
Vemurafenib reverses immunosuppression by myeloid derived suppressor cells

Current Cancer Drug Targets, 2008, 8, 207-222 207
1568-0096/08 $55.00+.00 © 2008 Bentham Science Publishers Ltd.
Deregulation of the Mitochondrial Apoptotic Machinery and Development
of Molecular Targeted Drugs in Acute Myeloid Leukemia
G. Del Poeta*, A. Bruno, M.I. Del Principe, A. Venditti, L. Maurillo, F. Buccisano, R. Stasi, B. Neri, F. Luciano, A. Siniscalchi, P. de Fabritiis and S. Amadori
Cattedra di Ematologia, Università Tor Vergata, Ospedale S.Eugenio, Roma, Italy
Abstract: Apoptosis plays a key role in the control of rapidly renewing tissues, such as the hematopoietic system and leu-
kemia cells invariably have abnormalities in one or more apoptotic pathways, determining a survival advantage of these
cells and the development of drug resistance. These defects are also frequently associated with a low rate of response to
standard chemotherapy and with a poor survival in acute myeloid leukemia (AML). The major form of apoptosis proceeds
through the mitochondrial pathway, with the mitochondrial outer membrane permeabilization, leading to the release of
proteins normally found in the space between the inner and outer mitochondrial membranes (cytochrome C, AIF and oth-
ers). Higher levels of anti-apoptosis proteins bcl-2, bcl-xL, Mcl-1 block permeabilization of the membrane and are re-
ported in AML patients presenting a poor outcome. On the contrary, activated pro-apoptotic bax or bad proteins allow this
permeabilization and are correlated to a good prognosis in AML. Defects in the mitochondrial pathway induce multidrug-
resistance and confer important prognostic information in AML. High ratios of bcl-2 to bax protein confer a poor progno-
sis with decreased rates of complete remission and overall survival. The prognostic information from the ratio of the pro-
teins is greater than bcl-2 levels alone. Recently, we confirmed the impressive impact of the bax/bcl-2 ratio, determined
by flow cytometry, on AML prognosis (complete remission and overall survival) in 255 AML patients. Bcl-2 down regu-
lation might lower the apoptotic threshold of leukemic cells and, through this mechanism, favor response to chemother-
apy. Phase II studies of oblimersen (antisense Bcl-2), cytarabine and daunorubicin or oblimersen plus gentuzumab, a cyto-
toxic antibody directed against CD33+ cells in relapsed AMLs, showed promising results. Defects in apoptosome pro-
teins, such as APAF-1, are frequent in AML and treatment with 5-aza-2’-deoxycytidine, a specific inhibitor of DNA
methylation, restored APAF-1 expression in leukemic cells. In conclusion, targeted therapies that are designed to induce
apoptosis in leukemic cells, are the most promising anti-leukemia strategies. The elucidation of the apoptotic machinery
and of its defects in AML lays the basis for developing new drugs able to trigger apoptosis and overcome therapy resis-
tance.
Keywords: Acute myeloid leukemia, Mitochondrial Apoptosis, Bcl-2, Apoptosome, Prognosis, Targeted therapy, Anti-sense oligonucleotides, Proteasome inhibitors.
INTRODUCTION
Acute myeloid leukaemia (AML) is a heterogeneous group of malignant diseases arising as the result of progres-sive genetic damage occurring in haemopoietic progenitor cells. Although current treatment strategies and supportive care are improved, the AML prognosis is still poor. In fact, patients with AML who are younger than 55 years of age and who have no adverse prognostic features have a com-plete remission rate of 70 to 85% with standard induction chemotherapy, but the long-term survival of patients is only 50% [1-2].
The development of AML is associated with ac-
cumulation of acquired genetic alterations and epigenetic changes in hematopoietic progenitor cells which alter normal mechanisms of cell growth, proliferation, death and differen-tiation. Multiple mechanisms may account for the reason why leukemic cells become resistant to chemotherapy, such as alterations in drug transport, drug metabolism and in cel-lular repair mechanisms. One common cause of resistance to chemotherapy includes defects in apoptotic pathways [3]. Studies carried out during the last ten years have signifi-cantly improved our understanding of apoptotic mechanisms
*Address correspondence to this author at the Cattedra di Ematologia, Uni-
versità Tor Vergata, Ospedale S.Eugenio,P.le Umanesimo, 10, 00144 Roma,
Italy; Tel: +390651002509; Fax: +39065914745; E-mail: [email protected]
in the control of differentiation, proliferation and survival of normal and leukemic cells. The major form of apoptosis seen in most settings in vertebrate cells proceeds from the mito-chondrial pathway, defined by a pivotal event in the process – mitochondrial outer membrane permeabilization (MOMP) [4]. MOMP occurs suddenly, leading to the release of pro-teins normally found in the space between the inner and outer mitochondrial membranes (including citochrome c, AIF, and others). MOMP precipitates cell death through as many as three general mechanisms, including the release of molecules involved in the activation of caspases that orches-trate downstream events often associated with apoptosis, the release of molecules involved in caspase-independent cell death and the loss of mitochondrial functions essential for cell survival.
CELL DEATH PATHWAYS
A common feature of the apoptotic process is the activa-
tion of caspases, a family of intracellular cysteine proteases (cysteine aspartyl-specific proteases). These enzymes are
present as inactive zymogens and can be triggered to assume
an active state, through proteolytic processing at conserved aspartic acid residues. Caspases are distinguished in initiator
caspases (caspase-8, -2, -9 and -10) and effector caspases
(caspase-3, -6 and -7): the former ones are involved in cell

208 Current Cancer Drug Targets, 2008, Vol. 8, No. 3 Del Poeta et al.
death initiation, while the latter are involved in cell death
execution. The effector caspases raise the proteolytic cleav-
age of a broad spectrum of cellular proteins, whose cleavage contributes to the morphological and functional changes as-
sociated with apoptosis [5-6]. Upstream or initiator caspases
contain long prodomains that interact with their specific ac-tivators. The downstream or effector caspases contain
smaller prodomains and are activated by proteolytic cleavage
by upstream caspase. There are two pathways by which caspase activation is induced, the extrinsic and the intrinsic
apoptotic pathways (Fig. (1)). Both pathways converge on
activation of effector caspases, but require different initiator caspases to start the process. The extrinsic pathway is acti-
vated by engagement of death receptors on the cell mem-
brane. Binding of ligands such as Fas ligand, tumor necrosis factor (TNF) and TNF-related apoptosis-inducing ligand
(TRAIL) to their cognate membrane receptors Fas, TNF-R
and TRAIL-R result in the formation of the oligomeric death-induced signaling complex (DISC). DISC promotes
caspase-8 activation and a cascade of activation of caspases
(caspase-3) that culminates with cell death. The activation of caspase-8 is antagonized by cellular FLICE inhibitory pro-
tein (c-FLIP), inactive and relative of caspase-8 and –10 that
binds to DISC. The intrinsic pathway is triggered by various extracellular and intracellular stresses, including growth fac-
tor deprivation, DNA damage, oncogene induction, hypoxia
and cytotoxic drugs (Fig. (1)). In response to exogenous stimuli, the intermediates (ceramide, p53, JNK/SAPK, c-
myc) activate the mitochondrial-mediated pathway of apop-
tosis. Mitochondrial membrane permeability is controlled by pro-apoptotic (Bax, Bak, Bad, Bid, Bim, Bmf, NOXA,
PUMA) and anti-apoptotic (Bcl-2, Bcl-xl, Mcl-1) members
of the Bcl-2 family, inducing or preventing heterodimeriza-tion of pro-apoptotic members. Bcl-2 proteins localize or
translocate to the mitochondrial membrane and modulate
apoptosis by permeabilization of the inner and/or outer membrane, leading to the release of cytochrome c or by sta-
bilizing barrier function. The bax family of death agonist act
at the mitochondria to induce apoptosis through three mecha-nisms: 1) at the outer membrane bax accelerates the opening
of the voltage-dependent anion channel (VDAC), with
sequential release of cytochrome c, 2) bax enhances the permeability of the inner mitochondrial membrane and 3)
bax is capable of forming pores by a direct oligomerization
mechanism that induces cytochrome c release [7-8]. The bcl-2 class of antiapoptotic proteins are important inhibitors of
the mitochondrial apoptotic pathway. Bcl-2 can inhibit the
action of bax/bak by forming inactivating heterodimers. Be-sides, bcl-2 binds VDAC and stabilizes it, thereby preventing
permeabilization of the outer mitochondrial membrane. Bcl-
2 and bcl-xl dimerize with several members of bcl-2 family of proteins thereby changing the ratio of these members and
altering the threshold of cell death [9]. Bcl-2 can inhibit the
action of bax/bak by forming inactivating heterodimers. The damage of the outer mitochondrial membrane and the conse-
quent release of cytochrome c and other pro-apoptotic mole-
cules, such as Smac/DIABLO from the inner membrane into the cytosol enable the formation of the apoptosome, a large
molecular complex formed by cytochrome c, apoptotic pro-
tease activating factor 1 (APAF-1) and caspase-9, and mas-sive activation of caspases (Fig. (1)).
The pro-survival Bcl-2 family members are characterized by four Bcl-2 homology domains (BH1-BH4) which play crucial roles for cell survival and the loss of any of these proteins causes major deregulation of survival of some cell types. The pro-apoptotic members of the Bcl-2 family can be subdivided into two groups: 1) Bax-like proteins (Bax, Bak, Bok) contain three BH regions; 2) BH3-only proteins (Bad, Bik, Bid, Bim, Noxa, PUMA, Bmf) possess only the BH3 domain. BH3-only proteins are crucial for cell death initia-tion through the intrinsic pathway, whereas the Bax/Bak like proteins play a key role in the progression of the apoptotic signaling at the level of permeabilization of the outer mito-chondrial membrane.
GENERAL MECHANISMS OF APOPTOSIS DE-REGULATION IN ACUTE MYELOID LEUKEMIA
Deregulation of apoptosis disrupts the frail balance be-tween cell proliferation, cell survival and cell death playing a major role in the development of cancers such as leukemia. Moreover, tumors that have alterations in proteins involved in cell death signaling are often resistant to chemotherapy that primary acts by inducing apoptosis. In an attempt to overcome drug resistance and improve clinical outcomes, attention has turned to develop therapeutic agents that over-come defects in apoptosis pathways.
Leukemia development is regarded as a multistep process characterized by progressive genetic alterations that drives the transformation of normal human hematopoietic stem/progenitor cells into leukemic derivatives. Leukemia originates from a single cell that has undergone malignant transformation driven by frequent genetic mutations. These events are followed by a clonal selection of mutated cells that display an increasingly aggressive behavior.
About 50–55% of AML are characterized by the pres-ence of specific balanced chromosome abnormalities in-volved in the formation of specific fusion genes that encode for fusion proteins, playing a key role in the development of leukemia through their effect on cell proliferation, survival and apoptosis [10]. Simultaneous alterations in the normal regulation of cell proliferation and apoptosis induce leuke-mic cells to accumulate, thus promoting further genetic al-terations and leukemogenesis. The key role of apoptosis in the pathogenesis of AML has been elucidated over the last years. Some fusion proteins interact with mediators of apop-tosis, sending anti-apoptotic signals that favor the preferen-tial survival of leukemic cells: PML/RAR-alpha or CBFbeta/SMMHC through the p53 pathway or AML1/ETO through the Bcl-2-related pathway [11-13]. Furthermore, it has been suggested that MLL fusion proteins modify the PPP1R15A function and inhibit apoptosis [14]. Finally, the PML/RAR-alpha fusion protein was shown to exert an anti-apoptotic activity by down modulating the expression of some death-inducing genes, such as TNF-R1 [15] and TRAIL-R1/-R2 [16]. Interestingly, a high frequency of mu-tations of nucleophosmin was observed in AML that do not display chromosomal translocations [17]. In fact, mutations affecting the carboxyterminal domain of nucleophosmin oc-cur in a high percentage of adult patients with AML not as-sociated with chromosomal translocations and these altera-tions create an additional nuclear export signal that relocal-

