
1º Mendel segun Mendel(color)
190
[1] Prólogo Sobre los experimentos de Mendel se ha escrito mucho, no cabe duda, pero siempre interpretándolos, no exponiendo sus experimentos como tales. Pero cabría decir a esto que para eso ya está el Versuche über Pflanzen- Hybriden, donde son expuestos los experimentos de Mendel tal como los concibió. Es verdad. Pero las interpretaciones son tales, que se le atribuyen a Mendel cosas, ideas o procesos, hoy llamados genéticos, que jamás los concibió ni pudo hacerlo, por la falta de información que en aquella época podía obtener, respecto de la constitución celular. Y de esta podríamos atrevernos a afirmar que la más importante era la existencia de los cromosomas. Y en cambio no se describe, no se explica aquellas otras ideas y conceptos científicos que en aquella época fueron verdaderamente adelantadas a su tiempo. El presente libro versa sobre la monografía de Mendel titulada Versuche über Pflanzen-Hybriden, en la cual, como es sabido, expone su autor los experimentos y la explicación de los experimentos, reconocidos actualmente como el origen histórico y el fundamento primero, tanto teórico como experimental, de la moderna ciencia de la genética. Más concretamente, el presente libro, o trabajo científico histórico, es un estudio de índole teórico- crítica, que comprende cinco partes principales: La primera, relativa a dar una breve semblanza de Mendel como abad y prelado; la segunda, la más importante, está subdividida, a su vez, en otras dos: una, relativa a reconocer y definir el significado de los entes biológicos considerados por Mendel en sus experimentos y en la explicación de ellos; la otra, destinada a esclarecer el significado preciso de las fórmulas y las expresiones formales empleadas por Mendel para simbolizar, a la vez que aquellos entes, las relaciones biológicas entre ellos, necesarios tanto para exponer cuanto para explicar los resultados de los propios experimentos. En la parte tercera, exponemos sensu
-
Upload
independent -
Category
Documents
-
view
6 -
download
0
Transcript of 1º Mendel segun Mendel(color)

[1]
Prólogo
Sobre los experimentos de Mendel se ha escrito mucho, no cabe duda, pero
siempre interpretándolos, no exponiendo sus experimentos como tales. Pero
cabría decir a esto que para eso ya está el Versuche über Pflanzen-
Hybriden, donde son expuestos los experimentos de Mendel tal como los
concibió. Es verdad. Pero las interpretaciones son tales, que se le atribuyen a
Mendel cosas, ideas o procesos, hoy llamados genéticos, que jamás los
concibió ni pudo hacerlo, por la falta de información que en aquella época
podía obtener, respecto de la constitución celular. Y de esta podríamos
atrevernos a afirmar que la más importante era la existencia de los
cromosomas. Y en cambio no se describe, no se explica aquellas otras ideas y
conceptos científicos que en aquella época fueron verdaderamente
adelantadas a su tiempo.
El presente libro versa sobre la monografía de Mendel titulada Versuche über
Pflanzen-Hybriden, en la cual, como es sabido, expone su autor los
experimentos y la explicación de los experimentos, reconocidos actualmente
como el origen histórico y el fundamento primero, tanto teórico como
experimental, de la moderna ciencia de la genética. Más concretamente, el
presente libro, o trabajo científico histórico, es un estudio de índole teórico-
crítica, que comprende cinco partes principales: La primera, relativa a dar una
breve semblanza de Mendel como abad y prelado; la segunda, la más
importante, está subdividida, a su vez, en otras dos: una, relativa a reconocer y
definir el significado de los entes biológicos considerados por Mendel en sus
experimentos y en la explicación de ellos; la otra, destinada a esclarecer el
significado preciso de las fórmulas y las expresiones formales empleadas por
Mendel para simbolizar, a la vez que aquellos entes, las relaciones biológicas
entre ellos, necesarios tanto para exponer cuanto para explicar los resultados
de los propios experimentos. En la parte tercera, exponemos sensu

[2]
matematico, a estilo de Mendel, lo que debe ser el significado de fenotipo y de
genotipo de forma rigurosa, análogo a lo que hace las matemáticas con el
significado, por ejemplo, del signo de ∫; la integral definida, cuyo significado
huye, se escapa a las ideas y conceptos subjetivos. Ya que esta podrá ser
explicada -su significado- con conceptos más o menos acertados, pero el
significado es uno y solo uno. Pues lo mismo creemos que se debe proceder
con el significado de fenotipo y genotipo.
En la parte anteúltima, la cuarta, exponemos de la forma más clara posible
aquel proceder teórico-práctico en los experimentos de Mendel en el
Versuche über Pflanzen-Hybriden, y que hoy viene a ser llamado el Método
Científico.
Y para terminar, una pequeña descripción histórica, que aportará alguna
claridad a la información de ciertos artículos y libros, sobre la introducción de la
genética en España.
En fin, el presente libro está principalmente concebido con objeto de aclarar y
puntualizar cuál es propiamente la auténtica teoría sostenida por Mendel en
aquella monografía suya, en realidad más famosa que verdaderamente
conocida.
Así que, este libro que hemos realizado tiene mucho más interés que el
meramente histórico o el puramente epistemológico que al pronto se advierte.
Cuando nuestras ya clásicas y tradicionales ideas acerca del gen se hallan,
como ocurre en el momento actual, en evidente y profunda crisis, es
sumamente importante, incluso para el progreso de la ciencia misma -no solo
de la teoría de la ciencia-, averiguar hasta qué punto las fundamentales
llamadas leyes de Mendel implican o presuponen la noción de algo como gen,
factor o determinante, conforme se ha venido creyendo, o lo que realmente
implican o presuponen. Y no digamos de ese otro casi inexistente modo de
investigar en las Universidades, donde casi todos los jóvenes y los no tan
jóvenes están ayunos de lo que es el método científico.
Y en esta tarea de exponer el Versuche über Pflanzen-Hybriden según
Mendel, es obvio que el reconocimiento de los conceptos y relaciones bilógicas

[3]
considerados por él, y el estudio del significado de los símbolos y fórmulas
utilizados también por Mendel importa sobremanera llevarlos a cabo a través
de las propias palabras de Mendel. Y esto es lo que justifica el título del libro
“Mendel según Mendel”. Para ello, me he servido del “Versuche Ubber
Pflanzenhybriden” en los Verhandlungen des naturforschenden Vereines in
Brünnn, Band IV,1865 (publicado en 1866), Abhandlungen pag. 3 a 47
(reproducción facsímil editada por la revista The journal of Heredity, en el num.
1 del Vol. 42, correspondiente al año 1951). Pero, además, he tenido en
cuenta, como no debía ser de otra manera, las páginas web:
http://www.mendel-museum.com/, http://himetop.wikidot.com/tutorials-how-to-add-new-items-to-himetop,
http://www.mendelmuseum.muni.cz/en/news/, http://archive.fieldmuseum.org/mendel/story_life.asp,
http://www.mendelmuseum.muni.cz/en/contact/ http://economictimes.indiatimes.com/topic/Mendel-
Museum-of-Genetics/photos, http://www.mendelmuseum.muni.cz/en/photogallery/garden/ y
http://www.mendelweb.org/, entre otras. También en nuestro estudio hemos tenido en
cuenta el libro de Sinnot, Dunn y Dobzhansky “Principios of Genetica”, 5ª
edición 1958, págs. 419 a 443. Asimismo, he tenido en cuenta la versión
española, de aquella misma memoria de Mendel, hecha por Arturo Burkart y
publicada, primero en la Revista Argentina de Agronomía, 1934, 1,3; y luego,
en el tomo titulado “Cuatro estudios sobre Genética” de la colección Maestros
de la Ciencia de EMECË Editores, S.A. Buenos Aires, 1946.
Todos estos lugares, así como libros de la memoria de Mendel, me han
ayudado sobremanera, aunque no siempre los haya encontrado por completo
de mi gusto y, en consecuencia, haya optado por insertar - particularmente al
final del libro- la traducción hecha por nosotros, la cual creemos que es
rigurosa, entre otras cosas, por ser del alemán al español.
Y para terminar y no cansar al lector, no puedo ni podemos por menos que
recordar con este libro a un gran maestro y científico. Es para nosotros el más
alto honor dedicar este libro a la memoria de un hombre, genial en la labor
científica, pero que fue un proscrito por la sociedad científica de su tiempo, el
Prof. Galán. Un hombre con una sed insaciable de saber y una ponderable
capacidad de trabajo que eclipsaba la labor de sus colaboradores. Pero
además, nos mostró ese anhelo por legalizar en su espíritu la innumerable

[4]
información sobre la ciencia que había acumulado en su memoria tras la
lectura y la observación. No solo fue un adelantado a su tiempo, sino también a
sus más íntimos colaboradores.
Y ahora me viene al pensamiento que <<.. desde la remota antigüedad la
enseñanza ha tenido dos problemas básicos: atinar acerca de qué
conocimientos se debían transmitir al discípulo y atinar acerca de cómo había
de hacerse esa transmisión… de la sabiduría a la ignorancia…>>.

[5]
PAGINA
1) Introducción…………………………………………………………………….6
2) El abad y prelado Gregor Johann Mendel………………………………...11
3) La obra científica de Mendel. Su índole teórica y experimental…………18
4) El significado del Fenotipo y Genotipo en la genética mendeliana…. 61
5) Concepto del método científico……………………………………………..80
6) Introductores de la Genética experimental en España………………….98
7) Versuche über Pflanzen-Hybriden por Gregor Mendel……………...122
(Traducción del Alemán al Español por F. Galán-Gutiérrez y Sonia Blasco Sancho).
BIBLIOGRAFÍA……………………………………………………………….170

[6]
INTRODUCCIÓN.
Este libro, que versa sobre la obra científica de Mendel titulada Versuche über
Pflanzen-Hybriden, constituye un estudio exhaustivo hecho con el doble
propósito: por un lado, de precisar exactamente la teoría genética formulada
por el propio Mendel, destacando -dentro de la explicación de los resultados
dada por Mendel- cuáles son los enunciados que tienen la categoría de
principios fundamentales, y cómo estos mismos enunciados que son principios
fundamentales se relacionan lógicamente con las proposiciones que se
expresan en los resultados experimentales; y de reconocer, por otro lado, la
forma de proceder experimental puesta en práctica por Mendel, la cual
constituye, a la postre, el paradigma de lo que hoy conocemos como Método
Científico.
A nuestros lectores podrá, en principio, parecerles plausible nuestro propósito
por que piensen, con razón, que los clásicos son siempre merecedores de
estudio, y Mendel lo es no solo de la Biología moderna, sino también del
método teórico experimental de la ciencia. Pero, quizá, ese mismo propósito
nuestro también pueda resultarles extraño y algo injustificado, ocioso y fatuo
por el hecho de que tratemos de explicar de nuevo la teoría genética
formulada por Mendel, sabedores de la existencia de numerosísima bibliografía
al respecto. Nosotros pensamos, sin embargo, que, aunque han pasado 147
años desde que Mendel dio a conocer la memoria de sus investigaciones,
todavía no se conocen de forma rigurosa, a pesar de haber sido objeto de

[7]
intenso estudio, pues todavía se atribuyen caprichosamente a Mendel
conocimientos, conceptos y datos experimentales que jamás propuso, tuvo o
realizó. Bibliografía de artículos, libros y direcciones web, ejemplo de esto
La memoria de Mendel, que figura con el número 121 en la colección de
monografías Ostwalds Klassikerf der exakten Wissenschaften, ha sido
traducida prácticamente a todos los idiomas. La traducción al español hecha
por el profesor Prevosti, publicada en el libro “Principios de Genética” de
Sinnott, Dunn y Dobzhansky de 1961, puede considerarse la más rigurosa y la
más acreditada de las españolas, aunque contiene numerosas incorrecciones
arrastradas de la traducción inglesa de la monografía de Mendel. Bibliografía
de algunas traducciones en español ordenadas según sus incorrecciones, de
menos a más.
A pesar de esta difusión, y aunque parezca increíble, no se conoce todavía
con el rigor que merece cuál es exactamente la teoría formulada por Mendel
para explicar los resultados de sus experimentos en los guisantes. Pruebas
indiciarias de lo poco riguroso que es todavía el conocimiento que se tiene de
la teoría de Mendel las hay en gran número. Baste citar que hay una gran
diversidad de sistemas de enunciados que, con el nombre de leyes de Mendel
o leyes mendelianas de la herencia, son ofrecidas al estudiante universitario
de genética por los tratadistas como propuestas por Mendel, cuando son una
simple interpretación actual de los experimentos de Mendel. Y no digamos del
empeño palpable de que Mendel formuló el concepto de factor mendeliano,
como hoy lo podemos entender, cuando, en realidad, jamás utilizó tal concepto,
pues no es necesario para entender sus experimentos ni sus hipótesis
explicativas (incorrectamente las llamadas teorías). Y entrando en este empeño

