
Autoradiographic distribution of TRH binding sites in the human hypothalamus
11
66 BRES 17103 Brain Research, 563 (1991 ) 60 -7~ © 1991 Elsevier Science Publishers B.V, All rights reserved. 0006-8993/91/$03.51) ADONIS 0006899391171031 ~: Autoradiographic distribution of TRH binding sites in the human hypothalamus M. Najimi, F. Chigr, J. Champier, A. Tabib, N. Kopp and D. Jodani Laboratoire d'Anatornie Pathologique, Facultd de Mddecine Alexis Carrel, Lyon (France) (Accepted 11 June 1991) Key words: Autoradiography; Hypothalamus; Human; Thyrotropin-releasing hormone binding sites Using in vitro quantitative autoradiography and [3H]3MeTRH, a selective high affinity radioligand, we examined the rostrocaudal distri- bution of TRH binding sites in both the infant and the adult human hypothalamus. The saturation curve shows that the [3H]3MeTRH binds with high affinity to a single class of TRH binding sites and is saturable, the apparent constant of dissociation is in the namomolar range. TRH binding sites showed a wide distribution, principally in the anterior and mediobasal levels of the hypothalamus. TRH binding site concentration was highest within the diagonal band of Broca, the lateral preoptic area, the infundibular and the tuberal nuclei. TRH binding site concentration was moderate in the ventromedial nucleus and the medial preoptic area, whereas we observed low densities in the periven- tricular, paraventricular and mammillary nuclei. The distribution in the infant and the adult is generally similar. However, it is noteworthy that the infant tuberal nuclei displayed a lower binding site density when compared to the adult. On the other hand, the diagonal band of Broca is relatively more labeled in infant. The analysis of the whole hypothalamus allows us to ascertain the absence of lateral asymmetric distribution both in the infant and the adult. No significant difference is noticed when considering as paremeters of variation age, sex or post mortem delay. INTRODUCTION Thyrotropin-releasing hormone (TRH) was initially isolated from bovine and porcine hypothalamus 2'6 and identified as the tripeptide amine: pGlu-His-Pro-NH 2. Using biochemical and immunohistochemical tech- niques, many studies have reported that the hypothala- mus is the brain region containing the highest concen- trations of this tripeptide in mammalian species 5'8-1°'12' 16,26,27,43 including human 13'3°. Its biological actions have been reported to be mediated by specific membrane re- ceptors TM. The location of these receptor sites has been well documented in the central nervous system of many laboratory animals 18'35'37'38'41'42. In the rat, the TRH binding sites are widely distributed in many brain re- gions 18'21'37'38. More precisely, numerous TRH binding sites have been documented in several hypothalamic structures 18,37. In contrast to the murine brain, much less is known about the presence and the characteristics of TRH bind- ing sites in the human brain. A few studies, performed both by binding on tissue homogenates and autoradio- graphic studies, have demonstrated the presence of TRH binding sites in the whole central nervous system 2°'3~. However, while the regional distribution of these bind- ing sites has been studied in the human spinal cord 19 and hippocampus 11, no study had investigated the detailed regional distribution of such binding sites in the human hypothalamus. A recent report from our laboratory indicates that the TRH levels, as measured by radioimmunoassay (RIA), was higher in 3 structures, namely the ventromedial, the paraventricular and the dorsal nuclei, on the left side of the human hypothalamus 3. The aim of the present study is to investigate the bio- chemical characteristics and the detailed anatomical lo- calization of TRH binding sites in the human hypothal- amus using the well characterized analog [3H]MeTRH, by in vitro quantitative autoradiography. Further, to in- vestigate the developmental aspect of these binding sites, both infant and adult hypothalami have been processed. Finally, in order to confirm an eventual lateralization of TRH binding sites, the distribution and densities of la- beling have been determined for the 2 hypothalamic sides. Correspondence: M. Najimi, Laboratoire d'Anatomie Pathologique, Facult6 de Mrdecine Alexis Carrel, Lyon Cedex 08, France.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Autoradiographic distribution of TRH binding sites in the human hypothalamus

66
BRES 17103
Brain Research, 563 (1991 ) 60 -7~ © 1991 Elsevier Science Publishers B.V, All rights reserved. 0006-8993/91/$03.51)
ADONIS 0006899391171031 ~:
Autoradiographic distribution of TRH binding sites in the human hypothalamus
M. Najimi, F. Chigr, J. Champier, A. Tabib, N. Kopp and D. Jodani
Laboratoire d'Anatornie Pathologique, Facultd de Mddecine Alexis Carrel, Lyon (France)
(Accepted 11 June 1991)
Key words: Autoradiography; Hypothalamus; Human; Thyrotropin-releasing hormone binding sites
Using in vitro quantitative autoradiography and [3H]3MeTRH, a selective high affinity radioligand, we examined the rostrocaudal distri- bution of TRH binding sites in both the infant and the adult human hypothalamus. The saturation curve shows that the [3H]3MeTRH binds with high affinity to a single class of TRH binding sites and is saturable, the apparent constant of dissociation is in the namomolar range. TRH binding sites showed a wide distribution, principally in the anterior and mediobasal levels of the hypothalamus. TRH binding site concentration was highest within the diagonal band of Broca, the lateral preoptic area, the infundibular and the tuberal nuclei. TRH binding site concentration was moderate in the ventromedial nucleus and the medial preoptic area, whereas we observed low densities in the periven- tricular, paraventricular and mammillary nuclei. The distribution in the infant and the adult is generally similar. However, it is noteworthy that the infant tuberal nuclei displayed a lower binding site density when compared to the adult. On the other hand, the diagonal band of Broca is relatively more labeled in infant. The analysis of the whole hypothalamus allows us to ascertain the absence of lateral asymmetric distribution both in the infant and the adult. No significant difference is noticed when considering as paremeters of variation age, sex or post mortem delay.
