
Cytokines affecting CD4+ T regulatory cells in transplant tolerance. Interleukin-4 does not maintain...
9
Cytokines affecting CD4 + T regulatory cells in transplant tolerance. Interleukin-4 does not maintain alloantigen specific CD4 + CD25 + Treg Karren M. Plain a,1 , Nirupama D. Verma a,2 , Giang T. Tran a , Masaru Nomura a,3 , Rochelle Boyd a,4 , Catherine M. Robinson a , Suzanne J. Hodgkinson a,b , Bruce M. Hall a, ⁎ a Immune Tolerance Laboratory Faculty of Medicine, University of New South Wales, Sydney, Australia b Department of Medicine, Liverpool Hospital, Liverpool, NSW, Australia abstract article info Article history: Received 19 July 2013 Received in revised form 3 October 2013 Accepted 3 October 2013 Keywords: CD4 + CD25 + FoxP3 + Treg Transplantation tolerance Regulatory T cell Interleukin 4 Th2 response IL-4 is thought to promote induction of transplantation tolerance and alloantigen-specific CD4 + CD25 + T regulatory cells (Treg). This study examined the effect of IL-4 on the induction and maintenance of the CD4 + T regulatory cells (Treg) that mediate transplantation tolerance. Tolerance was induced in DA rats with PVG heterotopic cardiac allografts by a short course of cyclosporine. Naïve and tolerant lymphocytes, including the CD4 + and CD4 + CD25 + T cell subsets, were assayed in mixed lymphocyte cultures with or without recombinant (r)IL-4 or other cytokines. The proliferation, cell surface and cytokine phenotype of these cells was examined, as was their capacity to adoptively transfer tolerance. rIL-4 enhanced the proliferation of naïve and tolerant lymphoid cells, including CD4 + and CD4 + CD25 + T cells, but this was not alloantigen specific. Naïve or tolerant CD4 + T cells cultured with rIL-4 and donor PVG antigen effected rapid graft rejection, even though before culture tolerant CD4 + T cells transferred antigen-specific tolerance. These rIL-4 cultured CD4 + T cells had a phenotype consistent with activated CD4 + CD25 + FoxP3 - Th2 cells. While naïve natural CD4 + CD25 + T cells (nTreg) cultured with alloantigen and rIL-4 had enhanced proliferation and capacity to suppress rejection in vivo, the culture of tolerant CD4 + CD25 + T cells with alloantigen and rIL-4 could not sustain their proliferation against specific donor, nor their capacity to transfer tolerance to specific donor allograft. Thus, IL-4 promotes both regulatory and effector T cells early in the immune response, but once alloimmune tolerance is established, IL- 4 promoted the activation of effector cells to mediate rejection and did not support alloantigen-specific Treg that could transfer specific tolerance. Crown Copyright © 2013 Published by Elsevier B.V. All rights reserved. 1. Introduction The mechanisms involved in the induction and maintenance of transplantation tolerance include a key role for regulatory or suppressor T cells (Treg) [1,2] especially within CD4 + T cells [3–8] and these include CD4 + CD25 + Treg [3,5,7,9,10]. Peripheral CD4 + T cells from animals with transplant tolerance can inhibit specific donor but not third party allograft rejection mediated by either CD4 + or CD8 + T effector cells [1,4,5]. Such alloantigen specific tolerance is demonstrated by the co- transfer of tolerant CD4 + T cells with naïve T cells that should mediate rejection in an immunocompromised host [1,4,5]. Precisely how tolerant antigen specific CD4 + T cells suppress naïve T cells is not precisely known, but there are many potential mechanisms, including inhibition of inflammation in the graft itself (reviewed in [11,12]). Transplant tolerance is alloantigen specific and mediated by alloantigen-specific CD4 + CD25 + FoxP3 + Treg [4,5,13] that have distinct difference to natural CD4 + CD25 + FoxP3 + Treg (nTreg) [5,13], though both are known to express FoxP3 [13]. nTreg are a separate lineage of T cells produced in the thymus that represent b 10% of peripheral CD4 + T cells, suppress in a non-antigen specific manner [14,15] and play a key role in induction of transplant tolerance [10,16–18]. nTreg suppress graft rejection only at non-physiological ratios of >1:1 [19] or when effector T cell responses are inhibited (reviewed in [20]). Induced CD4 + Treg (iTreg), derived from CD4 + CD25 - effector T lineage cells activated with antigen in the presence of TGF-β and not IL-1 and IL-6, also express CD25 and FoxP3 [21]. Transplant Immunology 29 (2013) 51–59 Abbreviations: CPM, Counts per minute; ConAsup, Supernatant from concanavalin A activated spleen cells; IFN-γ, Interferon-gamma; IFNGR, IFN-γ receptor; IL, Interleukin; IL-5Rα, IL-5 receptor alpha, specific for IL-5 binding; iTreg, T regulatory cells induced from CD4 + CD25 - T cells; MLC, Mixed lymphocyte culture; nTreg, Natural T regulatory cells (CD4 + CD25 + FoxP3 + T cell); rIL, Recombinant rat interleukin; Ts2, nTreg activated by IL-4 and alloantigen; Ts1, nTreg activated by IL-2 and alloantigen. ⁎ Corresponding author at: Suite 206, National Innovation Centre, 4 Cornwallis St., Australian Technology Park, Eveleigh 1430, Australia. Tel.: +61 2 92094955; fax: +61 2 92094957. E-mail address: [email protected] (B.M. Hall). 1 Current address: Faculty of Veterinary Science, University of Sydney, 425 Werombi Rd, Camden, NSW 2570, Australia. 2 Current address: ANZAC Institute, Concord Hospital and University of Sydney, Concord, NSW, Australia. 3 Current address: Keiwakai Ebetsu Hospital, Nopporo Yoyogi-cho 81-6, Ebetsu City, Hokkaido 069-0817, Japan. 4 Current address: The Australian School of Advanced Medicine, 2 Technology Place, Macquarie University, NSW 2109, Australia. 0966-3274/$ – see front matter. Crown Copyright © 2013 Published by Elsevier B.V. All rights reserved. http://dx.doi.org/10.1016/j.trim.2013.10.003 Contents lists available at ScienceDirect Transplant Immunology journal homepage: www.elsevier.com/locate/trim
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Cytokines affecting CD4+ T regulatory cells in transplant tolerance. Interleukin-4 does not maintain...

Transplant Immunology 29 (2013) 51–59
Contents lists available at ScienceDirect
Transplant Immunology
j ourna l homepage: www.e lsev ie r .com/ locate / t r im
Cytokines affecting CD4+ T regulatory cells in transplant tolerance. Interleukin-4 doesnot maintain alloantigen specific CD4+CD25+ Treg
Karren M. Plain a,1, Nirupama D. Verma a,2, Giang T. Tran a, Masaru Nomura a,3, Rochelle Boyd a,4,Catherine M. Robinson a, Suzanne J. Hodgkinson a,b, Bruce M. Hall a,⁎a Immune Tolerance Laboratory Faculty of Medicine, University of New South Wales, Sydney, Australiab Department of Medicine, Liverpool Hospital, Liverpool, NSW, Australia
Abbreviations: CPM, Counts per minute; ConAsup, Suactivated spleen cells; IFN-γ, Interferon-gamma; IFNGR,IL-5Rα, IL-5 receptor alpha, specific for IL-5 binding; iTfrom CD4+CD25− T cells; MLC, Mixed lymphocyte cultucells (CD4+CD25+FoxP3+ T cell); rIL, Recombinant rat inby IL-4 and alloantigen; Ts1, nTreg activated by IL-2 and a⁎ Corresponding author at: Suite 206, National Innov
Australian Technology Park, Eveleigh 1430, Australia. Tel.92094957.
E-mail address: [email protected] (B.M. Hall).1 Current address: Faculty of Veterinary Science, Univers
Camden, NSW 2570, Australia.2 Current address: ANZAC Institute, Concord Hospit
Concord, NSW, Australia.3 Current address: Keiwakai Ebetsu Hospital, Nopporo
Hokkaido 069-0817, Japan.4 Current address: The Australian School of Advanced
Macquarie University, NSW 2109, Australia.
