
Rapid modulation of TRH and TRH-like peptide release in rat brain, pancreas, and testis by a GSK-3β...
11
Peptides 31 (2010) 1083–1093 Contents lists available at ScienceDirect Peptides journal homepage: www.elsevier.com/locate/peptides Rapid modulation of TRH and TRH-like peptide release in rat brain, pancreas, and testis by a GSK-3 inhibitor Albert Eugene Pekary a,c,d,∗ , Schetema A. Stevens a , James D. Blood a , Albert Sattin b,e,f a Research Services, VA Greater Los Angeles Healthcare System, United States b Psychiatry Services, VA Greater Los Angeles Healthcare System, United States c Center for Ulcer Research and Education, VA Greater Los Angeles Healthcare System, United States d Department of Medicine, University of California, Los Angeles, CA 90073, United States e Department of Psychiatry and Biobehavioral Sciences, University of California, Los Angeles, CA 90073, United States f Brain Research Institute, University of California, Los Angeles, CA 90073, United States article info Article history: Received 6 March 2010 Accepted 15 March 2010 Available online 23 March 2010 Keywords: GSK-3 Thyrotropin-releasing hormone Limbic system Testis HPLC Radioimmunoassay abstract Antidepressants have been shown to be neuroprotective and able to reverse damage to glia and neurons. Thyrotropin-releasing hormone (TRH) is an endogenous antidepressant-like neuropeptide that reduces the expression of glycogen synthase kinase-3 (GSK-3), an enzyme that hyperphosphorylates tau and is implicated in bipolar disorder, diabetes and Alzheimer’s disease. In order to understand the potential role of GSK-3 in the modulation of depression by TRH and TRH-like peptides and the therapeutic potential of GSK-3 inhibitors for neuropsychiatric and metabolic diseases, young adult male Sprague–Dawley (SD) rats were (a) injected ip with 1.8 mg/kg of GSK-3 inhibitor VIII (GSKI) and sacrificed 0, 2, 4, 6, and 8 h later or (b) injected with 0, 0.018, 0.18 or 1.8 mg/kg GSKI and bled 4 h later. Levels of TRH and TRH-like peptides were measured in various brain regions involved in mood regulation, pancreas and reproductive tissues. Large, 3–15-fold, increases of TRH and TRH-like peptide levels in cerebellum, for example, as well as other brain regions were noted at 2 and 4 h. In contrast, a nearly complete loss of TRH and TRH-like peptides from testis within 2 h and pancreas by 4 h following GSKI injection was observed. We have previously reported similar acute effects of corticosterone in brain and peripheral tissues. Incubation of a decapsulated rat testis with either GSKI or corticosterone accelerated release of TRH, and TRH-like peptides. Glucocorticoids, via inhibition of GSK3- activity, may thus be involved in the inhibition of TRH and TRH-like peptide release in brain, thereby contributing to the depressogenic effect of this class of steroids. Corticosterone-induced acceleration of release of these peptides from testis may contribute to the decline in reproductive function and redirection of energy needed during life-threatening emergencies. These contrasting effects of glucocorticoid on peptide release appear to be mediated by GSK-3. Published by Elsevier Inc. 1. Introduction Glycogen synthase kinase-3 (GSK-3) phosphorylates a num- ber of key regulatory proteins that have been implicated in diabetes, stroke, Alzheimer’s disease, tumor progression, major depression, bipolar disorder, and microtubule assembly in neu- rons [3–5,12,14,21,26,29,35,48,58,59,62,63]. We and others have previously reported that TRH (pGlu-His-Pro-NH 2 ) and TRH-like peptides (pGlu-X-Pro-NH 2 or X-TRH, where “X” can be any ∗ Corresponding author at: VA Greater Los Angeles Healthcare System, Bldg. 114, Rm. 229, 11301 Wilshire Blvd., Los Angeles, CA 90073, United States. Tel.: +1 310 268 4430; fax: +1 310 441 1702. E-mail address: [email protected] (A.E. Pekary). amino acid residue), have neuroprotective, antidepressant, analep- tic, and anti-amnesic properties [16,23,24,29,37,42,44–46,61]. In the case of TRH, this is due, at least in part, to potent inhibition of GSK-3 gene expression [29]. TRH and TRH-like peptide levels in brain and peripheral tissues can be profoundly modulated by antidepressants, thyroid and steroid hormones, proinflammatory cytokines, neuropharmacologic agents, psy- chostimulants, electroconvulsive shock, and the photoperiod [16,18,37,40,42,44–47]. As part of a systematic exploration of the involvement of this family of endogenous neuropeptides in both the pathogenesis and treatment of psychiatric diseases, we hypothesized that (a) many of the extrapituitary effects of TRH and TRH-like peptides involve the modulation of GSK-3 activity, and (b) the well-established decline in levels of TRH and responsiveness to TRH in brain and periph- eral tissues with aging [1,17,36] contributes to age-related GSK-3 0196-9781/$ – see front matter. Published by Elsevier Inc. doi:10.1016/j.peptides.2010.03.020
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Rapid modulation of TRH and TRH-like peptide release in rat brain, pancreas, and testis by a GSK-3β...

Rt
Aa
b
c
d
e
f
a
ARAA
KGTLTHR
1
bddrpp
RT
0d
Peptides 31 (2010) 1083–1093
Contents lists available at ScienceDirect
Peptides
journa l homepage: www.e lsev ier .com/ locate /pept ides
apid modulation of TRH and TRH-like peptide release in rat brain, pancreas, andestis by a GSK-3� inhibitor
lbert Eugene Pekarya,c,d,∗, Schetema A. Stevensa, James D. Blooda, Albert Sattinb,e,f
Research Services, VA Greater Los Angeles Healthcare System, United StatesPsychiatry Services, VA Greater Los Angeles Healthcare System, United StatesCenter for Ulcer Research and Education, VA Greater Los Angeles Healthcare System, United StatesDepartment of Medicine, University of California, Los Angeles, CA 90073, United StatesDepartment of Psychiatry and Biobehavioral Sciences, University of California, Los Angeles, CA 90073, United StatesBrain Research Institute, University of California, Los Angeles, CA 90073, United States
r t i c l e i n f o
rticle history:eceived 6 March 2010ccepted 15 March 2010vailable online 23 March 2010
eywords:SK-3�hyrotropin-releasing hormoneimbic systemestisPLCadioimmunoassay
a b s t r a c t
Antidepressants have been shown to be neuroprotective and able to reverse damage to glia and neurons.Thyrotropin-releasing hormone (TRH) is an endogenous antidepressant-like neuropeptide that reducesthe expression of glycogen synthase kinase-3� (GSK-3�), an enzyme that hyperphosphorylates tau and isimplicated in bipolar disorder, diabetes and Alzheimer’s disease. In order to understand the potential roleof GSK-3� in the modulation of depression by TRH and TRH-like peptides and the therapeutic potentialof GSK-3� inhibitors for neuropsychiatric and metabolic diseases, young adult male Sprague–Dawley(SD) rats were (a) injected ip with 1.8 mg/kg of GSK-3� inhibitor VIII (GSKI) and sacrificed 0, 2, 4, 6,and 8 h later or (b) injected with 0, 0.018, 0.18 or 1.8 mg/kg GSKI and bled 4 h later. Levels of TRH andTRH-like peptides were measured in various brain regions involved in mood regulation, pancreas andreproductive tissues. Large, 3–15-fold, increases of TRH and TRH-like peptide levels in cerebellum, forexample, as well as other brain regions were noted at 2 and 4 h. In contrast, a nearly complete lossof TRH and TRH-like peptides from testis within 2 h and pancreas by 4 h following GSKI injection wasobserved. We have previously reported similar acute effects of corticosterone in brain and peripheraltissues. Incubation of a decapsulated rat testis with either GSKI or corticosterone accelerated release of
TRH, and TRH-like peptides. Glucocorticoids, via inhibition of GSK3-� activity, may thus be involved inthe inhibition of TRH and TRH-like peptide release in brain, thereby contributing to the depressogeniceffect of this class of steroids. Corticosterone-induced acceleration of release of these peptides fromtestis may contribute to the decline in reproductive function and redirection of energy needed duringlife-threatening emergencies. These contrasting effects of glucocorticoid on peptide release appear to be mediated by GSK-3�.. Introduction
Glycogen synthase kinase-3� (GSK-3�) phosphorylates a num-er of key regulatory proteins that have been implicated in
iabetes, stroke, Alzheimer’s disease, tumor progression, majorepression, bipolar disorder, and microtubule assembly in neu-ons [3–5,12,14,21,26,29,35,48,58,59,62,63]. We and others havereviously reported that TRH (pGlu-His-Pro-NH2) and TRH-likeeptides (pGlu-X-Pro-NH2 or X-TRH, where “X” can be any∗ Corresponding author at: VA Greater Los Angeles Healthcare System, Bldg. 114,m. 229, 11301 Wilshire Blvd., Los Angeles, CA 90073, United States.el.: +1 310 268 4430; fax: +1 310 441 1702.
E-mail address: [email protected] (A.E. Pekary).
