
Detection of total and hemolysin-producing Vibrio parahaemolyticus in shellfish using multiplex PCR...
11
Journal of Methods Microbiological Journal of Microbiological Methods 36 (1999) 215–225 Detection of total and hemolysin-producing Vibrio parahaemolyticus in shellfish using multiplex PCR amplification of tl, tdh and trh a, a a a * Asim K. Bej , Donald P. Patterson , Cynthia W. Brasher , Michael C.L. Vickery , a b Daniel D. Jones , Charles A. Kaysner a Department of Biology, The University of Alabama at Birmingham, 1300 University Boulevard, Birmingham, AL 35294-1170, USA b Seafood Products Research Center, U.S. Food and Drug Administration, Bothell, WA 98041-3012, USA Abstract Vibrio parahaemolyticus is an important human pathogen which can cause gastroenteritis when consumed in raw or partially-cooked seafood. A multiplex PCR amplification-based detection of total and virulent strains of V . parahaemolyticus was developed by targeting thermolabile hemolysin encoded by tl, thermostable direct hemolysin encoded by tdh, and thermostable direct hemolysin-related trh genes. Following optimization using oligonucleotide primers targeting tl, tdh and trh genes, the multiplex PCR was applied to V . parahaemolyticus from 27 clinical, 43 seafood, 15 environmental, 7 strains obtained from various laboratories and 19 from oyster plants. All 111 V . parahaemolyticus isolates showed PCR amplification of the tl gene; however, only 60 isolates showed amplification of tdh, and 43 isolates showed amplification of the trh gene. Also, 18 strains showed amplification of the tdh gene, but these strains did not show amplification of the trh gene. However, one strain exhibited amplification for the trh but not the tdh gene, suggesting both genes need to be targeted in a PCR amplification reaction to detect all hemolysin-producing strains of this pathogen. The multiplex PCR approach was successfully used to detect various strains of V . parahaemolyticus in seeded oyster tissue homogenate. Sensitivity of 1 2 detection for all three target gene segments was at least between 10 –10 cfu per 10 g of alkaline peptone water enriched seeded oyster tissue homogenate. This high level of sensitivity of detection of this pathogen within 8 h of pre-enrichment is 4 well within the action level (10 cfu per 1 g of shell stock) suggested by the National Seafood Sanitation Program guideline. Compared to conventional microbiological culture methods, this multiplex PCR approach is rapid and reliable for accomplishing a comprehensive detection of V . parahaemolyticus in shellfish. 1999 Elsevier Science B.V. All rights reserved. Keywords: Vibrio parahemolyticus; Multiplex PCR; Microorganism; Shellfish; Hemolysin 1. Introduction when consumed in raw, undercooked or mishandled seafood (DePaola et al., 1990). This pathogen, like Vibrio parahaemolytcus is an enteric pathogen that other members of the genus Vibrio, is a Gram- causes acute gastroenteritis in humans primarily negative halophilic bacterium distributed worldwide in the estuarine environment (Janda et al., 1988; Joseph et al., 1983; Kaysner et al., 1992). Recently, two outbreaks of V . parahemolyticus-related illnesses *Corresponding author. Tel.: 11-205-934-9857; fax: 11-205- in the US resulting from raw oyster consumption 975-6097. E-mail address: [email protected] (A.K. Bej) have heightened the need to develop a rapid and 0167-7012 / 99 / $ – see front matter 1999 Elsevier Science B.V. All rights reserved. PII: S0167-7012(99)00037-8
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Detection of total and hemolysin-producing Vibrio parahaemolyticus in shellfish using multiplex PCR...

Journalof
MethodsMicrobiological
Journal of Microbiological Methods 36 (1999) 215–225
Detection of total and hemolysin-producing Vibrio parahaemolyticusin shellfish using multiplex PCR amplification of tl, tdh and trh
a , a a a*Asim K. Bej , Donald P. Patterson , Cynthia W. Brasher , Michael C.L. Vickery ,a bDaniel D. Jones , Charles A. Kaysner
aDepartment of Biology, The University of Alabama at Birmingham, 1300 University Boulevard, Birmingham, AL 35294-1170, USAbSeafood Products Research Center, U.S. Food and Drug Administration, Bothell, WA 98041-3012, USA
Abstract
Vibrio parahaemolyticus is an important human pathogen which can cause gastroenteritis when consumed in raw orpartially-cooked seafood. A multiplex PCR amplification-based detection of total and virulent strains of V. parahaemolyticuswas developed by targeting thermolabile hemolysin encoded by tl, thermostable direct hemolysin encoded by tdh, andthermostable direct hemolysin-related trh genes. Following optimization using oligonucleotide primers targeting tl, tdh andtrh genes, the multiplex PCR was applied to V. parahaemolyticus from 27 clinical, 43 seafood, 15 environmental, 7 strainsobtained from various laboratories and 19 from oyster plants. All 111 V. parahaemolyticus isolates showed PCRamplification of the tl gene; however, only 60 isolates showed amplification of tdh, and 43 isolates showed amplification ofthe trh gene. Also, 18 strains showed amplification of the tdh gene, but these strains did not show amplification of the trhgene. However, one strain exhibited amplification for the trh but not the tdh gene, suggesting both genes need to be targetedin a PCR amplification reaction to detect all hemolysin-producing strains of this pathogen. The multiplex PCR approach wassuccessfully used to detect various strains of V. parahaemolyticus in seeded oyster tissue homogenate. Sensitivity of
1 2detection for all three target gene segments was at least between 10 –10 cfu per 10 g of alkaline peptone water enrichedseeded oyster tissue homogenate. This high level of sensitivity of detection of this pathogen within 8 h of pre-enrichment is
4well within the action level (10 cfu per 1 g of shell stock) suggested by the National Seafood Sanitation Program guideline.Compared to conventional microbiological culture methods, this multiplex PCR approach is rapid and reliable foraccomplishing a comprehensive detection of V. parahaemolyticus in shellfish. 1999 Elsevier Science B.V. All rightsreserved.
Keywords: Vibrio parahemolyticus; Multiplex PCR; Microorganism; Shellfish; Hemolysin
1. Introduction when consumed in raw, undercooked or mishandledseafood (DePaola et al., 1990). This pathogen, like
Vibrio parahaemolytcus is an enteric pathogen that other members of the genus Vibrio, is a Gram-causes acute gastroenteritis in humans primarily negative halophilic bacterium distributed worldwide
in the estuarine environment (Janda et al., 1988;Joseph et al., 1983; Kaysner et al., 1992). Recently,two outbreaks of V. parahemolyticus-related illnesses*Corresponding author. Tel.: 11-205-934-9857; fax: 11-205-in the US resulting from raw oyster consumption975-6097.
