
In Silico inhibitors for Plasmodium falciparum lactate dehydrogenase
Upload
sunderlandCategory
view
0download
0
Molecular & Biochemical Parasitology 127 (2003) 59–68
Apical location of a novel EGF-like domain-containing proteinof Plasmodium falciparum�
Casilda G. Black∗, Lina Wang, Tieqiao Wu, Ross L. CoppelDepartment of Microbiology and the Victorian Bioinformatics Consortium, P.O. Box 53, Monash University 3800, Vic., Australia
Received 6 September 2002; received in revised form 10 November 2002; accepted 15 November 2002
Abstract
Using bioinformatics analyses of the unfinished malaria genome sequence, we have identified a novel protein ofPlasmodium falciparumthat contains two epidermal growth factor (EGF)-like domains near the C-terminus of the protein. The sequence contains a single openreading frame of 1572 bp with the potential to encode a protein of 524 residues containing hydrophobic regions at the extreme N- andC-termini which appear to represent signal peptide and glycosylphosphatidylinositol (GPI)-attachment sites, respectively. RT–PCR analysishas confirmed that the novel gene is transcribed in asexual stages ofP. falciparum. Antibodies to the EGF-like domains of the novel proteinare highly specific and do not cross-react with the EGF-like domains of MSP1, MSP4, MSP5 or MSP8 expressed as GST fusion proteins.Antisera to the C-terminal fragments react with two bands of 80 and 36 kDa inP. falciparumparasite lysates whereas antisera to the mostN-terminal fusion protein only recognises the 80 kDa band, suggesting that the novel protein may undergo processing in a similar wayto MSP1 and MSP8, but with fewer cleavage events. Immunoblot analysis of stage-specific parasite samples reveals that the protein ispresent in trophozoites, schizonts and in isolated merozoites. The protein partitions in the detergent-enriched phase after Triton X-114fractionation and is localised to the surfaces of trophozoites, schizonts and free merozoites in an apical distribution. Based on the acceptednomenclature in the field we now designate this protein MSP10. We have shown that the MSP10 fusion proteins are in a conformation thatcan be recognised by human immune sera and that there is very limited sequence diversity in an∼l kb region of MSP10, encompassing thetwo EGF-like domains. A sequence similar to MSP10 can be identified in the availableP. yoeliigenomic sequence, offering the possibilityof ascertaining whether this novel protein can induce host protective responses in an in vivo model.© 2002 Elsevier Science B.V. All rights reserved.
Keywords: Plasmodium falciparum; Malaria; Merozoite surface protein; Epidermal growth factor-like domain; Bioinformatics
1. Introduction
The existence of a number of invasion pathways forPlas-modium falciparuminto human red blood cells[1–3] sug-gests that there may be multiple sets of proteins involvedat each step, some of which may be adapted for particularreceptors. Support for this idea comes from the observationthat a number of proteins with a role in red cell invasion suchas the erythrocyte binding protein EBA175, the reticulocytebinding proteins (PvRBP1 and 2), the serine repeat antigen(SERA) and merozoite surface protein 3 (MSP3) have been
Abbreviatioins:EGF-like, epidermal growth factor-like; GPI, glyco-sylphosphatidylinositol; MSP, merozoite surface protein
� Note: Nucleotide sequence data reported in this paper are availablein the GenBankTM, EMBL and DDBJ data bases under the accessionnumbers: AF403477 and AF432070–AF432079.
∗ Corresponding author. Tel.:+61-3-9905-1500; fax:+61-3-9905-4811.E-mail address:[email protected] (C.G. Black).
shown to have a number of paralogs in theP. falciparumgenome[4–12].
One of the important proteins involved in red cell in-vasion is merozoite surface protein 1 (MSP1), a largemulti-domain protein that is processed into a number ofcomponents that form a multi-protein complex on themerozoite surface[8,13–15]. An important structural fea-ture of MSP1 is the presence of two epidermal growthfactor (EGF)-like domains at the C-terminus which areinvolved in invasion. We hypothesised that the importanceof EGF-like domains in invasion and as protein interac-tion motifs useful in assembling multi-protein structures,may mean that there was a requirement for additional suchproteins at the merozoite surface. Our recent studies ofMSP8 [16], identified a second merozoite surface proteinthat contained two EGF-like domains, but we wonderedif there may be additional members of this protein familyas other merozoite proteins had between 2 and 6 paralogs.Accordingly we searched for potential new members of this
0166-6851/02/$ – see front matter © 2002 Elsevier Science B.V. All rights reserved.doi:10.1016/S0166-6851(02)00308-0

60 C.G. Black et al. / Molecular & Biochemical Parasitology 127 (2003) 59–68
protein family by screening the Malaria Genome Sequenc-ing Project databases. Using a series of degenerate motifqueries we identified a short sequence from an expressedsequence tag (EST) data base which, when extended byPCR-based techniques, was found to encode a protein withtwo EGF-like domains. We report here the identificationand characterisation of this novel merozoite surface protein,designated MSP10, a third member of a family of asexualstage proteins inP. falciparumthat are GPI-linked, undergopost-translational processing and contain two EGF-likedomains.
2. Materials and methods
2.1. Culture of malaria parasites
P. falciparumparasites were cultured in vitro by standardprocedures[17]. Infected erythrocytes were harvested fromcultures and the parasites were isolated by lysis with 0.15%(w/v) saponin, washed with phosphate-buffered saline[18], and stored at−70◦C until required. For analysis ofstage-specific expression, the cultures were synchronisedby treatment with sorbitol as previously described[19].Free merozoites were isolated from in vitro cultures ofP.falciparum3D7 as reported earlier[20].
2.2. DNA methods
Chromosome walking was performed using vectorettePCR [21] and inverse PCR[22]. Vectorette libraries wereconstructed fromP. falciparum3D7 genomic DNA digestedwith various restriction enzymes and ligated to compatiblevectorette ends provided by D. Peterson (University of Geor-gia, Athens, GA, USA). Subsequent PCR reactions wereperformed on these libraries using a primer derived fromthe known sequence and a vectorette-specific primer. Forinverse PCR, genomic DNA was digested with appropriaterestriction enzymes and religated under dilute conditionsto form circular molecules. PCRs were performed with in-verted primers derived from the known sequence. PCR reac-tions were performed using AmpliTaq Gold (Perkin–Elmer,Branchburg, NJ, USA) under the following temperatureconditions: one cycle of 94◦C for 9 min, 40 cycles (each)of 94◦C for 1 min, 50◦C for 1 min, 72◦C for 1 min and onecycle of 72◦C for 7 min. DNA sequencing of PCR productsand recombinant plasmids was performed using the PRISMReady Reaction DyeDeoxy Terminator Cycle SequencingKit (Applied Biosystems, Inc., Foster City, CA, USA) asdescribed by the manufacturer. Sequencing reaction prod-ucts were separated on a model 373A automated DNAsequencer (Applied Biosystems), and sequences analysedusing Sequencher software.
