
Cholesterol interaction with free radicals produced from carbon tetrachloride or...
14
ELSEVIER Chemico-BiologicalInteractions 98 (1995) 223-236 Cholesterol interaction with free radicals produced from carbon tetrachloride or bromotrichloromethane by either catalytic decomposition or via liver microsomal activation S.L. Fanelli, G.D. Castro, J.A. Castro* Centro de Investigaciones Toxicolbgicas (CEITOX) CITEFA/CONICET. Zufriategui 4380 (1603) Villa Martelli, Pcia. de Buenos Aires, Argentina Received 27 March 1995;revision received 12 June 1995;accepted 13 June 1995 Abstract The reaction between cholesterol (Ch) and trichloromethyl or trichloromethyl peroxyl radi- cals was studied. The latter were generated from Ccl, either by benzoyl peroxide (BP) catalysis or via thermal activation or by liver microsomal NADPH-dependent biotransforma- tion of CBrC13. The structure of the products formed was elucidated by gas chromatography-mass spectrometry (GC/MS). Under aerobic conditions and using thermal activation of Ccl,, the formation of 6 products was observed. Two (I and II) were dehydrated Ch derivatives (one also having a third double bond) (I). Another product was a As-3 ketone derivative of Ch (III). Two additional reaction products were determined as ketocholesterols (IV and V). One chloro Ch was also formed (VI). At low concentrations of BP, reaction was more extensive than under thermal activation, and the formation of peaks I to IV was also observed. When the reaction was conducted anaerobically and using thermal activation of Ccl, to generate radicals, only products I and II were formed in low yield. Under anaerobic conditions, but using catalyst, compounds I and III were produced plus two new isomeric ketocholesterol derivatives (VIII and IX) and also a compound having an extra hydroxyl group on the Ch structure (X). In order to check whether similar reactions are obser- vable under biological experimental conditions, we used activation of CBrCl, by liver micro- somes. The incubation using only microsomes (without CBrCl, or NADPH) showed two ketocholesterol peaks (A and B). In the presence of CBrCl, we could detect peak B and l Corresponding author, Tel.: 541 761 0031 Ext. 239; Fax: 541 761 591 I. E-mail: [email protected]. 0009-2797/95/Y&9.50 0 1995 Elsevier Science Ireland Ltd. All rights reserved SSDI 0009-2797(95)03648-6
-
Upload
conicet-ar -
Category
Documents
-
view
3 -
download
0
Transcript of Cholesterol interaction with free radicals produced from carbon tetrachloride or...

ELSEVIER Chemico-Biological Interactions 98 (1995) 223-236
Cholesterol interaction with free radicals produced from carbon tetrachloride or
bromotrichloromethane by either catalytic decomposition or via liver microsomal activation
S.L. Fanelli, G.D. Castro, J.A. Castro*
Centro de Investigaciones Toxicolbgicas (CEITOX) CITEFA/CONICET. Zufriategui 4380 (1603) Villa Martelli, Pcia. de Buenos Aires, Argentina
Received 27 March 1995; revision received 12 June 1995; accepted 13 June 1995
Abstract
The reaction between cholesterol (Ch) and trichloromethyl or trichloromethyl peroxyl radi- cals was studied. The latter were generated from Ccl, either by benzoyl peroxide (BP) catalysis or via thermal activation or by liver microsomal NADPH-dependent biotransforma- tion of CBrC13. The structure of the products formed was elucidated by gas chromatography-mass spectrometry (GC/MS). Under aerobic conditions and using thermal activation of Ccl,, the formation of 6 products was observed. Two (I and II) were dehydrated Ch derivatives (one also having a third double bond) (I). Another product was a As-3 ketone derivative of Ch (III). Two additional reaction products were determined as ketocholesterols (IV and V). One chloro Ch was also formed (VI). At low concentrations of BP, reaction was more extensive than under thermal activation, and the formation of peaks I to IV was also observed. When the reaction was conducted anaerobically and using thermal activation of Ccl, to generate radicals, only products I and II were formed in low yield. Under anaerobic conditions, but using catalyst, compounds I and III were produced plus two new isomeric ketocholesterol derivatives (VIII and IX) and also a compound having an extra hydroxyl group on the Ch structure (X). In order to check whether similar reactions are obser- vable under biological experimental conditions, we used activation of CBrCl, by liver micro- somes. The incubation using only microsomes (without CBrCl, or NADPH) showed two ketocholesterol peaks (A and B). In the presence of CBrCl, we could detect peak B and
l Corresponding author, Tel.: 541 761 0031 Ext. 239; Fax: 541 761 591 I. E-mail: [email protected].
0009-2797/95/Y&9.50 0 1995 Elsevier Science Ireland Ltd. All rights reserved SSDI 0009-2797(95)03648-6

224 S. L. Fanelli er al. / Chemico-Biological Interaclions 98 (1995) 223-236
hydroxycholesterol (C) and two others, ketocholesterols (D and E). D was the only peak showing close similarity (spectrum and retention time) to one of those observed in the chemi- cal reaction system (V). The reaction of CBrCl, in the presence of NADPH showed peaks B, C, D and E, in low abundance and a 7-ketocholesterol (F). If some of the reaction products reported here were formed during the intoxication with these haloalkanes, significant biologi- cal consequences might be expected.
Keywork Cholesterol; Free radicals; Carbon tetrachloride; Bromotrichloromethane; Mass spectrometry
1. Introduction
It is very well established that Ccl4 hepatotoxicity is related to its biotransforma-
tion in the liver to ’ Ccl, and CClsO,, * free radicals (l-41.