E-mail address: [email protected] (A.K. Bej) have heightened the need to develop a rapid and
0167-7012/99/$ – see front matter 1999 Elsevier Science B.V. All rights reserved.PI I : S0167-7012( 99 )00037-8

216 A.K. Bej et al. / Journal of Microbiological Methods 36 (1999) 215 –225
reliable method to detect this pathogen in shellfish negative, whereas some showed amplification for trh(Nishibuchi and Kaper, 1985). The pathogenic effect but not for tdh (Lee and Pan, 1993). DNA–DNAfor humans following infection has been associated hybridization, using tdh- or trh-specific probeswith those strains that produce thermostable direct (Nishibuchi et al., 1985; Nishibuchi et al., 1986;hemolysin (TDH) (Miyamoto et al., 1969; Honda Yamamoto et al., 1992), or use of restriction frag-and Iida, 1993). Production of TDH by a strain of ment length polymorphism (RFLP) of the tdh andthis pathogen is referred to as Kanagawa phenom- trh genes (Suthienkul et al., 1996) produced resultsenon positive and can be identified by b-type comparable to the PCR study. In addition, DNA–hemolysis on Wagatsuma blood agar (Wagatsuma, DNA hybridization using the tdh gene as a probe1968; Takeda, 1982; Kaysner et al., 1992; Morbidity showed positive hybridization with the tdh-like geneand Mortality Weekly Report, 1998). However, the in non-parahaemolyticus strains of vibrios such as V.testing for Kanagawa phenomenon is time consum- hollisae, V. mimicus and V. cholerae non-O1ing; the agar medium is cumbersome to prepare, and (Nishibuchi et al., 1985; Nishibuchi et al., 1986;the test may not be determinative since many clinical Yamamoto et al., 1992; Honda and Iida, 1993).isolates are Kanagawa negative and do not produce Therefore, using the tdh gene exclusively as a probeTDH (Miyamoto et al., 1969; Tada et al., 1992; for DNA–DNA hybridization is not reliable forOkuda and Nishibuchi, 1998). Similarly, immuno- specific detection of V. parahaemolyticus. In separatelogical methods, although shown to be highly sensi- studies, PCR amplification methodology was used totive for detecting TDH, fail to detect clinical strains detect V. parahaemolyticus isolates by targeting onlythat lack TDH (Miyamoto et al., 1969; Honda et al., the tdh gene fragment (Lee and Pan, 1993) or a1980; Tada et al., 1992; Lee and Pan, 1993; Okuda 0.71-kbp cloned segment of the chromosomal DNA,and Nishibuchi, 1998). Furthermore, DNA probes pR72H, of unknown function (Lee et al., 1995). Aand oligonucleotide probes specific for the gene separate gene, thermolabile hemolysin (tl), has beenencoding for thermostable hemolysin (TDH) and a characterized in this pathogen which does not seemtdh-related gene encoding for TRH (Honda et al., to cause hemolysis on Wagatsuma agar (Taniguchi et1991; Kishishita et al., 1992) were shown to react al., 1985, 1986). Also, the thermolabile hemolysinpositively for TDH-negative and several non- has not been reported to cause virulence in humansparahaemolyticus strains such as V. hollisae, V. as all human isolates are found to be either positivemimicus, and V. cholerae non-O1 (Tada et al., 1992; for thermostable direct hemolysin or thermostable-Lee et al., 1995). Subsequent studies using tdh or trh related hemolysin or both. However, this gene wasgene probes on V. parahaemolyticus strains showed a shown to be present in all of the V. parahaemolyticusstrong correlation between clinically significant strains tested previously (Taniguchi et al., 1985,strains and presence of either of these genes, sug- 1986; A. Bej, unpublished). Although in previouslygesting that both tdh and trh genes are virulence reported studies, the tdh or the trh genes were usedfactors in V. parahaemolyticus (Miyamoto et al., as targets, the PCR- and gene probe-based detection1969; Shirai et al., 1990). Presently, there is no in of this pathogen as a species specific target has notvitro test that will demonstrate the production of been sought so far. Therefore, a multiplex PCR-TRH in this pathogen. Genetic analysis showed that based detection system targeting tl gene for total, andthe trh gene is closely related to the tdh gene with both tdh and trh genes for hemolysin-producingapproximately 68% nucleotide sequence homology pathogenic strains of V. parahaemolyticus is neces-(Honda et al., 1991; Tada et al., 1992; Honda and sary for comprehensive detection of this pathogen inIida, 1993). Moreover, variability in nucleotide shellfish. In this study, we describe development ofsequences within the trh gene isolated from various such a multiplex PCR-amplification approach toclinical strains leads to the establishment of two detect total and hemolysin-producing pathogenicsubdivisions of this gene, trh1 and trh2 (Kishishita et strains of V. parahaemolyticus in shellfish by simul-al., 1992). In a PCR amplification-based study, a taneously targeting the tl, tdh and trh genes in anumber of virulent strains were tdh positive but trh single PCR reaction.
A.K. Bej et al. / Journal of Microbiological Methods 36 (1999) 215 –225 217
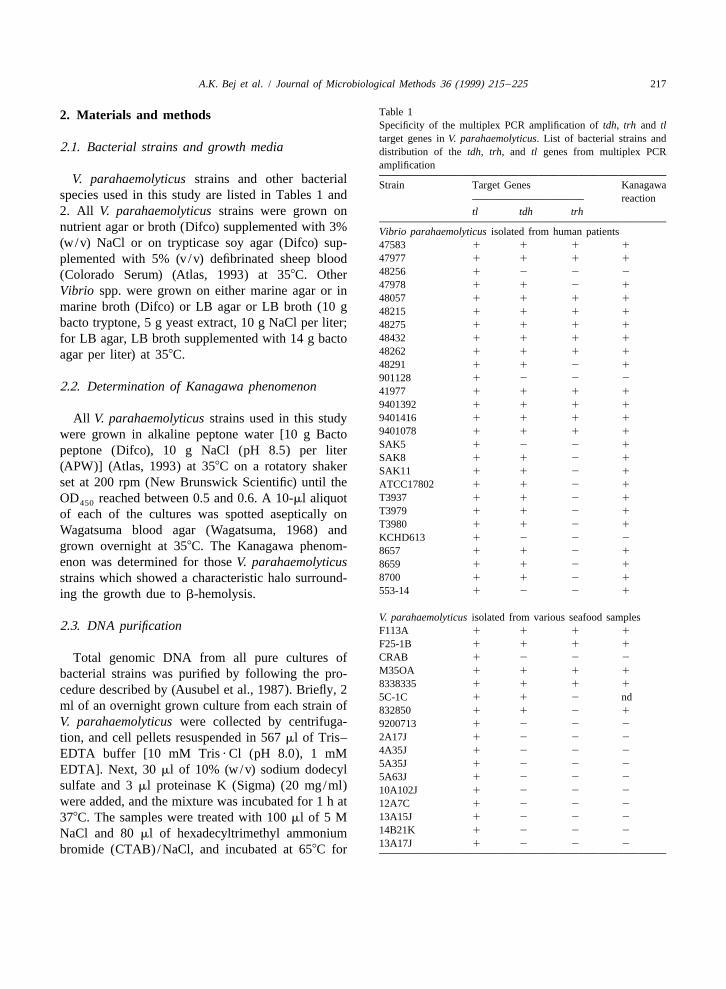
Table 12. Materials and methodsSpecificity of the multiplex PCR amplification of tdh, trh and tltarget genes in V. parahaemolyticus. List of bacterial strains and
2.1. Bacterial strains and growth media distribution of the tdh, trh, and tl genes from multiplex PCRamplification
V. parahaemolyticus strains and other bacterial Strain Target Genes Kanagawaspecies used in this study are listed in Tables 1 and reaction2. All V. parahaemolyticus strains were grown on tl tdh trh
nutrient agar or broth (Difco) supplemented with 3% Vibrio parahaemolyticus isolated from human patients(w/v) NaCl or on trypticase soy agar (Difco) sup- 47583 1 1 1 1
47977 1 1 1 1plemented with 5% (v/v) defibrinated sheep blood48256 1 2 2 2(Colorado Serum) (Atlas, 1993) at 358C. Other47978 1 1 2 1Vibrio spp. were grown on either marine agar or in48057 1 1 1 1
marine broth (Difco) or LB agar or LB broth (10 g 48215 1 1 1 1bacto tryptone, 5 g yeast extract, 10 g NaCl per liter; 48275 1 1 1 1
48432 1 1 1 1for LB agar, LB broth supplemented with 14 g bacto48262 1 1 1 1agar per liter) at 358C.48291 1 1 2 1
901128 1 2 2 22.2. Determination of Kanagawa phenomenon 41977 1 1 1 1
9401392 1 1 1 1
9401416 1 1 1 1All V. parahaemolyticus strains used in this study9401078 1 1 1 1were grown in alkaline peptone water [10 g BactoSAK5 1 2 2 1peptone (Difco), 10 g NaCl (pH 8.5) per literSAK8 1 1 2 1
(APW)] (Atlas, 1993) at 358C on a rotatory shaker SAK11 1 1 2 1set at 200 rpm (New Brunswick Scientific) until the ATCC17802 1 1 2 1
T3937 1 1 2 1OD reached between 0.5 and 0.6. A 10-ml aliquot450T3979 1 1 2 1of each of the cultures was spotted aseptically onT3980 1 1 2 1Wagatsuma blood agar (Wagatsuma, 1968) andKCHD613 1 2 2 2
grown overnight at 358C. The Kanagawa phenom- 8657 1 1 2 1enon was determined for those V. parahaemolyticus 8659 1 1 2 1
8700 1 1 2 1strains which showed a characteristic halo surround-553-14 1 2 2 1ing the growth due to b-hemolysis.