196-9781/$ – see front matter. Published by Elsevier Inc.oi:10.1016/j.peptides.2010.03.020
Published by Elsevier Inc.
amino acid residue), have neuroprotective, antidepressant, analep-tic, and anti-amnesic properties [16,23,24,29,37,42,44–46,61].In the case of TRH, this is due, at least in part, to potentinhibition of GSK-3� gene expression [29]. TRH and TRH-likepeptide levels in brain and peripheral tissues can be profoundlymodulated by antidepressants, thyroid and steroid hormones,proinflammatory cytokines, neuropharmacologic agents, psy-chostimulants, electroconvulsive shock, and the photoperiod[16,18,37,40,42,44–47].
As part of a systematic exploration of the involvement of thisfamily of endogenous neuropeptides in both the pathogenesis and
treatment of psychiatric diseases, we hypothesized that (a) many ofthe extrapituitary effects of TRH and TRH-like peptides involve themodulation of GSK-3� activity, and (b) the well-established declinein levels of TRH and responsiveness to TRH in brain and periph-eral tissues with aging [1,17,36] contributes to age-related GSK-3�
1 tides 3
hr
(ir
2
2
ua#alrbwAtrttmAtsap
2
oVa8idwaf[tG
2
1b4
2
fa((hd
084 A.E. Pekary et al. / Pep
yperactivity and consequent CNS and metabolic pathology andeproductive decline [54,63].
For these reasons we have used a specific GSK-3� inhibitorGSKI) to study both the in vivo and in vitro effects of acute GSK-3�nhibition on TRH and TRH-like peptide levels in, and release from,at brain and peripheral tissues.
. Materials and methods
.1. Animals
Male Sprague–Dawley (SD) rats (Harlan, Indianapolis, IN) weresed for all experiments. These animals were group housed (4nimals per cage), maintained with standard Purina rodent chow5001 and water ad libitum during a standard 1 week initial quar-ntine in a controlled temperature and humidity environment;ights on: 6 am to 6 pm. All animals were weighed on the day ofeceipt and on the morning of each experiment. Initial and finalody weights did not differ between experimental groups. Researchas approved by the VA Greater Los Angeles Healthcare Systemnimal Care and Use Committee and conducted in compliance with
he Animal Welfare Act and the federal statutes and regulationselated to animals and experiments involving animals and adhereso principles stated in the Guide for the Care and use of Labora-ory Animals, NRC Publication, 1996 edition. All efforts have been
ade to minimize the number of animals used and their suffering.ll animals were transferred from the Veterinary Medical Unit to
he laboratory 12 h before the start of experiments to minimize thetress of a novel environment. Decapitation occurred between 9 amnd 11 am to minimize the diurnal variation in TRH and TRH-likeeptide levels [45].
.2. Time-dependent in vivo effects of GSKI
Ten rats received a single 0.5 ml ip injection of 1.0 mg/ml DMSOf the cell permeable and antidepressant-like [15] GSK-3 inhibitorIII (GSKI, IC50 = 104 ± 27 nM, Calbiochem, San Diego, CA), whichcts in an ATP-competitive manner (Ki = 38 nM, [6]), at 0, 2, 4, 6, andh (n = 2 for each time point) before sacrifice. This does not signif-
cantly inhibit cdk2 or cdk5 (IC50 > 100 �M) or 26 other kinasesemonstrating high specificity for GSK3. The mean body weightsere 272 ± 12 g (mean ± SD). Thus 1.8 mg GSKI/kg body weight was
dministered. Uptake values for [11C]GSKI into rat brain, correctedor radioactivity in the vascular compartment, was 0.06% at 30 min57]. Even though this uptake is very poor, the GSKI concentra-ion in brain at this time would be 1.0 �M, 10 times the IC50 forSK-3.
.3. Concentration-dependent in vivo effects of GSKI
Sixteen rats were ip injected with 0.5 ml of 0, 0.01, 0.1 or.0 mg GSKI/ml DMSO (n = 4 for each concentration). The meanody weight was 281 ± 14 g (mean ± SD). All rats were decapitatedh after injection.
.4. Dissection of rat brain, pancreas and reproductive organs
After decapitation, Nucleus accumbens (NA), amygdala (AY),rontal cortex (FCX), cerebellum (CBL), medulla oblongata (MED),
nterior cingulate (ACNG), posterior cingulate (PCNG), striatumSTR), pyriform cortex (PYR), hippocampus (HC), entorhinal cortexENT), pancreas (PAN), prostate, epididymis, and testes (T) wereand dissected, weighed rapidly, and then extracted as previouslyescribed [37,42,44–47].1 (2010) 1083–1093
2.5. In vitro effects of GSKI and corticosterone
GSKI is extremely photosensitive requiring that all possiblemeasures be used to protect it from light exposure during the invitro experiments described below. Lights were turned out andwindow shades were closed. Bacitracin from Bacillus licheniformis(3.0 mg, Sigma) and 1.0 mg of a potent TRH-degrading ectoenzymeinhibitor (pGlu-Asn-Pro-d-Tyr-d-Trp-NH2, Phoenix Pharmaceuti-cals, Belmont, CA [52]) were dissolved in 40 ml of Ham’s F-10medium with HEPES buffer (MP Biomedicals, Irvine, CA) and equi-librated with 95% O2–5% CO2 for 15 min at room temperature.Maximum dissolved O2 of 31.2 mg/L was reached at 10 min as mea-sured with a Fisher Brand Portable Dissolved Oxygen Meter (FisherScientific, Pittsburg, PA). Three 20 ml glass scintillation vials werelabeled as A, B, and C. DMSO (150 �l) was added to vial A. Corti-costerone hemisuccinate (150 �l of 10 mg/ml DMSO) was addedto vial B (224 �M corticosterone final concentration) and 150 �l of1.0 mg GSKI/ml DMSO was added to vial C wrapped in aluminumfoil (37 �M GSKI final concentration). Thirteen ml of the Ham’sF-10 mixture were then added to each vial. Contents of vials Awas distributed equally between four 13 mm × 100 mm glass testtubes, 3.0 ml/vial; likewise for vials B and C. The “C” test tubes werewrapped in black paper. 95% O2–5% CO2 was bubbled into eachvial continuously to maintain the oxygenation and pH balance ofthe incubation mixture using a Thermolyne Dri-Bath evaporator(Barnstead, Dubuque, IA) set at 35 ◦C. One 350 g male SD rat wasthen decapitated. Each decapsulated testis was divided into 6 equalpieces. One testis fragment was transferred into each of the 12 glasstest tubes. A 100 �l aliquot from each of the four replicate glasstubes containing medium from Vial A were pooled together; simi-larly for tubes containing medium from Vial B and C. These pooledaliquots was centrifuged at 100 × g for 5 min at 4 ◦C to remove Ley-dig cells dislodged from tubules by the continuous oxygenationat 0.5, 1.0, 1.5, 2.0, and 2.5 h after the addition of the testes. Thesupernatant was decanted into a 12 mm × 75 mm glass test tubecontaining 2.0 ml of methanol. All tubes were then centrifuged at1000 × g, the supernatant decanted and dried on a heater block witha fan blowing air across the top of the tubes to accelerate drying.The dried residues were reconstituted with 0.5 ml of 0.02% NaN3prior to TRH RIA in duplicate.
2.6. HPLC and RIA procedures, HPLC peak identification andquantitation
HPLC and RIA procedures, peak identification, and quantitationby co-chromatography with synthetic TRH and TRH-like peptides,relative potency analysis of multiple antibodies to TRH and TRH-like peptides, mass spectrometry and resolution of overlappingpeaks by least squares fitting of a 2-Gaussian statistical model havebeen previously reported in detail [39,43].
Briefly, after boiling, tissues were dried, re-extracted withmethanol, dried and defatted by water–ethyl ether partitioning.Dried samples were dissolved in 0.1% trifluroacetic acid (TFA), andloaded onto reverse phase C18 Sep-Pak cartridges (Water, Milford,MA). TRH and TRH-like peptides were eluted with 30% methanol.Dried peptides were again dissolved in TFA, filtered and then frac-tionated by HPLC using a 4.6 mm × 150 mm Econosphere, 3 �mC18 reverse phase column (Alltech Associates, Deerfield, IL) and a0.33% min−1 gradient of acetonitrile. The 0.5 ml fractions collectedwere dried completely and reconstituted with 0.15 ml of 0.02%NaN3 just before RIA.
The antiserum used (8B9) cross-reacts with TRH and eight TRH-like peptides with a relative potency of displacement ranging from2.31 (Lys-TRH) to 0.288 (Ser-TRH) relative to Tyr-TRH (Table 2,[42]). Two of the regularly observed peaks (2a and 2b) consist ofa mixture of unidentified TRH-like peptides. Of the seven observed
tides 3
ptrrvR
awsfwTw
2h
GrLs
Fa
A.E. Pekary et al. / Pep
eptides three have so far been confirmed by mass spectrome-ry: TRH, Glu-TRH and Tyr-TRH [37] and mass spectrometry of theemaining four is planned. Tissue samples from the GSKI-treatedats dissected at each time point were pooled prior to HPLC to pro-ide the minimum amount of immunoreactivity needed for reliableIA measurements.
The mean recovery of TRH and TRH-like peptide immunore-ctivity from all tissues studied was 84 ± 15% (mean ± SD). Theithin-assay and between-assay coefficient of variation for mea-
uring 333 pg/ml TRH was 4.8% and 16.9%, respectively. All HPLCractions obtained from a given brain region or peripheral tissueere analyzed in the same RIA. The minimum detectable dose for
RH was 5 pg/ml. The maximum specific binding of [125I]TRH (Bo/T)as 25%.
