
Detection by 32P-postlabelling of 8-oxo-7,8-dihydro-2′-deoxyguanosine in DNA as biomarker of...
12
Mutation Research 564 (2004) 9–20 Detection by 32 P-postlabelling of 8-oxo-7,8-dihydro-2 -deoxyguanosine in DNA as biomarker of microcystin-LR- and nodularin-induced DNA damage in vitro in primary cultured rat hepatocytes and in vivo in rat liver Imed Maatouk a , Noureddine Boua¨ ıcha b,∗ , Marie Jos´ e Plessis a , Franc ¸ois P´ erin a a Laboratoire Raymond Latarjet, Institut Curie-Section Recherche, Bˆ atiment 112, Centre Universitaire, 91405 Orsay Cedex, France b Laboratoire de Sant´ e Publique-Environnement, EA3542, Universit´ e Paris-Sud, UFR de Pharmacie, 5 rue J.B. Cl´ ement, 92296 Ch ˆ atenay-Malabry, France Received 17 February 2004; received in revised form 1 June 2004; accepted 25 June 2004 Available online 15 September 2004 Abstract Microcystin-LR (MCYST-LR) and nodularin (NOD) produced by cyanobacteria are potent and specific hepatotoxins. The induction of free-radical formation, reduction of glutathione levels and induction of DNA damage are three major events found in rat hepatocytes treated with these hepatotoxins. However, the mechanism of MCYST-LR- and NOD-mediated induction of oxidative DNA damage has not been fully elucidated. The objective of this study was to determine whether MCYST-LR and NOD increase the formation of a DNA oxidative damage marker such as 8-oxo-7,8-dihydro-2 -deoxyguanosine (8-oxo-dG) in vitro in primary rat hepatocytes and in vivo in rat liver cells. Rat hepatocytes were exposed to MCYST-LR or NOD at low doses (2 and 10 ng/ml), at which there is no evidence of morphologically apparent cytotoxic effects, as well as an induced dose- and time-dependent formation of 8-oxo-dG. Moreover, MCYST-LR treatment of rats (50 g/kg, ip) resulted in a significant increase of 8-oxo-dG in liver DNA, at 24 h after treatment before decreasing at 48 h. However, NOD-induced DNA damage was increased both at 24 and 48h, in contrast to the MCYST-LR-induced effect. The effects on this oxidative DNA damage marker indicates that MCYST-LR and NOD do evoke oxidative stress, which may contribute, at least in part, to their liver toxicity and carcinogenicity during long-term exposure. © 2004 Elsevier B.V. All rights reserved. Keywords: Microcystin-LR; Nodularin; 8-Oxo-7,8-dihydro-2 -deoxyguanosine; 32 P postlabelling Abbreviations: MCYST-LR, microcystin-LR; NOD, nodularin; DNA, deoxyribonucleic acid; dAp, 2 -deoxyadenosine 3 -monophosphate; dGp, 2 -deoxyguanosine 3 -monophosphate; 8-oxo-dG, 8-oxo-7,8-dihydro-2 -deoxyguanosine; ROS, reactive oxygen species; GSH, reduced glutathione; PP-2A and PP-3, protein phosphatases type 1, 2A, 3; PEI, polyethyleneimine; TLC, thin-layer chromatography; RAL, relative adduct levels; dNps, normal nucleotides ∗ Corresponding author. Tel.: +33 1 468 35891; fax: +33 1 468 35732. E-mail address: [email protected] (N. Boua¨ ıcha). 1383-5718/$ – see front matter © 2004 Elsevier B.V. All rights reserved. doi:10.1016/j.mrgentox.2004.06.010
Transcript of Detection by 32P-postlabelling of 8-oxo-7,8-dihydro-2′-deoxyguanosine in DNA as biomarker of...

Mutation Research 564 (2004) 9–20
Detection by32P-postlabelling of8-oxo-7,8-dihydro-2′-deoxyguanosine in DNA as biomarker of
microcystin-LR- and nodularin-induced DNA damage in vitro inprimary cultured rat hepatocytes and in vivo in rat liver
Imed Maatouka, Noureddine Bouaıchab,∗, Marie Jose Plessisa, Francois Perina
a Laboratoire Raymond Latarjet, Institut Curie-Section Recherche, Bˆatiment 112, Centre Universitaire, 91405 Orsay Cedex, Franceb Laboratoire de Sant´e Publique-Environnement, EA3542, Universit´e Paris-Sud, UFR de Pharmacie,
5 rue J.B. Clement, 92296 Chˆatenay-Malabry, France
Received 17 February 2004; received in revised form 1 June 2004; accepted 25 June 2004Available online 15 September 2004
Abstract
Microcystin-LR (MCYST-LR) and nodularin (NOD) produced by cyanobacteria are potent and specific hepatotoxins. Theinduction of free-radical formation, reduction of glutathione levels and induction of DNA damage are three major events foundin rat hepatocytes treated with these hepatotoxins. However, the mechanism of MCYST-LR- and NOD-mediated induction of
LR andnat low
ced dose-tage wasmarker
ity and
;uced, relative
oxidative DNA damage has not been fully elucidated. The objective of this study was to determine whether MCYST-NOD increase the formation of a DNA oxidative damage marker such as 8-oxo-7,8-dihydro-2′-deoxyguanosine (8-oxo-dG) ivitro in primary rat hepatocytes and in vivo in rat liver cells. Rat hepatocytes were exposed to MCYST-LR or NODdoses (2 and 10 ng/ml), at which there is no evidence of morphologically apparent cytotoxic effects, as well as an induand time-dependent formation of 8-oxo-dG. Moreover, MCYST-LR treatment of rats (50�g/kg, ip) resulted in a significanincrease of 8-oxo-dG in liver DNA, at 24 h after treatment before decreasing at 48 h. However, NOD-induced DNA damincreased both at 24 and 48 h, in contrast to the MCYST-LR-induced effect. The effects on this oxidative DNA damageindicates that MCYST-LR and NOD do evoke oxidative stress, which may contribute, at least in part, to their liver toxiccarcinogenicity during long-term exposure.© 2004 Elsevier B.V. All rights reserved.
Keywords:Microcystin-LR; Nodularin; 8-Oxo-7,8-dihydro-2′-deoxyguanosine;32P postlabelling
Abbreviations: MCYST-LR, microcystin-LR; NOD, nodularin; DNA, deoxyribonucleic acid; dAp, 2′-deoxyadenosine 3′-monophosphatedGp, 2′-deoxyguanosine 3′-monophosphate; 8-oxo-dG, 8-oxo-7,8-dihydro-2′-deoxyguanosine; ROS, reactive oxygen species; GSH, redglutathione; PP-2A and PP-3, protein phosphatases type 1, 2A, 3; PEI, polyethyleneimine; TLC, thin-layer chromatography; RALadduct levels; dNps, normal nucleotides∗ Corresponding author. Tel.: +33 1 468 35891; fax: +33 1 468 35732.E-mail address:[email protected] (N. Bouaıcha).