V. parahaemolyticus isolated from various seafood samples2.3. DNA purification F113A 1 1 1 1
F25-1B 1 1 1 1
CRAB 1 2 2 2Total genomic DNA from all pure cultures ofM35OA 1 1 1 1bacterial strains was purified by following the pro-8338335 1 1 1 1cedure described by (Ausubel et al., 1987). Briefly, 25C-1C 1 1 2 nd
ml of an overnight grown culture from each strain of 832850 1 1 2 1V. parahaemolyticus were collected by centrifuga- 9200713 1 2 2 2
2A17J 1 2 2 2tion, and cell pellets resuspended in 567 ml of Tris–4A35J 1 2 2 2EDTA buffer [10 mM Tris ? Cl (pH 8.0), 1 mM5A35J 1 2 2 2EDTA]. Next, 30 ml of 10% (w/v) sodium dodecyl5A63J 1 2 2 2
sulfate and 3 ml proteinase K (Sigma) (20 mg/ml) 10A102J 1 2 2 2were added, and the mixture was incubated for 1 h at 12A7C 1 2 2 2
13A15J 1 2 2 2378C. The samples were treated with 100 ml of 5 M14B21K 1 2 2 2NaCl and 80 ml of hexadecyltrimethyl ammonium13A17J 1 2 2 2bromide (CTAB)/NaCl, and incubated at 658C for

218 A.K. Bej et al. / Journal of Microbiological Methods 36 (1999) 215 –225
Table 1. Continued Table 1. Continued
Strain Target Genes Kanagawa Strain Target Genes Kanagawareaction reaction
tl tdh trh tl tdh trh
855329-2 1 2 2 2 JJ41B1 1 1 1 1
855330-5B 1 2 2 2 JJ41B2 1 1 1 1
855330-1A 1 1 1 1 MMX4 1 2 2 2
9736165 1 2 2 2 BB5 1 1 1 1
95046527 1 1 1 1 FF5 1 1 1 1
95046528 1 2 2 2 JJ51A 1 1 1 1
18V10B 1 1 2 2 MMX5 1 1 1 1
3V0C 1 1 1 1 B71B 1 1 1 1
G181 1 1 1 1 M7 1 1 1 1
WR5 1 2 2 2 N7 1 1 1 1
83320964 1 2 2 2 B8 1 2 2 2
CO5-1A 1 2 2 2
33V1C 1 1 2 2 V. parahaemolyticus obtained from various laboratories6V-1A 1 1 1 1 14D1 1 1 1 1
8G5 1 2 2 2 14D10 1 1 2 1
96736341 1 2 2 2 14D13 1 2 2 2
27V10B 1 2 2 2 14D14 1 2 2 2
30V10A 1 1 1 2 15A17J 1 2 2 2
M25OB 1 2 2 2 NY 477 1 1 2 1
AOC1 1 2 2 2 AQ4037 1 2 1 2
AOC2 1 2 2 2
AOC3 1 2 2 2 Non-V. parahaemolyticus strainsAOC4 1 2 2 2 Vibrio hollisaeAOC5 1 2 2 2 89A1960 2 1 2 2
AOC6 1 2 2 2 89A1961 2 2 2 2
AOC7 1 2 2 2 89A7503 2 1 2 2
89A4206 2 2 2 2
V. parahaemolyticus isolated from environmental watersVibrio cholerae
W9OA 1 1 1 189A4555 2 2 2 2
WR1 1 2 2 2O145B 2 2 2 2
WR2 1 2 2 10138 2 2 2 2
VP89 1 1 1 1C153PY 2 2 2 2
VP89-1B 1 2 2 2154 (O1) 2 2 2 2
OR152 1 1 1 1569B (O1) 2 2 2 2
VP5G 1 1 1 1
VP60 1 1 1 1 V. mimicus 2 2 2 2
VP35 1 1 1 1 V. vulnificus 2 2 2 2
VP43-1A 1 1 1 1 V. alginolyticus 2 2 2 2
35V0A 1 2 2 2 V. damsela 2 1 2 2
VP53 1 2 2 2 V. fluvialis 2 2 2 2
VP99 1 2 2 2 V. furnissii 2 2 2 2
96Q 1 2 2 2 V. metschnikovii 2 2 2 2
D3 OA 1 2 2 2 V. proteolyticus 2 2 2 2
Plesiomonas spp. 2 2 2 2
V. parahaemolyticus isolated from oyster plants15presence.
B1A 1 2 2 225absence.
C2 1A 1 2 2 2nd5not determined
JJ2J1C 1 1 2 1
C3 1A 1 1 1 1
W3 1 1 1 1 10 min. Following incubation, the sample was firstMM3 1 2 2 2 extracted with an equal volume of chloroform–iso-C4-3B 1 1 1 1
amyl alcohol (24:1, v /v) and then with phenol–J4 1 1 1 1

A.K. Bej et al. / Journal of Microbiological Methods 36 (1999) 215 –225 219
Table 2Summary of distribution and occurence of tl, tdh and trh genes among V. parahaemolyticus and non-V. parahaemolyticus isolates tested bymultiplex PCR amplification
Target Sources of V. parahaemolyticus strains isolatesgenes
aClinical Food Environmental Oyster plant Obtained from Totalisolates isolates isolates isolates various labs.
