
Lamin A Ser404 Is a Nuclear Target of Akt Phosphorylation in C2C12 Cells
Upload
independentCategory
view
0download
0
C
SBa
b
c
a
ARRA
KASHSCE
1
ss
2
0d
Micron 41 (2010) 966–973
Contents lists available at ScienceDirect
Micron
journa l homepage: www.e lsev ier .com/ locate /micron
2C12 myoblast sensitivity to different apoptotic chemical triggers
ara Saluccia, Michela Battistelli a, Sabrina Burattinia, Cesare Squillacea,arbara Canonicoa,b, Pietro Gobbia, Stefano Papaa,b, Elisabetta Falcieri a,c,∗
DiSUAN, Università degli Studi di Urbino “Carlo Bo”, Urbino, ItalyCentro di Citometria e Citomorfologia, Università degli Studi di Urbino “Carlo Bo”, Urbino, ItalyIGM, CNR, Istituti Ortopedici Rizzoli, Bologna, Italy
r t i c l e i n f o
rticle history:eceived 23 April 2010eceived in revised form 2 July 2010ccepted 2 July 2010
eywords:poptosiskeletal muscle cellydrogen peroxidetaurosporineisplatinumtoposide
a b s t r a c t
Apoptosis is a form of cell death crucial for normal development and tissue homeostasis. Its typicalfeatures include chromatin changes, nuclear breakdown, plasma membrane blebbing and splitting ofcellular content into apoptotic bodies, that progressively undergo phagocytosis.
Apoptosis is considered essential for skeletal muscle development, where defective cells are deletedduring differentiation. In addition, it plays a relevant role in several muscle myopathies, as well as indenervation and disuse.
The aim of this study was to evaluate muscle cell sensitivity to different apoptotic triggers, actingthrough different mechanisms of action. Chemical agents, active against distinct intracellular targets, suchas mitochondrial respiratory chain and DNA, have been chosen to better highlight cell death mechanisms.To induce apoptosis, C2C12 myoblasts have been exposed to H2O2, staurosporine, cisplatin and etoposide,at different doses and incubation times, and they have been analysed by flow cytometry, scanning andtransmission electron microscopy.
Flow cytometry analysis revealed a certain subdiploid peak after all treatments. The best apoptoticeffect was observable, as confirmed at reverted microscope, at minimum doses and after the majorexposure time.
At ultrastructural level programmed cell death has been observed. Characteristic chromatin conden-sation and margination, as well as apoptotic bodies, frequently appeared, even if in the presence ofsecondary necrosis; surface blebs were also observed during scanning microscopic observation.
In particular, exposure to H2O2 or staurosporine showed the largest number of myoblasts in late apop-totic stages and in secondary necrosis. Cisplatin treatments revealed few early apoptotic cells. The analysisof etoposide-induced apoptosis was in agreement with data obtained from flow cytometry, indicating asignificant increase of apoptotic cell number.
These results suggest that all conditions are able to induce apoptosis in C2C12 myoblasts, which occurs,considering trigger mechanisms of action, mostly following the mitochondrial pathway, if not excludingthat due to DNA damage. Therefore, mitochondria permeability alteration is an important step in skeletalmuscle programmed cell death. This last conclusion seems to have a significant relevance in understand-ing the mechanisms involved in muscle disorders, denervation and chronic muscle disuse, conditionsfrequently characterized by a decline in mitochondrial content and by an increase of mitochondrial
apoptosis susceptibility.. Introduction
Apoptosis is a cell suicide program by which damaged orenescent cells are individually eliminated to maintain homeosta-is in multicellular organisms (Shiokawa et al., 2002). It plays a
∗ Corresponding author at: DiSUAN, Campus Scientifico Sogesta,Via Ca’ le Suore, 61029 Urbino (PU), Italy. Tel.: +39 0722304284; fax: +39 0722304244.
E-mail address: [email protected] (E. Falcieri).
968-4328/$ – see front matter © 2010 Elsevier Ltd. All rights reserved.oi:10.1016/j.micron.2010.07.002
© 2010 Elsevier Ltd. All rights reserved.
key role in development, reproduction, aging, immune functionand cell diseases. It is characterized by specific events, such asDNA fragmentation, chromatin margination and condensation, cellshrinkage, membrane blebbing and apoptotic bodies formation(Burattini et al., 2009).
Skeletal muscle tissue has long been considered highly resistant
to apoptotic cell death. Nevertheless, apoptosis frequently occurs inskeletal muscle biology, where it has been proposed to have a rolein development, both in proliferating myoblast and in postmitoticmuscle fiber control. Therefore, its deregulation is involved in anumber of muscle disorders.
cron 4
eUt(mfa
l
csmaHs
eieeMcd
aatoir
s
Fm
S. Salucci et al. / Mi
Signs of apoptosis have been detected in several muscular dis-ases, such as Duchenne Muscular Dystrophy (DMD), Becker andlrich myopathies (Rizzuto et al., 2004), Juvenile Dermatomyosi-
is (Zhao et al., 2007), Congenital Muscular Dystrophy type 1AGirgenrath et al., 2009) and others. Histological studies in many
uscle pathologies reported, in fact, that loss of skeletal muscleunction might be due to an excessive apoptotic cell death (Millernd Girgenrath, 2006).
