
Protective Role of Genistein in Acute Liver Damage Induced by Carbon Tetrachloride
Upload
independentCategory
view
1download
0
INTRODUCTION
ADAM (a disintegrin and metalloprotease, ADAM) proteins are expressed in a variety of cells and tissues, including brain, testis, epididymis, ovary, breast, placen-ta, liver, lung, bone, and muscle. ADAMs are a family of membrane-anchored glycoproteins that are involved in the proteolytic processing of membrane-bound precursors, the modulation of cell–cell and cell–matrix interactions, and issue morphogenesis (Matsuno et al., 2006). ADAM8 is an important member of ADAM family. It is a membrane-anchored glycoprotein consisting of an N-terminal pro-domain, followed by a metalloproteinase-, disintegrin-, cysteine-rich-, EGF-like- and transmembrane domain and cytoplasmic tail (Yoshida et al., 1990; Yoshiyama et al., 1997). ADAM8 was initially identified in macro-phages (Yoshida et al., 1990) and subsequently detected in neurons, osteoclasts, leukocytes, neutrophils, epitheli-
al cells, and cancer cells (Choi et al., 2001; Mandelin et al., 2003; Hodgkinson and Ye, 1993; Kelly et al., 2005). ADAM8 may regulate cellular functions by processing membrane receptors that are crucial for cellular signal-ing pathways, and its catalytic activity has been demon-strated in biochemical and cell-based assays towards sub-strates that include HB-EGF, EGF, CD23, and L-selectin. The dysregulation of ADAM8 could therefore influence pathological conditions such as rheumatoid arthritis, can-cer, and asthma (Guaiquil et al., 2010). Activated hepatic stellate cells (HSC) showed increased mRNA expression of ADAM8 compared to quiescent HSC (Schwettmann et al., 2008). But the concrete role of ADAM8 in acute liver injury is still unknown.
Carbon tetrachloride (CCl4)-induced acute liver inju-ry model is similar to the human disease, from the stand-point of morphology and biochemical aspects of colla-gen metabolism. A single dose or very few doses result
Neutralization of ADAM8 ameliorates liver injury and accelerates liver repair in carbon tetrachloride-induced
acute liver injurySan-Qiang Li1, Sha Zhu2, Xue-Dong Wan1, Zheng-Shun Xu1 and Zhao Ma1
1The Molecular Medicine Key Laboratory of Liver Injury and Repair, Medical College, Henan University of Science and Technology, Luoyang 471003, P.R. China
2Department of Microbiology Immunology, College of Basic Medical Sciences Zhengzhou University, 45001, China
(Received December 3, 2013; Accepted February 14, 2014)
ABSTRACT — Although some studies have described the function of ADAM8 (a disintegrin and met-alloprotease 8) related with rheumatoid arthritis, cancer and asthma, etc., the concrete role of ADAM8 in acute liver injury is still unknown. So mice respectively received anti-ADAM8 monoclonal antibody (mAb) of 100 μg/100 μl, 200 μg/100 μl or 300 μg/100 μl in PBS or PBS pre-injection. Then acute liv-er injury was induced in the mice by intraperitoneal (i.p.) injection of carbon tetrachloride (CCl4). Serum AST and ALT level, Haematoxylin-eosin (H&E) staining, the expression level of vascular endothelial growth factor (VEGF), cytochrome P450 1A2 (CYP1A2) and proliferating cell nuclear antigen (PCNA) were detected in the mice after CCl4 administration. Our results showed that anti-ADAM8 mAb pre-injec-tion could effectively lower AST and ALT levels (P < 0.05 or P < 0.01) and reduce liver injury (P < 0.05 or P < 0.01), induce the expression of VEGF, CYP1A2 and PCNA (P < 0.05 or P < 0.01) in dose-depend-ent manner compared with the control mice which received PBS pre-injection. In summary, our study suggested that ADAM8 might promote liver injury by inhibiting the proliferation of hepatocytes, angio-genesis and affecting the metabolism function of liver during acute liver injury induced by CCl4. Anti-ADAM8 mAb injection might be suitable as a potential method for acute liver injury therapy.
Key words: ADAM8, Monoclonal antibody, Acute liver injury, VEGF, CYP1A2, PCNA
Correspondence: San-Qiang Li (E-mail: [email protected])
Original Article
The Journal of Toxicological Sciences (J. Toxicol. Sci.)Vol.39, No.2, 339-351, 2014
Vol. 39 No. 2
339

in acute liver damage that has been widely employed to study morphological and biochemical features of the cel-lular lesions, especially steatosis and necrosis (Song et al., 2007).
Many evidence suggest that ADAMs plays import roles in pathogenesis of various kinds of liver injury. Selective TNF-α converting enzyme (a member of the ADAM family) inhibition could reduce hepatic inflamma-tion and fibrosis in auto-immune hepatitis (Sharma et al., 2013). ADAM17 was significantly up-regulated in liver injury and hepatocarcinogenesis (Berasain et al., 2012). ADAM8 is a negative regulator of retinal neovasculari-zation (Guaiquil et al., 2010). ADAM9 and ADAM12 expression is associated with tumor aggressiveness and progression in liver cancers (Le Pabic et al., 2003).
Although the functions of some ADAMs have been widely studied, there is limited information about the function of ADAM8 during acute liver injury. The pur-pose of the present study therefore, is to study the func-tion of ADAM8 during acute liver injury induced by CCl4 and help to further understand the functions of ADAMs in diseases.
MATERIALS AND METHODS
ChemicalsCarbon tetrachloride (CCl4, purity ≥ 99.5) were pur-
chased from Tianjin Fengchuan Chemical Reagent Co., Ltd. (Tianjin, China). All other reagents and solvents used in this study were purchased commercially and were of analytical pure grade.
AnimalsMale BALB/c mice (approximately 6-8 weeks old,
22 ± 2 g) were purchased from the experimental animals center of Henan Province and maintained in an air-condi-tioned animal room at 25°C with free access to water and food under 12 hr light/dark cycles for the experiments. All animals were allowed to adapt to the environment for 1 week before the experiment and fed on laboratory chow. All protocols conformed to the guidelines from the National Animal Care and Use Committee of China. All animals received care in compliance with the Principles of Laboratory Animal Care.