[8]
de los factores mendelianos, llegamos al craso error que cometen los grandes
tratadistas de la historia de la ciencia de creer que los “elementos” que utilizo
Mendel en sus “ Schluss-Bemerkungen / Concluding Remarks/
Consideraciones finales” vienen a ser los famosos factores mendelianos. Y lo
cierto es que Mendel, como decíamos y con el riesgo de ser reiterativos, no
necesitó de esa idea de la bolita o trocito en el cromosoma, que, sin embargo,
parece necesaria para nuestras pobres mentes.
Ciertamente, en el transcurso de los años, entre los autores se ha ido
acentuando cada vez más la tendencia a no enunciar como ley ninguna de las
expresiones que en los primeros tiempos del mendelismo se solían dar con los
nombres de “ley de dominancia” o "ley de la uniformidad de la 1ª generación
filial", reduciéndose, así, el conjunto de principios mendelianos, más o menos
formalmente enunciados, a dos: el de la ley de la segregación o disyunción (
como 1ª ley de Mendel); y el de la ley de la segregación independiente o de la
recombinación al azar (como 2ª ley de Mendel). Pero se sigue creyendo en los
factores mendelianos, y únicamente el concepto biológico que utilizó Mendel,
como ente biológico fundamental de sus experimentos, fue el carácter. Nada
parecido, pues, al concepto de factores que atribuyen a Mendel los genetistas
al tratar ellos mismos de entender los experimentos de Mendel mediante ese
concepto de factor mendeliano.
Importa mucho insistir sobre la índole no solo experimental, sino también
teórica de la obra científica de Mendel, por cuanto en libros de texto,
manuales, monografías, comentarios, etc., los autores destacan la parte
experimental de ella, pero pasan de puntillas por la parte teórica, sin dedicarle
la muy especial atención que requiere, y que desde luego merece. Porque la

[9]
parte teórica de la obra de Mendel es genuina y resulta ser una rigurosa teoría
científica, y no una vaga especulación pseudofilósofica. No olvidemos que las
hipótesis son el germen de las teorías (hipótesis explicativas en la filosofía de
la ciencia), y las teorías, a su vez, la esencia del conocimiento científico.
Conscientemente imbuidos de este reconocimiento acerca de la índole, no
menos teórica que experimental, de la obra de Mendel, hemos escrito este
libro, aspirando a contribuir a la pública y rigurosa divulgación de la obra de
Mendel. Recordemos que aquel sencillo, pero genial fraile agustino, cultivando
vulgares guisantes en el jardín de su convento y reflexionando more
mathemático sobre los resultados de estos cultivos, estableció los fundamentos
y concibió el método de una de las más nuevas, originales, heurísticas y
fecundas orientaciones de la Biología moderna, La Genética.
Merece la pena, para terminar esta introducción, hacer referencia a la genética
molecular por su impacto experimental y mediático. Y esto se debe a que,
siendo una ciencia metodológicamente tan distinta de la genética mendeliana -
aunque originariamente suscitada por esta misma-, que el nombre de genética
molecular con el que se la conoce no resulta, en verdad, ni el más apropiado ni
el más significativo. Sensacionales, resonantes y revolucionarios son los
hallazgos biológicos de la genética molecular, los más sensacionales y
resonantes quizá de la Biología de todos los tiempos, pero que, no obstante,
versan acerca de la química o bioquímica del material genético, por lo cual ya
no son genética propiamente dicha, sino Química o Bioquímica, más
concretamente hablando. Así, se da una vez más en la historia de las ciencias
el interesante caso de que de las manos de los más recientes o adelantados

[10]
cultivadores de una ciencia surge otra ciencia metodológica muy distinta de la
originaria.

[11]
El abad y el prelado Gregor Johann Mendel
El abad y prelado Gregor Johann Mendel
El 6 de enero de 1884, a los 61 años de edad (había nacido el 22 de julio de
1822), moría en el convento de Santo Tomás de Brünn, del que era abad y
prelado, el religioso agustino Gregor Johann Mendel, quien, algún tiempo
después, habría de pasar a la historia de la ciencia como el descubridor de las
leyes de la herencia que llevan su nombre; las famosas “leyes de Mendel”.
Dicho convento (el “Königns-Kloster de Brünn”), regido por la orden
agustiniana, estaba situado (dentro de la histórica región de Moravia) en la
ciudad que, perteneciente en tiempos de Mendel, primero al imperio austriaco y
luego al imperio austro húngaro, llevaba en lengua alemana el nombre de
Brünn, y que hoy, como perteneciente a la actual república Checa, lleva, en
cambio, en idioma checo, el nombre de Brno.
http://www.mendel-museum.com/eng/2visit/

[12]
El convento se cerró definitivamente en 1949, siendo destinado hasta hoy a
Museum Mendelianum.
Este monasterio fortificado se realizó a tal efecto para la defensa de los
habitantes de localidades cercanas. Los monjes cistercienses, que residían
desde 1322, tuvieron que abandonar el monasterio en el siglo XVIII, lo que
originó serios desperfectos en sus instalaciones. En 1793 fue rehabilitado por
los monjes agustinianos que procedían de un céntrico edificio en Brno, al ser
desalojados por el emperador austro-húngaro Francisco José, con el fin de
instalar allí su residencia y fijar sus oficinas.
Próximo a 1854, el monasterio de Santo Tomás estaba regido por el abad
Napp. La orden agustiniana venía precedida por una fama de excesivo
liberalismo, asunto éste que al abad no le preocupaba en demasía. Asimismo,
Napp presidía desde 1827 la más que prestigiosa Real Sociedad Imperial para
el desarrollo de las ciencias naturales y la mejora de la Agricultura en Moravia
(conocida popularmente como la sociedad para la agricultura, fundada por el
emperador Francisco I, el cual ordenó a los monjes del monasterio de santo
Tomás que enseñasen en el propio instituto local las asignaturas de religión y
matemáticas). El mismo año de 1854, comienzan los trabajos experimentales
del monje Gregor Mendel en los huertos del monasterio, con el consentimiento
total del abad.
Hasta la fecha el monje Gregor Mendel pasaba bastante desapercibido a los
ojos de muchos otros. Nacido en 1822, era el hermano mediano y único chico
de una familia de tres hijos, que crecieron en el seno de una familia de

[13]
agricultores de Moravia, que por aquel entonces pertenecía al imperio austriaco
y que hoy forma parte de la república checa.
La personalidad de Gregor Mendel era cambiante y oscilaba entre el
pesimismo del padre, Anton, de carácter rudo; al igual que su hermana mayor
Veronica, y la bondad de la madre Rosine, al igual que su hermana pequeña,
Teresa, que gozaban de un carácter más dulce y sosegado.
Gregor fue un niño brillante e incluso ambicioso, pero los escasos recursos
familiares hacían difícil que lograse una educación mayor que la obtenida en la
escuela primaria. Además, los continuos brotes de una enfermedad
psicosomática que padecía, hacían que tuviese que mantenerse en la cama
durante semanas. El padre y su hermana mayor no aceptaban esta clase de
comportamiento, pero su madre y la hermana pequeña eran bastante
indulgentes con él. Teresa, además, tuvo un papel fundamental en la
educación de su hermano, puesto que le cedió parte de la dote que le
correspondía para que, tras graduarse en la escuela secundaria, pudiese
realizar un curso de dos años, requerido para todos los estudiantes que
deseaban ir a la universidad, en el instituto de filosofía de la localidad de
Olomuc. Este sacrificio fue posteriormente recompensado por Gregor Mendel
cuando colaboró activa y económicamente en la educación de los tres
sobrinos, hijos de Teresa. Sin embargo, ni el obtener una beca escolar, ni sus
esfuerzos para ganar dinero proporcionaban los recursos suficientes si quería
recibir una formación superior (por aquel entonces, para una persona sin
recursos, el único modo de conseguirlo era ordenarse sacerdote).
Mendel tuvo la fortuna de tener como profesor de física en el instituto de
filosofía a Friedrich Franz, que además de monje era un gran amigo del abad

[14]
Napp. Así, con la recomendación del propio Franz por las habilidades
intelectuales de Mendel, el abad accedió a aceptarlo en el monasterio de Santo
Tomás en 1843, a la edad de 20 años, y en él estudió durante los siguientes
cinco años con el fin de ordenarse sacerdote, comenzando como novicio y
pasando por subdiácono y diácono. Ciertamente, estos pasos fueron más
rápidos de los habituales, debido a la ausencia de párrocos por la zona.
Los monjes de la época atendían a los pacientes terminales del hospital de
Santa Ana, de manera que acababan contrayendo enfermedades fatales, que
fueron las causantes de ese bajo número de monjes. Así, Mendel fue ordenado
sacerdote el 6 de agosto de 1847, a la edad de 25 años, y añadió un año más a
su formación antes de tomar responsabilidades pastorales. Pronto, su timidez e
inseguridad hicieron ver que no conectaba con sus parroquianos y, además,
tuvo un nuevo brote de esa extraña enfermedad que le mantenía en cama
durante tiempo indefinido. El abad Napp, consciente de esta situación, decidió
que sería más útil como profesor, de modo que el obispado le envió a la
localidad de Znojmo como instructor de matemáticas y de griego en la escuela
secundaria secular de esta ciudad. Siendo un acierto la decisión del abad Napp
ya que como profesor fue todo un éxito; y aquellas indisposiciones de Mendel
en su labor pastoral no aparecieron con los niños. Incluso estaba bien
considerado entre sus compañeros docentes.
Su deseo por entonces era obtener el título de profesor universitario de
ciencias, pero en 1850 no superó los exámenes oral y escrito necesarios para
esa acreditación. Muchos biógrafos del monje checo han especulado sobre
este fracaso y lo han atribuido a una enfermedad de ansiedad relacionada con
su trastorno psicosomático.

[15]
Seis años después se volvió a presentar y obtuvo el mismo resultado, a pesar
de haberse preparado durante dos años para ese examen en la Universidad de
Viena. Así que Mendel siguió enseñando el grado escolar durante muchos
años, pero su fracaso en los exámenes anteriores confirieron a sus trabajos un
cierto grado de amateurismo entre la gente de su época. Sin embargo, Mendel
recibió unos conocimientos matemáticos superiores, lo que le permitió aplicar el
rigor matemático a sus experimentos en guisantes como nadie lo había
practicado.
Podríamos decir que era un joven de gran talento, pero sin el grado académico
suficiente para su independencia económica. No obstante, cabe especular que,
si Gregor Mendel hubiese aprobado los exámenes para ser docente en la
universidad, probablemente no hubiese tenido tiempo para desarrollar esos
experimentos que le hicieron pasar a la posteridad.
Entre los intentos de ganar acreditación, el monje Mendel inició sus primeros
experimentos cruzando ratones. Sin embargo, el estamento eclesiástico
católico no comulgaba con ese tipo de trabajos ni con la ciencia experimental
en general. Pero las influyentes amistades del abad Napp y la reputación de
sus monjes como profesores impidieron que se parase la actividad científica del
monasterio; las cuales, por cierto, trató de impedir el obispo Antón Ernst
Schaffgostch .
A finales de 1854 Mendel dejó los ratones y comenzó sus experimentos con
guisantes en el jardín experimental del monasterio. De estos experimentos no
sabemos si Mendel guardó anotaciones, puesto que la única referencia que
tenemos es su artículo en la sociedad de agricultura de 1865. Se piensa que
dichas anotaciones fueron quemadas.

[16]
Los experimentos de Mendel finalizaron en el verano de 1863. En ese mismo
año podemos decir que la condición física de Gregor Mendel estaba bastante
deteriorada y comenzaba, además, a tener problemas de visión.
Durante dos años Mendel trabajó en un artículo que describiese sus
experimentos. Una vez realizado este, lo llamó Versuche über Pflanzen-
Hybriden (experimentos de hibridación en plantas), el cual lo presentó a la
sociedad agrícola de Brno en dos entregas: una el miércoles 8 de febrero y
otra, también un miércoles, el 8 de marzo. La sociedad lo publicó, con una
extensión de 44 páginas, y entregó a Mendel 40 copias que había solicitado
con la intención de enviarlo a los científicos europeos más prestigiosos,
incluyendo al propio Charles Darwin.
La primera impresión que se extrae de su trabajo de 1865 es que Mendel tenía
una mente extraordinariamente ordenada.Desgraciada y sorprendentemente, la
reacción a su trabajo no pudo ser más fría. Sin embargo, Mendel envió a
finales de 1866 una copia de su trabajo al profesor de botánica Karl von Nageli
de la Universidad de Munich, y dos meses después recibió respuesta del
científico alemán, en la que alaba su trabajo experimental, pero considera que
no es lo suficientemente explícito como para establecer a partir de él unas
leyes generales. Mendel decidió responder a esa carta con otra misiva que
contenía explicaciones aclaratorias de su trabajo. Al no recibir respuesta a esta
última carta, Mendel escribe una tercera y una cuarta carta solicitando que él
mismo podría establecerse como asistente de Nageli. Esta última carta no
recibió contestación pero la correspondencia continuó de manera intermitente
durante siete años.