0966-3274/$ – see front matter. Crown Copyright © 2013http://dx.doi.org/10.1016/j.trim.2013.10.003
a b s t r a c t
a r t i c l e i n f oArticle history:Received 19 July 2013Received in revised form 3 October 2013Accepted 3 October 2013
Keywords:CD4+CD25+FoxP3+ TregTransplantation toleranceRegulatory T cellInterleukin 4Th2 response
IL-4 is thought to promote induction of transplantation tolerance and alloantigen-specific CD4+CD25+ Tregulatory cells (Treg). This study examined the effect of IL-4 on the induction and maintenance of the CD4+ Tregulatory cells (Treg) that mediate transplantation tolerance. Tolerance was induced in DA rats with PVGheterotopic cardiac allografts by a short course of cyclosporine. Naïve and tolerant lymphocytes, including theCD4+ and CD4+CD25+ T cell subsets, were assayed inmixed lymphocyte cultures with or without recombinant(r)IL-4 or other cytokines. The proliferation, cell surface and cytokine phenotype of these cells was examined, aswas their capacity to adoptively transfer tolerance. rIL-4 enhanced the proliferation of naïve and tolerantlymphoid cells, including CD4+ and CD4+CD25+ T cells, but this was not alloantigen specific. Naïve or tolerantCD4+ T cells culturedwith rIL-4 and donor PVG antigen effected rapid graft rejection, even though before culturetolerant CD4+ T cells transferred antigen-specific tolerance. These rIL-4 cultured CD4+ T cells had a phenotypeconsistent with activated CD4+CD25+FoxP3− Th2 cells. While naïve natural CD4+CD25+ T cells (nTreg)cultured with alloantigen and rIL-4 had enhanced proliferation and capacity to suppress rejection in vivo, theculture of tolerant CD4+CD25+ T cells with alloantigen and rIL-4 could not sustain their proliferation againstspecific donor, nor their capacity to transfer tolerance to specific donor allograft. Thus, IL-4 promotes bothregulatory and effector T cells early in the immune response, but once alloimmune tolerance is established, IL-4 promoted the activation of effector cells to mediate rejection and did not support alloantigen-specific Tregthat could transfer specific tolerance.
Crown Copyright © 2013 Published by Elsevier B.V. All rights reserved.
1. Introduction
The mechanisms involved in the induction and maintenance oftransplantation tolerance include a key role for regulatory or suppressorT cells (Treg) [1,2] especially within CD4+ T cells [3–8] and these
pernatant from concanavalin AIFN-γ receptor; IL, Interleukin;reg, T regulatory cells inducedre; nTreg, Natural T regulatoryterleukin; Ts2, nTreg activatedlloantigen.ation Centre, 4 Cornwallis St.,: +61 2 92094955; fax: +61 2
ity of Sydney, 425Werombi Rd,
al and University of Sydney,
Yoyogi-cho 81-6, Ebetsu City,
Medicine, 2 Technology Place,
Published by Elsevier B.V. All rights
include CD4+CD25+ Treg [3,5,7,9,10]. Peripheral CD4+ T cells fromanimals with transplant tolerance can inhibit specific donor but notthird party allograft rejectionmediated by either CD4+ or CD8+T effectorcells [1,4,5]. Such alloantigen specific tolerance is demonstrated by the co-transfer of tolerant CD4+ T cells with naïve T cells that should mediaterejection in an immunocompromised host [1,4,5]. Precisely how tolerantantigen specific CD4+T cells suppress naïve T cells is not precisely known,but there are many potential mechanisms, including inhibition ofinflammation in the graft itself (reviewed in [11,12]).
Transplant tolerance is alloantigen specific and mediated byalloantigen-specific CD4+CD25+FoxP3+ Treg [4,5,13] that have distinctdifference to natural CD4+CD25+FoxP3+ Treg (nTreg) [5,13], thoughboth are known to express FoxP3 [13]. nTreg are a separate lineage of Tcells produced in the thymus that represent b10% of peripheral CD4+ Tcells, suppress in a non-antigen specific manner [14,15] and play a keyrole in induction of transplant tolerance [10,16–18]. nTreg suppressgraft rejection only at non-physiological ratios of >1:1 [19] or wheneffector T cell responses are inhibited (reviewed in [20]). Induced CD4+
Treg (iTreg), derived from CD4+CD25− effector T lineage cells activatedwith antigen in the presence of TGF-β and not IL-1 and IL-6, also expressCD25 and FoxP3 [21].
reserved.

52 K.M. Plain et al. / Transplant Immunology 29 (2013) 51–59
Alloantigen-specific Treg are lesswell studied than nTreg, to the pointwhere it has been suggested that iTreg and not nTreg produce antigen-specific Treg [22]. In transplant tolerance the CD4+CD25+FoxP3+ Tregthat mediate alloantigen specific tolerance potentially may be derivedfrom nTreg [23] or iTreg. In the peripheral lymphoid tissues of toleranthosts, CD4+CD25+FoxP3+ T cells are only present at a physiologicalratio of 1:10 [13] suggesting that the Treg cells in these animals are morepotent at suppressing responses to specific donor than nTreg [4,5,13,24].
Our previous studies showed that CD4+ T cells from animals tolerantto an allograft lost their capacity to transfer transplant tolerance afterculture for three days (d) with a donor alloantigen [6,7]. Cytokine richsupernatant from conconavalin A activated spleen cells (ConAsup)preserved CD4+ T cell's capacity to transfer tolerance [7], but IL-2 alonewas not sufficient. This effect was not due to ConA induced suppressorcells [25], as ConA in the supernatant was blocked by α- D mannosyl[7], thus cytokines other than IL-2 are required.We have set out to definewhich cytokines can promote survival of antigen specific Treg, and haveidentified that several T cell cytokines may promote antigen specificTreg (reviewed in [11,23]).
IL-4 can expand naïve nTreg [23,26,27]. We found that alloactivationof naïve nTreg with rIL-4, like rIL-2, leads to the induction of alloantigenspecific Treg that can suppress rejection and induce tolerance at a ratioof 1:10 to naïve CD4+T cells [23]. nTreg activated by rIL-4 and alloantigenexpressedmRNA for the specific IL-5 receptor alpha (IL-5Rα)whereas rIL-2 and alloantigen activated expression of mRNA for interferon gamma(IFN-γ) receptor (IFNGR). This showed that IL-4 activated a Th2 cytokinedependent antigen-specific Treg that we termed Ts2, whereas IL-2activatedaTh1cytokinedependent lineageof Treg thatwe termedTs1 [23].
This study examined whether rIL-4 promoted the survival of theantigen-specific CD4+ and CD4+CD25+ T cells that maintain transplanttolerance in animals with long standing tolerance induced by a shortperiod of immunosuppression by cyclosporine [2,4,5,7,28]. In thismodel, PVG cardiac allografts function without ongoing immuno-suppression and accept second grafts from specific donor but rejectthird party grafts, demonstrating that they have alloantigen specifictolerance to the original graft. Paradoxically CD4+ T cells from thesetolerant hosts proliferate to specific donor in the same manner as theyproliferate to third party alloantigen, and with a similar response to thatof cells from a normal naïve animal [28]. Thus while the host toleratesthe allogeneic grafts, they have specific alloreactive CD4+ T cells thatproliferate in vitro. These same CD4+ T cells from animals with tolerancewereunable to effect rejection of specific donor PVGgrafts in an irradiatedadoptive host, but could effect rejection of third party grafts [2,4,5].Further they suppress the capacity of co-transferred naïve CD4+ T cellsto mediate specific donor graft rejection [2,4,5].
In this study we examined the effect of culture of naïve and tolerantcells with rIL-4 and specific antigen. We found that rIL-4 promotedalloactivation of naïve and tolerant CD4+T cells into effectors of rejection.Although we previously reported that rIL-4 can promote induction ofalloantigen-specific Treg from nTreg, rIL-4 did not maintain the functionof tolerant alloantigen-specific CD4+CD25+FoxP3+ Treg in cultures withspecific donor stimulator cells.We concluded thatwhile IL-4 canpromotethe induction of antigen specific Treg from nTreg, it cannot support thegrowth and function of alloantigen specific Treg that have developed intolerant hosts.
2. Materials and methods
2.1. Animals
DA (RT1a), PVG (RT1c) and Lewis (RT1l) rats were bred andmaintained in the animalhouse, Liverpool Hospital [29]. Experiments were approved by the Animal EthicsCommittee, University of New South Wales.
2.2. Operative procedures
DAratswere graftedwith adult heart grafts heterotopically, as described [30]. Tolerancewas induced by a 10 d course of cyclosporine (10 mg/kg body weight) (Novartis, North
Ryde, Australia) [4,5] or anti-CD4 mAb, as described [29]. Adoptive transfer experimentsused irradiated hosts with heart grafts [4,5] and rejection scored as described [23]. Transferof tolerance occurs without rejection in about 60% of animals, the others have a transientrejection of b8 d before graft function recovers and they become tolerant so that the graftsfunction well for over 100 d. Controls, including irradiation only and reconstitution with5× 106 naïve CD4+ T cells, were rolling and include some previously published results.
