also among
all bryophytes. Examples of intercalary meristems
are known in other bryophytes: in the young
sporophyte of mosses (with the exception of
Sphagnum) (Roth, 1969; Crandall-Stotler, 1981,
1984); in the young gametangiophores of Reboulia
(Brown & Lemmon, 1990) and other marchantialean
liverworts (Marchantia and Preissia Cavers, 1904;
Goebel, 1905); and notably in the sporophyte of
anthocerotes (Crandall-Stotler, 1981). These, how-
ever, are instances of residual meristems of primary
origin rather than true secondary meristems.
In an evolutionary context, although at maturity
the central stem cells of Sphagnum cytologically
closely resemble the food-conducting cells of bryoid
mosses (Ligrone & Duckett, 1994), developmentally
they are not homologous. The transverse end walls
in Sphagnum clearly relate to the secondary origin of

Printed from the CJO service for personal use only by...
Meristems and shoot development in Sphagnum 593
these cells, whereas the inclined end walls in bryoid
conducting cells reflect their primary development
directly from derivatives of the apical cell.
On average, each secondary initial in Sphagnum
produces nine derivatives, each elongating to about
180 µm, with a resulting overall length of about
1600 µm for each file of cells. This accounts for the
considerable extension of the main stem recorded for
several Sphagnum spp. (Clymo, 1970; Sonesson et
al., 1980; Clymo & Hayward, 1982; Gerdol, Bonora
& Poli, 1994). By comparison, in the setae of
jungermannialean and metzgerialean liverworts,
which typically expand from 1 to 35–50 mm in a few
hours or days, elongation is caused solely by
vacuolation of individual cells with no cell divisions
(Thomas & Doyle, 1976; Schnepf & Deichgra$ ber,
1979). Single epidermal cells in the seta of Pellia
grow from 12 to about 600 µm in length within 24 h
(Schnepf & Deichgra$ ber, 1979); this figure, although
large, is much below the single-cell increment
achieved in Sphagnum by interposition of secondary
meristem activity.
Although this study focuses on only four species,
representing but two sections in the genus
Sphagnum, less-detailed observations on a range of
other species (Ligrone & Duckett, 1998) indicate
that the presence of a subapical secondary meristem
is ubiquitous in the genus. This unusual growth
mechanism is probably closely associated with the
equally unusual ecology of bog mosses and their
success in waterlogged oligotrophic habitats.
Sphagnum plants frequently keep growing through-
out the year, notably at temperate latitudes, although
generally the bulk of the growth occurs during
summer (Clymo, 1970; Clymo & Hayward, 1982).
The secondary meristem activity ensures that the
capitula, the most important part of the plants in
terms of photosynthetic activity (Johansson &
Linder, 1990), are kept at the surface; this has
benefits in terms of light interception and CO#
uptake. In addition, the secondary meristem might
be crucial for long-term maintenance of the growth
pattern of Sphagnum, because it permits rapid
elongation of old capitula when these are being
replaced by new ones after periods of dormancy
caused, for example, by winter freezing or summer
droughts. In striking contrast, other large perennial
mosses that grow in dense tufts (e.g. many members
of the Polytrichales), but which do not concentrate
their photosynthetic tissues at the stem apices, lack a
secondary meristem (He!bant, 1977).
A second remarkable aspect of leafy stem growth
in Sphagnum is the peculiar way the cells elongate.
At the onset of primary differentiation, moderate
cellular enlargement results in the formation of
distinct pitted areas. The concomitant decrease in
plasmodesmal frequency (Table 1) is fully consistent
with a dilution effect of cell elongation, suggesting
that the development of pitted areas involves the
confinement of pre-existing plasmodesmata rather
than the obliteration of old plasmodesmata and the
formation of new ones. Further cellular growth,
during both primary and secondary differentiation,
involves thickening of non-pitted areas followed by
expansion and thinning out. By contrast, the pitted
areas only undergo very limited thickening and
remain virtually unaltered in size. This is a re-
markable instance of differential expansion of ad-
jacent wall areas in the same cell.