Induction of acute liver injuryOur previous work showed that 200 μg/100 μl in PBS
containing anti-ADAM8 monoclonal antibody could effectively down-regulate the expression of ADAM8 in liver of mice. So the concentration gradient around 200 μg/100 μl in PBS of anti-ADAM8 monoclonal anti-
body was used to study the function of ADAM8 dur-ing acute liver injury induced by CCl4. Mice were divided into five groups. Every group contains thir-ty mice. 1 hr before CCl4 exposure, PBS, 100 μg/100 μl, 200 μg/100 μl or 300 μg/100 μl in PBS contain-ing anti-ADAM8 monoclonal antibody (mAb) (Santa Cruz, Santa Cruz, CA, USA) were respectively i.p. administered to the mice in the four groups. Sub-sequently mice were i.p. injected with 0.1% CCl4 (1 μl CCl4 in 1 ml mineral oil) to induced acute liv-er injury. Mice in the fifth group (as control group) only received anti-ADAM8 monoclonal antibody (300 μg/100 μl in PBS) pretreatment. In addition, we also detected the expression of ADAM8 in the liver of mice at 0 hr, 6 hr, 24 hr, 42 hr and 72 hr after only CCl4 injection by western blot (n = 30).
Serum AST and ALT activity detectionBlood was drawn through orbital vein of mice at 0,
6, 24, 42 and 72 hr in the mice of the five groups after CCl4 injection. Every time point contained six mice. The coagulated blood is left to clot at room temperature for approximately 15 to 30 min. After it is completely clot-ted, it is rimmed using an applicator stick and then cen-trifuged for approximately 5-10 min at 2,500 rpm. Then the supernatant fluid is separated. Serum AST and ALT activities were determined with a commercial assay kit (Nanjing Jiancheng Biological Technology, Inc., Nanjing, China) at 0, 6, 24, 42 and 72 hr in the mice of five groups after CCl4 injection. Enzyme activities were expressed as an international unit per liter (IU/l).
Histologic ExaminationLiver specimens were obtained from six mice at
each time point. Every liver specimen from one mouse was divided into three sections. They were respective-ly used for histologic examination, western blot and RT-PCR detection. Samples of liver were fixed in 10% for-maldehyde for 24 hr and then dehydrated and embedded in paraffin. Six micrometer sections were cut from each paraffin-embedded tissue and stained with haematoxy-lin-eosin (H&E). Necrotic areas were measured from tis-sue sections that were at least 10 mm2 for each mouse by ImagePro Plus 6.0 software (Media Cybernetics, Rockville, MD, USA). To evaluate the degree of necrosis after acute liver injury, we referred to an injury grading score (Grade 0-4) based on severity of necrotic lesions in the liver parenchyma (Zhu et al., 2010). The grades were as follows: Grade 0, Normal histology; Grade 1, presence of degenerated hepatocytes with only rare foci of necro-sis; Grade 2, Mild centrilobular necrosis around the cen-
Vol. 39 No. 2
340
S.-Q. Li et al.

tral vein, occupying only a part of Rappaport’s zone 3; Grade 3, Established necrosis limited to zone 3; Grade 4: Extensive, confluent centrilobular necrosis involving Rap-paport’s zone 3 and 2.
Immunohistochemistry of VEGF, CYP1A2 and PCNA
Six micrometer sections were cut from each paraf-fin-embedded tissue as prepared above. The sections of liver from the mice in different groups were immu-nostained with a monoclonal antibody to mouse VEGF, CYP1A2 and PCNA (dilution 1:300) (Santa Cruz) as described by Xu et al. (2000). The signal was detected using the Polink-2 plus polymer HRP detection system (Zhongshan, Beijing, China) using DAB. A negative con-trol was carried out on each slide by omitting the pri-mary antibody. Sections were examined microscopical-ly for specific staining and photographs were taken using a digital image-capture system (Olympus, Tokyo, Japan). Numbers of positive cells in the mice of five groups, at least 12 mm2 tissue sections were measured for each mouse by ImagePro Plus 6.0 software (ImagePro Plus (Media Cybernetics).
Western blot of VEGF, CYP1A2 and PCNA The expression level of VEGF, CYP1A2 and PCNA
were detected by western blot to compare liver blood ves-sel genesis, metabolism and cell proliferation in the liv-er of five groups after CCl4 administration. In addition, we also detected the expression of ADAM8 in the liver of mice at 0 hr, 6 hr, 24 hr, 42 hr and 72 hr after only CCl4 injection by western blot. Protein samples of 70 μg from the mice in different groups were adjusted to the compo-sition of the electrophoresis sample buffer (50 mM Tris, pH 6.8, 10% glycerol, 5% beta-mercaptoethanol, 2% SDS, 0.1% bromphenol blue) and boiled for 5 min pri-or to analysis. SDS-PAGE (10% polyacrylamide gels) in 1 mm slab gel was performed as described by Sambrook
and Russell (2001). The proteins were transferred from the gel to the nitrocellulose membranes. Then the mem-brane was probed with a monoclonal antibody to mouse VEGF, CYP1A2 or PCNA (Santa Cruz), respectively. The signal was detected by horseradish peroxidase detection system using DAB (Sigma, St. Louis, MO, USA). Protein bands were quantified with Gel Pro Analyzer software 4.0 (Media Cybernetics Inc., Bethesda, MD, USA) and the intensities of the bands were normalized against beta-ac-tin. Every experiment was repeated for three times.
Semi-quantitative RT-PCR To determine the effects of ADAM8 blocking on blood
vessel genesis, metabolism and cell proliferation of hepa-tocytes at gene level, semi-quantitative RT-PCR was con-ducted. The specific oligonucleotide primers were shown in Table 1. RT-PCR were performed and repeated at least three times on cDNA. Briefly, cDNA was synthesized from 1 μg of RNA in the presence of ribonuclease inhib-itor (BBI, Toronto, Canada), dNTPs, Oligo (dT) 18 prim-ers, and RevertAidTM M-Mulv reverse transcriptase (Fermentas, Hanover, MD, USA) in a total of 25 μl reac-tion mix. RT-PCR was performed using a TaKaRa mRNA Selective PCR kit (TaKaRa, Shiga, Japan). The PCR prod-ucts were visualized by ultraviolet illumination after elec-trophoresis for 30 min at 100 V through 1.5% agarose gels (COSMO BIO, Tokyo, Japan) and staining in Trisbo-rate/ EDTA buffer containing 0.5 mg/ml of ethidium bro-mide. The signal intensity of the bands with the expected size was measured using Gel Pro Analyzer software 4.0 software. The intensities of the bands were normalized against beta-actin. Experiments in the absence of reverse transcriptase were conducted as negative controls.