2.3. Monoclonal antibodies (mAb) and cytokines
Anti-rat mAb used were MRCOx35 (CD4), MRCOx8 (CD8), MRCOx39 (CD25, IL-2Rα),MRCOx6 (class II MHC) and MRCOx33 (CD45RA) (BD-PharMingen, San Diego, CA),MRCOx81 (blocks rat IL-4) (Dr. N. Barclay, School of Pathology, Oxford, UK), and FoxP3mAb (FJK-16s) (eBioscience, San Diego, CA).
rIL-4 was from an IL-4 transfected CHOK1 cell line [31] (Dr. N. Barclay) grown inserum free medium. rIL-4 was assayed for induction of class II MHC expression on Bcells [24,31]. We previously described the preparation of control supernatant from non-transfected CHOK1 cells (CHO-S), rIL-2, rIFN-γ, rIL-5 [23,32] and ConAsup [7]. ConAsupwas blocked with 0.05 mM/L of 1-o-methyl-alpha-mannopyranoside (Sigma), as ConAcan induce Treg [33]. The final concentrations in culture for rIL-2 and rIL-4 were 100 U/ml and 125 U/ml, respectively.
2.4. Lymphocyte subset isolation and cultures
LN and spleen cells were prepared as previously described and enriched for CD4+ Tcells or CD4+CD25+ and CD4+CD25− T cells as described [19,23]. Primary mixedlymphocyte cultures (MLC) and cultures of CD4+CD25+ T cells were as described [19,23].Secondary MLC were performed on cells stimulated in a primary MLC, washed twice andrested for 24 h prior to re-culture. Bulk MLC were seeded at 2.5 × 106 stimulator cells and2.5 × 105 responder cells/ml in 25 cm3 tissue culture flasks (Greiner, Frickenhausen,Germany). rIL-2 (100 U/ml), rIL-4 (125 U/ml) or other cytokines were added to cultures,as described [23]. Cultured cells were assessed; for proliferation at 3–6 d, at 48 h for RT-PCR for cytokines and cytokine receptors and at 72 h for FACS or adoptive transfer toirradiated heart grafted animals [19,23].
2.5. RT-PCR analysis
RNA extraction, cDNA synthesis, and semi-quantitative polymerase chain reaction(PCR) were performed as described [29]. For some experiments, a 1:10 dilution series ofcDNA was performed. Quantitative RT-PCR methods and primers were as described [23].
2.6. Statistical analyses
Parametric data were expressed as mean± standard deviation and compared usingStudent's t-test (Statview, Abacus). Graft survival was plotted using the Kaplan–Meiermethod, and analysed by Mann–Whitney log-rank test (Statview). Statistical significancewas set at p b 0.05.
3. Results
3.1. Comparison of naïve or tolerant lymphocytes cultured with IL-4 inMLC
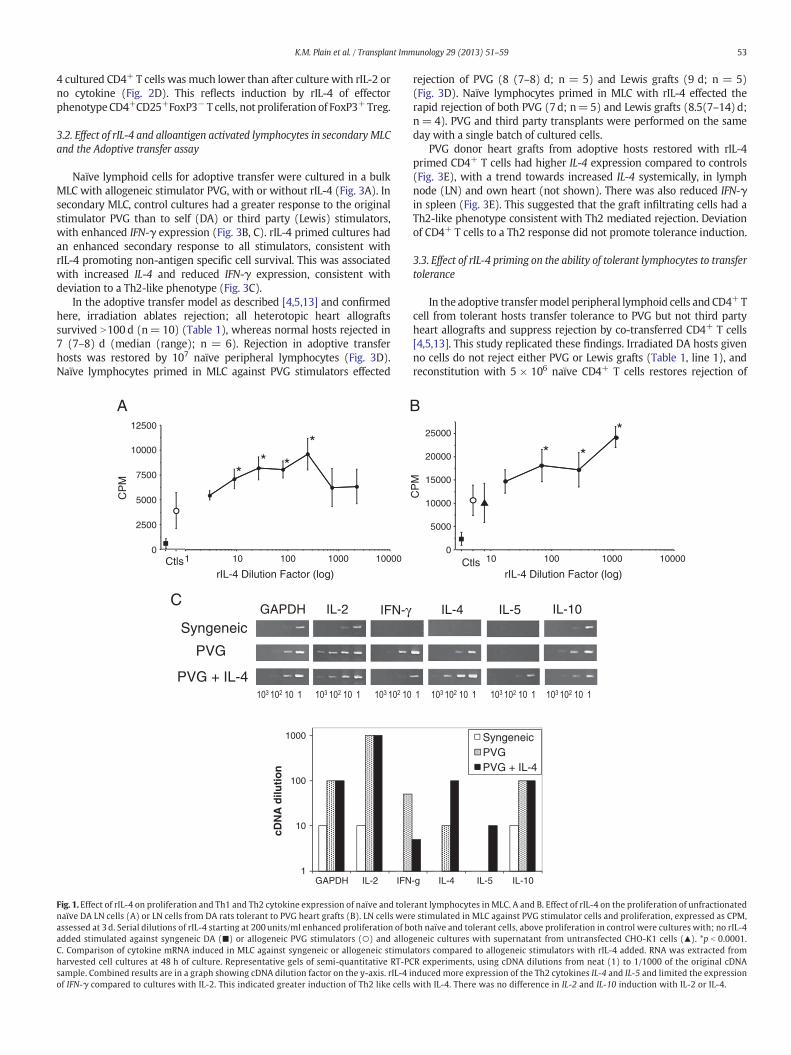
rIL-4 in vitro enhanced the proliferation of naïve lymphocytes in adose dependent fashion (Fig. 1A) associated with the induction of Th2cytokine mRNAs, IL-4 and IL-5 but reduced Th1 cytokine mRNA forIFN-γ, compared to cells cultured without cytokine (Fig. 1C). Similarly,tolerant lymphocytes from DA rats with long surviving allografts(N100 d), had enhanced proliferation with rIL-4 (Fig. 1B). The controlsupernatant from non-transfected CHO-K1 cells had no effect onproliferation or Th2 cytokine induction (Fig. 1B and data not shown).
Without cytokines, CD4+ T cells from naïve and tolerant ratsproliferated to a similar degree in MLC (Fig. 2A) similar to our previousreport [28]. Both rIL-2 and rIL-4 significantly enhanced the proliferationof naïve and tolerant CD4+ T cells to both self and alloantigen comparedto controls with no cytokine, whereas rIL-5 and rIFN-γ did not (Fig. 2A).
Culture of CD4+ T cell from both naïve and tolerant rats with rIL-4and alloantigen led to substantial induction of CD25 expression withup to 80% of cells being CD4+CD25+, compared to 10 and 14% in freshlyisolated naïve and tolerant populations respectively (Fig. 2B and C). rIL-4 did not markedly increase the proportion of FoxP3+ cells, indicatingthat there was activation of CD4+CD25− T cells to express CD25, notexpansion of CD4+CD25+FoxP3+ nTreg (Fig. 2B). Both naïve andtolerant CD4+ T cells had 2–3 fold higher expression of CD25 thanreplicate cultures supplemented with rIL-2 (Fig. 2C). IFN-γ and IL-5did not induce CD25 (data not shown). The FoxP3 to CD25 ratio in rIL-
53K.M. Plain et al. / Transplant Immunology 29 (2013) 51–59
4 cultured CD4+ T cells was much lower than after culture with rIL-2 orno cytokine (Fig. 2D). This reflects induction by rIL-4 of effectorphenotypeCD4+CD25+FoxP3− T cells, not proliferation of FoxP3+Treg.
3.2. Effect of rIL-4 and alloantigen activated lymphocytes in secondary MLCand the Adoptive transfer assay
Naïve lymphoid cells for adoptive transfer were cultured in a bulkMLC with allogeneic stimulator PVG, with or without rIL-4 (Fig. 3A). Insecondary MLC, control cultures had a greater response to the originalstimulator PVG than to self (DA) or third party (Lewis) stimulators,with enhanced IFN-γ expression (Fig. 3B, C). rIL-4 primed cultures hadan enhanced secondary response to all stimulators, consistent withrIL-4 promoting non-antigen specific cell survival. This was associatedwith increased IL-4 and reduced IFN-γ expression, consistent withdeviation to a Th2-like phenotype (Fig. 3C).