Fragments of the novel gene were amplified by PCR withthe Expand High Fidelity PCR System (Roche DiagnosticsAustralia Pty. Ltd., Vic., Australia) andP. falciparum3D7
genomic DNA as the template. Primers contained restrictionsites, and the inserts were digested with restriction endonu-cleases and ligated into appropriately cut pGEX-KG vector(Amersham Pharmacia Biotech Pty. Ltd., NSW, Australia).The recombinant plasmids were entirely sequenced to con-firm cloning in the correct reading frame and the absence ofmutations and then introduced intoE. coli BL21 (DE3) (No-vagen, Milwaukee, WI, USA) for protein expression. Theexpression constructs are designated A, B, C and D, andtheir relative positions are shown inFig. 1A. Expression inE. coli and purification of GST fusion proteins were per-formed as earlier described[23].
2.3. RNA methods
Messenger RNA (mRNA) was isolated fromP. falciparum3D7 parasites using the Quick-Prep mRNA Purification Kit(Amersham Pharmacia Biotech Pty. Ltd.) and treated withRQ1 RNase-free DNase (Promega, Madison, WI, USA). Re-verse transcriptase (RT) reactions were performed with Su-perscript II (Gibco BRL, Bethesda, MD, USA) accordingto the manufacturer’s instructions. RT–PCR reactions wereconducted withTaq DNA polymerase (Roche DiagnosticsAustralia Pty. Ltd.) under the following temperature condi-tions: one cycle of 94◦C for 3 min, 39 cycles (each) of 94◦Cfor 1 min, 50◦C for 1 min, 72◦C for 1 min and one cycle of72◦C for 7 min.
2.4. Animal and human antisera
For production of antisera to recombinant MSP10 frag-ments, female Balb/c mice (aged 6–8 weeks) or femaleNew Zealand White rabbits (aged 10–12 weeks) were pur-chased from Central Animal Services (Monash University,Australia) and used for immunisation. For each recombinantprotein, one rabbit and two mice were injected with 50�gsubcutaneously (for rabbits) or 25�g intraperitoneally (formice) of GST fusion protein emulsified in complete Freund’sadjuvant (Difco Laboratories, Detroit, MI, USA), followedby monthly boosting with the same amounts of GST fusionprotein in incomplete Freund’s adjuvant (Difco Laborato-ries). The antibody reactivity was tested by immunoblottingwith parasite-derived MSP10. Human immune sera were ob-tained with informed consent from residents of the Madangregion of Papua New Guinea[24] and the Khanh-Hoa regionof Vietnam[25] with documented exposure toP. falciparuminfection.
2.5. Protein and immunochemical methods
Proteins were analysed by SDS–PAGE, immunoblottedand detected with chemiluminescent reagents as earlier de-scribed[16]. To enrich for membrane-associated proteins,a Triton X-114 partitioning experiment was performed asearlier reported[23]. The indirect immunofluorescence as-says (IFA) were conducted as described earlier[16]. In

C.G. Black et al. / Molecular & Biochemical Parasitology 127 (2003) 59–68 61
Fig. 1. (A) Diagrammatic representation of the protein sequence encoded by the novel gene. Amino acid numbers are shown. The putative signal andGPI-attachment sequences are shown by solid black boxes. The two EGF-like domains are indicated by the striped box. The location of the block of eightdegenerate 6-mer tandem repeats with the sequence (N/I)EN(I/V/N)EN is indicated by the cross-hatched box. A, B, C, and D are four protein fragmentscloned in the pGEX-KG vector. The first and last amino acid residues included in each recombinant GST fusion protein are indicated. (B) Comparisonof MSP10 sequences from variousP. falciparum laboratory isolates. Type 1 includes 3D7, IMR143, IMR147, MAD71, NF7, Type 2 includes AA01,IMR144, ItG2, K1, V1 and Type 3 is FF7. Amino acid numbers are shown. Identical amino acids are indicated by dots and amino acid substitutions areshown. The region of MSP10 containing the 12 cysteine residues that constitute the two EGF-like domains is highlighted in bold-type.
the single colour IFA experiments, MSP10 was detectedusing rabbit antiserum to fragments A, B or D and AlexaFluor 488-labelled goat anti-rabbit antibodies (MolecularProbes, Inc., Eugene, OR, USA). In the two-colour IFAexperiments, MSP10 was detected using rabbit antiserumto fragment B and Alexa Fluor 488-labelled goat anti-rabbitantibodies. RAMA was detected using pooled mouse an-
tisera (S749–S752) and Alexa Fluor 568-labelled goatanti-mouse antibodies (Molecular Probes, Inc.). MSP119was detected with monoclonal antibody (mAb) 1E1[26] andAlexa Fluor 568-labelled goat anti-mouse antibodies. Thereactivity of human immune sera with recombinant MSP10proteins was examined by enzyme-linked immunosorbentassay (ELISA) as previously described[23].

62 C.G. Black et al. / Molecular & Biochemical Parasitology 127 (2003) 59–68
2.6. Nucleotide sequence data
Sequence data for chromosomes 6, 7 and 8 were obtainedfrom the Sanger Centre website athttp://www.sanger.ac.uk/Projects/Pfalciparum/. Sequencing ofP. falciparumchro-mosomes 6, 7 and 8 was accomplished as part of the MalariaGenome Project with support by the Wellcome Trust.