Deregulation of the Mitochondrial Apoptotic Machinery Current Cancer Drug Targets, 2008, Vol. 8, No. 3 209
Fig. (1). The principal pathways of apoptosis.
1) Receptor mediated-apoptosis. Apoptosis can be induced by external cell stimuli ((inflammation or drugs). These signals cause binding of
specific ligands to death receptors such as Fas which is the best studied members of the TNF receptor superfamily. The downstream signal
transduction involves formation of a death inducing signal complex (DISC) consisting of Fas, Fas-associated death domain (FADD) and pro-
caspase 8. In this case, 2 signal pathways are possible: caspase 8 can directly cleave procaspase 3 followed by activation of caspase 3 and
consequently DNA fragmentation or it can truncate Bid to tBid which acts on the outer mitochondrial membrane by pore formation and re-
lease of proapoptotic peptides. FLICE-inhibitory protein (FLIP) which contains two death effector domains and an inactive caspase domain,
bind to FADD and procaspase 8, and thereby inhibits death receptor-mediated apoptosis. 2) Mitochondria-mediated apoptosis. The mito-
chondrial pathway is activated by internal stimuli or cellular stress. The proapoptotic BCL-2 members (BAX and BAK) act on mitochondrial
level by pore formation and release of apoptogenic substances into the cytoplasm. The apoptosome complex (association of cyt c, APAF-1
and procaspase 9) activates caspases 9 and 3 and induces DNA fragmentation. IAPs (inhibitors of apoptosis protein) contribute to apoptosis
resistance by inhibition of caspases 9 and 3 and thereby rescue cells from apoptosis. Apoptosis can also proceed in the absence of caspase
activity when the mitochondria is damaged. In this case, release of AIF and Endonuclease G from mitochondria starts an apoptotic program
parallel to caspase activation. 3) Granzyme B. Cytolytic T cells, by secreting granule-associated serine proteases (granzyme B) can induce
apoptosis in target cells directly activating caspases.
izes much of the protein from its normal nucleolar stores to the cytoplasm [17]. Furthermore, nucleophosmin is also fre-quently found in chromosomal translocations associated with myelodysplasia/AML [18]. Nucleophosmin acts as a cellular p53 negative regulator to protect hematopoietic cells from stress-induced apoptosis [19]. The mutated nucleophosmin could perturb p53-dependent and independent activities of the ARF tumor suppressor protein, and through this mecha-nism could protect AML cells from apoptosis. Flt3 is over-expressed in the majority of AML and, importantly, is mu-tated in about 35% of AML. In about 25% of patients an internal tandem duplication (ITD) within the juxtamembrane domain of Flt3 is observed, while in about 10% of patients
mutations, small deletions or insertion within the activation loop of the second kinase domain of Flt3 are observed [20]. These mutations determine a constitutive activation and sig-naling of Flt3, resulting in the triggering of anti-apoptotic pathways. Flt3 ITD mutants constitutively activate MAPK, AKT and Stat5, leading to Pim-1 activation and Bcl-xL hy-perexpression [21]. Another membrane receptor tyrosine kinase frequently mutated in AML is c-kit. In fact, c-kit mu-tations at the level of exon 8 (encoding for a part of the ex-tracellular domain of the receptor) or mutations at the level of exon 17, codon 816 in the activation loop of the catalytic domain occur in about 20% and 30% of AML patients with t(8;21) or inv(16)t(16;26) [22]. Extracellular c-kit mutations

210 Current Cancer Drug Targets, 2008, Vol. 8, No. 3 Del Poeta et al.
resulted in c-kit receptor hyperactivation in response to kit ligand, with subsequent strong activation of MAPK and PI3K, while codon 816 c-kit mutations induced constitutive Stat-3 activation and upregulation of Bcl-xL and c-myc. The KITD816 mutations occur in t(8;21)/AML-1/ETO-positive AML and represent a reliable molecular marker identifying patients with a poor prognosis in an otherwise prognostically favorable AML group [23]. Similarly, both types of c-kit mutations have an adverse impact on relapse risk and overall survival in AML with inv(16) [24]. KRAS and NRAS muta-tions are present in approximately 15% and 25% of AML patients: these mutations are usually point mutations and determine constitutive GTPase activity [25]. NRAS muta-tions lead to increased activity of the RAS pathway, resulting in increased proliferation and decreased apoptosis [26]. Con-stitutive RAS activation in AML without RAS mutations is also frequently observed, probably due to deregulation of signaling proteins acting upstream of RAS. Animal models indicate that abnormalities in the apoptosis pathway may lead to the development of AML. Thus, Fas-deficient mice constitutively expressing Bcl-2 in myeloid cells develop a disease closely resembling the M2 FAB subtype of AML [27]. These observations indicate that molecules involved in the control of apoptosis may act as tumor suppressors to con-trol leukemogenic transformation in myeloid progenitor cells.
In addition to traditional cytogenetic analysis or the de-tection of mutations in individual genes, global gene expres-sion profiling through microarray technology has provided an additional tool to probe and to try to classify in a rational way the marked biological heterogeneity of AML [28]. This approach has enabled the identification of several clusters of AML, exhibiting different molecular signatures, prognoses and responses to treatment [29]. Cluster A is characterized by AML exhibiting a high frequency of NPM1 mutations, a so-called stem-cell molecular signature and overexpression of genes that promote apoptosis, such as LTBP1 and caspase-3. Cluster B, represented by a group of AML pa-tients with a very poor response to induction therapy and a very bad prognosis, is characterized by overexpression of genes inducing chemoresistance, such as ABCG2, almost complete absence of NPM1 mutations and often a normal cytogenetic analysis. AML patients in cluster C exhibit a high proliferative activity gene signature, a high incidence of karyotypic abnormalities, a low prevalence of NPM1 muta-tions and overexpression of IRF4 and IL10R. A high rate of response to induction therapy, but also a high rate of relapse have been observed in this cluster of patients. Cluster F is characterized by AML with monocytic features, with the highest white cell counts and a poor response to therapy. Some signaling pathways, including phosphoinositide 3-kinase (PI3K)/AKT, JAK/STAT, RAS/Raf, /MEK/ERK and protein kinase C , are crucial to many aspects of cell growth, survival and apoptosis, and their constitutive activation has been implicated in both the pathogenesis and progression of AML. These pathways have been extensively studied in AML. AKT was found to be constitutively phosphorylated in the majority of AML patients with high blast cell counts and was an adverse prognostic factor [30]. Phosphorylated AKT, in turn, phosphorylates caspases, Bad, mTOR, Waf1, and NF-kappaB, resulting in protection from apoptosis and in-
creased proliferation [31]. Activated AKT may promote leu-kemogenesis and inhibition of AKT activity induces apopto-sis of AML blasts [32]. The RAS/Raf/MEK/ERK pathway is a key signaling transduction pathway that induces increased cell proliferation and survival: constitutive activation of this pathway is observed in the majority of AML patients and its blockade with specific pharmacologic agents induces blast cell death [33]. Finally, recent studies have provided clear evidence that a significant proportion of AML exhibit the simultaneous activation of AKT, ERK and PKC and exhibit a worse prognosis, associated with short survival [34].
Dysregulation of apoptosis is due to abnormal expression or mutation of genes [35]. An important example also from the clinical point of view is the BCR–ABL fusion protein in chronic and acute myeloid leukemia with 9/22 translocation. This results in dysregulation of ABL tyrosine kinase increas-ing the transcription of the antiapoptotic Bcl-xL protein [36]. The dysfunction of the p53/Bax/caspase-3 apoptosis signal-ing pathway has been shown to play a role in tumorigenesis. Spontaneous apoptosis proved to be significantly higher in normal CD34+ cells compared to AML CD34+ cells exam-ined by annexin V binding, while it was lower, if all bone marrow mononuclear cells were examined. Apoptosis in acute lymphoid leukemia (ALL) is significantly lower than in AML and shows no significant difference from normal peripheral blood mononuclear cells [37-38]. Caspase-3 activ-ity was shown only in the cells of AML patients. The impor-tance of the methods used is shown by the finding that AML had four times higher caspase-3 activity than ALL, however the annexin V binding was similar. On the other hand, the overexpression of the inactive form of caspase-3 is fre-quently observed in the blast cells of patients with ALL as well, as in AML. In a study carried out on newly diagnosed AML patients increased levels of procaspase-3 in leukemic cells were correlated with a poor prognosis [39]. However, patients exhibiting spontaneously active caspases-3 at the time of diagnosis showed improved survival [39]. The poor prognosis among patients with high levels of procaspase-3 may be related to defects in caspase activation with subse-quent accumulation of uncleaved caspase-3. The better prog-nosis observed among patients with increased levels of cleaved caspase-3 may be related to the activation of apop-totic pathways in these leukemic cells.
MITOCHONDRIAL APOPTOSIS AS PROGNOSTIC MARKER IN AML
There are at least four pathways for initiation of caspase activation: (i) the mitochondrial pathway initiated by cyto-chrome c, (ii) the death receptor pathway initiated by the TNF-family of death receptors, (iii) direct caspase activation by the cytolytic T cell protease Granzyme B, and (iv) a pathway connected to the endoplasmic reticulum [40-41] (Fig. (1)).
Of these four pathways of caspase activation, the mito-chondrial pathway is the primary effector of chemotherapy-induced apoptosis [42]. This pathway for caspase activation is initiated by the release of cytochrome c from the mito-chondria [43]. Upon release, cytochrome c binds Apaf-1 in the presence of dATP and causes Apaf-1 to change confor-mation. This conformational change causes oligomerization

Deregulation of the Mitochondrial Apoptotic Machinery Current Cancer Drug Targets, 2008, Vol. 8, No. 3 211
Fig. (2). The formation of apoptosome.
a) The Apoptosis Protease Activating Factor-1 (APAF-1), a cytosolic protein, contains a caspase recruitment domain (CARD), a nucleotide
binding domain (NOD) and a WD-40 domain which are able to bind caspase 9, dATP/dADP and cytochrome c, respectively.
b) In absence of apoptotic signal, APAF-1 normally exists as an inactive monomer. The binding of cytochrome c to the WD-40 domain in-
duces a conformational change in APAF-1, allowing it to bind the nucleotide dATP/dADP. The nucleotide binding to the APAF-1-
cytochrome c complex triggers its oligomerization with a characteristic wheel aspect and recruits and activates procaspase-9 to form the apop-
tosome.
of Apaf-1, exposes its caspase-activation recruitment domain (CARD), and permits the recruitment of procaspase-9 [44] (Fig. (2)). Upon recruitment to the complex, procaspase-9 dimerizes and is activated. Active caspase- 9 then cleaves and activates effector caspases, such as procaspase-3 [45]. Thus, the mitochondrial pathway can be divided into three components: (i) pre-cytochrome c release, (ii) post-cytochrome c release, and (iii) the convergence point with the other caspase pathways at the level of effector caspases. Thus, defects at the convergence point, for example, can be due to lack of functional effector caspases or over-expression of inhibitors of effector caspases. Chemotherapeutic agents used in the treatment of AML, such as cytarabine and etoposide, induce apoptosis, in part, by disrupting mitochon-drial membrane potential and increasing reactive oxygen intermediates [45]. As a result, cytochrome c is released from the mitochondria and caspase activation is initiated. Blocks in this pathway can render cells resistant to cyta-rabine and etoposide-induced apoptosis [45]. Therefore,
identifying the occurrence and prevalence of defects in this caspase pathway can further improve our understanding of mechanisms of response and relapse in primary AML sam-ples. Impaired caspase activation was frequent in samples from patients with rapidly relapsing disease, but was rare in samples from patients with newly diagnosed AML. When defects in caspase activation were present, they usually af-fected a point at or below the convergence of the caspase pathways at effector caspases. To investigate the down-stream portion of the mitochondrial pathway of caspase acti-vation in patients with AML, cytosolic lysates were stimu-lated with cytochrome c and dATP and hydrolysis of Ac-DEVD-AFC by effector caspases was measured [46]. De-fects in the distal mitochondrial pathway were more common in samples from patients with AML that relapsed rapidly after induction chemotherapy compared to samples from treatment naive patients [46]. The incidence of blocked pathways did not differ based on response to induction chemotherapy, as even non responders generally had an intact pathway. When the distal mitochondrial pathway was