1383-5718/$ – see front matter © 2004 Elsevier B.V. All rights reserved.doi:10.1016/j.mrgentox.2004.06.010

10 I. Maatouk et al. / Mutation Research 564 (2004) 9–20
1. Introduction
Evidence of adverse effects on human health fromcyclic peptide microcystins and nodularin (NOD)results from a study of human populations showingsymptoms of poisoning or injury attributed to thepresence of these cyanotoxins in drinking water orother sources of water[1]. Microcystin-LR (MCYST-LR) and nodularin (NOD) with molecular weights of994 and 824, respectively, are specific liver poisonsin mammals. Following acute exposure at high doses,they cause death from liver haemorrhage[2], and theymay promote tumours following chronic exposureat low doses[3]. In addition, NOD is considered asa de novo liver carcinogen with both initiating andtumour-promoting activities, whereas MCYST-LRis a liver tumour promoter without initiating activity[4,5]. These health hazards have led the WorldHealth Organization (WHO) to establish a provisionalguideline value for MCYST-LR in drinking waterof 1�g/l [6]. Nevertheless, the damage provoked inhumans by repeated consumption of low doses ofmicrocystins and nodularins is probably more frequentthan acute intoxication. Epidemiological studies incertain areas of China have indicated that peopletaking their drinking water from ponds and ditchescontaminated with cyanobacterial blooms producingmicrocystins have a higher incidence of primary livercancer (PLC) than control populations. However, the
ee
rred
srn-
edxi-ge-A
an
R-edones
and Ong[17], reported that MCYST-LR-induced ox-idative stress plays an important role in mitochondrialchanges, reflected by the onset of mitochondrial perme-ability transition and the loss of mitochondrial mem-brane potential. However, the possible implication ofcyanobacterial hepatotoxins in oxidative DNA dam-age remains largely unknown. Recently, however, Ze-gura et al.[12] reported that MCYST-LR-induced DNAstrand breaks in HepG2 human hepatoma cells couldbe intermediates produced during the cellular repairof toxin-induced oxidative DNA damage. In this con-text, various biomarkers can be used to demonstrateoxidative DNA damage induced by reactive oxygenspecies (ROS). The most common of these is 8-oxo-7,8-dihydro-2′-deoxyguanosine (8-oxo-dG), resultingfrom the addition of a hydroxyl radical to deoxyguano-sine. The presence of 8-oxo-dG in DNA causes mis-reading and contributes to the oxidative mutationalload by inducing G:C to T:A transversions[18]. Thereare a number of analytical techniques used for mea-suring 8-oxo-dG including gas chromatography–massspectrometry (GC–MS)[19], high performance liq-uid chromatography with electrochemical detection(HPLC–ECD)[20], 32P-postlabelling[21,22]and var-ious immuno-assays[23].
The present study was carried out to investigate, forthe first time, whether MCYST-LR and NOD induceoxidative DNA damage such as 8-oxo-dG in vitro, inprimary cultured rat hepatocytes as well as in vivo, in ratliver. The 8-oxo-dG content was measured by the sen-s blesu ibleg erialh
2
2
b-t on,F 7-2 reds cro-c otatoan ne
itive 32P-postlabelling assay. This approach enas to provide additional information on the possenotoxic and carcinogenic potential of cyanobactepatotoxins.
. Materials and methods
.1. Chemicals and reagents
Nodularin (CAS no. 118399-22-7) was oained from Calbiochem-Novabiochem Co. (Meudrance). Microcystin-LR (CAS no. 101043-3), dimethyl sulfoxide (DMSO), phosphate-buffealine (PBS), RNaseA, RNaseTl, protease, mioccal nuclease, spleen phosphodiesterase, ppyrase, 8-hydroxyquinoline,N-tert-butyl-�-phenylitrone, trifluororacetic acid (TFA), polyethyleneimi
presence of aflatoxin Bl and hepatitis B virus in thdrinking water could explain the higher rate of thobserved PLC in China[7]. Thus, the evidence focarcinogenicity of these hepatotoxins is consideinadequate in humans and limited in animals[8].
Although, the hepatotoxicity of microcystins habeen very well documented, many aspects conceing these toxins are insufficiently known and indecontroversial, notably those relative to their genotocity. Nevertheless, microcystin-induced DNA damaand fragmentation in different in vitro or in vivo studies might be a consequence of endonucleolytic DNdegradation associated with cytotoxicity, rather ththe result of a direct toxin–DNA interaction[9–14].Furthermore, recent studies found that MCYST-Land NOD-induced oxidative stress may be involvin the pathogenesis of their hepatotoxicity, in additito or as a result of inhibition of protein phosphatas(type PP1, PP2A and PP3)[15–17]. Recently, Ding

I. Maatouk et al. / Mutation Research 564 (2004) 9–20 11
(PEI), dAp, dGp and calf thymus DNA were pur-chased from Sigma–Aldrich (Saint Quentin Fallavier,France). William’s E medium, foetal calf serum (FCS),l-glutamine, penicillin/streptomycin, and gentamicinwere from GibcoBRL (Cergy Pontoise, France). Hy-drazine hydrate (98%) was from Lancaster (England).[32P]orthophosphate (carrier free; 500 mCi/ml; sp. act.4–5000 Ci/mmol) for preparation of [�-32P] ATP, waspurchased from ICN Radiochemicals (Orsay, France)and T4 polynucleotide kinase from Appligene On-cor (Illkirch, France). Cellulose MN 301 A fromMacherey–Nagel (Germany) was used in a mixturewith PEI solution and ultra pure water to prepare PEI-cellulose TLC plates in our laboratory. Solvents andstandard solutions were prepared with Milli-Q water(Millipore, Saint Quentin Yvelines, France).
2.2. Synthesis of 8-oxo-dG
8-Oxo-7,8-dihydro-2′-deoxyguanosine (8-oxo-dG)was synthesized as described by Kasai and Nishimura[24], with the following modifications. dGp (10 mg)was dissolved in 10 ml of 0.1 M phosphate buffer(pH 6.8) containing 14 mM ascorbic acid, 6.5 mMethylenediamine tetra acetic acid and 1.3 mM FeSO4.The mixture was bubbled with oxygen at 37◦C for 4 h.Water was removed by lyophilization. The residue wassolubilized with 500�l of water, applied to a SephadexLH20 column (20 mm× 280 mm) and eluted withwater; 25 fractions (2 ml/fraction) were collected.A LCe sil 5O di6 reda erep zedw byH ing8 duew dt salt.T byH ro-2 ker( -2d ,8-d ve
identical ultraviolet spectra, but different retentiontimes.
2.3. Animals
Male Sprague–Dawley rats (40 days old; CharlesRiver, Saint Aubin-Les-Elbeuf, France) were suppliedwith food and water ad libitum and were allowed toacclimate in the animal facility. Rats were then anaes-thetised with an i.p. injection of sodium pentobarbital(60 mg/kg body weight) and sacrificed after 10 min,prior to isolation of hepatocytes.
