
Covalent binding of carbon tetrachloride reactive metabolites to liver microsomal and nuclear lipid...
12
Teratogenesis, Carcinogenesis, and Mutagenesis 15: 155-166 (1995) Covalent Binding of Carbon Tetrachloride Reactive Metabolites to Liver Microsomal and Nuclear Lipid and Phospholipid Classes From Sprague Dawley and Osborne Mendel Male Rats S.L. Fanelli and J.A. Castro Centro de lnvestigaciones Toxicologicas (CEITOX) CITEFNCONICET; Buenos Aires, Argentina The interaction between carbon tetrachloride (CCLJ-reactive metabolites and liver microsomal or nuclear lipids (covalent binding = CB) was studied in male rats from Osbome Mendel (OM) and Sprague Dawley (SD), strains with different cancer sus- ceptibility. The in vitro or in vivo CB was more intense in OM than in SD. Most of the CB was with phospholipid (PL; SD > OM). The CB to cholesterol (CH) and cholesterol esters (CHE) was OM > SD. We also observed the presence of specific adducts only present in lipids from either OM or SD strains. The results were related to the well-known role of PL and CH derivatives in gene function control and cell growth. o 1995 Wiley-Liss, Inc. Key words: covalent binding, CCI,, lipid, phospholipid, cholesterol, nuclei, microsomes INTRODUCTION Previous work from our laboratory evidenced the abilitj of liver nuclei to biotransform carbon tetrachloride (CC14) to CC13 and to reactive metabolites that either covalently bind to nuclear lipids, proteins, and DNA or spark a lipid peroxidation process [ 1-71. Further detailed studies on the deleterious consequences for liver nuclei of the above CC14 initiated processes revealed that as a result of the DNA attack by the CC13 as many as 20 altered bases might be formed [7], and that all nuclear protein fractions participate in the interaction [ 1,6], presumably by the formation of aminoacids adducts with the CC13 [4,8], or potentially through the generation of Address reprint requests and correspondence to Dr. J.A. Castro, Centro de InvestigacionesToxicologicas (CEITOX) CITEFA/CONICET,Zufriategui 4380, (1 603) Villa Martelli, Buenos Aires, Argentina. 0 1995 Wiley-Liss, Inc.
-
Upload
conicet-ar -
Category
Documents
-
view
5 -
download
0
Transcript of Covalent binding of carbon tetrachloride reactive metabolites to liver microsomal and nuclear lipid...

Teratogenesis, Carcinogenesis, and Mutagenesis 15: 155-166 (1995)
Covalent Binding of Carbon Tetrachloride Reactive Metabolites to Liver Microsomal and Nuclear Lipid and Phospholipid Classes From Sprague Dawley and Osborne Mendel Male Rats
S.L. Fanelli and J.A. Castro Centro de lnvestigaciones Toxicologicas (CEITOX) CITEFNCONICET; Buenos Aires, Argentina
The interaction between carbon tetrachloride (CCLJ-reactive metabolites and liver microsomal or nuclear lipids (covalent binding = CB) was studied in male rats from Osbome Mendel (OM) and Sprague Dawley (SD), strains with different cancer sus- ceptibility. The in vitro or in vivo CB was more intense in OM than in SD.
Most of the CB was with phospholipid (PL; SD > OM). The CB to cholesterol (CH) and cholesterol esters (CHE) was OM > SD. We also observed the presence of specific adducts only present in lipids from either OM or SD strains.
The results were related to the well-known role of PL and CH derivatives in gene function control and cell growth. o 1995 Wiley-Liss, Inc.
Key words: covalent binding, CCI,, lipid, phospholipid, cholesterol, nuclei, microsomes
INTRODUCTION
Previous work from our laboratory evidenced the abilitj of liver nuclei to biotransform carbon tetrachloride (CC14) to CC13 and to reactive metabolites that either covalently bind to nuclear lipids, proteins, and DNA or spark a lipid peroxidation process [ 1-71.