212 Current Cancer Drug Targets, 2008, Vol. 8, No. 3 Del Poeta et al.
pathway. When the distal mitochondrial pathway was blocked, defects were usually at the level of the effector caspases. Thus, functional defects in the distal portion of the mitochondrial pathway of caspase activation may help ex-plain the nature of response and relapse after treatment [46].
Defects in the mitochondrial pathway induce multidrug-resistance and confer important prognostic information. The Mcl-1 gene encodes a 36kDa protein with a carboxy terminal domain similar to Bcl-2. However, in contrast to Bcl-2, which is a long-lived protein, Mcl-1 has a short half-life. The analysis of Mcl-1 protein expression in AML showed great heterogeneity, but the levels of the protein do not seem to correlate with response to standard chemotherapy [47]. In a longitudinal study of patients with relapsed acute leukaemia,. Mcl-1 at relapse was 2-fold higher than at presentation in 53% of patients [47]. The increase in Mcl-1 may in part ex-plain the greater degree of drug resistance seen in relapsed leukaemia.
AML blasts frequently express elevated levels of the anti-apoptotic Bcl-2 family member, Bcl-xL. Elevated levels of Bc-xL have been observed in CD34+ AML cells and con-tribute to the chemoresistance of these cells [48]. Recent studies have shown that AML with Flt3 mutations are usu-ally associated with elevated Bc-XL levels, seemingly in-duced via constitutive Stat-5 activation [49].
The X-linked inhibitor of apoptosis protein (XIAP) was found to be expressed in the large majority of adult cases of AML [50-51]. In a significant proportion of patients high levels of XIAP were detected. An initial study carried out on 78 adult AML patients suggested a potential relationship between the level of XIAP expression and response to ther-apy, patients with high XIAP levels having a shorter survival than that observed in patients with low XIAP levels [50]. However, in a second study, carried out on 172 adult patients with primary AML, the same authors failed to observe a sig-nificant correlation between the level of XIAP expression in AML blasts and complete remission rate or overall survival or other clinical parameters [51]. Survivin expression was detected in the majority of AML, but there was no evidence that its levels in leukemic blasts correlated with the response to treatment or with survival [51].
It has been suggested that the bax/bcl-2 ratio may be more important than either promoter alone in determing apoptosis. Compared to conventional techniques, such as immunocytochemistry and western blot, flow cytometry may be particularly well suited for the precise, semi-quantitative measurement of bcl-2 and bax expression, for the determina-tion of bax/bcl-2 ratio and, finally, for the estimation of cel-lular heterogeneity that is frequently found in AML [52-53]. Moreover, the apoptosis pathways are positively and nega-tively regulated through signal transduction: bcl-2 and BAD are examples of proteins that are regulated by phosphoryla-tion and whose regulation is being exploited for drug devel-opment [54]. Some aspects of bcl-2 phosphorylation are con-troversial. One report indicates that phosphorylation of ser-70 by protein kinase C (PKC) activates the antiapoptotic function of bcl-2 [55], but another study reports that taxol treatment leads to phosphorylation on the same serine and inhibits its function [56]. All-trans retinoic acid (ATRA) inhibits bcl-2 by inducing phosphorylation on a serine dis-
tinct from the sites phosphorylated by PKC and the taxol-mediated pathway [57]. Pathways that dephosphorylate BAD increase its activity and promote apoptosis. Calcineurin, a calcium-dependent phosphatase, dephosphorylates BAD and promotes BAD-induced apoptosis [58].
Bcl-2 is an important predictor of survival in multivariate analysis over other known prognostic markers, including age and white blood cell count [53]. On the contrary, in patients with breast cancer, increased bcl-2 by immunostaining corre-lated with improved disease-free survival and in childhood ALL increased bcl-2 was associated with improved event-free survival [59-60]. These results may reflect the impor-tance of posttranslational modification of bcl-2. Under cer-tain circumstances PKC activates bcl-2, so in the presence of PKC, the bcl-2 that accumulates is functional, conferring a worse outcome. Infact, lower levels of PKC were associated with an improved prognosis in patients with AML [61]. In a study of 56 AML pts, increased levels of bax expression by immunoblotting correlated with improved rates of overall survival [62]. In contrast, a study of 165 AML de novo pts, reported that levels of bax expression did not correlate with response to induction chemotherapy or survival [61]. How-ever, high ratios of bcl-2 to bax protein conferred a poor prognosis with decreased rates of complete remission and overall survival. The prognostic information from the ratio of the proteins was greater than bcl-2 levels alone. The ratio also provided prognostic information above cytogenetics. We confirmed in our published study the impressive impact of the bax/bcl-2 ratio, determined by flow cytometry, on AML prognosis [63].
At AML relapse, it is not uncommon that blast cells dis-play more immature immunophenotypic features than they do at diagnosis. These findings suggest that immature blast cells may be more resistant to chemotherapy and more effi-ciently protected against undergoing apoptosis than differen-tiated blast cells. Bcl-2 is heterogeneously expressed in AML and its expression is often upregulated compared to levels in non-malignant CD34+ counterparts [64].
The study
by Banker et al. [65] demonstrated that AML samples with high Bcl-2 expression generally showed low apoptosis in vitro. Interestingly, van Stijn et al. [66] showed that CD34+ AML fractions are more resistant to apoptosis than are corre-sponding CD34-AML fractions, and that this paralleled by higher Bcl-2, Bcl-xl, Mcl-1, Pgp and lower Bax expression levels. The correlation of the other progenitor markers, CD133 and CD117, with an apoptosis-resistant protein pro-file supports the finding that immature cells are more apop-tosis-resistant than mature cells. Suarez et al. [67] reported that Bcl-2 and Bax differed according to different stages of maturation: CD34+ AML blast cells were defined as the most immature subpopulation, CD34-CD32-/+CD33+ as a less immature subpopulation, CD34-CD32+/++CD33++ as the most mature subpopulation. Delia et al. [68] reported that, in HL-60 cells induced to differentiate along the granu-locytic pathway by incubation with ATRA, bcl-2 expression was decreased. This study also showed that less mature mye-loblasts, metamyelocytes, myelocytes and CD34+ cells in bone marrow samples stained positively with anti-bcl-2 anti-bodies, as did splenic lymphocytes, while mature myeloid cells were negative for bcl-2 staining. Using flow cytometry, we have investigated the effects of all-trans-retinoic acid

Deregulation of the Mitochondrial Apoptotic Machinery Current Cancer Drug Targets, 2008, Vol. 8, No. 3 213
(ATRA) on bcl-2 expression in the blast cells of 25 AML patients and the HL-60 cell line [69]. The MFI ratio for the bcl-2 levels of the ATRA responders was significantly re-duced after incubation with ATRA compared to control sam-ples incubated without ATRA. Moreover, we performed a quantitative analysis of Fas and bcl-2 expression in CD34+ hematopoietic precursors obtained from bone marrow (BM), mobilized peripheral blood (MPB) and umbilical cord blood samples (UCB) [70]. Bcl-2 expression was significantly higher in BM than in MPB and UCB, and bcl-2 was in-versely related to CD34 expression. Bcl-2 expression was significantly higher in BM than in MPB and UCB and bcl-2 was inversely related to CD34 expression. Several reports confirmed the strict correlation between immaturity or CD34 expression and higher bcl-2 expression [64,71]: these find-ings are to be interpreted as a survival advantage of bcl-2 overexpressing progenitors and rapid elimination of cells with low bcl-2. In order to verify whether only the immatur-ity profile explains apoptosis levels and prognosis in AML, we studied a large series of 298 patients, affected by de novo AML, treated with intensive chemotherapy regimens [72]. Lower bax/bcl-2 ratio was strongly associated either with >20% CD34 (p<0.00001) or >10% CD133 (p=0.0002) or high CD117 (p<0.00001), confirming that higher primitive markers expression is consistent with an apoptosis resistant protein profile. A significant lower complete remission (CR) rate was found in patients with lower bax/bcl-2 ratio (42% vs 76%, p<0.00001). Overall survival (OS) and disease free survival (DFS) were significantly shorter in pts with lower bax/bcl-2 ratio (p<0.00001 and p=0.003). In order to confirm the independent prognostic value of bax/bcl-2 ratio from immaturity levels, we enucleated and investigated the CD34+ (168 patients), the CD117+ (119 patients) and the CD133+ (112 cases) AML subgroups. As a matter of fact, a lower CR rate was found in patients with lower bax/bcl-2 ratio either within CD34+ (39% vs 81%, p<0.00001) or CD117+ (30% vs 69%, p=0.0002) or CD133+ (34% vs 83%, p=0.0001) subsets. The strong independent prognostic value of bax/bcl-2 ratio was confirmed in multivariate analysis.
Expression profiling studies in blasts from relapsed AML showed mRNA expression changes in genes correlated with increased cellular proliferation and indicated by the overex-pression of the transferrin receptor, proliferating cell nuclear antigen and G1 cyclins [73]. Tumor cells in a high prolifera-tive state have a high density of transferrin receptors, as demonstrated in breast cancer cells [74] and in adult T-cell leukemia/lymphoma [75]. The present availability of an-tisense oligonucleotides targeted either to the transferrin re-ceptor genes or bcl-2 raised our interest to evaluate the real impact of proliferative and/or apoptotic pathways on AML prognosis [76]. Therefore, in order to verify whether the pro-liferative profile may explain both the apoptosis levels and prognosis in AML, we studied a large series of 325 patients, affected by de novo AML, treated with intensive chemother-apy regimens
1. The principal aims of our research were: 1) to
correlate bax/bcl-2 ratio, representing a measure of sponta-neous apoptosis, with the proliferative rate levels, deter-
1 Del Poeta, G.; Del Principe, M. I.; Venditti, A.; Maurillo, L.; Buccisano, F.; Irno
Consalvo, M.; Mazzone, C.; Cox, M.C.; Bruno, A.; Lo Coco, F.; Amadori, S. Clinical
significance of apoptosis is independent from proliferation in acute myeloid leukemia (AML). Blood 2004, 104, Abstract 61.
mined by the transferrin receptor (CD71) and 2) to demon-strate that the clinical significance of spontaneous apoptosis (bax/bcl-2 ratio) is independent from proliferation. No sig-nificant correlation was found between a higher bax/bcl-2 ratio and a lower CD71 (p=0.16), confirming that an apopto-sis resistant protein profile may have variable (high or low) proliferation levels. A significant lower complete remission (CR) rate was found in patients with lower bax/bcl-2 ratio (43% vs 72%, p<0.00001) or higher CD71 (47% vs 74%, p=0.00001). Overall survival (OS) was significantly shorter either in pts with lower bax/bcl-2 ratio (p=0.00001) or higher CD71 (p=0.002). Higher bax/bcl-2 ratio plus lower CD71 identified an AML subset at better prognosis with regard to CR (91% vs 32%; p<0.00001) and OS (p<0.00001).
In order to confirm the independent prognostic value of bax/bcl-2 ratio from proliferation levels, we investigated the high proliferative CD71 + (203 pts) and the low proliferative CD71- (121 pts) AML subgroups. As a matter of fact, a lower CR rate was found in patients with lower bax/bcl-2 ratio either within CD71+ (p=0.0009) or CD71- (p=0.0006). Also, a lower bax/bcl-2 ratio was associated with a shorter OS in CD71+ (p=0.008) and, more significantly, in CD71 negative (p=0.00008) patients. The independent prognostic value of bax/bcl-2 ratio was confirmed in multivariate analy-sis.
The overexpression of multidrug resistance (MDR) pro-teins (P-glycoprotein [PGP] [77], lung resistance protein [LRP] [78] and multidrug related protein [MRP] [79]) and the anti-apoptotic proteins (bcl-2, bcl-xl, Mcl-1) represent key mechanisms explaining the high rate of treatment failure in AML. Gene expression profiling studies have identified as discriminator genes for a poor prognosis genes involved ei-ther in multidrug resistance or in apoptosis block [29]. Moreover, the availability of both third-generation MDR inhibitors, such as zosuquidar [80] and pro-apoptotic drugs stimulated us to analyse the impact of multidrug resistance and apoptosis proteins on AML prognosis. Therefore, we tested a large series of 386 patients, affected by de novo AML, treated with intensive chemotherapy regimens
2-3.
The principal aims of our study were: 1) to correlate bax/bcl-2 ratio, as spontaneous apoptosis, with PGP, LRP and MRP expressions as multidrug resistance phenotype (MDR); 2) to demonstrate that bax/bcl-2 ratio and MDR are biologically and clinically critical events; and, finally 3) to clarify whether MDR overexpression is able to modify the favorable outcome of patients with high levels of spontane-ous apoptosis. A significant correlation was found between a higher PGP and a higher bax/bcl-2 ratio (p=0.02), demon-strating that a MDR1 phenotype is more frequent in patients with an elevated spontaneous apoptosis. On the contrary, MRP was more represented in patients with lower apoptosis levels (p=0.00005). With regard to clinical outcome, a sig-
2 Del Poeta, G.; Del Principe, M. I.; Venditti, A.; Buccisano, F.; Maurillo, L.; Mazzone,
C.; Neri, B.; Irno Consalvo, M.; Bruno, A.; Lo Coco, F.; Gattei, V.; Amadori, S. Clini-cal relevance of spontaneous apoptosis is independent from multidrug resistance in
acute myeloid leukemia (AML). Blood 2005, 106, Abstract 101. 3 Del Poeta, G.; Del Principe, M. I.; Buccisano, F.; Venditti, A.; Maurillo, L.; Ammatu-
na, E.; Bruno, A.; Neri, B.; Sarlo, C.; Lo Coco, F.; de Fabritiis, P.; Amadori, S. Multidrug resistance proteins overexpression worsens the favorable clinical impact of
an elevated spontaneous apoptosis in acute myeloid leukemia (AML). Blood 2006, 108, Abstract 656.
214 Current Cancer Drug Targets, 2008, Vol. 8, No. 3 Del Poeta et al.
nificant lower CR rate was found in patients with lower bax/bcl-2 ratio (p<0.00001) or higher MDR (p<0.00001). OS was significantly shorter either in patients with lower bax/bcl-2 ratio (p<0.00001) or higher MDR (p<0.00001). Bax/bcl-2 ratio and MDR showed additive prognostic im-pact, since higher bax/bcl-2 ratio plus lower MDR identified AML patients at better prognosis with regard to CR (p<0.00001), OS (p<0.00001) and DFS (p=0.0009). How-ever, AML patients showing lower bax/bcl-2 ratio without any MDR protein expression, presented also a short OS and DFS, confirming that an apoptosis block is enough to confer a worse prognosis alone. In order to establish whether MDR overexpression is able to worsen the favorable clinical out-come of AML pts with elevated apoptosis levels, we ana-lysed the AML subgroup with higher bax/bcl-2 ratio (224 pts). As a matter of fact, a lower CR rate was found in pa-tients with higher MDR expression (p=0.0005). The superior independent prognostic value of bax/bcl-2 ratio over MDR proteins was confirmed in multivariate analysis with regard to CR, OS and DFS. Therefore, apoptosis and multidrug re-sistance pathways are confirmed to be crucial prognostic factors in AML. In our opinion, low apoptosis and higher MDR in patients with elevated apoptosis identify the two main different biologic AML subsets at worse prognosis. Moreover, both multivariate analysis and capacity of lower bax/bcl-2 ratio without any MDR overexpression of identify-ing patients at worse prognosis implies that apoptosis has a more relevant clinical impact. However, it has to be taken in account that the favorable prognostic impact of a higher bax/bcl-2 ratio is greatly reduced by MDR positivity. Actu-ally, we observe that we have to focus future research and therapeutic strategies targeting in the first place apoptosis and then MDR proteins in AML. In a recent study, simulta-neous quantification by the reverse transcriptase multiplex
ligation-dependent probe amplification technique of 31 apoptosis-related transcripts in viable blasts from BM aspi-rates of 120 newly diagnosed AML patients was used both to construct an accurate prediction model for patient survival and to guide therapeutic modalities. High expression of seven genes, 3 anti-apoptotic (Bcl-2L2, BIRC4 and BIRC3) and 4 pro-apoptotic (BAXs, BAK1, MAP1 and BAX1) was associated with poor OS. However, the average expression of all 12 anti-apoptotic genes was associated with poor OS (p=0.029). The anti-apoptotic gene BIRC3 and the pro-apoptotic genes BAX1 and BMF optimally predicted OS [81].
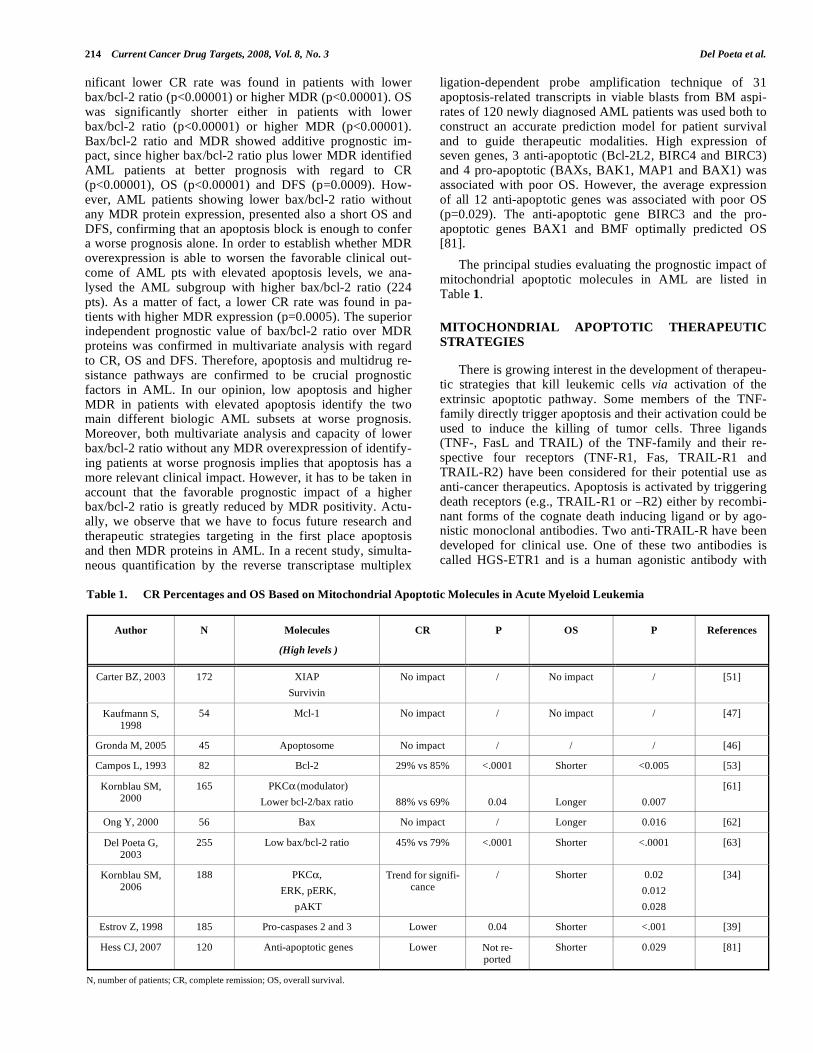
The principal studies evaluating the prognostic impact of mitochondrial apoptotic molecules in AML are listed in Table 1.
MITOCHONDRIAL APOPTOTIC THERAPEUTIC
STRATEGIES
There is growing interest in the development of therapeu-tic strategies that kill leukemic cells via activation of the extrinsic apoptotic pathway. Some members of the TNF-family directly trigger apoptosis and their activation could be used to induce the killing of tumor cells. Three ligands (TNF-, FasL and TRAIL) of the TNF-family and their re-spective four receptors (TNF-R1, Fas, TRAIL-R1 and TRAIL-R2) have been considered for their potential use as anti-cancer therapeutics. Apoptosis is activated by triggering death receptors (e.g., TRAIL-R1 or –R2) either by recombi-nant forms of the cognate death inducing ligand or by ago-nistic monoclonal antibodies. Two anti-TRAIL-R have been developed for clinical use. One of these two antibodies is called HGS-ETR1 and is a human agonistic antibody with
Table 1. CR Percentages and OS Based on Mitochondrial Apoptotic Molecules in Acute Myeloid Leukemia
Author N Molecules
(High levels )
CR P OS P References
Carter BZ, 2003 172 XIAP
Survivin
No impact / No impact / [51]
Kaufmann S, 1998
54 Mcl-1 No impact / No impact / [47]
Gronda M, 2005 45 Apoptosome No impact / / / [46]
Campos L, 1993 82 Bcl-2 29% vs 85% <.0001 Shorter <0.005 [53]
Kornblau SM, 2000
165 PKC (modulator)
Lower bcl-2/bax ratio
88% vs 69%
0.04
Longer
0.007
[61]
Ong Y, 2000 56 Bax No impact / Longer 0.016 [62]
Del Poeta G, 2003
255 Low bax/bcl-2 ratio 45% vs 79% <.0001 Shorter <.0001 [63]
Kornblau SM, 2006
188 PKC ,
ERK, pERK,
pAKT
Trend for signifi-cance
/ Shorter 0.02
0.012
0.028
[34]
Estrov Z, 1998 185 Pro-caspases 2 and 3 Lower 0.04 Shorter <.001 [39]
Hess CJ, 2007 120 Anti-apoptotic genes Lower Not re-ported
Shorter 0.029 [81]
N, number of patients; CR, complete remission; OS, overall survival.