Statistical AnalysisAll data were presented as the mean ± standard devia-
tion (SD). Statistical comparisons were made using one-way and two-way (without interaction) ANOVA with the
Table 1 Oligonucleotide primers used in RT-PCR
Gene name GenBank Accession No. Forward primer Reverse primer Expected size (bp)
vascular endothelial growth factor (VEGF) NM_001025250.3 5’- GGCAGACTATTCAGCGGACTC 5’- CTCAAACCGTTGGCACGA 154
cytochrome P450, family 1, subfamily a, polypeptide 2 (CYP1A2) NM_009993 5’- CATCGGCTCCACTCCTGT 5’- GCATCTCCTCGCTCTTCC 413
proliferating cell nuclear antigen (PCNA) NM_011045 5’- GTAGTTGTCGCTGTAGGC 5’- GTTGCTCCACATCTAAGTC 492
beta-actin (Actb) NM_007393 5’- CTGTCCCTGTATGCCTCTG 5’- ATGTCACGCACGATTTCC 218
Vol. 39 No. 2
341
The role of ADAM8 in acute liver injury

Tukey post-hoc test for multiple comparisons. All statisti-cal analyses were performed using SPSS 13.0 (SPSS Inc., Chicago, IL, USA).
RESULTS
Alterations of serum AST, ALT level in the mice of five groups post-CCl4 injection
Fig. 1 shows the levels of serum AST and ALT in
the mice of the five groups. AST and ALT levels in the mice with anti-ADAM8 mAb pre-injection at 6 hr, 24 hr, 42 hr and 72 hr post CCl4 injection were significant-ly lower than that in the mice injected with PBS pre-in-jection (P < 0.05 or P < 0.01). With the concentration increasing of anti-ADAM8 mAb pre-injection, the levels of AST and ALT in the mice were down-regulated in a dose-dependent manner post CCl4 injection. The levels of AST and ALT in the mice with only 300 μg/100 μl anti-
Fig. 1. Serum AST (A) and ALT (B) levels in the mice with PBS or anti-ADAM8 mAb of different concentrations pre-in-jection at 0, 6, 24, 42, and 72 hr after administration of 0.1 ml/10 g 0.1% CCl4 (1 μl CCl4 in 1 ml mineral oil). Serum AST (A) and ALT (B) levels in the normal mice with only 300 μg/100 μl ADAM8 mAb pre-injection were also detect-ed as controls. The Data are expressed as means ± S.D. (n = 6). PBS+CCl4, 100ADAM8+CCl4, 200ADAM8+CCl4 and 300ADAM8+CCl4: mice respectively received PBS or anti-ADAM8 mAb pre-injection (100 μg/100 μl, 200 μg/100 μl or 300 μg/100 μl in PBS), and then recovered at room temperature for 1 hr under normal feeding conditions; 0.1% CCl4 was subsequently administered to the mice. 300ADAM8: mice only received 300 μg/100 μl ADAM8 mAb pre-injection. **P < 0.01 or *P < 0.05: significant difference in the each ADAM8 mAb+CCl4 group or 300ADAM8 group as com-pared with the PBS+CCl4 group. #P < 0.05: significant difference in the 300ADAM8+CCl4 group as compared with the 100ADAM8+CCl4 group or 200ADAM8+CCl4 group. & P < 0.05: significant difference in the 300ADAM8+CCl4 group as compared with the 300ADAM8 group.
Vol. 39 No. 2
342
S.-Q. Li et al.

ADAM8 mAb pre-injection were significantly lower than those with 300 μg/100 μl anti-ADAM8 mAb pre-injec-tion and CCl4 injection at 6 hr, 24 hr and 42 hr post CCl4 injection (P < 0.05).
Determination of treatment effect by histological analysis of liver from CCl4 challenged and unchallenged mice
To evaluate liver injury degree, haematoxylin-eosin staining was performed in the liver of mice. Table 2 and Fig. 2 show that anti-ADAM8 mAb pre-injection sig-nificantly reduced liver injury degree compared with the mice which only received PBS pre-injection at 6 hr, 24 hr, 42 hr and 72 hr after CCl4 injection (P < 0.05 or P < 0.01). Anti-ADAM8 mAb of 300 μg/100 μl pre-injec-tion significantly ameliorates liver injury compared with anti-ADAM8 mAb of 200 μg/100 μl or 100 μg/100 μl pre-injection in the mice after CCl4 injection (P < 0.05). Only receiving anti-ADAM8 mAb of 300 μg/100 μl pre-injection did not induce liver injury in the mice (Table 2 and Fig. 2).
Determination of treatment effect by immunohistochemistry of VEGF, CYP1A2 and PCNA in the liver from CCl4 challenged and unchallenged mice
Fig. 3 and Fig. 4 shows that VEGF and CYP1A2 were mostly expressed in the cytoplasm of hepatocytes of mice. The expression of VEGF and CYP1A2 were significantly increased in the hepatocytes of mice with anti-ADAM8
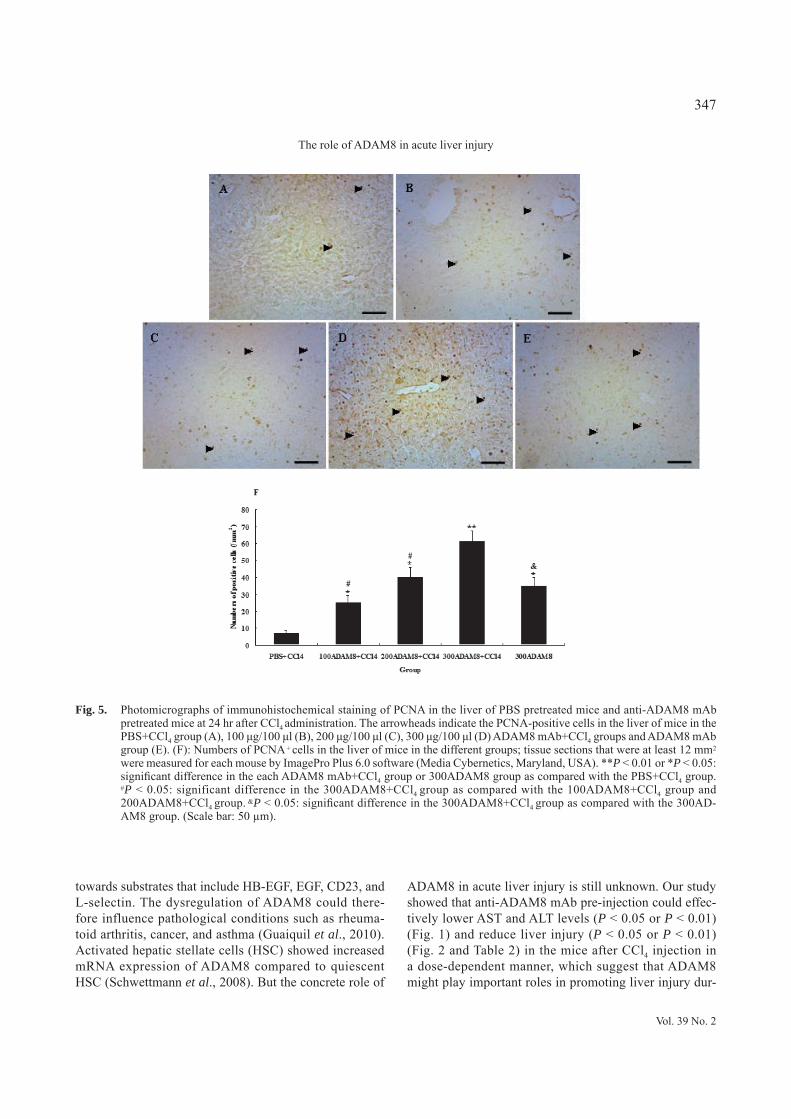