3. Results
3.1. A novel protein of P. falciparum that contains twoEGF-like domains
We searched the unfinished Malaria Genome SequencingProject databases at the NCBI Malaria Genetics and Ge-nomics website (http://www.ncbi.nlm.nih.gov/Malaria/) us-ing a series of EGF-like domains, particularly that of MSP4and a number of motifs based on spaced cysteines[24] asthe BLAST query sequence. We identified a short nucleotidesequence derived from aP. falciparumDd2 expressed se-quence tag (EST) database generated by Chakrabarti et al.[27]. This novel sequence encoded a cluster of seven cysteineresidues preceding a stretch of predominantly hydropho-bic residues that is typical of a glycosylphosphatidylinositol(GPI) attachment motif. The quality of EST sequence in thisparticular database is quite variable and it was contaminatedwith sequences from other organisms, so we wanted to con-firm this was an authenticP. falciparumsequence and that itencoded an EGF-like domain. Additionally, since EGF-likedomains typically contain six cysteine residues, we set outto determine whether the seventh cysteine could constitutepart of a second adjacent motif. We extended the nucleotidesequence in the 5′ direction by chromosome walking onP. falciparum3D7 genomic DNA template using the tech-niques of vectorette and inverse PCR. Sequence analysisof the resulting PCR products revealed the presence of anadditional five cysteine residues. Comparison of these twoEGF-like domains of the novel protein with those of MSP8showed 34.2% identity in the first EGF-like domain and44.4% identity in the second EGF-like domain. Compari-son with MSP1 revealed 31.4% identity in the first EGF-likedomain and 40.0% identity in the second EGF-like domain(data not shown). The spacing between the cysteines is con-sistent with that found in other malaria proteins. The se-quence of the extreme 5′ end of the gene encodes a stretchof predominantly hydrophobic residues and shows similarityto the predicted signal peptide of MSP8 (data not shown).The complete sequence of the novel gene lies within a singleORF of 1572 bp and encodes a polypeptide of 524 aminoacids with a predicted molecular mass of 61.2 kDa. The se-quence was not identical to any known proteins ofP. falci-parum. Similar to MSP8, the novel protein sequence has ahighly asparagine-rich region near the N-terminus.
Even though the initial sequence of the coding region wasderived from a putative EST sequence, it was still important
to confirm that the gene was transcribed in asexual stages ofP. falciparum. Accordingly, mRNA was extracted from ma-ture blood stageP. falciparumparasites, treated with DNaseand subjected to reverse transcription and PCR with a num-ber of primers that collectively encompassed the entire pre-dicted open reading frame (data not shown). In every case,a single product of the expected size was generated in thereverse transcriptase (RT)-containing sample but no prod-uct was observed in the RT-negative sample. Sequencing ofthe products confirmed that they contained the expected nu-cleotide sequence. The amplified cDNA product in all caseswas identical in size to that produced when genomic DNAwas the template suggesting that no cryptic introns werepresent in the sequence. This experiment proved that thepredicted gene is transcribed in late blood stage parasites.
3.2. Specificity of antisera raised to recombinantfragments of the novel protein
Four contiguous, non-overlapping fragments of the novelprotein sequence, designated A, B, C, and D, excluding thenucleotides encoding the putative signal sequence and GPIattachment site, were amplified by PCR fromP. falciparum3D7 genomic DNA and ligated in frame into the pGEX-KGvector (Fig. 1A). The GST fusion proteins were expressedin E. coli and subsequently purified. Panels consisting ofimmunoblotted recombinant proteins of the EGF-like do-mains of MSP1, MSP4, MSP5, MSP8 and the novel pro-tein were probed separately with rabbit antiserum raisedagainst the EGF-like domains of this new protein follow-ing pre-absorption against GST (Fig. 2). The results show
Fig. 2. Lack of cross-reactivity between the antibodies to the EGF-likedomain(s) of MSP1, MSP4, MSP5, MSP8 and the novel protein. Equiva-lent amounts (0.01�g) of GST fusion proteins containing MSP119 (lane1), MSP4D (lane 2), MSP5D (lane 3), MSP8D (lane 4), novel protein(lane 5) and GST alone (lane 6) were probed with rabbit antisera raisedto GST-MSP119 (A), GST-MSP4D (B), GST-MSP5D (C), GST-MSP8D(D) and GST-fragment D of the novel protein (E). All of the antisera werepre-absorbed against GST. Molecular mass standards (in kilodaltons) aregiven at the left of each panel.

C.G. Black et al. / Molecular & Biochemical Parasitology 127 (2003) 59–68 63
that the antiserum is highly specific and does not cross-reactwith the EGF-like domains of knownP. falciparummero-zoite surface proteins. Neither did antisera raised against theEGF-like domains of MSP1, MSP4, MSP5 and MSP8 reactwith recombinant protein of the EGF-like domains encodedby this new gene (Fig. 2). These results provide evidencethat we have identified a novel protein ofP. falciparum, andthat the resultant antisera can be used to provide informationabout its properties.
3.3. The novel protein appears to undergo processingin P. falciparum
In order to study the properties of the protein encoded bythe novel gene, antisera raised against the various recom-binant proteins were used in a number of immunochemicalexperiments. Mature stage parasites were harvested fromP. falciparum 3D7, lysed and subjected to SDS–PAGEfollowed by immunoblotting. Rabbit antibodies raisedagainst fragment A reacted with a single band of 80 kDa(Fig. 3A). Antisera to fragments B, C and D reacted withthe 80 kDa protein and also with a smaller 36 kDa protein(Fig. 3A). The lack of reactivity to the 36 kDa band byantiserum raised to the most N-terminal fragment (anti-A)is consistent with the proposition that this smaller proteinis a C-terminal processing product of the full length pro-
Fig. 3. (A) P. falciparum3D7 parasite lysates were separated by SDS–PAGE, electroblotted and probed with rabbit antisera to fragments A (lane 1), B(lane 2), and C (lane 3). Rabbit antiserum to fragment D was used to immunoprecipitate the novel protein from parasite lysate. The eluted sample waselectroblotted and probed with mouse antiserum to recombinant fragment D of the novel protein (lane 4). (B) Immunoblot analysis ofP. falciparum3D7parasites at various time points (in hours) post-synchronization to ring-stage parasites. Parasite extracts were separated by SDS–PAGE, electroblotted andprobed with rabbit antiserum to fragment B of the novel protein. Uninfected red blood cells (RBC) were used as a negative control. (C) Phase-separationstudies ofP. falciparum. Asynchronous parasites were lysed in the presence of Triton X-114 and after phase separation at 37◦C, aliquots of varioussamples were subjected to SDS–PAGE and electroblotted. Immunoblots were probed with rabbit antiserum raised against recombinant fragment B of thenovel protein. The fractions shown are the Triton X-114 depleted aqueous phase (lane 1) and the Triton X-114 detergent phase (lane 2). Molecular massstandards (in kilodaltons) are indicated on the left of each panel.
tein, that is cleaved in the region between residues 250and 320.
To investigate the stage-specific expression of this pro-tein, asynchronous parasites were cultured in vitro, harvestedand treated with sorbitol to enrich for ring forms. The syn-chronised parasites were then grown for a further 48 h andsampled at 0, 4, 8, 18, 21, 24, 27, 30, 33, 44 and 47 h,representing various stages of the parasite asexual life cy-cle. Parasite lysates were subjected to SDS–PAGE and elec-troblotted. Presence of the protein was first detected as avery weak band of 80 kDa at 30 h post-synchronisation (inearly trophozoites) with the highest levels of expression de-tected at 44 and 47 h, in late trophozoites and schizonts. The36 kDa band was first detected at 33 h post-synchronisation(Fig. 3B). The protein was also detected in isolated mero-zoites (data not shown).