INTRODUCTION
Thyrotropin-re leas ing hormone (TRH) was initially
isolated from bovine and porcine hypothalamus 2'6 and
identified as the t r ipept ide amine: pGlu-His -Pro -NH 2.
Using biochemical and immunohis tochemical tech-
niques, many studies have repor ted that the hypothala-
mus is the brain region containing the highest concen-
trations of this t r ipept ide in mammal ian species 5'8-1°'12' 16,26,27,43 including human 13'3°. Its biological actions have
been repor ted to be media ted by specific membrane re-
ceptors TM. The locat ion of these receptor sites has been
well documented in the central nervous system of many labora tory animals 18'35'37'38'41'42. In the rat, the T R H
binding sites are widely dis t r ibuted in many brain re- gions 18'21'37'38. More precisely, numerous T R H binding
sites have been documented in several hypothalamic structures 18,37.
In contrast to the murine brain, much less is known
about the presence and the characteristics of T R H bind-
ing sites in the human brain. A few studies, per formed
both by binding on tissue homogenates and autoradio-
graphic studies, have demons t ra ted the presence of T R H
binding sites in the whole central nervous system 2°'3~.
However , while the regional distr ibution of these bind-
ing sites has been studied in the human spinal cord 19 and
hippocampus 11, no study had investigated the detai led
regional distr ibution of such binding sites in the human
hypothalamus.
A recent repor t from our labora tory indicates that the
T R H levels, as measured by radio immunoassay (RIA) ,
was higher in 3 structures, namely the ventromedial , the
paraventr icular and the dorsal nuclei, on the left side of
the human hypothalamus 3.
The aim of the present study is to investigate the bio-
chemical characterist ics and the detai led anatomical lo-
calization of T R H binding sites in the human hypothal-
amus using the well character ized analog [3H]MeTRH,
by in vitro quanti tat ive autoradiography. Fur ther , to in-
vestigate the developmenta l aspect of these binding sites,
both infant and adult hypothalami have been processed.
Finally, in order to confirm an eventual lateral izat ion of
T R H binding sites, the distr ibution and densities of la-
beling have been de te rmined for the 2 hypothalamic
sides.
Correspondence: M. Najimi, Laboratoire d'Anatomie Pathologique, Facult6 de Mrdecine Alexis Carrel, Lyon Cedex 08, France.

TABLE I
Source of brain tissues
Cases Sex Age Post Cause of mortem death delay
Adults A M 22 y B F 27 y C M 34 y D F 42 y E F 45 y F M 47 y G M 67 y H F 82 y
Neonates I F J M
K M
L M M M N F
30 h Cardiac failure 25 h Cardiac failure
28.5 h Sudden death 12 h Sudden death 20 h Coronary thrombosis 12 h Cardiac failure 7 h Infarctus
2.5 h Cardiac failure
2 h 14 h Pulmonary hypoplasia 35 h 34 h Amniotic inhalation
with gastric regurgitation
1 day I0 h Oedematic alveolitis with refractory hypoplasia
1 month 20 h Liver lesions 1 month 5 h Enterocolitis 1 year 7 h Hepatic necrosis
MATERIALS AND METHODS
Fourteen brains (8 males and 6 females, 8 adults and 6 neo- nates), obtained from Edouard Herriot and Lyon Sud Hospitals, Lyon, France, were used in this study (Table I). None of them presented clinical, neurological or neuroendocrinological signs. Fur- thermore, no pathological lesions were observed after macroscopic and microscopic examination of the brains. After removing the brain at autopsy, the hypothalamus was dissected at 4 °C and stored at -80 °C. The hypothalamus sample was dissected, taking the parallel plane joining the optic chiasm and anterior commissure as frontal plane and the caudal plane just behind the mammillary complex. The sample remained 1-6 days at -20 °C before cutting into 20 pm-thick sections on a Micron cryostat at -20 °C. The sec- tions were mounted on gelatine chromalun coated glass slides and stored at -20 °C until they were processed for autoradiography.
For anatomical localization of the hypothalamic nuclei and areas, adjacent sections to those used for autoradiography were stained with Cresyl violet. Identification of the hypothalamic nuclei and areas was made according to an atlas of the human hypothalamus developed in our laboratory.
In the autoradiography procedure we used, the sections were preincubated for 20 rain at room temperature in 0.05 M Tris HCI buffer (pH 7.4) containing 0.2 % bovine serum albumin (BSA). After drying, they were incubated at 4 °C in the same medium to which [3H]3MeTRH (10 nM) (specific activity: 87 Ci/mmol), MgC12 (5 x 10 -6 M) and bacitracin (2 × 10 -s M) had been added. Non- specific binding was determined by incubation of adjacent sections in the same medium containing 10 -6 M non-radioactive TRH. Af- ter a 2 h incubation period, the sections were washed 4 times in ice cold buffer (30 s/bath) and once in distilled water (15 s) and dried immediately at room temperature.
The labeled sections, together with standards (Amersham), were apposed to 3H ultrofilm (Amersham) for 2 months at 4 °C. The films were processed in Dektol (Kodak) developer and the optical density of the autoradiographic images quantitated in terms of fmol/mg tissue using an Imstar-Starwise computer and image anal- ysis system.
67
For saturation studies, sections from the anterior and mediobasal hypothalamus from 4 different human hypothalami (2 adults and 2 infants) were incubated with ligand concentrations ranging from 1 to 30 nM. For displacement experiments, serial sections from the preoptic area and infundibular nucleus were incubated in the same incubation medium containing graded concentrations of unlabeled TRH and 3Me-TRH (10-12-10 -6 M). Kinetic analysis were com- puted by the method of Parker and Waud 29, in which the comput- erized process is based on the conventional method of the least squares, including the calculation of the variance-covariance ma- trix. Standard errors on the best estimates of the Imax and IC50 values were calculated from the coefficients of the variance-cova- fiance matrix.