mAb pre-injection (P < 0.05 or P < 0.01) when compared with the control mice that only received PBS pre-injection at 24 hr after CCl4 injection. PCNA was mainly expressed in the nucleus of hepatocytes and was found significantly higher in the hepatocytes of mice with anti-ADAM8 mAb pre-injection than that in the control group which only received PBS pre-injection at 24 hr after CCl4 injection (P < 0.05 or P < 0.01) (Fig. 5). Among three doses, anti-ADAM8 mAb pre-injection of 300 μg/100 μl could more significantly induce the expression of VEGF, CYP1A2 and PCNA than other two doses (P < 0.05). Anti-AD-AM8 mAb pre-injection could also induce the expression of VEGF, CYP1A2 and PCNA at 6 hr and 42 hr (P < 0.05 or P < 0.01), but not significantly induce the expression of those molecules at 72 hr (P > 0.05) after CCl4 injection compared with the control mice that only received PBS pre-injection (data not shown).
Detection the expression of ADAM8 in the liver of mice after only CCl4 injection by Western blot
Fig. 6 shows that the expression of ADAM8 in the liver of mice was significantly increased at 6 hr, 24 hr, 42 hr and 72 hr when compared with 0 h after CCl4 injec-tion (P < 0.05 or P < 0.01).
Determination of treatment effect by Western blot of VEGF, CYP1A2 and PCNA in the liver from CCl4 challenged and unchallenged mice
The expression of VEGF, CYP1A2 and PCNA was analyzed by western blot analysis in the liver of mice at
Table 2 Liver injury degreeGroup of mice 0 hr 6 hr 24 hr 42 hr 72 hrPBS+CCl4 0 2 4 3 2100ADAM8+CCl4 0 1-2*# 3*# 2-3*# 1-2*#
200ADAM8+CCl4 0 1*# 2-3*# 2*# 1*#
300ADAM8+CCl4 0 0-1** 2** 1-2** 0-1**
300ADAM8 0 0** & 0** & 0** & 0** &
Time (hr) after 0.1% CCl4 (1 μl CCl4 in 1 ml mineral oil) i.p. injection in the mice. Injury degree according to severity of necrosis in the liver parenchyma: Grade 0, Normal histology; Grade 1, presence of degenerated hepatocytes with only rare foci of necrosis; Grade 2, Mild centrilobular necrosis around the central vein, occupying only a part of Rappaport’s zone 3; Grade 3, Established necrosis limited to zone 3; Grade 4: Extensive, confluent centrilobular necrosis involving Rappaport’s zone 3 and 2. PBS+CCl4, 100ADAM8+CCl4, 200ADAM8+CCl4 and 300ADAM8+CCl4: mice respectively received PBS or anti-ADAM8 mAb pre-injection (100 μg/100 μl, 200 μg/100 μl or 300 μg/100 μl in PBS), and then recovered at room temperature for 1 hr under normal feeding conditions; 0.1% CCl4 was subsequently administered to the mice. 300ADAM8: mice only received 300 μg/100 μl ADAM8 mAb pre-injection. **P < 0.01 or *P < 0.05: significant difference in the each ADAM8 mAb+CCl4 group or 300ADAM8 group as compared with the PBS+CCl4 group. #P < 0.05: significant difference in the 300ADAM8+CCl4 group as compared with the 100ADAM8+CCl4 group and 200ADAM8+CCl4 group. &P < 0.05: significant difference in the 300ADAM8+CCl4 group as compared with the 300ADAM8 group.
Vol. 39 No. 2
343
The role of ADAM8 in acute liver injury

24 hr after CCl4 injection (Fig. 7). As shown in Fig. 7, anti-ADAM8 mAb pre-injection with different concen-trations significantly induced the expression of VEGF, CYP1A2 and PCNA in the liver of experimental mice compared with the control mice which only received PBS pre-injection at 24 hr after CCl4 injection (P < 0.05 or P < 0.01). Anti-ADAM8 mAb pre-injection of 300 μg/ 100 μl could more significantly induce the expression of VEGF, CYP1A2 and PCNA than other two doses (P < 0.05). At 24 hr after CCl4 injection, anti-ADAM8
mAb pre-injection of 300 μg/100 μl could significantly induce the expression of VEGF, CYP1A2 and PCNA in the liver of mice than those with only anti-ADAM8 mAb pre-injection of 300 μg/100 μl (P < 0.05).
Determination of treatment effect by RT-PCR of VEGF, CYP1A2 and PCNA in the liver from CCl4 challenged and unchallenged mice
In order to assess the effect of anti-ADAM8 mAb on the mRNA expression of VEGF, CYP1A2 and PCNA,
Fig. 2. Histologic examination of liver injury in mice by H&E staining at 24 hr after CCl4 administration. A, B, C, D and E respec-tively represent the degree of liver injury of mice in the PBS+ CCl4 group, 100 μg/100 μl, 200 μg/100 μl and 300 μg/100 μl ADAM8 mAb+CCl4 groups and 300 μg/100 μl ADAM8 mAb group. F: Necrotic areas. Representative findings from meas-ure tissue sections that were at least 10 mm2 for each mouse by ImagePro Plus 6.0 software (Media Cybernetics, Maryland, USA). **P < 0.01 or *P < 0.05: significant difference in the each ADAM8 mAb+CCl4 group or 300ADAM8 group as compared with the PBS+CCl4 group. #P < 0.05: significant difference in the 300ADAM8+CCl4 group as compared with the 100ADAM8+CCl4 group and 200ADAM8+CCl4 group. &P < 0.05: significant difference in the 300ADAM8+CCl4 group as compared with the 300ADAM8 group. (Scale bar: 50 μm).