Percent distribution of individual genestl 27/27 43/43 15/15 19/19 7/7 111/111
(100%) (100%) (100%) (100%) (100%) (100%)tdh 22/27 14/43 7/15 14/19 3/7 60/111
(81.48%) (32.56%) (46.66%) (73.68%) (42.8%) (54%)trh 11/27 10/43 7/15 13/19 2/7 43/111
(40.74%) (2.3%) (46.66%) (68.4%) (28.57%) (38.73%)Percent distribution of genes in various combinations
tl 1 tdh 1 trh 1 11/27 10/43 7/15 13/19 1/7 42/111(40.74%) (23.25%) (46.66%) (68.4%) (14.28%) (37.8%)
tl 1 tdh 1 trh 2 11/27 4/43 0/15 1/19 2/7 18/111(40.74%) (9.3%) (0%) (5.2%) (28.5%) (16.2%)
tl 1 tdh 2 trh 1 0/27 0/43 0/15 0/19 1/7 1/111(0%) (0%) (0%) (0%) (14.28%) (0.9%)
tl 1 tdh 2 trh 2 5/27 29/43 8/15 5/19 3/7 50/111(18.51%) (67.44%) (53.33%) (26.3%) (42.85%) (45%)
a Distribution of individual genes or in combinations in all 111 V. parahaemolyticus isolates tested in this study.
chloroform–isoamyl alcohol (25:24:2, v /v). The 2.5. Multiplex PCR amplificationsDNA was precipitated with 0.6 volume of 100% coldisopropanol and washed with 1 ml of 70% cold ethyl Multiplex PCR amplification was optimized in aalcohol. The DNA pellet was dried in a DNA 120 50-ml reaction consisting of 0.5 mg of purifiedspeedvac (Savant) for 10 min and resuspended in genomic DNA from V. parahaemolyticus F113ATris ? EDTA buffer (pH 8.0). An aliquot (typically strain, 1 mM of each of the oligonucleotide primers1–2 ml) of the sample was subjected to spectro- for tl, tdh, trh (2.5 ml of each of the primers from aphotometric analysis at optical densities of 260 and 20 mM stock suspension), 5 ml of a 103 PCR280 nm wavelengths to determine the purity and reaction buffer (103 buffer consisted of 500 mMefficiency of recovery. Tris?Cl, pH 8.9, 500 mM KCl and 25 mM, 30 mM
or 40 mM MgCl ; final concentration of 13), 2002
mM of each of the dNTPs (8 ml from a 5 mM stock2.4. Oligonucleotide primers dNTP) (Pharmacia), 2.5 units AmpliTaq DNA poly-
merase (Perkin Elmer) and an appropriate volume ofNucleotide sequences, locations, melting tempera- sterile MilliQ water (Millipore).
tures (T ) of oligonucleotide primers specific for the All multiplex PCR amplifications were performedm
tl, tdh, and trh genes, and size of the amplicons in a DNA thermal cycler (Perkin Elmer, Model 480)following PCR amplification are described in Table using the following temperature-cycling parameters:3. The T value for each of the primers was initial denaturation at 948C for 3 min followed by 30m
estimated by using the equation, T (8C)52(A1 cycles of amplification; each cycle consisted ofm
T)14(G1C) (Suggs et al., 1981). All oligonucleo- denaturation at 948C for 1 min, primer annealing attide primers were custom-synthesized by Integrated 588C for 1 min, and primer extension at 728C for 1DNA Technology, Inc., Coralville, IA. min. Following the amplification cycles, samples
220 A.K. Bej et al. / Journal of Microbiological Methods 36 (1999) 215 –225
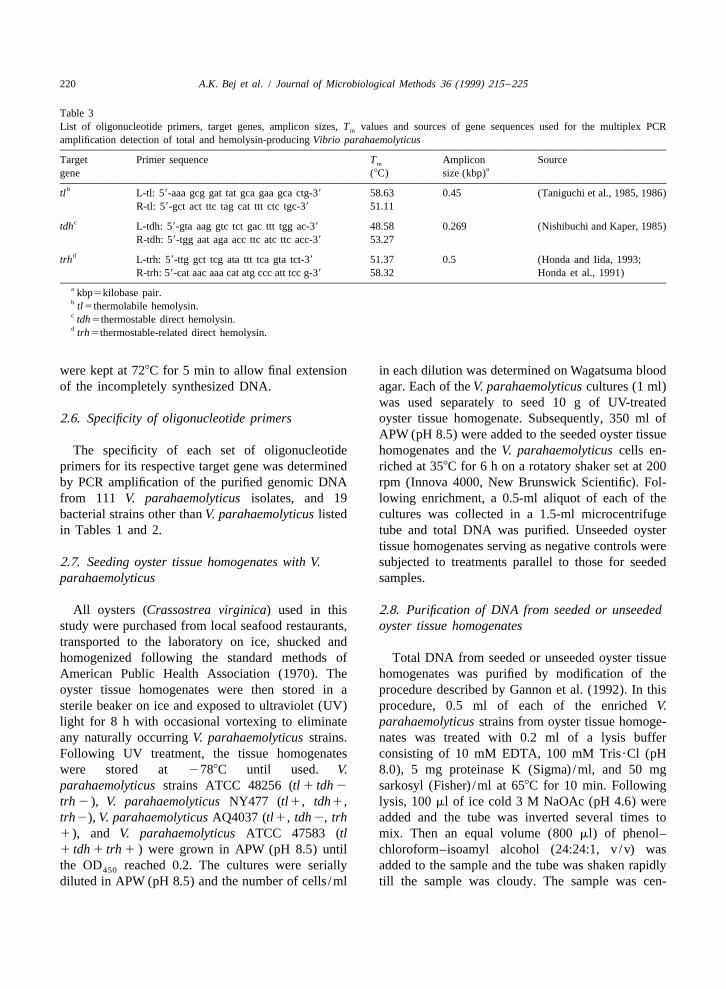
Table 3List of oligonucleotide primers, target genes, amplicon sizes, T values and sources of gene sequences used for the multiplex PCRm
amplification detection of total and hemolysin-producing Vibrio parahaemolyticus
Target Primer sequence T Amplicon Sourcemagene (8C) size (kbp)
btl L-tl: 59-aaa gcg gat tat gca gaa gca ctg-39 58.63 0.45 (Taniguchi et al., 1985, 1986)R-tl: 59-gct act ttc tag cat ttt ctc tgc-39 51.11
ctdh L-tdh: 59-gta aag gtc tct gac ttt tgg ac-39 48.58 0.269 (Nishibuchi and Kaper, 1985)R-tdh: 59-tgg aat aga acc ttc atc ttc acc-39 53.27
dtrh L-trh: 59-ttg gct tcg ata ttt tca gta tct-39 51.37 0.5 (Honda and Iida, 1993;R-trh: 59-cat aac aaa cat atg ccc att tcc g-39 58.32 Honda et al., 1991)
a kbp5kilobase pair.b tl5thermolabile hemolysin.c tdh5thermostable direct hemolysin.d trh5thermostable-related direct hemolysin.