[17]
En marzo de 1868 fue elegido abad del monasterio de Santo Tomás en una
turbulenta segunda vuelta, sucediendo así al abad Napp, que había fallecido a
la edad de 75 años. Sus obligaciones administrativas le hicieron abandonar el
trabajo experimental, y, además, un tornado en 1870 destruyó el invernadero
que había construido para el cultivo de sus plantas.
Finalmente, continuó al servicio del monasterio y de la orden agustiniana hasta
su muerte, el 6 de enero de 1884. En sus últimos años de vida Mendel comenta
a algunos amigos -respecto del anonimato de sus trabajos experimentales-
que “su día llegaría”. Posteriormente, Anselm Rambousek, al que Mendel
había vencido en las elecciones a abad, fue nombrado nuevo abad y todas las
notas y apuntes de su predecesor, Mendel, fueron quemadas.

[18]
La obra científica de Mendel.
Glosario de algunos términos
FPf: forma de plantas (en el sentido menos utilizado por Mendel) = lo que hoy se “
llama” fenotipo.
FPg: forma de plantas (en el sentido más utilizado por Mendel) = lo que hoy se “ llama” genotipo.
CE: carácter esencial
CD: carácter diferente = caracteres que se excluyen recíprocamente, caracteres alelomorfos
CED: carácter esencialmente diferente = caracteres no alelomorfos
CP: células polen.
CG: células germen.
Su índole teórica y experimental.
Los principalísimos y básicos conceptos para entender la obra de
Mendel.
1). El principal ente biológico considerado por Mendel.
Antes de entrar en la cuestión del significado de algunas palabras utilizadas por
Mendel, conviene conocer los diversos entes, cosas, elementos, factores
biológicos -en rigor, las diversas clases o géneros de entes biológicos- que
Mendel considera en la explicación de sus resultados experimentales:

[19]
Entre todos los géneros de entes biológicos considerados por Mendel, el más
elemental y fundamental de todos, es el de los caracteres. Todos los demás
géneros de entes biológicos considerados, lo son de entes definidos o
definibles por referencia más o menos mediata a entes del género carácter, es
decir, por referencia a algún carácter.
entes biológicos considerados = ƒ (carácter)
El género carácter, o cualquiera de los caracteres considerados por Mendel,
resultan indefinibles porque no pueden ser definidos por referencia a otros
entes biológicos de género alguno.
2). Entes biológicos definidos o definibles por referencia inmediata a
algún carácter. Usados por Mendel como formas de plantas; esto es
“clases” de plantas que pueden observarse un carácter o varios, en ella,
directamente.

[20]
De los entes biológicos definidos o definibles por referencia inmediata
(contigua) a entes del género carácter, es decir, por referencia inmediata a
algún carácter, Mendel considera dos géneros de ellos distintos, y de muy
diferente importancia, a los cuales, sin embargo, no distingue nominalmente, y
habla en todo caso de formas de plantas al referirse a estos entes. Nosotros
los distinguiremos diciendo “formas de plantas”, FP, en el sentido menos usado
por Mendel, que vendría a ser lo que hoy llamamos fenotipo; y que lo
simbolizaremos por FPf, esto es, vendría a ser la clase de planta definida por el
carácter que es observado en esa misma, planta;
<< Eintheilung und Ordnung der Versuche................ Erbsenformen zeigten
Unterschiede in der Länge und Färbung des Stengels, in der Grösse und Gestalt
der Blätter, in der Stellung, Farbe und Grösse der Blüthen, in der Länge der
Blüthenstiele, in der Farbe, Gestalt und Grösse der Hülsen, in der Gestalt und
Grösse der Samen, in der Färbung der Samenschale und des
Albumens...............>>

[21]
<< Clasificación y orden de los experimentos……….las diferentes formas de
guisantes elegidas para fecundación mostraron diferencias en la longitud y coloración
del tallo, en el tamaño y forma de las hojas, en la posición, color y tamaño de las
flores, en la longitud del pedúnculo floral, en el color, forma y tamaño de las
legumbres,…>>>
Pero debemos de hacer una puntualización en honor al rigor de esta clase de
plantas. Forma de plantas en el sentido menos usado por Mendel, que hoy es
equivalente al fenotipo de las plantas, como ya hemos dicho; puede ser unas
veces de fenotipo simple, respecto a un carácter, o fenotipo complejo, respecto
a los caracteres 1,2,.....n. En estas expresiones verbales que Mendel utiliza de manera
inmediata con referencia a los entes del género carácter, no distingue entre
formas y clases. Así, todas las plantas que tienen la misma forma, para
Mendel, son todas las plantas que están incluidas en la misma clase. Por tanto
es más claro entender clase de plantas que por “formas”.
3). Entes biológicos usados por Mendel como formas/clases de plantas
que se definen en función de los caracteres observados en su
descendencia obtenida por autopolinización.
Y por “formas de plantas” FP en el sentido más usado por Mendel, vendría a
ser lo que hoy designamos como genotipo mendeliano, y podemos simbolizarlo
como, FPg, y por ende, podemos verlo esto es, comprobarlo en cualquiera de
los tantos párrafos en los que se refiere a esta clase de formas de plantas; en
el sentido que acabamos de apuntar;
<<..Bei jedem dieser Versuche wird eine bestimmte Anzahl Pflanzen mit dem
dominirenden Merkmale constant. Für die Beurtheilung des Verhältnisses, in
welchem die Ausscheidung der Formen mit dem constant bleibenden Merkmale
erfolgt, sind die beiden ersten Versuche von besonderem Gewichte, weil bei
diesen eine grössere Anzahl Pflanzen verglichen werden konnte. Die

[22]
Verhaltnisse 1.93 :1 und 2.13 : 1 geben zusammen fast genau das Durchschnitts-
Verhältniss 2 :1..>>
<<… En cada uno de estos experimentos cierto número de plantas fueron constantes
con el carácter dominante. Para apreciar la proporción según la cual ocurre la
separación de las formas con el carácter constante, son de particular importancia los
dos primeros experimentos porque en ellos cabe comparar gran número de plantas.
Las proporciones 1'93:1–2´13:1 dan reunidas casi exactamente la proporción-
promedio 2:1..>>.
<<..Das Durchschnitts-Verhältniss 2:1 erscheint demnach als gesichert. Es ist
damit erwiesen, dass von jenen Formen, welche in der ersten Generation das
dominirende Merkmal besitzen, zwei Theile den Hybriden Character an sich
tragen, ein Theil aber mit dem dominirenden Merkmale constant bleibt...>>
<<… La relación promedio 2:1 parece, pues, como segura. Con esto resulta probado
que, de aquellas formas que en la primera generación poseen el carácter dominante,
dos terceras partes llevan el carácter híbrido, pero una permanece constante con el
carácter dominante.…..>>
Donde se puede comprobar que el carácter de ser “plantas constantes” o
“aquellas formas que en la primera generación poseen el carácter
dominante” es algo que no puede ser observado directamente, pero si
observando la proporción de las formas de plantas que se obtenga, de dicha
planta, en su descendencia por autopolinización. ¡¡ No por el carácter que
pueda verse en ella directamente !!.
Por tanto, podemos afirmar, que la forma de plantas en el sentido menos usado
por Mendel; equivaldría hoy al fenotipo, FPf. Y formas de plantas en el sentido
más usado por Mendel; es hoy a lo que llamamos genotipo, FPg .
Forma de plantas en el sentido más usado por Mendel significa: clase de
plantas, cada una de las cuales tiene la propiedad que, entre todos sus hijos
originados por autopolinización, se presenta un determinado carácter, en la
proporción determinada

[23]
.
Ya que, entre todos sus hijos, aparecen otras plantas, en su respectiva
proporción, hijas de la autofecundación1, en las que no se observa tal carácter,
sino, el respectivo “carácter diferente”.
______________________________________________________________
1. Aquí la autofecundación esta contemplado según el lenguaje bilógico de hoy, ya que según Mendel autopolinización.

[24]
En el preciso sentido mendeliano del término “diferente”. Esto es, refiriéndose a
dos caracteres “alelomorfos”, según el lenguaje de hoy. Pero hemos de aclarar,
que lo que tienen en común todas las plantas pertenecientes a una forma de
plantas respecto a un carácter, en el sentido más usado por Mendel, y que, por
tanto, sirve para definir a esta forma, no es un carácter cualquiera observable
en aquellas plantas, sino únicamente aquel carácter observable en una
proporción determinada para cada forma específica de plantas de que se trate,
en los hijos, producto de la autopolinización
<<.. In der weiteren Besprechung werden jene Merkmale, welche ganz oder fast
unverändert in die Hybride-Verbindung übergehen, somit selbst die Hybriden-
Merkmale repräsentiren, als dominirende, und jene, welche in der Verbindung
latent werden, als recessive bezeichnet. Der Ausdruck "recessiv" wurde
desshalb gewählt, weil die damit benannten Merkmale an den Hybriden
zurücktreten oder ganz verschwinden, jedoch unter den Nachkommen
derselben, wie später gezeigt wird, wieder unverändert zum Vorscheine
kommen.………>>
<<.. En lo sucesivo aquellos caracteres que se transmiten completamente o casi
completamente invariables, a la combinación híbrida puesto que propiamente
representan el carácter híbrido, serán designados como dominantes, y aquellos que en
dicha combinación vienen a ser latentes, como recesivos……………..>>

[25]
.
Y a pesar de correr el peligro de ser reiterativos en la explicación sobre esta
clase o forma de plantas más utilizada por Mendel, es justificado, por su
importancia científica, por un lado, y por otro, por el craso error que se comete
por muchos genéticos, unas veces por no haberse percatado ya por una lectura
poco cuidadosa del “Versuche” y otras por no haber leído el “Versuche”; pero
hoy día se llega a afirma que Mendel no tenía claro lo que hoy llamamos
fenotipo y genotipo. Así que sigamos en este tema sobre esa clase de
formas/plantas que Mendel definió su significado como lo que hoy llamamos
genotipo de forma tan clara, que hoy día sigue siendo válido para las plantas
hermafroditas.
Pues bien, entonces, si el carácter observable, respecto al cual se define
expresamente una forma específica de plantas en el sentido más usado por
Mendel, es un carácter dominante, la proporción determinada de hijos por
autofecundación de la planta perteneciente a la forma que se define, y que se

[26]
observa tal carácter, o es 1 o es ¾ (también según la forma específica de la
planta de que se trate). Por el contrario, si el carácter observable, definido
expresamente por la forma específica de plantas en el sentido más usado por
Mendel, es un carácter recesivo, entonces la proporción determinada de hijos
por autofecundación de la planta perteneciente a la forma que se define es 1, o
es ¼ .Urge señalar que la forma/clase de plantas, en el sentido más usado por
Mendel, está definida únicamente con referencia a plantas hermafroditas,
esto es, que sus flores son perfectas; que tienen gineceo y androceo
completamente desarrollados. Y que ya las formas hibridas como las
constantes únicamente se denominan así por la proporción matemática de las
formas obtenidas por autopolinización. No por cruzamientos por emasculación,
por accidentes ambientales o insectos. Ni en plantas dioicas.
Así Mendel no contempló, entre otras, las plantas dioicas. Como ya
señalábamos anteriormente. Las cuales no es aplicable dicha definición. Es por

[27]
tanto que con rigor el significado de forma de plantas, en el sentido más usado
por Mendel, no es exactamente equivalente al significado actual de “genotipo”,
de modo que no se puede afirmar que todo genotipo sea una “forma de
plantas”.. Así en el caso de las plantas dioicas el genotipo híbrido se obtiene
de padres constantes (exactamente equivalente al concepto actual de
homocigóticas) como de padres híbridos; análogos a los cruzamientos por
emasculación que podríamos efectuar con Pisum. Podemos observar en la Fig.
12, primer caso, planta femenina, constante, con células germen ‘a’ o ‘A’ y la
planta macho, con células polen ‘A’ y ‘a’; y el caso al contrario (planta
femenina, con células polen ‘A’ y ‘a’ y la planta macho, constante, con células
germen ‘a’ o ‘A’). Así como las dos, planta hembra y planta macho, híbridas.
Donde su descendencia seria plantas híbridas con el carácter dominante ya
para plantas constantes como con el carácter dominante como para el
recesivo. Cosa que en las plantas hermafroditas sólo cabría explicarlo
suponiendo que los padres fueron dos plantas constantes para un carácter
diferente, uno dominante y otro recesivo.
4.) Otras clases de entes biológicos para el carácter; el carácter esencial,
carácter esencialmente diferente y los caracteres diferentes.
Aunque como ya hemos dicho para los entes del género carácter, no tiene
definición mediata ni inmediata con referencia a otros entes biológicos
definidos por Mendel. Cabe decir, por tanto, algo que apunte hacia el
significado de este término mendeliano, del carácter.
Mendel emplea como sinónimos el vocablo germánico Merkmal y el de origen
griego Character, si bien aquel con un poco más de frecuencia que este otro.