Deregulation of the Mitochondrial Apoptotic Machinery Current Cancer Drug Targets, 2008, Vol. 8, No. 3 215
high affinity and specificity for TRAIL-R1 [82]. The HGS-ETR1 antibody (Human Genome Sciences) was evaluated in phase I/II clinical trial in patients with advanced solid or hematologic tumors, revealing little toxicity [83].
Moreover, apoptosis is induced or sensitivity to apoptotic stimuli is greatly increased by lowering the levels of the caspase-8 inhibitor c-FLIP through the addition of various drugs, such as the synthetic triterpenoid CDDO. CDDO has been shown to inhibit cellular proliferation and induce apop-tosis of leukemic cells [84].
A phase I study using an antisense to Bcl-2 in elderly AML patients showed promising results: in this study AML patients received induction with oblimersen, cytabarine and daunorubicin and, on achievement of complete remission, consolidation with oblimersen (antisense Bcl-2) and high-dose cytabarine. Of the 29 treated patients, 14 achieved a complete remission; during 12.6 months of follow-up, seven of these patients relapsed [85].
A phase II multicenter study of oblimersen plus gentu-zumab, a cytotoxic antibody directed against CD33+ cells, in relapsed AML patients has been reported: 12 out of 39 treated patients achieved a complete remission
4. Ten of the
12 responders survived more than 6 months. Finally, phar-macodynamic studies showed that therapy with oblimersen produced intracellular concentrations of the drug sufficient to down-regulate Bcl-2 [86]. The findings from these studies have been extended into an ongoing phase III study of about 400 patients conducted by CALGB with untreated AML who are receiving daunorubicin plus cytosine arabinoside with or without oblimersen.
Inhibition of BCL-2, using the oligonucleotide ODN Genasense, and inhibition of Bcl-2/Bcl-xL using small-molecule inhibitor ABT-737, results in a block of the anti-apoptotic function of these molecules and induction of mito-chondrial apoptosis [87]. A novel small molecule inhibitor, 4 - (3-methoxy-phenylsulphannyl) -7-nitro-benzofurazan-3-oxide (MNB), competes with the Bak BH3 peptide to bind Bcl-2 protein: 85% of HL-60 cells treated with 5 microM of MNB for 6 h were detected to undergo apoptosis. Further experiments have illustrated that MNB inhibits the het-erodimerization of Bcl-2 with Bax or Bid, reduces the mito-chondrial membrane potential (DeltaPsimt) and induces cy-tochrome c release from mitochondria in HL-60 cells. MNB seems to induce apoptosis in HL-60 cells by inhibiting the heterodimerization of Bcl-2 with pro-apoptosis Bcl-2 mem-bers, resulting in cytochrome c release, activation of caspases and PARP. MNB potently kills fresh AML and ALL cells, indicating that it has the potential to be developed to treat leukemia [88].
Disruption of Mdm2-p53 interaction activates p53 signal-ing, disrupts the balance of antiapoptotic and proapoptotic bcl-2 family proteins and induces apoptosis in AML [89]. Thus, functional inactivation of antiapoptotic bcl-2 proteins may enhance apoptogenic effects of Mdm2 inhibition. The combined targeting of Mdm2 by Nutlin-3a and Bcl-2 by
4 Moore, L. O.; Siter, K.; Kolitz, J. E.; Stock, W.; Yu, R.; Frankel, S. R. Phase 2 study
of oblimersen sodium G3139; Bcl-2 antisense; Genasense plus gemtuzumab ozogami-cin (Myelotarg) in elderly patients with relapsed acute myeloid leukaemia (AML).
Blood 2004, 104, Abstract 865.
ABT-737 were recently investigated by Kojima et al. [90]. Nutlin-3a and AB-737 induced Bax conformational change and mitochondrial apoptosis in AML cells in a strikingly synergistic fashion. Nutlin-3a induced p53-mediated apopto-sis predominantly in S and G2/M cells, while ABT-737 in-duced apoptosis predominantly in G1. The complementary effects of Nutlin-3a and ABT-737 in different cell cycle phases could account for their synergistic activity. The com-bined targeting of Mdm2 and Bcl-2 proteins could offer con-siderable therapeutic promise in AML [90].
Nitric oxide (NO) induces differentiation and apoptosis in AML cells. The NO prodrug JS-K has potent antileukemic activity inducing apoptosis in HL-60 cells by a caspase-dependent mechanism. JS-K alters DeltaPsim and induces cytochrome c release from mitochondria into the cytoplasm. Treatment with JS-K resulted in activation of Caspase (Casp) 9, Casp 3 and Casp 8. JS-K constitutes a promising lead for a new class of anti-leukemic agents [91].
Following cytochrome c release, the next step in the apoptotic process is apoptosome formation. Correct apopto-some formation requires APAF-1. The primary structure of APAF-1, a key member of the apoptosome, consists of an amino-terminal domain with 12 or 13 WD-40 repeats, a cen-tral ATPase domain with homology to CED-4, and a car-boxy-terminal caspase recruitment domain (CARD). Four major isoforms of the protein have been identified: only iso-forms containing an additional WD-40 repeat between re-peats 5 and 6 are able to activate caspase-9 in response to cytochrome c and ATP.
Defects in apoptosome proteins, such as APAF-1, have been involved in the development of some human malignan-cies, including AML. Studies carried out in leukemic cell lines, as well as in few cases of fresh primary leukemic cells, have shown that leukemic cells express low levels of APAF-1 in a significant proportion of cases and that their pattern of APAF-1 expression directly correlates with ultraviolet light-induced apoptosis [92]. Recent studies showed that 25% of human leukemic cell lines and 42% of primary AML cells express low levels of APAF-1 mRNA, due to methylation silencing of the APAF-1 gene.121(104) Treatment of leuke-mic cells with 5-aza-2’-deoxycytidine, a specific inhibitor of DNA methylation, restored APAF-1 expression in leukemic cells [93].
Two approaches are currently being used to develop XIAP inhibitors: (i) antisense oligonucleotides that are in clinical trials and (ii) small molecule inhibitors that are in preclinical development [94]. Inhibition of the XIAP-mediated block on active caspase-9, using synthetic SMAC peptides, markedly increases the sensitivity of AML cells to apoptotic stimuli. Interestingly, polyphenylurea-based small-molecule XIAP antagonists induce apoptosis of AML cells at low micromolar concentrations through a mechanism involv-ing activation of downstream caspase-3 and -7, independ-ently of Bcl-2 levels [95]. Triptolide, a diterpenoid isolated from the Chinese herb Tripterygium Wilfordii, decreases XIAP and potently induces caspase-dependent apoptosis of primary AML blasts mediated through the mitochondrial pathway, at low nanomolar concentrations. Triptolide also decreases Mcl-1, but not Bcl-2 and Bcl-xl levels. Further, triptolide induces loss of the mitochondrial membrane poten-