Moreover, it has been described in loaded muscle after pro-onged exercise (Sandri et al., 1997).
In addition, chronic heart failure, motor neuron disorders, spinalord injury and skeletal muscle atrophy due to hind limb suspen-ion or immobilization, have been all associated to an increase ofuscle apoptosis as revealed by TUNEL or by DNA fragmentation
nalysis after gel electrophoresis (Smith et al., 2000; Adhihetty andood, 2003; Dupont-Versteegden, 2006; Davatz et al., 2007), both
uggesting apoptotic DNA cleavage.Apoptosis is also an important component in the pathogen-
sis of post-denervation muscle atrophy. Morphological studiesn rat denervated muscle (soleus and extensor digitorum longus)vidence, besides atrophy, significant myofibril loss and pres-nce of myofibril-free zones, mostly located in perinuclear areas.oreover, numerous nuclei containing large patches of condensed
hromatin, sharply separated from diffuse small areas, a widelyescribed apoptotic feature, appear (Borisov and Carlson, 2000).
It is known that external stimuli such as cytokines, radiationsnd several chemotherapic agents and drugs are able to inducepoptosis. In a recent work (D’Emilio et al., 2010) we demonstratedhat UV-B treatment induces it in both myoblasts and myotubes
f C2C12 cells. In particular, apoptotic morphology was revealedn the presence of DNA fragmentation evidenced in situ by TUNELeaction.The purpose of this work is to investigate C2C12 myoblast sen-itivity to a variety of different chemical apoptotic triggers, in order
ig. 1. Control cells at RM (A), SEM (B) and TEM (C). Myoblasts reveal a well-preserveditochondria with a good morphology appeared. A and B bars 10 �m; C bar 1 �m; D bar
1 (2010) 966–973 967
to highlight mechanisms underlying muscle cell death. Hydrogenperoxide (H2O2), an oxidative stress mediator (Siu et al., 2009) andstaurosporine, a PKC inhibitor and a ROS generator too (Falcieri etal., 1993; Basset et al., 2006) were utilized.
In addition, two chemotherapic agents were chosen. Cisplatin,forming a spectrum of intra- and interstrand DNA cross-links,leading to DNA damage, followed by transcription and replica-tion inhibition, was used (Todd and Lippard, 2009). The effect ofetoposide, a topoisomerase II inhibitor, inducing DNA double strandbreaks, leading to p53 activation and apoptosis (Karpinich et al.,2002), was finally investigated.
2. Materials and methods
2.1. Cell culture and apoptosis induction
Mouse C2C12 myoblasts, a murine model derived from satel-lite cells, and generally considered a muscle progenitor lineage(Burattini et al., 2004), were grown in flasks or on coverslips indishes, in the presence of Dulbecco’s modified Eagle’s medium sup-plemented with 10% heat-inactivated fetal bovine serum, 2 mMglutamine at 37 ◦C and 5% CO2.
To induce apoptosis, myoblasts were exposed to:
- 500 �M and 2 mM H2O2,- 0.5 �M and 1 �M staurosporine,- 30 �M and 60 �M cisplatin,- 25 �M and 50 �M etoposide.
For each trigger two incubation times, 12 h and 24 h, have beenchosen.
Control and treated cell behaviours were monitored by meansof a TE 2000-S Nikon reverted microscope (RM), equipped with adigital Nikon DN100 system (D’Emilio et al., 2010).
cell morphology with star-like or fusiform shape and smooth cell surface. In (D)0.5 �m.