In the adoptive transfer model as described [4,5,13] and confirmedhere, irradiation ablates rejection; all heterotopic heart allograftssurvived N100 d (n=10) (Table 1), whereas normal hosts rejected in7 (7–8) d (median (range); n = 6). Rejection in adoptive transferhosts was restored by 107 naïve peripheral lymphocytes (Fig. 3D).Naïve lymphocytes primed in MLC against PVG stimulators effected
BA
0
2500
5000
7500
10000
12500
100001000100101
rIL-4 Dilution Factor (log)
** *
*
Ctls
CP
M
C
1
10
100
1000
GAPDH IL-2 IFN
cDN
A d
iluti
on
IFN-γGAPDH IL-2
Syngeneic
PVG
PVG + IL-4
Fig. 1.Effect of rIL-4 on proliferation and Th1 and Th2 cytokine expression of naïve and tolenaïve DA LN cells (A) or LN cells from DA rats tolerant to PVG heart grafts (B). LN cells werassessed at 3 d. Serial dilutions of rIL-4 starting at 200units/ml enhanced proliferation of boadded stimulated against syngeneic DA (■) or allogeneic PVG stimulators (○) and allogC. Comparison of cytokine mRNA induced in MLC against syngeneic or allogeneic stimulharvested cell cultures at 48 h of culture. Representative gels of semi-quantitative RT-PCsample. Combined results are in a graph showing cDNA dilution factor on the y-axis. rIL-4 iof IFN-γ compared to cultures with IL-2. This indicated greater induction of Th2 like cells
rejection of PVG (8 (7–8) d; n = 5) and Lewis grafts (9 d; n = 5)(Fig. 3D). Naïve lymphocytes primed in MLC with rIL-4 effected therapid rejection of both PVG (7d; n=5) and Lewis grafts (8.5(7–14) d;n= 4). PVG and third party transplants were performed on the sameday with a single batch of cultured cells.
PVG donor heart grafts from adoptive hosts restored with rIL-4primed CD4+ T cells had higher IL-4 expression compared to controls(Fig. 3E), with a trend towards increased IL-4 systemically, in lymphnode (LN) and own heart (not shown). There was also reduced IFN-γin spleen (Fig. 3E). This suggested that the graft infiltrating cells had aTh2-like phenotype consistent with Th2 mediated rejection. Deviationof CD4+ T cells to a Th2 response did not promote tolerance induction.
3.3. Effect of rIL-4 priming on the ability of tolerant lymphocytes to transfertolerance
In the adoptive transfermodel peripheral lymphoid cells and CD4+ Tcell from tolerant hosts transfer tolerance to PVG but not third partyheart allografts and suppress rejection by co-transferred CD4+ T cells[4,5,13]. This study replicated these findings. Irradiated DA hosts givenno cells do not reject either PVG or Lewis grafts (Table 1, line 1), andreconstitution with 5 × 106 naïve CD4+ T cells restores rejection of
0
5000
10000
15000
20000
25000
Ctls 10000100010010
rIL-4 Dilution Factor (log)
*
**
CP
M
-g IL-4 IL-5 IL-10
SyngeneicPVGPVG + IL-4
IL-4 IL-5 IL-10
rant lymphocytes in MLC. A and B. Effect of rIL-4 on the proliferation of unfractionatede stimulated in MLC against PVG stimulator cells and proliferation, expressed as CPM,th naïve and tolerant cells, above proliferation in control were cultures with; no rIL-4eneic cultures with supernatant from untransfected CHO-K1 cells (▲). *p b 0.0001.ators compared to allogeneic stimulators with rIL-4 added. RNA was extracted fromR experiments, using cDNA dilutions from neat (1) to 1/1000 of the original cDNAnduced more expression of the Th2 cytokines IL-4 and IL-5 and limited the expressionwith IL-4. There was no difference in IL-2 and IL-10 induction with IL-2 or IL-4.

12.2 13.5 14.4 64.7
Fresh CD4+ Control cultureCD4+
Culture + rIL-2CD4+
Culture + rIL-4CD4+
12.0 15.0 19.3 14.4
CD
4-C
y
0
20
40
60
80
CD
4+ CD
25+ (
%)
0
0.2
0.4
0.6
0.8
1
Fo
xp3+
% /
CD
25+ %
CD
4-C
y
CD25-PE
Foxp3-FITC
B
C
D
0
20
40
60
80
Fresh Control +IL-2 +IL-4
Fresh Control +IL-2 +IL-4
Fresh Control +IL-2 +IL-4
Fresh Control +IL-2 +IL-4
CD
4+ CD
25+
(%)
A
0
500
1000
1500
2000
2500
3000
3500
4000
CTL +IL-4 +IL-2 +IL-5 +IFN-g +CHO-S
/DA
/PVG*
*
*0
500
1000
1500
2000
2500
3000
CTL +IL-4 +IL-2 +IL-5 +IFN-g +CHO-S
/DA
/PVG* *
Naïve CD4+ Tolerant CD4+
Naïve Tolerant
Naïve
0
0.4
0.8
1.2
1.6
2
Fo
xp3+
% /
CD
25+ %
Tolerant
CP
M
Fig. 2. rIL-4 induces increased expression of CD25 but not FoxP3 on CD4+ T cells from naïve and tolerant animals in MLC. A. Proliferation in MLC of CD4+ T cells from a naïve DA or DAtolerant to PVG heart allograft. MLC against syngeneic DA or allogeneic PVG stimulator cells with no cytokines added (CTL) or in the presence of rIL-4, rIL-2, rIL-5, rIFN-γ or supernatantfromuntransfectedCHO-K1 cells (CHO-S). Proliferation, expressed as CPM,was assessed at 3d. *pb0.05 compared control cultureswithout cytokine against the same stimulator cells. rIL-2increased proliferation of both naïve and tolerant cells to DA (p b 0.001) and PVG (p b 0.009) as did IL-4 (p b 0.013 and 0.001). rIL-5, rIFN-γ and CHOK supernatant did not enhanceproliferation compared to control, IFN-γ significantly inhibit naïve cell proliferation (p b .001). A similar enhancement of naïve and tolerant CD4+ T cell proliferation has been observedin all other experiments. B–D. FACS results of naïve and tolerant CD4+ T cells, from one experiment comparing freshly isolated (Fresh) with cells post-MLC against specific allogeneic PVGstimulator cells, without cytokines added (Control culture) orwith rIL-2 or rIL-4 added. The resultswere reproducible in other experiments. B. Tolerant cell FACS; CD4-Cy5 shownony-axisand CD25-PE (top panels) or FoxP3-FITC (bottom panels) on the x-axis. C. Graph representing the results from panel B, showing the proportion of CD4+CD25+ T cells post-culture of bothnaïve and tolerant cells weremarkedly increasedwith rIL-4 but notwith rIL-2, though the proportion of FoxP3+ cells was similar. D. Graph representing the results from panel B, showingthat the proportion of CD4+FoxP3+ compared to CD4+CD25+ cells was reduced in cultures with rIL-4, leading to a low FoxP3+/CD25+ ratio.
54 K.M. Plain et al. / Transplant Immunology 29 (2013) 51–59

B
PVG or Lewis rat
Adoptive host
Donorheart
Naïve DA
7 Gy
Control or IL-4 cultured Graft survival
(severe rejection)
Spl&LN cells
A
ControlIL-4 Treated
Donor heart2425
2627
2829
3031
3233
IL-2 IFN-γ IL-4 IL-10
Spleen25
26
27
28
29
30
31
32
33IL-2 IFN-γ IL-4 IL-10P
CR
cyc
le o
f app
eara
nce
D
a,b
Control rIL-4
CP
M (
x10-3
)
0
20
40
60
80
c
a
DAPVGLewis
IL-4
Ban
d D
ensi
ty (
ng)
0
20
40
60
IFN-γ
Control rIL-4 Control rIL-4
DAPVGLewis
CE
Control MLCMLC +IL-4
Irradiation ctl Naive cells
504030201000
20
40
60
80
100
PVG graft
Days post-transplantation
Sur
viva
l to
seve
re r
ejec
tion
(%)
504030201000
20
40
60
80
100
Lewis grafta,b
d
dd
e
Fig. 3. Effect of IL-4 on the ability of naïve DA spleen and LN cells, cultured against PVG stimulator cells, to effect rejection or induce tolerance. A. Diagram ofmodel showing cells that weretaken fromnaïve DA rats and stimulated by PVG cells in cultureswith andwithout rIL-4, then tested in the adoptive transfer assaywith specific donor PVG or third party Lewis heart grafts.B Proliferation ofDA cells in a secondaryMLC, followingprimary stimulation inMLC against PVG stimulatorswith no cytokine added (Control) orwith rIL-4. Proliferation (CPM) to DA, PVGor third party Lewis stimulators assessed at 2 d. a. p b 0.0001 compared to control cultures against all stimulator types; b. p b 0.0001 compared to rIL-4 MLC versus DA stimulators. c.p b 0.0001 compared to control MLC versus DA or Lewis stimulators C. RT-PCR shows marked induction of IL-4 after culture with rIL-4 (d; p b 0.05), but suppression of IFN-γ comparedto controls (nearly significant). GAPDH expression was comparable in all samples (data not shown). D. Graft survival for PVG specific and Lewis third party cardiac allografts in adoptiverecipients givenno cells (Irradiation CTL), 1×107 naïveDA cells (naïve cells), 1×107DA/PVGMLC cellswith no rIL-4 (ControlMLC) or 1×107 rIL-4 treatedDA/PVGMLC cells (MLC& rIL-4).n = 4–11 rats per group. rIL-4 cultured cells effected rejection and did not induce tolerance E. Comparison of cytokine expression mRNA in tissues from adoptive transfer recipientsrestored with cells from control MLC or MLC with rIL-4 as described in A. Collated semi-quantitative RT-PCR data (mean± range) from n=5–8 rats per group. GAPDH expression wassimilar (data not shown) thus results were directly compared. There was more IL-4 in rejecting hearts mediated by naïve cells that had been cultured in MLC with rIL-4 than in controlsgiven cells cultured in MLC with no rIL-4 (e; p=0.05), and a trend for less IFN-γ. Thus the rejection had Th2-like IL-4 expressing cells in the grafts.