.7. Glucose, rat insulin, rat leptin, corticosterone, thyroidormone, and testosterone assays
Serum glucose was measured with the OneTouch Ultra Bloodlucose Monitoring System, Life Scan, Milpitas, CA. Rat insulin and
at leptin were quantitated with the Sensitive Rat Insulin and Rateptin RIA Kits (Millipore, Billerica, MA). Corticosterone was mea-ured by MP Biomedical RIA (Irvine, CA) and serum free T4, total
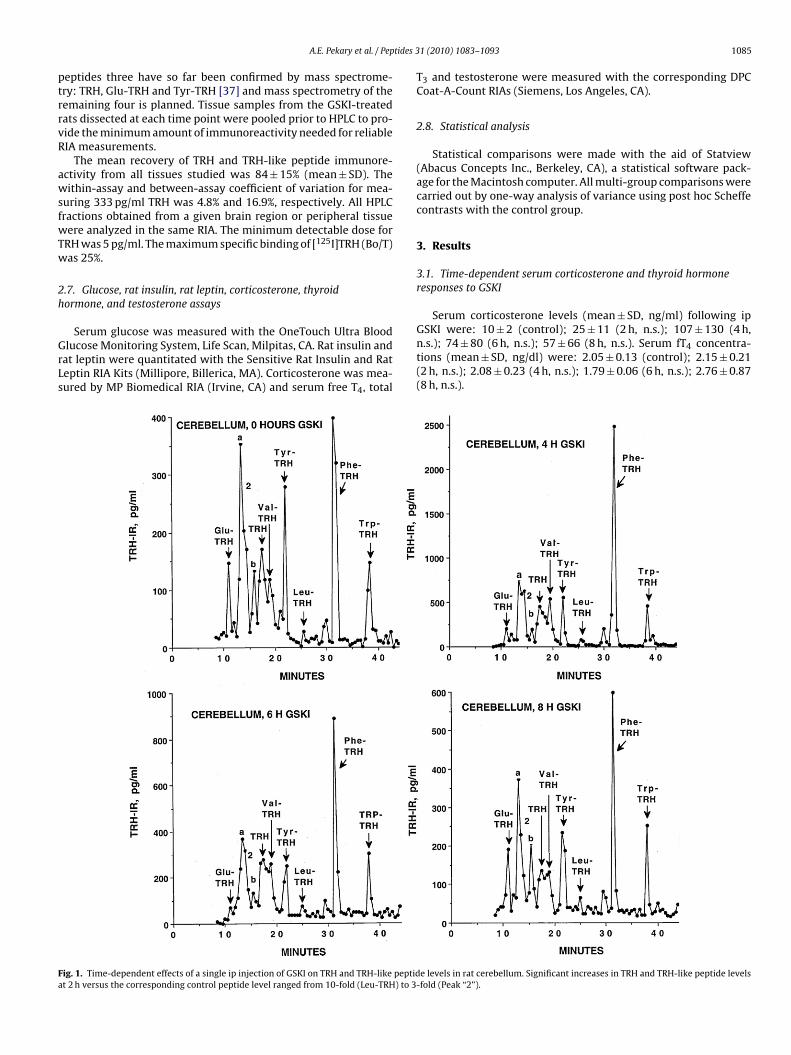
ig. 1. Time-dependent effects of a single ip injection of GSKI on TRH and TRH-like peptidt 2 h versus the corresponding control peptide level ranged from 10-fold (Leu-TRH) to 3
1 (2010) 1083–1093 1085
T3 and testosterone were measured with the corresponding DPCCoat-A-Count RIAs (Siemens, Los Angeles, CA).
2.8. Statistical analysis
Statistical comparisons were made with the aid of Statview(Abacus Concepts Inc., Berkeley, CA), a statistical software pack-age for the Macintosh computer. All multi-group comparisons werecarried out by one-way analysis of variance using post hoc Scheffecontrasts with the control group.
3. Results
3.1. Time-dependent serum corticosterone and thyroid hormoneresponses to GSKI
Serum corticosterone levels (mean ± SD, ng/ml) following ip
GSKI were: 10 ± 2 (control); 25 ± 11 (2 h, n.s.); 107 ± 130 (4 h,n.s.); 74 ± 80 (6 h, n.s.); 57 ± 66 (8 h, n.s.). Serum fT4 concentra-tions (mean ± SD, ng/dl) were: 2.05 ± 0.13 (control); 2.15 ± 0.21(2 h, n.s.); 2.08 ± 0.23 (4 h, n.s.); 1.79 ± 0.06 (6 h, n.s.); 2.76 ± 0.87(8 h, n.s.).e levels in rat cerebellum. Significant increases in TRH and TRH-like peptide levels-fold (Peak “2”).
1 tides 3
3c
w(i(
TA
R
086 A.E. Pekary et al. / Pep
.2. Concentration-dependent serum glucose, insulin, leptin,orticosterone and thyroid hormone responses to GSKI
Serum glucose levels (mean ± SD, mg/dl) following ip GSKIere: 138 ± 39 (0 mg/kg); 201 ± 9 (0.018 mg/kg, p < 0.05); 101 ± 15
0.18 mg/kg, p < 0.05) and 146 ± 13 (1.8 mg/kg, n.s.). Serum ratnsulin concentrations (mean ± SD, ng/ml) were: 0.078 ± 0.0050 mg/kg); 0.228 ± 0.120 (0.018 mg/kg, p < 0.05), 0.095 ± 0.030
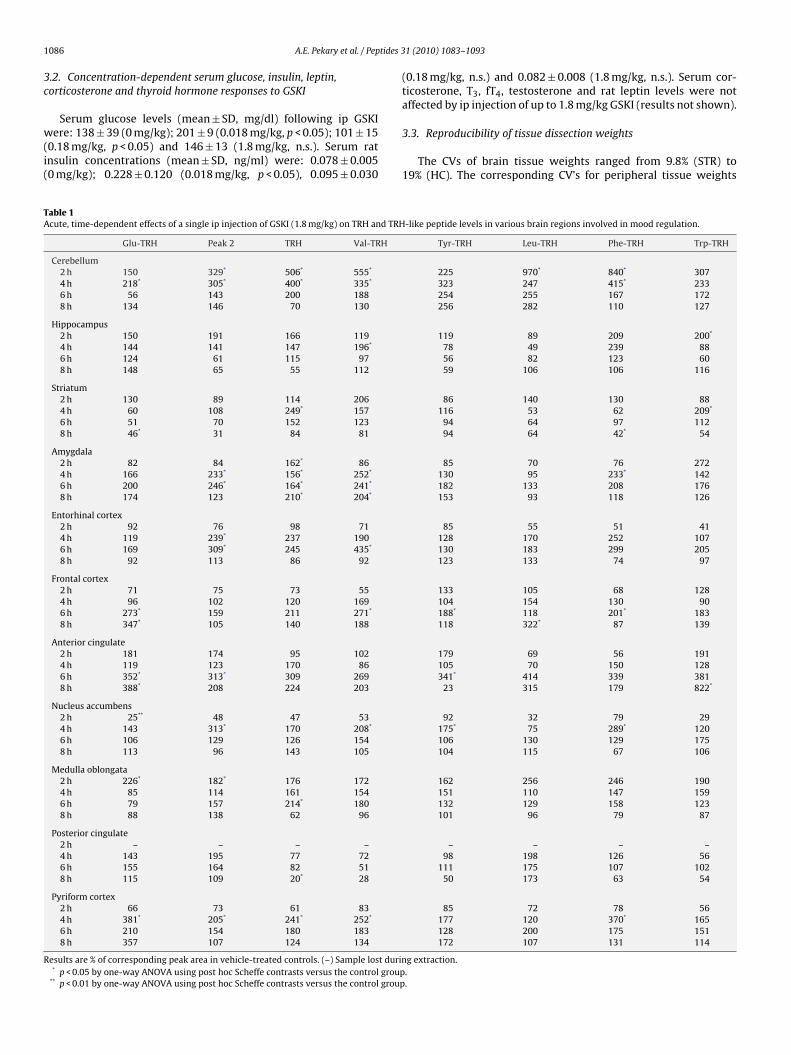
able 1cute, time-dependent effects of a single ip injection of GSKI (1.8 mg/kg) on TRH and TRH
Glu-TRH Peak 2 TRH Val-TRH
Cerebellum2 h 150 329* 506* 555*
4 h 218* 305* 400* 335*
6 h 56 143 200 1888 h 134 146 70 130
Hippocampus2 h 150 191 166 1194 h 144 141 147 196*
6 h 124 61 115 978 h 148 65 55 112
Striatum2 h 130 89 114 2064 h 60 108 249* 1576 h 51 70 152 1238 h 46* 31 84 81
Amygdala2 h 82 84 162* 864 h 166 233* 156* 252*
6 h 200 246* 164* 241*
8 h 174 123 210* 204*
Entorhinal cortex2 h 92 76 98 714 h 119 239* 237 1906 h 169 309* 245 435*
8 h 92 113 86 92
Frontal cortex2 h 71 75 73 554 h 96 102 120 1696 h 273* 159 211 271*
8 h 347* 105 140 188
Anterior cingulate2 h 181 174 95 1024 h 119 123 170 866 h 352* 313* 309 2698 h 388* 208 224 203
Nucleus accumbens2 h 25** 48 47 534 h 143 313* 170 208*
6 h 106 129 126 1548 h 113 96 143 105
Medulla oblongata2 h 226* 182* 176 1724 h 85 114 161 1546 h 79 157 214* 1808 h 88 138 62 96
Posterior cingulate2 h – – – –4 h 143 195 77 726 h 155 164 82 518 h 115 109 20* 28
Pyriform cortex2 h 66 73 61 834 h 381* 205* 241* 252*
6 h 210 154 180 1838 h 357 107 124 134
esults are % of corresponding peak area in vehicle-treated controls. (–) Sample lost durin* p < 0.05 by one-way ANOVA using post hoc Scheffe contrasts versus the control group
** p < 0.01 by one-way ANOVA using post hoc Scheffe contrasts versus the control group
1 (2010) 1083–1093
(0.18 mg/kg, n.s.) and 0.082 ± 0.008 (1.8 mg/kg, n.s.). Serum cor-ticosterone, T3, fT4, testosterone and rat leptin levels were notaffected by ip injection of up to 1.8 mg/kg GSKI (results not shown).