3.4. Evidence for the location of the novel protein at themerozoite apex and on the parasite surface
Parasitised erythrocytes were harvested fromP. falci-parumcultures and subjected to Triton X-114 fractionationto separate proteins into detergent-soluble and aqueous-phase proteins. Phase-separated proteins were electroblot-ted and probed with rabbit anti-B antiserum (Fig. 3C). Thepartitioning of the C-terminal 36 kDa portion of the novel
64 C.G. Black et al. / Molecular & Biochemical Parasitology 127 (2003) 59–68
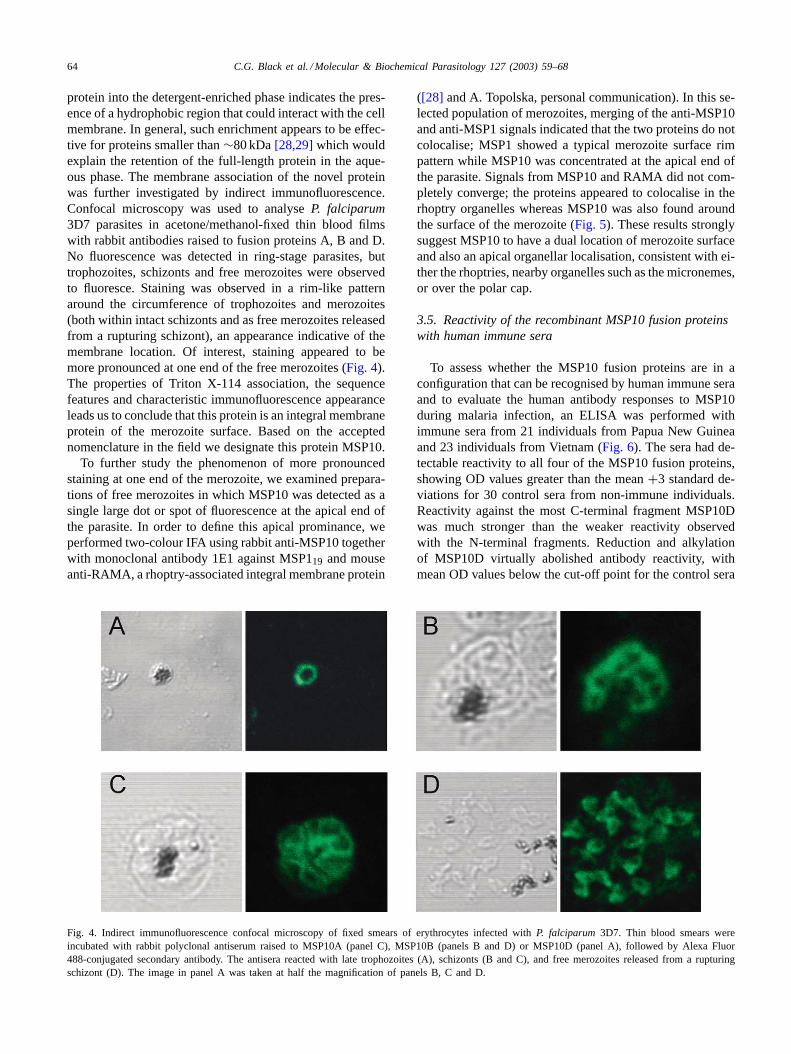
protein into the detergent-enriched phase indicates the pres-ence of a hydrophobic region that could interact with the cellmembrane. In general, such enrichment appears to be effec-tive for proteins smaller than∼80 kDa[28,29]which wouldexplain the retention of the full-length protein in the aque-ous phase. The membrane association of the novel proteinwas further investigated by indirect immunofluorescence.Confocal microscopy was used to analyseP. falciparum3D7 parasites in acetone/methanol-fixed thin blood filmswith rabbit antibodies raised to fusion proteins A, B and D.No fluorescence was detected in ring-stage parasites, buttrophozoites, schizonts and free merozoites were observedto fluoresce. Staining was observed in a rim-like patternaround the circumference of trophozoites and merozoites(both within intact schizonts and as free merozoites releasedfrom a rupturing schizont), an appearance indicative of themembrane location. Of interest, staining appeared to bemore pronounced at one end of the free merozoites (Fig. 4).The properties of Triton X-114 association, the sequencefeatures and characteristic immunofluorescence appearanceleads us to conclude that this protein is an integral membraneprotein of the merozoite surface. Based on the acceptednomenclature in the field we designate this protein MSP10.
To further study the phenomenon of more pronouncedstaining at one end of the merozoite, we examined prepara-tions of free merozoites in which MSP10 was detected as asingle large dot or spot of fluorescence at the apical end ofthe parasite. In order to define this apical prominance, weperformed two-colour IFA using rabbit anti-MSP10 togetherwith monoclonal antibody 1E1 against MSP119 and mouseanti-RAMA, a rhoptry-associated integral membrane protein
Fig. 4. Indirect immunofluorescence confocal microscopy of fixed smears of erythrocytes infected withP. falciparum 3D7. Thin blood smears wereincubated with rabbit polyclonal antiserum raised to MSP10A (panel C), MSP10B (panels B and D) or MSP10D (panel A), followed by Alexa Fluor488-conjugated secondary antibody. The antisera reacted with late trophozoites (A), schizonts (B and C), and free merozoites released from a rupturingschizont (D). The image in panel A was taken at half the magnification of panels B, C and D.