Further detailed studies on the deleterious consequences for liver nuclei of the above CC14 initiated processes revealed that as a result of the DNA attack by the CC13 as many as 20 altered bases might be formed [7], and that all nuclear protein fractions participate in the interaction [ 1,6], presumably by the formation of aminoacids adducts with the CC13 [4,8], or potentially through the generation of
Address reprint requests and correspondence to Dr. J.A. Castro, Centro de Investigaciones Toxicologicas (CEITOX) CITEFA/CONICET, Zufriategui 4380, (1 603) Villa Martelli, Buenos Aires, Argentina.
0 1995 Wiley-Liss, Inc.

156 Fanelli and Castro
aminoacid breakdown products [2,3]. Positive correlation between CB to acidic non- histone proteins and liver susceptibility to CC14 carcinogenic effects in different spe- cies was found in those studies [2]. No apparent correlation was demonstrated between carcinogenic response and covalent binding (CB) to DNA or with CCl, promoted lipid peroxidation [2,3]. No equivalent detailed studies are available for the CB to liver nuclear or microsomal lipid classes and their correlation or not with CCl, liver cancer susceptibility.
The present work describes initial efforts to gain knowledge about the CB of CC1,-reactive metabolites to different nuclear and microsomal lipids from rat strains having a marked difference in the carcinogenic response to CCl,, the Sprague Dawley (non-susceptible), and the Osborne Mendel (responsive) male rats [9].
MATERIALS AND METHODS Chemicals
['4C]CC14 (specific activity: 4.10 mCi/mmol) was purchased from DuPont New England Nuclear, Boston, Massachusetts, USA. The chemical was greater than 99% pure when determined by gas liquid chromatography using a Carbowax 400 Amapak Porasil C column at 65°C. Experiments were performed within the 6 months follow- ing this analysis. The rate of radiolytic decomposition of ['4C]CC14 is less than 1% for 18 months stored as a liquid at -20°C. In our laboratory we had it at -70°C in the darkness. Consequently, expected rates of radiolysis of the [ ''C]CC14 employed should be nonsignificant. Other chemicals employed were of the best available quality.
Animals and Treatments Non-inbred male rats of the following strains were used: Sprague Dawley (SD;
240-250 g) and Osborne Mendel (OM; 240-260 g). The animals were starved for 12-14 h before CCl, administration or before being killed (nuclei or microsomal isolation experiments). Water was available ad libitum. Temperature in the animal room was 23 k 2°C and the relative humidity was between 45 and 65%. Light in the animal room was on from 6 A.M. to 6 P.M. In the study on in vivo covalent binding of [I4C] from ['4C]CC14 to liver lipids and phospholipids, ['4C]CC14 (6.86 x lo7 dpm/ ml) was given ip as a 5% (v/v) solution in olive oil (5 ml of this solution per kg). The animals were killed by decapitation 3 h after ['4C]CC14 administration, and their livers were rapidly removed and processed.
Methods Microsomal fractions and nuclear preparations were obtained as previously de-
scribed. Purity of the preparations was routinely checked by phase contrast micros- copy. Only traces of contamination with endoplasmic reticulum or other organelles were found in preparations via this procedure when checked by electron microscopy t3,6,101.
The in vitro binding of ["C] from ['4C]CC14 to nuclear or microsomal lipids and phospholipids was performed as follows: Incubation mixtures containing purified nuclei [( 10-1 2 mg protein/ml) in 0.25 M sucrose/50 mM Tris-HC1 (pH 7.5)/2.5 mM KC1/5 mM MgC12 (final volume 3 ml)] or microsomal suspensions [(3.34.3 mg protein/ml) in 50 mM Tris-HC1 (pH 7.5)/MgC12 (final volume 3 ml)]; with NADPH generating system [ 1,5,6] and 0.2 mM ['4C]CC14 (2 x lo6 dpm/ml) were conducted 1 h

Covalent Binding to Lipids 157
at 37°C under anaerobic atmosphere in a Dubnoff shaker septum stoppered glass vials. The reaction was interrupted by putting incubation flaks in ice cold.
The irreversible binding of 14C to lipids was assessed by extracting the incuba- tion mixtures with chloroform-methanol [(2: 1) v/v, 0.01 % p/v 2,6-diterbutyl-pocresol (BHT)]; labeled lipids isolated were separated into classes by Silica gel thin layer chromatography. Phospholipids were further separated into different fractions [ 1 1,121. All of the lipid and phospholipid fractions were quantitatively transferred to scintil- lation counting vials and counted.