216 Current Cancer Drug Targets, 2008, Vol. 8, No. 3 Del Poeta et al.
tial and cytochrome c release [96]. Moreover, melissoidesin G, a diterpenoid purified from Isodon melissoides, a plant used in Chinese traditional medicine as antitumor and anti-inflammatory agent, was shown to specifically inhibit the growth of leukemia cell lines and primary acute myeloid leukemia blasts via induction of apoptosis, with evidence of mitochondrial DeltaPsim loss, reactive oxygen species pro-duction, caspase activation and nuclear fragmentation [97].
Combined exposure of U937 leukemia cells to toxic con-centrations of histone deacetylase inhibitor vorinostat and the cyclin-dependent kinase inhibitor flavopiridol resulted in a marked increase in mitochondrial damage and apoptosis accompanied by pronounced reductions in XIAP and Mcl-1 mRNA and protein. Down-regulation of Mcl-1 and XIAP expression by vorinostat/flavopiridol was associated with enhanced inhibition of phosphorylation of RNA polymerase II and was amplified by caspase-mediated protein degrada-tion. Flavopiridol/vorinostat-mediated transcriptional repres-sion of XIAP, Mcl-1 enhanced apoptosis also occurred in primary acute myelogenous leukemia blasts [98].
The proapototic IAP inhibitor, LBW242 was shown in proliferation studies done in vitro to enhance the killing of PKC412-sensitive and PKC412-resistant cell lines express-ing mutant FLT3 when combined with either PKC412 or standard cytotoxic agents (doxorubicin and Ara-C) [99].
Noteworthy, Carter et al. [100] demonstrated that target-ing surviving expression with a survivin anti-sense oligonu-cleotide (sur-AS-ODN (ISIS 23722) causes increases in sub-G1 phase and annexin V+ cells with subsequent cell death involving mitochondrial release of cytochrome c and Smac and the nuclear translocation of AIF.
Inhibition of the proteasome, using proteasome inhibitors such as bortezomib, deregulates protein homeostasis, leading to activation of mitochondrial apoptosis through stabilization and activation of the BH3-only proteins Bim and Bik. Pro-teasome inhibitors might increase sensitivity to chemother-apy for AML. Idarubicin with a low concentration of MG132 or bortezomib interacted synergistically to induce apoptosis in THP-1 leukemic cells. Western blot analysis of Bax and Bim show an accumulation of these pro-apoptotic proteins in THP-1 treated cells. Proteasome inhibition also potentiated IDA-induced apoptosis in primary blast cells from 22 AML patients while no such effect was found on normal lympho-cytes [101].
The tyrphostin adaphostin interacts synergistically with proteasome inhibitors (eg, MG-132 and bortezomib) induc-ing mitochondrial damage (eg, cytochrome c release), caspase activation, and apoptosis in human leukemia cells. These interactions were associated with a marked increase in oxidative damage (eg, ROS generation), down-regulation of the Raf/MEK/ERK pathway, and JNK activation. The adaphostin/MG-132 regimen displayed toxicity toward 5 primary AML samples, but not normal hematopoietic pro-genitors [102]. CD34+ bone marrow blasts from higk-risk myelodysplastic syndrome (MDS) or AML patients constitu-tively activate the nuclear factor-kappaB (NF-kappaB) path-way and undergo apoptosis when NF-kappaB is inhibited. BAY11-7082, which targets the NF-kappaB-activating I-kappaB kinase complex, combined with nutrient (glucose,
insulin, etc) depletion led primary CD34+ bone marrow blasts from AML to cell death accompanied by all hallmark of apoptosis, including an early loss of the mitochondrial transmembrane potential, release of cytochrome c and AIF from mitochondria, activation of caspase-3, phosphatidylser-ine exposure on the plasma membrane surface and nuclear chromatin condensation [103]. In fact, AML cells are charac-terized by a constitutive and abnormal activation of the NF-kappaB transcription factor. Targeting NF-kappaB activation via pharmacologic inhibition of IKB chinase 2 (IKK2) with AS602868 led to apoptosis of human primary AML cells. Moreover, AS602868 potentiated the apoptotic response induced by doxorubicin, cytarabine or etoposide. AS602868-induced cell death was associated with the rupture of the mitochondrial transmembrane potential and activation of cellular caspases [104]. NF-kappaB inhibition did not affect normal CD34+ hematopoietic precursors, suggesting that it could represent a new adjuvant strategy for AML. Moreover, inhibition of NEMO, the regulatory subunit of the IkappaB kinase (IKK) complex crucial for NF-kappaB activation, by a cell-permeable peptide induces apoptosis in high-risk mye-lodysplastic syndrome and AML. The apoptotic mechanism involved the caspase-independent loss of the mitochondrial transmembrane potential as well as signs of outer mitochon-drial membrane permeabilization with the consequent release of cytochrome c, apoptosis-inducing factor and endonuclease G [105].
The aberrant function of transcription factors and kinase-based signaling pathways that regulate the ability of hema-topoietic cells to proliferate, differentiate and escape apopto-sis accounts for the leukemic transformation of myeloid pro-genitors. Milella et al. [106] demonstrated that simultaneous retinoid receptor ligation and blockade of the MEK/ERK signaling module, using the small molecule inhibitor CI-1040, result in a synergistic induction of apoptosis in both AML and acute promyelocytic leukemia (APL) cells with constitutive ERK activation. The Authors identified disrup-tion of Bcl-2-dependent mitochondrial homeostasis as a pos-sible point of convergence for the proapoptotic synergism observed with retinoids and MEK inhibitors.
Gemtuzumab ozogamicin (GO, Mylotarg ) is an immmunoconiugate targeted against CD33, which is ex-pressed on more than 90% of myeloid leukemic blasts. The antibody is attached to calicheamicin, a potent cytotoxic enediyne antibiotic that inhibits DNA synthesis and induces apoptosis activating caspase-3 [107]. This agent is currently being investigated in many clinical trials as front-line ap-proach in previously untreated AML. Major cooperative groups are conducting phase-3 studies that incorporate gen-tuzumab ozogamicin as part of standard chemotherapy in patients newly diagnosed with AML [108].
Erlotinib, an inhibitor of the epidermal growth factor re-ceptor (EGFR) induces differentiation, cell cycle arrest and apoptosis of EGFR-negative myeloblasts of patients with myelodysplastic syndrome (MDS) and acute myeloid leuke-mia (AML), as well as in EGFR-negative cell lines repre-senting these diseases (P39, KG-1 and HL-60). In apoptosis-sensitive AML cells, erlotinib caused a rapid nucleo-cytoplasmic translocation of nucleophosmin-1 (NPM-1) and p14 (ARF). Apoptosis-insensitive myeloblasts failed to

Deregulation of the Mitochondrial Apoptotic Machinery Current Cancer Drug Targets, 2008, Vol. 8, No. 3 217
manifest this translocation, yet they became sensitive to apoptosis induction by erlotinib when NPM-1 was depleted by RNA interference. Erlotinib also killed CD34+ bone mar-row blasts from MDS and AML patients while sparing nor-mal CD34+ progenitors. This ex vivo therapeutic effect was once more associated with the nucleocytoplasmic transloca-tion of NPM-1 and p14(ARF). One patient afflicted with both MDS and non-small cell lung cancer manifested hema-tological improvement in response to erlotinib [109].
Finally, the proto-oncogene c-Myc plays an important role in the control of cell proliferation, apoptosis and differ-entiation, and its aberrant expression is frequently seen in multiple human cancers, including AML. C-Myc heterodi-merizes with Max to transactivate target genes in leu-kemogenesis. A small molecule compound, 10058-F4 inhib-its c-My/Max heterodimerization and represents a potential antileukemic approach. Huang et al. [110] showed that 10058-F4 arrested AML cells at G0/G1 phase, downregu-lated c-Myc and upregulated CDK inhibitors, p21 and p27. Meanwhile, 10058-F4 induced apoptois through activation of mitochondrial pathway shown by downregulation of Bcl-2, upregulation of Bax, release of cyoplasmic cytochrome c, and cleavage of caspase 3, 7 and 9.
The main molecules or drugs which trigger apoptotic pathways are summarized in Table 2.
CONCLUSIONS
AML is an aggressive hematologic malignancy. The de-velopment and progression of the leukemic disease invaria-bly involve deregulation in the apoptotic response. Despite
major efforts during the past 40 years, limited improvements in disease survival have been made. The current primary treatment for AML is chemotherapy, which induces cell death by apoptosis mainly mediated through the intrinsic mitochondrial pathway but also through the extrinsic death receptor pathway, both of which lead to caspase activation and cell death. The progress made in our understanding of the central role of apoptosis deregulation in the pathogenesis of leukemia and in the development of chemoresistance has launched the academic community, as well as the pharma-ceutical and biotechnology industries into a search to iden-tify new compounds for the treatment of leukemia. Targeted therapies that are designed to induce apoptosis in leukemic cells are currently the most promising anti-leukemic strate-gies. These strategies are aimed at targeting and killing the leukemia cells specifically with no or limited collateral dam-age to normal hematopoietic progenitor cells. The elucida-tion of the molecular apoptotic machinery and of its defects in AML lays the basis for developing new drugs able to trig-ger apoptosis of leukemia cells.
ACKNOWLEDGEMENTS
This study was in part supported by a Finalized Project, 2005 of Italian Ministry of University and Research. We realize that, due to the limitation of space, we could not cite all the excellent contribution published above the prognostic and therapeutic implications of molecules correlated with mitochondrial apoptosis, and we apologize for omission of some papers and reviews from our national and international colleagues.
Table 2. Apoptosis-Triggering Strategies in Acute Myeloid Leukemia
Pathway Target Author Drug Stage of development
References
TRAIL Pukac HGS-ETR1 Phase I [82] Receptor-mediated
Caspase-8
c-FLIP
Konopleva CDDO Preclinical [84]
Bcl-2 Marcucci ODN Genasense Phase I/II/III [85]
Bak Zhang MNB Preclinical [88]
Bcl-2/Bcl-xl
MDM-2 and Bcl-2
Kojima
ABT-737
Nutlin-3a +
Phase I [89]
Cytochrome c and caspases 3, 9,8
Udupi JS-K (NO prodrug) Preclinical [91]
Mitochondrial-
mediated
APAF-1 (Apoptosome) Furukawa 5-aza-2’-deoxycytidine Phase II [93]
XIAP
Schimmer
Carter
Rosato
Antisense oligonucleotides
Triptolide
Flavopiridol/Vorinostat
Preclinical
Preclinical
Preclinical
[94]
[96]
[98]
IAP Weisberg LBW242 Preclinical [99]
Inhibitors apoptosis
proteins
Survivin Carter ODN-survivin Preclinical [100]
Proteasome and cy-tocrome c
Dasmahapatra PS-341+ MG132 Prceclinical [102]
NF- B Fabre BAY11-7082 Preclinical [103]
NPM-1/ARF
Boherer Erlotinib Preclinical [109]
Oncogenes
c-Myc Huang 10058-F4 Preclinical [110]

218 Current Cancer Drug Targets, 2008, Vol. 8, No. 3 Del Poeta et al.
ABBREVIATIONS
ABCG2 = ATP binding cassette G2 or BCRP (breast cancer resistance protein)
AIF = apoptosis inducing factor
AKT = protein kinase B
ALL = acute lymphoid leukemia
AML = acute myeloid leukemia
APAF-1 = apoptosis protease activating factor 1
APK = activated protein kinases
APL = acute promyelocytic leukemia
ARF = ADP ribosylation factors
ATRA = all-trans retinoic acid
Bad = bcl-2 associated death promoter
Bak = bcl-2 homologous antagonist killer
Bax = bcl-2 associated x protein
Bcl-2 = B-cell lymphoma 2
BCR-ABL = breakpoint cluster gene-Abelson
BH = bcl-2 homology domains
Bid = BH3 interacting domain death agonist
Bik = bcl-2 interacting killer
Bim = bcl-2 –like 11 apoptosis facilitator
BIRC = Baculoviral IAP repeat-containing
BM = bone marrow
Bmf = bcl-2 modifying factor
CALGB = Cancer and Leukemia Group B
CARD = caspase activation recruitment domain
CASP = Caspase
CBF-Beta/ = core binding factor beta/smooth muscle my SMMHC osin heavy chain
PPP1R15A = protein phosphatase 1, regulatory (inhibitor) subunit 15A
CDDO = 2-cyano-3,12-dioxooleana-1,9(11)-dien-28-oic acid
CDK = Cyclin dependent kinase
CED-4 = caEnorhabditis Death -4
c-FLIP = cellular FLICE [FADD-like IL-1beta con-verting enzyme] inhibitory protein
CR = complete remission
dATP = deoxyadenosine-5-triphosphate
DEVD-AFC = 7-amino-4 T Trifluoromethylcoumarin,N-CBZ-L aspartyl-L-glutamyl-valyl-l-aspartic acid amide
DFS = disease free survival
DISC = death-induced signaling complex
EGFR = epidermal growth factor receptor
ERK = extracellular signal-related kinase
FasL = Fas ligand
FLICE = Fas-associated death domain like interleukin 1beta converting enzyme
FLT3 = FMS-like tyrosine kinase 3
GO = gentuzumab ozogamicin
GTPase = guanosin triphosphate hydrolases
HL-60 = human promyelocytic leukemia cell line
IAP = inhibitor of apoptosis protein
IKK = IkappaB kinase
IL10R = interleukin 10 receptor
IRF4 = interferon regulatory factor 4
ITD = internal tandem duplication
JAK = Janus kinase
JNK = Jun N-terminal kinase
JS-K = O2-(2,4-dinitrophenyl)1-[(4-ethoxycarbonyl)piperazin-1-yl]diazen-1-ium-1,2-diolate
LRP = lung resistance protein
LTBP1 = Latent TGF [transforming growth factor] beta binding protein
M2 FAB = acute myeloid leukemia subtype according to the french-american-british classification
MAP1 = modulator of apoptosis
MAPK = mitogen-activated protein kinase
Max = myc associated factor X
Mcl-1 = myeloid cell leucemia sequence 1
MDM = mouse double minute 2
MDR = multidrug resistance
MDS = myelodysplastic syndrome
MEK = mitogen activated protein kinase kinase
MFI = mean fluorescence intensity
MLL = mixed lineage leukaemia gene
MNB = 4-(3 methoxy-phenylsulphannyl)-7-nitro-be-nzofurazan-3-oxide
MOMP = mitochondrial outer membrane permeabili-zation
MPB = mobilized peripheral blood
MRP = multidrug related protein
mTOR = mammalian target of rapamycin
NEMO = NF-kappaB-essential modulator
NF-kappaB = nuclear factor-kappaB
NO = nitric oxide
NOXA = latin for damage