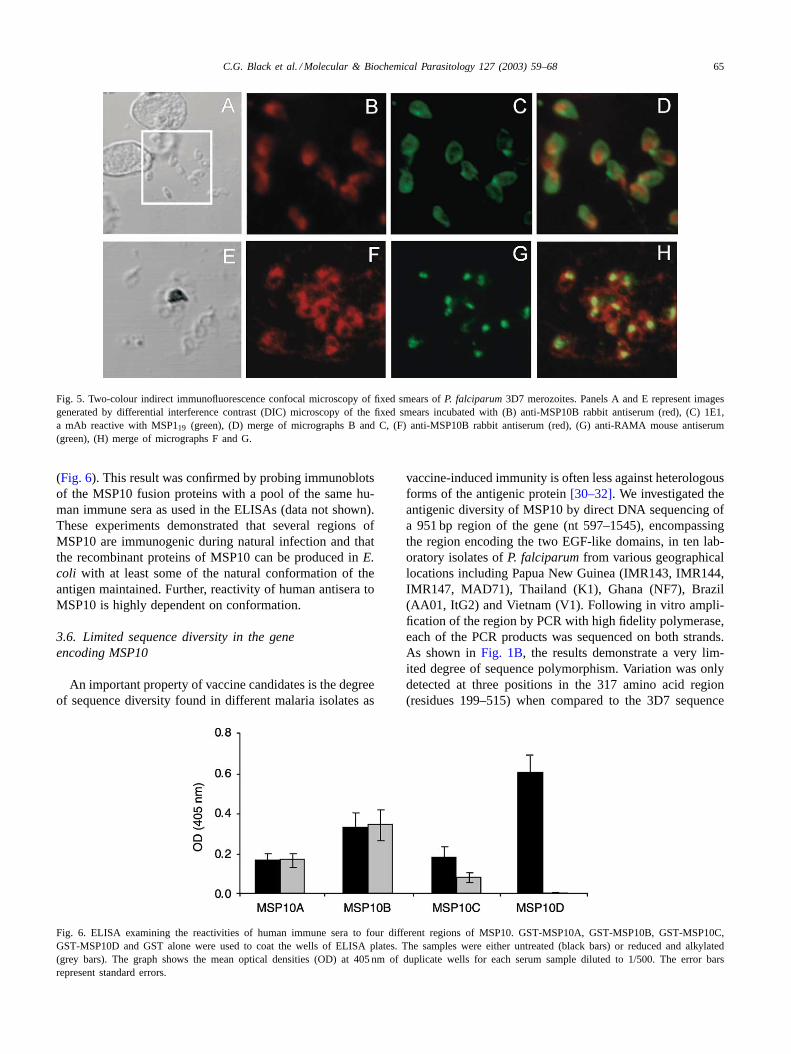
([28] and A. Topolska, personal communication). In this se-lected population of merozoites, merging of the anti-MSP10and anti-MSP1 signals indicated that the two proteins do notcolocalise; MSP1 showed a typical merozoite surface rimpattern while MSP10 was concentrated at the apical end ofthe parasite. Signals from MSP10 and RAMA did not com-pletely converge; the proteins appeared to colocalise in therhoptry organelles whereas MSP10 was also found aroundthe surface of the merozoite (Fig. 5). These results stronglysuggest MSP10 to have a dual location of merozoite surfaceand also an apical organellar localisation, consistent with ei-ther the rhoptries, nearby organelles such as the micronemes,or over the polar cap.
3.5. Reactivity of the recombinant MSP10 fusion proteinswith human immune sera
To assess whether the MSP10 fusion proteins are in aconfiguration that can be recognised by human immune seraand to evaluate the human antibody responses to MSP10during malaria infection, an ELISA was performed withimmune sera from 21 individuals from Papua New Guineaand 23 individuals from Vietnam (Fig. 6). The sera had de-tectable reactivity to all four of the MSP10 fusion proteins,showing OD values greater than the mean+3 standard de-viations for 30 control sera from non-immune individuals.Reactivity against the most C-terminal fragment MSP10Dwas much stronger than the weaker reactivity observedwith the N-terminal fragments. Reduction and alkylationof MSP10D virtually abolished antibody reactivity, withmean OD values below the cut-off point for the control sera
C.G. Black et al. / Molecular & Biochemical Parasitology 127 (2003) 59–68 65
Fig. 5. Two-colour indirect immunofluorescence confocal microscopy of fixed smears ofP. falciparum3D7 merozoites. Panels A and E represent imagesgenerated by differential interference contrast (DIC) microscopy of the fixed smears incubated with (B) anti-MSP10B rabbit antiserum (red), (C) 1E1,a mAb reactive with MSP119 (green), (D) merge of micrographs B and C, (F) anti-MSP10B rabbit antiserum (red), (G) anti-RAMA mouse antiserum(green), (H) merge of micrographs F and G.
(Fig. 6). This result was confirmed by probing immunoblotsof the MSP10 fusion proteins with a pool of the same hu-man immune sera as used in the ELISAs (data not shown).These experiments demonstrated that several regions ofMSP10 are immunogenic during natural infection and thatthe recombinant proteins of MSP10 can be produced inE.coli with at least some of the natural conformation of theantigen maintained. Further, reactivity of human antisera toMSP10 is highly dependent on conformation.
3.6. Limited sequence diversity in the geneencoding MSP10
An important property of vaccine candidates is the degreeof sequence diversity found in different malaria isolates as
Fig. 6. ELISA examining the reactivities of human immune sera to four different regions of MSP10. GST-MSP10A, GST-MSP10B, GST-MSP10C,GST-MSP10D and GST alone were used to coat the wells of ELISA plates. The samples were either untreated (black bars) or reduced and alkylated(grey bars). The graph shows the mean optical densities (OD) at 405 nm of duplicate wells for each serum sample diluted to 1/500. The error barsrepresent standard errors.
vaccine-induced immunity is often less against heterologousforms of the antigenic protein[30–32]. We investigated theantigenic diversity of MSP10 by direct DNA sequencing ofa 951 bp region of the gene (nt 597–1545), encompassingthe region encoding the two EGF-like domains, in ten lab-oratory isolates ofP. falciparumfrom various geographicallocations including Papua New Guinea (IMR143, IMR144,IMR147, MAD71), Thailand (K1), Ghana (NF7), Brazil(AA01, ItG2) and Vietnam (V1). Following in vitro ampli-fication of the region by PCR with high fidelity polymerase,each of the PCR products was sequenced on both strands.As shown inFig. 1B, the results demonstrate a very lim-ited degree of sequence polymorphism. Variation was onlydetected at three positions in the 317 amino acid region(residues 199–515) when compared to the 3D7 sequence

66 C.G. Black et al. / Molecular & Biochemical Parasitology 127 (2003) 59–68
(L325S, R406K and H489Q). The latter amino acid substi-tution is located within the second EGF-like domain ofMSP10. The isolates can be divided into three allelic typesbased on their pattern of substitutions in this region. Type 1(IMR143, IMR147, MAD71, NF7) has the same sequenceas 3D7, Type 2 (AA01, IMR144, ItG2, K1, V1) has the se-quence S325, K406, Q489 and Type 3 (FF7) has the sequenceS325, K406, H489. Importantly, all 12 cysteine residues inthe sequenced region are conserved, allowing the formationof the two EGF-like structures.