55K.M. Plain et al. / Transplant Immunology 29 (2013) 51–59
both PVG or Lewis grafts (Table 1, line 3). Freshly isolated tolerant CD4+
T cells transferred tolerance to PVG but rejected Lewis allografts(p b 0.05) (Table 1, line 2) and if mixed with 5 × 106 naïve CD4+ Tcells suppress rejection of PVG but not Lewis grafts (p b 0.05) (Table 1,line 4). We have previously shown that tolerant CD4+ T cells need tobe at a ratio of 1:4 to naïve CD4+ T cells to suppress rejection[2,4,5,13]. Thus this ratio was used in this study.
Tolerant CD4+ T cells cultured in MLC with specific donor PVGstimulators and no cytokine supplement for 3 d lost the capacity totransfer tolerance and effected rejection with a median rejection timeof 15d (Table 1, line 5) significantly different (pb0.001) to fresh tolerantcells (Table 1, line 2) and as described [7]. The tolerance transferringcapacity of these CD4+ T cells to PVG allograft was retained in MLCwith PVG stimulators and with ConAsup as a source of cytokines.These ConAsup supported tolerant cells suppressed PVG graft rejectionby co-transferred naïve CD4+ T cells (N90 (11- >100) d; n = 11)(Table 1, line 6) which was significantly different to tolerant CD4+ Tcells cultured with no cytokines which rejected PVG grafts (p b 0.05)(Table 1, line 5). Tolerant cells cultured with PVG and ConAsup rejectedthird party Lewis grafts with a median rejection time of 18 d (Table 1,line 6) and not significantly different to the rejection time with naïveCD4+ T cells alone (Table 1, line 3). Culture of naïve cells with ConAsup
and PVG alloantigen did not induce tolerance transferring cells, as thesecells effected both PVG and Lewis graft rejection (Table 1, line 10). Thisindicated that ConAsup did not induce non-antigen specific Treg [33]and confirmed our previous study of the effects of ConAsup on tolerantCD4+ T cells in culture [7].
Tolerant cells cultured with rIL-4 and PVG alloantigen effectedrejection of both PVG (8 (7–9) d; n= 7) and Lewis third party grafts(7 (7–11) d; n = 5) (NSD) (Table 1, line 7) and were significantlydifferent to survival with fresh tolerant cells (p b 0.001) or tolerantcells cultured with ConAsup and PVG stimulator cells (p b 0.001).Taken together these results showed that rIL-4 alone was not sufficientto promote survival of tolerance transferring CD4+ T cells.
As IL-4 could be one of the many cytokines in ConAsup thatpromoted the survival of tolerant cells, we cultured tolerant cells withdonor alloantigen and ConAsup blocked with MRCOx81, an anti-IL-4mAb. MRCOx81 inhibited cell proliferation (data not shown). Alone,these cells transferred tolerance (Table 1, line 9) and retained thecapacity to suppress naïve CD4+ T cell mediated rejection (Table 1,line 8).
These results indicate that cytokines in ConAsup, other than IL-4,were sufficient to sustain the antigen specific Treg within the CD4+ Tcells, and therefore IL-4 is not essential for themaintenance of tolerance

Table 1Examination of the capacity of CD4+T cells fromDA ratswith tolerance to PVG cardiac to transfer tolerance to irradiatedDA ratswith either PVGor Lewis cardiac allografts. Effect of culturewith PVG stimulators and ConA supernatant (ConAsup) with IL-4 blocking mAb.
Restorative innocula PVG graft rejection time (days) Lewis graft rejection time (days) Significance(p)
2 × 107 tolerant CD4+ 5× 106 naïve CD4+ Median Actual Median Actual
1 – − N100 N100 (10) N100 25, N100 (3) NSD2 Fresh − N100 N100 (6) 11.5 9,10, 13 (2) p b 0.013 – + 13 12 (6), 13 (2), 14 (4), N100 13 9, 10 (3), 13, 17, 25, N100 NSD4 Fresh + N100 11, 12, N100 (6) 13 9, 10, 13, 25, 39 p b 0.055 Cultured
No cytokine15 11, 14, 15 (3), N100 ND
6 Cultured with ConAsup + N90 11, N32 (4), N90 (2), N100 (4) 18 8, 10 (2), 18 (3), 41, N47, 100 (3) p b 0.017 Cultured with rIL-4 − 7 7 (5), 8, 9 7 7 (3), 8, 11 NSD8 Cultured with ConAsup & anti-IL-4
mAb⁎⁎+ N100 8, 10, 14, N30, N100 (6) ND
9 Cultured with ConAsup & anti-IL-4 mAb⁎ − N100 N100 (3) ND10 – +Culturedwith ConA sup 10.5 8 (2), 9, 10 (2), 11, 12, 13, 14, N100 ND
ND: not done, NSD: not significantly different.⁎ Some control results are shared or rolling controls already published.⁎⁎ Some cells were prepared from anti-CD4 monoclonal antibody induced tolerance as described [45].
56 K.M. Plain et al. / Transplant Immunology 29 (2013) 51–59
mediating cells. It was possible that the rIL-4 was promoting T effectorcells, as there are more CD4+CD25+FoxP3− T cells after culture inMLC with IL-4 (Fig. 2). Thus we next examined the effect of rIL-4 onthe CD4+CD25+ T cells, which in fresh tolerant CD4+ T cells are 80%FoxP3+.
3.4. Effect of rIL-4 on CD4+CD25+ and CD4+CD25− subsets from toleranthosts
Naïve and tolerant CD4+CD25+ T cells were enriched to N95% purityand both have 70–80% FoxP3+ cells, as described [13,23,34]. Cultures ofnaïve CD4+CD25+FoxP3+ Treg with rIL-4 and PVG are activated toalloantigen-specific Ts2 cells that express IL-5Rα and suppress CD4+ Tcell mediated rejection of PVG but not Lewis [23].
The proliferative responses of naïve (Fig. 4A) and tolerant (Fig. 4B).CD4+CD25+ T cells to self and alloantigens with or without rIL-4 wereexamined. Naïve CD4+CD25+ T cells had a small response toalloantigen with no cytokines added and proliferation was markedlyenhanced by rIL-4 (Fig. 4A), as described [23].
Tolerant CD4+CD25+ T cells responded differently to naïve nTreg, inthat their proliferation against specific donor PVG stimulators in theabsence of cytokines was similar to self and at background (Fig. 4B),but they responded to third party allogeneic stimulators similar tonTreg. Tolerant CD4+CD25+ T cell response to PVG and third partystimulators was enhanced by rIL-4 (Fig. 4B). Thus rIL-4 enhanced theresponse of both naïve and tolerant CD4+CD25+ T cells in MLC.
Tolerant CD4+CD25+ T cells that had been activated by PVG and rIL-4 in MLC were examined in a secondary MLC (Fig. 4C). Both controlCD4+CD25+ T cell cultures and rIL-4 primed cultures had a similarsecondary response (Fig. 4C), as proliferation to no stimulator cells(background), self (DA) and PVG stimulators were similar (NSD), butthey proliferated against third party to a greater degree than to nostimulator cells, self (DA) and specific donor stimulator cells (PVG)(pb0.05).
As naïve nTreg alloactivated with rIL-4 are induced to Ts2 thatexpress IL-5Rα and IFN-γ, but do not express IFNGR and IL-5 [23], weexamined if similar changes were induced by culture of tolerantCD4+CD25+ T cells with alloantigen and rIL-4. Pre-culture, tolerantCD4+CD25+ T cells expressed FoxP3 and markers of both Ts1 and Ts2cells, in that they expressed IFNGR and IL-5Rα (Fig. 4D). However,after culture with specific donor alloantigen and rIL-4, the expression ofboth IFNGR and IL-5Rα was lost (Fig. 4E). This was consistent with theloss of both Ts1 and Ts2 antigen-specific CD4+CD25+ Treg. Concurrentculture of nTreg with alloantigen and rIL-4 induced IL-5Rα, the Ts2phenotype [23] (not shown).