Deregulation of the Mitochondrial Apoptotic Machinery Current Cancer Drug Targets, 2008, Vol. 8, No. 3 219
NPM1 = nucleophosmin 1
ODN = oligonucleotide antisense
OS = overall survival
PARP = Poly (ADP-ribose) polymerase
PGP = p-glycoprotein
PI3K = Phosphatidylinositol 3’-kinase
Pim-1 = Pim-1 oncogene
PKC = protein kinase C
PML = promyelocytic leukemia protein
pts = patients
PUMA = p53 upregulated modulator of apoptosis
RAR = retinoid acid receptor
ROS = reactive oxygen species
Smac/ = second mitochondria derived activator of DIABLO caspases
STAT = signal transducer S and activators of tran-scription protein
THP-1 = human acute monocytic leucemia cell line
TNF = tumor necrosis factor
TNF-R1 = Tumor necrosis factor-receptor 1
TRAIL = tumor related apoptosis-inducing ligand
TRAIL-R1 = Tumor necrosis factor related apoptosis in-ducing ligand-receptor 1
UCB = umbilical cord blood
VDAC = voltage dependent anion channel
WAF1 = cell cycle regulatory gene p21
WD 40 = Apaf-1 isoform
XIAP = X-linked inhibitor of apoptosis protein
REFERENCES
[1] Champlin, R.; Gale, R. P. Acute myelogenous leukemia: Recent
advances in therapy. Blood 1987, 69, 1551-1562.
[2] Taylor, P. R.; Reid, M. M.; Stark, A. N.; Brown, N.; Hamilton, P.
J.; Proctor, S. J. De novo acute myeloid leukemia in patients over
55-years old: a population based study of incidence, treatment and
outcome. Leukemia 1995, 9, 231-237.
[3] Kerr, J. F.; Winterford, C. M.; Harmon, B. V. Apoptosis: its sig-
nificance in cancer and cancer therapy. Cancer 1994, 73, 2013-
2026.
[4] Goldstein, J. C.; Waterhouse, N. J.; Juin, P.; Evan, G. I.; Green, D.
R. The coordinate release of cytochrome c during apoptosis is
rapid, complete and kinetically invariant. Nat. Cell. Biol. 2000, 2,
156-162.
[5] Thornberry, N. A.; Lazebnik, Y. Caspases:enemies within. Science
1998, 281, 1312-1316.
[6] Green, D. R.; Reed, J. C. Mitochondria and apoptosis. Science
1998, 281, 1309-1312.
[7] Narita, M.; Shimizu, S.; Ito, T.; Chittenden, T.; Lutz, R. J.; Ma-
tsuda, H.; Tsujimoto, Y. Bax interacts with the permeability transi-
tion pore to induce permeability transition and cytochrome c re-
lease in isolated mitochondria. Proc. Natl. Acad. Sci. USA 1998,
95, 14681-14686.
[8] Brenner, C.; Cadiou, H.; Vieira, H. L.; Zamzani, N.; Marzo, I.; Xie,
Z.; Leber, B.; Andrews, D.; Duclohier, H.; Reed, J. C.; Kroemer,
G. Bcl-2 and Bax regulate the channel activity of the mitochondrial
adenine nucleotide translocator. Oncogene 2000, 19, 329-336.
[9] Oltvai, Z. N.; Milliman, C. L.; Korsmeyer, K. Bcl-2 heterodimer-
izes in vivo with a conserved homolog, Bax, that accelerates pro-
grammed cell death. Cell 1993, 74, 609-619.
[10] Frohling, S.; Scholl, C.; Gilliland, D. G.; Levine, R. L. Genetics of
myeloid malignancies: pathogenetic and clinical implications. J.
Clin. Oncol. 2005, 23, 6285-6295.
[11] Klampfer, L.; Zhang, J.; Zelenetz, A. O.; Uchida, H.; Nimer, S. D.
The AML1/ETO fusion protein activates transcription of BCL-2.
Proc. Natl. Acad. Sci. USA 1996, 95, 11863-11868.
[12] Britos-Bray, M.; Ramirez, M.; Cao, W.; Wang, X.; Liu, P. P.;
Civin, C. I.; Friedman, A. D. CBF-SMMHC, expressed in M4eo
acute myeloid leukaemia, reduces p53 induction and slows apopto-
sis in hematopoietic cells exposed to DNA-damaging agents. Blood
1998, 92, 4344-4352.
[13] Pandolfi, P. P. Oncogenes and tumor suppressors in the molecular
pathogenesis of acute promyelocytic leukaemia. Hum. Mol. Genet.
2001, 10, 769-775.
[14] Adler, H. T.; Chinery, R.; Wu, D. Y.; Kussick, S. J.; Payne, J. M.;
Fornace, A. J. Jr.; Tkachuk, D. C. Leukemic HRX fusion proteins
inhibit GADD34-induced apoptosis and associate with the
GADD34 and hSNF5/INH proteins. Mol. Cell. Biol. 1999, 19,
7050-7060.
[15] Testa, U.; Grignani, F.; Samoggia, P.; Zanetti, C.; Riccioni, R.; Lo
Coco, F.; Diverio, D.; Felli, N.; Passerini, C. G.; Grell, M.; Pelicci,
P. G.; Peschle, C. The PML/RAR_ fusion protein inhibits tumor
necrosis factor--induced apoptosis in U937 cells and acute promye-
locytic leukemia blasts. J. Clin. Invest. 1998, 101, 2278-2289.
[16] Riccioni, R.; Pasquini, L.; Mariani, G.; Saulle, E.; Rossigni, A.;
Diverio, D.; Pelosi, E.; Vitale, A.; Chierichini, A.; Cedrone, M.;
Foà, R.; Lo Coco, F.; Peschle, C.; Testa, U. TRAIL decoy receptors
mediate resistance of acute myeloid leukemia cells to TRAIL.
Haematologica 2005, 90, 612-624.
[17] Falini, B.; Mecucci, C.; Tiacci, E.; Alcalay, M.; Rosati, R.; Pasqua-
lucci, L.; La Starza, R.; Diverio, D.; Colombo, E.; Cantucci, A.;
Bigerna, B.; Pacini, R.; Pucciarini, A.; Liso, A.; Vignetti, M.; Fazi,
P.; Meani, N.; Pettirossi, V.; Saglio, G.; Mandelli, F.; Lo-Coco, F.;
Pelicci, P. G.; Martelli, M. F. GIMEMA Acute Leukemia Working
Party. Cytoplasmic nucleophosmin in acute myelogenous leukemia
with a normal karyotype. N. Engl. J. Med. 2005, 352, 254-266.
[18] Verhaak, R. G.; Goudswaard, C. S.; Lowenberg, B.; Valk, P. J.
Mutations in nucleophosmin (NPM1) in acute myeloid leukaemia
(AML): association with other gene abnormalities and previously
established gene expression signatures and their favourable prog-
nostic significance. Blood 2005, 106, 3747-3754.
[19] Lambert, B.; Buckle, M. Characterization of the interface between
nucleophosmin (NPM) and p53: potential role in p53 stabilization.
FEBS Lett. 2006, 6580, 345-350.
[20] Stirewalt, D. L.; Radich, J. P. The role of FLT3 in hemopoietic
malignancies. Nat. Rev. Cancer 2003, 3, 650-665.
[21] Minami, Y.; Yamamoto, K.; Kiyoi, H.; Ueda, R.; Saito, H.; Naoe,
T. Different antiapoptotic pathways between wild-type and mutated
FLT3: insights into therapeutic targets in leukemia. Blood 2003,
102, 2969-2975.
[22] Kohl, T. M.; Schnittger, S.; Ellwart, J. W. Kit exon 8 mutations
associated with core-binding factor (CBF)-acute myeloid leukemia
(AML) cause hyperactivation of the receptor in response to stem
cell factor. Blood 2005, 105, 3319-3321.
[23] Schnittger, S.; Kohl, T. M.; Haferlach, T.; Kern, W.; Hiddemann,
W.; Spiekermann, K.; Schoch, C. KIT-816 mutations in AML1-
ETO-positive AML are associated with impaired event-free and
overall survival. Blood 2006, 107, 1790-1799.
[24] Paschka, P.; Marcucci, G.; Ruppert, A. S.; Mrózek, K.; Chen, H.;
Kittles, R. A.; Vukosavljevic, T.; Perrotti, D.; Vardiman, J. W.;
Carroll, A. J.; Kolitz, J. E.; Larson, R. A.; Bloomfield, C. D.
Cancer and Leukemia Group B. Adverse prognostic significance of
KIT mutations in adult acute myeloid leukaemia with inv(16) and
t(8;21): a cancer and leukaemia group B study. J. Clin. Oncol.
2006, 24, 3904-3911.
[25] Reuter, C. W.; Morgan, M. A.; Bergmann, L. Targeting the Ras
signaling pathway. A rational mechanism-based treatment for he-
matologic malignancies? Blood 2000, 96, 1665-1669.