These radicals can then interact with cellular components including DNA, pro- teins, lipids and heme via addition, abstraction and other reactions to initiate
deleterious processes ending in liver necrosis, fatty liver, cirrhosis and cancer as well as many manifestations of cell injury [l-4]. There is limited information, however, about the nature of the chemical interactions between the molecules from the dif-
ferent cellular components and the ’ Ccl, or the CClsOz ’ free radicals. Reactions of these radicals with lipids have been by far better studied [2,5- 131, although, most detailed structural studies concerned the polyunsaturated fatty acids (PUFA) from
phospholipids [ 12,131. Studies by others and by our laboratory, however, provided evidence that cholesterol, either free or esterilied, might be significant targets for Ccl,, derived free radical attack [5,7,9,12]. Those studies were not accompanied by identification of the reaction products formed. That information, however, is of
relevance because cholesterol is a critical component of biological membranes and a precursor of major biologically active steroids [ 14-171. Consequently, its altera- tion via the attack by reactive metabolites of CCL might profoundly perturb not only its structure but also its function. In this work we describe our initial attempt
to shed some light into the chemistry of the interaction between cholesterol and the ’ Ccl3 and CClsOz’ free radicals, when the latter are either thermally or
catalytically generated in a chemical model system or when they are enzymatically produced in liver microsomal incubation mixtures.
2. Materials and methods
2.1. Chemicals
Ccl, was purchased from Mallinkrodt and CBrCls from Aldrich and both were distilled before use. The original commercial CBrCls contained several additives obscuring the chromatographic analysis with mass spectrometric identification (GC/MS). Mass spectrometric analysis of the freshly distilled Ccl, or CBrCls did
not reveal the presence of any of the products reported or any other impurity. Benzo- yl peroxide, BSTFA (bis(trimethylsilyl)trifluoro-acetamide) and cholesterol were

S.L. Fanelli et al. / Chemico-Biological Interactions 98 (1995) 223-236 225
purchased from Sigma Chemical Co. Benzoyl peroxide contains about 30% water as stabilizer. GLYME (1,2-dimethoxy ethane) was from Merck, Germany.
2.2. Procedures Chemical reaction. The study of the in vitro chemical interaction of Ccl, derived
free radicals with cholesterol was carried out as follows: 0.6 mg of cholesterol in 0.5 ml of Ccl, was heated at 85°C for 5 h in the absence or the presence of benzoyl per- oxide (1 mg or 0.1 mg as indicated) in an ampoule. The reaction mixture and am- poule were purged with either nitrogen or air in each experiment before sealing. At the end of this period, the solvent was evaporated under nitrogen. The residue was then silylated with a mixture of BSTFA:acetonitrile (I: l), 0.2 ml at 70°C for 15 min. Blanks without benzoyl peroxide or without cholesterol were run simultaneously.
It is not possible to run blanks without Ccl,, because it is the only solvent in the reaction mixture. Further, it is not possible to run the reaction in solvents other than CC& (e.g. methanol, 1,2-dimethoxy ethane, acetone and others) because they themselves generate free radicals which obscure product identification by production of many artifacts.
Microsomal incubations. Non-inbred male Sprague-Dawley rats (240-260 g) were used. Animals were fasted for 12-14 h before sacrifice. Microsomal fractions were obtained as previously described [22,23]. Incubation mixtures containing micro- somal suspensions (6.2-6.8 mg protein/ml), NADPH generating system [23] and 7 ~1 of CBrC13 in 0.15 M KH2P04 (pH 7.4) 5 mM MgC12 (final volume 3 ml) were conducted under aerobic atmosphere for 1 h at 37°C in aluminium-sealed-neoprene septum-stoppered glass vials. CBrC13 was chosen for study because of its ready me- tabolism to form ’ CCls radicals [24]. The reaction was interrupted by putting the incubation flasks on cold ice. The incubation mixtures were extracted with 12 ml of acetone, filtered and evaporated under nitrogen. Blanks without CBrCl, or without generating system were performed simultaneously.
2.3. Chromatographic analysis The evaporated extract was derivatized for chromatographic analysis as described
above. The gas chromatographic/mass spectrometric identifications of reaction products
were performed on a Hewlett Packard Model 5970 B mass selective detector interfac- ed to a HP 5890 gas chromatograph. Chromatographic conditions were as follows: injection port temperature, 265°C in the splitless injection mode; column, fused silica capillary column (12.5 m x 0.20 i.d.) crosslinked with 5% phenylmethylsilicone 0.33 pm film thickness (Hewlett Packard, Palo Alto, CA); carrier gas, helium (column head pressure, 100 kPa); temperature program, 70°C for 2 min and then increased to 140°C at a ramp velocity of 70Wmin and then to 280°C at a ramp velocity of 10Wmin.
The GUMS interface temperature was 280°C and ion source was - 200°C. Spec- tra were obtained at 70 eV ionizing energy and the mass range scanned was loo-670 a.m.u., at 0.75 s/scan.
It is important to emphasize that we have to rely entirely on MS for identification
226 S. L. Fanelli et al. / Chemico-Biological Interactions 98 (1995) 223-236
because the very low product yield from the reaction mixture, under the experimen- tal conditions employed, prevented the use of additional NMR methods to further elucidate the structures.
3. Results
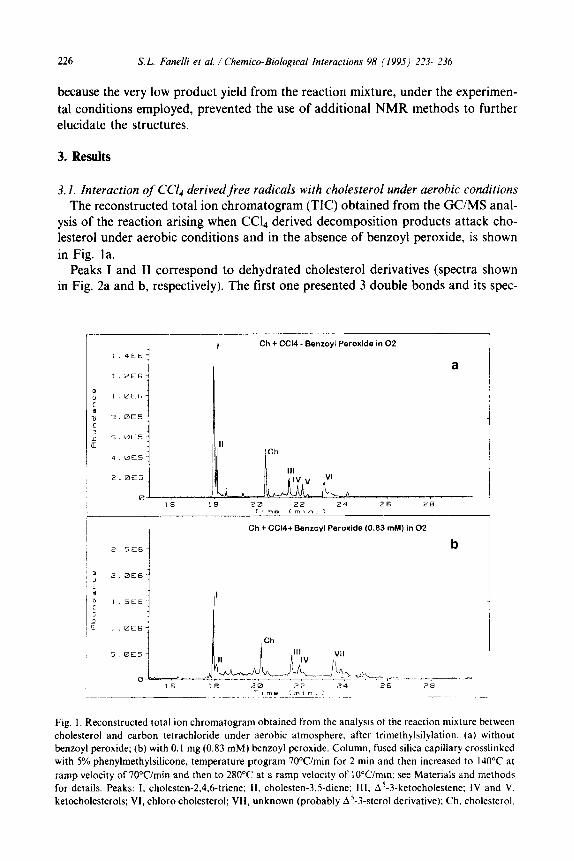
3.1. Interaction of Ccl4 derivedfree radicals with cholesterol under aerobic conditions The reconstructed total ion chromatogram (TIC) obtained from the GUMS anal-
ysis of the reaction arising when CCL derived decomposition products attack cho- lesterol under aerobic conditions and in the absence of benzoyl peroxide, is shown in Fig. la.