Statistics A decision tree for selecting statistical procedures [13] was applied to results
from every experiment. The appropriate parametric (unpaired t) or non-parametric (Mann-Whitney) tests were applied. Calculations were performed using adequate com- puter programs [ 141.
RESULTS Covalent Binding of [14C] (CB) From I4CCl4 to Microsomal and Nuclear Lipids From Sprague Dawley (SD) and Osborne Mendel (OM) Male Rats
Microsomal suspensions from SD and OM rats were able to activate CC14 to reactive metabolites that bind covalently to lipids both in the absence and in the presence of NADPH. Activation in the presence of NADPH is 26.45 times more intense than in the absence of cofactor in the SD and 42.22 times in the OM rats (Table I). In turn, the ratio between microsomal activation leading to CB in the pres- ence of NADPH was 1.659 times higher in OM than in SD rats while that in its absence was 1.039 times more intense (Table I).
Both nuclear suspensions from SD and OM rats activate CCl, to metabolites giving CB to lipids either in the presence or absence of NADPH. CB in the presence of NADPH is 6.45 times higher than in its absence for the case of SD and 9.41 times in that of OM rats (Table I). Nuclear activation in the presence of NADPH was 2.17 times more intense in OM than in SD rats, whereas in the absence of NADPH it was 1.49 higher in the former than in the latter (Table I).
The relation between microsomal to nuclear activation capacity leading to CB to lipids in the presence of NADPH is 13.29 in SD and 10.14 in OM rats. These parameters in the absence of NADPH are 3.24 and 2.26, respectively, in the case of SD and OM male rats (Table I).
Pattern of Distribution of ["C] From 14CC14 Covalently Bound In Vitro to Microsomal and Nuclear Lipids From Sprague Dawley (SD) or Osborne Mendel (OM) Rats
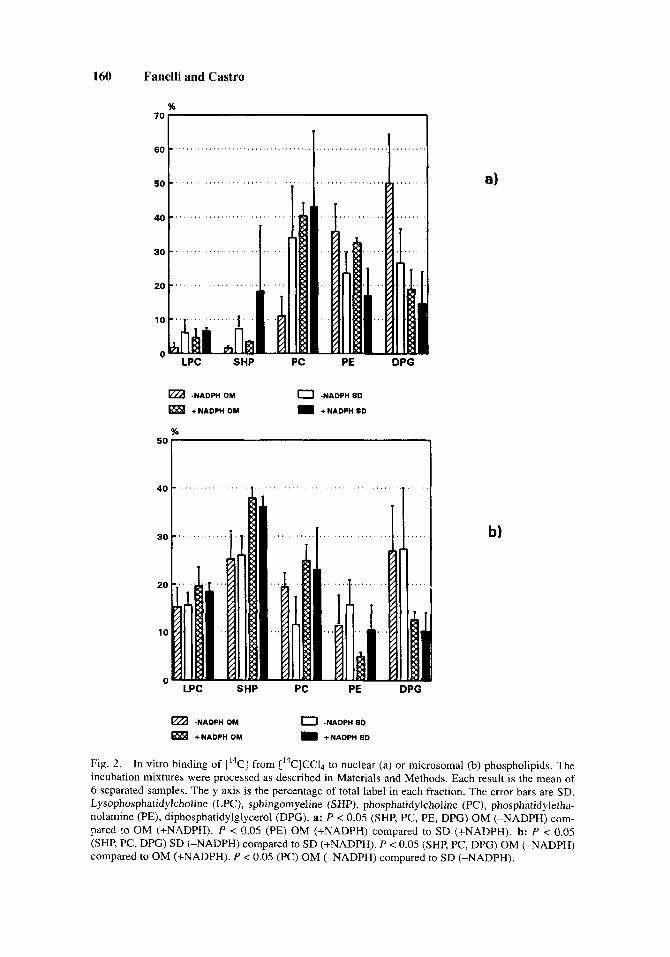
CC4-reactive metabolites produced by liver nuclear or microsomal preparations from SD or OM rats either in the presence or absence of NADPH covalently bind to all classes of lipids from both organelles (Fig. la,b). In all cases, except that of nuclear preparations from OM rats in the absence of NADPH, most of the attack by the reactive metabolites was on the phospholipid fraction. In turn, all phospholipid classes interacted with the CC1,-reactive metabolites to lead to CB (Fig. 2a,b).