[28]
Cosa que puede ser comprobada leyendo uno de los párrafos de Versuche
über Planzen-Hybriden,
<<….in dieser generation treten nebst den dominirenden Merkmalen auch die
recessiven in ihrer vollen eigenthümlichkeit wieder auf, und zwar in dem
entschieden ausgesprochenen durchschnitts-verhältnisse 3: 1, so dass unter je
4 pflanzen aus dieser generation 3 den dominirenden und eine den recessiven
Character erhalten.....>>
<<..en esta generación, reaparecen, al lado de los caracteres dominantes, también los
recesivos en su completa peculiaridad, y, por cierto en la decisiva y expresiva
proporción media 3:1, de modo que, de cuatro plantas de esta generación, tres
ostentan el carácter dominante y una el recesivo....>>.
También podemos comprobar, de esta manera, que Mendel considera pares de
caracteres, y cada miembro del par está relacionado por dominancia o por su
inversa, la de recesividad.
Continuando acerca de los caracteres según Mendel, unas veces se refiere al
carácter esencial (wesentliche Merkmale) CE; otras, a los caracteres
esencialmente diferentes (wesentlich verschiedene Merkmale) CED, y otras
a los caracteres diferentes (differierende Merkmale) CD, naturalmente con
significados distintos.
En alemán: CE (wesentliche Merkmale); CED (wesentlich verschiedene
Merkmale); y CD (differirende MerKmale).
En Ingles; CE (essential character); CED (essentially different characters))
y CD (differentiating characters).

[29]
Importa, pues, aclarar estos distintos significados. Empecemos por el
significado que parece más fundamental y es un poco más dificultoso de
explicar: el carácter esencial, CE ; el cual es un ente algo abstracto, algo así
como un hueco o sitio que puede ser llenado de manera determinada. Son
ejemplos, en Pisum, de carácter esencial la forma de los cotiledones (en cuanto
pueden ser redondas o rugosas), el color de las flores (en cuanto pueden ser
rojo-violetas o blancos), etc. El carácter esencial CE de Mendel es el mismo
concepto de la expresión “cualquiera de los dos miembros de un par
mendeliano de caracteres”, pero no debe confundirse jamás con la posible
expresión “alguno de los dos miembros de un par mendeliano de caracteres”.
En cambio, lo que Mendel entiende por caracteres esencialmente diferentes,
CED y por caracteres diferentes, CD es mucho más fácil de explicar. Lo que
entiende por caracteres diferentes son dos caracteres que se excluyen
recíprocamente, de modo que no hay ningún individuo planta en el que se
observen ambos caracteres a la vez. Desde el punto de vista puramente
teórico, el número de caracteres que puede haber siendo entre sí excluyentes
cualesquiera dos de ellos, puede ser dos o mayor que dos. Un número mayor
que dos de caracteres entre sí recíprocamente excluyentes, ocurre realmente
en los casos de no dominancia (dominancia incompleta, como suele decirse), y
en los que hoy llamamos de alelomorfismo múltiple, que no es el caso de los
experimentos de Mendel. Aquellos (los de no dominancia), aunque conocidos
por Mendel, no fueron experimentados por él. Por tanto un ejemplo de
caracteres diferentes CD sería rugoso y liso, tallo cortó y tallo largo, etc. Por
último, los caracteres esencialmente diferentes, CED son aquellos caracteres
considerados conjuntamente, tal que cualesquiera dos de estos caracteres

[30]
están en el caso de ser observados ambos a la vez en algún individuo planta,
y asimismo, en algún otro individuo planta en el que alguno de los dos
caracteres no es observable, porque en vez de él, se observa otro. Este otro es
precisamente dicho con la propia terminología de Mendel, el carácter diferente
CD (dominante o recesivo) de aquél, del que se deja de observar. Así por
ejemplo, en las plantas empleadas por Mendel en sus experimentos, son
caracteres esencialmente diferentes, CED; rugoso, rojo-violeta, axilares, corto,
etc. que son los correspondientes caracteres esenciales, CE, forma de los
cotiledones, color de las flores, posición de las inflorescencias y longitud del
tallo, respectivamente. Entonces estos caracteres esencialmente diferentes,
CED son los entes biológicos que llenarían esos huecos de los que hablamos
al definir los caracteres esenciales, CE.
Para terminar con lo relativo al significado de estas expresiones de Mendel,
veamos que, refiriéndolas a términos de la genética pos mendeliana actual,

[31]
caracteres diferentes CD casi equivale a caracteres alelomorfos. Decimos
“casi” y no “equivale”, ya que los caracteres alelomorfos pueden ser más de
dos, y los caracteres diferentes, para Mendel no pueden ser más carácter
dominante y el recesivo. Caracteres esencialmente diferentes, CED equivalen
a caracteres no alelomorfos, y en el caso de los caracteres esenciales CE no
existe equivalencia en el léxico genético actual.
5) Otros entes utilizados por Mendel por referencia mediata de algún
carácter.
Los otros entes utilizados por Mendel y definidos o definibles por referencia no
inmediata, pero si mediata, en función de algún carácter, esto es, por referencia
indirecta o directa a algún carácter.
Mendel considera sólo un género más, al que denomina formas de células-
germen, o formas de células-polen. En algunas ocasiones emplea la
expresión clases de células-polen/células-germen en vez de formas. Así
podemos leer en su Versuche über Planzen-Hybriden;

[32]
<<..War die obige Annahme richtig, so mussten sich an den Hybriden Keim- und
Pollenzellen von den Formen AB, Ab, aB, ab entwickeln, und es wurden
verbunden:
1. Die Keimzellen AB, Ab, aB, ab mit den Pollenzellen AB.
2. " AB, Ab, aB, ab " ab.....>>
<<…si la anterior suposición era cierta tenían que desarrollarse en el híbrido células-
germen y células-polen de las formas AB, Ab, aB, ab, y unirse:
1. las células–germen AB, Ab, aB, ab con las células-polen AB.
2. las células–germen AB, Ab, aB, ab “ “ ab……..>>.
<<.....Auch beschränkt sich die gemachte Voraussetzung darauf, dass bei der
Bildung der verschiedenartigen Keim- und Pollenzellen die gleiche Anzahl
angestrebt werde, ohne dass diese an jeder einzelnen Hybride mit
mathematischer Genauigkeit erreicht werden müsste...>>
<<…….Después de todo la precedente suposición se limita a [afirmar] que en la
formación de las distintas clases de células-germen y células-polen se tiende a
iguales números [pero] sin que esto se tenga que ser alcanzado con exactitud
matemática….>>

[33]
La memoria de Mendel; un paradigma de comunicación
científico-natural y teórico-experimental.
El método científico en Versuche über Pflanzen-Hybriden.
En este momento reparemos -desde luego muy sumariamente- en aquellas
razones metodológicas que hay que reconocer en el texto de la memoria de
Mendel, los enunciados o proposiciones del discurso, los cuales procedería
considerar como el más auténtico paradigma de lo que hoy día llamamos el
método científico (ver capítulo 5º). Respecto a la teoría propia de una ciencia
natural experimental, se distinguen en todo caso, en cualquier ciencia tres
clases principales (de enunciados que son: 1º) enunciados protocolarios o
de observación (que describen concretamente los resultados experimentales
observados), proposiciones protocolarias; 2º) generalizaciones inductivas
(hablando con más rigor, hipótesis inductivas, obtenidas por inducción a
partir de los enunciados protocolarios de observación); 3º) hipótesis
explicativas (ideadas ad hoc, a fin de deducir de ellas, como conclusiones
lógicamente necesarias, las generalizaciones inductivas, y, explicar los
resultados experimentales observados). En base a estos criterios, la memoria
de Mendel es un ejemplo verdaderamente modélico de comunicación científico-
natural y teórico-experimental del método científico. En Versuche über
Plfanzen Hybriden aparecen nítidamente deslindados y bien caracterizados,
según su respectiva condición metodológica con arreglo a las tres susodichas
clases principales de enunciados esenciales a toda teoría científica natural,
cuantos párrafos importantes del discurso lo requieren. Ahora bien, los
enunciados que en las ciencias naturales, sobre todo en las llamadas ciencias

[34]
naturales exactas (como la Astronomía, la Física, la Química, etc.) merecen la
consideración de leyes -termino éste, en el léxico de las ciencias y de la teoría
de ellas, sobrecargado de connotaciones extra científicas- son siempre o bien
generalizaciones inductivas (hipótesis inductivas), o bien hipótesis explicativas.
El Método Científico en el “Versuche”
1). Proposiciones protocolarias o de observación que pueden ser
consideradas en Versuche über Pflanzen-Hybriden.
Como ya decíamos antes, una proposición protocolaria es, en cualquier
ciencia natural, un enunciado -constituido por una oración o por varias- que
tiene como objeto de expresión o referencia objetiva un hecho particular
observado, un hecho experimental. En Genética, el tipo de hecho a que se
refiere cualquier proposición protocolaria es un conjunto de individuos (de
hombres, de animales, de plantas, de bacterias o de virus); pero -y esto es lo
más importante ahora- a un conjunto de individuos observables, manejable, y
numerable, no sólo en el sentido teórico de la palabra, sino en el práctico, por
cuanto tal conjunto ha sido efectivamente observado, manejado y contado por
un observador, o por varios. En este sentido diremos, como característica más
distintiva de las proposiciones protocolarias en Genética, que éstas se refieren
concretamente a conjuntos asequibles, esto es, efectivamente practicable,
accesible en la práctica al observador.
Como ejemplos de proposiciones protocolarias en Versuche über
Pflanzen-Hybriden, podemos considerar las siguientes:

[35]
1. Versuch. Gestalt der Samem Von 253 Hybriden wurden im zweiten Versuchsjahre 7324 Samen erhalten. Darunter waren rund oder rundlich 5474, und kantig runzlig 1850 Samen. Daraus ergibt sich das Verhältniss 2.96: 1.
1. Experimento. Forma de las semillas. De 253 híbridos se obtuvieron en el segundo año 7324 semillas. De ellas, 5474 fueron redondas o redondeadas; y 1850, rugosas y angulosas. De lo que resulta la relación 2'96: 1.
2. Versuch. Färbung des Albumens. 258 Pflanzen gaben 8023 Samen, 6022
gelbe und 2001 grüne; daher stehen jene zu diesen im Verhältllisse 3.01: 1.
2. Experimento. Coloración del albumen. 258 plantas dieron 8023 semillas, 6022 amarillas y 2001 verdes; por lo que aquéllas están con éstas en la relación 3'01 : 1
3. ………………………………………………………………………………………………………………………………………………………………………
Las proposiciones protocolarias relatan de forma más o menos sumaria,
pero concretísima, lo que ocurrió en un tiempo o fecha perfectamente
determinado, y en un sitio asimismo determinado, según observación
personal de uno o varios autores también determinados; en nuestro caso
las observaciones realizadas por Mendel en su jardín del monasterio de
Santo Tomas de Brüno, hoy Brno.