2.4. Isolation and treatment of rat hepatocytes
Rat hepatocytes were isolated from rats weighing200–250 g by a modified two-step collagenase perfu-sion method[25]. Viability of hepatocytes exceeded85% as determined by trypan-blue exclusion. The cellswere plated on a collagen-coated (1.27�g/cm2), 10 cmdiameter plastic dish at a density of 200,000 cells/ml(2 million cells per dish) in William’s E medium sup-plemented with antibiotics (mixture of penicillin andstreptomycin (1000 U/ml), gentamicin (10 mg/ml),l-glutamine (200 mM)), and foetal calf serum (FCS).Incubation was for 3 h at 37◦C in a 5% CO2 incubatorfor adhesion of the cells. Unattached cells were thenremoved and the medium was replaced with freshmedium containing all supplements as above, but only2% FCS and two concentrations (2 and 10 ng/ml)o ins ide( l ofm )f atedo fort ellsw BS)t erals
2
int alsw LRoa al.
liquots of each fraction were then analyzed by HPquipped with a diode-array detector on an InertDS-2 column (2 mm× 150 mm, Chrompack), elute
socratically at 0.5 ml/min with 10 mM NaH2PO4 (pH.0) containing 2% methanol. UV detection occurt 293 nm. Fractions containing 8-oxo-dG wooled and lyophilized. The residue was solubiliith 200�l of water and 8-oxo-dG was purifiedPLC as indicated above. Again, fractions contain-oxo-dG were pooled and lyophilized. The resias solubilized with 500�l of water and then applie
o Sephadex LH20, as described above to removehe concentration of 8-oxo-dG was determinedPLC as indicated above using 8-oxo-7,8-dihyd′-deoxyguanosine 5′-monophosphate as a marpurchased from Sigma). 8-Oxo-7,8-dihydro′-eoxyguanosine 3′-monophosphate and 8-oxo-7ihydro-2′-deoxyguanosine 5′-monophosphate ha
f MCYST-LR and NOD. Different cyanotoxolutions were mixed (1:1) in dimethyl sulphoxDMSO). Cells were exposed to 2 and 10 ng/microcystin-LR (MCYST-LR) or nodularin (NOD
or 3, 6 and 24 h. The control hepatocytes were trenly with DMSO (0.1%) at the same volume as
oxin-treatment. At the end of each incubation, cere washed twice in phosphate-buffered saline (P
o remove detached dead cells, divided in sevamples and stored at−80◦C, until DNA isolation.
.5. Animal treatment with toxins
After 4–6 days of acclimation, rats were dividedhree experimental groups (four per group). Animere given an intraperitoneal injection of MCYST-r NOD at 50�g/kg body weight (LD50 in the rat isbout 120�g/kg body weight according to Miura et

12 I. Maatouk et al. / Mutation Research 564 (2004) 9–20
[26]) dissolved in sterile saline solution. The group ofcontrol rats received only the sterile saline solution atthe same volume as for the toxin-treated group. Tworats from each group were sacrificed after 24 and 48 h,respectively. Livers were quickly removed, minced andstored frozen at−80◦C, until DNA isolation. Housingand treatment of rats were in accordance with nationaland institutional guidelines.
2.6. DNA isolation
In the absence of fluorescent light (cool white), DNAwas isolated from rat hepatocytes or from 200–300 mgof frozen liver as described by Arif et al.[27], withminor modifications. After RNA and protein removal,DNA was extracted twice with an equal volume of coldsolution of Sevag (chloroform:isoamyl alcohol, 24:1),recovered from the aqueous phase by adding 0.1 vol.of 5 M Nal and 1 vol. of isopropanol and dissolved in0.01× SSC (0.15 M sodium chloride, 0.015 M sodiumcitrate, 0.1 mM EDTA). DNA was then re-precipitatedwith 0.1 vol. of 5 M NaCl and 1 vol. of chilled ethanol.After centrifugation (10,000 rpm, 15 min), DNA wasre-suspended in an appropriate volume of 0.01× SSCsolution. 8-Hydroxyquinoline (6.8 mM) andN-tert-butyl-�-phenyl nitrone (500�M) were included duringtissue homogenization, solvent extractions and DNAprecipitation to avoid artefactual 8-oxo-dG generation.DNA content was determined by spectrophotometricanalysis assuming that 1 A260 unit equals 50�g DNA;a s am
2
u-c ith0p and18 d asdt tor ichc
thes rmalp
2.8. 32P-postlabelling analysis
Using enriched synthesized standard 8-oxo-7,8-dihydro-2′-deoxyguanosine 3′-monophosphate, the32P-postlabelling assay reported in the literature[21,22]was adapted, with modifications for 8-oxo-dGanalysis without nuclease P1 enrichment. The invitro and/or in vivo DNA digests (either untreatedor treated with TFA and hydrazine) were labelled ina reaction mixture containing kinase buffer (0.2 Mbicine-NaOH, 0.1 M MgCl2, 0.1 M dithiothreitol,0.01 M spermidine, pH 9.6), freshly prepared [�-32P]ATP (20�Ci or 80�Ci for DNA or modified nu-cleotides, respectively), 10 mM bicine-NaOH, pH 9.6and T4 polynucleotide kinase (0.5 U/20�Ci [�-32P]ATP). A volume of 2.5�l of 10 mM bicine-NaOH wasadded to adjust the pH for the labelling step. Afterincubation at 37◦C for 60 min, 1�l solution of 20 mUpotato apyrase was supplemented to remove excessATP and the incubation was continued for 30 min.
2.9. Thin layer chromatography andco-chromatograhpy
Separation of p(8OH)dGp, called 8-oxo-dG, fromlabelled samples was performed by three-directionalanion-exchange thin-layer chromatography (TLC) onpre-washed home made polyethyleneimine-cellulose(PEI) thin-layer plates. After application of enriched-labelled DNA or mononucleotides, the excess of32Pi( entw iums opo di-l liedu atelyo les,i e toit con-t potY areac em reshs ot-t 8 Ma mt 5 M
n A260:A280 ratio of 1.70–1.90 was considered, aeasure of acceptable purity of DNA.
.7. DNA digestion and enrichment of 8-oxo-dG
DNA (4 �g) was hydrolyzed to deoxyribonleoside 3′-monophosphates by incubation w.5 U micrococcal nuclease and 4.8�g of spleenhosphodiesterase in 30 mM sodium succinate0 mM calcium chloride (pH 6.0) at 37◦C for 4 h. The-oxo-dG content in DNA digest was then enricheescribed by Devanaboyina and Gupta[21], with 90%
rifluoroacetic acid (TFA) and hydrazine in orderemove a maximum of unmodified nucleotides whould be oxidized during postlabelling.
An 8-oxo-dG standard solution was treated iname manner as above, alone or with addition of nourine nucleotides (dAp, dGp).