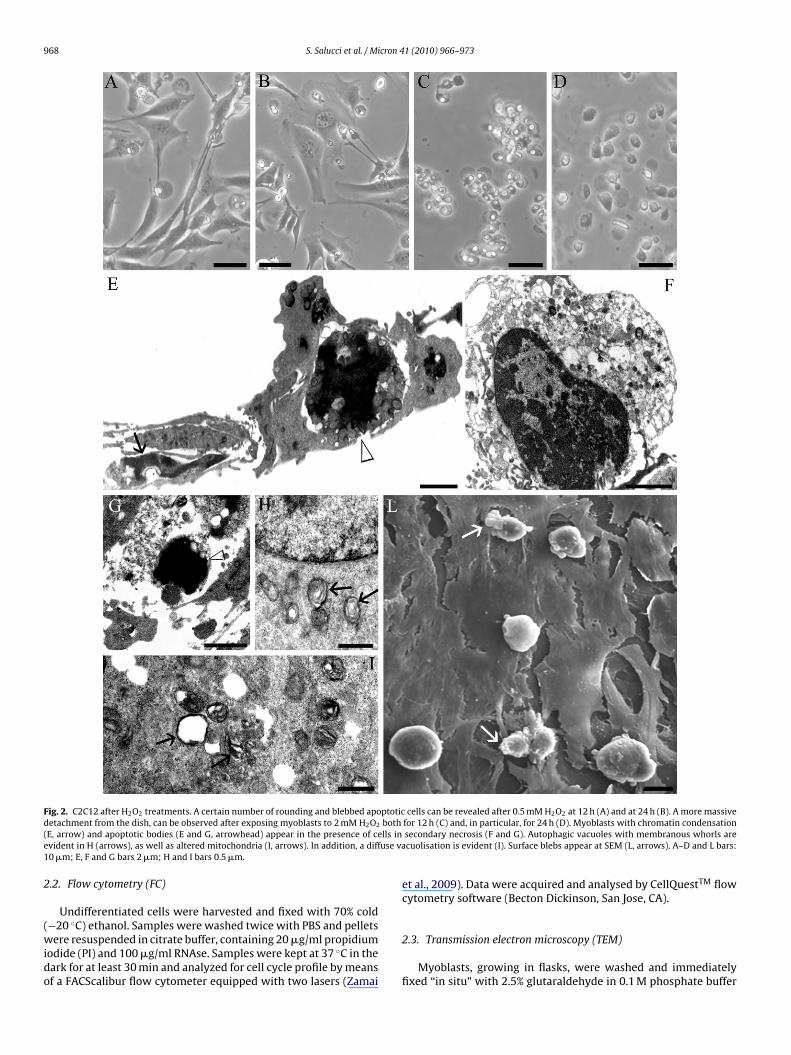
968 S. Salucci et al. / Micron 41 (2010) 966–973
Fig. 2. C2C12 after H2O2 treatments. A certain number of rounding and blebbed apoptotic cells can be revealed after 0.5 mM H2O2 at 12 h (A) and at 24 h (B). A more massived both( lls ine use va1
2
(wido
etachment from the dish, can be observed after exposing myoblasts to 2 mM H2O2
E, arrow) and apoptotic bodies (E and G, arrowhead) appear in the presence of cevident in H (arrows), as well as altered mitochondria (I, arrows). In addition, a diff0 �m; E, F and G bars 2 �m; H and I bars 0.5 �m.
.2. Flow cytometry (FC)
Undifferentiated cells were harvested and fixed with 70% cold
−20 ◦C) ethanol. Samples were washed twice with PBS and pelletsere resuspended in citrate buffer, containing 20 �g/ml propidiumodide (PI) and 100 �g/ml RNAse. Samples were kept at 37 ◦C in theark for at least 30 min and analyzed for cell cycle profile by meansf a FACScalibur flow cytometer equipped with two lasers (Zamai
for 12 h (C) and, in particular, for 24 h (D). Myoblasts with chromatin condensationsecondary necrosis (F and G). Autophagic vacuoles with membranous whorls arecuolisation is evident (I). Surface blebs appear at SEM (L, arrows). A–D and L bars:
et al., 2009). Data were acquired and analysed by CellQuestTM flowcytometry software (Becton Dickinson, San Jose, CA).
2.3. Transmission electron microscopy (TEM)
Myoblasts, growing in flasks, were washed and immediatelyfixed “in situ” with 2.5% glutaraldehyde in 0.1 M phosphate buffer

cron 4
fwi
swa
2
cp01w5
3
tMd
C
bq
atc
eDia13s
qtd3aad
q6
TC
S. Salucci et al. / Mi
or 15 min, gently scraped and centrifuged at 1200 rpm. Pelletsere additionally fixed for 1 h, alcohol dehydrated, and embedded
n araldite (Salucci et al., 2010).Thin sectioning was preceded by the analysis of toluidine blue-
tained semithin sections at light microscopy (LM). Thin sectionsere stained with uranyl acetate and lead citrate and analysed withPhilips CM10 electron microscope.
.4. Scanning electron microscopy (SEM)
Both control and treated myoblasts were directly cultured onover slides, inside Petri dishes. After careful washing with phos-hate buffer, monolayers were fixed with 2.5% glutaraldehyde in.1 M phosphate buffer for 1 h, quickly washed and post-fixed with% OsO4 in the same buffer for 1 h. After alcohol dehydration, theyere critical point dried, gold sputtered and observed with a Philips
15 scanning electron microscope.
. Results
FC (Table 1) evidenced that all triggers were able to induce a cer-ain DNA cleavage, quantified by the presence of hypodiploid peak.
oreover, all of them, except H2O2, used at high doses presented aecrease of cell amount, due to the increase of necrotic cell death.