According to our evaluation, the expansion and
thinning-out of pre-thickened wall areas can account
completely for cellular elongation during the tran-
sition from stage 3 to stage 4, with no need for
concomitant synthesis of new wall material. By
contrast, the same mechanism can be responsible for
only about 27% of cellular elongation during
secondary differentiation. The expansion of pre-
thickened wall areas is restricted to the initial phase
of secondary differentiation, resulting in the
formation of very thin uniform walls whose further
elongation must require the synthesis of new wall
material. Like longitudinal growth, the cellular
growth in diameter also depends on the expansion of
the non-pitted area in transverse walls and is
therefore restricted to primary differentiation, be-
cause secondarily formed septa apparently lack a rim
of expandable wall.
As in Sphagnum, cellular expansion in the seta of
Pellia also involves the initial thinning out of pre-
thickened walls, followed by an expansion phase
entirely sustained by the synthesis of new wall
material (Schnepf & Deichgra$ ber, 1979). In Pellia,
however, the process of wall thickening and ex-
pansion is not restricted to specific areas, but instead
affects all the whole longitudinal walls. In both
systems, the pre-thickened walls have a pre-
dominately transverse orientation of microfibrils,
with no visible layering, and are consistently
associated with cortical microtubules showing the
same orientation as the microfibrils.
The plasmodesmal frequency decreases linearly
with cellular elongation during the whole devel-
opmental sequence in stem cells of S. cuspidatum,
indicating that at no stage in differentiation do new
plasmodesmata arise. The same is true during
elongation of the water-conducting cells in the
liverwort Symphyogyna (Ligrone & Duckett, 1996).
By contrast, an analysis of plasmodesmal frequency
in expanding leaflet cells of S. palustre supports de
novo formation of plasmodesmata (Schnepf & Sych,
1983).
A major difference between pitted and non-pitted
areas is the absence of microtubules (MTs) in the
cortical cytoplasm adjoining the former. Here the
MTs are replaced with a network of tubular ER
continuous with the desmotubules. A similar con-
dition was recently reported in developing water-
conducting cells of the liverwort Symphyogyna,

Printed from the CJO service for personal use only by...
594 R. Ligrone and J. G. Duckett
where the dense array of cortical MTs associated
with the longitudinal walls avoids the wall areas
surrounding single plasmodesmata; these areas also
exhibit different staining properties from the rest
of the wall (Ligrone & Duckett, 1996). In
Symphyogyna, however, local differences in the wall
structure arise along with the deposition of a
secondary wall (i.e. after the completion of cellular
elongation) and affect the differential lysis of the wall
after the death of the cells rather than wall expansion.
This is not the case in Sphagnum, where pitted and
non-pitted areas are both part of the primary wall
and clearly recognizable since a very early de-
velopmental stage. The different thickening proper-
ties of pitted and non-pitted areas are presumably
linked to the uneven distribution of MTs and cortical
ER and probably reflect a differential control of
cellulose deposition and of wall-loosening and wall-
stiffening processes (Cosgrove, 1993a, b ; Cyr, 1994).
Apart from tip-growing protonemal cells (Sievers
& Schnepf, 1981), other precedents for localized cell
wall expansion is intercalary growth in young thallus
cells in Marchantia (Apostolakos & Galatis, 1993)
and in the initial cells of gemmae in jungerman-
nialean liverworts (Duckett & Ligrone, 1995) and the
moss Aulacomnium androgynum (Hedw.) Schwaegr.
(Ligrone, Duckett & Gambardella, 1996b). In con-
trast to Sphagnum, here the expanding wall areas do
not undergo preliminary thickening, although they
are consistently associated with cortical MTs. Also
different from the mechanism observed in Sphagnum
is the process of localized wall expansion in fully
differentiated cells of mosses such as the initials of
foliar gemmae in Tortula (Ligrone, Duckett &
Gambardella, 1996a) or protonemal cells during the
development of lateral branches (Schmiedel &
Schnepf, 1979; Ligrone, Duckett & Egunyomi,
1992). Here expansion involves the deposition of a
highly plastic new wall layer and entails breaking off
of the old wall.
The occurrence of highly pitted cells in the leafy
shoot apex of Sphagnum was probably first reported
by Schimper (1858), who assumed that the apparent
absence of pits in cells of mature parts of the stem
resulted from obliteration by new wall material. The
same view was expressed in a more recent study
using scanning electron microscopy (Baker, 1988).