were kept at 728C for 5 min to allow final extension in each dilution was determined on Wagatsuma bloodof the incompletely synthesized DNA. agar. Each of the V. parahaemolyticus cultures (1 ml)
was used separately to seed 10 g of UV-treated2.6. Specificity of oligonucleotide primers oyster tissue homogenate. Subsequently, 350 ml of
APW (pH 8.5) were added to the seeded oyster tissueThe specificity of each set of oligonucleotide homogenates and the V. parahaemolyticus cells en-
primers for its respective target gene was determined riched at 358C for 6 h on a rotatory shaker set at 200by PCR amplification of the purified genomic DNA rpm (Innova 4000, New Brunswick Scientific). Fol-from 111 V. parahaemolyticus isolates, and 19 lowing enrichment, a 0.5-ml aliquot of each of thebacterial strains other than V. parahaemolyticus listed cultures was collected in a 1.5-ml microcentrifugein Tables 1 and 2. tube and total DNA was purified. Unseeded oyster
tissue homogenates serving as negative controls were2.7. Seeding oyster tissue homogenates with V. subjected to treatments parallel to those for seededparahaemolyticus samples.
All oysters (Crassostrea virginica) used in this 2.8. Purification of DNA from seeded or unseededstudy were purchased from local seafood restaurants, oyster tissue homogenatestransported to the laboratory on ice, shucked andhomogenized following the standard methods of Total DNA from seeded or unseeded oyster tissueAmerican Public Health Association (1970). The homogenates was purified by modification of theoyster tissue homogenates were then stored in a procedure described by Gannon et al. (1992). In thissterile beaker on ice and exposed to ultraviolet (UV) procedure, 0.5 ml of each of the enriched V.light for 8 h with occasional vortexing to eliminate parahaemolyticus strains from oyster tissue homoge-any naturally occurring V. parahaemolyticus strains. nates was treated with 0.2 ml of a lysis bufferFollowing UV treatment, the tissue homogenates consisting of 10 mM EDTA, 100 mM Tris?Cl (pHwere stored at 2788C until used. V. 8.0), 5 mg proteinase K (Sigma) /ml, and 50 mgparahaemolyticus strains ATCC 48256 (tl 1 tdh 2 sarkosyl (Fisher) /ml at 658C for 10 min. Followingtrh 2 ), V. parahaemolyticus NY477 (tl1, tdh1, lysis, 100 ml of ice cold 3 M NaOAc (pH 4.6) weretrh2), V. parahaemolyticus AQ4037 (tl1, tdh2, trh added and the tube was inverted several times to1), and V. parahaemolyticus ATCC 47583 (tl mix. Then an equal volume (800 ml) of phenol–1 tdh 1 trh 1 ) were grown in APW (pH 8.5) until chloroform–isoamyl alcohol (24:24:1, v /v) wasthe OD reached 0.2. The cultures were serially added to the sample and the tube was shaken rapidly450
diluted in APW (pH 8.5) and the number of cells /ml till the sample was cloudy. The sample was cen-

A.K. Bej et al. / Journal of Microbiological Methods 36 (1999) 215 –225 221
trifuged at 10 0003g for 10 min at room tempera- achieved with equimolar quantities of each of theture and the clear supernatant was transferred to a oligonucleotide primers (2.5 mM of each of thenew microcentrifuge tube. Total DNA was precipi- primers), 25 mM MgCl in PCR reaction buffer2
tated with 0.8 ml of ice-cold isopropanol, the tubes (103), and a primer annealing temperature of 558Cinverted several times to mix and centrifuged at (Fig. 1).12 0003g for 10 min. The supernatant was decantedand the pellet washed with 0.5 ml of ice-cold 70% 3.2. Distribution of tl, tdh, and trh genes in V.(v /v) alcohol, centrifuged, and the pellet dried in a parahaemolyticus isolates and correlation with thespeedvac DNA 20 (Savant). The DNA sample was Kanagawa reactionresuspended in 10 ml of TE buffer (pH 8.0) andincubated at 658C for 30 min before being subjected The results from the multiplex PCR showedto PCR amplification. positive amplification of the tl gene segment in all
111 (100%) V. parahaemolyticus isolates whereas,2.9. Sensitivity of the multiplex PCR amplification genomic DNAs from four V. hollisae, six V. cholerae,detection in seeded oysters eight Vibrio spp., and one Plesiomonas sp. did not
V. parahaemolyticus F113A (tl1 tdh1 trh1) wasgrown exponentially (OD 50.17) and serially600
diluted. The number of cells per ml of culture wasdetermined by viable plate count on Wagatsumablood agar plates. An aliquot (1 ml) of each of theserially diluted V. parahaemolyticus F113A cultures
4 0consisting of 10 to 10 cells was added to 10 g ofindividual oyster tissue homogenates and pre-en-riched at 358C for 6 h. Following pre-enrichment, 0.5ml of each of the samples was subjected to DNApurification by the procedure described above.
2.10. Detection of the PCR amplified DNAs
PCR-amplified DNAs (10-ml aliquot) were sepa-rated in a 1% (w/v) NuSieve 3:1 agarose gel (FMC
24Bioproducts) in which 2310 mg per ml ofethidium bromide were added. Electrophoresis wasperformed using 13 TAE buffer [40 mM Tris?Cl(pH 8.0), 1.18 ml acetic acid, 2 mM Na EDTA per2
liter] (Taniguchi et al., 1985) and a constant voltageof 5 V/cm. Following electrophoretic separation,DNA fragments in the gels were visualized under a
Fig. 1. Agarose gel electrophoresis showing the results from PCRPhotoprep I UV transilluminator (Fotodyne) andamplification of genomic DNA from V. parahaemolyticus F113A.photographed using Type 55 polaroid film.Lane 1, 123-bp DNA ladder (Gibco) as size marker enhanced byPRORFLP (Nashville, TN, USA) computer software; lane 2, multip-lex PCR DNA ProScan amplification using oligonucleotide prim-
3. Results and discussion ers specific for the tl, tdh and the trh genes showing three bandsof DNA with expected molecular weights of 0.450 kbp, 0.269kbp, and 0.5 kbp, respectively; lane 3, PCR amplification using3.1. Optimization of the multiplex PCRoligonucleotide primers specific for the trh gene; lane 4, PCRamplification using oligonucleotide primers specific for the tl
In multiplex PCR, the amplification of all three gene; lane 5, PCR amplification using oligonucleotide primersgenes, tl, tdh, and trh, with equal intensities was specific for the tdh gene.