FC analysis showed only 2% of necrotic-apoptotic cells in control2C12, revealing a good cell viability.
The treatment with 500 �M H2O2, after 12 h and 24 h of incu-ation, presented, at FC, a similar behaviour and a low subG1 peak,uantified as 16% and 17%, respectively, was observed.
After 2 mM H2O2 exposure, a 16% hypodiploid peak appearedt 12 h of incubation, and it increased to 62% after 24 h incubationime. The last peak identifies a mixture of necrotic and apoptoticells.
Staurosporine treatments evidenced an hypodiploid peak pres-nce for all doses used and for each incubation time. In particular,NA cleavage appeared increased using a lower dose, correspond-
ng to a 0.5 �M concentration. Apoptotic presence was quantifieds 25% after 12 h and 38.4% after 24 h of treatment. Incubation with�M staurosporine presented an hypodiploid peak quantified as3% after 12 h and 27% after 24 h, being this decrease, due to thetrong effect that induce a massive necrotic cell death.
Cisplatin treatment revealed a certain DNA cleavage presence,uantified, after an incubation of 24 h at 30 �M, as 28%. On the con-rary, after 12 h, a strong resistance to cell death appeared, its effectecreased and only a 10% hypodiploid peak, after incubation with0 �M cisplatin, was revealed. The incubation with 60 �M cisplatin,fter 12 h and 24 h, showed a DNA cleavage presence, quantifieds 15% for both situations. In the last case the apoptotic response
ecreased in the presence of necrotic cell death.After 25 �M etoposide exposure, a certain hypodiploid peak,uantified as 15%, appeared after 12 h and its effect increased to0% after 24 h of incubation time.
able 1ell death percentage at different concentrations and times.
12 h 24 h
CTRL 2% 2%H2O2 0.5 mM 16% 17%H2O2 2 mM 16% 62%Staurosporina 0.5 �M 25% 38.4%Staurosporina 1 �M 33% 27%Cisplatin 30 �M 10% 28%Cisplatin 60 �M 15% 15%Etoposide 25 �M 15% 60%Etoposide 50 �M 14% 31%
1 (2010) 966–973 969
Treatment with 50 �M etoposide showed a minor effect, if com-parable to the dose described above, and apoptotic cell death wasquantifiead as 14% after 12 h and 31% after 24 h. In this last situationa necrotic and apoptotic cell deaths coexist.
All experimental conditions, analysed at FC, have been also mon-itored at RM.
Control cells presented a good viability, evidenced by RM, SEMand TEM (Fig. 1). Myoblasts appeared mononucleated and flat, withan elongated or star-shaped shape (A). SEM and TEM observationsrevealed an uniform smooth cell surface, with rare microvilli (B andC), and a good preservation of mitochondria (D).
Morphological analyses indicated that all triggers induced a cer-tain apoptotic effect.
After H2O2 treatments, cell death and some alterations in cellmorphology appeared (Fig. 2). Myoblasts treated with 0.5 mM H2O2for 12 h (A) and 24 h (B) or with 2 mM H2O2 for 12 h (C) and 24 h (D),monitored at RM, appeared progressively modified and rounding.They lost the anchorage to the underlying substrate, evidencing adiffuse cell detachment, surface blebs and, at high doses (2 mM),an increase of necrosis. TEM analyses revealed late apoptotic mor-phological events. Chromatin margination and condensation, aswell as apoptotic bodies with clumped chromatin, were observed(E) even if in the presence of a certain necrotic cell death (F andG).
Cytoplasmic vesicles, containing rearranged membranouswhorls, similar to myelin figures, were also observed, in proximityof nuclei with normal chromatin (H). These structures, suggestive ofautophagic vacuoles (Martinet et al., 2005), will be further inves-tigated. In addition, mitochondria occasionally lost their internalorganisation and some of them appeared completely swollen (I).Sometimes a diffuse vacuolisation can be revealed (I), as frequentlydescribed in cell oxidative stress. SEM observations showed a num-ber of rounding, non-adherent myoblasts with frequent surfaceblebs (L).
Staurosporine treatment had a strong effect on myoblasts(Fig. 3). In fact, a massive detachment with diffuse cell roundingand blebbing was evident exposing cells to 0.5 �M for 12 h (A)and 24 h (B) or 1 �M for 12 h (C) and 24 h (D). In particular, sur-face blebs and other apoptotic signs were better observable usingthis trigger at minimum dosage. Apoptotic bodies with clumpedchromatin and cells in secondary necrosis appeared at TEM (E andF).