4. Discussion
TheMSP10andMSP8genes encode proteins of 524 and597 amino acids, respectively which is much smaller thanthe protein encoded byMSP1 (∼1700 amino acids)[33].The extreme N-termini of the two proteins, including theputative hydrophobic signal peptides, show 52.2% identity(78.3% similarity) over a 23 amino acid stretch (data notshown). Together with the predominantly hydrophobic na-ture of the sequence, this finding provides further evidenceto suggest that we have correctly designated the initiatingmethionine in both proteins. As with MSP8, the MSP10 pro-tein has a highly asparagine rich region near the N-terminus.In addition, MSP10 has a block of eight degenerate 6-mertandem repeats with the sequence (N/I)EN(I/V/N)EN span-ning amino acids 126–173 (Fig. 1A).
The lack of cross-reactivity of rabbit antisera raised toMSP10D with the recombinant EGF-like domains of theknown asexual stage merozoite surface proteins ofP. fal-ciparumprovides evidence that we have identified a novelprotein. Immunoprecipitation experiments using antiserumto MSP10D have confirmed this lack of cross-reactivity un-der native conditions in the parasite (Fig. 3A). Furthermore,co-precipitation was not detected with other known mero-zoite surface proteins (MSP1, MSP4, MSP5 and MSP8) pro-viding no evidence to suggest that these proteins form acomplex inP. falciparum(data not shown).
MSP1 undergoes a series of proteolytic processing eventsto generate four polypeptides that form a non-covalentcomplex on the merozoite surface. The 19 kDa C-terminalfragment (MSP119) contains two EGF-like domains, andis the only part of MSP1 carried into the newly infectederythrocyte [35]. We have recently reported that MSP8appears to undergo a similar processing event to liberatea C-terminal fragment that is carried into the developingrings [16]. The results of the present study indicate that theMSP10 protein may also be processed to generate a 36 kDaC-terminal polypeptide that contains the two EGF-like do-mains. Antibodies to MSP10A reacts with the full lengthprotein (80 kDa) but not to the smaller fragment recognisedby antisera to MSP10B, MSP10C and MSP10D. We havenot been able to detect any C-terminal fragment of MSP10in newly invaded rings, thus it would appear that no part ofMSP10 is carried into the ring, a feature shared with MSP2,
MSP4, MSP5 and MSP8[16,20,24,34]. Pulse-chase andprocessing inhibition experiments would be useful to morefully define the presence and nature of MSP10 processing.
Although the data do not formally demonstrate a precur-sor product relationship between the 80 and 36 kDa peptidesrecognised by the MSP10 antisera, the stage-specific im-munoblots show clear evidence of MSP10 expression andpotential processing by 33 h, with maximal expression at44–47 h post-synchronisation. This corresponds to the timeof appearance of trophozoites and schizonts inP. falciparumcultures. We have shown previously that cleavage of fulllength MSP8 commences at 21 h and continues through therest of the asexual life cycle[16]. Expression and processingof MSP1 would start somewhat later than that of MSP10,occurring around the time of schizogony at 44 h onwards(data not shown). This result suggests that the malaria para-site has a functional requirement for both MSP10 and MSP8in the time leading up to schizogony and re-invasion. It isnot known whether the same protease that cleaves MSP1would also be responsible for cleavage of MSP8 and MSP10,but since this protease is produced late in schizont matu-ration and is found in small vesicles, it seems more likelythat a different protease is involved. Candidates for this roleinclude a number of related proteases present in the de-veloping parasite that have been found by genomic search-ing [36–38]. Consistent with the presence of a C-terminalGPI moiety, MSP10 was found to partition in the TritonX-114-soluble phase, a property shared by several integralmembrane proteins ofP. falciparumincluding MSP2[28],MSP4 [24], MSP5 [20,34], MSP8 [16] and AMA1 [39].Confocal microscopy of indirect immunofluorescence as-says demonstrated a rim-like staining pattern for MSP10around the circumference of trophozoites and merozoites,both within intact schizonts and as free merozoites releasedfrom the rupturing schizont. The combination of these find-ings provides strong evidence that MSP10 is present on thesurface of the merozoite. Interestingly, in merozoites MSP10was also found to be associated with an apical organelle con-sistent with the rhoptries or, alternatively, located over theapical polar prominence. The finding is analogous to MSP5which has a complex pattern of distribution being foundboth as small dots or evenly distributed over the surface ofmerozoites ([20,40] and our unpublished observations).
We found that MSP10D was more strongly recognised byhuman immune sera than the other three regions of MSP10.This finding contrasts with results obtained for MSP4[25],MSP8 [16] and MSP119 [41] which indicated that thoseEGF-like domains are poorly immunogenic in humans. Thereduction sensitivity of MSP10D does however agree withthe findings of Egan et al.[42], which demonstrated that an-tibodies to MSP119 in immune human sera tend to recognisedisulphide bond-dependent epitopes, although minor linearepitopes are also present.
The results of this study indicate a very limited degreeof sequence diversity between MSP10 sequences fromvarious laboratory isolates ofP. falciparum. Only three

C.G. Black et al. / Molecular & Biochemical Parasitology 127 (2003) 59–68 67
non-synonymous mutations were detected in a 951 bp re-gion of the gene. One of the resulting amino acid substi-tutions is located within the sequence encoding the secondEGF-like domain of MSP10. The degree of variability inMSP1 is in general much higher. Amino acid substitutionshave been shown to occur at four positions in the EGF-likedomains of MSP1[43,44] and there are multiple sequencedifferences elsewhere in the protein. Sequence diversityin MSP8 is quite limited and no variation in either of theEGF-like domains has yet been reported[16].
The exposed location of MSP10 and its similarity in struc-tural features to MSP1, a protein known to have an importantrole in red cell invasion suggests the possibility that MSP10may have some similar role. To investigate this hypothe-sis, antisera raised against recombinant GST fusion proteinsMSP10A, MSP10B, MSP10C and MSP10D were assessedfor the ability to inhibit P. falciparum3D7 multiplicationin vitro. Each of the four anti-MSP10 antisera was capableof significantly inhibiting parasite growth to similar extentswhen compared to the anti-GST control serum, which hadno effect upon parasite multiplication (data not shown). Itshould be noted, however, that the growth inhibition assayswere conducted using whole serum and will need to be re-peated with purified IgG before any definitive conclusionscan be drawn.
Based on the highly conserved nature of EGF-like do-mains across different species ofPlasmodium[16,45–49],we decided to use a bioinformatics approach in an attemptto identify the Plasmodium yoeliihomologue of MSP10.Searches of the unfinishedP. yoelii genome project data re-leased at the TIGR website (http://www.tigr.org/tdb/edb2/pya1/htmls/) revealed a complete ORF showing significantsimilarity to PfMSP10, particularly within the two EGF-likedomains. We believe that we have identified the PyMSP10protein and we have commenced immunisation and chal-lenge studies in mice to provide further information aboutits importance in the invasion process and to ascertain theprotective efficacy of this novel malarial antigen.