222 A.K. Bej et al. / Journal of Microbiological Methods 36 (1999) 215 –225
show PCR amplification of this gene suggesting that the isolates were negative for the tl gene but positivethe L-TL and R-TL oligonucleotide primers and the for the tdh and/or the trh gene, suggesting thetl gene are specific for the detection of total V. occurrence of the tdh gene in combination with the tlparahaemolyticus (Tables 1 and 2). However, only gene is more prevalent than with the trh gene60 (54%) of the total 111 V. parahaemolyticus segment. Also, the majority of clinical isolates wereisolates showed positive PCR amplification of the positive for the tdh gene segment, but none weretdh gene segment, and only 43 (38.73%) isolates positive only for the trh gene and negative for theshowed amplification of the trh gene segment, tdh gene. Therefore, the role of the trh gene insuggesting that not all isolates tested in this study virulence to humans needs to be investigated.were b-hemolytic. Also, 42 (37.8%) isolates showed Tests of these isolates on Wagatsuma blood agarpositive PCR amplification of both tdh and trh genes showed a total of 58 isolates as b-hemolytic, demon-(Tables 1 and 2), suggesting that some V. strating absolute correlation with positive PCR am-parahaemolyticus strains can carry both hemolytic plification of the tdh or the trh gene, or both. Thesegenes. Whether the presence of both tdh and trh results suggest the tdh gene is more prevalent thangenes in these strains increases their virulence to the trh gene in these isolates. Since several of thesehumans is not known. Interestingly, three isolates (V. isolates showed positive amplifications of tdh but notparahaemolyticus SAK5, 553-14, and WR2) showed the trh gene, as well as the reverse pattern, it isno PCR amplification of the tdh gene segment but important that a multiplex PCR be used to achieve awere Kanagawa positive. All three of these isolates rapid but comprehensive analysis of the V.showed negative PCR amplification of both tdh and parahaemolyticus isolates with b-hemolytic prop-trh gene segments. Among these three isolates, two erties.(SAK5 and 553-14) were human-patient isolates andthe third (WR2) was an environmental isolate. In 3.3. Detection of V. parahaemolyticus in seededcontrast, three other isolates (V. parahaemolyticus oyster tissue homogenates33V1C, 30V10A, and 18V10B) showed positiveamplification of the tdh gene segment but produced a PCR amplification targeting the tl, tdh, and trhnegative Kanagawa reaction. Among these latter genes individually or in combination from variousthree isolates, PCR amplification results showed the strains of V. parahaemolyticus showed amplificationspresence of both tdh and trh gene segments in V. of DNA segments of expected molecular weights ofparahaemolyticus 30V10A whereas, 18V10B and 0.45 kbp, 0.269 kbp, and 0.5 kbp, respectively (Fig.33V1C showed positive PCR amplification of only 2). This suggests that the oligonucleotide primersthe tdh gene segment. All three of these V. used for the multiplex PCR-based assay were spe-parahaemolyticus isolates were from seafood sam- cific for their targeted gene segments enablingples. The majority of the V. parahaemolyticus strains detection in shellfish of V. parahaemolyticus throughexhibited a direct correlation between the presence of one or both of these virulence-associated genes (tdhthe thermostable hemolysin genes and positive re- and trh) in combination with the genus-specific tlaction for Kanagawa pehnomenon on Wagatsuma gene.blood agar. However, the reasons why the six strains Detectable levels of PCR-amplified DNA bandsdid not show the same correlation is currently not from all three target genes in agarose gel is evi-understood. denced from the oyster tissue homogenates to which
2The distribution of the tl, tdh, and trh gene 10 cells of V. parahaemolyticus were added (Fig. 3).showed the majority (45%) of the 111 isolates were In addition, positive PCR amplification of the trh andpositive for the tl gene but negative for both tdh and the tl gene segments was detected in oyster tissue
1trh genes. Forty-two (37.8%) isolates were positive homogenates in which 10 V. parahaemolyticus cellsfor all three genes (Tables 1 and 2). Also, 16.2% of were added (Fig 3). The entire procedure includingthe isolates were positive for the tl and the tdh genes, the purification of DNA from oyster tissue homoge-but not the trh gene. Only one isolate was positive nate, PCR amplification and detection of the am-for tl and trh genes, but not the tdh gene. None of plified DNA can be finished within 8 h of the
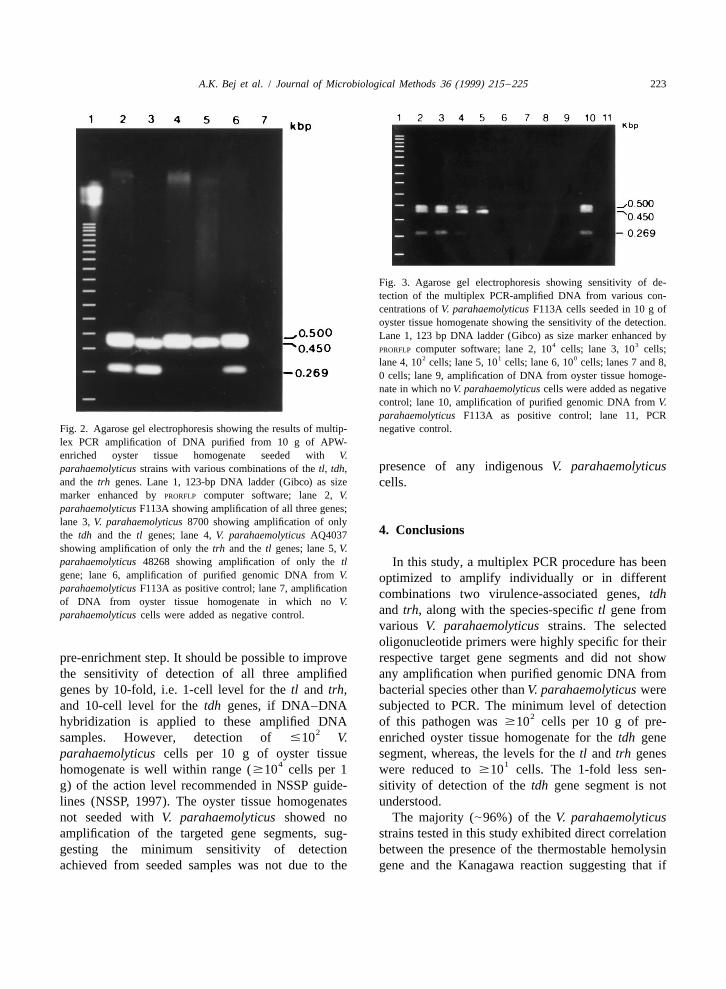
A.K. Bej et al. / Journal of Microbiological Methods 36 (1999) 215 –225 223
Fig. 3. Agarose gel electrophoresis showing sensitivity of de-tection of the multiplex PCR-amplified DNA from various con-centrations of V. parahaemolyticus F113A cells seeded in 10 g ofoyster tissue homogenate showing the sensitivity of the detection.Lane 1, 123 bp DNA ladder (Gibco) as size marker enhanced by
4 3PRORFLP computer software; lane 2, 10 cells; lane 3, 10 cells;
2 1 0lane 4, 10 cells; lane 5, 10 cells; lane 6, 10 cells; lanes 7 and 8,0 cells; lane 9, amplification of DNA from oyster tissue homoge-nate in which no V. parahaemolyticus cells were added as negativecontrol; lane 10, amplification of purified genomic DNA from V.parahaemolyticus F113A as positive control; lane 11, PCR
Fig. 2. Agarose gel electrophoresis showing the results of multip- negative control.lex PCR amplification of DNA purified from 10 g of APW-enriched oyster tissue homogenate seeded with V.