3.3. Reproducibility of tissue dissection weights
The CVs of brain tissue weights ranged from 9.8% (STR) to19% (HC). The corresponding CV’s for peripheral tissue weights
-like peptide levels in various brain regions involved in mood regulation.
Tyr-TRH Leu-TRH Phe-TRH Trp-TRH
225 970* 840* 307323 247 415* 233254 255 167 172256 282 110 127
119 89 209 200*
78 49 239 8856 82 123 6059 106 106 116
86 140 130 88116 53 62 209*
94 64 97 11294 64 42* 54
85 70 76 272130 95 233* 142182 133 208 176153 93 118 126
85 55 51 41128 170 252 107130 183 299 205123 133 74 97
133 105 68 128104 154 130 90188* 118 201* 183118 322* 87 139
179 69 56 191105 70 150 128341* 414 339 381
23 315 179 822*
92 32 79 29175* 75 289* 120106 130 129 175104 115 67 106
162 256 246 190151 110 147 159132 129 158 123101 96 79 87
– – – –98 198 126 56
111 175 107 10250 173 63 54
85 72 78 56177 120 370* 165128 200 175 151172 107 131 114
g extraction...

A.E. Pekary et al. / Peptides 31 (2010) 1083–1093 1087
F pepto
vwl
3
a[TlcTG
3G
0r
ig. 2. Time-dependent effects of a single ip injection of GSKI on TRH and TRH-likebserved for Trp-TRH at 2 h and Val-TRH at 4 h.
aried from 3.6% (epididymis) to 38.0% (prostate). Prostateeights vary markedly in normal, untreated, young adult male
ittermates.
.4. Mean within-group CV for TRH and TRH-like peptide levels
The mean within-group CV’s for TRH and TRH-like peptides,veraged over 4 equally spaced intervals within a 24 h photoperiod45] ranged from 6.3% for TRH levels of AY to 48% for Leu-RH levels of ACNG. These CVs were then used to estimate theevel of significance, by one-way ANOVA, of time-dependent andoncentration-dependent changes in the pooled mean values ofRH and TRH-like peptide levels following a single ip dose ofSKI.
.5. Overview of time-dependent HPLC results in brain after ip
SKIThe response of TRH and TRH-like peptide levels in brain at, 2, 4, 6 and 8 h after a single ip injection of GSKI are summa-ized in Table 1 and Figs. 1 and 2. With 4 exceptions: 75% decrease
ide levels in rat hippocampus. Significant (2-fold) increases in peptide levels were
in Glu-TRH at 2 h in NA; 80% reduction of TRH at 8 h in PCNG;decreases in Glu-TRH and Phe-TRH of 54% and 58%, respectively,at 8 h in STR. GSKI either had no effect or increased levels of TRHand TRH-like peptides 2–10-fold during the 8 h following injec-tion.
3.6. Effects at 2 h in brain
Effects at 2 h are considered particularly informative since veryrapid responses are not likely to be the result of changes in geneexpression which require more than 2 h to influence the pool sizeof fully processed and releasable TRH and TRH-like tripeptides.Increases, for example, in peptide content at this time are mostlike due to an acute decrease in the rate of peptide release andclearance by TRH-degrading enzymes. Of particular interest wasthe large number and magnitude of peptide level increases in CBL
at 2 h. Significant increases ranged from 3.3-fold for Peak ‘2’ to 9.7-fold for Leu-TRH (Table 1). These levels then fell gradually duringthe remaining 8 h after injection. Other brain areas with significanteffects at 2 h were HC: 2-fold increase in Trp-TRH; MED: 2.3-foldincrease in Glu-TRH and 1.8-fold increase in Peak ‘2’.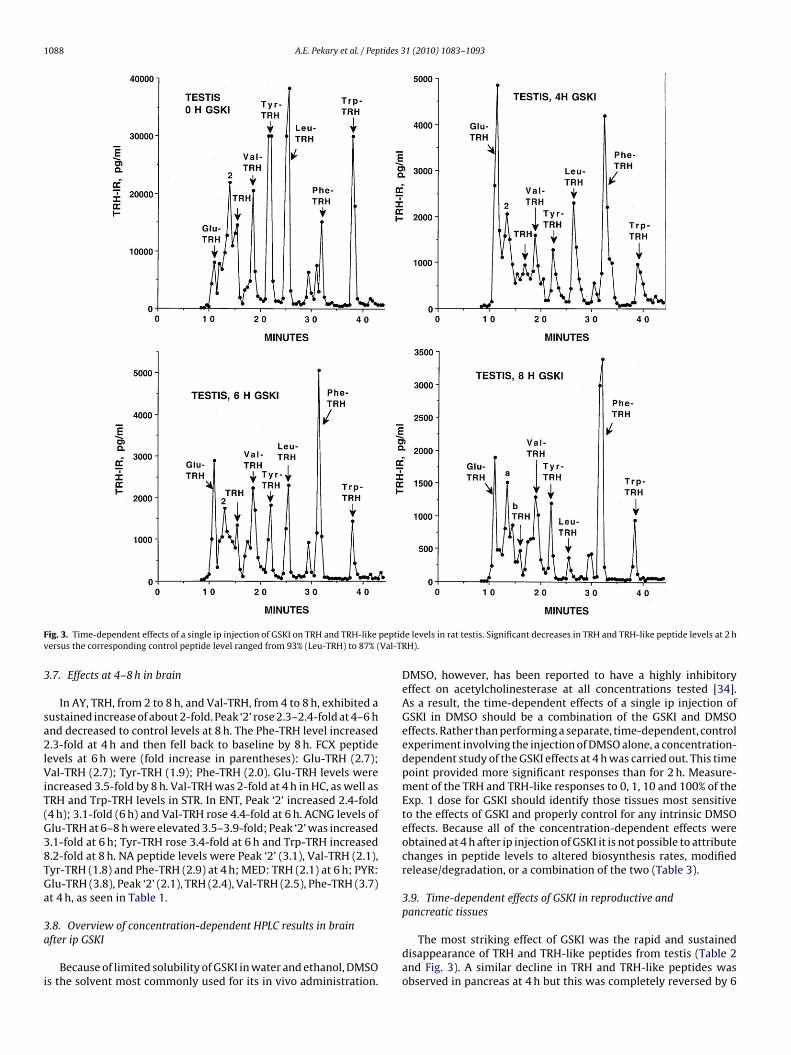
1088 A.E. Pekary et al. / Peptides 31 (2010) 1083–1093
F peptidv Val-TR
3
sa2lViT(G38TGa
3a
i
ig. 3. Time-dependent effects of a single ip injection of GSKI on TRH and TRH-likeersus the corresponding control peptide level ranged from 93% (Leu-TRH) to 87% (
.7. Effects at 4–8 h in brain
In AY, TRH, from 2 to 8 h, and Val-TRH, from 4 to 8 h, exhibited austained increase of about 2-fold. Peak ‘2’ rose 2.3–2.4-fold at 4–6 hnd decreased to control levels at 8 h. The Phe-TRH level increased.3-fold at 4 h and then fell back to baseline by 8 h. FCX peptide
evels at 6 h were (fold increase in parentheses): Glu-TRH (2.7);al-TRH (2.7); Tyr-TRH (1.9); Phe-TRH (2.0). Glu-TRH levels were
ncreased 3.5-fold by 8 h. Val-TRH was 2-fold at 4 h in HC, as well asRH and Trp-TRH levels in STR. In ENT, Peak ‘2’ increased 2.4-fold4 h); 3.1-fold (6 h) and Val-TRH rose 4.4-fold at 6 h. ACNG levels oflu-TRH at 6–8 h were elevated 3.5–3.9-fold; Peak ‘2’ was increased.1-fold at 6 h; Tyr-TRH rose 3.4-fold at 6 h and Trp-TRH increased.2-fold at 8 h. NA peptide levels were Peak ‘2’ (3.1), Val-TRH (2.1),yr-TRH (1.8) and Phe-TRH (2.9) at 4 h; MED: TRH (2.1) at 6 h; PYR:lu-TRH (3.8), Peak ‘2’ (2.1), TRH (2.4), Val-TRH (2.5), Phe-TRH (3.7)t 4 h, as seen in Table 1.