Vol. 39 No. 2
344
S.-Q. Li et al.

semi-quantitative RT-PCR was performed (Fig. 8). Fig. 8 shows that anti-ADAM8 mAb pre-injection of different concentrations caused a great increase in VEGF, CYP1A2 and PCNA mRNA expression in comparison with the controls which only received PBS pre-injection at 24 hr after CCl4 injection (P < 0.05 or P < 0.01). Anti-ADAM8 mAb pre-injection of 300 μg/100 μl could more signifi-cantly induce the mRNA expression of VEGF, CYP1A2 and PCNA than other two doses (P < 0.05). At 24 hr
after CCl4 injection, anti-ADAM8 mAb pre-injection of 300 μg/100 μl could significantly induce the mRNA expression of VEGF, CYP1A2 and PCNA in the liver of mice than those with only anti-ADAM8 mAb pre-in-jection of 300 μg/100 μl (P < 0.05). These are consistent with the results of protein level.
Fig. 3. Photomicrographs of immunohistochemical staining of VEGF in the liver of PBS pretreated mice and anti-ADAM8 mAb pretreated mice at 24 hr after CCl4 administration. The arrowheads indicate the VEGF-positive cells in the liver of mice in the PBS+CCl4 group (A), 100 μg/100 μl (B), 200 μg/100 μl (C), 300 μg/100 μl (D) ADAM8 mAb+CCl4 groups and ADAM8 mAb group (E). (F): Numbers of VEGF + cells in the liver of mice in the different groups; tissue sections that were at least 12 mm2 were measured for each mouse by ImagePro Plus 6.0 software (Media Cybernetics, Maryland, USA). **P < 0.01 or *P < 0.05: significant difference in the each ADAM8 mAb+CCl4 group or 300ADAM8 group as compared with the PBS+CCl4 group. #P < 0.05: significant difference in the 300ADAM8+CCl4 group as compared with the 100ADAM8+CCl4 group and 200ADAM8+CCl4 group. &P < 0.05: significant difference in the 300ADAM8+CCl4 group as compared with the 300ADAM8 group. (Scale bar: 50 μm).
Vol. 39 No. 2
345
The role of ADAM8 in acute liver injury

DISCUSSION
In this study, we first investigated the role of ADAM8 during acute liver injury in the mice induced by CCl4. We found that only CCl4 injection could significantly induce the expression of ADAM8 (Fig. 6) and anti-ADAM8 mAb pre-injection could effectively ameliorate the liv-
er injury and adverse the effect of ADAM8 in acute liver injury (Figs. 1-5, 7, 8 and Table 2).
Some studies have been performed to dissect the func-tion of ADAM8. ADAM8 may regulate cellular functions by processing membrane receptors that are crucial for cellular signaling pathways, and its catalytic activity has been demonstrated in biochemical and cell-based assays
Fig. 4. Photomicrographs of immunohistochemical staining of CYP1A2 in the liver of PBS pretreated mice and anti-ADAM8 mAb pretreated mice at 24 hr after CCl4 administration. The arrowheads indicate the CYP1A2-positive cells in the liver of mice in the PBS+CCl4 group (A), 100 μg/100 μl (B), 200 μg/100 μl (C), 300 μg/100 μl (D) ADAM8 mAb+CCl4 groups and AD-AM8 mAb group (E). (F): Numbers of CYP1A2 + cells in the liver of mice in the different groups; tissue sections that were at least 12 mm2 were measured for each mouse by ImagePro Plus 6.0 software (Media Cybernetics, Maryland, USA). **P < 0.01 or *P < 0.05: significant difference in the each ADAM8 mAb+CCl4 group or 300ADAM8 group as compared with the PBS+CCl4 group. #P < 0.05: significant difference in the 300ADAM8+CCl4 group as compared with the 100ADAM8+CCl4 group and 200ADAM8+CCl4 group. &P < 0.05: significant difference in the 300ADAM8+CCl4 group as compared with the 300ADAM8 group. (Scale bar: 50 μm).
Vol. 39 No. 2
346
S.-Q. Li et al.
towards substrates that include HB-EGF, EGF, CD23, and L-selectin. The dysregulation of ADAM8 could there-fore influence pathological conditions such as rheuma-toid arthritis, cancer, and asthma (Guaiquil et al., 2010). Activated hepatic stellate cells (HSC) showed increased mRNA expression of ADAM8 compared to quiescent HSC (Schwettmann et al., 2008). But the concrete role of
ADAM8 in acute liver injury is still unknown. Our study showed that anti-ADAM8 mAb pre-injection could effec-tively lower AST and ALT levels (P < 0.05 or P < 0.01) (Fig. 1) and reduce liver injury (P < 0.05 or P < 0.01) (Fig. 2 and Table 2) in the mice after CCl4 injection in a dose-dependent manner, which suggest that ADAM8 might play important roles in promoting liver injury dur-
Fig. 5. Photomicrographs of immunohistochemical staining of PCNA in the liver of PBS pretreated mice and anti-ADAM8 mAb pretreated mice at 24 hr after CCl4 administration. The arrowheads indicate the PCNA-positive cells in the liver of mice in the PBS+CCl4 group (A), 100 μg/100 μl (B), 200 μg/100 μl (C), 300 μg/100 μl (D) ADAM8 mAb+CCl4 groups and ADAM8 mAb group (E). (F): Numbers of PCNA + cells in the liver of mice in the different groups; tissue sections that were at least 12 mm2 were measured for each mouse by ImagePro Plus 6.0 software (Media Cybernetics, Maryland, USA). **P < 0.01 or *P < 0.05: significant difference in the each ADAM8 mAb+CCl4 group or 300ADAM8 group as compared with the PBS+CCl4 group. #P < 0.05: significant difference in the 300ADAM8+CCl4 group as compared with the 100ADAM8+CCl4 group and 200ADAM8+CCl4 group. &P < 0.05: significant difference in the 300ADAM8+CCl4 group as compared with the 300AD-AM8 group. (Scale bar: 50 μm).
Vol. 39 No. 2
347
The role of ADAM8 in acute liver injury

Fig. 6. The expression of ADAM8 in the liver of mice at 0 hr, 6 hr, 24 hr, 42 hr and 72 hr after CCl4 injection. The expression of ADAM8 was detected by Western blot (A). The protein bands were quantified for ADAM8 (B) with the Gel-Pro Analyzer 4.0 software (Media Cybernetics Inc.) and the intensities of the bands were normalized against beta-actin. AU represents arbitrary unit. Every experiment was repeated three times. All data were presented as the mean ± standard deviation (SD). **P < 0.01 or *P < 0.05: significant difference when compared with the expression of ADAM8 at 0 hr after CCl4 injection.