In the presence of NADPH, the pattern of distribution of CB to lipid fractions changes in relation to that observed when it is absent in both microsomes and nuclei

158 Fanelli and Castro
TABLE I. Covalent Binding of "C From 14CC14 to Microsomal and Nuclear Lipids in Sprague Dawley (SD) and Osborne Mendel (OM) Rats in Vitro'
Covalent binding of I4C from ['4C]CC1, (dpm/mg lipids)
Microsomes Nuclei
- NADPH SD 453.2 f 154.4" 139.8 f 6.7h OM 471.0 f 32.6"' 208.4 f 54.1f,L
SD 11 989.3 f 1 322.9" 902.0 f 137.7d.' + NADPH
OM 19 888.8 f 1 383.1R 1961.4 f 244.Ih
'Incubation mixtures containing purified liver nuclei or microsomal fractions were processed as de- scribed in Materials and Methods. Each result is the mean f standard deviation of 6 separated samples. *SD: "P < 0.001 compared to +NADPH; bP < 0.01 compared to microsomes; ' P < 0.001 compared to nuclei; dP < 0.001 compared to -NADPH. OM: "P < 0.001 compared to + NADPH; 'P < 0.01 com- pared to microsomes; < 0.01 compared to nuclei; hP < 0.01 compared to -NADPH. Microsomes: 'P > 0.05 compared to -NADPH SD; 'P < 0.001 compared to +NADPH OM. Nuclei: kP > 0.05 compared to -NADPH SD; ' P < 0.01 compared to +NADPH OM.
and evidences a significant tendency to increase CB in phospholipids (PL), free fatty acids (FFA), and triglycerides (TG), while it dramatically decreases the CB to cho- lesterol esters (CHE) (Fig. la,b).
The CB to monoglycerides (MG) and diglycerides (DG) tends to decrease in microsomal preparations from SD and OM rats (Fig. 1 b), while that to nuclear frac- tions either slightly increases (OM rats) or remains unchanged (SD rats) (Fig. la). The CB to free cholesterol (CH) tends to decrease in the presence of NADPH for the case of microsomal preparations from both rat strains (Fig. lb), and markedly in- creases in nuclear preparations from OM rats while it remains unchanged in the case of SD rats (Fig. la).
The analysis of phospholipids showed that in the absence of NADPH the prin- cipal nuclear phospholipid targets were diphosphatidylglycerol (DPG; OM rats) and phosphatidylcholine (PC, SD rats) (Fig. 2a). When NADPH was added, the CB to DPG decreased and the CB associated with PC increased, in both rat strains (Fig. 2a).
In the case of microsomes in the absence of NADPH, most of the attack was on the DPG and sphingomyeline (SHP) fractions (from OM and SD rats; Fig. 2b). In the presence of NADPH, while the CB to DPG decrease, we observed an increase of CB to SHP, PC, and in a less extension lysophosphatidylcholine (LPC) in both rat strains (Fig. 2b).
Covalent Binding of 14C From l4CCI4 to Microsomal and Nuclear Lipids Isolated From I4CCI4-Treated SD or OM Male Rats
As reported above for the case of in vitro experiments, we could observe CB to microsomal and nuclear lipids from both SD and OM rats. In vivo, the intensity of the CB to total microsomal lipids in OM rats was significantly higher than in SD rats. The statistical analysis of data from experiments with nuclei only showed a neat but not statistically significant tendency to be so (Table 11). The CC14 reactive me- tabolites produced by nuclear or microsomal fractions covalently bind to all lipid classes in both organelles. Most of the attack was on the phospholipid fraction (Fig. 3a,b).