After exposure (Fig. 4) to 30 �M cisplatin for 12 h (A) and 24 h(B) or 60 �M cisplatin for 12 h (C) and 24 h (D), a few numberof detached myoblasts could be identified, some of which show-ing progressive cytoplasmic vacuolisation, better observable at LM(E) and TEM (F and G). In addition, mitochondria appeared wellpreserved after this type of treatment (G). TEM analyses revealedcells with an initial chromatin margination at nuclear periphery(F), apoptotic bodies with a strongly condensed chromatin (H) andnecrotic cell death (I and L).
After etoposide exposure (Fig. 5), myoblasts treated with dif-ferent doses and at different incubation times – 25 �M for 12 h(A) and 24 h (B) or 50 �M for 12 h (C) or 24 h (D) – displayedbleb formation and appeared mostly fusiform, even if in the pres-ence of ultrastructural changes. In particular, a major apoptoticresponse was observed exposing myoblasts to 25 �M etoposideat 24 h of incubation and at this condition apoptotic cell numberfurther increased. Surface blebs were better observable at LM andTEM (E and F) and a diffuse cytoplamic vacuolisation was alsorevealed (Fig. 5E, F, G and I). Moreover, mitochondria appeared
occasionally altered and their internal organisation appeared com-pletely lost (G). Chromatin condensation (H and I), organized inlarge dense masses, different from the classic cup-shaped apoptoticones, could be observed and cells in secondary necrosis appeared(L).
970 S. Salucci et al. / Micron 41 (2010) 966–973
F h furt( h (C) aa pen aa
4
mm(tt2
vm
rcha
csof
m
ig. 3. Staurosporine treatments. A diffuse cell detachment from the substrate, witA) and 24 h (B). Cell death increases after exposure to 1 �M staurosporine for 12nalysis reveals myoblasts in late apoptotic stages: cells in secondary necrosis (E, ond D bars 12 �m; E and F bars 2 �m.
. Discussion
Apoptosis plays an important role in skeletal muscle develop-ent, by controlling the size of the population of proliferatingyoblasts which undergo differentiation into mature myotubes
Vasconsuelo et al., 2008). It is known, both in vivo and in vitro,hat a large fraction of cells is deleted by apoptosis during skele-al muscle development (Sandri and Carraro, 1999; Huppertz et al.,001).
This study suggests that apoptosis underlies muscle cell death initro and demonstrates a certain dependence on apoptotic inducerechanism of action.Apoptotic response was repeatedly monitored by FC, that
eveals a moderate cleaved DNA content after H2O2, staurosporine,isplatin and etoposide treatments. In particular, a significantypodiploid peak can be observed after cell exposure to etoposidet minimum dose and after 24 h of incubation time.
Ultrastructural observations confirm the apoptotic response:hromatin margination and condensation, cytoplasmic vacuoli-
ation, mitochondrial alterations and surface blebs were in factbserved. Also in terms of morphology, etoposide is responsibleor a significant apoptotic cell number.Literature data demonstrated that H2O2 induces apoptosis viaitochondrial pathway and in our model these organelles appear
her rounding and blebbing, appears after exposure to 0.5 �M staurosporine at 12 hnd 24 h (D), and cell amount decreases in the presence of a diffuse necrosis. TEM
rrow) and apoptotic bodies (E and F, arrow) can be observed. A and B bars 6 �m; C
strongly modified by H2O2 treatment. Mitochondria, at ultrastruc-tural analysis, appear in fact altered and in some cases completelyswollen. It is known that mitochondrial changes could be relatedto the release of their proteins during apoptosis (Granville et al.,2001; Kroemer and Zitvogel, 2007), suggesting that mitochondriaplays a pivotal role in ROS-induced apoptosis. ROS generation, afterH2O2 treatment, lead to cytochrome c and Smac/DIABLO release, byactivating or enhancing the levels of proapoptotic proteins, such asp53 or Bax (Green and Reed, 1998; Siu et al., 2009). Successively,the activation of caspase-9 and caspase-3, and DNA fragmentation(Jiang et al., 2005) take place. This is in agreement with our data: infact, myoblasts exposed to ROS increase show, at FC, a DNA cleav-age presence and, at ultrastructural analysis, all apoptotic markers,described above. In the present study, the appearence of apoptosiswas demonstrated in C2C12 myoblasts after exposure to relativelyhigh doses of H2O2.
Some studies showed that ROS increase is involved in musculardamage and dysfunction after ischemia followed by reperfusion(Adhihetty and Hood, 2003). ROS induced-apoptosis was also
observed in aging muscle, where proapototic events, such asincreases in Bax/Bcl-2, cytochrome c release, AIF, p53, caspasesand/or DNA fragmentation, were detected (Pistilli et al., 2006;Always and Siu, 2008). For this reason the accelerated rate of apop-tosis in aging muscle may be attributed to oxidative stress.