presence of any indigenous V. parahaemolyticusparahaemolyticus strains with various combinations of the tl, tdh,and the trh genes. Lane 1, 123-bp DNA ladder (Gibco) as size cells.marker enhanced by PRORFLP computer software; lane 2, V.parahaemolyticus F113A showing amplification of all three genes;lane 3, V. parahaemolyticus 8700 showing amplification of only
4. Conclusionsthe tdh and the tl genes; lane 4, V. parahaemolyticus AQ4037showing amplification of only the trh and the tl genes; lane 5, V.parahaemolyticus 48268 showing amplification of only the tl In this study, a multiplex PCR procedure has beengene; lane 6, amplification of purified genomic DNA from V. optimized to amplify individually or in differentparahaemolyticus F113A as positive control; lane 7, amplification combinations two virulence-associated genes, tdhof DNA from oyster tissue homogenate in which no V.
and trh, along with the species-specific tl gene fromparahaemolyticus cells were added as negative control.various V. parahaemolyticus strains. The selectedoligonucleotide primers were highly specific for their
pre-enrichment step. It should be possible to improve respective target gene segments and did not showthe sensitivity of detection of all three amplified any amplification when purified genomic DNA fromgenes by 10-fold, i.e. 1-cell level for the tl and trh, bacterial species other than V. parahaemolyticus wereand 10-cell level for the tdh genes, if DNA–DNA subjected to PCR. The minimum level of detection
2hybridization is applied to these amplified DNA of this pathogen was $10 cells per 10 g of pre-2samples. However, detection of #10 V. enriched oyster tissue homogenate for the tdh gene
parahaemolyticus cells per 10 g of oyster tissue segment, whereas, the levels for the tl and trh genes4 1homogenate is well within range ($10 cells per 1 were reduced to $10 cells. The 1-fold less sen-
g) of the action level recommended in NSSP guide- sitivity of detection of the tdh gene segment is notlines (NSSP, 1997). The oyster tissue homogenates understood.not seeded with V. parahaemolyticus showed no The majority (|96%) of the V. parahaemolyticusamplification of the targeted gene segments, sug- strains tested in this study exhibited direct correlationgesting the minimum sensitivity of detection between the presence of the thermostable hemolysinachieved from seeded samples was not due to the gene and the Kanagawa reaction suggesting that if

224 A.K. Bej et al. / Journal of Microbiological Methods 36 (1999) 215 –225
necessary, both tests can be performed simultaneous- the potential to reduce V. parahaemolyticus-associ-ly to confirm the presence of potentially virulent ated illnesses in humans consuming seafood.strains of V. parahaemolyticus in shellfish. In previ-ously reported studies, the presence of the Kanagawaphenomenon has been directly correlated with the
Acknowledgementspositive detection of urease by urea hydrolysis in thispathogen (Kaysner et al., 1994; Okuda et al., 1997a
This research was supported by the Nationaland b; Ghosh and Sehgal, 1998; Iida et al., 1998).
Oceanic and Atmospheric Administration, the Mis-However, all V. parahaemolyticus strains isolated
sissippi Alabama Sea Grant Consortium and thefrom infected human patients from recent outbreaks
University of Alabama at Birmingham (Grant no.(Okuda et al., 1997a and b; Morbidity and Mortality
R-LR 26). We thank A. DePaola for valuable sug-Weekly Report, 1998), exhibited negative reaction
gestions, and L. Beasely for technical assistance.for urease, although positive for the tdh or the trhgenes. Therefore, the test for urease may not besuitable for correlating the presence of the virulencestrains of this pathogen (Okuda et al., 1997a and b). References
PCR amplification of the tl gene segment occurredin all 111 V. haemolyticus strains tested in this study. APHA, 1970. Recommended procedures for the examination of
sea water and shellfish, 4th edition, American Public HealthThis suggests the tl gene is V. parahaemolyticus-Association, New York, NY.specific, and can be used as a genus-specific marker
Atlas, R.M., 1993. Handbook of Microbiological Media, CRCfor PCR amplification detection of this pathogen. Press, Boca Raton, FL.Since some V. parahaemolyticus strains are trh2 but Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Sideman,tdh1 and others the reverse, targeting these two J., Smith, J., Struhl, K., 1987. Current Protocols in Molecular
Biology, Wiley, New York.genes simultaneously in a single PCR reactionDePaola, A., Hopkins, L.H., Peeler, J.T., Wentz, B., McPhearson,should provide comprehensive information about the
R.M., 1990. Incidence of Vibrio parahaemolyticus in USoccurrence and distribution of the virulence-associ- coastal waters and oysters. Appl. Environ. Microbiol. 56,ated genes in this pathogen. The significance of the 2299–2302.presence of the trh gene in a limited number of the V. Gannon, V.P., King, R., Kim, J., Thomas, E., 1992. Rapid and
sensitive method for detection of shiga-like toxin-producingparahaemolyticus isolates as compared to the tdhEscherichia coli in ground beef using the polymerase chaingene is currently not perceived.reaction reaction. Appl. Environ. Microbiol. 58, 3809–3815.
This multiplex PCR method, which targets three Ghosh, A.R., Sehgal, S.C., 1998. Detection of tdh and trh genes ingene segments simultaneously and enables detection a urea-hydrolysing environmental isolate of Vibrioof total and hemolysin-producing V. parahaemolyticus from the Andamans. J. Diarrhoeal Dis. Res.
16, 87–90.parahaemolyticus, is less time-consuming and moreHonda, T., Chearskul, S., Takeda, Y., Miwatani, T., 1980. Immu-cost-effective than the conventional PCR approach,
nological methods for the detection of Kanagawa phenomenonin which each of the three target genes is amplified of Vibrio parahaemolyticus. J. Clin. Microbiol. 11, 600–603.in three separate PCR reactions. The multiplex PCR- Honda, T., Abad-Lapuebla, M.A., Ni, Y., Yamamoto, K.,based detection described in this report could be Miwatani, T., 1991. Characterization of a new thermostable
direct haemolysin produced by a Kanagawa-phenomenon-nega-completed within 8 h of pre-enrichment with the1 2 tive clinical isolate of Vibrio parahaemolyticus. J. Gen. Mi-sensitivity of detection of 10 –10 cells per 10 g of
crobiol. 137, 253–259.shellstock. This result corresponded with the de- Honda, T., Iida, T., 1993. The pathogenicity of Vibriotection of the virulent strains of V. parahaemolyticus parahaemolyticus and the role of the thermostable directcarrying the tdh gene in marine fish (Karunasagar et haemolysin and related haemolysins. Rev. Med. Microbiol. 4,
106–113.al., 1996). The availability of a rapid and reliableIida, T., Park, K.S., Suthienkul, O., Kozawa, J., Yamaichi, Y.,multiplex PCR that comprehensively detects V.