inorganic phosphate) was removed by developmith 0.05 M sodium phosphate and 0.28 M ammonulfate (pH 6.7) in the first direction (D1) to the tf the 18 cm sheet. Two microliters of a suitable
ution of synthetic standard p(8OH)dGp was appnder these conditions, in the same plate separr in co-chromatography with enriched DNA samp
n order to detect its migration zone after exposurntensifying-screen enhanced X-ray film.Fig. 1showshat 8-oxo-dG migrates between the non-specificaminant spot X (situated above the origin) and s, which represents also a non-specific spot. Theontaining 8-oxo-dG (1 cm× 1.5 cm) was cut out, thaterial was transferred by magnetic contact to a f
heet (12 cm× 18 cm) and resolved in D2 again (bom to top) with 0.05 M sodium phosphate and 0.2mmonium sulfate (pH 6.7). After cutting 12 cm fro
he bottom, the plate was further developed in 1.2
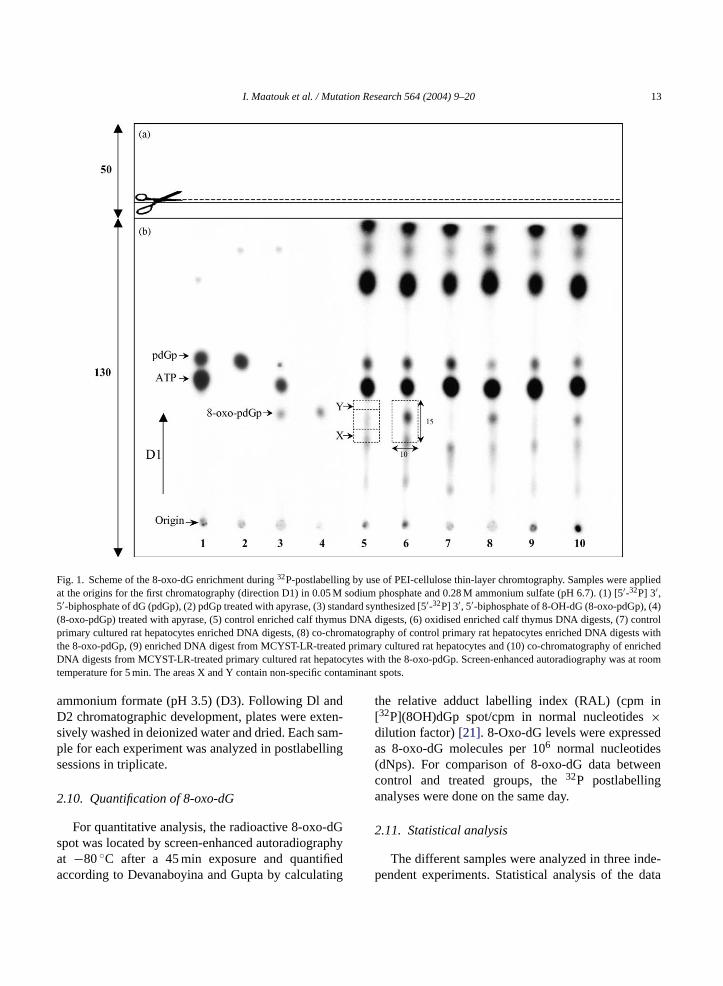
I. Maatouk et al. / Mutation Research 564 (2004) 9–20 13
Fig. 1. Scheme of the 8-oxo-dG enrichment during32P-postlabelling by use of PEI-cellulose thin-layer chromtography. Samples were appliedat the origins for the first chromatography (direction D1) in 0.05 M sodium phosphate and 0.28 M ammonium sulfate (pH 6.7). (1) [5′-32P] 3′,5′-biphosphate of dG (pdGp), (2) pdGp treated with apyrase, (3) standard synthesized [5′-32P] 3′, 5′-biphosphate of 8-OH-dG (8-oxo-pdGp), (4)(8-oxo-pdGp) treated with apyrase, (5) control enriched calf thymus DNA digests, (6) oxidised enriched calf thymus DNA digests, (7) controlprimary cultured rat hepatocytes enriched DNA digests, (8) co-chromatography of control primary rat hepatocytes enriched DNA digests withthe 8-oxo-pdGp, (9) enriched DNA digest from MCYST-LR-treated primary cultured rat hepatocytes and (10) co-chromatography of enrichedDNA digests from MCYST-LR-treated primary cultured rat hepatocytes with the 8-oxo-pdGp. Screen-enhanced autoradiography was at roomtemperature for 5 min. The areas X and Y contain non-specific contaminant spots.
ammonium formate (pH 3.5) (D3). Following Dl andD2 chromatographic development, plates were exten-sively washed in deionized water and dried. Each sam-ple for each experiment was analyzed in postlabellingsessions in triplicate.
2.10. Quantification of 8-oxo-dG
For quantitative analysis, the radioactive 8-oxo-dGspot was located by screen-enhanced autoradiographyat −80◦C after a 45 min exposure and quantifiedaccording to Devanaboyina and Gupta by calculating
the relative adduct labelling index (RAL) (cpm in[32P](8OH)dGp spot/cpm in normal nucleotides×dilution factor)[21]. 8-Oxo-dG levels were expressedas 8-oxo-dG molecules per 106 normal nucleotides(dNps). For comparison of 8-oxo-dG data betweencontrol and treated groups, the32P postlabellinganalyses were done on the same day.
2.11. Statistical analysis
The different samples were analyzed in three inde-pendent experiments. Statistical analysis of the data

14 I. Maatouk et al. / Mutation Research 564 (2004) 9–20
(means± S.D.) was performed by non-parametricMann–WhitneyU-test. Statistical significance for com-parison of rat hepatocytes or animals treated with cor-responding controls was indicated by theP-values:∗P< 0.05;∗∗P < 0.001.
3. Results
3.1. Evaluation of 8-oxo-dG in primary culturedrat hepatocytes
The determination of oxidative DNA damage in pri-mary cultured rat hepatocytes treated with MCYST-LR
F oxo-dG ytesD88Db
or NOD was achieved by the adaptation to sensitive32P-postlabelling assays in order to detect and quan-tify the 8-oxo-dG contents. The presence of 8-oxo-dGin treated and untreated cells is illustrated inFig. 2. Theauto-radiograms presented in this figure show, in addi-tion to the 8-oxo-dG ([32P](8OH)dGp) spot, the pres-ence of non-specific spots X and Y that could containunspecific contaminants, which could be either artefac-tual or indigenous. The chemical enrichment of DNAwith TFA and hydrazine, and the action of T4 polynu-cleotide kinase in postlabelling procedure could con-tribute to their formation. Indeed, the polynucleotidekinase can label non-nucleic acid components, such assome hydroxylated metabolites[28].
ig. 2. Autoradiograms of PEI-cellulose TLC maps of the [5′-32P] 8-
NA induced after 3 h treatment, (A) DNA of control cells, (B) contro-oxo-dG, (C) 10 ng/ml NOD exposure and (D) 10 ng/ml MCYST-LR e-oxo-dG spot was detected by screen-enhanced autoradiography us2, 0.05 M sodium phosphate and 0.28 M ammonium sulfate (pH 6.7)y the arrow.(3′, 5′-biphosphate of 8-OH-dG) in primary cultured rat hepatoc
l DNA in co-chromatography with the enriched synthesized standardxposure. Enriched 50 ng of digested DNA was chromatographed and theing X-ray films at−80◦C for 45 min. Chromatographic conditions: D1 and; D3, 1.25 M ammonium formate, pH 3.5. The 8-oxo-dG spot is indicated

I. Maatouk et al. / Mutation Research 564 (2004) 9–20 15
Fig. 3. Kinetics of 8-oxo-dG formation in primary cultured rat hepatocytes treated with MCYST-LR and NOD. Cell cultures were treated withdifferent doses of MCYST-LR or NOD (2 and 10 ng/ml) for 3, 6 and 24 h. Data shows the number of modified bases (8-oxo-dG) per 105 DNAbases. Values are means± S.D. of three independent experiments. Significant difference from the control is designated by∗P < 0.05,∗∗P <0.001 (Mann–WhitneyU-test).
Kinetic studies revealed a dose-dependent 8-oxo-dGformation for 3 and 6 h exposure times in both MCTSY-LR- and NOD-treated primary cultured rat hepatocytes,when compared with untreated cells (Fig. 3). Treat-ment with high dose of NOD (10 ng/ml) significantlyenhanced (P < 0.001,U-test), the 8-oxo-dG level to17.05± 1.131 8-oxo-dG per 106 normal nucleotides af-ter 3 h, approximately 3.5 and 1.5 higher than the levelsnoted in untreated cells and in cells treated with the lowdose of NOD, respectively. Treatment with MCYST-LR, both at high and low dose (2 and 10 ng/ml) sig-nificantly increased 8-oxo-dG formation, which washigher in particular with the high toxin dose. The 8-oxo-dG levels went on increasing during the 6 h incubationperiod in the presence of MCYST-LR, attaining a valueof 16.00± 1.29 8-oxo-dG per 106 normal nucleotidesat the high toxin dose. In contrast, DNA oxidation haddecreased after a 6 h exposure to the two concentra-tions of NOD (2 and 10 ng/ml). Thus, NOD seemsto induce 8-oxo-dG formation in primary cultured rathepatocytes more rapidly than MCYST-LR. At 24 h
post-treatment the reduction in 8-oxo-dG content ob-served in the presence of NOD was higher than thatafter MCYST-LR-exposure. Indeed, 8-oxo-dG levelswere noted to be at control levels after a 24 h treatmentwith NOD, in contrast to the MCYST-LR exposure forwhich the 8-oxo-dG values remain significantly higher.