S. Salucci et al. / Micron 41 (2010) 966–973 971
Fig. 4. Cisplatin treatments. Few rounding and blebbed myoblasts can be identified after 30 �M cisplatin treatment for 12 h (A) and 24 h (B). Myoblasts exposed to 60 �Mc r 24 hT ith ana and I b
toroceaasi
i
isplatin for 12 h (C) and 24 h (D) presented a necrotic aspect, better observable afteEM (F). Generally mitochondria present a good morphology (F). Moreover, cells wnd L) can be also observed. A–D bars 10 �m; E bar 4 �m; F and L bars 0.5 �m G, H
Moreover, autophagy signs were detected in C2C12 exposedo H2O2. TEM analyses revealed indeed the occasional appearancef vacuoles organized in membranous whorls, resulting from dis-uption of cellular components and organelle deletion. Autophagyccurs in many eukaryotic cell types, where organelles and otherell components are degraded inside phago-lysosomes, like a gen-ral defense mechanism against anomalous cellular componentsccumulated following toxic insults or during cell death (Lockshin
nd Zakeri, 2004). It could represent a type of cell death – or atage of it – in dying skeletal muscle fiber too. Further studies aren progress to better address this hypothesis.Staurosporine, a PKC inhibitor, is a therapeutic agent able tonhibit tumor cell growth by inducing cell death via intrinsic apop-
of incubation. A general cytoplasmic vacuolisation is evident at LM (E, arrows) andinitial chromatin margination (G), apoptotic bodies (H and I) and necrotic cells (Iars 1 �m.
totic pathway (Antonsson and Persson, 2009) and cytochromec release. It stimulates ROS production in myoblasts and, toa minor extent, in myotubes (Camara et al., 2007). In thepresent work, staurosporine treatments were strong for cellsthat became rounding and diffusely blebbed. In addition, sev-eral apoptotic bodies and myoblasts in secondary necrosis can beobserved at TEM. This work demonstrates muscle cell sensitiv-ity to staurosporine-induced apoptosis. This type of cell death is
involved also in muscle degeneration occuring in DMD patients:Basset et al. (2006) showed in fact that dystrophic myotubeswere more sensitive to staurosporine-induced stress, indicatingthat dystrophin-deficient cells are more susceptible to free radi-cals.
972 S. Salucci et al. / Micron 41 (2010) 966–973
Fig. 5. Etoposide treatments. RM evidences cell rounding and bleb formation (arrows) after exposure to 25 �M for 12 h (A) and 24 h (B). Cells with blebs appear also after5 d TEMc rion ic ecrosa
sDRsoht
0 �M at 12 h (C) and 24 h (D), even if in the presence of necrotic process. At LM anytoplasmic vacuoles are evident (G and H). In G, a completely swollen mitochondhromatin condensation (H and I) can be observed. In addition, cells in secondary nnd L bars 1 �m.
Cisplatin reacts with DNA to form intrastrand and/or inter-trand cross-links of platinum adducts. This behaviour blocksNA replication and transcription, because of RNA synthesis or
NA polymerases inhibition (Todd and Lippard, 2009). Tran-cription block leads to apoptosis through either p53-dependentr -independent pathways. The role of p53 in this process isighly controversial: the p53-dependent pathway may involveranslocation of the protein to mitochondria, where it can bind anti-, fusiform cells appear, showing a characteristic blebbing (E–F, arrows) and diffuses evidenced (arrow). A diffuse presence of apoptotic cells characterized by strongis can be detected (L). A–D bars 10 �m; E bar 5 �m; F bar 2 �m; G bar 0.5 �m; H, I
apoptotic Bcl-2 family proteins and induce cell death (Marchenkoet al., 2000). After cisplatin treatments, in our experiments chro-matin margination and condensation, as well as DNA cleavage,
were detected, even if in the presence of necrotic cells. However,this trigger induces apoptosis in C2C12 myoblasts without mito-chondrial damage.Etoposide, a topoisomerasi II inhibitor, is a widely used anti-neoplastic drug, involved in DNA damage. It stabilizes the complex

cron 4
fi(baspmIlbspmu
o
hdcmatttd
pc
A
Dt
R
A
A
A
B
B
B
B
C
D
S. Salucci et al. / Mi
ormed by topoisomerase II and the 5′-cleaved DNA end, thus form-ng stable and irreversible protein-linked DNA double strand breaksMeresse et al., 2004). In the present study this drug is responsi-le for the major apoptotic effect on myoblasts (if used at 25 �Mnd after 24 h of incubation time), as revealed by FC that showed aignificant hypodyploid peak. Moreover, at microscopy level, cellsresented apoptotic features, such as a strongly condensed chro-atin, a diffuse vacuolisation and, sometimes, secondary necrosis.