Acknowledgements
This work was supported by research project grants fromthe National Health and Medical Research Council, theUNDP/World Bank/WHO Special Programme for Researchand Training in Tropical Diseases, the Burroughs WellcomeFund and the Howard Hughes Medical Institute Interna-tional Scholars in Infectious Diseases and ParasitologyProgram. Sequence data for chromosomes 6, 7 and 8 wereobtained from the Sanger Centre website athttp://www.sanger.ac.uk/Projects/Pfalciparum/. Sequencing ofP. falci-parumchromosomes 6, 7 and 8 was accomplished as partof the Malaria Genome Project with support by the Well-come Trust. We thank Tony Holder for kindly providing theanti-MSP119 mAb 1E1 and Agnieszka Topolska for kindlyproviding the anti-RAMA antisera.
References
[1] Chitnis CE. Molecular insights into receptors used by malaria para-sites for erythrocyte invasion. Curr Opin Hematol 2001;8:85–91.
[2] Hadley TJ, Klotz FW, Pasvol G, Haynes JD, McGinniss MH. Fal-ciparum malaria parasites invade erythrocytes that lack glycophorinA and B (MkMk). Strain differences indicate receptor heterogeneityand two pathways for invasion. J Clin Invest 1987;80:1190–3.
[3] Okoyeh JN, Pillai CR, Chitnis CE.Plasmodium falciparumfieldisolates commonly use erythrocyte invasion pathways that are in-dependent of sialic acid residues of glycophorin A. Infect Immun1999;67:5784–91.
[4] Gardner MJ, Tettelin H, Carucci DJ, et al. Chromosome 2 se-quence of the human malaria parasitePlasmodium falciparum. Sci-ence 1998;282:1126–32.
[5] Gardner MJ, Tettelin H, Carucci DJ, et al. The malaria genomesequencing project: complete sequence ofPlasmodium falciparumchromosome 2. Parasitologia 1999;41:69–75.
[6] Rayner JC, Vargas-Serrato E, Huber CS, Galinski MR, Barnwell JW.A Plasmodium falciparumhomologue ofPlasmodium vivaxreticulo-cyte binding protein (PvRBP1) defines a trypsin-resistant erythrocyteinvasion pathway. J Exp Med 2001;194:1571–81.
[7] Rayner JC, Galinski MR, Ingravallo P, Barnwell JW. TwoPlasmod-ium falciparum genes express merozoite proteins that are relatedto Plasmodium vivaxand Plasmodium yoeliiadhesive proteins in-volved in host cell selection and invasion. Proc Natl Acad Sci USA2000;97:9648–53.
[8] Trucco C, Fernandez-Reyes D, Howell S, et al. The merozoite surfaceprotein 6 gene codes for a 36 kDa protein associated with thePlasmodium falciparummerozoite surface protein-1 complex. MolBiochem Parasitol 2001;112:91–101.
[9] McColl DJ, Anders RF. Conservation of structural motifs and anti-genic diversity in thePlasmodium falciparummerozoite surfaceprotein-3 (MSP-3). Mol Biochem Parasitol 1997;90:21–31.
[10] Cowman AF, Baldi DL, Healer J, et al. Functional analysis of proteinsinvolved in Plasmodium falciparummerozoite invasion of red bloodcells. FEBS Lett 2000;476:84–8.
[11] Cowman AF, Baldi DL, Duraisingh M, et al. Functional analysis ofPlasmodium falciparummerozoite antigens: implications for erythro-cyte invasion and vaccine development. Philos Trans R Soc Lond BBiol Sci 2002;357:25–33.
[12] Mills KE, Pearce JA, Crabb BS, Cowman AF. Truncation of mero-zoite surface protein 3 disrupts its trafficking and that of acidic-basicrepeat protein to the surface ofPlasmodium falciparummerozoites.Mol Microbiol 2002;43:1401–11.
[13] Blackman MJ, Holder AA. Secondary processing of thePlas-modium falciparum merozoite surface protein-1 (MSP1) by acalcium-dependent membrane-bound serine protease: shedding ofMSP133 as a noncovalently associated complex with other fragmentsof the MSP1. Mol Biochem Parasitol 1992;50:307–15.
[14] Stafford WH, Gunder B, Harris A, Heidrich HG, Holder AA, Black-man MJ. A 22 kDa protein associated with thePlasmodium falci-parummerozoite surface protein-1 complex. Mol Biochem Parasitol1996;80:159–69.
[15] Pachebat JA, Ling IT, Grainger M, et al. The 22 kDa componentof the protein complex on the surface of Plasmodium falciparummerozoites is derived from a larger precursor merozoite surfaceprotein 7. Mol Biochem Parasitol 2001;117:83–9.
[16] Black CG, Wu T, Wang L, Hibbs AR, Coppel RL. Merozoite surfaceprotein 8 ofPlasmodium falciparumcontains two epidermal growthfactor-like domains. Mol Biochem Parasitol 2001;114:217–26.
[17] Trager W, Jensen JB. Human malaria parasites in continuous culture.Science 1976;193:673–5.
[18] Rosenthal PJ.Plasmodium falciparum: effects of proteinase inhibitorson globin hydrolysis by cultured malaria parasites. Exp Parasitol1995;80:272–81.

68 C.G. Black et al. / Molecular & Biochemical Parasitology 127 (2003) 59–68
[19] Holder AA, Freeman RR. Biosynthesis and processing of aPlasmodium falciparumschizont antigen recognized by immuneserum and a monoclonal antibody. J Exp Med 1982;156:1528–38.
[20] Wu T, Black CG, Wang L, Hibbs AR, Coppel RL. Lack of sequencediversity in the gene encoding merozoite surface protein 5 ofPlas-modium falciparum. Mol Biochem Parasitol 1999;103:243–50.
[21] Allen MJ, Collick A, Jeffreys AJ. Use of vectorette and subvec-torette PCR to isolate transgene flanking DNA. PCR Methods Appl1994;4:71–5.
[22] Ochman H, Gerber AS, Hartl DL. Genetic applications of an inversepolymerase chain reaction. Genetics 1988;120:621–3.
[23] Wang L, Black CG, Marshall VM, Coppel RL. Structural and anti-genic properties of merozoite surface protein 4 ofPlasmodium fal-ciparum. Infect Immun 1999;67:2193–200.