Fig. 7. The expression of VEGF, CYP1A2 and PCNA in the liver of PBS pretreated mice and anti-ADAM8 mAb pre-treated mice at 24 hr after CCl4 administration. The expression of VEGF, CYP1A2 and PCNA were detect-ed by Western blot (A). 1: PBS+CCl4 group; 2: 100 μg/100 μl ADAM8 mAb+CCl4 group; 3: 200 μg/100 μl AD-AM8 mAb+CCl4 group; 4: 300 μg/100 μl ADAM8 mAb+CCl4 group; 5: ADAM8 mAb group. The protein bands were quantified for VEGF (B), CYP1A2 (C) and PCNA (D) with the Gel-Pro Analyzer 4.0 software (Media Cybernetics Inc.) and the intensities of the bands were normalized against beta-actin. AU represents arbitrary unit. Every experiment was repeated three times. All data were presented as the mean ± standard deviation (SD). **P < 0.01 or *P < 0.05: significant difference in the each ADAM8 mAb+CCl4 group or 300ADAM8 group as com-pared with the PBS+CCl4 group. #P < 0.05: significant difference in the 300ADAM8+CCl4 group as compared with the 100ADAM8+CCl4 group and 200ADAM8+CCl4 group. &P < 0.05: significant difference in the 300ADAM8+CCl4 group as compared with the 300ADAM8 group.
Vol. 39 No. 2
348
S.-Q. Li et al.

ing acute liver injury in the mice induced by CCl4. The formation of new blood vessels is critical for nor-
mal development and tissue repair, as well as for patho-logical events such as retinal neovascularization, rheuma-toid arthritis, and tumor growth (Friedlander et al., 2007; Bradley et al., 2007; Chen and Smith, 2007; Olsson et al., 2006; Khong et al., 2007; Sherris, 2007). During physi-ological angiogenesis, vascular endothelial growth factor (VEGF) expression in response to local hypoxia drives the development of new blood vessels (Ferrara, 2002; Holderfield and Hughes, 2008; Carmeliet and Jain, 2000). In addition, Angiogenesis is known to play a critical role in pathological settings like chronic inflammatory and tumor growth (Medina et al., 2004; Tugues et al., 2007; Taura et al., 2008). VEGF is considered to be the cen-tral angiogenic factor during chronic liver injury. It has
been demonstrated that expression of VEGF is up-regu-lated during liver fibrosis, and its expression is increased in activated hepatic stellate cells (HSCs) (Medina et al., 2004; Tugues et al., 2007; Rosmorduc et al., 1999; Maria De Souza et al., 2006). Activated HSCs can express VEGF and VEGF receptors in the liver after CCl4 intox-ication (Ishikawa et al., 1999; Ankoma-Sey et al., 2000). Inflammatory mediators cause the HSCs to differenti-ate into myofibroblasts. They play a role in angiogenesis and act by releasing the proangiogenic mediators VEGF and angiopoietin-1 during the development of liver fibro-sis (Aleffi et al., 2005; Novo et al., 2007). Our results showed that anti-ADAM8 mAb could induce more VEGF expression with the concentration increasing of anti-AD-AM8 mAb (Figs. 3, 7 and 8). So we think that VEGF may play important roles in liver repair during acute liver inju-
Fig. 8. The mRNA expression of VEGF, CYP1A2 and PCNA in the liver of PBS pretreated mice and anti-ADAM8 mAb pre-treated mice at 24 hr after CCl4 administration. The mRNA expression of VEGF, CYP1A2 and PCNA were detected by semi-quantitative RT-PCR (A). 1: PBS+CCl4 group; 2: 100 μg/100 μl ADAM8 mAb+CCl4 group; 3: 200 μg/100 μl ADAM8 mAb+CCl4 group; 4: 300 μg/100 μl ADAM8 mAb+CCl4 group; 5: ADAM8 mAb group. The DNA bands were quanti-fied for VEGF (B), CYP1A2 (C) and PCNA (D) with the Gel-Pro Analyzer 4.0 software (Media Cybernetics Inc.) and the intensities of the bands were normalized against beta-actin. AU represents arbitrary unit. Every experiment was repeated three times. All data were presented as the mean ± standard deviation (SD). **P < 0.01 or *P < 0.05: significant difference in the each ADAM8 mAb+CCl4 group or 300ADAM8 group as compared with the PBS+CCl4 group. #P < 0.05: significant difference in the 300ADAM8+CCl4 group as compared with the 100ADAM8+CCl4 group and 200ADAM8+CCl4 group. &P < 0.05: significant difference in the 300ADAM8+CCl4 group as compared with the 300ADAM8 group.
Vol. 39 No. 2
349
The role of ADAM8 in acute liver injury

ry. Anti-ADAM8 mAb pre-injection could induce VEGF expression to accelerate liver repair after CCl4 injection, which indicated that ADAM8 may decrease the expres-sion of VEGF and aggravate liver injury during acute liv-er injury induced by CCl4.
Our results also indicated that CYP1A2 was signif-icantly up-regulated in the liver of mice with anti-AD-AM8 mAb pre-injection compared with control group which received PBS injection (P < 0.05 or P < 0.01) (Figs. 4, 7 and 8). CYP1A2 is one of the major CYPs in human liver (approximately 13%) and metabolises a vari-ety of clinically important drugs. This enzyme also metab-olizes several important endogenous compounds includ-ing steroids, retinols, melatonin, uroporphyrinogen and arachidonic acid. Like many of other CYPs, CYP1A2 is subject to induction and inhibition by a number of com-pounds (Zhou et al., 2009). CYP1A2 could be used as an important marker for metabolic effects (Filimonova et al., 2011; Yan and Caldwell, 2013). Our results demon-strated that anti-ADAM8 mAb pre-injection could induce CYP1A2 expression to improve the metabolism func-tion in the liver of mice in dose-dependent manner, which suggest that ADAM8 may regulate the expression of CYP1A2 to affect the metabolism function of liver, which result in aggravating liver damage.
ADAM8 is characterized by a metalloprotease and a disintegrin domain, are membrane-anchored glycopro-teins involved in proteolysis and cell adhesion (Matsuno et al., 2006). CYP1A2 and VEGF shedding was mediated by the ADAM8 metalloprotease. Thus, inhibiting expres-sion of ADAM8 in hepatic cells in vivo could potential-ly down-regulate the processing of these proteins. So anti-ADAM8 antibody could induce the expression of CYP1A2 and VEGF during acute liver injury induced by CCl4 in the mice.