Chemicals [3H]MeTRH was purchased from NEN. All peptides were ob-
tained from Peninsula; bovine serum albumin, gelatin, Tris and ba- citracin from Sigma; MgCI 2 from Merck.
RESULTS
Scatchard analysis of [3H]MeTRH binding sites
The experiments have focused on 2 hypothalamic
structures: the preoptic area in the anterior hypothala-
mus and the infundibular nucleus in the mediobasal hy-
pothalamus. The saturation curve showed that specific
[3H]MeTRH binding to the 2 structures analyzed is sat-
urable (Fig. 1). Scatchard analysis revealed that the ap-
parent kDa values and maximal binding capacity are
equivalent both in infant and adult (Table II).
Inhibition analysis of [3H]MeTRH binding sites
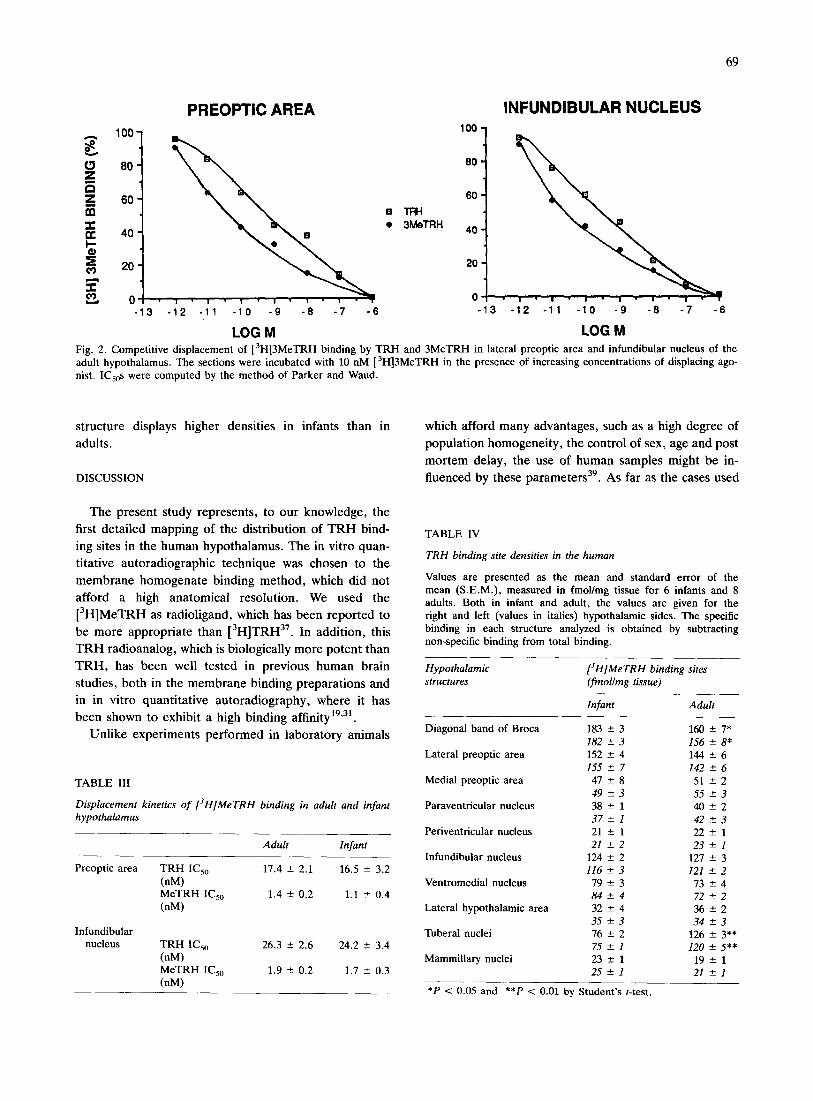
In these experiments, we have tested the ability of
many T R H analogs to inhibit the binding of
[3H]MeTRH. Among the analogs used, only M e T R H
and T R H inhibited competitively the specific binding in
both preoptic area and infundibular nucleus by 50% with
nanomolar potency. M e T R H was more potent than T R H
(Fig. 2, Table III). By contrast, T R H - O H , DKP, His-
Pro-Glu, as well as LH R H , VIP, neurotensin, somato-
statin and substance P, did not displace [3H]MeTRH
binding in any of the regions studied (data not shown).
Autoradiographic distribution
The data obtained from the quantitative autoradio-
graphic study of the binding of [3H]MeTRH in the dif-
ferent hypothalamic structures are summarized in Table
IV and Figs. 3-5. The resulting detailed mapping in the
adult and infant hypothalamus appeared as follows:
Adult hypothalamus. A. Anter ior hypothalamus. This
hypothalamic region (Figs. 3 A - C ; 4 A - C ) is very rich in
T R H binding sites. The highest concentrations were lo-
cated principally in the diagonal band of Broca and the
preoptic area. The T R H binding sites are distributed in
the horizontal and the vertical limbs of the diagonal
band of Broca with the same density and without sharp

68
"~" PREOPTIC AREA 1 2 0 -
1oo"
60
"N 40
[3H] MeTRH (nM)
INFUNDIBULAR NUCLEUS
_g
g
,~. [3H] Me TRN (nM)
Fig. I. Kinetics of [3H]3MeTRH binding to slide-mounted human hypothalamic sections as a function of [3H]3MeTRH concentration. Hypothalamic sections representing lateral preoptic area and in- fundibular nucleus were incubated for 2 h at 4 °C with increasing concentrations of tritiated 3MeTRH. Sections were apposed under ultrofilm and the binding was quantitated after autoradiography. Specific binding is defined by subtracting non-specific from total binding. Points are average of triplicate separate determinations. Average data from scatchard analysis are reported in Table II.
delimitation. Anteriorly, the labeling covers homoge- neously the whole anatomical extent of the structure.