The present study reveals that the pits arise by
differential deposition of primary wall material
during primary differentiation. The pits are then
allocated to separate cells during secondary divisions
and eventually become scarcely visible as a conse-
quence of the expansion and thinning out of the cell
walls during secondary differentiation; however,
they are never obliterated by new wall material. In
striking contrast, primary plasmodesmata and the
adjacent walls are modified profoundly during
phloem differentiation (Glockmann & Kollmann,
1996).
From the functional standpoint, the changes in
plasmodesmal distribution in the stems of Sphagnum
almost certainly reflect changes in the major
symplastic pathways. Lateral transport, involving
the growing branches, probably predominates within
the capitulum, but below the capitulum translocation
will be exclusively axial. Thus, plasmodesmata are
approximately equally distributed between lateral
and end walls in the capitulum, but below the
capitulum those in the end walls outnumber the
laterals by at least one order of magnitude.
R.L. thanks Queen Mary & Westfield College for
laboratory facilities in 1992, 1993 and 1996. The obser-
vations were in part performed at the CIRUB (University
of Naples ‘Federico II’).
Apostolakos P, Galatis B. 1993. Interphase and preprophase
microtubule organization in some polarized cell types of the
liverwort Marchantia paleacea Bert. New Phytologist 124 :
409–421.
Baker RGE. 1988. The morphology and distribution of pits in the
cell walls of Sphagnum. Journal of the Hattori Botanical
Laboratory 64 : 359–365.
Brown RC, Lemmon BE. 1990. Polar organizers mark division
axis prior to preprophase band formation in mitosis of the
hepatic Reboulia hemisphaerica (Bryophyta). Protoplasma 156 :
74–81.
Cavers F. 1904. On the structure and biology of Fegatella conica.
Annals of Botany 18 : 87–119.
Cavers F. 1911. The inter-relationships of the Bryophyta. VI.
Sphagnales. New Phytologist 10 : 1–21.
Clymo RS. 1970. The growth of Sphagnum : methods of
measurement. Journal of Ecology 58 : 13–49.
Clymo RS. 1984. Sphagnum-dominated peat bog: a naturally acid
ecosystem. Philosophical Transactions of the Royal Society of
London B305 : 487–499.
Clymo RS. 1987. Interactions of Sphagnum with water and air.
In: Hutchinson TC, Meema KM, eds. Effects of Atmospheric
Pollutants on Forests, Wetlands and Agricultural Ecosystems.
Berlin, Germany: Springer Verlag, 513–529.
Clymo RS, Duckett JG. 1986. Regeneration of Sphagnum. New
Phytologist 102 : 589–614.
Clymo RS, Hayward PM. 1982. The ecology of Sphagnum. In:
Smith AJE, ed. Bryophyte Ecology. London, UK: Chapman and
Hall, 229–289.
Cosgrove DJ. 1993a. How do plant cell walls extend? Plant
Physiology 102 : 1–6.
Cosgrove DJ. 1993b. Wall extensibility : its nature, measurement
and relationship to plant cell growth. New Phytologist 124 :
1–23.
Crandall-Stotler B. 1981. Morphology}anatomy of hepatics and
anthocerotes. Advances in Bryology 1 : 315–398.
Crandall-Stotler B. 1984. Musci, hepatics and anthocerotes – an
essay on analogues. In: Schuster RM, ed. New Manual of
Bryology, vol. 2. Nichinan, Japan: Hattori Botanical Lab-
oratory, 1093–1129.
Crosby MR. 1980. The diversity and relationships in mosses. In:
Taylor RJ, Leviton AE, eds. The Mosses of North America. San
Francisco CA, USA: Pacific Division, AAAS, 115–129.
Cyr RJ. 1994. Microtubules in plant morphogenesis : role of the
cortical array. Annual Review of Cell Biology 10 : 153–180.
Duckett JG, Ligrone R. 1995. The formation of catenate foliar
gemmae and the origin of oil bodies in the liverwort
Odontoschisma denudatum (Mart.) Dum. (Jungermanniales) : a
light and electron microscope study. Annals of Botany 76 :
405–419.

Printed from the CJO service for personal use only by...
Meristems and shoot development in Sphagnum 595
Gerdol R, Bonora A, Poli F. 1994. The vertical pattern of
pigment concentration in chloroplasts of Sphagnum
capillifolium. Bryologist 97 : 158–161.
Gignac LD. 1993. Distribution of Sphagnum species, com-
munities, and habitats in relation to climate. Advances in
Bryology 5 : 187–222.