.8. Overview of concentration-dependent HPLC results in brainfter ip GSKI
Because of limited solubility of GSKI in water and ethanol, DMSOs the solvent most commonly used for its in vivo administration.
e levels in rat testis. Significant decreases in TRH and TRH-like peptide levels at 2 hH).
DMSO, however, has been reported to have a highly inhibitoryeffect on acetylcholinesterase at all concentrations tested [34].As a result, the time-dependent effects of a single ip injection ofGSKI in DMSO should be a combination of the GSKI and DMSOeffects. Rather than performing a separate, time-dependent, controlexperiment involving the injection of DMSO alone, a concentration-dependent study of the GSKI effects at 4 h was carried out. This timepoint provided more significant responses than for 2 h. Measure-ment of the TRH and TRH-like responses to 0, 1, 10 and 100% of theExp. 1 dose for GSKI should identify those tissues most sensitiveto the effects of GSKI and properly control for any intrinsic DMSOeffects. Because all of the concentration-dependent effects wereobtained at 4 h after ip injection of GSKI it is not possible to attributechanges in peptide levels to altered biosynthesis rates, modifiedrelease/degradation, or a combination of the two (Table 3).
3.9. Time-dependent effects of GSKI in reproductive andpancreatic tissues
The most striking effect of GSKI was the rapid and sustaineddisappearance of TRH and TRH-like peptides from testis (Table 2and Fig. 3). A similar decline in TRH and TRH-like peptides wasobserved in pancreas at 4 h but this was completely reversed by 6

A.E. Pekary et al. / Peptides 31 (2010) 1083–1093 1089
Table 2Acute, time-dependent effects of a single ip injection of GSKI (1.8 mg/kg) on TRH and TRH-like peptide levels in pancreas and reproductive tissues.
Glu-TRH Peak 2 TRH Val-TRH Tyr-TRH Leu-TRH Phe-TRH Trp-TRH
Pancreas2 h 115 178 82 158 127 80 126 1804 h 45* 63 32* 50* 43* 26* 17** 16**
6 h 63 110 64 97 83 139 106 1168 h 369* 212 133 296* 197 431** 229* 211*
Testis2 h 67 12** 70 13** 10*** 7*** 8*** 884 h 22** 19** 10*** 14** 8*** 10*** 11*** 8***
8 h 17** 11*** 3*** 12*** 4*** 2*** 10*** 5***
Epididymis2 h 38* 96 128 114 138 120 90 1724 h 67 140 107 128 91 111 126 846 h 98 126 127 114 98 152 165 938 h 191 156 153 111 127 154 203* 120
Prostate2 h 250* – 359* 274* 174 284* 198 1884 h 213* 101 59 193 270* 185 92 806 h 18** 139 86 65 47 50* 43* 24*
8 h 202* 165 139 99 151 165 107 84
R tectable.group.group.l group.
aEbTf(lTvP
3a
tawai(tT(
3c
f1
3c
mmto
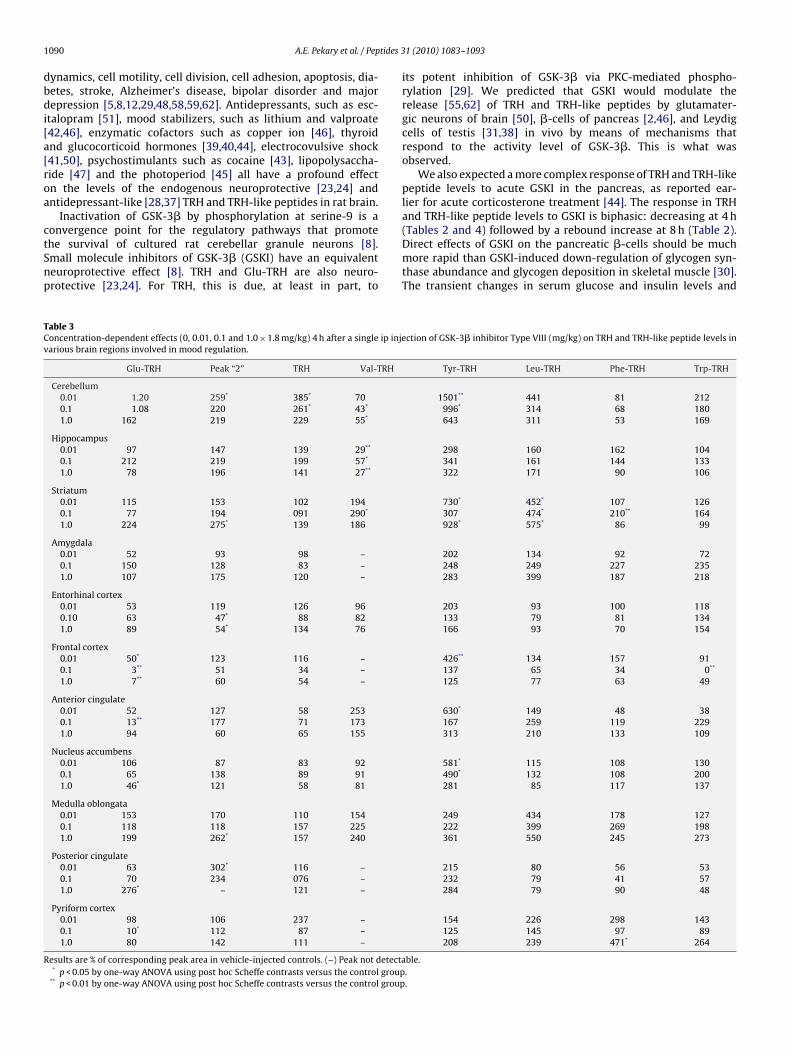
Fig. 4. TRH immunoreactivity (TRH-IR) released from rat testis during incubation
esults are % of corresponding peak area in vehicle-treated controls. (–) Peak not de* p < 0.05 by one-way ANOVA using post hoc Scheffe contrasts versus the control
** p < 0.01 by one-way ANOVA using post hoc Scheffe contrasts versus the control*** p < 0.001 by one-way ANOVA using post hoc Scheffe contrasts versus the contro
nd nearly all TRH-like peptides were significantly increased by 8 h.pididymal levels of TRH and TRH-like peptides were unaffectedy GSKI except for Glu-TRH which declined 62% at 2 h and Phe-RH which doubled at 8 h. At 2 h the prostate peptide levels as aold increase above the corresponding control values were Glu-TRH2.5), TRH (3.6); Val-TRH (2.7); Leu-TRH (2.8). At 4 h most peptideevels had returned to baseline except for Glu-TRH (2.1), and Tyr-RH (2.7). By 6 h, most peptide levels had decreased below controlalues (decrease in parentheses): Glu-TRH (82%), Leu-TRH (50%),he-TRH (57%), and Trp-TRH (76%) (Table 2).
.10. Concentration-dependent effects of GSKI in reproductivend pancreatic tissues
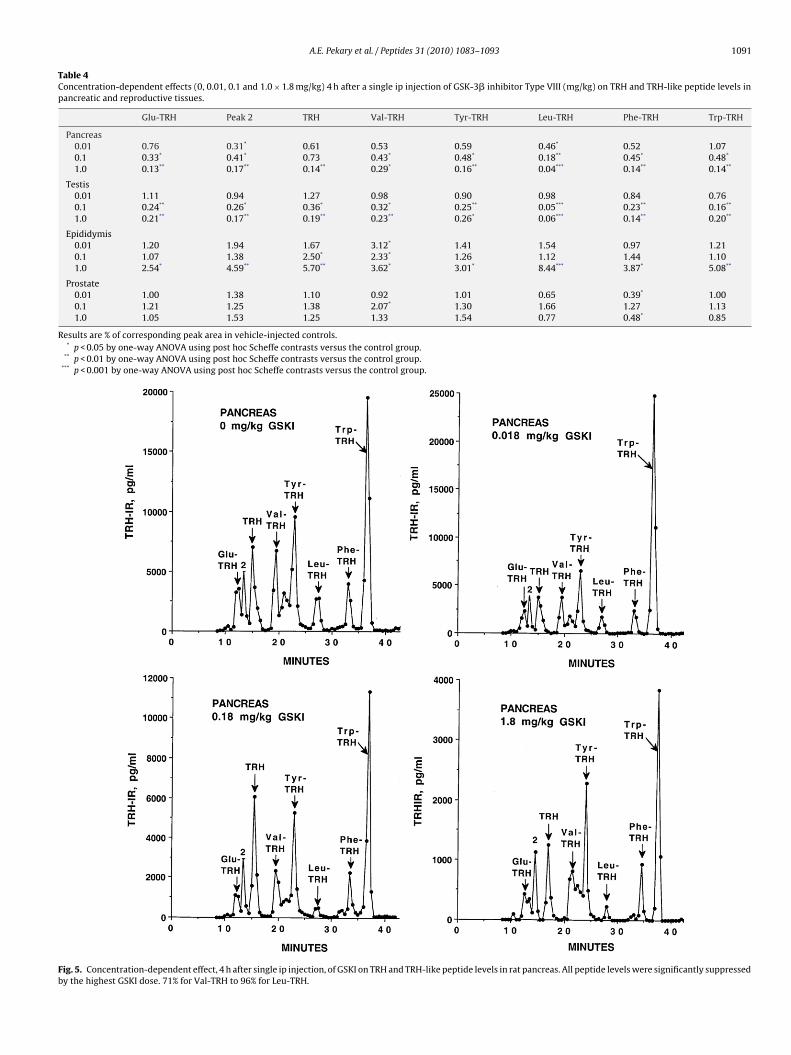
GSKI (0.18 and 1.8 mg/kg) had a profound suppressive effect onhe levels of TRH and TRH-like peptides in testis and pancreas 4 hfter ip injection (Table 4 and Fig. 5). These results were consistentith the corresponding time-dependent effects at 4 h summarized
bove and in Table 2. The 1.8 mg/kg dose of GSKI significantlyncreased the levels of TRH and all TRH-like peptides in epididymisTable 4). The 0.018 and 0.18 mg/kg doses of GSKI also increasedhe Val-TRH levels in epididymis. GSKI (0.18 mg/kg) increased Val-RH and 0.018 and 1.8 mg/kg GSKI suppressed Phe-TRH in prostateTable 4).