This labeling does not change from the rostral to the caudal extent. Very high concentrations were also found
TABLE II
Kinetics at equilibrium of [3H]3MeTRH binding in adult and in- fant hypothalamus
Adult Infant
Preoptic area kDa (nM) 4.8 ± 0.6 5.3 -+ 0.5 Bma x 147 ± 14 135 -+ 12 (fmol/mg tissue)
Infundibular kDa (nM) 7.6 ± 0.5 6.5 ± 0.5 nucleus Bran x 114 ± 5 120 ± 16
(fmol/mg tissue)
in the lateral preoptic area. This labeling is continuous
with that present in the horizontal limb of the diagonal
band of Broca. It decreases caudally. Thc medial part ot the preoptic area, in contrast to the lateral part, displays
low densities. The paraventricular nucleus shows low but
significant binding site densities. The labeling is distrib-
uted homogeneously in both the parvocellular and mag-
nocellular parts of the nucleus. Further, similar densities were measured in the anterior and mediobasal levels of
the nucleus. The suprachiasmatic and supraoptic nuclei
do not display any significant labeling. Finally, the lat-
eral hypothalamic area displays low densities. B. Mediobasal hypothalamus. Most parts of the me-
diobasal hypothalamus exhibit high to intermediate den-
sities (Figs. 3D,E; 4D,E). The infundibular nucleus is
one of the mediobasal structures displaying high densi- ties. However, the labeling does not cover whole the
nucleus, it is mainly concentrated in the ventral part.
Along the anteroposterior extent of the nucleus, the au-
toradiographic grain density displays a heterogeneous distribution. While the T R H binding sites are very dense
in the anterior part, they decrease in the posterior level and become rare. Caudally, the medial and lateral tu-
beral nuclei display very high densities. Labeling is ho-
mogeneous within these 2 structures. As for the in- fundibular nucleus, TRH binding sites were localized
only in the ventral part. In contrast, no heterogeneous
distribution was observed. More dorsally in the mediobasal hypothalamus, the
ventromedial nucleus displayed intermediate T R H bind-
ing site densities. Labeling is distributed throughout the nucleus. As in the anterior region, low densities of T R H
binding sites are observed in the lateral hypothalamic
area. The median eminence as well as the fornix column do
not display any significant labeling. C. Posterior hypothalamus. This hypothalamic level
displays generally low T R H binding site concentrations
(Figs, 3F; 4F). Low levels of T R H binding sites could
be found in the medial mammillary nucleus. However, the lateral part of the mammillary nucleus displays in-
termediate T R H binding site densities. Autoradiographic labeling was absent further caudally. Dorsally, the sub- thalamic nucleus displays no autoradiographic labeling.
Infant hypothalamus. The same general autoradio- graphic distribution of T R H binding sites (Fig. 5A-F) is observed in both the infant and adult hypothalamus. However, in the mediobasal hypothalamic level, while the adult tuberal nuclei display high densities, only low T R H binding site concentrations were detected in these nuclei in infants. Another discrepancy between the T R H binding site density distribution in the infant and the adult was observed in the diagonal band of Broca: this
69
1°° t
t 6O
4 0 -
20
0 - 3
PREOPTIC AREA 100
80
6O
3MeTRH 40
20
0 I " I " •
- ,2 -11 - ; -'s -'7 -6 -
L O G M
INFUNDIBULAR NUCLEUS
-12 -11 -10 -9 -8 -7 -6
L O G M
Fig. 2. Competitive displacement of [3H]3MeTRH binding by TRH and 3MeTRH in lateral preoptic area and infundibular nucleus of the adult hypothalamus. The sections were incubated with 10 nM [3H]3MeTRH in the presence of increasing concentrations of displacing ago- hist. ICs0s were computed by the method of Parker and Waud.
structure displays higher densities in infants than in
adults.
DISCUSSION
The present study represents, to our knowledge, the
first detailed mapping of the distribution of T R H bind-
ing sites in the human hypothalamus. The in vitro quan-
titative autoradiographic technique was chosen to the
membrane homogenate binding method, which did not
afford a high anatomical resolution. We used the
[3H]MeTRH as radioligand, which has been reported to
be more appropriate than [3H]TRH37. In addition, this
T R H radioanalog, which is biologically more potent than
T R H , has been well tested in previous human brain
studies, both in the membrane binding preparations and
in in vitro quantitative autoradiography, where it has
been shown to exhibit a high binding affinity 19"31.
Unlike experiments performed in laboratory animals
TABLE III
Displacement kinetics o f [3H]MeTRH binding in adult and infant hypothalamus
Adult Infant
Preoptic area TRH IC50 17.4 +- 2.1 16.5 -+ 3.2 (nM) MeTRH IC5o 1.4 --- 0.2 1.1 +- 0.4 (nM)
Infundibular nucleus TRH IC5o 26.3 -+ 2.6 24.2 -+ 3.4
(nM) MeTRH IC5o 1.9 -+ 0.2 1.7 -+ 0.3 (nM)
which afford many advantages, such as a high degree of
populat ion homogeneity, the control of sex, age and post
mortem delay, the use of human samples might be in-
fluenced by these parameters 39. As far as the cases used
TABLE IV
TRH binding site densities in the human
Values are presented as the mean and standard error of the mean (S.E.M.), measured in fmol/mg tissue for 6 infants and 8 adults. Both in infant and adult, the values are given for the fight and left (values in italics) hypothalamic sides. The specific binding in each structure analyzed is obtained by subtracting non-specific binding from total binding.