Glockman C, Kollmann R. 1996. Structure and development of
cell connections in the phloem of Metasequoia glyptostroboides
needles. I. Ultrastructural aspects of modified primary plasmo-
desmata in Strasburger cells. Protoplasma 193 : 191–203.
Goebel K. 1905. Organography of Plants. Trans. Balfour IB.
Oxford, UK: Clarendon Press.
He! bant C. 1977. The conducting tissues of bryophytes. Bryo-
phtorum Bibliotheca 10 : 1–157.
Johansson LG, Linder S. 1990. Photosynthesis of Sphagnum in
different microhabitats on a subarctic mire. In: Sonesson M,
ed. Ecology of a Subarctic Mire. Stockholm, Sweden: Ecological
Bulletins 30 : 181–190.
Ligrone R, Duckett JG. 1994. Cytoplasmic polarity and
endoplasmic microtubules associated with the nucleus and
organelles are ubiquitous features of food-conducting cells in
bryoid mosses (Bryophyta). New Phytologist 127 : 601–614.
Ligrone R, Duckett JG. 1996. Development of water-conducting
cells in the antipodal liverwort Symphyogyna brasiliensis
(Metzgeriales). New Phytologist 132 : 603–615.
Ligrone R, Duckett JG. 1998. The leafy stems of Sphagnum
(Bryophyta) contain highly differentiated polarized cells with
axial arrays of endoplasmic microtubules. New Phytologist 140 :
567–579.
Ligrone R, Duckett, JG, Egunyomi A. 1992. Foliar and
protonemal gemmae in the tropical moss Calymperes
(Calymperaceae) : an ultrastructural study. Cryptogamic Botany
2 : 317–329.
Ligrone R, Duckett JG, Gambardella R. 1996a. Serial
development of foliar gemmae in Tortula (Pottiales, Musci), an
ultrastructural study. Annals of Botany 78 : 305–315.
Ligrone R, Duckett JG, Gambardella R. 1996b. Development
and liberation of cauline gemmae in the moss Aulacomnium
androgynum (Hedw.) Schwaegr. (Bryales) : an ultrastructural
study. Annals of Botany 78 : 559–568.
Roth D. 1969. Embryo und embryotheca bei den Laubmoosen.
Eine histogenetische und morphologische Untersuchung.
Bibliotheca Botanica 129 : 1–49.
Rydin H, Clymo RS. 1989. Transport of carbon and phos-
phorous compounds about Sphagnum. Proceedings of the Royal
Society of London B237 : 63–84.
Schimper WP. 1858. Versuch einer Entwicklungsgeschichte der
Torfmoose (Sphagnum) und einer Monographie in der Europa
verkommenden Arten dieser Gattung. Stuttgart, Germany:
Schweitzerbart’s Verlagshandlung.
Schmiedel G, Schnepf E. 1979. Side branch formation and
orientation in the caulonema of the moss, Funaria hygrometrica :
normal development and fine structure. Protoplasma 100 :
367–383.
Schnepf E, Sych A. 1983. Distribution of plasmodesmata in
developing Sphagnum leaflets. Protoplasma 116 : 51–56.
Schnepf E, Deichgra$ ber G. 1979. Elongation growth of setae of
Pellia (Bryophyta) : fine structural analysis. Zeitschrift fuX rPflanzenphysiologie 94 : 283–297.
Sievers A, Schnepf E. 1981. Morphogenesis and polarity of
tubular cells with tip growth. In: Kiermayer O, ed. Cyto-
morphogenesis in Plants. Wien, Austria : Springer Verlag,
265–299.
Smith GM. 1955. Cryptogamic Botany. New York, USA:
McGraw-Hill Book Company.
Sonesson M, Persson S, Basilier K, Stenstrøm TA. 1980.Growth of Sphagnum riparium Angstr. In relation to some
environmental factors in the Stordalen mire. In: Sonesson M,
ed. Ecology of a Subarctic Mire. Stockholm, Sweden: Swedish
Natural Science Research Council, 191–207.
Thomas RJ, Doyle WT. 1976. Changes in the carbohydrate
constituents of elongating Lophocolea heterophylla setae
(Hepaticae). American Journal of Botany 63 : 1054–1059.
Copyright © 2022 FDOKUMEN




