220 Current Cancer Drug Targets, 2008, Vol. 8, No. 3 Del Poeta et al.
[26] Bacher, U.; Haferlach, T.; Schoch, C.; Kern, W.; Schnittger, S.
Implications of NRAS mutations in AML: a study of 2502 patients.
Blood 2006, 107, 3847-3853.
[27] Traver, D.; Akashi, K.; Weissman, I. L.; Lagasse, E. Mice defec-
tive in two apoptosis pathways in the myeloid lineage develop
acute myeloblastric leukaemia. Immunity 1998, 9, 47-57.
[28] Bullinger, L.; Döhner, K.; Bair, E.; Fröhling, S.; Schlenk, R. F.;
Tibshirani, R.; Döhner, H.; Pollack, J. R. Use of gene-expression
profiling to identify prognostic subclasses in adult myeloid leu-
kaemia. N. Engl. J. Med. 2004, 350, 1605-1616.
[29] Wilson, C. S.; Davidson, G. S.; Martin, S. B.; Andries, E.; Potter,
J.; Harvey, R.; Ar, K.; Xu, Y.; Kopecky, K. J.; Ankerst, D. P.;
Gundacker, H.; Slovak, M. L.; Mosquera-Caro, M.; Chen, I. M.;
Stirewalt, D. L.; Murphy, M.; Schultz, F. A.; Kang, H.; Wang, X.;
Radich, J. P.; Appelbaum, F. R.; Atlas, S. R.; Godwin, J.; Willman,
C. L. Gene expression profiling of adult acute myeloid leukaemia
identifies novel biologic clusters for risk classification and outcome
prediction. Blood 2006, 108, 685-696.
[30] Min, Y. H.; Cheong, J. W.; Kim, J. Y. Cytoplasmic mislocalization
of p27 Kip 1 protein is associated with constitutive phosphorylation
of Akt or protein kinase B and poor prognosis in acute myeloge-
nous leukaemia. Cancer Res. 2004, 64, 5225-5231.
[31] Martelli, A. M.; Nyakjern, M.; Bortul, R.; Tarzani, P. L.; Eva-
lengelisti, C.; Cocco, L. Phosphoinositide 3-kinase/Aky signaling
pathway and its therapeutical implications for human acute myeloid
leucemia. Leukemia 2006, 20, 911-928.
[32] Xu, Q.; Simpson, S. E.; Scialla, T. J.; Bagg, A.; Carroll, M. Sur-
vival of acute myeloid leukemia cell requires PI3 kinase activation.
Blood 2003, 102, 972-980.
[33] Milella, M.; Kornblau, S. M.; Estrov, Z. Therapeutic targeting of
the MEK/MAPK signal transduction module in acute myeloid leu-
kemia. J. Clin. Invest. 2001, 108, 851-859.
[34] Kornblau, S. M.; Womble, M.; Qiu, Y. H.; Jackson, C. E.; Chen,
W.; Konopleva, M.; Estey, E. H.; Andreeff, M. Simultaneous acti-
vation of multiple signal transduction pathways confers poor prog-
nosis in acute myelogenous leukemia. Blood 2006, 108, 2358-2365.
[35] Moriai, R.; Asanuma, K.; Kobayashi, D.; Yajima, T.; Yagihashi,
A.; Yamada, M.; Lin, C. W.; Manshouri, T.; Jilani, I.; Neuberg, D.;
Patel, K.; Kantarjian, H.; Andreeff, M.; Estrov, Z.; Beran, M.;
Keating, M.; Estey, E.; Albitar, M. Quantitative analysis of the the
anti-apoptotic gene survivin expression in malignant haema-
topoietic cells. Anticancer Res. 2001, 21, 595-600.
[36] Gesbert, F.; Griffin, J. Bcr/Abl activates transcription of Bcl-X
gene through STAT 5. Blood 2000, 96, 2269-2276.
[37] Invernizzi, R.; Pecci, A.; Bellotti, L.; Ascari, E. Expression of p53,
bcl-2 and ras oncoproteins and apoptosis levels in acute leukaemias
and myelodysplastic syndromes. Leuk. Lymphoma 2001, 42, 481-
489.
[38] Lin, C. W.; Manshouri, T.; Jilani, I.; Neuberg, D.; Patel, K.; Kan-
tarjian, H.; Andreeff, M.; Estrov, Z.; Beran, M.; Keating, M.;
Estey, E.; Albitar, M. Proliferation and apoptosis in acute and
chronic leukemias and myelodysplastic syndrome. Leuk. Res. 2002,
26, 551-559.
[39] Estrov, Z.; Thall, P. F.; Talpaz, M.; Estey, E. H.; Kantarjian, H. M.;
Andreeff, M.; Harris, D.; Van, Q.; Walterscheid, M.; Kornblau, S.
M. Caspase 2 and caspase 3 protein levels as predictors of survival
in acute myelogenous leukaemia. Blood 1998, 92, 3090-3097.
[40] Green, D. R.; Evan, G. I. A matter of life and death. Cancer Cell
2002, 1, 19-30.
[41] Barry, M.; Heibein, J. A.; Pinkoski, M. J.; Lee, S. F.; Moyer, R.
W.; Green, D. R.; Bleackley, R. C. Granzyme B short-circuits the
need for Caspase 8 activity during granulemediated cytotoxic T-
lymphocyte killing by directly cleaving Bid. Mol. Cell Biol. 2000,
20, 3781-3794.
[42] Debatin, K. M.; Poncet, D.; Kroemer, G. Chemotherapy: Targeting
the mitochondrial cell death pathway. Oncogene 2002, 21, 8786-
8803.
[43] Kluck, R. M.; Bossy-Wetzel, E.; Green, D. R.; Newmeyer, D. D.
The release of cytochrome c from mitochondria: A primary site for
Bcl-2 regulation of apoptosis. Science 1997, 275, 1132-1136.
[44] Hu, Y.; Benedict, M.; Ding, L.; Nunez, G. Role of cytochrome c
and dATP/ATP hydrolysis in Apaf-1-mediated caspase-9 activation
and apoptosis. EMBO J. 1999, 18, 3586-3595.
[45] Saleh, A.; Srinivasula, S.; Acharya, S.; Fishel, R.; Alnemri, E.
Cytochrome c and dATP-mediated oligomerization of Apaf-1 is a
prerequisite for procaspase-9 activation. J. Biol. Chem. 1999, 274,
17941-17945.
[46] Gronda, M.; Brandwein, J.; Minden, M. D.; Pond, G. R.; Schuh, A.
C.; Wells, R. A.; Messner, H.; Chun, K.; Schimmer, A. D. Assess-
ment of the downstream portion of the mitochondrial pathway of
caspase activation in patients with acute myeloid leukemia. Apop-
tosis 2005, 10, 1285-1294.
[47] Kaufmann, S. H.; Karp, J. E.; Svingen, P. A.; Krajewski, S.; Burke,
P. J.; Gore, S. D.; Reed, J. C. Elevated expression of the apoptotic
regulator Mcl-1 at the time of leukemic relapse. Blood 1998, 91,
991-1000.
[48] Konopleva, M.; Zhao, S.; Hu, W.; Jiang, S.; Snell, V.; Weidner, D.;
Jackson, C. E.; Zhang, X.; Champlin, R.; Estey, E.; Reed, J. C.;
Andreeff, M. The antiapoptotic genes Bcl-X(L) and Bcl-2 are over-
expressed and contribute to chemoresistance of non-proliferating
leukaemic CD34+ cells. Br. J. Haematol. 2002, 118, 521-534.
[49] Bagrintseva, K.; Geisenhof, S.; Kern, R.; Eichenlaub, S.; Reindl,
C.; Ellwart, J. W.; Hiddemann, W.; Spiekermann, K. FLT3-ITD-
TKD dual mutants associated with AML confer resistance to FLT3
PTK inhibitors and cytotoxic agents by overexpression of Bcl-
X(L). Blood 2005, 105, 3679-3685.
[50] Tamm, I.; Kornblau, S. M.; Segall, H.; Krajewski, S.; Welsh, K.;
Kitada, S.; Scudiero, D. A.; Tudor, G.; Qui, Y. H.; Monks, A.; An-
dreeff, M.; Reed, J. C. Expression and prognostic significance of
IAP-family genes in human cancers and myeloid leukemias. Clin.
Cancer Res. 2000, 6, 1796-1803.
[51] Carter, B. Z.; Kornblau, S. M.; Tsao, T.; Wang, R. Y.; Schober, W.
D.; Milella, M.; Sung, H. G.; Reed, J. C.; Andreeff, M. Caspase-
independent cell death in AML: caspase inhibition in vitro with
pan-caspase inhibitors or in vivo by XIAP or Survivin does not af-
fect cell survival or prognosis. Blood 2003, 102, 4179-4186.
[52] Stoetzer, O. J.; Nüssler, V.; Darsow, M.; Gullis, E.; Pelka-
Fleischer, R.; Scheel, U.; Wilmanns, W. Association of bcl-2, bax,
bcl-xl and interleukin-1-beta-converting enzyme expression with
initial response to chemotherapy in acute myeloid leukemia. Leu-
kemia 1996, 10, S18-S22.
[53] Campos, L.; Rouault, J. P.; Sabido, O.; Oriol, P.; Roubi, N.; Vas-
selon, C.; Archimbaud, E.; Magaud, J. P.; Guyotat, D. High expres-
sion of bcl-2 protein in acute myeloid leukemia is associated with
poor response to chemotherapy. Blood 1993, 81, 3091-3096.
[54] Haldar, S.; Jena, N.; Croce, C. M. Inactivation of Bcl-2 by phos-
phorylation. Proc. Natl. Acad. Sci. USA 1995, 92, 4507-4511.
[55] Ito, T.; Deng, X.; Carr, B.; May, W. S. Bcl-2 phosphorylation re-
quired for anti-apoptosis function. J. Biol. Chem. 1997, 272,
11671-11673.
[56] Srivastava, R.; Mi, Q.; Hardwick, J.; Longo, D. Deletion of the
loop region of Bcl-2 completely blocks palcitaxel-induced apopto-
sis. Proc. Natl. Acad. Sci. USA 1999, 96, 3775-3780.
[57] Hu, Z. B.; Minden, M. D.; McCulloch, E. A. Phosphorylation of
Bcl-2 after exposure of human leukemic cells to retinoic acid.
Blood 1998, 92, 1768-1775.
[58] Wang, H. G.; Pathan, N.; Ethell, I. M.; Krajewski, S.; Yamaguchi,
Y.; Shibasaki, F.; McKeon, F.; Bobo, T.; Franke, T. F.; Reed, J. C.
Ca2+-induced apoptosis through calcineurin dephosphorylation of
BAD. Science 1999, 284, 339-343.
[59] Charpin, C.; Garcia, S.; Bonnier, P.; Martini, F.; Andrac, L.; Hor-
schowski, N.; Lavaut, M. N.; Allasia, C. Bcl-2 automated and
quantitative immunocytochemical assays in breast carcinomas :
correlations with 10-year follow-up. J. Clin. Oncol. 1998, 16,
2025-2031.
[60] Del Principe, M. I.; Del Poeta, G.; Maurillo, L.; Buccisano, F.;
Venditti, A.; Tamburini, A.; Bruno, A.; Cox, M. C.; Suppo, G.;
Tendas, A.; Giannì, L.; Postorino, M.; Masi, M.; Del Principe, D.;
Amadori, S. P-glycoprotein and BCL-2 levels predict outcome in
adult acute lymphoblastic leukaemia. Br. J. Haematol. 2003, 121,
730-738.
[61] Kornblau, S. M.; Vu, H. T.; Ruvolo, P.; Estrov, Z.; O'Brien, S.;
Cortes, J.; Kantarjian, H.; Andreeff, M.; May, W. S. Bax and PKCa
modulate the prognostic impact of Bcl-2 expression in acute mye-
logenous leukaemia. Clin. Cancer Res. 2000, 6, 1401-1409.