Peaks I and II correspond to dehydrated cholesterol derivatives (spectra shown in Fig. 2a and b, respectively). The first one presented 3 double bonds and its spec-
r I Ch + CC14 - Benroyl Peroxide in 02
a
I Ch
III
AlV,V hV’
I 4 Ch + CCl4+ Benzovl Peroxide (0.83 mM) in 02
Ch
III
d ,lV
VII
Ii
Fig. I. Reconstructed total ion chromatogram obtained from the analysis of the reaction mixture between
cholesterol and carbon tetrachloride under aerobic atmosphere. after trimethylsilylation: (a) without
benzoyl peroxide; (b) with 0.1 mg (0.83 mM) benzoyl peroxide. Column, fused silica capillary crosslinked with 5% phenylmethylsilicone, temperature program 7O”Cimin for 2 min and then increased to 140°C at
ramp velocity of 70Wmin and then to 28O“C at a ramp velocity of IOWmin; see Materials and methods
for details. Peaks: I, cholesten-2,4.6-triene; II, cholesten-3.5-diene; III, A’-3-ketocholestene; IV and V,
ketocholesterols; VI, chloro cholesterol; VII, unknown (probably A’-3-sterol derivative); Ch, cholesterol.

XL. Fanelli et al. / Chemico-Biological Interactions 98 (1995) 223-236 221
t
Scan 667 (17.970 min) of peak I in Fig la
SUBTRRCTED SCRLELJ
, I
366
a
200 250 300 350 MISG/ChlPQO
Scan 673 (16.125 min) of peak II in Fig la
SUBTRRCTEO SCRLEO I 366
b
J
,
1 I 1
Fig. 2. (a) Mass spectrum obtained for peak I Fig. la. M+ is observed at m/z 366, confirmed by (M-IS)+
at m/z 351. (b) Mass spectrum obtained for peak II in Fig. la. M+ is observed at m/z 368. confirmed by
(M-15)+ at m/z 353.
9000
7000
,” 6000
; u 5000
:
? 4000
3000
24
Scan 604 (21.451
SUBI-RRC
lin) of peak Ill in Fig la
;m SCRLED
280
239
; 3f7
200 250 300 350 .1 .^I
369 \
304 /
Fig. 3. Mass spectrum obtained for peak III Fig. la. M+ and (M-15)+ are observed at ml: 384 and 369,
respectively. Loss of ketene from M+ leads to m/z 342. Other fragments are m/z 327 (loss of ketene from
(M-15)). m/z 260 (M-124) and m/z 124 (characteristic of A4-3-ketones). The base peak at m/z 229 cor-
responds to loss of 155 a.m.u. from M+, possibly a loss of the lateral chain plus D ring (M-CsH,,-42).
228 S. L. Faneili et al. / Chemico-Biological Interactions 98 (1995) 223-236
Scan 820 (21.859 min) of peak IV in Fig la
SUBTRRtTED SCRLED z’47
3
a
368 ‘1 I
‘.
1.1 Ii -- ._ 200 300 4GGd .1 1
Scan 829 (22.088 min) of peak V m Fig la h
k \
385
b
253 /
,’
/ 1 474
329 \ \
i
459,
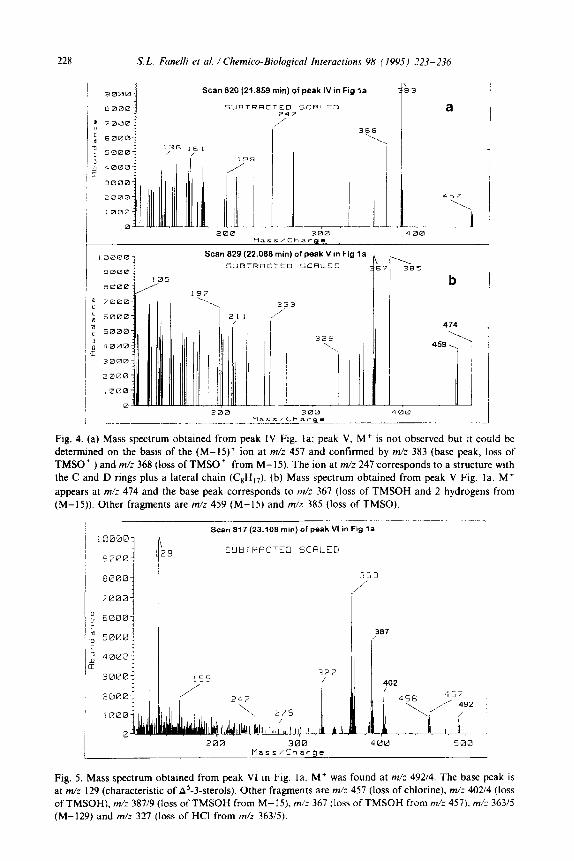
Fig. 4. (a) Mass spectrum obtained from peak IV Fig. la; peak V, M+ is not observed but it could be
determined on the basis of the (M-IS)+ ion at m/z 457 and confirmed by m/: 383 (base peak, loss of TMSO ’ ) and m/z 368 (loss of TMSO ’ from M-15). The ion at m/z 247 corresponds to a structure with the C and D rings plus a lateral chain (CsH,,). (b) Mass spectrum obtained from peak V Fig. la. M+
appears at m/z 474 and the base peak corresponds to m/z 367 (loss of TMSOH and 2 hydrogens from (M-15)). Other fragments are m/z 459 (M-15) and m/z 385 (loss of TMSO).