Hypothalamic structures
[3H]MeTRH binding sites (fmol/mg tissue)
Infant Adult
Diagonal band of Broca 183 +- 3 160 182 +- 3 156
Lateral preoptic area 152 -+ 4 144 155 +- 7 142
Medial preoptic area 47 +- 8 51 49 -+3 55
Paraventricular nucleus 38 +- 1 40 37++-1 42
Pefiventricular nucleus 21 -+ 1 22 21 -+2 23
Infundibular nucleus 124 -+ 2 127 116 +- 3 121
Ventromedial nucleus 79 +- 3 73 84++-4 72
Lateral hypothalamic area 32 +- 4 36 35 +-3 34
Tuberal nuclei 76 +- 2 126 75 +- 1 120
Mammillary nuclei 23 -+ 1 19 25+-1 21
- 7* +- 8* +-6 +-6 -+2 -+3 +-2 -+3 -+1 +-1 +-3 +-2 -+4 +-2 -+2 -+3 -+ 3** -+ 5"* +-1 +-1
*P < 0.05 and **P < 0.01 by Student's t-test.

70
m
M , !,
A
r /
.2
A
¢ - - ' - v
Fig. 3. Schematic coronal sections of the human hypothalamus drawn from adult, representing the distribution and densities of TRH binding sites at different hypothalamic levels (A-C: anterior level; D,E: mediobasal level; F: posterior level). 1~-~1, >140 fmol/mg tissue; [ ] 101-140 fmol/mg tissue; [K-I, 45-100 fmol/mg tissue; [7], <45 fmol/mg tissue.
Fig. 4. Regional variation of TRH binding sites in the human adult hypothalamus. Coronal sections were labeled with 10 nM [ 3H]MeTRH. The panels progress from rostral to caudal and are the level of: A-C: anterior hypothalamus; D and E: mediobasal hypothalamus; and F: posterior hypothalamus.

71

72
A~
D
C\
m
~, !~i i~ iii~ • AC
i
I
/
P I
Fig. 5. Autoradiographic distribution of TRH binding sites in the human infant hypothalamus. Rostrocaudally, the labeling is present in the anterior (A-C), the mediobasal (D-E) and posterior (F) levels•

in this study are concerned, we did not notice any sig- nificant fluctuation of binding site densities which could
be related to these parameters. This has also been re- ported for other neuropeptides or neurotransmitter bind- ing sites (D 2 and benzodiazepine receptors 28"44) (so- matostatin and L H R H binding sites32"33).
In our experimental conditions, the saturation curve shows that [3H]3MeTRH binds with high affinity to a
single class of saturable TRH binding sites. The appar- ent constant of dissociation is in the nanomolar range in the preoptic area and infundibular nucleus. These results are in good agreement with the biochemical kinetics of [3H]MeTRH which we previously reported for the hu- man hippocampus n. Furthermore, the displacement ki- netics show that 3MeTRH is the more potent analog for displacement of [3H]MeTRH binding. These character-
istics agree well with the main biochemical findings in the rat and human brain 18"31"37.
The TRH binding sites in the human hypothalamus, as reported in this study by quantitative autoradiogra- phy, displayed a wide and heterogeneous distribution. In particular, the highest densities of T R H binding sites are localized in the anterior and mediobasal hypothalamic levels. Within the anterior hypothalamic level, the bind- ing sites are principally concentrated in the diagonal band of Broca and the lateral part of the preoptic area. In the mediobasal hypothalamic level, the highest den- sities have been observed in the infundibular, the ven- tromedial and the tuberal nuclei.
In this study, we have examined the distribution of TRH binding sites in the hypothalamus both in the in-
fant and in the adult. The distribution in both cases is in general similar. However, it is noteworthy that the in- fant tuberal nuclei displayed less binding site densities when compared to the adult. We have recently reported a similar observation for the distribution of somatostatin binding sites in the tuberal nuclei 2s. It seems that the
first occurrence of TRH binding sites in the infant tu- beral hypothalamic nuclei begins during the first postna- tal year and probably increases to reach the adult level. The differences in TRH binding site densities as sug- gested for SRIF binding sites could be explained by a developmental process. Another explanation of these differences is the fact that these nuclei are well differ- entiated only in humans and not in lower mammals 7. Their later ontogenic appearance could reflect their re- cent phylogenic evolution. Another interesting differ- ence between infant and adult is illustrated by the pres- ence of high TRH binding site densities in the infant diagonal band of Broca. These postnatal changes suggest that TRH, as other neurosubstances, may act as a neuro- transmitter or a neuromodulator.
Recently, we have investigated, by radioimmunologi-
73
cal studies, the lateralization of endogenous T R H in the adult human hypothalamus 3. The comparison of T R H
concentrations between the 2 hypothalamic sides, re-
vealed the presence of highest levels in the right hypo- thalamic ventromedial, paraventricular and dorsal nu- clei. However, in the present study, the comparison of TRH binding site densities on the same structures in the 2 hypothalamic sides did not reveal any structure later- alization.
The rare previous studies describing the presence of TRH binding sites in the human hypothalamus 2°m, ei-
ther by an in vitro homogenate binding or a quantitative autoradiography method, reported TRH receptor levels in the hypothalamus and mammillary bodies only, with- out specifying their anatomical distribution. Thus, it is difficult to establish any anatomical comparison of our distribution of TRH binding sites in the human hypo- thalamus with these studies.