3.2. 8-oxo-dG formation in rat liver DNA
DNA oxidation was investigated in rat liver aftertreatment of the animals with MCYST-LR and NODat 50�g/kg body weight. The results obtained by the32P-postlabelling assay were significantly (P < 0.001)different from those obtained with untreated controlanimals. The auto-radiograms (Fig. 4), show that theprofile of the different radioactive spots varied slightlyfrom that observed previously in in vitro analysis, no-tably regarding the non-specific spot X which couldcomprise more unspecific contaminants.
In kinetic studies (Fig. 5), the 8-oxo-dG levelsappeared twice as high as in controls at 24 h post-
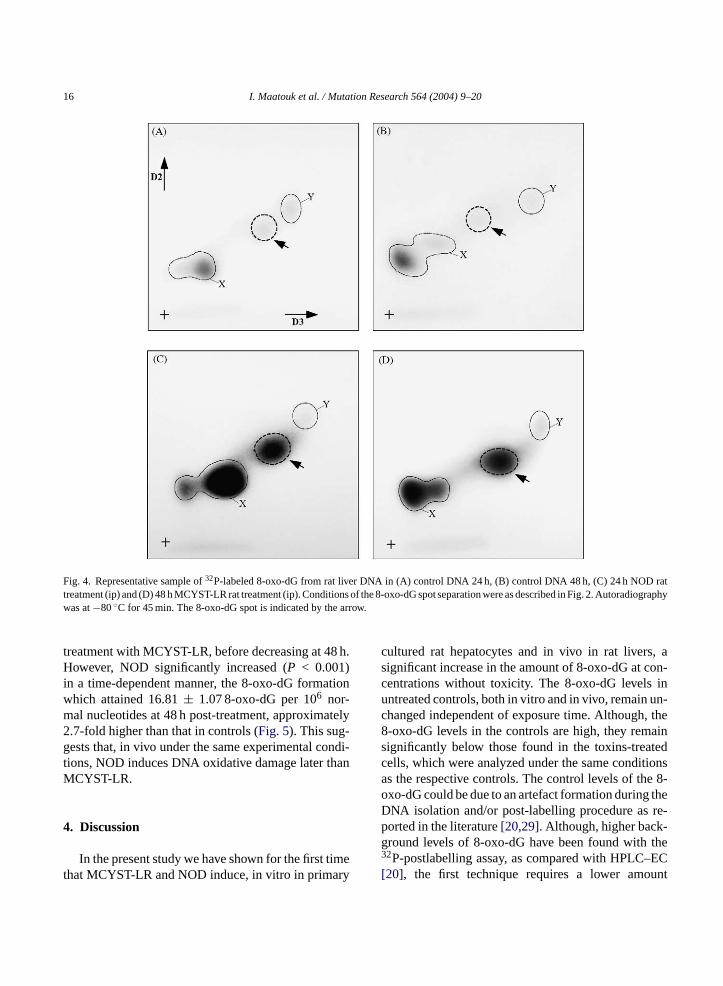
16 I. Maatouk et al. / Mutation Research 564 (2004) 9–20
Fig. 4. Representative sample of32P-labeled 8-oxo-dG from rat liver DNA in (A) control DNA 24 h, (B) control DNA 48 h, (C) 24 h NOD rattreatment (ip) and (D) 48 h MCYST-LR rat treatment (ip). Conditions of the 8-oxo-dG spot separation were as described in Fig. 2. Autoradiographywas at−80◦C for 45 min. The 8-oxo-dG spot is indicated by the arrow.
treatment with MCYST-LR, before decreasing at 48 h.However, NOD significantly increased (P < 0.001)in a time-dependent manner, the 8-oxo-dG formationwhich attained 16.81± 1.07 8-oxo-dG per 106 nor-mal nucleotides at 48 h post-treatment, approximately2.7-fold higher than that in controls (Fig. 5). This sug-gests that, in vivo under the same experimental condi-tions, NOD induces DNA oxidative damage later thanMCYST-LR.
4. Discussion
In the present study we have shown for the first timethat MCYST-LR and NOD induce, in vitro in primary
cultured rat hepatocytes and in vivo in rat livers, asignificant increase in the amount of 8-oxo-dG at con-centrations without toxicity. The 8-oxo-dG levels inuntreated controls, both in vitro and in vivo, remain un-changed independent of exposure time. Although, the8-oxo-dG levels in the controls are high, they remainsignificantly below those found in the toxins-treatedcells, which were analyzed under the same conditionsas the respective controls. The control levels of the 8-oxo-dG could be due to an artefact formation during theDNA isolation and/or post-labelling procedure as re-ported in the literature[20,29]. Although, higher back-ground levels of 8-oxo-dG have been found with the32P-postlabelling assay, as compared with HPLC–EC[20], the first technique requires a lower amount
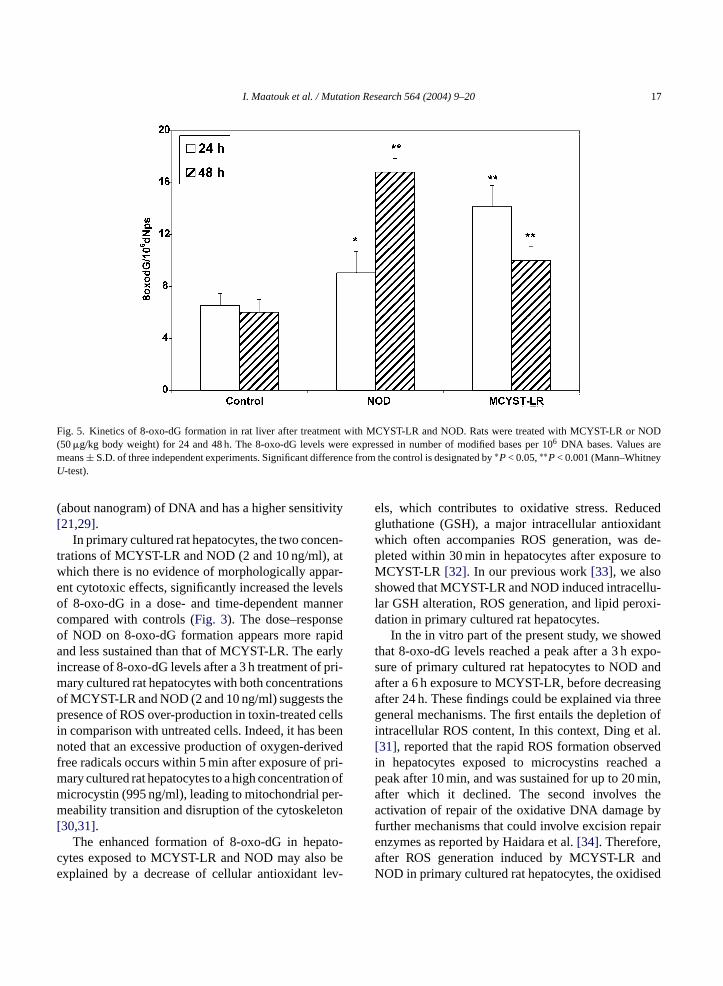
I. Maatouk et al. / Mutation Research 564 (2004) 9–20 17
Fig. 5. Kinetics of 8-oxo-dG formation in rat liver after treatment with MCYST-LR and NOD. Rats were treated with MCYST-LR or NOD(50�g/kg body weight) for 24 and 48 h. The 8-oxo-dG levels were expressed in number of modified bases per 106 DNA bases. Values aremeans± S.D. of three independent experiments. Significant difference from the control is designated by∗P< 0.05,∗∗P< 0.001 (Mann–WhitneyU-test).