.11. Comparison of in vivo effects of GSKI and high-doseorticosterone
The time-dependent results above are strikingly similar to thoseollowing injection of Sprague–Dawley rats with a single ip dose of4 mg/kg corticosterone [44].
.12. Effects of GSKI and corticosterone on in vitro release andellular content of TRH-IR for decapsulated testis
The onsets of rapid release of TRH-IR from the testis frag-ents incubated in control, corticosterone- and GSKI-containingedium were at 1.28 ± 0.09 h; 0.59 ± 0.11 h (p < 0.01 versus con-
rol), and 0.85 ± 0.04 h (p < 0.05 versus control), respectively, basedn extrapolation of the maximum slopes (Fig. 4). The maximum
with 224 �M corticosterone hemisuccinate (corticosterone), 37 �M GSKI, or controlmedium versus time. Results are representative of two independent experiments.Error bars are 1 SD of duplicate measurements.
release rate was 438 ± 46 pg TRH-IR/ml/h (average for the 3 treat-ment groups) and did not differ between groups. TRH-IR representsthe combined immunoreactivity of TRH and TRH-like peptidesrelease by Leydig cells during the in vitro incubation [31,38,46].
4. Discussion
This is the first report that GSK-3� participates in the
corticosterone-induced regulation of TRH and TRH-like peptiderelease from rat brain, pancreas, and testis.GSK-3� is an enzyme that plays a pivotal role in regulating pro-tein synthesis, cell proliferation, and differentiation, microtubule
1 tides 3
dbdi[a[roa
ctSnp
TCv
R
090 A.E. Pekary et al. / Pep
ynamics, cell motility, cell division, cell adhesion, apoptosis, dia-etes, stroke, Alzheimer’s disease, bipolar disorder and majorepression [5,8,12,29,48,58,59,62]. Antidepressants, such as esc-
talopram [51], mood stabilizers, such as lithium and valproate42,46], enzymatic cofactors such as copper ion [46], thyroidnd glucocorticoid hormones [39,40,44], electrocovulsive shock41,50], psychostimulants such as cocaine [43], lipopolysaccha-ide [47] and the photoperiod [45] all have a profound effectn the levels of the endogenous neuroprotective [23,24] andntidepressant-like [28,37] TRH and TRH-like peptides in rat brain.
Inactivation of GSK-3� by phosphorylation at serine-9 is aonvergence point for the regulatory pathways that promote
he survival of cultured rat cerebellar granule neurons [8].mall molecule inhibitors of GSK-3� (GSKI) have an equivalenteuroprotective effect [8]. TRH and Glu-TRH are also neuro-rotective [23,24]. For TRH, this is due, at least in part, toable 3oncentration-dependent effects (0, 0.01, 0.1 and 1.0 × 1.8 mg/kg) 4 h after a single ip injearious brain regions involved in mood regulation.
Glu-TRH Peak “2” TRH Val-TRH
Cerebellum0.01 1.20 259* 385* 700.1 1.08 220 261* 43*
1.0 162 219 229 55*
Hippocampus0.01 97 147 139 29**
0.1 212 219 199 57*
1.0 78 196 141 27**
Striatum0.01 115 153 102 1940.1 77 194 091 290*
1.0 224 275* 139 186
Amygdala0.01 52 93 98 –0.1 150 128 83 –1.0 107 175 120 –
Entorhinal cortex0.01 53 119 126 960.10 63 47* 88 821.0 89 54* 134 76
Frontal cortex0.01 50* 123 116 –0.1 3** 51 34 –1.0 7** 60 54 –
Anterior cingulate0.01 52 127 58 2530.1 13** 177 71 1731.0 94 60 65 155
Nucleus accumbens0.01 106 87 83 920.1 65 138 89 911.0 46* 121 58 81
Medulla oblongata0.01 153 170 110 1540.1 118 118 157 2251.0 199 262* 157 240
Posterior cingulate0.01 63 302* 116 –0.1 70 234 076 –1.0 276* – 121 –
Pyriform cortex0.01 98 106 237 –0.1 10* 112 87 –1.0 80 142 111 –
esults are % of corresponding peak area in vehicle-injected controls. (–) Peak not detecta* p < 0.05 by one-way ANOVA using post hoc Scheffe contrasts versus the control group
** p < 0.01 by one-way ANOVA using post hoc Scheffe contrasts versus the control group
1 (2010) 1083–1093
its potent inhibition of GSK-3� via PKC-mediated phospho-rylation [29]. We predicted that GSKI would modulate therelease [55,62] of TRH and TRH-like peptides by glutamater-gic neurons of brain [50], �-cells of pancreas [2,46], and Leydigcells of testis [31,38] in vivo by means of mechanisms thatrespond to the activity level of GSK-3�. This is what wasobserved.
We also expected a more complex response of TRH and TRH-likepeptide levels to acute GSKI in the pancreas, as reported ear-lier for acute corticosterone treatment [44]. The response in TRHand TRH-like peptide levels to GSKI is biphasic: decreasing at 4 h(Tables 2 and 4) followed by a rebound increase at 8 h (Table 2).
Direct effects of GSKI on the pancreatic �-cells should be muchmore rapid than GSKI-induced down-regulation of glycogen syn-thase abundance and glycogen deposition in skeletal muscle [30].The transient changes in serum glucose and insulin levels andction of GSK-3� inhibitor Type VIII (mg/kg) on TRH and TRH-like peptide levels in
Tyr-TRH Leu-TRH Phe-TRH Trp-TRH
1501** 441 81 212996* 314 68 180643 311 53 169
298 160 162 104341 161 144 133322 171 90 106
730* 452* 107 126307 474* 210** 164928* 575* 86 99
202 134 92 72248 249 227 235283 399 187 218
203 93 100 118133 79 81 134166 93 70 154
426** 134 157 91137 65 34 0**
125 77 63 49
630* 149 48 38167 259 119 229313 210 133 109
581* 115 108 130490* 132 108 200281 85 117 137
249 434 178 127222 399 269 198361 550 245 273
215 80 56 53232 79 41 57284 79 90 48
154 226 298 143125 145 97 89208 239 471* 264
ble...
A.E. Pekary et al. / Peptides 31 (2010) 1083–1093 1091
Table 4Concentration-dependent effects (0, 0.01, 0.1 and 1.0 × 1.8 mg/kg) 4 h after a single ip injection of GSK-3� inhibitor Type VIII (mg/kg) on TRH and TRH-like peptide levels inpancreatic and reproductive tissues.
Glu-TRH Peak 2 TRH Val-TRH Tyr-TRH Leu-TRH Phe-TRH Trp-TRH
Pancreas0.01 0.76 0.31* 0.61 0.53 0.59 0.46* 0.52 1.070.1 0.33* 0.41* 0.73 0.43* 0.48* 0.18** 0.45* 0.48*
1.0 0.13** 0.17** 0.14** 0.29* 0.16** 0.04*** 0.14** 0.14**
Testis0.01 1.11 0.94 1.27 0.98 0.90 0.98 0.84 0.760.1 0.24** 0.26* 0.36* 0.32* 0.25** 0.05*** 0.23** 0.16**
1.0 0.21** 0.17** 0.19** 0.23** 0.26* 0.06*** 0.14** 0.20**
Epididymis0.01 1.20 1.94 1.67 3.12* 1.41 1.54 0.97 1.210.1 1.07 1.38 2.50* 2.33* 1.26 1.12 1.44 1.101.0 2.54* 4.59** 5.70** 3.62* 3.01* 8.44*** 3.87* 5.08**
Prostate0.01 1.00 1.38 1.10 0.92 1.01 0.65 0.39* 1.000.1 1.21 1.25 1.38 2.07* 1.30 1.66 1.27 1.131.0 1.05 1.53 1.25 1.33 1.54 0.77 0.48* 0.85
Results are % of corresponding peak area in vehicle-injected controls.* p < 0.05 by one-way ANOVA using post hoc Scheffe contrasts versus the control group.
** p < 0.01 by one-way ANOVA using post hoc Scheffe contrasts versus the control group.*** p < 0.001 by one-way ANOVA using post hoc Scheffe contrasts versus the control group.