n addition, mitochondria appear occasionally altered and seem toose their internal organisation. This last result could be explainedy the fact that the drug causes an accumulation of doubletrand breaks, generating a sequence that proceeds, trough p53hosphorylation, to Bax up-regulation. This last translocates toitochondria, with the consequent release of cytochrome c, and,
ltimately, apoptotic cell death (Karpinich et al., 2002).Our data show C2C12 myoblast sensitivity to apoptosis, based
n FC evaluation of DNA cleavage and on morphological features.In our previous work, by D’Emilio et al. (2010), UV-B radiation,
as been used to induce apoptosis in C2C12 cells. In this case celleath was due to mitochondrial damage followed by cytochromerelease and caspase-9 activation. The present work shows C2C12yoblast sensitivity to different chemical agents, confirming thatvariety of external stimuli can initiate the apoptotic process. All
he utilized triggers have in common, at least indirectly, the execu-ive apoptotic step which follows the intrinsic pathway, suggestinghat mitochondria exert a fundamental role in muscle apoptotic celleath.
Morphological, biochemical and functional studies are inrogress to further investigate mitochondrial involvement in mus-le apoptosis.
cknowledgements
The research was supported by Urbino University “Carlo Bo”.r. Davide Curzi and Mr. Aurelio Valmori are thanked for excellent
echnical assistance.
eferences
dhihetty, P.J., Hood, D.A., 2003. Mechanisms of Apoptosis in Skeletal Muscle. BasicAppl. Myol. 13, 171–179.
lways, S.E., Siu, P.M., 2008. Nuclear apoptosis contributes to sarcopenia. Exerc.Sport Sci. Rev. 36, 51–57.
ntonsson, A., Persson, J.L., 2009. Induction of apoptosis by staurosporine involvesthe inhibition of expression of the major cell cycle proteins at the G(2)/m check-point accompanied by alterations in Erk and Akt kinase activities. AnticancerRes. 29, 2893–2898.
asset, O., Boittin, F.X., Cognard, C., Constantin, B., Ruegg, U.T., 2006. Bcl-2 over-expression prevents calcium overload and subsequent apoptosis in dystrophicmyotubes. Biochem. J. 395, 267–276.
orisov, A.B., Carlson, B.M., 2000. Cell death in denervated skeletal muscle is distinctfrom classical apoptosis. Anat. Rec. 258, 305–318.
urattini, S., Ferre, P., Battistelli, M., Curci, R., Luchetti, F., Falcieri, E., 2004.C2C12 murine myoblasts as a model of skeletal muscle development: morpho-functional characterization. Eur. J. Histochem. 48, 223–234.
urattini, S., Ferri, P., Battistelli, M., D’Emilio, A., Biagiotti, L., Sestili, P., Rocchi, M.B.,Falcieri, E., 2009. Apoptotic DNA fragmentation can be revealed in situ: an ultra-
structural approach. Microsc. Res. Tech. 72, 913–923.amara, Y., Duval, C., Sibille, B., Villarroya, F., 2007. Activation of mitochondrial-driven apoptosis in skeletal muscle cells is not mediated by reactive oxygenspecies production. Int. J. Biochem. Cell. Biol. 39, 146–160.
avatz, C.G., Andreo, J.C., Rodrigues, C.A., Rosa Junior, G.M., Moraes, R.L.H., 2007.Apoptosis in denervated skeletal muscle. Int. J. Morphol. 25, 529–536.
1 (2010) 966–973 973
D’Emilio, A., Biagiotti, L., Burattini, S., Battistelli, M., Canonico, B., Evangelisti, C.,Ferri, P., Papa, S., Martelli, A.M., Falcieri, E., 2010. Morphological and biochemicalpatterns in skeletal muscle apoptosis. Histol. Histopathol. 25, 21–32.
Dupont-Versteegden, E.E., 2006. Apoptosis in skeletal muscle and its relevance toatrophy. World J. Gastroenterol. 12, 7463–7466.
Falcieri, E., Martelli, A.M., Bareggi, R., Cataldi, A., Cocco, L., 1993. The protein kinaseinhibitor staurosporine induces morphological changes typical of apoptosis inMolt-4 cells without concomitant DNA fragmentation. Biochem. Biophys. Res.Commun. 193, 19–25.
Girgenrath, M., Beermann, M.L., Vishnudas, V.K., Homma, S., Miller, J.B., 2009.Pathology is alleviated by doxycycline in a laminin-á2-null model of congenitalmuscular dystrophy. Ann. Neurol. 65, 47–56.
Granville, D.J., Cassidy, B.A., Ruehlmann, D.O., Choy, J.C., Brenner, C., Kroemer, G.,van Breemen, C., Margaron, P., Hunt, D.W., McManus, B.M., 2001. Mitochondrialrelease of apoptosis-inducing factor and cytochrome c during smooth musclecell apoptosis. Am. J. Pathol. 159, 305–311.
Green, D.R., Reed, J.C., 1998. Mitochondria and apoptosis. Science 281, 1309–1312,Review.