[36]
Las proposiciones protocolarias llámense, por otro nombre, enunciados de
observación
2). Enunciados subrayados por Mendel que son generalizaciones
inductivas, o hipótesis inductivas, así como las hipótesis explicativas de
su teoría.
Y siguiendo en nuestra lectura de Versuche über Pflanzen-Hybriden. Su
primer subrayado corresponde a la generalización inductiva/hipótesis
inductiva sobre las proporciones numéricas de la forma híbrida y de las formas
constantes en la generación hija por autofecundación de una planta híbrida
respecto a un carácter esencial, CE. Podemos leer:
<<...wird es nun ersichtlich, dass die Hybriden je zweier differirender Merkmale
Samen bilden, von denen die eine Hälfte wieder die Hybridform entwickelt,
während die andere Pflanzen gibt, welche constant bleiben, und zu gleichen
Theilen den dominirenden und recessiven Character erhalten…>>
<<… cuando el carácter dominante se distingue, según su significación, como carácter
híbrido y como carácter parental. Puesto que los miembros de la primera generación
se producen inmediatamente de las semillas de los híbridos, es pues evidente que los
híbridos de cada dos caracteres diferentes producen semillas, de las cuales una mitad
desarrolla nuevamente la forma híbrida, mientras que la otra da plantas que
permanecen constantes y que en igual número reciben respectivamente el carácter
dominante y el recesivo >>.
En este párrafo, Mendel, se refiere a las plantas que define como híbridas,
(formas híbridas), y que son tales porque en su descendencia por
autopolinización (y no antes de observar la descendencia) el número de estos
distintos individuos que cabe pensar que compongan el conjunto ideal, están

[37]
en las proporciones tales que: ½ lo compone el conjunto de formas de plantas
híbridas del total de individuos del conjunto; ¼ lo componen las formas de
plantas constantes uno de los caracteres distintos del que hemos obtenido el
híbrido, del número total de individuos del conjunto ideal; ¼ lo componen las
formas de plantas constantes para el otro carácter. Como se ve por el análisis
que se acaba de exponer, del enunciado de Mendel que nos ocupa, y por cierto
uno de los más importantes, por no decir el más importante, se trata de
caracteres observables en plantas, de formas de plantas definidas respecto a
estos caracteres y de proporciones de estas formas de plantas dentro de
ciertos conjuntos de plantas que no son sino determinadas generaciones. Y por
otro lado estos valores no se refieren a un experimento en concreto; estos
valores son los que suponemos que se aproximarían todos los resultados de
los experimentos que podríamos realizar en las mismas condiciones, pero que
no hemos hecho todavía,…pues aun no ha desaparecido la vida. Los demás
conceptos que intervienen en el enunciado, son conceptos, como formar,
desarrollar, tener, etc. En resumen: esta generalización empírica/hipótesis
inductiva de Mendel, la primera de su monografía, está formulada en términos
que no se refieren a entes que sean lo que en genética actual llama genotipos,
sino, términos de formas de plantas que se refieren a entes que si son
propiamente genotipos; pero -y esto es todavía más notable- no se hace
referencia a ningún ente, explícitamente ni implícitamente, que sean como los
genes, factores, determinantes, etc., esto es, que sean partes componentes de
la planta o de sus células; los más elementales entes biológicos a que se
hace referencia es a caracteres observables en las plantas, que se obtienen
por autopolinización de aquellas. Además, hemos de añadir que no se trata

[38]
explícitamente, ni implícitamente en el enunciado de Mendel, de otros entes
biológicos que no sean los que hacen referencia a caracteres observables
directamente en las plantas, o caracteres observables, también directamente
en plantas, pero que se han obtenido por autofecundación de aquellas.
Consideremos ahora otro enunciado de los subrayados por Mendel, que
también es una generalización inductiva/hipótesis inductiva y que podemos
leer:
<<..... die Nachkommen der Hybriden, in welchen mehrere wesentlich
verschiedene Merkmale vereinigt sind, stellen die Glieder einer
Combinationsreihe vor, in welchen die Entwicklungsreihen für je zwei
differirende Merkmale verbunden sind. Damit ist zugleich erwiesen, dass das
Verhalten je zweier differirender Merkmale in Hybrider Verbindung unabhängig
ist von den anderweitigen Unterschieden an den beiden Stammpflanzen..... >>
<<…. los descendientes de los híbridos en los cuales están reunidos varios
caracteres esencialmente distintos, representan los miembros de una serie de
combinación, en la cual están reunidas las series de desarrollo, de cada dos
caracteres diferentes [contrapuestos]. Con esto resulta probado al mismo tiempo que
el comportamiento de cada dos caracteres diferentes en combinación híbrida es
independiente de los demás diferencias en ambas plantas genitoras.......>>
Este enunciado que se acaba de transcribir, lo formula Mendel inmediatamente
después de exponer la descripción y los resultados de aquellos experimentos
suyos a los que designa como “primer experimento” y “segundo experimento”.
Hemos de reconocer, que en este enunciado, la expresión de Mendel no es tan
clara como en el enunciado considerado anteriormente. El lector, ante estas
expresiones, no puede menos de preguntarse, ¿qué es exactamente lo que

[39]
quiere decir Mendel con ellas ? Nosotros creemos que lo que quiere decir con
la expresión
<<....... los descendientes de los híbridos........., representan (vorstallen) los
términos de una serie de combinación.....>>
es que el tipo, la clase, de conjuntos al cual pertenecen los descendientes del
híbrido que se trate, está definido por cierta expresión formal, simbólica, que es
la que Mendel llama “serie de combinación”. A su vez, media la expresión:
<<...en la cual están combinadas las series de desarrollo para cada dos
caracteres diferentes....>> y que Mendel aclara, que en esa expresión
simbólica, se obtiene o cabe deducirla, mediante una operación de calculo, a
partir de varias expresiones también simbólicas, pero sencillas y elementales
tales que definen las formas de plantas constante (A y a) y formas de
plantas hibridas (Aa más Aa) que se obtienen por autofecundación de un
híbrido respecto a un par de caracteres esenciales, no de caracteres
esencialmente diferentes. Esto es, la expresión simbólica sencilla
A + 2Aa + a
Serán tantas como pares de CD en los cuales es híbrida la planta, de la cual
tratamos los descendientes, por autopolinización. Así pues, de nuevo es una
hipótesis inductiva, una generalización inductiva, que deducimos de los datos
protocolarios, estructurada matemáticamente, y a lo que Mendel llama “serie de
combinación”, y que suponemos que por ahora y “siempre” podremos utilizar
para interpretar aquellos casos particulares que fueron los datos protocolarios.
Y que con esta “serie de combinación”, baste decir que en el caso de realizar

[40]
un experimento para varios pares de CED, la hipótesis inductiva o
generalización inductiva, es precisamente lo que se obtiene al multiplicar
matemáticamente las distintas, expresiones sencillas y elementales, o
trinomios, que son las series de desarrollo para cada dos CD para cada par de
caracteres distintos CD, obteniéndose, lo que podemos llamar polinomios con
un número determinado de términos en función de los trinomios a multiplicar.
La expresión de cálculo mediante la cual, a partir de las varias expresiones
sencillas y elementales que son las series de desarrollo para cada dos CD, se
obtiene la expresión relativamente compleja que es la serie de combinación
cuyos términos representan los descendientes del híbrido para varios pares de
CED, trátese concretamente de la operación ordinaria de multiplicar “n”
trinomios que sean, la cual da como producto un polinomio de “3n ” términos.
<<.. (A+2Aa+a) X (B+2Bb+b) = para un híbrido con dos pares de CED, y
(A+2Aa+a) X (B+2Bb+b) X (C+2Cc+c) = para un híbrido con tres pares de
CED...>>
Lo que se acaba de explicar acerca del enunciado subrayado por Mendel, <<.....
die Nachkommen der Hybriden, in welchen mehrere wesentlich verschiedene
Merkmale vereinigt sind, stellen die Glieder einer Combinationsreihe vor, in
welchen die Entwicklungsreihen für je zwei differirende Merkmale verbunden
sind. Damit ist zugleich erwiesen, dass das Verhalten je zweier differirender
Merkmale in Hybrider Verbindung unabhängig ist von den anderweitigen
Unterschieden an den beiden Stammpflanzen.>>,
<<..los descendientes de los híbridos en los cuales están reunidos varios caracteres
esencialmente distintos representan los miembros de una serie de combinación en la
cual están reunidas las series de desarrollo de cada dos caracteres diferentes

[41]
[contrapuestos]. Con esto resulta probado, al mismo tiempo, que el comportamiento de
cada dos caracteres diferentes en combinación híbrida es independiente de los demás
diferencias en ambas plantas genitoras.>>
se resume proponiendo la siguiente versión más explícita del mismo
enunciado:
“cualquier generación hija por autopolinización de una planta híbrida para
varios caracteres esenciales distintos, (para varios pares de caracteres entre sí
diferentes, A, a; B, b; y C, c; etc.), pertenece a un tipo de conjuntos de plantas,
el cual esta expresado y definido, tanto en su composición cualitativa como en
su composición cuantitativa, por el polinomio producto que resulta de efectuar
la multiplicación de los trinomios (A+2Aa +a) * (B+2Ba +b) * (C+2Cc +c) =
.................”.
Por último Mendel deduce, que de esta serie combinatoria, los varios pares de
CED de un híbrido, son independientes cuando obtenemos los hijos de este.
Esta relación de independencia es, sin duda, lo más importante de cuanto
afirma el enunciado subrayado por Mendel
<<..... die Nachkommen der Hybriden, in welchen mehrere wesentlich
verschiedene Merkmale vereinigt sind, stellen die Glieder einer
Combinationsreihe vor, in welchen die Entwicklungsreihen für je zwei
differirende Merkmale verbunden sind. Damit ist zugleich erwiesen, dass das
Verhalten je zweier differirender Merkmale in Hybrider Verbindung unabhängig
ist von den anderweitigen Unterschieden an den beiden Stammpflanzen....>>.
<<…..los descendientes de los híbridos en los cuales están reunidos varios caracteres
esencialmente distintos representan los miembros de una serie de combinación en la
cual están reunidas las series de desarrollo de cada dos caracteres diferentes

[42]
[contrapuestos]. Con esto resulta probado, al mismo tiempo, que el comportamiento de
cada dos caracteres diferentes en combinación híbrida es independiente de los demás
diferencias en ambas plantas genitoras…>>
La Genética considera esta relación desde luego como una de las leyes
mendelianas de la herencia; la ley de la independencia de los caracteres
mendelianos. Sin embargo, como es sabido, tal relación de independencia, y,
por lo tanto, tal ley de independencia de los caracteres, carece de validez
experimental en general, ya que es el reverso de la existencia o validez
experimental para las relaciones o “leyes” de ligamiento y recombinación
genética.
Y por ultimo podemos citar otra de las hipótesis inductivas propuestas por
Mendel, y que en ella se define de modo claro lo que hoy llamamos genotipo y
lo que llamamos fenotipo. Baste decir que lo que hoy llamamos genotipo viene
definido por una relación matemática, la famosa relación 2:1:1 que propone
Mendel observando la F3 los descendientes por autofecundación de la F2
<<..Das dominirende Merkmal kann hier eine doppelte Bedeutung haben,
nämlich die des Stamm-Characters oder des Hybriden-Merkmales. In welcher
von beiden Bedeutungen dasselbe in jedem einzelnen Falle vorkommt, darüber
kann nur die nächste Generation entscheiden. Als Stamm-Merkmal muss
dasselbe unverändert auf sämmtliche Nachkommen übergehen, als Hybrides-
Merlkmal hingegen ein gleiches Verhalten wie in der ersten Generation
beobachten...>>
<<..El carácter dominante puede tener aquí un doble significado, a saber; la de
carácter paternal o la de carácter híbrido. Acerca de cual de ambas

[43]
significaciones tenga el carácter en cada caso particular, sólo puede permitir
decidir la generación siguiente…………..>>
<<..Das Verhältniss 3:1, nach welchem die Vertheilung des dominirenden und
recessiven Characters in der ersten Generation erfolgt, löst sich demnach für
alle Versuche in die Verhältnisse 2:1:1 auf, wenn man zugleich das dominirende
Merkmal in seiner Bedeutung als Hybrides Merkmal und als Stamm-Character
unterscheidet. Da die Gliedcr der ersten Generation unmittelbar aus den Samen
der Hybriden hervorgehen, wird es nun ersichtlich, dass die Hybriden je zweier
differirender Merkmale Samen bilden, von denen die eine Hälfte wieder die
Hybridform entwickelt, während die andere Pflanzen gibt, welche constant
bleiben, und zu gleichen Theilen den dominirenden und recessiven Character
erhalten.>>
<<.. La razón 3:1, según la cual ocurre la distribución del carácter dominante y del
recesivo en la primera generación, se descompone, por tanto, para todos los
experimentos en la relación 2:1:1, cuando el carácter dominante se distingue, según
su significación, como carácter híbrido y como carácter parental. Puesto que los
miembros de la primera generación se producen inmediatamente de las semillas de
los híbridos, es pues evidente que los híbridos de cada dos caracteres diferentes
producen semillas, de las cuales una mitad desarrolla nuevamente la forma híbrida,
mientras que la otra da plantas que permanecen constantes y que en igual número
reciben respectivamente el carácter dominante y el recesivo.>>
<<.Si se reúnen los resultados de todos los experimentos, -como
Mendel afirma- entre el número, de formas con el carácter dominante
y el de formas con el carácter recesivo, resulta como proporción media
2´98:1, o sea, 3:1. ..>> y sigue diciendo Mendel <<…El carácter dominante
puede tener aquí un doble significado, a saber; la de carácter paternal o
la de carácter híbrido. Acerca de cual de ambas significaciones tenga el
carácter en cada caso particular, sólo puede permitir decidir la
generación siguiente..>>
Esto es, el carácter de una forma de planta, respecto de la rugosidad y liso de
las semillas u otro de los seis CDs utilizados, puede ser paternal o híbrido esto
es un carácter que se tiene que definir en función de la descendencia de esa
planta (por autopolinización).Y no decimos autógamia ya que Mendel jamás