[24] Marshall VM, Silva A, Foley M, et al. A second merozoite surfaceprotein (MSP-4) ofPlasmodium falciparumthat contains an epider-mal growth factor-like domain. Infect Immun 1997;65:4460–7.
[25] Wang L, Richie TL, Stowers A, Nhan DH, Coppel RL. Naturallyacquired antibody responses toPlasmodium falciparummerozoitesurface protein 4 in a population living in an area of endemicity inVietnam. Infect Immun 2001;69:4390–7.
[26] Uthaipibull C, Aufiero B, Syed SE, et al. Inhibitory and blockingmonoclonal antibody epitopes on merozoite surface protein 1 ofthe malaria parasitePlasmodium falciparum. J Mol Biol 2001;307:1381–94.
[27] Chakrabarti D, Reddy GR, Dame JB, et al. Analysis of expressedsequence tags fromPlasmodium falciparum. Mol Biochem Parasitol1994;66:97–104.
[28] Smythe JA, Coppel RL, Brown GV, Ramasamy R, Kemp DJ, AndersRF. Identification of two integral membrane proteins ofPlasmodiumfalciparum. Proc Natl Acad Sci USA 1988;85:5195–9.
[29] Smythe JA, Murray PJ, Anders RF. Improved temperature-dependentphase separation using Triton X-114: isolation of integral mem-brane proteins of pathogenic parasites. J Methods Cell Mol Biol1990;2:133–7.
[30] Renia L, Ling IT, Marussig M, Miltgen F, Holder AA, Mazier D. Im-munization with a recombinant C-terminal fragment ofPlasmodiumyoelii merozoite surface protein 1 protects mice against homologousbut not heterologousP. yoelii sporozoite challenge. Infect Immun1997;65:4419–23.
[31] Crewther PE, Matthew ML, Flegg RH, Anders RF. Protective im-mune responses to apical membrane antigen 1 ofPlasmodiumchabaudi involve recognition of strain-specific epitopes. Infect Im-mun 1996;64:3310–7.
[32] Rotman HL, Daly TM, Long CA. Plasmodium: immunization withcarboxyl-terminal regions of MSP-1 protects against homologousbut not heterologous blood-stage parasite challenge. Exp Parasitol1999;91:78–85.
[33] Holder AA, Blackman MJ, Burghaus PA, et al. A malaria merozoitesurface protein (MSP1)-structure, processing and function. Mem InstOswaldo Cruz 1992;87:37–42.
[34] Marshall VM, Tieqiao W, Coppel RL. Close linkage of three mero-zoite surface protein genes on chromosome 2 ofPlasmodium falci-parum. Mol Biochem Parasitol 1998;94:13–25.
[35] Blackman MJ, Heidrich HG, Donachie S, McBride JS, HolderAA. A single fragment of a malaria merozoite surface protein re-mains on the parasite during red cell invasion and is the target ofinvasion-inhibiting antibodies. J Exp Med 1990;172:379–82.
[36] Blackman MJ, Fujioka H, Stafford WH, et al. A subtilisin-like proteinin secretory organelles ofPlasmodium falciparummerozoites. J BiolChem 1998;273:23398–409.
[37] Sajid M, Withers-Martinez C, Blackman MJ. Maturation and speci-ficity of Plasmodium falciparumsubtilisin-like protease-1, a malariamerozoite subtilisin-like serine protease. J Biol Chem 2000;275:631–41.
[38] Hackett F, Sajid M, Withers-Martinez C, Grainger M, Blackman MJ.PfSUB-2: a second subtilisin-like protein inPlasmodium falciparummerozoites. Mol Biochem Parasitol 1999;103:183–95.
[39] Crewther PE, Culvenor JG, Silva A, Cooper JA, Anders RF.Plas-modium falciparum: two antigens of similar size are located in dif-ferent compartments of the rhoptry. Exp Parasitol 1990;70:193–206.
[40] Black CG, Barnwell JW, Huber CS, Galinski MR, Coppel RL. ThePlasmodium vivaxhomologues of merozoite surface proteins 4 and5 from Plasmodium falciparumare expressed at different locationsin the merozoite. Mol Biochem Parasitol 2002;120:215–24.
[41] Egan AF, Waterfall M, Pinder M, Holder AA, Riley E. Characteri-zation of human T- and B-cell epitopes in the C terminus ofPlas-modium falciparummerozoite surface protein 1: evidence for poorT- cell recognition of polypeptides with numerous disulfide bonds.Infect Immun 1997;65:3024–31.
[42] Egan AF, Chappel JA, Burghaus PA, et al. Serum antibodiesfrom malaria-exposed people recognize conserved epitopes formedby the two epidermal growth factor motifs of MSP1(19), thecarboxy-terminal fragment of the major merozoite surface protein ofPlasmodium falciparum. Infect Immun 1995;63:456–66.
[43] Miller LH, Roberts T, Shahabuddin M, McCutchan TF. Analysis ofsequence diversity in thePlasmodium falciparummerozoite surfaceprotein-1 (MSP-1). Mol Biochem Parasitol 1993;59:1–14.
[44] Qari SH, Shi YP, Goldman IF, Nahlen BL, Tibayrenc M, Lal AA.Predicted and observed alleles ofPlasmodium falciparummerozoitesurface protein-1 (MSP-1), a potential malaria vaccine antigen. MolBiochem Parasitol 1998;92:241–52.
[45] Black CG, Wang L, Hibbs AR, Werner E, Coppel RL. Identifi-cation of thePlasmodium chabaudihomologue of merozoite sur-face proteins 4 and 5 ofPlasmodium falciparum. Infect Immun1999;67:2075–81.
[46] Kedzierski L, Black CG, Coppel RL. Characterization of the mero-zoite surface protein 4/5 gene ofPlasmodium bergheiandPlasmod-ium yoelii. Mol Biochem Parasitol 2000;105:137–47.
[47] Burns JM, Belk CC, Dunn PD. A protective glycosylphospha-tidylinositol-anchored membrane protein of Plasmodium yoeliitrophozoites and merozoites contains two epidermal growth factor-like domains. Infect Immun 2000;68:6189–95.
[48] McKean PG, O’Dea K, Brown KN. Nucleotide sequence analysis andepitope mapping of the merozoite surface protein 1 fromPlasmodiumchabaudi chabaudiAS. Mol Biochem Parasitol 1993;62:199–209.
[49] Zhong H, Fan JY, Yang S, Davidson EA. Cloning and characterizationof the merozoite surface antigen 1 gene ofPlasmodium berghei. AmJ Trop Med Hyg 1999;60:994–9.
Copyright © 2022 FDOKUMEN




