' O 0 I
. . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
60t...l. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . 50
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 40
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30
20
. . . . . . . . . . . . . . . . . 10
0 PL MG DG CH FFA TO CHE
" PL MG DO CH FFA TG CHE
-NADPHOM 0 -NADMSD
+NADM OM = +NADM SD
b)
-NADPHOM 0 -NADPHSD
+NADPH OM +NADPHSD
Fig. 1. In vitro binding of [14C] from ['4C]CC14 to nuclear (a) or microsomal (b) lipids. The incuba- tion mixtures were processed as described in Materials and Methods. Each result is the mean of 6 separated samples. The y axis is the percentage of total label in each fraction. The error bars are SD. Phospholipids (PL), monoglycerides (MG), diglycerides (DG), cholesterol (CH), free fatty acids (FFA), triglycerides (TG), cholesterol esters (CHE). a: P < 0.0001 (PL, CHE) SD (-NADPH) compared to SD (+NADPH). P < 0.01 (DG, TG, CHE) OM (-NADPH) compared to OM (+NADPH). P < 0.01 (PL, MG, CHE) OM (-NADPH) compared to SD (-NADPH). P < 0.05 (MG, DG, TG, CHE) OM (+NADPH) compared to SD (+NADPH). b: P < 0.01 (PL, MG, DG, CH, FFA, TG, CHE) SD (-NADPH) com- pared to SD (+NADPH). P < 0.05 (PL, DG, CH, FFA, TG, CHE) OM (-NADPH) compared to OM (-NADPH). P < 0.05 (CH, TG) OM (-NADPH) compared to SD (-NADPH). P < 0.05 (PL, CHE) OM (+NADPH) compared to SD (+NADPH).
160 Fanelli and Castro
% 70 I
, . . . . . , . . . . .
1.
SHP
EZ2 - N A D M ~ M 0 -NADPHSD
+NADMOM +NADPH OD
40
" LPC SHP PC
a -NADPHOM 0 -NADPHSD
+NADM OM w +NADM SD
Fig. 2. In vitro binding of [I4C] from [14C]CC14 to nuclear (a) or microsomal (b) phospholipids. The incubation mixtures were processed as described in Materials and Methods. Each result is the mean of 6 separated samples. The y axis is the percentage of total label in each fraction. The error bars are SD. Lysophosphatidylcholine (LPC), sphingomyeline (SHP), phosphatidylcholine (PC), phosphatidyletha- nolamine (PE), diphosphatidylglycerol (DPG). a: P < 0.05 (SHP, PC, PE, DPG) OM (-NADPH) com- pared to OM (+NADPH). P < 0.05 (PE) OM (+NADPH) compared to SD (+NADPH). b: P < 0.05 (SHP, PC, DPG) SD (-NADPH) compared to SD (+NADPH). P < 0.05 (SHP, PC, DPG) OM (-NADPH) compared to OM (+NADPH). P < 0.05 (PC) OM (-NADPH) compared to SD (-NADPH).

Covalent Binding to Lipids 161
TABLE 11. Covalent Binding of 14C From l4Ccl4 to Microsomal and Nuclear Lipids in Sprague Dawley (SD) and Osborne Mendel (OM) Rats in Vivo'
Covalent binding of I4C from ['4C]CC14 (dpm/mg lipids) Microsomes Nuclei
SD 129.9 f 18.5 126.3 k 14.0 OM 116.1 f 26.4* 198.4 f 58.1
'The samples were processed as described in Materials and Methods. Each result is the mean f stan- dard deviation of 6 separated samples. *P < 0.05 compared to microsomes SD.
The CB to triglycerydes (TG) and cholesterol esters (CHE) of the microsomal preparations of OM rats were significantly lower than SD rats (Fig. 3b). In nuclear preparations the CB to phospholipids (PL) and monoglycerydes (MG) fractions were significantly higher in OM than in SD rats (Fig. 3a).
We were able to observe a lipid fraction of unknown nature (X) between FFA and TG (Fig. 3a,b) which suffered CB of CCl,-reactive metabolites in nuclear lipids of SD rats and microsomal lipids from both rat strains. That did not happen in our corresponding experiments in vitro. The analysis of the different classes of phospho- lipids attacked by reactive metabolites show that the larger proportion of the radio- activity was associated to the diphosphatidylglycerol (DPG) fraction of liver nuclear phospholipids from both OM and SD rats (OM < SD) (Fig. 4a) and to sphingomyelin (SHP) in microsomal phospholipids (Fig. 4b). In the case of OM rats we also ob- served an unknown labeled lipid fraction (Y), between phosphatidylcholine (PC) and phosphatidylethanolamine (PE), not appearing in the equivalent in vitro experiences (Fig. 4a,b).