(about nanogram) of DNA and has a higher sensitivity[21,29].
In primary cultured rat hepatocytes, the two concen-trations of MCYST-LR and NOD (2 and 10 ng/ml), atwhich there is no evidence of morphologically appar-ent cytotoxic effects, significantly increased the levelsof 8-oxo-dG in a dose- and time-dependent mannercompared with controls (Fig. 3). The dose–responseof NOD on 8-oxo-dG formation appears more rapidand less sustained than that of MCYST-LR. The earlyincrease of 8-oxo-dG levels after a 3 h treatment of pri-mary cultured rat hepatocytes with both concentrationsof MCYST-LR and NOD (2 and 10 ng/ml) suggests thepresence of ROS over-production in toxin-treated cellsin comparison with untreated cells. Indeed, it has beennoted that an excessive production of oxygen-derivedfree radicals occurs within 5 min after exposure of pri-mary cultured rat hepatocytes to a high concentration ofmicrocystin (995 ng/ml), leading to mitochondrial per-meability transition and disruption of the cytoskeleton[30,31].
The enhanced formation of 8-oxo-dG in hepato-cytes exposed to MCYST-LR and NOD may also beexplained by a decrease of cellular antioxidant lev-
els, which contributes to oxidative stress. Reducedgluthatione (GSH), a major intracellular antioxidantwhich often accompanies ROS generation, was de-pleted within 30 min in hepatocytes after exposure toMCYST-LR [32]. In our previous work[33], we alsoshowed that MCYST-LR and NOD induced intracellu-lar GSH alteration, ROS generation, and lipid peroxi-dation in primary cultured rat hepatocytes.
In the in vitro part of the present study, we showedthat 8-oxo-dG levels reached a peak after a 3 h expo-sure of primary cultured rat hepatocytes to NOD andafter a 6 h exposure to MCYST-LR, before decreasingafter 24 h. These findings could be explained via threegeneral mechanisms. The first entails the depletion ofintracellular ROS content, In this context, Ding et al.[31], reported that the rapid ROS formation observedin hepatocytes exposed to microcystins reached apeak after 10 min, and was sustained for up to 20 min,after which it declined. The second involves theactivation of repair of the oxidative DNA damage byfurther mechanisms that could involve excision repairenzymes as reported by Haidara et al.[34]. Therefore,after ROS generation induced by MCYST-LR andNOD in primary cultured rat hepatocytes, the oxidised

18 I. Maatouk et al. / Mutation Research 564 (2004) 9–20
DNA bases were formed rapidly and could then berepaired during prolonged toxin exposure. In thiscase, the repair mechanism seems to be more efficientafter NOD exposure than after MCYST-LR exposure,especially as the early 8-oxo-dG formation was notedin NOD treated cells. Similarly, Zegura et al.[12]found that oxidative DNA damage in the HepG2human hepatoma cell line was induced early afterexposure to MCYST-LR and was subjected to cellularDNA repair. Using the comet assay, these authors pro-vided evidence that MCYST-LR at low concentrations(0.01–1 ng/ml) induced DNA strand breaks, reachinga maximum after 4 h of exposure and then declining.The third mechanism entails the stimulation of DNAsynthesis by cyanotoxins in hepatocytes, which wouldbe expected to dilute the 8-oxo-dG levels. Mellgrenet al. [35] showed that microcystin did not inhibitthe rat hepatocytes DNA synthesis, while Humpageand Falconer[36] reported that MCYST-LR leads tothe formation of mononuclear tetraploid hepatocytes.The formation of polyploid cells was also observedin MCYST-LR-exposed Chinese hamster ovary cells(CHO-Kl), as a result of functional mitotic spindledamage and DNA replication[37].
We studied the MCYST-LR- and NOD-inducedDNA oxidative damage in vivo in order to confirmthe effects of these toxins observed in vitro. The 32P-postlabelling assay adapted as above was applied tomeasure 8-oxo-dG in the liver of rats treated by in-traperitoneal (ip) administration with each toxin at5 sedsW fterN r theM neds
-a vedi OSg nti-o iono ivoh kedbh icalsi rons rotonn used
to demonstrate lipid-derived radicals in rat liversfollowing in vivo exposure to MCYST-LR, eventuallyderived from oxidative lipid alterations[15], whichreflect previous induction of ROS. MCYST-LR-induced DNA damage in vivo in mouse liver hasbeen shown to be associated with a decrease in thepercentage of double-stranded DNA as a result ofstrand breakage[9], which is known to be associatedwith ROS production[38]. Lankoff et al.[16] reportedthat ip treatment of mice with NOD decreased theenzymatic activity of superoxide dismutase, catalaseand glutathione peroxidase, also suggesting that NODcould induce oxidative stress by ROS formation.
In contrast, in the in vitro studies described here,the effect of NOD on 8-oxo-dG formation was laterand lasted longer. NOD could induce free radicalsin the liver for a prolonged period of time[16]. Thekinetic studies described above show that NOD isdifferent from MCYST-LR in inducing 8-oxo-dGin primary cultured rat hepatocytes as well as in ratlivers. This is likely due to the difference reported inthe literature between these toxins, in spite of theirclose resemblance in terms of structure and toxicity.The bio-availability and the hepatic accumulation ofNOD could differ from that of MCYST-LR. The NODpenetrates more easily into the hepatocytes than doesMCYST-LR [4,5]. Moreover, although these two hep-atotoxins inhibit protein phosphatases (1 and 2A) withthe same potency, only MCYST-LR bind covalently tocysteine residues on protein phosphatases 1 and 2A[4].T e( ODc atea les,e rins[ thw isk tora R,w ,M ds,s ,r atedt
elsf e ox-i hi d to
0�g/kg body weight. The 8-oxo-dG levels increaignificantly, after either MCYST-LR or NOD (Fig. 5).hile the levels in rat liver continued to increase aOD treatment, they were decreased at 48 h afteCYST-LR dose, although in this case they remai
ignificantly higher than the control values.As in the in vitro experiments, the MCYST-LR
nd NOD-induced 8-oxo-dG formation obsern vivo could also be caused by an excessive Reneration or a disturbance of metabolic and axidative activities. Although, the direct detectf cyanobacterial hepatotoxin-induced ROS in vas not been achieved, formation of ROS provoy microcystins in rat liver cells[33], support theypothesis suggesting the generation of free rad
n vivo under the influence of cyanotoxins. Electpin resonance spectroscopy and image-guided puclear magnetic resonance spectroscopy were
he different positions ofN-methyldehydrobutyrinMdhb) at the surface of the protein phosphatase–Nomplex compared with microcystins may facilitchemical interaction with further macromolecu
xplaining the carcinogenic properties of nodula4]. Indeed, the NOD promotes liver tumour growith a higher activity than microcystin-LR. Itnown as a de novo liver carcinogen, i.e. initiand promoter of tumours, in contrast to MCYST-Lhich is known only as a promoter[4,5]. FurthermoreCYST-LR can bind covalently to SH compoun
uch asl-cysteine and GSH[39,40]. Neverthelessecent studies showed that NOD can also be conjugo GSH by enzymatic activities[41,42].