Taken together these results were consistent with rIL-4 notsupporting the survival and proliferation of the antigen specific Tregthat mediate specific tolerance, but still enhancing polyclonalproliferation of nTreg. These results suggested that specific donorreactive naïve nTreg may have all been activated and thus depletedthe nTreg pool of cells that can respond to specific donor. All wouldhave been activated to alloantigen specific Treg that do not depend onIL-4, but need other cytokines found in ConAsup to survive andproliferate.
3.5. Capacity of tolerant CD4+CD25+ T cells MLC with rIL-4 to suppressrejection
When tolerant CD4+CD25+ T cells were cultured in MLC againstspecific donor PVG stimulators with rIL-4, the ability to transfer specifictolerance was lost. PVG allografts were rejected when 2×106 rIL-4 andPVG stimulated tolerant CD4+CD25+ T cells were co-transferred with5 × 106 naïve CD4+ T cells, which is in contrast to the effect oftransferring 20 × 106 freshly isolated tolerant CD4+ T cells (Fig. 4F).2 × 106 CD4+CD25+ T cells were given as this represents the fraction(10%) of CD4+CD25+ cells present in 20 × 106 tolerant CD4+ T cells.However, 2 × 106 tolerant CD4+CD25+ T cells cultured with rIL-4 andspecific donor alloantigen (PVG), prevented rejection of third partyLewis grafts (Fig. 4G). This was consistentwith non-specific suppressionby nTreg manifest at high ratios of nTreg to effector CD4+ T cells [19].This suggested that tolerant activated alloantigen-specific Treg died inculture with rIL-4 and specific donor alloantigen, whereas naïve nTregthat can suppress non-specific donor alloantigen survived due topolyclonal activation by rIL-4.
4. Discussion
The function of alloantigen-specific CD4+CD25+ Treg in transplanttolerance is poorly understood and the role of IL-4 in the developmentand maintenance of this tolerance is also unclear [35]. We hypothesisethat alloantigen-specific Treg have different characteristics and functionsto that of nTreg, some of which have been identified in this study. Wepreviously identified that these specific Treg are present in the CD4+ Tcell fraction [2,4,5,13,36] and are short-lived in vitro [6,7], even whenstimulated with specific donor alloantigen, but can survive iflymphocyte-derived cytokines are added in the form of ConAsup[5,7,13]. IL-2 alone was insufficient to promote full survival [7].
Different effects of IL-4 on the induction and maintenance oftransplant tolerance were found. IL-4 promotes the expansion ofnTreg [23,26,27] as we previously found that co-culture with IL-4 andalloantigen induced CD4+CD25+FoxP3+ Ts2 cells that had enhanced

A Tolerant
0
500
1000
1500
2000
2500
B Secondary MLC Tolerant CD25+
PrimingCD25+/PVG
0
2000
4000
6000No stimulators
0
1000
2000
3000
4000
5000
6000
CP
M
Naive
DA
PVG
D
0
1000
2000
3000
4000
5000
Cop
y nu
mbe
r (n
orm
alis
ed to
GA
PD
H)
Freshly isolated Tolerant CD4+CD25+
Lewis
DAPVG LewisDA
PVG
0
1 x 107
2 x 107
3 x 107
4 x 107
5 x 107
6 x 107
Untreated rIL-4 CD25+/PVG +IL-4Untreated rIL-4
FoxP3 IFNg IFNGR IL-5 IL-5RA Gata 3 T-betFoxP3 IFNg IFNGR IL-5 IL-5RA Gata 3 T-bet
MLC Tolerant CD4+CD25+ with IL-4DA
PVGLewis
E
C
1008060402000
20
40
60
80
100 Specific donor
Days post-transplantation
Allo
graf
t sur
viva
l (%
)
F
1008060402000
20
40
60
80
100 Third party
Days post-transplantation
B
G
5x106 naïve CD4+
20x106 tol CD4+ & 5x106 naïve CD4+
2x106 tol CD25+ MLC/IL-4 & 5x106 naïve CD4+
ee
g
g
g
f
a
a
c
b
b
d
d
Fig. 4. Responses of tolerant CD4+CD25+ T cell cultured with rIL-4 in MLC. A. rIL-4 increase the proliferation of naïve CD4+CD25+ T cells to both self and PVG alloantigen, compared tocontrolswith no cytokine (a; pb0.01). B. rIL-4 increased proliferation of CD4+CD25+ T cells from a DA rat tolerant to a PVG graft to specific donor and third party stimulators (b; pb0.01),and to a lesser extent self. c. Untreated response to Lewis was significantly different (p b 0.05) to DA and PVG. C. Proliferation of CD4+CD25+ T cells from DA rats tolerant to PVG insecondary MLC. Cells were cultured in a primary MLC with specific donor PVG stimulators, either with rIL-4 (right group) or with no cytokine added (left group). TolerantCD4+CD25+ T cells culturedwith or without IL-4 response in secondaryMLC to PVGwas not different to DA (self) or no stimulator cells (background), however they retained a responseto third party Lewiswhichwas greater than their response to either DA, PVG andno stimulator cells (d,pb0.05). Quantitative RT-PCR forD. freshly isolated tolerant CD4+CD25+T cells andE. tolerant CD4+CD25+ cells cultured inMLCwith rIL-4. Normalized copynumber expressed as the ratio of specific geneper 103 copies ofGAPDH. e.pb0.01 compared to allMLC treatmentgroups for the same gene; f. pb0.01 compared to freshly tolerant cells for all other genes g. pb0.01 compared toMLC cells versusDA stimulators. Fresh tolerant cells had high expression ofIFNGR and expressed IL-5Rα respectively consistent with the presence of Ts1 and Ts2 cells. Therewas low expression of Tbet andGata-3 but the expression of FoxP3 confirm that theywereTreg, not Th1 or Th2 cells. Tolerant CD4+CD25+T cells culturedwith rIL-4 and PVG, had no expression of IFNGR or IL-5Rα consistentwith the loss of antigen specific Ts1 and Ts2 cells. Therewas low IFN-γ expression. F and G. Capacity of tolerant CD4+CD25+ T cells cultured with rIL-4 and alloantigen to transfer transplant tolerance to specific donor and third party grafts. Allgroups were reconstituted with 5× 106 naïve CD4+ T cells which restored rejection (n=12). 20× 106 tolerant freshly isolated CD4+ T cells suppressed rejection of specific donor graftsthat should have beenmediated by co-transferred 5×106 naïve CD4+ T cells (n=6). 2×106 tolerant CD4+CD25+ T cells cultured inMLCwith added rIL-4 (ratio 1:2.5 with CD4+ T cells)did not transfer tolerance to specific donor. Paradoxically 2×106 rIL-4 culturedCD4+CD25+T cells suppressed rejection of third party Lewis grafts by the co-transferred naïve CD4+T cells.These results suggested that antigen specific tolerance mediating CD4+CD25+FoxP3+ T cells did not survive in culture with IL-4, whereas the nTreg did and could suppress third partygraft rejection at a ratio of 1:2.5.
57K.M. Plain et al. / Transplant Immunology 29 (2013) 51–59
capacity to suppress naïve CD4+ T cells in MLC at 1:32–1:64. Ts2 cellssuppress rejection of specific donor but not third party allografts inadoptive hosts at a ratio of 1:10 [23]. This suggested that short-termactivation of nTreg with alloantigen and rIL-4 induced a more potent
alloantigen-specific Ts2 cell that may be similar to CD4+CD25+FoxP3+
Treg that maintain and transfer transplant tolerance.As rIL-4 therapy delays neonatal heart allograft rejection [37], we
examined if culture of naïve lymphocytes, containing both CD4+CD25+

58 K.M. Plain et al. / Transplant Immunology 29 (2013) 51–59
T cells and CD4+CD25− T cells, could develop a regulatory function ifactivated by specific alloantigen in the presence of rIL-4. A Th2-likedominant phenotype was shown in vitro and in vivo, however thesecells effected specific donor rejection on adoptive transfer, consistentwith other reports that Th2 alloantigen-specific cells effect rejection,not tolerance [38–41]. Interestingly, rIL-4 alloactivated CD4+ T cellsexpressed very high CD25 levels with an increased proportion ofCD4+CD25+FoxP3− T cells, consistent with IL-4 preferentially activatingeffector cells, not CD4+CD25+FoxP3+ Treg. IL-4 induces Th2 cells toresist Treg [42] and inhibits induction of iTreg [43,44]. These findingssuggest that IL-4 promotes cells that effect rejection as well as activatingnTreg to become alloantigen-specific Ts2. Rejection usually dominatesunless effector T cell activation is limited by immunosuppression.