Our results also showed that anti-ADAM8 mAb pre-injection could significantly induce PCNA expression in the liver of mice after CCl4 injection (P < 0.05 or P < 0.01) (Figs. 5, 7 and 8). PCNA is a subunit of the mammalian DNA polymerase delta and is synthesized primarily during the S phase of the cell cycle (Celis et al., 1987). PCNA is a relay or anchoring molecule that func-tions as a molecular integrator for proteins involved in the control of the cell cycle, DNA replication, DNA repair, and cell death (Kelman, 1997; Morrow et al., 2004). PCNA has been shown to be a good marker to distinguish proliferating cells (Xu et al., 2000; Assy et al., 1998). Our results explained that anti-ADAM8 mAb pre-injec-tion could promote the proliferation of hepatocytes in the mice after liver injury, which indicated that ADAM8 may promote liver injury by inhibiting the proliferation of
hepatocytes in the mice during acute liver injury induced by CCl4.
In conclusion, our study suggested that ADAM8 might promote liver injury by inhibiting the proliferation of hepatocytes, angiogenesis and affecting the metabolism function of liver during acute liver injury induced by CCl4. Anti-ADAM8 mAb injection might be suitable as a potential method for acute liver injury therapy.
ACKNOWLEDGMENTS
This work was supported by grants from the Nation-al Natural Science Foundation of China (#81201558) to SQL, Program for Science & Technology Innovation Talents in Universities of Henan Province (#13HAS-TIT025) to SQL, Foundation for Henan Province’s Key Project to Tackle Key Problems of Science and Tech-nology (#122102310030) to SQL and the Key Projects for Science and Technology Research of the Education Department of Henan Province (12B310003) to SQL. The authors thank all the members in the laboratory when this work was carried out. The experiments complied with the current laws of China.
REFERENCES
Aleffi, S., Petrai, I., Bertolani, C., Parola, M., Colombatto, S., Novo, E., Vizzutti, F., Anania, F.A., Milani, S., Rombouts, K., Laffi, G., Pinzani, M. and Marra, F. (2005): Upregulation of proinflam-matory and proangiogenic cytokines by leptin in human hepatic stellate cells. Hepatology, 42, 1339-1348.
Ankoma-Sey, V., Wang, Y. and Dai, Z. (2000): Hypoxic stimulation of vascular endothelial growth factor expression in activated rat hepatic stellate cells. Hepatology, 31, 141-148.
Assy, N., Gong, Y., Zhang, M., Pettigrew, N.M., Pashniak, D. and Minuk, G.Y. (1998): Use of proliferating cell nuclear antigen as a marker of liver regeneration after partial hepatectomy in rats. J. Lab. Clin. Med., 131, 251-256.
Berasain, C., Nicou, A., Garcia-Irigoyen, O., Latasa, M.U., Urtasun, R., Elizalde, M., Salis, F., Perugorría, M.J., Prieto, J., Recio, J.A., Corrales, F.J. and Avila, M.A. (2012): Epidermal growth factor receptor signaling in hepatocellular carcinoma: inflam-matory activation and a new intracellular regulatory mechanism. Dig. Dis., 30, 524-531.
Bradley, J., Ju, M. and Robinson, G.S. (2007): Combination thera-py for the treatment of ocular neovascularization. Angiogenesis, 10, 141-148.
Carmeliet, P. and Jain, R.K. (2000): Angiogenesis in cancer and oth-er diseases. Nature, 407, 249-257.
Celis, J.E., Madsen, P., Celis, A., Nielsen, H.V. and Gesser, B. (1987): Cyclin (PCNA, auxiliary protein of DNA polymerase delta) is a central component of the pathway(s) leading to DNA replication and cell division. FEBS. Lett., 220, 1-7.
Chen, J. and Smith, L.E. (2007): Retinopathy of prematurity. Angiogenesis, 10, 133-140.
Choi, S.J., Han, J.H. and Roodman, G.D. (2001): ADAM8: a novel
Vol. 39 No. 2
350
S.-Q. Li et al.

osteoclast stimulating factor. J. Bone. Miner. Res., 16, 814-822.Ferrara, N. (2002): Role of vascular endothelial growth factor in
physiologic and pathologic angiogenesis: therapeutic implica-tions. Semin. Oncol., 29, 10-14.
Filimonova, A.A., Ziganshina, L.E., Ziganshin, A.U. and Chichirov, A.A. (2011): New specific marker of cytochrome P450 1A2 activity. Bull. Exp. Biol. Med., 150, 762-764.
Friedlander, M., Dorrell, M.I., Ritter, M.R., Marchetti, V., Moreno, S.K. and El-Kalay, M. (2007): Progenitor cells and retinal angio-genesis. Angiogenesis, 10, 89-101.
Guaiquil, V.H., Swendeman, S., Zhou, W., Guaiquil, P., Weskamp, G., Bartsch, J.W. and Blobel, C.P. (2010): ADAM8 is a negative regulator of retinal neovascularization and of the growth of het-erotopically injected tumor cells in mice. J. Mol. Med. (Berl)., 88, 497-505.
Hodgkinson, C.P. and Ye, S. (2003): Microarray analysis of per-oxisome proliferator-activated receptor-gamma induced chang-es in gene expression in macrophages. Biochem. Biophys. Res. Commun., 308, 505-510.
Holderfield, M.T. and Hughes, C.C. (2008): Crosstalk between vas-cular endothelial growth factor, notch, and transforming growth factor-beta in vascular morphogenesis. Circ. Res., 102, 637-652.
Ishikawa, K., Mochida, S., Mashiba, S., Inao, M., Matsui, A., Ikeda, H., Ohno, A., Shibuya, M. and Fujiwara, K. (1999): Expres-sions of vascular endothelial growth factor in nonparenchymal as well as parenchymal cells in rat liver after necrosis. Biochem. Biophys. Res. Commun., 254, 587-593.
Kelly, K., Hutchinson, G., Nebenius-Oosthuizen, D., Smith, A., Bartsch, J.W., Horiuchi, K., Rittger, A., Manova, K., Docherty, A.J. and Blobel, C.P. (2005): Metalloprotease–disintegrin ADAM8: expression analysis and targeted deletion in mice. Dev. Dyn., 232, 221-231.
Kelman, Z. (1997): PCNA: structure, functions and interactions. Oncogene, 14, 629-640.