DISCUSSION AND CONCLUSIONS
Both liver microsomal and nuclear suspensions from SD and OM rats were able to activate C C , to reactive metabolites that bind covalently to lipids either in the absence or in the presence of NADPH. Interestingly, this CB was higher in the OM than in the SD strain of rats for the microsomal lipid but particularly for the case of nuclear lipid. This might be of relevance, for CB to lipids merely reflects the intensity of CC14 activation process in the given organelle. The more intense ability for .CC13 production in the DNA/nuclear proteins neighbourhood of the OM than in SD rats, in turn, might be also significant in relation to the very well known susceptibility of the OM rats to CC14 carcinogenic effects. In effect, DNA and nuclear proteins are believed to be the critical targets for the reaction of ultimate forms of chemical carcinogens in susceptible tissues [ 15-17]. SD rats, in contrast, have a cirrhogenic response but do not get liver cancer by exposure to CCl, [9]. As in the case of the in vitro experiments, in vivo the CB to microsomal lipids was also more intense in OM than in SD rats. In contrast, in the case of the in vivo experiments with nuclear lipids, we failed to observe equivalent significant differences despite the fact that there was a suggestive ten- dency for values in OM rats to be higher than those ir. SD animals. Failure might be attributed to a larger dispersion in the in vivo values from OM rats when compared to those in SD rats. Besides these unproven but suggestive quantita-

162 Fanelli and C a s h
% 70 1 I
I 1 6
a)
PL MG DG CH FFA X TG CHE
= O M =SD
PL MG DG CH FFA X TO CHE
Fig. 3. In vivo binding of [I4C] from ['4C]CC14 to nuclear (a) or microsomal (b) lipids. See Materials and Methods for more details. Each result is the mean of 6 separated samples. The y axis is the percentage of total label in each fraction. The error bars are SD. Phospholipids (PL), monoglycerides (MG), diglycerides (DG), cholesterol (CH), free fatty acids (FFA), triglycerides (TG), cholesterol es- ters (CHE). a: P < 0.05 (PL, MG) OM compared to SD. b: P < 0.005 (TG, CHE) OM compared to SD.

Covalent Binding to Lipids 163
LPC SHP PC Y PE DPG
m O M m S D
50m
LPC SHP PC Y PE DPG
Fig. 4. In vivo binding of [I4C] from ['4]CC14 to nuclear (a) or microsomal (b) phospholipids. See Materials and Methods for more details. Each result is the mean of 6 separated samples. The y axis is the percentage of total label in each fraction. The error bars are SD. Lysophosphatidylcholine (LPC), sphingomyeline (SHP), phosphatidylcholine (PC), phosphatidylethanolamine (PE), diphosphatidylglycerol (DPG). a: P < 0.05 (Y, DPG) OM compared to SD. b: P < 0.05 (Y, DPG) OM compared to SD.

164 Fanelli and Castro
tive differences in the abilities of both nuclear preparations to activate CCl, to reactive metabolites that covalently bind to lipids we also observed some quali- tative differences in their pattern that might have some significance.