Scan 817 (23.108 min) of peak VI in Fig la
2 I 3 SIJBTEACTCD SCRLED
! 363
/’
327 30001 155
400 500
Fig. 5. Mass spectrum obtained from peak VI tn big. la. M+ was found at m/z 49214. The base peak is at m/z 129 (characteristic of A5-3-sterols). Other fragments are m/: 457 (loss of chlorine), m/z 40214 (loss of TMSOH), m/z 387/9 (loss of TMSOH from M-15). m/z 367 (loss of TMSOH from m/z 457). m/z 363/5 (M-129) and m/z 327 (loss of HCI from mh 36315).
S. L. Fanelli et al. / Chemieo-Biological fnieractions 98 ( 199s) 223-236 229
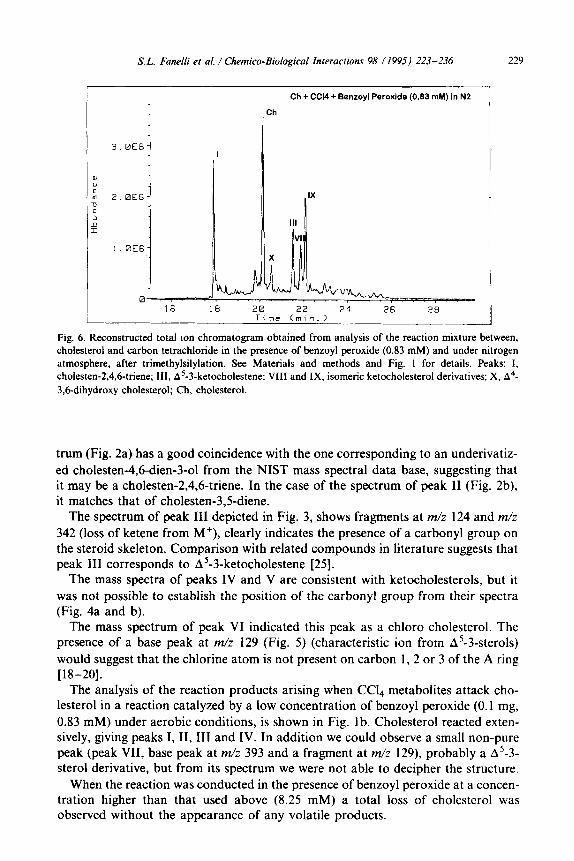
Ch + CC14 + Benzoyl Peroxide (0.83 mM) in N2
0 16 18 20 22 24 26 28
Time (min.) .__ L
Fig. 6. Reconstructed total ion chromatogram obtained from analysis of the reaction mixture between, cholesterol and carbon tetrachloride in the presence of benzoyl peroxide (0.83 mM) and under nitrogen
atmosphere, after trimethylsilylation. See Materials and methods and Fig. 1 for details. Peaks: I,
cholesten-2,4,6-triene; III, A’-3-ketocholestene; VIII and IX, isomeric ketocholesterol derivatives; X, A4-
3,6-dihydroxy cholesterol; Ch, cholesterol.
trum (Fig. 2a) has a good coincidence with the one corresponding to an underivatiz-
ed cholesten-4,6-dien-3-01 from the NIST mass spectral data base, suggesting that it may be a cholesten-2,4,6-triene. In the case of the spectrum of peak II (Fig. 2b), it matches that of cholesten-3,5-diene.
The spectrum of peak III depicted in Fig. 3, shows fragments at m/z 124 and m/z
342 (loss of ketene from M+), clearly indicates the presence of a carbonyl group on the steroid skeleton. Comparison with related compounds in literature suggests that peak III corresponds to A5-3-ketocholestene [25].
The mass spectra of peaks IV and V are consistent with ketocholesterols, but it
was not possible to establish the position of the carbonyl group from their spectra (Fig. 4a and b).
The mass spectrum of peak VI indicated this peak as a chloro cholesterol. The presence of a base peak at m/z 129 (Fig. 5) (characteristic ion from A5-3-sterols)
would suggest that the chlorine atom is not present on carbon 1, 2 or 3 of the A ring [l&20].
The analysis of the reaction products arising when Ccl4 metabolites attack cho- lesterol in a reaction catalyzed by a low concentration of benzoyl peroxide (0.1 mg,
0.83 mM) under aerobic conditions, is shown in Fig. lb. Cholesterol reacted exten- sively, giving peaks I, II, III and IV. In addition we could observe a small non-pure peak (peak VII, base peak at m/z 393 and a fragment at m/z 129), probably a A5-3- sterol derivative, but from its spectrum we were not able to decipher the structure.
When the reaction was conducted in the presence of benzoyl peroxide at a concen- tration higher than that used above (8.25 mM) a total loss of cholesterol was observed without the appearance of any volatile products.
230 XL. Faneili et al. / Chemieo-Biological Interactions 98 (1995) 223-236
3.2. Interaction of CC?4 derived free radicals with cholesterol under anaerobic con- ditions
When the previously described reaction was performed under nitrogen, a different pattern of reaction products was obtained as revealed by capillary GUMS analysis. In all cases the reaction appeared to be less extensive than under an aerobic atmosphere.
When peroxide was omitted from the reaction mixture, only the two dehydrated cholesterols were observed as in the case of the reaction proceeding under air (peaks I and II). They were produced, however, in lower abundance.