Fig. 5. Concentration-dependent effect, 4 h after single ip injection, of GSKI on TRH and TRH-like peptide levels in rat pancreas. All peptide levels were significantly suppressedby the highest GSKI dose. 71% for Val-TRH to 96% for Leu-TRH.

1 tides 3
rrfhio3i
tt(nftsb[pGs
ianip4dcgcclr
htTk[p
paavtpapbTb
orist
A
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
092 A.E. Pekary et al. / Pep
esulting alteration in pancreatic insulin and TRH production andelease would follow later. (See serum glucose and insulin resultsor concentration-dependent experiment and Ref. [35]). Phe-TRHas previously been reported to oppose the insulin releasing activ-
ty of TRH [25]. Insulin inhibits GSK3 by promoting phosphorylationf serine residue-21 in GSK-3� and Ser-9 in GSK-3� [27]. GSK-attenuates insulin signaling by serine 332 phosphorylation of
nsulin receptor substrate-1 [26].Mature axons lack protein synthesis machinery so neuropep-
ides such as TRH and TRH-like peptides must be synthesized inhe cell body and then packaged into large dense core vesiclesLDCV) [19,20] for fast anterograde axonal transport to presy-aptic terminals [32]. The molecular motor kinesin is required
or fast anterograde transport of LDCV [7]. Kinesin is a heterote-ramer with two heavy and two light chains. Kinesin light chainubunits (KLCs) are essential for kinesin binding to membrane-ounded organelles (MBO) and provide MBO binding specificity7,9,22,26,27,30,32,53,56] and regulation of mobility [32,56]. GSK3hosphorylates KLCs and promotes kinesin release of MBOs [32].SK-3 modulates AMPA receptor trafficking via the kinesin cargoystem [11,33].
Glucocorticoids are responsible for maintaining the availabil-ty of energy for critical functions such as brain glucose levelsnd shutting off other functions, such as reproduction, that areot essential for survival during an emergency [40,44]. A strik-
ng observation from the present study was that TRH and TRH-likeeptides nearly disappeared from testis by 2 h after 1.8 mg/kg, orh after 0.18 mg/kg, GSKI. Acute, high-dose corticosterone pro-uced a nearly identical result for TRH-like peptides [44]. This wasonfirmed by in vitro experiments, summarized in Fig. 4, that sug-est that much of the rapid decrease in TRH and TRH-like peptideontent of rat testis, following a single high-dose ip injection oforticosterone or GSKI, is due to accelerated release and extracel-ular degradation. TRH is involved in the regulation of testosteroneelease by Leydig cells [13].
Several glucocorticoid and mineralocorticoid-binding proteinsave been described that can regulate the degree of phosphoryla-ion, and therefore the catalytic activity of GSK-3� and GSK-3�.hese include serum and glucocorticoid-inducible kinase-likeinase (SGKL) [10] or serum- and glucocorticoid-regulated kinase49]. PKB [60], CDK5 [32], PI3K, neurotrophin, Wnts and Netrin allhosphorylate GSK-3� [62].
In summary, GSKI can rapidly alter levels of TRH and TRH-likeeptides in brain and peripheral tissues, in part, through changingcutely the rate of secretion of these peptides. GSK3� and GSK3�re involved in the regulation of anterograde transport of secretoryesicles via phosphorylation of microtubules and adaptor proteinshat couple vesicles to transport motors such as kenesin. TRH is aotent inhibitor of GSK3�, suggesting that, in addition to its medi-tion of the neuroprotective effects of TRH and possibly TRH-likeeptides, GSK3� may alter TRH and TRH-like peptide release inrain. Evidence was also obtained that corticosterone modulatesRH, and TRH-like peptide release from testis via glucocorticoid-inding enzymes that inactivate GSK3�.
We conclude that some of the depressogenic effects of chronicr episodic treatment with glucocorticoids [40] are the result ofeductions in TRH and TRH-like peptide release in brain mediated,n part, by GSK-3�. In addition, this enzyme likely participates in thetress-induced suppression of pancreatic and reproductive func-ions through acceleration of depletion of TRH and related peptides.
cknowledgements
This work was supported by Veterans Administration Medicalesearch Funds (AEP and AS) and the Pekary Family Trust.
[
[
1 (2010) 1083–1093
References
[1] Amir S. Aging blocks the thermoregulatory action of thyrotropin-releasing hor-mone in anaesthetized rats. Brain Res 1988;440:181–4.
[2] Bacova Z, Najvirtova M, Krizanova O, Hudecova S, Zorad S, Strbak V, et al.Effect of neonatal streptozotocin and thyrotropin-releasing hormone treat-ments on insulin secretion in adult rats. Gen Physiol Biophys 2005;24:181–97.
[3] Benedetti F, Serretti A, Colombo C, Lorenzi C, Tubazio V, Smeraldi E. A glyco-gen synthase kinase 3-beta promoter gene single nucleotide polymorphism isassociated with age at onset and response to total sleep deprivation in bipolardepression. Neurosci Lett 2004;368:123–6.
[4] Benedetti F, Serretti A, Pontiggia A, Bernasconi A, Lorenzi C, Colombo C, etal. Long-term response to lithium salts in bipolar illness is influenced bythe glycogen synthase kinase 3-beta -50 T/C SNP. Neurosci Lett 2005;376:51–5.
[5] Bhat RV, Budd Haeberlein SL, Avila J. Glycogen synthase kinase: a drug targetfor CNS therapies. J Neurochem 2004;89:1212–317.
[6] Bhat RV, Xue Y, Berg S, Hellberg S, Ormo M, Nilsson Y, et al. Structural insightsand biological effects of glycogen synthase kinase 3-specific inhibitor AR-A014418. J Biol Chem 2003;278:45937–45.
[7] Brady ST. A kinesin medley: biochemical and functional heterogeneity. TrendsCell Biol 1995;5:159–64.
[8] Chin PC, Majadzadeh N, D’Mello SR. Inhibition of GSK3beta is a commonevent in neuroprotection by different survival factors. Brain Res Mol Brain Res2005;137:193–201.
[9] Cyr JL, Pfister KK, Bloom GS, Slaughter CA, Brady ST. Molecular genetics ofkinesin light chains: generation of isoforms by alternative splicing. Proc NatlAcad Sci USA 1991;88:10114–8.
10] Dai F, Yu L, He H, Chen Y, Yu J, Yang Y, et al. Human serum and glucocorticoid-inducible kinase-like kinase (SGKL) phosphorylates glycogen syntheses kinase3� (GSK-3�) at serine-9 through direct interaction. Biochem Biophys Res Com-mun 2002;293:1191–6.
11] Du J, Wei Y, Liu L, Khairova R, Wang Y, Blumenthal R, et al. GSK-3 kinasemodulates AMPA receptor trafficking via kinesin cargo system: molecularmechanism for mood stabilizer? Biol Psychiat 2009;65:16S. Abst. #52.
12] Frame S, Cohen P. GSK3 takes center stage more than 20 years after its discovery.Biochem J 2001;359(Pt. 1):1–16.
13] Gerendai I, Sziebert P, Gorcs T, Csaba Z, Csernus V. Effect of intratesticularadministration of TRH or anti-TRH antiserum on function of rat testis. Life Sci2000;67:269–81.
14] Gould TD, Einat H, Bhat R, Manji HK. AR-A014418, a selective GSK-3 inhibitor,produces antidepressant-like effects in the forced swim test. Int J Neuropsy-chopharmacol 2004;7:387.
15] Gould TD, Picchini AM, Einat H, Manji HK. Targeting glycogen synthase kinase-3 in the CNS: implications for the development of new treatments for mooddisorders. Curr Drug Targets 2006;7:1399–409.
16] Hinkle PM, Pekary AE, Senanayake S, Sattin A. Role of TRH receptors as possiblemediators of analeptic actions of TRH-like peptides. Brain Res 2002;935:59–64.
17] Inanami O, Ohno K, Sato A. Responses of regional cerebral blood flow to intra-venous administration of thyrotropin releasing hormone in aged rats. NeurosciLett 1992;143:151–4.
18] Jackson IM, Luo LG. Antidepressants inhibit glucocorticoid stimulation of thy-rotropin releasing hormone expression in cultured hypothalamic neurons. JInvest Med 1998;46:470–4.
19] Johansson O, Hokfelt T. Thyrotropin releasing hormone, somatostatin, andenkephalin: distribution studies using immunohistochemical techniques. J His-tochem Cytochem 1980;28:364–6.
20] Johansson O, Hokfelt T, Jeffcoate SL, White N, Sternberger LA. Ultrastructurallocalization of TRH-like immunoreactivity. Exp Brain Res 1980;38:1–10.
21] Kelly S, Zhao H, Sun GH, Cheng D, Qiao Y, Luo J, et al. Glycogen synthasekinase 3� inhibitor Chir025 reduces neuronal death resulting from oxygen-glucose deprivation, glutamate excitotoxicity, and cerebral ischemia. ExpNeurol 2004;188:378–86.
22] Khodjakov A, Lizunova EM, Minin AA, Koonce MP, Gyoeva FK. A specific lightchain of kinesin associates with mitochondria in cultured cells. Mol Biol Cell1998;9:333–43.