It is interesting to note that the 8-oxo-dG levound in the present study are higher than one basdation product per 106 DNA bases, which is still hign terms of the amounts of other DNA lesions relate

I. Maatouk et al. / Mutation Research 564 (2004) 9–20 19
cancer[43]. Therefore, the risk posed by MCYST-LR-and NOD-induced ROS formation is considerable. Thepresence of 8-oxo-dG is recognized as being highlymutagenic, creating G to T transversions, as a resultof miscoding[18]. These transversions have also beenobserved in Ki-rasand H-rasby mis-replication of 8-oxo-dG lesions induced by ROS[44]. Nevertheless,Suzuki et al.[45] found that MCYST-LR induced base-substitution mutations in the K-rascodon 12 in humanRsa cells. A strong mutagenic response induced by anextract of the cyanobacteriumMicrocystissp. was ob-served using the Ames test with or without metabolicactivation[11]. The oxidative DNA damage could thusbe implicated in the mutagenic potential of cyanobac-terial hepatotoxins.
In conclusion, 8-oxo-dG formation in both primarycultured rat hepatocytes and in rat livers in vivo indi-cates clearly that MCYST-LR and NOD at low concen-trations induce DNA damage. Therefore, they couldact as tumour initiators, providing evidence of theircarcinogenicity. The observed oxidative DNA damagemay be a consequence of the cytotoxicity and the oxida-tive stress caused by these toxins. Thus, by induction ofDNA oxidative damage and subsequently the possibleexistence of errors in DNA replication following theDNA repair activation, the exposure to low concentra-tions of microcystins and nodularins could contributeto an increased risk for cancer. Further investigationsinto the metabolic activity and the induction and re-pair of DNA damages brought about by these toxinsw n-c ndc
A
ni-v ingo
R
S.T.ho,il-
alysis
[2] T. Kuiper-Goodman, I. Falconer, J. Fitzgerald, Human healthaspects, E & FN SPON, 1999.
[3] I.R. Falconer, A. Humpage, Tumour promotion by cyanobacte-rial toxins, Phycologia 35 (1996) 74–79.
[4] T. Ohta, E. Sueoka, N. Iida, A. Komori, M. Suganuma, R. Nishi-waki, M. Tatematsu, S.J. Kim, W.W. Carmichael, H. Fujiki,Nodularin, a potent inhibitor of protein phosphatases 1 and 2A,is a new environmental carcinogen in male F344 rat liver, Can-cer Res. 54 (1994) 6402–6406.
[5] H. Fujiki, E. Sueoka, M. Suganuma, Carcinogenesis of micro-cystins, CRC Press, New York, 1996.
[6] WHO, Guidelines for drinking-water quality, Addendum to vol.2 Geneva, World Health Organisation, 1998.
[7] Y. Ueno, S. Nagata, T. Tsutsumi, A. Hasegawa, M.F. Watan-abe, H.D. Park, G.C. Chen, G. Chen, S.Z. Yu, Detection ofmicrocystins, a blue-green algal hepatotoxin, in drinking watersampled in Haimen and Fusui, endemic areas of primary livercancer in China, by highly sensitive immunoassay, Carcinogen-esis 17 (1996) 1317–1321.
[8] I. Chorus, J. Bartram, Toxic Cyanobacteria in water: A guide totheir public health consequences, monitoring and management,WHO, E & FN SPON, 1999.
[9] P.V. Rao, R. Bhattacharya, The cyanobacterial toxinmicrocystin-LR induced DNA damage in mouse liver in vivo,Toxicology 114 (1996) 29–36.
[10] P.V. Lakshmana Rao, R. Bhattacharya, M.M. Parida, A.M. Jana,A.S.B. Bhaskar, Freswater cyanobacteriumMicrocystis aerug-inosa(UTEX 2385) induced DNA damage in vivo and in vitro,Environ. Toxicol. Pharmacol. 5 (1998) 1–6.
[11] W.X. Ding, H.M. Shen, H.G. Zhu, B.L. Lee, C.N. Ong, Geno-toxicity of microcystic cyanobacteria extract of a water sourcein China, Mutat. Res. 442 (1999) 69–77.
[12] B. Zegura, B. Sedmak, M. Filipic, Microcystin-LR induces ox-idative DNA damage in human hepatoma cell line HepG2, Tox-icon 41 (2003) 41–48.
[ para-xin88
[ g, T.ne-04)
[ f inin-002)
[ la-icol.
[ on-pato-
[ , 8-age,992)
ill make it possible to provide more information coerning their toxicity and their possible genotoxic aarcinogenic activity.
cknowledgement
We thank S. Ghouti (The language Department, Uersity Paris-Sud, UFR Pharmacy) for critical readf the manuscript.
eferences
[1] E.M. Jochimsen, W.W. Carmichael, J.S. An, D.M. Cardo,Cookson, C.E. Holmes, M.B. Antunes, D.A. de Melo FilT.M. Lyra, V.S. Barreto, S.M. Azevedo, W.R. Jarvis, Liver faure and death after exposure to microcystins at a hemodicenter in Brazil, N. Engl. J. Med. 338 (1998) 873–878.
13] N. Gupta, S.C. Pant, R. Vijayaraghavan, P.V. Rao, Comtive toxicity evaluation of cyanobacterial cyclic peptide tomicrocystin variants (LR, RR, YR) in mice, Toxicology 1(2003) 285–296.
14] L. Zhan, H. Sakamoto, M. Sakuraba, D.-S. Wu, L.-S. ZhanSuzuki, M. Hayashi, M. Honma, Genotoxicity of microcystiLR in human lymphoblastoid TK6 cells, Mutat. Res. 557 (201–6.
15] R.A. Towner, S.A. Sturgeon, K.E. Hore, Assessment ovivo oxidative lipid metabolism following acute microcystLR-induced hepatotoxicity in rats, Free Radic. Res. 36 (263–71.
16] A. Lankoff, A. Banasik, M. Nowak, Protective effect of metonin against nodularin-induced oxidative stress, Arch. Tox76 (2002) 158–165.
17] W.X. Ding, C. Nam Ong, Role of oxidative stress and mitochdrial changes in cyanobacteria-induced apoptosis and hetoxicity, FEMS Microbiol. Lett. 220 (2003) 1–7.
18] K.C. Cheng, D.S. Cahill, H. Kasai, S. Nishimura, L.A. LoebHydroxyguanine, an abundant form of oxidative DNA damcauses G-T and A-C substitutions, J. Biol. Chem. 267 (1166–172.

20 I. Maatouk et al. / Mutation Research 564 (2004) 9–20
[19] M. Dizdaroglu, P. Jaruga, H. Rodriguez, Measurement of 8-hydroxy-2′-deoxyguanosine in DNA by high-performance liq-uid chromatography–mass spectrometry: comparison with mea-surement by gas chromatography–mass spectrometry, NucleicAcids Res. 29 (2001) e12.