The comparison of our data with the distribution of TRH binding sites in rat hypothalamus is difficult to es- tablish. This is due to the fact that these studies have screened all the brain and no report has been focused on the hypothalamus as for somatostatin binding sites for example 14. The recent autoradiographic studies per-
formed in the rat hypothalamus did not give a precise and detailed mapping in the hypothalamus 18"35"37. How-
ever, the analysis of these data leads us to consider that overall the same distribution is present in human and rat hypothalamus. Thus, high T R H binding site densities have been found in the preoptic region and ventrome- dial nucleus. However, these studies do not mention in which anatomical portion of the preoptic area the stain- ing is localized, or the presence of the T R H binding sites in the paraventricular 37 or infundibular nuclei 37. In con-
trast to the previous studies in rat, reporting moderate densities of T R H binding sites in the posterior mammil- lary nucleus 37, we cannot detect any autoradiographic
labeling in this structure. In the mammillary nuclei, we can detect low densities present in the lateral and me- dial parts. Concerning the difference between the 2 structures, it could reflect species differences. Thus, it is interesting to note that the anatomical subdivisions, re- ported for the rat mammillary nucleus, are not detect- able cytoarchitecturally in the human 34. However, since the cytoarchitectural features of the mammillary com- plex is different in various mammals studied, it is diffi- cult to establish comparison between the anatomical sub- divisions labeled in the mammillary complex in rats and in humans. Another interesting discrepancy between the 2 species is illustrated by the labeling of the tuberal nu- clei. These structures, subdivided in medial and lateral parts, display high TRH binding site densities in the hu- man whereas an absence of labeling is reported in the

74
rat. However, previous cytoarchitectural studies in rat delineate these nuclei more laterally than in the human1; this might impair the comparison. In conclusion, inter-
specific differences in patterns of TRH binding site dis- tribution illustrate difficulties encountered when attempt-
ing to extrapolate from laboratory animal to human, and they emphasize the importance of studying human tis- sues.
The comparison between the distribution of T R H binding site densities and that of endogenous T R H re- vealed by biochemical or immunohistochemical studies
principally evidenced the presence of a discrepancy be- tween the 2 distributions in most rat brain areas 1s'21'35.
Unfortunately, a comparison of TRH binding site distri-
bution as presented in this study, with TRH immunore- active content (TRH-IR) is impossible to establish. In- deed, there are no immunohistochemical data relating the distribution of TRH-IR in human hypothalamus. We have tried to demonstrate the presence of TRH-IR in human hypothalamus but our different attempts (with
varying fixative solutions) failed in various tissues where, on the other hand, we demonstrated the presence of other hypophysiotropic peptides as somatostatin and L H R H 23'24. Thus, comparison can only be done with re-
ports dealing with microdissection of the hypothalamus and measurements of endogenous TRH as revealed by RIA 3'13'21. However, such comparison is restricted by
the fact that the dissection techniques of the human hy- pothalamus for RIA cannot avoid a possible contamina- tion of some hypothalamic structures by other neigh- bouring anatomical structures. In the mediobasal hypothalamus, a very good correlation is found for the ventromedial and infundibular nuclei. These nuclei dis- play both high densities of TRH binding sites and en- dogenous RIA TRH. A good correlation is also present for the preoptic region. Furthermore, the mammillary complex, where we detect relatively low densities, has also been reported to display low amounts of TRH. However, some structures like the paraventricular nu- cleus and the dorsomedial nucleus showing high densi- ties of TRH immunoreactivity by RIA exhibited a low density and an absence of labeling of T R H binding sites,
respectively. Several lines of evidence suggest that TRH is involved
in the control of thermoregulation. Indeed, previous physiological studies, using intracerebroventricular or peripheral injections of TRH or TRH analogs reported an elevation of central temperature 15'17. It seems that the anterior hypothalamic region is responsible for such effects. Indeed, it has been reported that after preoptic area lesions, a change in the amplitude of body temper- ature is observed in rat 4'36'4°. Moreover, microiontopho- retic application of TRH resulted in excitation of the
majority of cold-responsive neurons recorded in the p r e optic area 17. These studies support the involvement of
TRH and the preoptic area in thermoregulatory control. The presence of high densities of TRH binding sites in the preoptic region support the hypothesis that TRH might be involved in thermoregulation in humans and act as a neurotransmitter or neuromodulator in the pre- optic region, since early ages.
Interestingly, the lack of TRH binding sites in the median eminence is in good agreement with the hy- pophysiotropic role of TRH. Furthermore, the absence of corresponding binding sites of other hypophysiotropic peptides such as somatostatin in the median eminence has also been reported 14'25.
In the present study, we reported that the infundibu- lar nucleus displays high densities of TRH binding sites. Recently, we reported in immunohistochemistry studies that this nucleus also contains high densities of so- matostatin immunoreactive cell bodies 23. These findings
suggest that there may be interactions between TRH and
somatostatin. It is noteworthy that the results presented in this study
indicate an association, although not exclusive, of TRH binding sites with several structures involved in the pro- cess of memory, for instance, the diagonal band of Broca and the mammillary complex. Furthermore, we have also recently demonstrated the presence of important densities of such sites in human hippocampus II, an other anatomical structure involved in memory. Moreover, previous pharmacological studies indicated that TRH and TRH analogs could decrease memory impairment, caused by bilateral lesions of the basal forebrain, in some animal models 22.
In conclusion, in this report we described the regional localization of TRH binding sites, both in infant and adult human hypothalamus. This approach complements previous studies of TRH binding sites in other human brain structures such as the spinal cord and the hippo- campus. This study also demonstrates the usefulness of [3H]MeTRH for autoradiography in human studies. The results obtained provide a detailed anatomical mapping of TRH binding sites in the human hypothalamus. This could be of value for future study concerning the pathol- ogies involving TRH and the hypothalamus such as sud- den infant death syndrome and depression (works in
progress).
Acknowledgements. This work was supported by Grants from INSERM no. 876013, CR INSERM 884669 and Biologie Humaine no. 925-358. The authors would like to thank Pr. R. Gilly and Dr. R. Bouvier for their efficient help in obtaining human tissue sam- pies, Drs. J. Epelbaum and A. Sarrieau and Pr. B. Bloch for their helpful suggestions, and V. Thivolle for her secretarial help.