[44]
tenia información del concepto ni del significado de gametos.
Por lo tanto, el “genotipo” de una semilla lisa de la planta hibrida puede ser
homocigótico, esto es, según Mendel, carácter paternal “CP” o heterocigótico,
carácter hibrido “CI”.
Y atribuirle, por tanto, a Mendel la existencia tanto de los gametos femeninos y
masculinos quizás haga más fácil su entendimiento pero es un error e
incorrecto. Lo importante y obligado es reconocer que aun así, Mendel, sin
conocer los cromosomas y la meiosis, distinguió lo que hoy llamamos genotipo
y, además, de forma rigurosa. Craso error que cometen algunos renombrados
genéticos, cuando afirman que el concepto y significado de lo que hoy
llamamos genotipo -Mendel- no lo tenía claro.
3d).Las hipótesis explicativas.
En las proposiciones que vienen a ser el enunciado o enunciados de su propia
hipótesis explicativa, dadas por Mendel, (la que explica las generalizaciones
inductivas/hipótesis inductivas y, mediante éstas, los resultados experimentales
resumidos por ellas) son, y siguen siendo, sin duda alguna, un hito en la
forma de proceder en la ciencia. Para proponer aquella hipótesis
explicativa, que marcó un antes y un después en la forma de investigar en la
ciencia, Mendel habla de varias clases de células-polen (vielerlei Pollenzellen),
CP y de varias clases de células-germen (vielerlei Keimzellen) CG estando
toda clase de células-polen, como asimismo toda clase de células-germen,
perfectamente definida según expone claramente Mendel a lo largo del
discurso explicativo
. <<Die Befruchtungs-Zellen der Hybriden........ Einen wichtigen Anhaltspunct
bietet bei Pisum der Umstand, dass unter den Nachkommen der Hybriden

[45]
constante Formen auftreten, und zwar in allen Combinirungen der verbundenen
Merkmale........>>
<<Las células reproductoras de los híbridos........ En Pisum ofrece un importante
punto de apoyo la circunstancia de que entre los descendientes de los híbridos
parecen formas constantes -y, por cierto, en todas las combinaciones [posibles ]- de
los caracteres reunidos…….>>
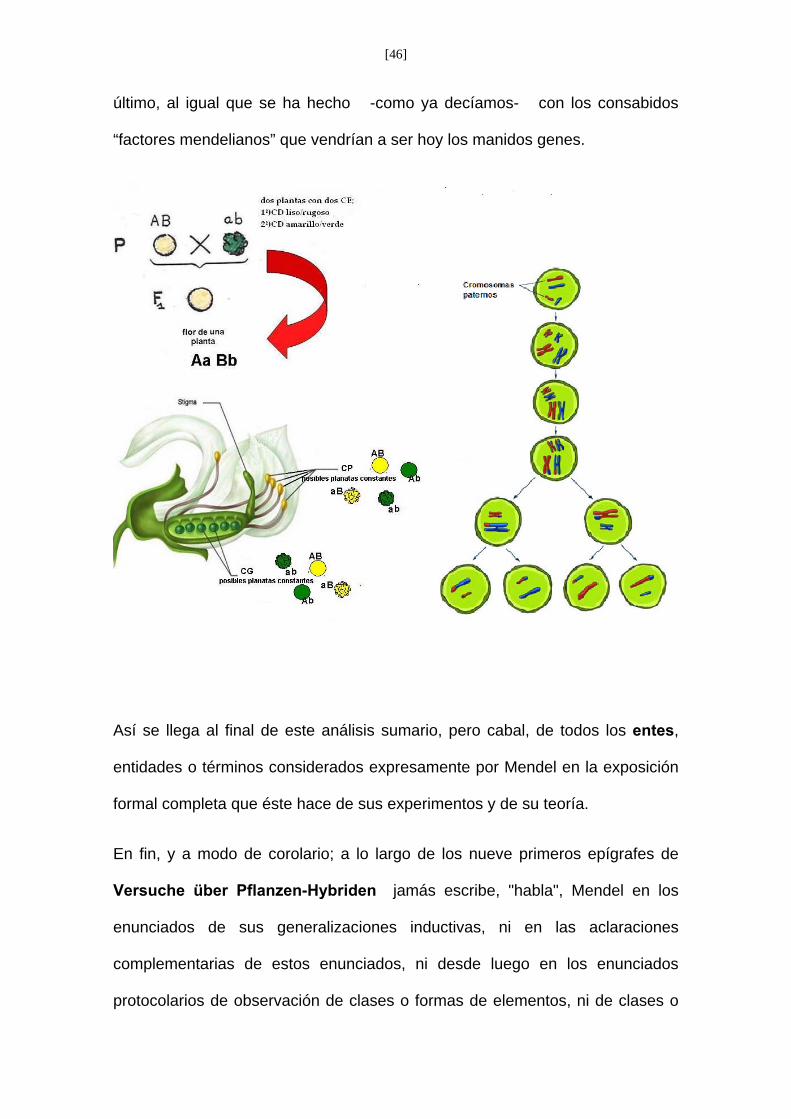
Esto es la CP y CG las define respecto a la clase, forma de plantas, que se
desarrollo del producto -hoy diríamos zigoto- originado por la unión o
fecundación, a su vez, de una célula-polen y una célula-germen. Pues bien, tal
hipótesis explicativa queda planteada con perfecta claridad en el epígrafe del
“Versuche” cuando leemos:
<<Die Befruchtungs-Zellen der Hybriden......Es ist daher auch auf
experimentellem Wege die Annahme gerechtfertigt, dass die Erbsen-Hybriden
Keim- und Pollen-zellen bilden, welche ihrer …. Merkmale hervborgehen……>>
<< Las células reproductoras de los híbridos ....(Es por tanto justificada, por via
experimental, la suposición de que los híbridos) o de esta otra forma (Resulta, pues,
confirmada por vía experimental la hipótesis, de que las plantas de guisante híbridas)
…..forman células-germen y células-polen que, por su constitución, corresponden en
igual número a todas las formas constantes que resultan de la combinación de los
caracteres reunidos por la fecundación>>.
Esta hipótesis explicativa única en la historia de la ciencia, podría ser
interpretada, como se ha hecho numerosas veces para algunas hipótesis
inductivas, como que Mendel propuso el importantísimo proceso de la meiosis.
Pero no menospreciemos a Mendel, atribuyéndole el haber propuesto el
proceso biológico de la meiosis porque coincida en muchos conceptos con este
[46]
último, al igual que se ha hecho -como ya decíamos- con los consabidos
“factores mendelianos” que vendrían a ser hoy los manidos genes.
Así se llega al final de este análisis sumario, pero cabal, de todos los entes,
entidades o términos considerados expresamente por Mendel en la exposición
formal completa que éste hace de sus experimentos y de su teoría.
En fin, y a modo de corolario; a lo largo de los nueve primeros epígrafes de
Versuche über Pflanzen-Hybriden jamás escribe, "habla", Mendel en los
enunciados de sus generalizaciones inductivas, ni en las aclaraciones
complementarias de estos enunciados, ni desde luego en los enunciados
protocolarios de observación de clases o formas de elementos, ni de clases o

[47]
formas de factores, ni mucho menos de elementos o de factores. Así que la
suposición de la existencia de las CP y CG, que como ya hemos dicho, son
corpúsculos específicos que siendo partículas del organismo actúan dentro de
éste condicionando la aparición de caracteres hereditarios observables, y que
pasan de los genitores a los hijos, es una suposición desde luego compatible
con los resultados experimentales obtenidos por Mendel, y que por otra parte,
forman parte principalísima de las hipótesis explicativas. No debe ser
interpretado, con el craso error de los que hoy llamamos factores mendelianos.
Pues, repetimos, que tal suposición no es correcta, pues Mendel no tenía
conocimiento de los cromosomas, ni de la mitosis ni de la meiosis. Todavía
más, nos atreveríamos a afirmar, que hoy no harían falta ni resultarían
necesarios mientras no se trate de explicar el fenómeno de ligamiento y
recombinación. La cuestión no es sólo que Mendel no haga tales afirmaciones
(acerca de la existencia de corpúsculo-partícula de las células-polen y de las
células-germen los cuales sean factores hereditarios o genes), sino que ni
entonces, ni hoy día, con referencia a los resultados experimentales obtenidos
por Mendel procede en rigor hacer tales afirmaciones.
Después de haber indicado los diversos y graves malentendidos de tantos
autores a la hora de interpretar a Mendel, creo que es obligado ofrecer algo
obviamente más objetivo que la mera expresión de nuestras opiniones,
exceptuando la afirmación comprobable, ya que es un posible hecho
experimental, de que no existe en toda la monografía de Mendel, ni una sola
vez, la palabra factor, para poder concebir, intuir, deducir que esa posible
palabra pudiera tener el posible concepto que en el sentido de partícula, de
tamaño finito y ubicada en un lugar concreto (que hoy lo concebimos como

[48]
gen). La única vez que Mendel emplea la palabra “factoren” es con el concepto
de circunstancia. Y esto lo encontramos en:
<<...die befruchtungs-zellen der hybbryden. díe resultate,..gleiche factoren
zusammenwirken...>>;
que es traducido equivocadamente en la mayoría de las traducciones
españolas, las cuales son, a su vez, realizadas de una traducción inglesa
efectuada por Horticultural Society of London.
<<...the reproductive cells of the hybrids. The results of the previously…that
exactly similar factors must be at work also in production of the constant forms
in hybrid plants…>>
<<... las células reproductoras de los híbridos. Los resultados a que llevaron los
precedentes experimentos... que considerar como necesario que también en la
producción de formas constantes a partir de planta híbrida cooperen factores
(ambientales, circunstanciales) completamente iguales...>>
Al mismo tiempo, existe en la monografía de Mendel, una parte que
actualmente es válida para el orden científico moderno, y repetimos una parte
que, ¡además!, es actualmente valida todavía, por cierto, de la obra de Mendel,
sin la mediación de interpretaciones adulteradas. Todo esto cabe hallarlo
magníficamente reunido en los cinco párrafos (522 palabras) con el título:
<< die weiteren Generationen der Hybriden... Bezeichnet a das eine der beiden
constanten Merkmale...>>.
<< las siguientes generaciones de los híbridos....Si a denomina uno de los dos
caracteres constantes...>>

[49]
En estos párrafos podemos comprobar cómo Mendel, con un rigor intelectual y
un sentido experimental verdaderamente admirables, se cuida muy mucho de
afirmar, ni siquiera implícitamente, que las letras A y a, que introduce en el
discurso simbolizando con ellas, respectivamente, dos visibles caracteres
contrapuestos (dominante y recesivo), las mismas letras que luego, englobadas
en el famoso trinomio <A +2Aa +a>, para simbolizar lo que llama “formas
constantes de plantas” (forma constante A de plantas, forma constante a de
plantas) y “forma híbrida de plantas” (forma Aa de plantas), y de las cuales
mismas letras hace, ahora uso explícito, para simbolizar respectivamente las
dos células-polen, y a su vez, las dos clases o formas de células-germen, se
cuida muy mucho -repito- de decir en ningún momento que estas letras A y a
simbolizan respectivamente de cualquier manera dos clases de factores o
elementos de índole alguna, y, por tanto, en absoluto, de corpúsculos-
partículas que estuviesen localizados en ningún sitio de éste bionte, Woodger
(1978) el Pisum sativum; ni en las células-polen, ni en las células-germen, ni en
las células productoras de la unión de aquéllas y éstas. Y, esa parte valida, que
apuntábamos antes, no es ni mas ni menos, que la hipótesis explicativa que
propone Mendel, inventada y después comprobada, de las CP y CG que hacen
de los experimentos un paradigma del método científico:
<< für jeden von diesen 4 versuchen wurden an 3 pflanzen sämmtliche blüthen
befruchtet. war die obige annahme richtig, so mussten sich an den hybriden
keim- und pollenzellen von den formen AB, Ab, aB, ab entwickeln, und es
wurden verbunden:>>

[50]
<<… Para cada uno de estos 4 experimentos fueron fecundadas todas las flores de 3
plantas. Si la anterior suposición era cierta tenían que desarrollarse en el híbrido
células-germen y células-polen de las formas AB, Ab, aB, ab, y unirse:…………>>
Comentario sobre las Schluss- Bemerkungen
(Consideraciones finales) de Mendel.
Si la primera parte indicada de Versuche über Plfanzen Hybriden que
comprende los primeros epígrafes de la monografía, es una pieza teórica al
mismo tiempo que experimental, concisa a la vez que suficientemente explícita,
y, sobre todo, eminentemente analítica y formal, la tercera y última parte (que
sólo comprende el epígrafe Schluss-Bemerkungen) es erudita y especulativa,
prolija sin llegar a ser suficientemente explícita, y, sobre todo, primordialmente
sintética y material. Pero de no igual importancia a la primera parte.
Como manifiesta su autor, esta parte que comprende las Schluss-
Bemerkungen, trata de comparar las observaciones hechas en Pisum, con los
resultados a que habían llegado en sus investigaciones las autoridades de
entonces sobre la cuestión, como era Kölrauter, Wichura y Gärtner.
Considerando los distintos tipos y subtipos de híbridos interespecíficos
reconocidos, al parecer, unánimemente por Gärtner y Kólrauter, cree -Mendel-
advertir una diversidad esencial (“einer wesentlinchen verschiedenheit”):
a) híbridos variables (hv) o que no permanecen constantes en sus
descendientes, y
b) aquellos otros que sí permanecen constantes en sus descendientes o
híbridos constantes (hc).