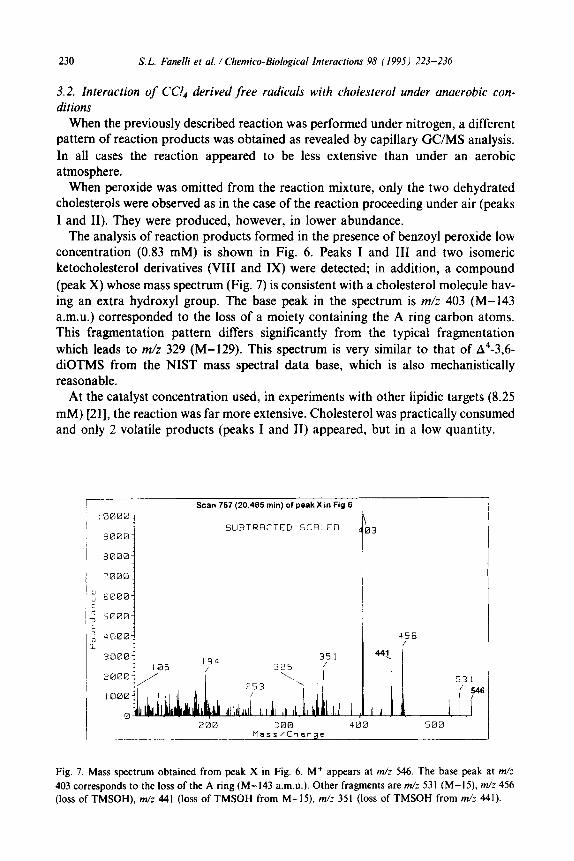
The analysis of reaction products formed in the presence of benzoyl peroxide low concentration (0.83 mM) is shown in Fig. 6. Peaks I and III and two isomeric ketocholesterol derivatives (VIII and IX) were detected; in addition, a compound (peak X) whose mass spectrum (Fig. 7) is consistent with a cholesterol molecule hav- ing an extra hydroxyl group. The base peak in the spectrum is m/z 403 (M-143 a.m.u.) corresponded to the loss of a moiety containing the A ring carbon atoms. This fragmentation pattern differs significantly from the typical fragmentation which leads to m/z 329 (M-129). This spectrum is very similar to that of A4-3,6- diOTMS from the NIST mass spectral data base, which is also mechanistically reasonable.
At the catalyst concentration used, in experiments with other lipidic targets (8.25 mM) [21], the reaction was far more extensive. Cholesterol was practically consumed and only 2 volatile products (peaks I and II) appeared, but in a low quantity.
Scan 757 (20.495 min) of peak X in Fig 6
: 0000
3 9000 j
SUBTRRCTED SCRLED
J
8000:
'0001
I 74”
I.1 I ! ll 1 . .
300 400 500 Mass/Charge
Fig. 7. Mass spectrum obtained from peak X in Fig. 6. M+ appears at m/z 546. The base peak at m/z
403 corresponds to the loss of the A ring (M-143 a.m.&). Other fragments are m/z 531 (M-15). m/z 456
(loss of TMSOH), m/z 441 (loss of TMSOH from M-15). m/z 351 (loss of TMSOH from m/z 441).
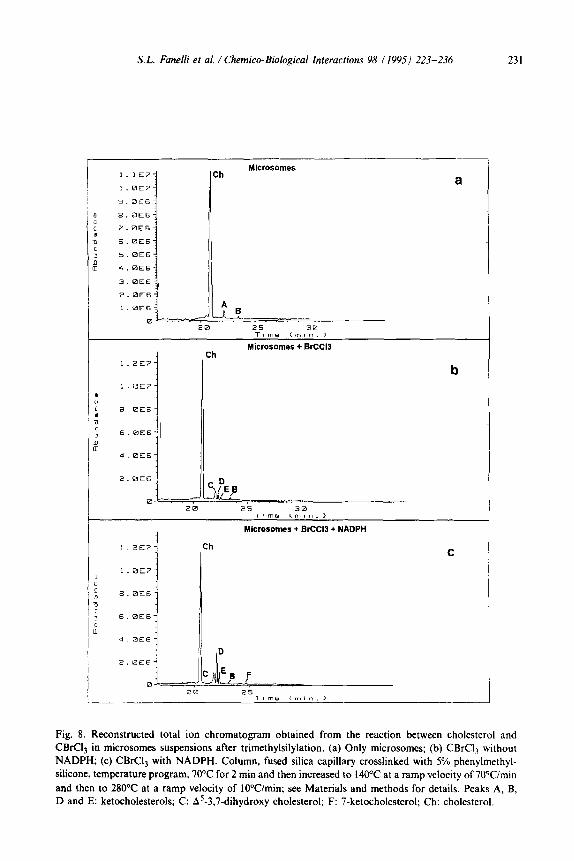
S.L. Fanelli et al. / Chemico-Biological Interactions 98 1199.5) 223-236 231
Mlcrosomes Ch a
2.OE6
-1. D
=, I,’ ,B
0 - 20 25 JB
I )rne cm?“. >
Microsotnes + BrCCl3 + NADPH
1.2YE>- Ch C
1.0E7-
Fig. 8. Reconstructed total ion chromatogram obtained from the reaction between cholesterol and
CBrCI3 in microsomes suspensions after trimethylsilylation. (a) Only microsomes; (b) CBrC13 without
NADPH; (c) CBrCl, with NADPH. Column, fused silica capillary crosslinked with 5% phenylmethyl-
silicone, temperature program, 70°C for 2 min and then increased to 140°C at a ramp velocity of 70Wmin
and then to 280°C at a ramp velocity of lOWmitt; see Materials and methods for details. Peaks A, B,
D and E: ketocholesterols; C: A5-3,7-dihydroxy cholesterol; F: 7-ketocholesterol; Ch: cholesterol.