ABBREVIATIONS
AC DBBh DDBv DM F IN LH LM LP LT
Anterior Commissure Diagonal Band of Broca horizontalis Diagonal Band of Broca verticalis Dorsomedial nucleus Fornix Infundibular nucleus Lateral Hypothalamic area Lateral Mammillary nucleus Lateral Preoptic area Lateral Tuberal nucleus
REFERENCES
1 Bleier, R., Cohn, P. and Siggelkow, I., A cytoarchitectonic at- las of the hypothalamus and hypothalamic third ventricle of the rat. In J. Morgane and J. Panksep (Eds.), Handbook of the Hypothalamus, 1. Anatomy of the Hypothalamus, 1 (1981) 137- 220.
2 Boler, R.M., Enzman, E, Folkers, K., Bowers, C.Y. and Schally, A.V., The identity of chemical and hormonal proper- ties of the thyrotropin-releasing hormone and pyroglutamyl-his- tidyl-proline amide, Biochem. Biophys. Res. Commun., 37 (1969) 705-710.
3 Borson-Chazot, E, Jordan, D., F6vre-Montagne, M., Kopp, N., Tourniaire, J., Rouzioux, J., Veisseire, M. and Mornex, R., TRH and LHRH distribution in discrete nuclei of hypothala- mus: evidence for a left prominence of TRH, Brain Research, 382 (1986) 433-436.
4 Briese, E., Do medial preoptic lesions interfere with the set- point of temperature regulation?, Brain Res. Bull., 23 (1989) 137-144.
5 Brownstein, M.J., Palkovits, M., Saavedra, J.M., Bassiri, R.M. and Utiger, R.D., Thyrotropin releasing hormone in specific nuclei of the rat brain, Science, 185 (1974) 267-269.
6 Burgus, R., Dunn, T.E, Desiderio, D. and Guillemin, R.G., Structure mol6culaire du facteur hypothalamique hypophysio- trope, TRI-I d'origine ovine: mise en 6vidence pour spectrom- 6trie de masse de la s6quence PCA-His-Pro-NH2, C.R. Acad. Sci. (Paris), 269 (1969) 1870-1873.
7 Grfinthal, E., Der Zellaufbau des Hypothalamus beim Hund, Z. Ges. Neurol. Psychiat., 120 (1929) 157-177.
8 H6kfelt, T., Fuxe, K., Johansson, O., Jeffcoate, S. and White N., Distribution of thyrotropin releasing hormone (TRH) in the central nervous system as revealed with immunohistochemistry, Eur. J. Pharmacol., 34 (1975) 389-392.
9 H6kfelt, T., Tsuro, Y., Ulfhake, B., Cullheim, S., Arvidsson, U., Foster, G.A., Schultzberg, M., Stalling, M., Arborelius, L., Freedman, J., Post, C. and Visser, T., Distribution of TRH- like immunoreactivity with special reference to coexistence with other neuroactive compounds, Ann. N.Y. Acad. Sci., 553 (1989) 76-105.
10 Johansson, O. and H6kfelt, T., Thyrotropin releasing hormone, somatostatin and enkephalin: distribution studies using immu- nohistochemical techniques, J. Histochem. Cytochem., 28 (1980) 364-366.
I1 Jordan, D., Kopp, N., Latouche, J., Crumeyrolle, M., Pas- quier, B., Bouvier, R., Borson-Chazot, E, Najimi, M., Fillion, G. and Haour, E, Localization and biochemical characteriza- tion of [3H]MeTRH binding sites in the human hippocampus, Ann. N.Y. Acad. Sci., 553 (1989) 521-523.
12 Kreider, M.S., Engber, T.M., Nilaver, G., Zimmermann, E.A. and Winokur, A., Immunohistochemical localization of TRH in rat CNS: comparison with RIA studies, Peptides, 6 (1985) 997- 1000.
13 Kubek, M.J., Lorincz, A. and Wilber, J.E, The identification of thyrotropin releasing hormone (TRH) in hypothalamic and extrahypothalamic loci of the human nervous system, Brain Re-
MM Medial Mammillary nucleus MP Medial Preoptic area MT Medial Tuberal nucleus OC Optic Chiasma ON Optic Nerve OT Optic Tract PA Posterior Hypothalamic Area PE Periventricular nucleus PV Paraventricular nucleus SN Supraoptic Nucleus VM Ventromedial Nucleus V Third ventricle
75
search, 126 (1977) 196-200. 14 Leroux, P., Gonzal~s, B.J., Laquerri~re, A., Bodenant, C. and
Vaudry, H., Autoradiographic study of somatostatin receptors in the rat hypothalamus: validation of GTP-induced desatura- tion procedure, Neuroendocrinology, 57 (1988) 533-544.
15 Lipton, J.M. and Glyn, J.R., Central administration of peptides alters thermoregutation in the rabbit, Peptides, 1 (1980) 15-18.
16 Lehan, R.M. and Jackson, I.M.D., Immunohistoehemical lo- calization of thyrotropin releasing hormone in the rat hypothal- amus and pituitary, Endocrinology, 111 (1982) 55-65.
17 Lin, M.T. and Yang, C.P., Effects of thyrotropin releasing hor- mone on thermoregulatory responses and hypothalamic neu- ronal activity in rats, Ann. N.Y. Acad. Sci., 553 (1989) 579-581.
18 Manaker, S., Winokur, A., Rhodes, C.H. and Rainbow, T.C., Autoradiographic localization of thyrotropin releasing hormone receptors in the rat central nervous system, J. Neurosci., 5 (1985) 167-174.
19 Manaker, S., Winokur, A., Rost6ne, W.H. and Rainbow, T.C., Autoradiographic localization of thyrotropin releasing hormone (TRH) receptors in the human spinal cord, Neurology, 35 (1985) 328-332.
20 Manaker, S., Eichan, A., Winokur, A., Rhodes, C.H. and Rainbow, T.C., Autoradiographic localization of thyrotropin re- leasing hormone receptors in the human brain, Neurology, 36 (1986) 641-648.