[51]
Con visión certera aplica Mendel la teoría ideada y comprobada por él en sus
experimentos con Pisum para explicar satisfactoriamente, en términos que hoy
diríamos estadísticos, los fenómenos inherentes a los hv: la aparente
irregularidad de las formas observables en los hijos por autofecundación de
estos híbridos; los llamados experimentos de transformación de una especie en
otra mediante fecundación artificial, los cuales refuta Gärtner como los más
difíciles entre los de hibridación. Y así Mendel razona consecuentemente, ante
esta clase de hv:
<<Die Befruchtungs-Zellen der Hybriden........ Einen wichtigen Anhaltspunct
bietet bei Pisum der Umstand, dass unter den Nachkommen der Hybriden
constante Formen auftreten, und zwar in allen Combinirungen der verbundenen
Merkmale........>>
<<Las células reproductoras de los híbridos........ En Pisum ofrece un importante
punto de apoyo la circunstancia de que entre los descendientes de los híbridos
parecen formas constantes -y, por cierto, en todas las combinaciones [posibles ]- de
los caracteres reunidos…….>>
diciendo que los fenómenos inherentes a los híbridos variables, hv, encuentra
explicación satisfactoria admitiendo que el híbrido produce varias clases de
células polen (CP) y las mismas clases que estas, de células germen (CG).
Pero las cosas para Mendel se vuelven más complejas cuando trata de seguir
explicando estos otros fenómenos propios de los híbridos constantes hc. Estos
otros deben de encontrar su explicación admitiendo, ahora, al contrario que
antes, que el híbrido produce una sola clase de CG y CP, y que esta clase,
será además, la misma clase de célula fundamental originaria del híbrido.
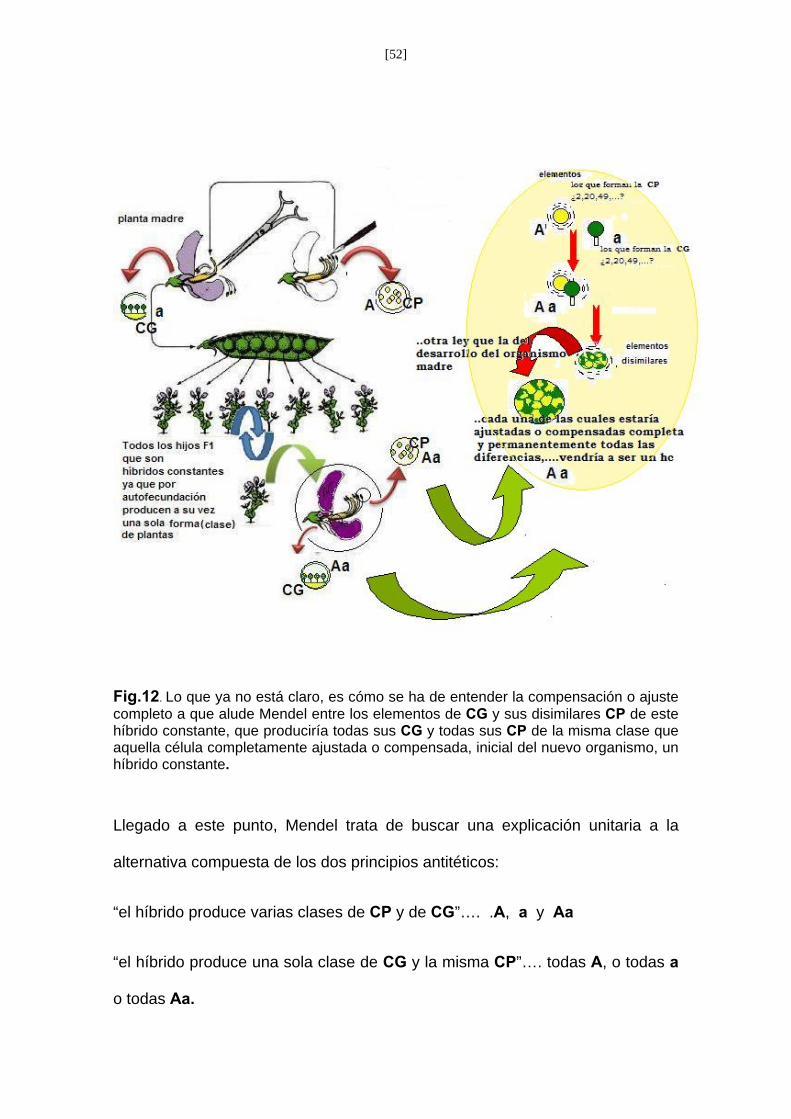
[52]
sss
Fig.12. Lo que ya no está claro, es cómo se ha de entender la compensación o ajuste completo a que alude Mendel entre los elementos de CG y sus disimilares CP de este híbrido constante, que produciría todas sus CG y todas sus CP de la misma clase que aquella célula completamente ajustada o compensada, inicial del nuevo organismo, un híbrido constante.
Llegado a este punto, Mendel trata de buscar una explicación unitaria a la
alternativa compuesta de los dos principios antitéticos:
“el híbrido produce varias clases de CP y de CG”…. .A, a y Aa
“el híbrido produce una sola clase de CG y la misma CP”…. todas A, o todas a
o todas Aa.

[53]
Para ello, apela a los más recientes conocimientos anatómicos y fisiológicos de
aquella época acerca de la fecundación y reproducción de las fanerógamas.
Sugerida por la consideración de que el organismo de la planta hija, se
desarrolla a partir de una sola célula que es, resultado de la unión de una
célula producida por la planta madre y otra producida por la planta padre,
Mendel expone en demasiadas pocas palabras una teoría de los procesos
materiales del desarrollo y de la herencia. En esta exposición aparecen
pensamientos claros y explícitos, otros notoriamente obscuros, cuya carencia
de claridad no cabe suplir mediante el contexto de sus expresiones, y otros
muchos que, sin ser todo lo explícitos que fuera de desear, son susceptibles de
una interpretación o significado claro, si bien no unívoca.
Se dice y entiende claramente, que el desarrollo del organismo cuya célula es
la célula resultante de la reunión de una CG y una CP, está determinada por la
constitución material y por la disposición de unos elementos aportados a
aquella célula por estas otras dos, CP + CG. Así mismo, se dice y entiende con
claridad que, si la CG y CP, que se reúnen originando la célula inicial del nuevo
organismo, son similares entre sí y similares a la que fue célula inicial del
organismo madre, el desarrollo del nuevo organismo hijo, ha de realizarse con
arreglo a la misma ley, según la cual se desarrolló el organismo madre; y que
en el caso contrario, si la CG y CP no son similares, el desarrollo del nuevo
organismo hijo ha de ser -si lo hay- según esto otro, así porque, entre los
disimilares elementos de la CG y CP reunidos en la célula inicial del nuevo
organismo, se habrá llegado a establecer una suerte de arreglo o ajuste viable.
Lo que ya no está claro, o al menos no está suficientemente explicado, es
cómo se ha de entender la compensación o ajuste a que alude Mendel

[54]
calificándolo de completo entre los elementos provenientes de la CG y sus
disimilares provenientes de la CP, reunidos unos y otros en la célula que va a
ser inicial del nuevo organismo; compensación o ajuste completo por el cual,
precisamente, el embrión de este organismo habría de estar formado de
células todas similares y en cada una de las cuales estarían ajustadas o
compensadas completa y permanentemente todas la diferencias, de esos
elementos disimilares y por lo que, en fin, este nuevo organismo vendría a ser
un híbrido constante, que produciría todas sus CG (Aa) y todas sus CP (Aa)
de la misma clase que aquella célula completamente ajustada o compensada
inicial del nuevo organismo, híbrido constante.
Es justo recordar, en este momento, que Tschemak-Seysseneg propuso una
explicación o tesis citomorfologica, la cual por cierto, comprobada
experimentalmente, para explicar los híbridos constantes en los que Mendel,
sin embargo, tuvo que inventarse esa idea/hipótesis tan engorrosa y poco clara,
así como desacertada, de la “compensación y ajuste de los elementos de las
CP y CG”.
Tschemak-Seysseneg en la explicación o tesis que da para tales híbridos
constantes, los hc, es que los gametos producidos por un híbrido F1, genitor
de una F2 es sin segregación cromosomica, esto es, que se asimilen o igualen
a la célula fundamental origen de este mismo híbrido, la teoría llamada de la
“adición de cromosomas” sin reducción; que explica la existencia de los
híbridos constantes como en Aegilotriticum y otros híbridos fértiles constantes.
Pero esto no pudo pensarlo Mendel, ni mucho menos podemos atrivuirselo a
Mendel ya que el no tener conocimiento de la existencia de los cromosomas le

[55]
condujo a dar una explicación mediante la invención de unos elementos que se
ajustaban y compensaban de tal manera que la hipótesis explicativa de que:
<<Die Befruchtungs-Zellen der Hybriden......Es ist daher auch auf
experimentellem Wege die Annahme gerechtfertigt, dass die Erbsen-Hybriden
Keim- und Pollen-zellen bilden, welche ihrer …. Merkmale hervborgehen……>>
<< Las células reproductoras de los híbridos ....(Es por tanto justificada, por vía
experimental, la suposición de que los híbridos)
o de esta otra forma ….
<< Resulta, pues, confirmada por vía experimental la hipótesis, de que las plantas de
guisante híbridas) …………………………………………………..forman células-germen
y células-polen que, por su constitución, corresponden en igual número a todas las
formas constantes que resultan de la combinación de los caracteres reunidos por la
fecundación>>
Hoy día todas sus afirmaciones en “Versuche” pueden ser explicadas
experimentalmente de forma más rigurosa y explicita.
Pero urge añadir, en aras al rigor, algo más, que la teoría de la “adición de
cromosomas sin reducción”, que implica toda la precisa y rigurosa teoría
cromosómica, no contradiga las expresiones de Mendel que se discuten es una
cosa y otra que Mendel predijese, propusiese la “adición de cromosomas sin
reducción” de Tschemak-Seysseneg. Pues si no llegaremos a esas
confusiones, tan generalizadas, de atribuirle a Mendel cosas, conceptos,
experimentos, cta. Como la machacona idea que tienen, no solo los biólogos,
sino muchos genéticos sobre los famosos e inventados factores mendelianos,
y que a Mendel ni los necesito ni los concibió.