Deregulation of the Mitochondrial Apoptotic Machinery Current Cancer Drug Targets, 2008, Vol. 8, No. 3 221
[62] Ong, Y.; McMullin, M.; Bailie, K.; Lappin, T. R. J.; Jones, F.;
Irvine, A. High bax expression is a good prognostic indicator in
acute myeloid leukaemia. Br. J. Hematol. 2000, 111, 182-189.
[63] Del Poeta, G.; Venditti, A.; Del Principe, M. I.; Maurillo, L.; Buc-
cisano, F.; Tamburini, A.; Cox, M. C.; Franchi, A.; Bruno, A.;
Mazzone, C.; Panetta, P.; Suppo, G.; Masi, M.; Amadori, S.
Amount of spontaneous apoptosis detected by Bax/Bcl-2 ratio pre-
dicts outcome in acute myeloid leucemia (AML). Blood 2003, 101,
2125-2131.
[64] Andreeff, M.; Jiang, S.; Zhang, X.; Konopleva, M.; Estrov, Z.;
Snell, V. E.; Xie, Z.; Okcu, M. F.; Sanchez-Williams, G.; Dong, J.;
Estey, E. H.; Champlin, R. C.; Kornblau, S. M.; Reed, J. C.; Zhao,
S. Expression of Bcl-2 related genes in normal and AML progeni-
tors: changes induced by chemotherapy and retinoic acid. Leukemia
1999, 13, 1881-1892.
[65] Banker, D. E.; Groudine, M.; Norwood, T.; Appelbaum, F. R.
Measurement of spontaneous and therapeutic agent-induced apop-
tosis with Bcl-2 protein expression in acute myeloid leukaemia.
Blood 1997, 89, 243-255.
[66] Van Stijn, A.; Van der Pol, M. A.; Kok, A.; Bontje, P. M.; Roemen,
G. M.; Beelen, R. H.; Ossenkoppele, G. J.; Schuurhuis, G. J. Dif-
ferences between the CD34+ and CD34- blast compartments in
apoptosis resistance in acute myeloid leucemia. Haematologica
2003, 88, 497-508.
[67] Suárez, L.; Vidriales, B.; García-Laraña, J.; López, A.; Martínez,
R.; Martín-Reina, V.; Tormo, M.; González-San Miguel, J. D.;
Lavilla, E.; García-Boyero, R.; Orfao, A.; San Miguel, J. F.;
PETHEMA Cooperative Group. Multiparametric analysis of apop-
totic and multidrug resistano phenotypes according to the blast cell
maturation stage in elderly patients with acute myeloid leukaemia.
Haematologica 2001, 86, 1287-1295.
[68] Delia, D.; Aiello, A.; Soligo, D.; Fontanella, E.; Melani, C.; Pezzel-
la, F.; Pienotti, M. A.; Della Porta, G. Bcl-2 proto-oncogene e-
xpression in normal and neoplastic human myeloid cells. Blood
1992, 79, 1291-1298.
[69] Pisani, F.; Del Poeta, G.; Aronica, G.; Venditti, A.; Caravita, T.;
Amadori, S. In vitro down-regulation of bcl-2 expression by all-
trans retinoic acid in AML blasts. Ann. Hematol. 1997, 75, 145-
147.
[70] Maurillo, L.; Del Poeta, G.; Venditti, A.; Buccisano, F.; Battaglia,
A.; Santinelli, S.; Caravita, T.; Epiceno, A. M.; Del Moro, B.;
Tamburini, A.; Picardi, A.; Suppo, G.; Catalano, G.; Bruno, A.;
Amadori S. Quantitative analysis of Fas and bcl-2 expression in
hematopoietic precursors. Haematologica 2001, 86, 237-243.
[71] Campos, L.; Sapido, O.; Viallet, A.; Vasselon, C.; Guyotat, D.
Expression of apoptosis-controlling proteins in acute leukemia
cells. Leuk. Lymphoma 1999, 33, 499-509.
[72] Del Principe, M. I.; Del Poeta, G.; Venditti, A.; Buccisano, F.;
Maurillo, L.; Mazzone, C.; Bruno, A.; Neri, B.; Irno Consalvo, M.;
Lo Coco, F.; Amadori, S. Apoptosis and immaturity in acute mye-
loid leucemia. Hematology 2005, 10, 25-34.
[73] Staber, P. B.; Linkesch, W.; Zauner, D.; Beham-Schmid, C.;
Guelly, C.; Schauer, S.; Hoefler, G. Common alterations in gene
expression and increased proliferation in recurrent acute myeloid
leukemia. Oncogene 2004, 23, 894-904.
[74] Yang, D. C.; Jiang, X. P.; Elliott, R. L.; Head, J. F. Inhibition of
growth of human breast carcinoma cells by an antisense oligonu-
cleotide targeted to the transferrin receptor gene. Anticancer Res.
2001, 21, 1777-1787.
[75] Moura, I. C.; Lepelletier, Y.; Arnulf, B.; England, P.; Baude, C.;
Beaumont, C.; Barzabachi, A.; Benhamou, M.; Monteiro, R. C.;
Hermine, O. A neutralizing monoclonal antibody (mAb A24) di-
rected against the transferrin receptor induces apoptosis of tumor T
lymphocytes from ATL patients. Blood 2004, 103, 1838-1845.
[76] Konopleva, M.; Tari, A. M.; Estrov, Z.; Harris, D.; Xie, Z.; Zhao,
S.; López-Berestein, G.; Andreeff, M. Liposomal Bcl-2 antisense
oligonucleotides enhance proliferation, sensitize acute myeloid
leukemia to cytosine-arabinoside, and induce apoptosis independ-
ent of other antiapoptotic proteins. Blood 2000, 95, 3929-3938.
[77] Del Poeta, G.; Stasi, R.; Aronica, G.; Venditti, A.; Cox, M. C.;
Bruno, A.; Buccisano, F.; Masi, M.; Tribalto, M.; Amadori, S.; Pa-
pa, G. Clinical relevance of P-glycoprotein expression in de novo
acute myeloid leukemia. Blood 1996, 87, 1997-2004.
[78] Filipits, M.; Pohl, G.; Stranzl, T.; Suchomel, R. W.; Scheper, R. J.;
Jager, U.; Geissler, K.; Lechner, K.; Pirker, R. Expression of the
lung resistance protein predicts poor outcome in de novo acute
myeloid leukemia. Blood 1998, 91, 1508-1513.
[79] Schaich, M.; Soucek, S.; Thiede, C.; Ehninger, G.; Illmer, T.; SHG
AML96 Study Group. MDR1 and MRP1 gene expression are inde-
pendent predictors for treatment outcome in adult acute myeloid
leukaemia. Br. J. Haematol. 2005, 128, 324-332.
[80] Gerrard, G.; Payne, E.; Baker, R. J.; Jones, D. T.; Potter, M.; Pren-
tice, H. G.; Ethell, M.; McCullough, H.; Burgess, M.; Mehta, A. B.;
Ganeshaguru, K. Clinical effects and P-glycoprotein inhibition in
patients with acute myeloid leukemia treated with zosuquidar tri-
hydrochloride, daunorubicin and cytarabine. Haematologica 2004,
89, 782-790.
[81] Hess, C. J.; Berkhof, J.; Denkers, F.; Ossenkoppele, G. J.;
Schouten, J. P.; Oudejans, J. J.; Waisfisz, Q.; Schuurhuis, G. J. Ac-
tivated intrinsic apoptosis pathway is a key related prognostic pa-
rameter in acute myeloid leukemia. J. Clin. Oncol. 2007, 25, 1209-
1215.
[82] Pukac, L.; Kanakaraj, P.; Humphreys, R.; Alderson, R.; Bloom, M.;
Sung, C.; Riccobene, T.; Johnson, R.; Fiscella, M.; Mahoney, A.;
Carrel, J.; Boyd, E.; Yao, X. T.; Zhang, L.; Zhong, L.; von Ker-
czek, A.; Shepard, L.; Vaughan, T.; Edwards, B.; Dobson, C.; Sal-
cedo, T.; Albert, V. HGS-ETR1, a fully human TRAIL-receptor 1
monoclonal antibody, induces cell death in multiple tumour types
in vitro and in vivo. Br. J. Cancer 2005, 92, 1430-1441.
[83] Goy, A. New directions in the treatment of mantle cell lymphoma:
an overview. Clin. Lymphoma Myeloma 2006, 1, S24-32.
[84] Konopleva, M.; Tsao, T.; Estrov, Z.; Lee, R. M.; Wang, R. Y.;
Jackson, C. E.; McQueen, T.; Monaco, G.; Munsell, M.; Belmont,
J.; Kantarjian, H.; Sporn, M. B.; Andreeff, M. The synthetic triter-
penoid 2-cyano-3,12-dioxooleana-1,9-dien-28-oic acid induces
caspase-dependent and -independent apoptosis in acute myeloge-
nous leukemia. Cancer Res. 2004, 64, 7927-7933.
[85] Marcucci, G.; Stock, W.; Dai, G. Phase I study of oblimersen so-
dium, an antisense to Bcl-2, in untreated older patients with acute
myeloid leukaemia: pharmacokinetics, pharmacodynamics, and
clinical activity. J. Clin. Oncol. 2005, 23, 3404-3411.
[86] Dai, G.; Chan, K. K.; Liu, S. Cellular uptake and intracellular lev-
els of the Bcl-2 antisense G3139 in cultured cells and treated pa-
tients with acute myeloid leukemia. Clin. Cancer Res. 2005, 11,
2998-3008.
[87] Kohl, T. M.; Hellinger, C.; Ahmed, F.; Buske, C.; Hiddemann, W.;
Bohlander, S. K.; Spiekermann, K. BH3 mimetic ABT-737 neutral-
izes resistance to FLT3 inhibitor treatment mediated by FLT3-
independent expression of BCL2 in primary AML blasts. Leukemia
2007, 21, 1763-1772.
[88] Zhang, M.; ling, Y.; Yang, C. Y.; Liu, H.; Wang, R.; Wu, X.; Ding,
K.; Zhu, F.; Griffith, B. N.; Mohammad, R. M.; Wang, S.; Yang,
D. A novel Bcl-2 small molecule inhibitor 4-(3-methoxy-
phenylsulfannyl)-7-nitro-benzofurazan-3-oxide (MNB)-induced ap-
optosis in leukemia cells. Ann. Hematol. 2007, 86, 471-481.
[89] Kojima, K.; Konopleva, M.; Samudio, I. J.; Shicami, M.; Cabreira-
Hansen, M.; McQueen, T.; Ruvolo, V.; Tsao, T.; Zeng, Z.; Vas-
silev, L. T.; Andreeff, M. MDM2 antagonists induce p53-
dependent apoptosis in AML:implications for leukemia therapy.
Blood 2005, 106, 3150-3159.
[90] Kojima, K.; Konopleva, M.; Samudio, I. J.; Schober, W. D.; Born-
mann, W. G.; Andreeff, M. Concomitant inhibition of MDM2 and
Bcl-2 protein function synergistically induce mitochondrial apopto-
sis in AML. Cell Cycle 2006, 5, 2778-2786.
[91] Udupi, V.; Yu, M.; Malaviya, S.; Saavedra, J. E.; Shami, P. J. JS-
K, a nitric oxid prodrug, induces cytochrome c release and caspase
activation in HL-60 myeloid leukaemia cells. Leuk. Res. 2006, 30,
1279-1283.
[92] Jia, L.; Srinivasula, S. M.; Liu, F. T.; Newland, A. C.; Fernandes-
Alnemri, T.; Alnemri, E. S.; Kelsey, S. M. Apaf-1 protein defi-
ciency confers resistance to cytochrome c-dependent apoptosis in
human leukemic cells. Blood 2001, 98, 414-421.
[93] Furukawa, Y.; Sutheesophon, K.; Wada, T.; Nishimura, M.; Saito,
Y.; Ishii, H.; Furukawa, Y. Methylation silencing of the Apaf-1
gene in acute leukemia. Mol. Cancer Res. 2005, 3, 325-334.

222 Current Cancer Drug Targets, 2008, Vol. 8, No. 3 Del Poeta et al.
[94] Schimmer, A. D.; Dalili, S.; Batey, R. A.; Riedl, S. J. Targeting.
XIAP for the treatment of malignancy. Cell Death Differ. 2006, 13,
179-188.
[95] Carter, B. Z.; Gronda, M.; Wang, Z.; Chun, K.; Reed, J. C.;
Schimmer, A. D. Small-molecule XIAP inhibitors derepress down-
stream effector casapases and induce apoptosis of acute myeloid
leukaemia cells. Blood 2005, 105, 4043-4050.
[96] Carter, B. Z.; Mak, D. H.; Schober, W. D.; McQueen, T.; Harris,
D.; Estrov, Z.; Evans, R. L.; Andreef, M. Triptolide induces
caspase-dependent cell death mediated via the mitochondrial path-
way in leukemic cells. Blood 2006, 108, 630-637.
[97] Yu, Z. Y.; Liang, Y. G.; Xiao, H.; Shan, Y. J.; Dong, B.; Huang,
R.; Fu, Y. L.; Zhao, Z. H.; Liu, Z. Y.; Zhao, Q. S.; Wang, S. Q.;
Chen, J. P.; Mao, B. Z.; Cong, Y. W. Melissoidesin G, a diterpe-
noid purified from Isodon melissoides, induces leukemic-cell apop-
tosis through induction of redox imbalance and exhibits synergy
with other anticancer agents. Int. J. Cancer 2007, 121, 2084-2094.
[98] Rosato, R. R.; Almenara, J. A.; Kolla, S. S.; Maggio, S. C.; Coe, S.;
Gimenez, M. S.; Dent, P.; Grant, S. Mechanism and functional role
of XIAP and Mcl-1 down-regulation in flavopiridol/vorinostat an-
tileukemic interactions. Mol. Cancer Ther. 2007, 6, 692-702.
[99] Weisberg, E.; Kung, A. L.; Wright, R. D.; Moreno, D.; Catley, L.;
Ray, A.; Zawel, L.; Tran, M.; Cools, J.; Gilliland, G.; Mitsiades,
C.; McMillin, D. W.; Jiang, J.; Hall-Meyers, E.; Griffin, J. D. Po-
tentiation of antileukemic therapies by Smac mimetic, LBW242:
effects on mutant FLT3-expressing cells. Mol. Cancer Ther. 2007,
6, 1951-1961.
[100] Carter, B. Z.; Wang, R. Y.; Schober, W. D.; Milella, M.; Chism,
D.; Andreeff, M. Targeting Survivin expression induces cell prolif-
eration defect and subsequent cell death involving mitochondrial
pathway in myeloid leukemic cells. Cell Cycle 2003, 2, 488-493.
[101] Pigneux, A.; Mahon, F. X.; Moreau-Gaudry, F.; Uhalde, M. ;
Verneuil, H. D.; Lacombe, F.; Reiffers, J.; Milpied, N.; Praloran,
V.; Belloc, F. Proteasome inhibition specifically sensitises leuke-
mic cells to anthracyclin-induced apoptosis through the accumula-
tion of Bim and Bax pro-apoptotic proteins. Cancer Biol. Ther.
2007, 4, 603-611.
[102] Dasmahapatra, G.; Almenara, J. S.; Grant, S. Flavopiridol and
histone deacetylase inhibitors promote mitochondrial injury and
cell death in huma leukemia cells that overexpress Bcl-2. Mol.
Pharmacol. 2006, 69, 288-298.
[103] Fabre, C.; Carvalho, G.; Tasdemir, E.; Braun, T.; Ades, L.; Gros-
jean, J.; Boehrer, S.; Metivier, D.; Souquere, S.; Pierron, G.; Fen-
aux, P.; Kroemer, G. NF-kappaB inhibition sensitizes to starvation-
induced cell death in high-risk myelodysplastic syndrome and acute
myeloid leukemia. Oncogene 2007, 26, 4071-4083.
[104] Frelin, C.; Imbert, V.; Griessinger, E.; Peyron, A. C.; Rochet, N.;
Philip, P.; Dageville, C.; Sirvent, A.; Hummelsberger, M.; Bérard,
E.; Dreano, M.; Sirvent, N.; Peyron, J. F. Targeting NF-kappaB ac-
tivation via pharmacologic inhibition of IKK2-induced apoptosis of
human acute myeloid leukemia cells. Blood 2005, 105, 804-811.
[105] Carvalho, G.; Fabre, C.; Braun, T.; Grosjean, J.; Ades, L.; Agou,
F.; Tasdemir, E.; Boherer, S.; Israel, A.; Veron, M.; Fenaux, P.;
Kroemer, G. Inhibition of NEMO, the regulatory subunit of the
IKK complex, induces apoptosis in high-risk myelodysplastic syn-
drome and acute myeloid leukemia. Oncogene 2007, 26, 2299-
2307.
[106] Milella, M.; Konopleva, M.; Precupanu, C. M.; Tabe, Y.; Ricciardi,
M. R.; Gregorj, C.; Collins, S. J.; Carter, B. Z.; D’Angelo, C.;
Petrucci, M. T.; Foà, R.; Cognetti, F.; Tafuri, A.; Andreeff, M.
MEK blockade converts AML differentiating response to retinoids
into extensive apoptosis. Blood 2007, 109, 2121-2129.
[107] Amico, D.; Barbui, A. M.; Erba, E.; Rambaldi, A.; Introna, M.;
Golay, J. Differential response of human acute myeloid leukemia
cells to gemtuzumab ozogamicin in vitro: role of Chk1 and Chk2
phosphorylation and caspase 3. Blood 2003, 101, 4589-4597.
[108] Kell, W. J.; Burnett, A. K.; Chopra, R.; Yin, J. A.; Clark, R. E.;
Rohatiner, A.; Culligan, D.; Hunter, A.; Prentice, A. G.; Milligan,
D. W. A feasibility study of simultaneous administration of gemtu-
zumab ozogamicin with intensive chemotherapy in induction and
consolidation in younger patients with acute myeloid leukemia.
Blood 2003, 102, 4277-4283.
[109] Boehrer, S.; Ades, L.; Braun, T.; Galluzzi, L.; Grosjean, J.; Fabre,
C.; Le Roux, G.; Gardin, C.; Martin, A.; de Botton, S.; Fenaux, P.;
Kroemer, G. Erlotinib exhibits anti-neoplastic off-target effects in
AML and MDS : a preclinical study. Blood 2008, 111, 2170-2180.
[110] Huang, M. J.; Cheng, Y. C.; Liu, C. R.; Lin, S.; Liu, H. E. A small-
molecule c-Myc inhibitor, 10058-F4, induces cell-cycle arrest,
apoptosis, and myeloid differentiation of human acute myeloid leu-
kaemia. Exp. Hematol. 2006, 34, 1480-1489.
Received: December 04, 2007 Revised: March 08, 2008 Accepted: March 12, 2008

Copyright © 2022 FDOKUMEN




