21 Mantyh, EW. and Hunt, S.P., Thyrotropin releasing hormone (TRH) receptors localization by light microscopic autoradiogra- phy in rat brain using 3H(3Me-His2) TRH as the radioligand, J. Neurosci., 5 (1985) 551-561.
22 Miyamoto, M., Yamazaki, N., Nagaoka, A. and Nagawa, Y., Effects of TRH and its analogue, DN-1417, on memory impair- ment in animal models, Ann. N.Y. Acad. Sci., 553 (1989) 508- 510.
23 Najimi, M., Chigr, E, Leduque, E, Jordan, D., Charnay, Y., Chayvialle, J.A., Tohyama, M. and Kopp, N., Immunohis- tochemical distribution of somatostatin in the infant hypothal- amus, Brain Research, 483 (1989) 205-221.
24 Najimi, M., Chigr, E, Jordan, D., Leduque, P., Bloch, B., Re- baud, R., Tommasi, M. and Kopp, N., Anatomical distribution of LHRH immunoreactive neurons in the human infant hypo- thalamus, Brain Research, 516 (1990) 280-291.
25 Najimi, M., Jordan, D., Chigr, F., Champier, J., Kopp, N., Slama, A., Bertherat, J., Videau, C. and Epelbaum, J., Re- gional distribution of somatostatin binding sites in the human hypothalamus. A quantitative autoradiographic study, Neuro- science, 40 (1991) 321-335.
26 Nishiyama, T., Heike, Y., Matsuzaki, T., Kawano, H., Daikoku, S. and Suzuki, M., Immunoreactive TRH-containing neurons in the rat hypothalamus, Biomed. Res. Suppl., 4 (1983) 65-74.
27 Nishiyama, T., Kawano, H., Tsurio, Y., Maegawa, M., Hisano, S., Adachi, T., Daikoku, S. and Suzuki, M., Hypothalamic thy- rotropin releasing hormone TRH-containing neurons involved in the hypothalamic-hypophysial-thyroid axis. Light microscopic immunohisto-chemistry, Brain Research, 345 (1985) 205-218.
28 Palacios, J.M., Camps, M., Cortes, R. and Probst, A., Map- ping dopamine receptors in the human brain, J. Neural.

76
Transm., 27 (1988) 227-235. 29 Parker, R.B. and Waud, D.R., Pharmacological estimation of
drug receptor dissociation constants. I. Agonists, J. Pharrnacol. Exp. Ther., 177 (1971) 1-12.
30 Parker, C.R. and Porter, J.C., Regional localization and sub- cellular compartmentalization of thyrotropin-releasing hormone in adult human brain, J. Neurochem., 40 (1983) 1614-1622.
31 Parker, C.R. and Capdevila, A., Thyrotropin releasing hor- mone (TRH) binding sites in the adult human brain: localiza- tion in the adult human brain: an autoradiographic study, Neu- roscience, 18 (1984) 329-346.
32 Reubi, J.C., Cortes, R., Maurer, R. and Probst, A., Distribu- tion of somatostatin receptors in the human brain: an autora- diographic study, Neuroscience, 18 (1986) 329-346.
33 Reubi, J.C., Palacios, J.M. and Maurer, R., Specific luteinizing hormone-releasing hormone receptor binding sites in hippocam- pus and pituitary: an autoradiographic study, Neuroscience, 21 (1987) 847-856.
34 Rose, J.E., The cell structure of the mammillary body in the mammals and in man, J. Anat., 74 (1940) 91-115.
35 Rost~ne, W.H., Morgat, J.L., Dussaillant, M., Rainbow, T.C., Sarrieau, A. and Rosselin, G., In vitro biochemical character- ization and autoradiographic distribution of 3H-thyrotropin re- leasing hormone binding sites in the rat brain sections, Neu- roendocrinology, 39 (1984) 81-86.
36 Satinoff, E., Liran, J. and Clapman, R., Aberrations of circa- dian body temperature rhythms in rats with preoptic lesions,
Am. J. Physiol., 242 (1982) 352-357. 37 Sharif, N.A., Quantitative autoradiography of TRH receptors
in discrete brain regions of different mammalian species, Ann. N.~( Acad. Sci., 553 (1989) 147-175.
38 Simasko, S.M. and Horita, A., Characterization and distribu- tion of 3H-(3Me Hise)-thyrotropin releasing hormone receptors in the rat brain, Life Sci., 30 (1982) 1793-1799.
39 Swaab, D.F. and Uylings, H.B.M., Potentialities and pitfalls in the use of human brain material in molecular neuroanatomy. In M. Van Leeuwen, S. Buijs, B.M. Pool and D. Pach (Eds.), Molecular Neuroanatomy, 1988 pp. 403-416.
40 Szymusiak, R., De Mory, A., Kittrel, E.M.W. and Satinoff, E., Diurnal changes in thermoregulatory in rats with medial preop- tic lesions, Am. J. Physiol., 249 (1981) 219-227.
41 Taylor, R.L. and Burt, D.R., Properties of 3H (3 Me-His z) TRH binding to apparent TRH receptors in the sheep central nervous system, Brain Research, 218 (1981) 207-217.
42 Taylor, R.L. and Burt, D.R., Species differences in the brain regional distribution of receptor binding for thyrotropin releas- ing hormone, J. Neurochem., 38 (1982) 1649-1656.
43 Winokur, A. and Utiger, R.D., Thyrotropin releasing hor- mone: regional distribution in rat brain, Science, 185 (1974) 265 -267.
44 Zezula, J., Cortes, R., Probst, A. and Palacios, J.M., Benzo- diazepine receptor sites in the human brain: autoradiographic mapping, Neuroscience, 25 (1988) 771-795.




