Most of the CB of CCl,-reactive metabolites in both rat strains occurs in the phospholipid and in the (cholesterol + cholesterol esters) fractions. The relative con- tribution of each of these lipids varies, depending on the in vitro conditions and on the presence or absence of NADPH. The reason for this different behaviour might be related to the already established different CC14 enzymatic activation systems present in both microsomes and nuclei. The NADPH- and P450-dependent CC14 activation system is immersed in a phospholipid (mostly phosphatidylcholine) environment, and consequently the highly reactive ''~CC13 formed by this pathway would have more of a tendency to interact with them than with other lipids [11,12]. Conversely, the non-P450-non-NADPH CC1, activation pathway might be in closer or equivalent to that of other lipids not phospholipidic in nature. The CB to phospholipids is higher in the cirrhosis-sensitive strain (Sprague Dawley), while that to CH or to CHE is more intense in the cancer-sensitive strain (Osbome Mendel). These correlations might be of some future interest in view of the known abilities of phospholipids to regulate gene function, nucleosome structure, RNA synthesis, and activation of DNA poly- merase alpha [ 18-24]. Since nuclear lipids may be part of an intracellular signaling system modulating protein kinase C, an enzyme involved in phosphorylation of nuclear proteins [22], it is interesting that we observed CB to all nuclear lipid frac- tions in vivo and in vitro and that we now report that specific altered lipids appeared associated to nuclear lipids from the cirrhogenic susceptible strain (SD) (peak X in Fig. 3a) or with those from a carcinogenesis susceptible strain (OM) (peak Y in Fig. 4a). The structure of these altered lipid moieties remains to be established. In addi- tion, more CB to DPG was observed in the SD strain of rats. Past studies from our laboratory showed that in the case of the microsomal lipids CB is mainly to PUFA and that as result of that interaction, their physico-chemical properties became dra- matically altered [25]. If similar or equivalent reactions occurred at the nuclear mem- brane, their properties might also be altered. There is no reason to envisage a different behaviour in this respect between nuclear and endoplasmic reticulum membranes, for the former derive from the latter [26]. This in turn might be of some significance to carcinogenic effects of CCI4 for the case of OM rats. In effect, it has been recently reported that the composition of polyunsaturated fatty acids (PUFA) in nuclear phospholipids may modulate cell proliferation in mam- malian cells [27]. Concerning the interactions evidenced here with cholesterol and its esters, it is interesting that recent preliminary studies from our laboratory [28] revealed that cholesterol moieties are oxidized during .CC13/CC1302. attack, to give keto derivatives potentially having structures resembling phorbol or ac- tive hormones [29]. This might also be of significance in light of the fact that present studies evidenced that cholesterol was preferentially attacked by CC1,- derived reactive metabolites (.CCl3) and potentially to CC1302. as well. In effect, i t is known that cholesterol is important to the processes of cell growth and that it directly or indirectly plays a role in gene expression [30,31].
All these preliminary observations require further studies to learn whether they might be considered of additional interest. Some of them are at present being ana- lyzed in our laboratory.

Covalent Binding to Lipids 165
REFERENCES 1. Castro GD, Diaz G6mez MI, Castro JA: Species differences in the interaction between CC14 reac-
tive metabolites and liver DNA and nuclear protein fractions. Carcinogenesis 10:289-294, 1989. 2. Castro GD, Diaz G6mez MI, Castro JA: Dimethyldisulfide formation during trichloromethyl radi-
cal attack on methionine. Biochem Pharmacol 38:41454147, 1989. 3. Castro GD, Diaz G6mez MI, Castro JA: Biotransformation of carbon tetrachloride and lipid
peroxidation promotion by liver nuclear preparations from different animal species. Cancer Lett
4. Castro GD, Diaz G6mez MI, Castro JA: Interaction of trichloromethyl free radicals with phenyla- lanine. Arch Toxicol 65:340-343, 1991.
5. Diaz G6mez MI, Castro JA: Nuclear activation of carbon tetrachloride and chloroform. Res Commun Chem Path01 Pharmacol27: 191-194, 1980.
6. Diaz G6mez MI, Castro JA: Covalent binding of carbon tetrachloride to liver nuclear DNA, pro- teins and lipids. Toxicol Appl Pharmacol 56: 199-206, 1980.
7. Diaz G6mez MI, Castro JA: Reaction of trichloromethyl free radicals with deoxyribonucleic acid bases. Res Commun Chem Pathol Pharmacol 32:147-153, 1981.
8. Villarruel MC, Castro JA: Reactions of trichloromethyl free radicals with aminoacids. Res Commun Chem Pathol Pharmacol28:79-85, 1980.
9. Reuber MD, Glover EL: Cirrhosis and carcinoma of the liver in male rats given subcutaneous carbon tetrachloride. J Natl Cancer Inst 44:419-427, 1970.
10. Fanelli SL, Castro JA: Carbon tetrachloride promoted malondialdehyde formation in liver mi- crosomal and nuclear preparations from Sprague Dawley or Osbome Mendel male rats. Res Commun Chem Pathol Pharmacol 82:233-236, 1993.