Huppertz, B., Tews, D.S., Kaufmann, P., 2001. Apoptosis and syncytial fusion in humanplacental trophoblast and skeletal muscle. Int. Rev. Cytol. 205, 215–253.
Jiang, B., Xiao, W., Shi, Y., Liu, M., Xiao, X., 2005. Heat shock pretreatment inhibitedthe release of Smac/DIABLO from mitochondria and apoptosis induced by hydro-gen peroxide in cardiomyocytes and C2C12 myogenic cells. Cell Stress Chap. 10,252–262.
Karpinich, N.O., Tafani, M., Rothman, R.J., Russo, M.A., Farber, J.L., 2002. The courseof etoposide-induced apoptosis from damage to DNA and p53 activation tomitochondrial release of cytochrome c. J. Biol. Chem. 277, 16547–16552.
Kroemer, G., Zitvogel, L., 2007. Death, danger, and immunity: an infernal trio.Immunol. Rev. 220, 5–7.
Lockshin, R.A., Zakeri, Z., 2004. Apoptosis, autophagy, and more. Int. J. Biochem. Cell.Biol. 36, 2405–2419.
Marchenko, N.D., Zaika, A., Moll, U.M., 2000. Death signal-induced localization of p53protein to mitochondria: a potential role in apoptotic signaling. J. Biol. Chem.275, 16202–16212.
Martinet, W., De Meyer, G.R.Y., Herman, A.G., Kockx, M.M., 2005. Amino acid depri-vation induces both apoptosis and autophagy in murine C2C12 muscle cells.Biotechnol. Lett. 27, 1157–1163.
Meresse, P., Dechaux, E., Monneret, C., Bertounesque, E., 2004. Etoposide: Discoveryand medicinal chemistry. Curr. Med. Chem. 11, 2443–2466.
Miller, J.B., Girgenrath, M., 2006. Role of apoptosis in neuromuscular diseasesand potential usefulness of anti-apoptosis therapy. Trends Mol. Med. 12, 279–286.
Pistilli, E.E., Jackson, J.R., Always, S.E., 2006. Death receptor-associated pro-apoptoticsignaling in aged skeletal muscle. Apoptosis 11, 2115–2126.
Rizzuto, R., Duchen, M.R., Pozzan, T., 2004. Flirting in little space: theER/mitochondria Ca2+ liaison. Sci. STKE 13, re1.
Salucci, S., Burattini, S., Battistelli, M., Chiarini, F., Martelli, A.M., Sestili, P., Gobbi, P.,Falcieri, E., 2010. Melatonin prevents hydrogen peroxide-induced apoptotic celldeath. Microscopie 1, 47–53.
Sandri, M., Podhorska-Okolow, M., Geromel, V., Rizzi, C., Arslan, P., Franceschi,C., Carraio, U., 1997. Exercise induces myonuclear ubiquitination and apop-tosis in dystrophin-deficient muscle of mice. J. Neuropathol. Exp. Neur. 56,45–57.
Sandri, M., Carraro, U., 1999. Apoptosis of skeletal muscles during development anddisease. Int. J. Biochem. Cell. Biol. 31, 1373–1390.
Shiokawa, D., Kobayashi, T., Tanuma, S., 2002. Involvement of DNase gamma in Apop-tosis Associated with Myogenic Differentiation of C2C12 Cells. J. Biochem. Chem.277, 31031–31037.
Siu, P.M., Wang, Y., Always, S.E., 2009. Apoptotic signaling induced by H2O2-mediated oxidative stress in differentiated C2C12 myotubes. Life Sci. 84,468–481.
Smith, H.K., Maxwell, L., Martyn, J.A., Bass, J.J., 2000. Nuclear DNA fragmentationand morphological alterations in adult rabbit skeletal muscle after short-termimmobilization. Cell Tissue Res. 302, 235–241.
Todd, R.C., Lippard, S.J., 2009. Inhibition of transcription by platinum antitumorcompounds. Metallomics 1, 280–291.
Vasconsuelo, A., Milanesi, L., Boland, R., 2008. 17B-Estradiol abrogates apoptosisin murine skeletal muscle cells through estrogen receptors: role of the phos-phatidylinositol 3-kinase/Akt pathway. J. Endocrinol. 196, 385–397.
Zamai, L., Galeotti, L., Zotto, G.D., Canonico, B., Mirandola, P., Papa, S., 2009. Identifica-tion of a NCR + /NKG2D + /LFA-1(low)/CD94(-) immature human NK cell subset.Cytometry A 75, 893–901.
Zhao, Y., Fedczyna, T.O., McVicker, V., Caliendo, J., Li, H., Pachman, L.M., 2007. Apopto-sis in the skeletal muscle of untreated children with juvenile dermatomyositis:Impact of duration of untreated disease. Clin. Immunol. 125, 165–172.
Copyright © 2022 FDOKUMEN




