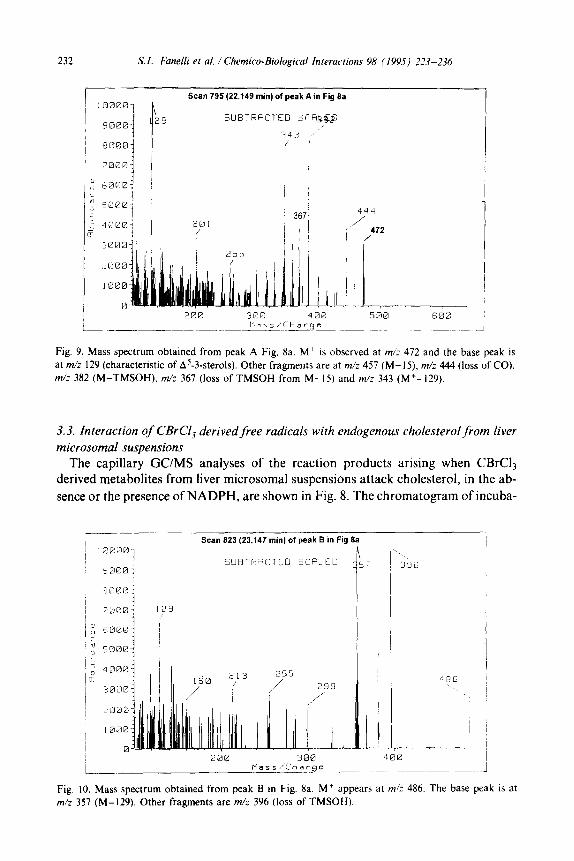
232 S. L. Fanelli er al. / Chemico-Biological Interactions 98 (1995) 223-236
Scan 795 (22.149 min) of peak A in Fig 8a
i I
! !
367, 444
201 / (,( j /’ 472
i /
Fig. 9. Mass spectrum obtained from peak A Fig. 8a. M+ is observed at m/z 472 and the base peak is
at m/z 129 (characteristic of A5-3.sterols). Other fragments are at m/: 457 (M-IS), m/z 444 (loss of CO).
m/z 382 (M-TMSOH), m/z 367 (loss of TMSOH from M-15) and m/z 343 (M+-129).
3.3. Interaction of CBrCI, derivedfree radicals with endogenous cholesterol from liver microsomal suspensions
The capillary GUMS analyses of the reaction products arising when CBrQ derived metabolites from liver microsomal suspensions attack cholesterol, in the ab- sence or the presence of NADPH, are shown in Fig. 8. The chromatogram of incuba-
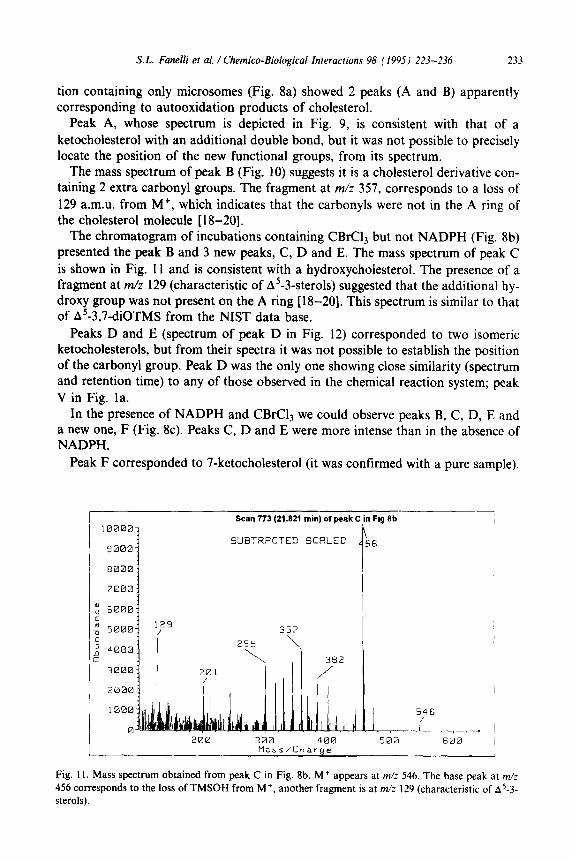
I Scan 823 (23.147 min) of peak B in Fig 8a
sa00! EjUEl~PCiCTED SCFI- CL
1 Mass/Charge .J
Fig. 10. Mass spectrum obtained from peak B in Fig. 8a. M+ appears at m/z 486. The base peak is at
m/z 357 (M-129). Other fragments are m/z 396 (loss of TMSOH).
XL. Fanelli et al. / Chemico-Biological Interactions 98 (1995) 223-236 233
tion containing only microsomes (Fig. 8a) showed 2 peaks (A and B) apparently corresponding to autooxidation products of cholesterol.
Peak A, whose spectrum is depicted in Fig. 9, is consistent with that of a ketocholesterol with an additional double bond, but it was not possible to precisely locate the position of the new functional groups, from its spectrum.
The mass spectrum of peak B (Fig. 10) suggests it is a cholesterol derivative con- taining 2 extra carbonyl groups. The fragment at m/z 357, corresponds to a loss of 129 a.m.u. from M+, which indicates that the carbonyls were not in the A ring of the cholesterol molecule [ 18-201.
The chromatogram of incubations containing CBrC13 but not NADPH (Fig. 8b) presented the peak B and 3 new peaks, C, D and E. The mass spectrum of peak C is shown in Fig. 11 and is consistent with a hydroxycholesterol. The presence of a fragment at m/z 129 (characteristic of A5-3-sterols) suggested that the additional hy- droxy group was not present on the A ring [ 18-201. This spectrum is similar to that of A5-3,7-diOTMS from the NIST data base.
Peaks D and E (spectrum of peak D in Fig. 12) corresponded to two isomeric ketocholesterols, but from their spectra it was not possible to establish the position of the carbonyl group. Peak D was the only one showing close similarity (spectrum and retention time) to any of those observed in the chemical reaction system; peak V in Fig. la.