This study confirmed that the capacity of tolerant CD4+ T cells totransfer specific tolerance to an adoptive host is lost in culture within3 d, even when they are cultured with specific donor alloantigen. Wehypothesise that the tolerance transferring Treg die in culture and theeffector T cells survive and mediate rejection of specific donor and thirdparty grafts. We also confirmed that ConAsup, a mixture of cytokinesproduced by activated lymphocytes, preserves the suppressor functionof tolerant CD4+ T cells. However culture with rIL-4 alone, even withspecific donor alloantigen, did not preserve the suppressor function oftolerant CD4+ T cells. Tolerant CD4+ T cells cultured with IL-4 andspecific donor alloantigen lost the capacity to transfer tolerance andrapidly effected specific donor rejection. Blocking IL-4 in the ConAsupdid not reduce the capacity of the ConAsup to preserve the suppressorfunction of tolerant CD4+ T cells. Thus, when tolerant CD4+ T cellswere cultured with specific alloantigen and ConAsup, their capacity totransfer tolerance was partially preserved when IL-4 was blocked. InDA rats with PVG grafts, blockade of IL-4 also does not impede inductionof transplant tolerance [45], nor transfer of tolerance [24,46], althoughin other models it partially [13,47] or fully [48], blocked transfer oftolerance. Taken together these findings suggested that survival ofalloantigen specific tolerancemediating CD4+ T cells was not dependenton IL-4, and that IL-4 alone could not promote survival of these cells.
We demonstrated that rIL-4 promotes activation of FoxP3− CD4+ Tcells and induces them to express the activation marker CD25. Whentolerant CD4+ T cells were stimulated with specific donor alloantigen,they had an enhanced proliferation in the presence of rIL-4 and themajority was CD4+CD25+FoxP3− T cells. Thus rIL-4 may preferentiallypromote effector Th2 CD4+ cells. rIL-4 could not promote survival oftolerant CD4+CD25+FoxP3+ T cells, and they also did not respondin vitro to specific donor alloantigen, or transfer tolerance to specificdonor. The cultured tolerant CD4+CD25+ T cells appeared to lacknTreg that respond to donor alloantigen upon re-challenge, suggestingthat all of the donor reactive Treg had been activated and depletedfrom the nTreg pool. Culture of tolerant CD4+CD25+ T cells with rIL-4and specific donor alloantigen did not enhance their capacity to transfertolerance and suppress rejection by naïve CD4+ T cells, though they didsuppress rejection of third party grafts when given at a high ratiocompared to naïve CD4+ T cells. This is consistent with IL-4 supportingnTreg that could suppress third party rejection rather than alloantigen-specific Treg, which can suppress at high ratios to effector cells, asdescribed [19].
Gene expression studies supported these findings of loss of functionof alloantigen-specific Treg when cultured with rIL-4. Freshly isolatedtolerant CD4+CD25+ T cells expressed both IFNGR and IL-5Rα,consistent with the presence of both Ts1 and Ts2 [23]. IFNGR andIL-5Rα expression was lost during culture with alloantigen and rIL-4,consistent with the failure of rIL-4 alone to sustain tolerancetransferring antigen specific CD4+CD25+FoxP3+ Treg. Taken together,these results showed that IL-4 alone was unable to promote survivalof alloantigen specific CD4+ T cells especially the CD4+CD25+FoxP3+
Treg that maintain tolerance to a specific allograft [5,13].Our hypothesis, summarized in reviews [11,12] is that the survival of
antigen-specific CD4+CD25+FoxP3+ Treg that maintain tolerance
depends upon cytokines produced by activated effector CD4+FoxP3−
T cells reacting to the persistent alloantigen in the graft (reviewed in[40]). Our previous studies showed that the initial activation of nTregby antigen can be promoted by either IL-2 or IL-4, and that separatelineages of antigen specific Ts1 and Ts2 cells are induced that respec-tively express IFNGR and IL-5Rα. After this initial activation, the effectorcells no longer produce IL-2 or IL-4, but produce later Th1 and Th2cytokines such as IFN-γ and IL-5 respectively [20,40].
Although rIL-4 promotes the initial activation of alloantigen-specificCD4+CD25+FoxP3+ Treg, once tolerance develops rIL-4 did not supportthe antigen-specific CD4+CD25+Treg thatmaintain this tolerance. ThusIL-4 may play little or no role in the maintenance of CD4+ T cellmediated tolerance. The expression of IL-5Rα by Ts2 cells and the abilityof IL-5 to promote auto-antigen-specific CD4+CD25+FoxP3+ Ts2in vitro and in vivo [49], makes IL-5 a candidate for promoting transplanttolerant CD4+CD25+ T cells.
Acknowledgements
This work was supported by grants from Bob and Jack Ingham,Liverpool, Australia, NH&MRC Australia, UNSW, Sydney South WestArea Health Research Foundation andNSWHealth. At Liverpool Hospitalthe authorswould like to acknowledge the expert technical assistance ofMr. Botros in the animal house, the Haematology Department for use ofan irradiator and the Radiotherapy Department for irradiation ofanimals. IL-4 transfected cell line and the anti-IL-4monoclonal antibodyclone were kindly provided by Dr. Neil Barclay, and the William DunnSchool of Pathology Oxford UK. We greatly appreciate the work of Mr.Austin Spinelli, Dr. Steve Merten and Dr. J. Chen who performed theheterotopic heart grafts.
References
[1] Hall B. The cellular basis of the maintenance of passive enhancement. TransplantProc 1981;13:403–6.
[2] Hall BM, Jelbart ME, Dorsch SE. Suppressor T cells in rats with prolonged cardiacallograft survival after treatmentwith cyclosporine. Transplantation 1984;37:595–600.
[3] Graca L, Thompson S, Lin CY, Adams E, Cobbold SP, Waldmann H. Both CD4+CD25+
and CD4+CD25− regulatory cells mediate dominant transplantation tolerance. JImmunol 2002;168:5558–65.
[4] Hall BM, Jelbart ME, Gurley KE, Dorsch SE. Specific unresponsiveness in rats withprolonged cardiac allograft survival after treatment with cyclosporine. Mediationof specific suppression by T helper/inducer cells. J Exp Med 1985;162:1683–94.
[5] Hall BM, Pearce NW, Gurley KE, Dorsch SE. Specific unresponsiveness in rats withprolonged cardiac allograft survival after treatment with cyclosporine. III. Furthercharacterization of the CD4+ suppressor cell and its mechanisms of action. J ExpMed 1990;171:141–57.
[6] Pearce NW, Spinelli A, Gurley KE, Dorsch SE, Hall BM. Mechanisms maintainingantibody-induced enhancement of allografts. II. Mediation of specific suppressionby short lived CD4+ T cells. J Immunol 1989;143:499–506.
[7] Pearce NW, Spinelli A, Gurley KE, Hall BM. Specific unresponsiveness in rats withprolonged cardiac allograft survival after treatment with cyclosporin V. Dependenceof the CD4+ suppressor cell on the presence of alloantigen and cytokines, includinginterleukin-2. Transplantation 1993;55:374–80.
[8] Qin S, Cobbold SP, Pope H, Elliott J, Kioussis D, Davies J, et al. “Infectious”transplantation tolerance. Science 1993;259:974–7.
[9] Dai Z, Li Q, Wang Y, Gao G, Diggs LS, Tellides G, et al. CD4+CD25+ regulatory T cellssuppress allograft rejection mediated by memory CD8+ T cells via a CD30-dependent mechanism. J Clin Invest 2004;113:310–7.
[10] Kingsley CI, Karim M, Bushell AR, Wood KJ. CD25+CD4+ regulatory T cells preventgraft rejection: CTLA-4- and IL-10-dependent immunoregulation of alloresponses. JImmunol 2002;168:1080–6.
[11] Hall BM, Tran GT, Verma ND, Plain KM, Robinson CM, Nomura M, et al. Do natural Tregulatory cells become activated to antigen specific T regulatory cells intransplantation and in autoimmunity? Front Immunol 2013;4:208.
[12] Hall BM, Verma ND, Tran GT, Hodgkinson SJ. Distinct regulatory CD4+ T cell subsets;differences between naïve and antigen specific T regulatory cells. Curr OpinImmunol 2011;23:1–7.
[13] Hall BM, Plain KM, Verma ND, Tran G, Boyd R, Robinson CM, et al. Transfer ofallograft-specific tolerance requires CD4 + CD25 + T cells, but not IL-4 or TGF-βand cannot induce tolerance to linked antigens. Transplantation 2007;83:1075–84.