11. Villanuel MC, Castro JA: Carbon tetrachloride target lipids in rat liver microsomes. Effect of cystamine administration on their pattern of labeling by CC14. Biochem Biophys Res Commun
12. Villarruel MC, Diaz G6mez MI, Castro JA: The nature of the in vitro irreversible binding of
13. Gad SC, Weil CS: Statistics for toxicologist. In Hayes AW “Principles and Methods of Toxicol-
14. Graph Pad Software: “Graph Pad Instat. Instant Biostatistics.” San Diego: Graph Pad Software,
15. Harris CC: Chemical and physical carcinogenesis: advances and perspectives for the 1990s. Can-
16. Miller JA: Brief history of chemical carcinogenesis. Cancer Lett 83:9-14, 1994. 17. Pitot H: The molecular biology of carcinogenesis. Cancer 72:962-970, 1993. 18. Chapkin RS, Davidson LD, Davidson LA: Phospholipid molecular species composition of mouse
liver nuclei. Biochem J 287:237-240, 1992. 19. Cocco L, Gilmour RS, Ognibene A, Letcher AJ, Manzoli FA, Irvine RF: Synthesis of poly-
phosphoinositides in nuclei of friend cells. Biochem J 248:765-770, 1987. 20. Herlan G, Giese G, Wanderlick F Influence of nuclear membrane lipid fluidity on nuclear RNA
release. Exp Cell Res 118:305-309, 1979. 21. Manzoli FA, Martelli AM, Capiteni S , Maraldi NM, Rizzoli R, Bamabi 0, Cocco L: Nuclear
polyphosphoinositides during cell growth and differentiation. Adv Enzyme Regul28:25-34, 1989. 22. Sylvia VL, Joe CO, Norman JO, Curtin GM, Busbee DL: Phosphatidylinositol-dependent activa-
tion of DNA polymerase alpha. Biochem Biophys Res Commun 135:880-885, 1986. 23. Venkataman JT, Lefibure YA, Clandinin MT: Diet fat alters the structure and function of the nuclear
envelope: modulation of membrane fatty acid composition, NTPAse activity and binding of triiodothyrosine. Biochem Biophys Res Commun 135:655-661, 1986.
24. Venkatraman JT, Clandinin M T Ribonucleic acid efflux from isolated mouse liver nuclei is al- tered by diet and genotypically determined change in nuclear envelope composition. Biochim Biophys Acta 940:33-42, 1988.
25. Villarruel MC, Castro JA: Irreversible binding of carbon tetrachloride to microsomal phospholip- ids. Free radical nature of the reactive species and alterations in the physico-chemical properties of the target fatty acid. Res Commun Chem Pathol Pharmacol 10:105-116, 1975.
26. De Robertis ED, De Robertis EM: “Cell and molecular biology.” PhiIadelphia: Lea and Febiger,
53~9-15, 1990.
54:108-115, 1973.
carbon tetrachloride to microsomal lipids. Toxicol Appl Pharmacol 33: 106-1 14, 1975.
ogy.” New York: Raven Press, 1982, pp 273-320.
1992, pp 1-127.
cer Res (Suppl) 5 1 :5023-5044, 1991.
1987, pp 358-370.

166 Fanelli and Castro
27. Isihara H, Tamiya-Koizumi K, Kuriki H, Yoshida S, Kojima K: Growth-associated changes in fatty acid compositions of nuclear phospholipids of liver cells. Biochim Biophys Acta 1084:53-59, 1991.
28. Fanelli SL, Castro GD, Castro JA: Cholesterol interaction with free radicals produced from carbon tetrachloride or bromotrichloromethane by either catalytic decomposition or via liver microsomal activation. Chem Biol Interact, 1995 (in press).
29. Endo Y, Fukasawa H, Hashimoto Y, Shudo K: Synthesis of oxygenated cholesterols as structural mimics of phorbol ester-type tumor promoters. Chem Pharm Bull (Tokyo) 42:462469, 1994.
30. Eriksson LC, Anderson GN: Membrane biochemistry and chemical hepatocarcinogenesis. Crit Rev Biochem Mol Biol 27:l-55, 1992.
31. Yeagle PL: Cholesterol and the cell membrane. Biochim Biophys Acta 822:267-287, 1985.




