In the presence of NADPH and CBrC13 we could observe peaks B, C, D, E and a new one, F (Fig. 8~). Peaks C, D and E were more intense than in the absence of NADPH.
Peak F corresponded to 7-ketocholesterol (it was confirmed with a pure sample).
100001
9000:
6000:
7000:
“, 6000 ii u 5000: c
’ 4000: f
3000:
Scan 773 (21.1121 min) of peak ( Fig ilb
382
/
300 400 Mass/Charge L
Fig. II. Mass spectrum obtained from peak C in Fig. 8b. M+ appears at m/z 546. The base peak at m/z
456 corresponds to the loss of TMSOH from M+, another fragment is at m/z 129 (characteristic of AS-3- sterols).
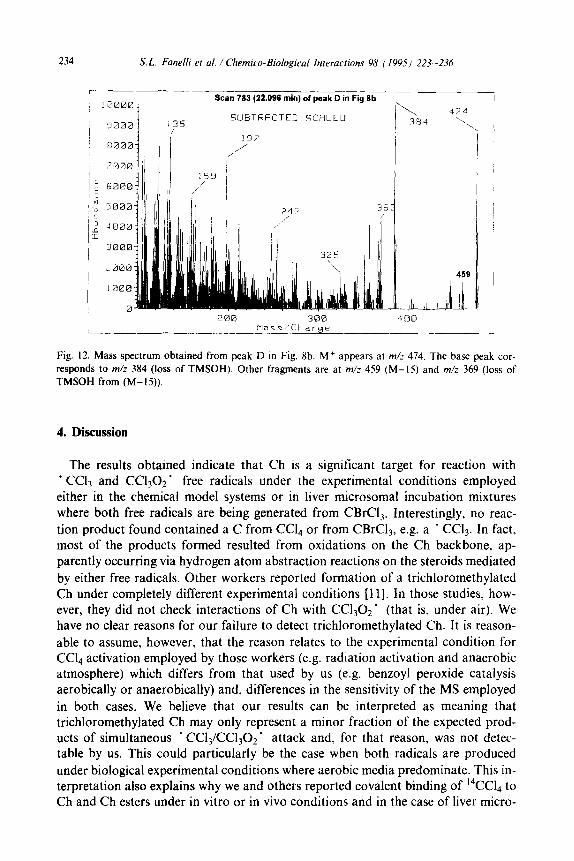
234 S. L. Fanelli et al. / Chemico-Biological Interactions 98 ( 199.5) 223-236
r : 0Q00_?
Scan 783 (22.098 min) of peak D in Fig 8b
SUBTRACTED SCRLEO ( 9000 135 ,
7000
j: 6000
‘; 5000 i
‘2 4000
IE 3000
..a00
I I000 0 250 300 400
L_.^ Mass/Charge I_
Fig. 12. Mass spectrum obtained from peak D in Fig. 8b. M+ appears at m/z 474. The base peak cor- responds to m/z 384 (loss of TMSOH). Other fragments are at m/z 459 (M-15) and m/z 369 (loss of TMSOH from (M-15)).
4. Discussion
The results obtained indicate that Ch is a significant target for reaction with ’ Ccl3 and Ccl,02 ’ free radicals under the experimental conditions employed either in the chemical model systems or in liver microsomal incubation mixtures
where both free radicals are being generated from CBrCl,. Interestingly, no reac-
tion product found contained a C from CCL, or from CBrCls, e.g. a . CCl3. In fact, most of the products formed resulted from oxidations on the Ch backbone, ap- parently occurring via hydrogen atom abstraction reactions on the steroids mediated
by either free radicals. Other workers reported formation of a trichloromethylated Ch under completely different experimental conditions [1 11. In those studies, how- ever, they did not check interactions of Ch with CC1302’ (that is, under air). We have no clear reasons for our failure to detect trichloromethylated Ch. It is reason-
able to assume, however, that the reason relates to the experimental condition for CC& activation employed by those workers (e.g. radiation activation and anaerobic atmosphere) which differs from that used by us (e.g. benzoyl peroxide catalysis aerobically or anaerobically) and, differences in the sensitivity of the MS employed
in both cases. We believe that our results can be interpreted as meaning that trichloromethylated Ch may only represent a minor fraction of the expected prod- ucts of simultaneous ’ CC13/CCl,02’ attack and, for that reason, was not detec- table by us. This could particularly be the case when both radicals are produced
under biological experimental conditions where aerobic media predominate. This in- terpretation also explains why we and others reported covalent binding of 14CC14 to Ch and Ch esters under in vitro or in vivo conditions and in the case of liver micro-

S.L. Fanelli et al. / Chemico-Biological Interactions 98 (1995) 223-236 235
somal [5,7,1 l] or mitochondrial lipids [12] and in liver nuclear lipids [9]. Radiolabel detection is far more sensitive than our MS detection procedure. If that detectable fraction represented only a negligible part of the actual cell alterations, as our pres- ent results might suggest, then Ch could be a significant target for damage during Ccl4 or CBrCl, intoxication. Expected biological alterations resulting from these CCl&BrCl, derived free radical-promoted alterations in Ch structure might in- clude perturbation of biological membrane functions. In effect, Ch is a relevant com- ponent of membranes; influences its permeability and stability and modulates membrane protein activity [ 14- 171. Ch is also involved in the metabolism of biliary acids, steroidal and sex hormones and progesterone [ 14- 171. Consequently, if altera- tions in Ch structure such as those here described occurred in vivo during CCL or CBrCl, poisoning, then relevant deleterious consequences might be expected. Stud- ies are in course in our laboratory directed to sensitively detect these altered Ch moieties in vivo during haloalkane poisoning.
Acknowledgements
This work was supported by Grant ES05619-03 from the National Institutes of Health (NIH), USA.
References
[I] T.F. Slater, Activation of carbon tetrachloride: chemical principles and biological significance, in:
D.C.H. MC Brien and T.F. Slater (Eds.), Free Radicals Lipid Peroxidation and Cancer, Academic
Press, New York, 1982, pp. 243-274.