Yamamoto, K., Honda, T., 1998. Close proximity of he tdh,parahaemolyticus in shellfish by simultaneously trh, and ure genes on chromosome of Vibrio parahaemolyticus.amplifying the genus-specific tl gene and hemolysin- Microbiology 144, 2517–2523.producing tdh and trh genes in a single reaction has Janda, J.M., Powers, C., Bryant, R.G., Abbot, S., 1988. Current

A.K. Bej et al. / Journal of Microbiological Methods 36 (1999) 215 –225 225
perspectives on the epidemiology and pathogenesis of clinical- phenomenon, the virulence-associated phenotype, of Vibrioly significant Vibrio spp. Clin. Microbiol. Rev. 1, 245–267. parahaemolyticus depends on a particular single base change in
Joseph, S.W., Colwell, R.R., Kaper, J.B., 1983. Vibrio the promoter of the thermostable direct hemolysin gene. Mol.parahaemolyticus and related halophilic vibrios. CRC Crit. Microbiol. 30, 499–511.Rev. Microbiol. 10, 77–124. Okuda, J., Ishibashi, M., Abbott, S.L., Janda, J.M., Nishibuchi,
Karunasagar, I., Sugumar, G., Karunasagar, I., Reilly, P.J., 1996. M., 1997a. Analysis of the thermostable direct hemolysin (tdh)Rapid polymerase chain reaction method for detection of gene and the tdh related hemolysin (trh) genes in urease-Kanagawa positive Vibrio parahaemolyticus in seafoods. Int. J. positive strains of Vibrio parahaemolyticus isolated on theFood Microbiol. 31, 317–323. West Coast of the United States. J. Clin. Microbiol. 35, 1965–
Kaysner, C.A., Abeyta, C., Trost, P.A., Wetherington, J.H., 1971.Jinneman, K.C., Hill, W.E., Wekell, M.M., 1994. Urea hy- Okuda, J., Ishibahi, M., Hayakawa, E., Nishino, T., Takeda, Y.,drolysis can predict the potential pathogenicity of Vibrio Mukhopadhyay, A.K., Garg, S., Bhattacharya, S.K., Nair, G.B.,parahaemolyticus strains isolated in the Pacific Northwest. Nishibuchi, M., 1997b. Emergence of a unique O3:K6 clone ofAppl. Environ. Microbiol. 60, 3020–3022. Vibrio parahaemolyticus in Calcutta, India, and isolation of
Kaysner, C.A., Tamplin, M.L., Twedt, R.M., 1992. Vibrio. In: strains from the same cloncal group Southeast Asian travellersVanderzant, C., Splittstoesser, D.F. (Eds.), Compendium of arriving in Japan. J. Clin. Microbiol. 35, 3150–3155.Methods for the Microbiological Examination of Foods, Amer- Shirai, H., Ito, H., Hirramaya, T., Nakamoto, T., Nakabayashi, N.,ican Public Health Association, Washington, DC, pp. 451–473. Kumagai, K., Takeda, Y., Nishibuchi, M., 1990. Molecular
Kishishita, M., Matsouka, N., Kumagai, K., Yamasaki, S., Takeda,epidemiologic evidence for association of thermostable direct
Y., Nishibuchi, M., 1992. Sequence variation in the thermo-hemolysin (TDH) and TDH-related hemolysin of Vibrio
stable direct hemolysin-related hemolysin (trh) gene in Vibrioparahaemolyticus with gastroenteritis. Infec. Immun. 58,
parahaemolyticus. Appl. Environ. Microbiol. 58, 2449–2457.3568–3573.
Lee, T., Pan, S.F., 1993. Rapid and specific detection of theSuggs, S.B., Hirose, T., Miyake, T., Kawashima, E.H., Johnson,
thermostable direct hemolysin gene in Vibrio parahaemolyticusM.J., Itakara, K., Wallace, R.B., 1981. Use of syntheticby polymerase chain reaction. J. Gen. Microbiol. 139, 3225–oligodeoxyribonucleotides for the isolation of specific clone3231.DNA sequences. ICN–UCLA Symp. Mol. Cell. Biol. 23,Lee, C.Y., Pan, S.F., Chen, C.H., 1995. Sequence of a cloned683–693.pR72H fragment and its use for detection of Vibrio
Suthienkul, O., Iida, T., Park, K.S., Ishibashi, M., Supavej, S.,parahaemolyticus in shellfish with the PCR. Appl. Environ.Yamamoto, K., Honda, T., 1996. Restriction fragment lengthMicrobiol. 61, 1311–1317.polymorphism of the tdh and trh genes in clinical VibrioMiyamoto, Y., Kato, T., Obra, S., Akiyama, S., Takizawa, K.,parahaemolyticus strains. J. Clin. Microbiol. 34, 1293–1295.Yamai, S., 1969. In vitro characteristic of Vibrio
Tada, J., Ohashi, T., Nishimura, N., Shirasaki, Y., Ozaki, H.,parahaemolyticus: its close correlation with human patho-Fukushima, S., Takano, J., Nishibuchi, M., Takeda, Y., 1992.genicity. J. Bacteriol. 100, 1147–1149.Detection of the thermostable direct hemolysin gene (tdh) and1998. Outbreak of Vibrio parahaemolyticus infections associatedthe thermostable direct hemolysin-related hemolysin gene (trh)with eating raw oysters – Pacific Northwest, 1997. Morb.of Vibrio parahaemolyticus by polymerase chain reaction. Mol.Mortal. Weekly Rep. 47, 12–16.Cell. Prob. 6, 477–487.NSSP: National Shellfish Sanitation Program, 1997. Sanitation of
Takeda, Y., 1982. Thermostable direct hemolysin of Vibriothe harvesting, processing and distribution of shellfish. In:parahaemolyticus. Pharmocol. Ther. 19, 123–146.Manual of Operations Part II, U.S. Department of Health and
Taniguchi, H., Ohta, H., Ogawa, M., Mizuguchi, Y., 1985. CloningHuman Services, Food and Drug Administration, Washington,DC. and expression of Escherichia coli of Vibrio parahaemolyticus
Nishibuchi, M., Kaper, J.B., 1985. Nucleotide sequence of the thermostable direct hemolysin and thermolabile hemolysinthermostable direct hemolysin gene of Vibrio genes. J. Bacteriol. 162, 510–515.parahaemolyticus. J. Bacteriol. 162, 558–564. Taniguchi, H., Hirano, H., Kubomura, S., Higashi, K., Mizuguchi,
Nishibuchi, M., Ishibashi, M., Takeda, Y., Kaper, J.B., 1985. Y., 1986. Comparison of the nucleotide sequences of the genesDetection of the thermostable direct hemolysin gene and for the thermolabile hemolysin from Vibrio parahaemolyticus.related DNA sequences in Vibrio parahaemolyticus and other Microb. Pathogen. 1, 425–432.Vibrio species by the DNA colony hybridization test. Infect. Wagatsuma, S., 1968. A medium for the test of the hemolyticImmun. 49, 481–486. activity of Vibrio parahaemolyticus. Media Circle 13, 159.
Nishibuchi, M., Hill, W.E., Zon, G., Payne, W.L., Kaper, J.B., Yamamoto, K., Honda, T., Miwatani, T., Tamatsukuri, S., Shibata,1986. Synthetic oligodeoxyribonucleotide probes to detect S., 1992. Enzyme-labeled oligonucleotide probe for the de-Kanagawa phenomenon-positive Vibrio parahaemolyticus. J. tection of the genes for thermostable direct hemolysin (TDH)Clin. Microbiol. 23, 1091–1095. and TDH-related hemolysin (TRH) of Vibrio
Okuda, J., Nishibuchi, M., 1998. Manifestation of the Kanagawa parahaemolyticus. Can. J. Microbiol. 38, 410–416.




