Khong, T.L., Larsen, H., Raatz, Y. and Paleolog, E. (2007): Angio-genesis as a therapeutic target in arthritis: learning the lessons of the colorectal cancer experience. Angiogenesis, 10, 243-258.
Le Pabic, H., Bonnier, D., Wewer, U.M., Coutand, A., Musso, O. and Baffet, G. (2003): ADAM12 in human liver cancers: TGF-beta-regulated expression in stellate cells is associated with matrix remodeling. Hepatology, 37, 1056-1066.
Mandelin, J., Li, T.F., Hukkanen, M.V., Liljestrom, M., Chen, Z.K., Santavirta, S., Kitti, U. and Konttinen, Y.T. (2003): Increased expression of a novel osteoclast-stimulating factor, ADAM8, in interface tissue around loosened hip prostheses. J. Rheumatol., 30, 2033-2038.
Maria De Souza, M., Tolentino, M.Jr., Assis, B.C., Cristina De Oliveira Gonzalez, A., Maria Correia Silva, T. and Andrade, Z.A. (2006): Pathogenesis of septal fibrosis of the liver. (An experimental study with a new model.). Pathol. Res. Pract., 202, 883-889.
Matsuno, O., Miyazaki, E., Nureki, S., Ueno, T., Kumamoto, T. and Higuchi, Y. (2006): Role of ADAM8 in experimental asthma. Immunol. Lett., 102, 67-73.
Medina, J., Arroyo, A.G., Sanchez-Madrid, F. and Moreno-Otero, R. (2004): Angiogenesis in chronic inflammatory liver disease. Hepatology, 39, 1185-1195.
Morrow, P.W., Tung, H.Y. and Hemmings, H.C. (2004): Rapamy-cin causes activation of protein phosphatase-2A1 and nuclear translocation of PCNA in CD4+ T cells. Biochem. Biophys. Res. Commun., 323, 645-651.
Novo, E., Cannito, S., Zamara, E., Valfrè di Bonzo, L., Caligiuri, A., Cravanzola, C., Compagnone, A., Colombatto, S., Marra,
F., Pinzani, M. and Parola, M. (2007): Proangiogenic cytokines as hypoxia-dependent factors stimulating migration of human hepatic stellate cells. Am. J. Pathol., 170, 1942-1953.
Olsson, A.K., Dimberg, A., Kreuger, J. and Claesson-Welsh, L. (2006): VEGF receptor signalling - in control of vascular func-tion. Nat. Rev. Mol. Cell. Biol., 7, 359-371.
Rosmorduc, O., Wendum, D., Corpechot, C., Galy, B., Sebbagh, N., Raleigh, J., Housset, C. and Poupon, R. (1999): Hepatocel-lular hypoxia-induced vascular endothelial growth factor expres-sion and angiogenesis in experimental biliary cirrhosis. Am. J. Pathol., 155, 1065-1073.
Sambrook, J. and Russell, D.W. (2001): Molecular cloning - a lab-oratory manual, 4rd ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor.
Schwettmann, L., Wehmeier, M., Jokovic, D., Aleksandrova, K., Brand, K., Manns, M.P., Lichtinghagen, R. and Bahr, M.J. (2008): Hepatic expression of A disintegrin and metallopro-teinase (ADAM) and ADAMs with thrombospondin motives (ADAM-TS) enzymes in patients with chronic liver diseases. J. Hepatol., 49, 243-250.
Sharma, M., Mohapatra, J., Malik, U., Wagh, A., Singh, A., Patel, H.M., Pandey, D., Kadam, S., Shah, G.B., Chatterjee, A. and Jain, M.R. (2013): Selective inhibition of tumor necrosis factor-α converting enzyme attenuates liver toxicity in a murine model of concanavalin A induced auto-immune hepatitis. Int. Immunop-harmacol., 17, 229-236.
Sherris, D. (2007): Ocular drug development—future directions. Angiogenesis, 10, 71-76.
Song, J.Y., Li, L., Ahn, J.B., Park, J.G., Jo, J.S. and Park, D.H. (2007): Acute liver toxicity by carbon tetrachloride in HSP70 knock out mice. Exp. Toxicol. Pathol., 59, 29-34.
Taura, K., De Minicis, S., Seki, E., Hatano, E., Iwaisako, K., Osterreicher, C.H., Kodama, Y., Miura, K., Ikai, I., Uemoto, S. and Brenner, D.A. (2008): Hepatic stellate cells secrete angiopoie-tin 1 that induces angiogenesis in liver fibrosis. Gastroenterology, 135, 1729-1738.
Tugues, S., Fernandez-Varo, G., Muñoz-Luque, J., Ros, J., Arroyo, V., Rodés, J., Friedman, S.L., Carmeliet, P., Jiménez, W. and Morales-Ruiz, M. (2007): Antiangiogenic treatment with sunitinib ameliorates inflammatory infiltrate, fibrosis, and portal pressure in cirrhotic rats. Hepatology, 46, 1919-1926.
Xu, C.S., Lu, A.L., Xiong, L. and Li, G.W. (2000): Changes of ACP, AKP, HSC70/HSP68 and PCNA in growth and development of rat Liver. Dev. & Reprod. Biol., 9, 1-14.
Yan, Z. and Caldwell, G.W. (2013): In vitro identification of cyto-chrome P450 enzymes responsible for drug metabolism. Methods Mol. Biol., 1015, 251-261.
Yoshida, S., Setoguchi, M., Higuchi, Y., Akizuki, S. and Yamamoto, S. (1990): Molecular cloning of cDNA encoding MS2 antigen, a novel cell surface antigen strongly expressed in murine mono-cytic lineage. Int. Immunol., 2, 586-591.
Yoshiyama, K., Higuchi, Y., Kataoka, M., Matsuura, K. and Yamamoto, S. (1997): CD156 (human ADAM8): expression, primary amino acid sequence, and gene location. Genomics, 41, 56-62.
Zhou, S.F., Yang, L.P., Zhou, Z.W., Liu, Y.H. and Chan, E. (2009): Insights into the substrate specificity, inhibitors, regulation, and polymorphisms and the clinical impact of human cytochrome P450 1A2. AAPS. J., 11, 481-494.
Zhu, R.Z., Xiang, D., Xie, C., Li, J.J., Hu, J.J., He, H.L., Yuan, Y.S., Gao, J., Han, W. and Yu, Y. (2010): Protective effect of recom-binant human IL-1Ra on CCl4-induced acute liver injury in mice. World J. Gastroenterol., 16, 2771-2779.
Vol. 39 No. 2
351
The role of ADAM8 in acute liver injury
Copyright © 2022 FDOKUMEN




