23] Koenig ML, Sgarlat CM, Yourick DL, Long JB, Meyerhoff JL. In vitro neuroprotec-tion against glutamate-induced toxicity by pGlu-Glu-Pro-NH2 (EEP). Peptides2001;22:2091–7.
24] Koenig ML, Yourick DL, Meyerhoff JL. Thyrotropin-releasing hormone (TRH)attenuates glutamate-stimulated increases in calcium in primary neuronal cul-tures. Brain Res 1996;730:143–9.
25] Kulkarni RN, Wang ZL, Akinsanya KO, Bennet WM, Wang RM, Smith DM, etal. Pyroglutamyl-phenylalanyl-proline amide attenuates thyrotropin-releasinghormone-stimulated insulin secretion in perifused rat islets and insulin-secreting clonal beta-cell lines. Endocrinology 1995;136:5155–64.
26] Liberman Z, Eldar-Finkelman H. Serine332 phosphorylation of insulin receptorsubstrate-1 byglycogen synthase kinase-3 attenuates insulin signaling. J BiolChem 2005;280:4422–8.
27] Lipina C, Huang X, Finlay D, McManus EJ, Alessi DR, Sutherland C. Analy-sis of hepatic gene transcription in mice expressing insulin-insensitive GSK3.Biochem J 2005;392:633–9.
28] Lloyd RL, Pekary AE, Sattin A, Amundson T. Antidepressant effects ofthyrotropin-releasing hormone analogues using a rodent model of depression.Pharmacol Biochem Behav 2001;70:15–22.

tides 3
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
A.E. Pekary et al. / Pep
29] Luo L, Stopa EG. Thyrotropin releasing hormone inhibits tau phosphoryla-tion by dual signaling pathways in hippocampal neurons. J Alzheimers Dis2004;6:527–36.
30] MacAulay K, Blair AS, Hajduch E, Terashima T, Baba O, Sutherland C, et al. Con-stitutive activation of GSK3 down-regulates glycogen synthase abundance andglycogen deposition in rat skeletal muscle cells. J Biol Chem 2005;280:9509–18.
31] Montagne JJ, Ladram A, Grouselle D, Nicolas P, Bulant M. Identification andcellular localization of thyrotropin-releasing hormone-related peptides in rattestis. Endocrinology 1996;137:185–91.
32] Morfini G, Szebenyi G, Brown H, Pat HC, Pigino G, DeBoer S, et al. A novel CDK5-dependent pathway for regulating GSK3 activity and kinesin-driven motility inneurons. EMBO J 2004;23:2235–45.
33] Nishimoto T, Kihara T, Akaike A, Niidome T, Sugimoto H. AMPA reducessurface expression of NR1 through regulation of GSK3beta. Neuroreport2009;20:161–5.
34] Obregon AD, Schetinger MR, Correa MM, Morscch VM, da Silva JE, Martins MA,et al. Effects per se of organic solvents in the cerebral acetylcholinesterase ofrats. Neurochem Res 2005;30:379–84.
35] Pearce NJ, Arch JRS, Clapham JC, Coghlan MP, Corcoran SL, Lister CA, et al. Devel-opment of glucose intolerance in male transgenic mice overexpressing humanglycogen synthase kinase-3� on a muscle-specific promoter. Metabolism2004;53:1322–30.
36] Pekary AE, Carlson HC, Yamada T, Sharp B, Walfish PG, Hershman JM.Thyrotropin-releasing hormone levels decrease in hypothalamus of aging rats.Neurobiol Aging 1984;5:221–6.
37] Pekary AE, Faull KF, Paulson M, Lloyd RL, Sattin A. TRH-like antidepressantpeptide, pyroglutamyltyrosylprolineamide, occurs in rat brain. J Mass Spectrom2005;40:1232–6.
38] Pekary AE, Meyer NV, Vaillant C, Hershman JM. Thyrotropin-releasing hor-mone and a homologous peptide in the male rat reproductive system. BiochemBiophys Res Commun 1980;95:993–1000.
39] Pekary AE, Sattin A. Regulation of TRH and TRH-like peptides in rat brain bythyroid and steroid hormones. Peptides 2001;22:1161–73.
40] Pekary AE, Sattin A, Blood J, Furst S. TRH and TRH-like peptide expression in ratfollowing episodic or continuous corticosterone. Psychoneuroendocrinology2008;33:1183–97.
41] Pekary AE, Sattin A, Lloyd RL. Electroconvulsive seizures increase levels of pGlu-Glu-Pro-NH2 (EEP) in rat brain. Peptides 1999;20:107–19.
42] Pekary AE, Sattin A, Meyerhoff JL, Chilingar M. Valproate modulates TRH recep-tor, TRH and TRH-like peptide levels in rat brain. Peptides 2004;25:647–58.
43] Pekary AE, Senanayake S, Sattin A. Cocaine regulates TRH-related peptides inrat brain. Neurochem Int 2002;41:415–28.
44] Pekary AE, Stevens SA, Sattin A. Rapid modulation of TRH and TRH-like pep-tide levels in rat brain and peripheral tissues by corticosterone. Neurochem Int
2006;48:208–17.45] Pekary AE, Stevens SA, Sattin A. Circadian rhythms of TRH-like peptide levelsin rat brain. Brain Res 2006;1125:67–76.
46] Pekary AE, Stevens SA, Sattin A. Valproate and copper accelerate TRH-likepeptide synthesis in male rat pancreas and reproductive tissues. Peptides2006;27:2901–11.
[
[
1 (2010) 1083–1093 1093
47] Pekary AE, Stevens SA, Sattin A. Lipopolysaccharide modulation of thyrotropin-releasing hormone (TRH) and TRH-like peptide levels in rat brain and endocrineorgans. J Mol Neurosci 2007;31:245–59.
48] Ring DB, Johnson KW, Henriksen EJ, Nuss JM, Goff D, Kinnick TR, et al. Selectiveglycogen synthase kinase 3 inhibitors potentiate insulin activation of glucosetransport and utilization in vitro and in vivo. Diabetes 2003;52:588–95.
49] Sakoda H, Gotoh Y, Katagiri H, Kurokawa M, Ono H, Onishi Y, et al. Differing rolesof Akt and serum-and glucocorticoid-regulated kinase in glucose metabolism,DNA synthesis, and oncogenic activity. J Biol Chem 2003;278:25802–7.
50] Sattin A. The role of TRH and related peptides in the mechanism of action ofECT. J ECT 1999;15:76–92.
51] Sattin A, Pekary AE, Blood J. Escitalopram regulates expression of TRH andTRH-like peptides in rat brain and peripheral tissues. Neuroendocrinology2008;88:135–46.
52] Scalabrino GA, Hogan N, O’Boyle KM, Slator GR, Gregg DJ, Fitchett CM, et al.Discovery of a dual action first-in-class peptide that mimics and enhancesCNS-mediated actions of thyrotropin-releasing hormone. Neuropharmacology2007;52:1472–81.
53] Stenoien DS, Brady ST. Immunochemical analysis of kinesin light chain func-tion. Mol Biol Cell 1997;8:675–89.
54] Takashima A. GSK-3 is essential in the pathogenesis of Alzheimer’s disease. JAlzheimers Dis 2006;9(3 Suppl.):309–17.
55] Tatebayashi Y, Haque N, Tung YC, Iqbal K, Grundke-Iqbal I. Role of tau phospho-rylation by glycogen synthase kinase 3� in the regulation of organelle transport.J Cell Sci 2004;117:1653–63.
56] Tsai M-Y, Morfini G, Szebenyi G, Brady ST. Modulation of kinesin-vesicle inter-actions by Hsc70: implications for regulation of fast axonal transport. Mol BiolCell 2000;11:2161–73.
57] Vasdev N, Garcia A, Stableford WT, Young AB, Meyer JH, Houle S, et al.Synthesis and ex vivo evaluation of carbon-11 labelled N-(4-methoxybenzyl)-N′-(5-nitro-1,3-thiazol-2-yl)urea ([11C]AR-A014418): A radiolabelled glycogensynthase kinase-3� specific inhibitor for PET studies. Bioorg Med Chem Lett2005;15:5270–3.
58] Wagman AS, Johnson KW, Bussiere DE. Discovery and development ofGSK3 inhibitors for the treatment of type 2 diabetes. Curr Pharm Des2004;10:1105–37.
59] Woodgett JR. Physiological roles of GSK-3 potential as a therapeutic target fordiabetes and other disorders. Curr Drug Targets Immune Endocr Metabol Disord2003;3:281–90.
60] Wyatt AW, Hussain A, Amann K, Klingel K, Kandolf R, Artunc F, et al. DOCA-induced phosphorylation of glycogen synthase kinase 3beta. Cell PhysiolBiochem 2006;17:137–44.
61] Zeng H, Schimpf BA, Rohde AD, Pavlova MN, Gragerov A, Bergmann JE.Thyrotropin-releasing hormone receptor 1-deficient mice display increased
depression and anxiety-like behavior. Mol Endocrinol 2007;21:2795–804.62] Zhou F-Q, Snider WD. GSK-3� and microtubule assembly in axons. Science2005;308:211–4.
63] Zmijewski JW, Jope RS. Nuclear accumulation of glycogen synthase kinase-3 during replicative senescence of human fibroblasts. Aging Cell 2004;3:309–17.




