[20] D. Schuler, M. Otteneder, P. Sagelsdorff, E. Erwin, R.C. Gupta,W.K. Lutz, Comparative analysis of 8-oxo-2′-deoxyguanosinein DNA by 32P-and33P-postlabeling and electrochemical de-tection, Carcinogenesis 18 (1997) 2367–2371.
[21] U. Devanaboyina, R.C. Gupta, Sensitive detection of 8-hydroxy-2′deoxyguanosine in DNA by32P-postlabeling assayand the basal levels in rat tissues, Carcinogenesis 17 (1996)917–924.
[22] R.C. Gupta, J.M. Arif, An improved32P-postlabeling assay forthe sensitive detection of 8-oxodeoxyguanosine in tissue DNA,Chem. Res. Toxicol. 14 (2001) 951–957.
[23] H. Ide, Y.W. Kow, B.X. Chen, B.F. Erlanger, S.S. Wallace, Anti-bodies to oxidative DNA damage: characterization of antibodiesto 8-oxopurines, Cell Biol. Toxicol. 13 (1997) 405–417.
[24] H. Kasai, S. Nishimura, Hydroxylation of deoxydeoxyguanoo-sine by ascorbic acid and other reducing agents, Nucleic AcidsRes. 12 (1984) 2137–2145.
[25] P.O. Seglen, Preparation of isolated rat liver cells, Methods Cell.Biol. 13 (1976) 29–83.
[26] G.A. Miura, N.A. Robinson, W.B. Lawrence, J.G. Pace, Hepa-totoxicity of microcystin-LR in fed and fasted rats, Toxicon 29(1991) 337–346.
[27] J.M. Arif, L.W. Robertson, R.C. Gupta, Artifactual productionof 8-oxodeoxyguanosine (8-oxo-dG) during tissue DNA pro-cessing and its inhibition, Proc. Am. Assoc. Cancer Res. 40(1999) 645.
[28] D.K. Scates, A.D. Spigelman, S. Venitt, Appearance of arte-facts when using32P-postlabelling to investigate DNA adductformation by bile acidsin vitro: lack of evidence for covalentbinding, Carcinogenesis 16 (1995) 1489–1491.
[ al-e446
[ ven inHep-
[ y-dis-on.
[ jurybac-–
[33] N. Bouaıcha, I. Maatouk, Microcystin-LR, Nodularin induceintracellular glutathione alteration, reactive oxygen species pro-duction and lipid peroxidation in primary cultured rat hepato-cytes, Toxicol. Lett. 148 (2004) 53–63.
[34] K. Haidara, I. Morel, V. Abalea, M. Gascon Barre, F. Denizeau,Mechanism oftert-butylhydroperoxide induced apoptosis inrat hepatocytes: involvement of mitochondria and endoplasmicreticulum, Biochim. Biophys. Acta 1542 (2002) 173–185.
[35] G. Mellgren, O.K. Vintermyr, R. Boe, S.O. Doskeland, Hepato-cyte DNA replication is abolished by inhibitors selecting proteinphosphatase 2A rather than phosphatase 1, Exp. Cell Res. 205(1993) 293–301.
[36] A.R. Humpage, I.R. Falconer, Microcystin-LR and liver tu-mour promotion: effects on cytokinresis, ploidy, and apopto-sis in cultured hepatocytes, Environ. Toxicol. Water Qual. 14(1999) 61–75.
[37] A. Lankoff, A. Banasik, G. Obe, M. Deperas, K. Kuzminski,M. Tarczynska, T. Jurczak, A. Wojcik, Effect of microcystin-LR and cyanobacterial extract from Polish reservoir of drinkingwater on cell cycle progression, mitotic spindle, and apoptosis inCHO-K1 cells, Toxicol. Appl. Pharmacol. 189 (2003) 204–213.
[38] H. Wiseman, B. Halliwell, Damage to DNA by reactive oxygenand nitrogen species: role in inflammatory disease and progres-sion to cancer, Biochem. J. 313 (1996) 17–29.
[39] S. Pflugmacher, C. Wiegand, A. Oberemm, K.A. Beattie, E.Krause, G.A. Codd, C.E. Steinberg, Identification of an en-zymatically formed glutathione conjugate of the cyanobacte-rial hepatotoxin microcystin-LR: the first step of detoxication,Biochim. Biophys. Acta 1425 (1998) 527–533.
[40] S. Takenaka, Covalent glutathione conjugation to cyanobacte-rial hepatotoxin microcystin LR by F344 rat cytosolic and mi-crosomal glutathione S-transferases, Environ. Toxicol. Pharma-col. 9 (2001) 135–139.
[41] K.A. Beattie, J. Ressler, C. Wiegand, E. Krause, G.A. Codd,C.E. Steinberg, S. Pflugmacher, Comparative effects and
rine
[ -altic
[ e toea-
Res.
[ l in-uman
[ to,y of98)
29] A. Izzotti, C. Cartiglia, M. Taningher, S. De Flora, R. Bansky, Age-related increases of 8-hydroxy-2′-deoxyguanosinand DNA-protein crosslinks in mouse organs, Mutat. Res.(1999) 215–223.
30] W.X. Ding, H.M. Shen, C.N. Ong, Critical role of reactioxygen species and mitochondrial permeability transitiomicrocystin-induced rapid apoptosis in rat hepatocytes,atology 32 (2000) 547–555.
31] W.X. Ding, H.M. Shen, C.N. Ong, Critical role of reactive oxgen species formation in microcystin-induced cytoskeletonruption in primary cultured hepatocytes, J. Toxicol. EnvirHealth A. 64 (2001) 507–519.
32] M.T. Runnegar, J. Andrews, R.G. Gerdes, I.R. Falconer, Into hepatocytes induced by a peptide toxin from the cyanoterium Microcystis aeruginosa, Toxicon 25 (1987) 12351239.
metabolism of two microcystins and nodularin in the bshrimpArtemia salina, Aquat. Toxicol. 62 (2003) 219–226.
42] V. Sipia, H. Kankaanpaa, K. Lahti, W.W. Carmichael, J. Meriluoto, Detection of nodularin in flounders and cod from the BSea, Environ. Toxicol. 16 (2001) 121–126.
43] B. Halliwell, Oxygen nitrogen are pro-carcinogens. DamagDNA by reactive oxygen, chlorine and nitrogen species: msurement, mechanism and the effects of nutrition, Mutat.443 (1999) 37–52.
44] S.P. Hussain, F. Aguilar, P. Amstad, P. Cerutti, Oxy-radicaduced mutagenesis of hotspot codons 248 and 249 of the hp53 gene, Oncogene 9 (1994) 2277–2281.
45] H. Suzuki, M.F. Watanabe, Y. Wu, T. Sugita, K. Kita, T. SaX. Wang, H. Tanzawa, S. Sekiya, N. Suzuki, Mutagenicitmicrocystin-LR in human RSa cells, Int. J. Mol. Med. 2 (19109–112.





![4-[(2,4-Difluorophenyl)hydrazinylidene]-3-methyl-5-oxo-4,5-dihydro-1 H -pyrazole-1-carbothioamide](https://static.fdokumen.com/doc/165x107/63448600596bdb97a9087f01/4-24-difluorophenylhydrazinylidene-3-methyl-5-oxo-45-dihydro-1-h-pyrazole-1-carbothioamide.jpg)