[14] Sakaguchi S, Hori S, Fukui Y, Sasazuki T, Sakaguchi N, Takahashi T. Thymic gen-eration and selection of CD25+CD4+ regulatory T cells: implications of their broadrepertoire and high self-reactivity for the maintenance of immunological self-tolerance. Novartis Found Symp 2003;252:6–16 [discussion -23, 106–14].
[15] Shevach EM. Regulatory T cells. Introduction. Semin Immunol 2004;16:69–71.

59K.M. Plain et al. / Transplant Immunology 29 (2013) 51–59
[16] Jiang X, Morita M, Sugioka A, Harada M, Kojo S, Wakao H, et al. The importance ofCD25+CD4+ regulatory T cells in mouse hepatic allograft tolerance. Liver Transpl2006;12:1112–8.
[17] Kies MW. Chemical studies on an encephalitogenic protein from guinea pig brain.Ann N Y Acad Sci 1965;122:161–72.
[18] Taylor PA, Noelle RJ, Blazar BR. CD4+CD25+ immune regulatory cells are requiredfor induction of tolerance to alloantigen via costimulatory blockade. J Exp Med2001;193:1311–7.
[19] Nomura M, Plain KM, Verma N, Robinson C, Boyd R, Hodgkinson SJ, et al. The cellularbasis of cardiac allograft rejection. IX. Ratio of naive CD4+CD25+ T cells/CD4+CD25− T cells determines rejection or tolerance. Transplant Immunol 2006;15:311–8.
[20] Hall BM. Mechanisms of induction of tolerance to organ allografts. Crit Rev Immunol2000;20:267–324.
[21] Shevach EM, Davidson TS, Huter EN, Dipaolo RA, Andersson J. Role of TGF-beta in theinduction of Foxp3 expression and T regulatory cell function. J Clin Immunol2008;28:640–6.
[22] McClymont SA, Putnam AL, LeeMR, Esensten JH, LiuW, HulmeMA, et al. Plasticity ofhuman regulatory T cells in healthy subjects and patients with type 1 diabetes. JImmunol 2011;186:3918–26.
[23] Verma ND, Plain KM, Nomura M, Tran GT, Robinson CM, Boyd R, et al. CD4+CD25+Tcells alloactivated ex vivo by IL-2 or IL-4, become potent alloantigen specificinhibitors of rejection with different phenotypes, suggesting Th1 and Th2 responsesactivate by separate pathways. Blood 2009;113:479–87.
[24] Hall BM, Fava L, Chen J, Plain KM, Boyd RA, Spicer ST, et al. Anti-CD4 monoclonalantibody-induced tolerance to MHC-incompatible cardiac allografts maintained byCD4+suppressor T cells that arenot dependentupon IL-4. J Immunol 1998;161:5147–56.
[25] Dwyer JM, Johnson C. The use of concanavalin A to study the immunoregulation ofhuman T cells. Clin Exp Immunol 1981;46:237–49.
[26] Pillai AB, George TI, Dutt S, Strober S. Host natural killer T cells induce an interleukin-4-dependent expansion of donor CD4+CD25+Foxp3+ T regulatory cells thatprotects against graft-versus-host disease. Blood 2009;113:4458–67.
[27] Thornton AM, Piccirillo CA, Shevach EM. Activation requirements for the inductionof CD4+CD25+ T cell suppressor function. Eur J Immunol 2004;34:366–76.
[28] Pearce NW, Berger MF, Gurley KE, Spinelli A, Hall BM. Specific unresponsiveness inrats with prolonged cardiac allograft survival after treatment with cyclosporine. VI.In vitro alloreactivity of T cell subsets from rats with long-surviving allografts.Transplantation 1993;55:380–9.
[29] Plain KM, Fava L, Spinelli A, He XY, Chen J, Boyd R, et al. Induction of tolerance withnondepleting anti-CD4 monoclonal antibodies is associated with down regulation ofTh2 cytokines. Transplantation 1997;64:1559–67.
[30] Hall BM, Dorsch S, Roser B. The cellular basis of allograft rejection in vivo. I. The cellularrequirements for first-set rejection of heart grafts. J Exp Med 1978;148:878–89.
[31] McKnight AJ, Barclay AN, Mason DW. Molecular cloning of rat interleukin 4 cDNAand analysis of the cytokine repertoire of subsets of CD4+ T cells. Eur J Immunol1991;21:1187–94.
[32] Verma ND, Boyd R, Robinson C, Plain KM, Tran GT, Hall BM. Interleukin-12p70prolongs allograft survival by induction of interferon gamma and nitric oxideproduction. Transplantation 2006;82:1324–33.
[33] Goodwin JS. Modulation of conconavalin A induced suppressor cell activation byprostaglandin E2. Cell Immunol 1980;49:421–5.
[34] Hall BM, Robinson CM, Plain KM, Verma ND, Carter N, Boyd R, et al. Studies on naïveCD4+CD25+T cells inhibition of naïve CD4+CD25− T cells in mixed lymphocytecultures. Transpl Immunol 2008;18:291–300.
[35] Tay SS, Plain KM, Bishop GA. Role of IL-4 and Th2 responses in allograft rejection andtolerance. Curr Opin Organ Transplant 2009;14:16–22.
[36] Aversa GG, Waugh JA, Bishop GA, Hall BM. Use of monoclonal antibodiesto study in vivo and in vitro-activated lymphocytes. Transplant Proc1989;21:349–50.
[37] He XY, Chen J, Verma N, Plain K, Tran G, Hall BM. Treatment with interleukin-4prolongs allogeneic neonatal heart graft survival by inducing T helper 2 responses.Transplantation 1998;65:1145–52.
[38] Barbara JA, Turvey SE, Kingsley CI, Spriewald BM, Hara M, Witzke O, et al.Islet allograft rejection can be mediated by CD4+, alloantigen experienced,direct pathway T cells of Th1 and Th2 cytokine phenotype. Transplantation2000;70:1641–9.
[39] Matesic D, Valujskikh A, Pearlman E, Higgins AW, Gilliam AC, Heeger PS. Type 2immune deviation has differential effects on alloreactive CD4+ and CD8+ T cells. JImmunol 1998;161:5236–44.
[40] VanBuskirk AM, Wakely ME, Orosz CG. Transfusion of polarized Th2-like cellpopulations into SCID mouse cardiac allograft recipients results in acute allograftrejection. Transplantation 1996;62:229–38.
[41] Zelenika D, Adams E, Mellor A, Simpson E, Chandler P, Stockinger B, et al. Rejectionof H-Y disparate skin grafts by monospecific CD4+ Th1 and Th2 cells: norequirement for CD8+ T cells or B cells. J Immunol 1998;161:1868–74.
[42] Pillemer BB, Qi Z, Melgert B, Oriss TB, Ray P, Ray A. STAT6 activation confers uponT helper cells resistance to suppression by regulatory T cells. J Immunol2009;183:155–63.
[43] Dardalhon V, Awasthi A, Kwon H, Galileos G, Gao W, Sobel RA, et al. IL-4 inhibitsTGF-beta-induced Foxp3+ T cells and, together with TGF-beta, generates IL-9+ IL-10+ Foxp3(−) effector T cells. Nat Immunol 2008;9:1347–55.
[44] Veldhoen M, Uyttenhove C, van Snick J, Helmby H, Westendorf A, Buer J, et al.Transforming growth factor-beta ‘reprograms’ the differentiation of T helper 2cells and promotes an interleukin 9-producing subset. Nat Immunol 2008;9:1341–6.
[45] Plain KM, Boyd R, Verma N, Robinson C, Tran G, Hodgkinson SJ, et al. Transplanttolerance associated with a Th1 response and not broken by IL-4. IL-5 and TGF-beta blockade or Th1 cytokine administration. Transplantation 2007;83:764–73.
[46] Chen ZK, Cobbold SP, Waldmann H, Metcalfe S. Amplification of natural regulatoryimmunemechanisms for transplantation tolerance. Transplantation 1996;62:1200–6.
[47] Davies JD, Martin G, Phillips J, Marshall SE, Cobbold SP, Waldmann H. T cellregulation in adult transplant tolerance. J Immunol 1996;156:529–33.
[48] Bushell A, Niimi M, Morris PJ, Wood KJ. Evidence for immune regulation in theinduction of transplantation tolerance: a conditional but limited role for IL-4. JImmunol 1999;162:1359–66.
[49] Tran GT, Hodgkinson SJ, Carter NM, Verma ND, Plain KM, Boyd R, et al. Interleukin-5(IL-5) promotes induction of antigen specific CD4+ CD25+ T regulatory cells thatsuppress autoimmunity. Blood 2012;119:4441–50.




