[2] J.A. Castro, Mechanistical studies and prevention of free radical cell injury, in: Proceedings
IUPHAR 9th International Congress in Pharmacology, Vol. 2, The MacMillan Press Ltd., London,
1984, pp. 243-250.
[3] R.O. Recknagel, E.A. Glende, J.A. Dolak and R.L. Walter, Mechanism of carbon tetrachloride
toxicity, Pharmacol. Ther., 43 (I 986) 139- 154.
[4] M.U. Dianzani, The role of free radicals in liver damage, Proc. Nutr. Sot., 46 (1987) 43-52.
[S] ES. Reynolds, Liver parenchymal cell injury: IV. Pattern of incorporation of carbon and chlorine
from carbon tetrachloride into chemical constituents of liver in vivo, J. Pharmacol. Exp. Ther., 155
(1967) 117-126.
[6] J.A. Castro, M.I. Diaz Gomez, EC. de Ferreyra, C.R. de Castro, N. D’Acosta and O.M. de Fenos.
Carbon tetrachloride effect on rat liver and adrenals related to their mixed function oxygenase con-
tent, Biochem. Biophys. Res. Commun., 47 (1972) 315-321.
[7] M.C. Villarruel and J.A. Castro, Carbon tetrachloride target lipids in rat liver microsomes. Effect
of cystamine administration in their pattern of labelling by Ccl,, Biochem. Biophys. Res. Com-
mun., 54 (1973) 108-115.
[8] M.C. Villarruel, E.G.D. de Toranzo and J.A. Castro, Carbon tetrachloride activation, lipid peroxi-
dation and the mixed function oxygenase activity of various rat tissues, Toxicol. Appl. Pharmacol.,
41 (1977) 337-344.
[9] M.I. Diaz Gomez and J.A. Castro, Covalent binding of carbon tetrachloride metabolites to liver
nuclear DNA, proteins and lipids, Toxicol. Appl. Pharmacol., 56 (1980) 199-206.
[IO] J.R. Trudell, R. Bijsterling and A.J. Trevor, Reductive metabolism of carbon tetrachloride by
human cytochrome P450 reconstituted in phospholipid vesicles: mass spectral identification of
trichloromethyl radicals bound to dioleyl phosphatidylcholine, Proc. Natl. Acad. Sci. USA, 79
(1982) 2678-2686.

236 S. L. Fanelli et al. / Chemico-Biological Interactions 98 (1995) 223-236
[l I] G.A. Ansari, MI. Moslen and E.S. Reynolds, Evidence for in vivo covalent binding of ’ Ccl, de-
rived from CC14 to cholesterol of rat liver, Biochem. Pharmacol., 31 (1982) 3509-3510.
[l2] M.C. Villarruel, G. Fernandez, E.C. Aguilar and J.A. Castro, Early biochemical alterations in liver
mitochondria from carbon tetrachloride poisoned rats, J. Appl. Toxicol., 7 (1987) 173-177.
1131 H. Frank, D. Thiel and J. Mac Lead, Mass spectrometric detection of cross-linked fatty acids form-
ed during radical-induced lesion of lipid membranes, Biochem. J., 260 (1989) 873-878.
[I41 P.L. Yeagle, Cholesterol and the cell membrane, Biochim. Biophys. Acta, 822 (1985) 267-287.
1151 R.R. Brenner, Role of cholesterol in the microsomal membrane, Lipids, 25 (1990) 581-585.
[I61 K.1. Okuda, Liver mitochondrial P450 involved in cholesterol catabolism and vitamin D activation,
J. Lipid Res., 35 (1994) 361-372.
[17] J.D. Watson, N.H. Hopkins, J.W. Roberts, J.A. Steitz and A.M. Weiner, Molecular Biology of the
Gene, Vol. 1, The Benjamin/Cummings Publishing Co. Inc., Menlo Park, CA, 1987, pp. 54-59.
(181 G.R. Wailer, Biochemical Applications of Mass Spectrometry, Wiley Interscience, New York, 1972,
pp. 254-265.
[19] P.B. Addis, H.A. Emanuel, S.D. Bergmann, J.H. Zavoral, Capillary GC quantification of choles-
terol oxidation products in plasma lipoproteins of fasted human. Free Rad. Biol. Med., 7 (1989)
179-182.
120) L. Pizzoferrato, S. Nicoli and C. Lintas, GC-MS characterization and quantification of sterols and
cholesterol oxidation products, Chromatographia, 35 (1993) 269-274.
[21] M.C. Villarruel and J.A. Castro, Irreversible binding of carbon tetrachloride to microsomal
phospholipids. Free radical nature of the reactive species and alterations in the physicochemical
properties of the target fatty acids, Res. Commun. Chem. Pathol. Pharmacol., 10(l) (1975)
105-l 16.
[22] J.A. Castro, E.V. Cignoli, C.R. de Castro and O.M. de Fenos, Prevention by cystamine of liver
necrosis and early biochemical alterations induced by carbon tetrachloride, B&hem. Pharmacol.,
21 (1972) 102-114.
[23] M.C. Villarruel, M.1. Diaz Gomez and J.A. Castro, The nature of the in vitro irreversible binding
of carbon tetrachloride to microsomal lipids, Toxicol. Appl. Pharmacol., 33 (1975) 106-I 14.
124) R.O. Recknagel and E.A. Glende, Carbon tetrachloride hepatotoxicity: an example of lethal
cleavage, Crit. Rev. Toxicol., 2 (1973) 263-297.
1251 H. Budzikiewicz, C. Djerassi and D.H. Williams, Structure Elucidation of Natural Products by
Mass Spectrometry. Vol. 2: Steroids, Terpenoids. Sugars and Miscellaneous Classes, Holden-Day
Inc., San Francisco, 1964, pp. 64-101.




