[56]
Pues bien, para finalizar, los Schluss-Bemerkungen, en el párrafo que Mendel
dedica a los híbridos variables, exponiendo su teoría de los elementos
celulares como substrato de la herencia y determinantes del desarrollo, es, sin
el menor género de duda, el más importante de todos los párrafos relativos a
esta teoría suya. Y es el más importante, el párrafo aludido, no sólo porque
desde luego atañe al caso de Pisum, cuya teoría o explicación formal ya ha
sido dada y comprobada por Mendel como lo más substancial de sus
investigaciones, sino porque es en este párrafo de su teoría de los elementos
celulares donde la expresión resulta más significativa acerca de cómo
intervengan éstos en el organismo. Toda conexión que se quiera buscar entre
la teoría de Mendel de los elementos celulares y las leyes formales
comprobadas experimentalmente por él mismo en Pisum, se encuentran
concentradas en este párrafo al que se viene aludiendo, y, aun más, en estas
dos oraciones:
<<... Bei der Bildung dieser Zellen betheiligen sich alle vorhandenen Elemente in
völlig freier und gieichmässiger Anordnung, wobei nur die differirenden sich
gegenseitig auschliessen. Auf diese Weise würde die Entstehung so vielerlei
Keim- und Pollenzellen ermöglicht, als die bildungsfähigen Elemente
Combinationen zulassen...>>.
<<...En la creación de éstas células toman parte en un orden libre y regular todos los
elementos existentes, todos los elementos participan en orden completamente libre y
equiproporcional en la formación de las células de fecundación dónde sólo los
elementos diferentes se excluyen recíprocamente ….>>
Tratemos de entender a Mendel con la tozuda idea de que Mendel concebia los
factores mendelianos, no ya con los caracteres, sino ahora con los tales
elementos que alude, en este caso, para tratar de explicar los hibridos

[57]
constantes. ¿Qué se ha de entender exactamente por “<<..todos los elementos
participan en orden completamente libre y equiproporcional en la formación de
las células de fecundación..>>” ?. Quizás podríamos razonar que la
probabilidad de que un elemento x participe en la formación de cualquier célula
de fecundación sea igual -gieichmässinger Anordnung- a la probabilidad de
que participe un elemento y (x e y son cualesquiera dos elementos); y que si el
número que mide esta probabilidad es p y si el número total de elementos es n,
entonces las probabilidades de las distintas formas de células de fecundación
(según participe o no en ellas cada elemento) sera igual a los términos del
desarrollo <<…völlig freier… Anordnung…>> de (p+q)n; siendo q = (1-p)
la probabilidad de que un elemento no participe en la formación de cualquier
célula de fecundación. Así que mediante esta interpretación podría confirmarse
con lo que mucho más claramente dice/escribe después Mendel, cuando afirma
que
<<.. Nur die unterschiedlichen Merkmale schliessen sich gegenseitig aus…>>
<<..solo los elementos diferentes se excluyen recíprocamente..>>.
Tan claramente, por cierto, que apenas necesita aclaración, la cual, sin
embargo, hela aquí: si un elemento x condiciona un carácter, y un elemento y
condiciona lo que Mendel llama el carácter diferente de aquel, entonces, en
toda célula de fecundación en la que participe el elemento x no participa el
elemento y. Sin embargo, a pesar de esta aparente confirmación, la
interpretación antes insinuada no resulta admisible por cuanto de ella se
sigue el absurdo de que se podrían formar algunas células de fecundación sin
elemento alguno para uno, varios, o inclusive todos los caracteres. Todo esto

[58]
para cada dos elementos que condicionan respectivamente un carácter
diferente entre sí, ya que si solo los elementos diferentes se excluyen
recíprocamente, los elementos no diferentes no se excluyen entre sí. El que
participen en la formación de la célula de fecundación producida por el híbrido,
todos aquellos elementos reunidos en la célula fundamental del híbrido de los
cuales ninguno sea condicionante de un carácter diferente del condicionado
por otro, es cabalmente contrario a la idea esencial acerca de los genes, que
consisten, primero, en admitir que los genes existen en la célula fundamental
del híbrido -como asimismo en la célula fundamental de todo individuo- por
pares, ya sean estos pares homogéneos (esto es, constituidos por genes
condicionantes ambos de un mismo carácter), ya sea heterocigóticos; y
segundo, en que uno cualquiera, pero sólo uno de los dos elementos de cada
par, participe en la formación de toda célula de fecundación.
En ninguno de éstos, ni de los demás periodos en que Mendel trata de su
teoría de los elementos, es bastante explícito para permitir al lector descartar
que el autor está pensando más concretamente en que lo que existe, en una
célula de fecundación o en la célula fundamental relacionada con un carácter
observable en el organismo, no es tan sólo un elemento o un par de elementos,
sino muchos más elementos que uno o dos, una multitud de ellos. Esta idea de
que cada carácter observable en el organismo tenga relacionados con él
muchos elementos, es consistente -con el propósito de dar explicación
material a las leyes formales comprobadas en Pisum- con la otra idea de
Mendel que cualesquiera dos elementos existentes en célula fundamental que
condicionen, o intervengan en el condicionamiento, de dos caracteres entre sí
diferentes, se excluyen al participar en la formación de cualquier célula de

[59]
fecundación, mientras todos los demás elementos existen en la célula
fundamental participen en tal formación de modo completamente libre y
proporcional.
Notemos en cambio, que la idea de muchos elementos por cada carácter
condicionando a éste y excluyendo, al momento de participar en la formación
de cada célula de fecundación, a otros muchos elementos condicionantes del
carácter diferente de este mismo, es una idea que contradice esencialmente la
identificación de tales elementos con los genes actuales o los factores
hereditarios de los primeros tiempos de la Genética postmendeliana e
inventados ad hoc para dar mayor amplitud a las teorías de Mendel.
Así que, Mendel no creó el concepto de factor hereditario, y jamás tuvo la más
mínima idea clara sobre este concepto de factores, o pares de elementos -
homogéneos o heterogéneos-, tal que en cada fecundación de la célula
fundamental del individuo, se recompusiesen -recomponiéndose como pares
homogéneos aquellos que vinieren a condicionar caracteres constantes en este
individuo, y como heterogéneos aquellos otros que vinieren a condicionarlos
híbridos- y, en cambio, al formarse las células de fecundación, se
descompusiesen pasando al azar uno cualquiera, pero sólo uno, de los dos
elementos de cada par a cualquier célula de fecundación (células germen o
células polen) que se forme.
Mendel -es lógico- no habría tenido que formular el más explícito, de todas las
explicaciones dadas por él, acerca de su teoría de los elementos, que venimos
discutiendo. Más aún, cuando dice “….que solo los elementos diferentes se
excluyen recíprocamente…” está claro que estos tales elementos no pueden

[60]
tener ni el más mínimo atisbo a algún carácter de los utilizados por Mendel.
¿Como si no los individuos híbridos?, que tanta importancia tienen en los
trabajos experimentales de Mendel. No cabe duda que la interpretación que ha
venido dándose a los trabajos experimentales de Mendel, se han infravalorado,
al tratar de buscar esa vulgar idea de la necesaria existencia de una “cosa”,
factor hereditario, trozo del ADN, gen, etc. Mendel no necesito el concepto
de lo que hoy llamamos gen, para crear la base de la Genética.

[61]
Hacia un riguroso concepto de Fenotipo y Genotipo;
El objetivo principal de la lengua es que "fundamentalmente... sirva como un
instrumento de comunicación" (Ducrot y Todorov 1974 : 29) y no sea una obra
de arte. Esto es, que después de la realización de los complejísimos y
delicados experimentos que realizamos en la investigación científica tras la
búsqueda de los ansiados datos protocolarios, estos, y estos precisamente, los
tenemos que convertir, mediante el lenguaje, en palabras escritas para ser
dados a conocer. Es lo que en el Método Científico ahora serán las
generalizaciones inductivas, los mal llamados hechos generales, así como las
posibles hipótesis explicativas de dicha investigación científica.
El lenguaje, en la Ciencia, no es un fin en sí, sino un medio para un fin. Este es
el motivo principal/ fundamental por el cual los significados de algunas palabras
deben ser escrupulosamente rigurosas, en cuanto estas traten de describir algo
concreto, y no encontrarse en función de las interpretaciones personales o del
contexto de la oración gramatical. Quizás sería en este último caso cuando el
análisis lingüístico tendría cabida. Por eso, no hay que confundir la búsqueda
de los conceptos y definiciones de los significados rigurosos y claros, de las
palabras en el leguaje escrito o coloquial que utiliza la Ciencia, con el objetivo
que persigue el análisis lingüístico, que es el estudio científico de las palabras
en el contexto gramatical de esa lengua, ya que, sin duda alguna, el análisis
lingüístico principalmente se ocupa de descubrir y entender la naturaleza y las
leyes que gobiernan el lenguaje. Esto es, estudia sistemáticamente y con
mucho detalle las estructuras del discurso como objeto verbal (texto,

[62]
argumentación, narración), como los temas, la coherencia local y global,
los pronombres, el estilo, etc.
Pues bien, dicho esto, nosotros aquí hablaremos del significado de unas
palabras que tratan de describir ciertas cosas que necesitamos identificar para
poderlas distinguir en la praxis de la Ciencia, y que en nuestro caso es el
concepto riguroso del fenotipo y genotipo de un bionte (organismo vivo J. H
Woodger. 1978.); rigor que casi debe rayar al matemático y que debería
exigirse cuando se tratan los temas científicos. ¿A quién se le ocurriría que con
el lenguaje coloquial al tratar definir a la par de dar un concepto, como así se
hace, del signo esdivagaren cuestiones lingüísticas? Ni mucho menos, su
significado ha sido definido, de forma explícita, así su concepto; aunque, unas
veces de manera más clara que otras; según el autor que lo exponga.
Además, debemos recalcar que el fenotipo y genotipo no tienen ni deben tener
su significado según el entorno lingüístico en que estas palabras se utilicen,
sino que ellas mismas deben tener su significado y concepto de forma objetiva
y rigurosa. Si acaso, su explicación y descripción serán las que deben ser
acompañadas de las expresiones lingüísticas que expresen y lo expliquen con
más acierto y rigor.
Urge decir de nuevo, aunque seamos recurrentes, que una característica
esencial, primordial, del lenguaje, al transformarse en algo de carácter utilitario
para la Ciencia, es que las palabras han de ser precisas para aquello que
tratan de explicar y no prestarse a confusión alguna. Se necesitan definiciones
exactas y cabales acatables por todos los lectores y, más particularmente en
este caso, por los eruditos de la Genética y por la comunidad científica, de
manera que no den lugar a conflictos o discusiones interminables en sus

[63]
respectivos campos científicos, posibilitando así que el uso práctico del
lenguaje sea una herramienta para el conocimiento científico. Bühler expone
que el lenguaje científico obedece a intenciones pragmáticas o a servicios para
el hombre; esto es, la función de informar, o de transmitir conocimiento de
forma objetiva, por medio de la función representativa que se funda en uno de
los factores de la comunicación; el mensaje.
Por tanto, retomando nuestro primordial objetivo, podríamos decir que los
términos genotipo y fenotipo forman parte de los significados más elementales
de la Biología. Sin embargo, y a pesar de ser términos muy utilizados, podemos
encontrar tantas definiciones como libros de Genética se publican. Y, sobre
todo, en cuanto nos fijemos en el significado y la explicación de este, esto es,
del concepto que se da de fenotipo. Así pues, es necesario hacerlo, aunque
somos conscientes de lo muy arriesgado que es dar un concepto claro y
riguroso del significado, y definición, exacto -desde la Genética- de estos
términos y no volver a cometer los errores que uno trata de evitar.
Para ello partiremos de distintos conceptos de los significados dados por los
estudiosos y científicos que han tratado de forma rigurosa qué es
lo que se entiende por fenotipo y genotipo. ¡He aquí algunas!, entre los
numerosísimos conceptos de los significados de fenotipo y genotipo:
A Dictionary of Genetics: Seventh Edition. ROBERT C. KING (Emeritus Professor, Northwestern University) WILLIAM D. STANSFIELD (Emeritus Professor, California Polytechnic State University ). PAMELA K. MULLIGAN. 2006. ISBN-13 978-0-19-530762-7; 978-0-19-530761-0 Phenotype: he observable characteristics of a cell or an organism, such as its size and shape, its meta-bolic functions, and its behavior. The genotype (q.v.) is the underlying basis of the phenotype, and the term is commonly used to describe the effect a particular gene produces, in comparison with its mutant alleles. Some genes control the behavior of the organism.

[64]
Fenotipo: las características observables de una célula o un organismo, tales como su tamaño y forma, sus funciones metabólicas, y su comportamiento. El genotipo (q.v) es la base fundamental del fenotipo, término que es comúnmente utilizado para describir el efecto que produce un gen particular, en comparación con sus alelos mutantes. Algunos genes controlan el comportamiento del organismo, …… Recursos on-line para el curso Licenciatura de Biología, UAB. Prof. Antonio Barbadilla. http://biologia.uab.es/base/base.asp?sitio=cursogenetica&anar=concep&item=genfen Fenotipo: La clase de la que se es miembro según las cualidades físicas observables en un organismo incluyendo su morfología, fisiología y conducta a todos los niveles de descripción. Genotipo: La clase de la que es miembro según el estado de los factores hereditarios internos de un organismo, sus genes. http://bioinformatica.uab.es/base/base.asp?sitio=cursogenetica&anar=concep&item=g
enfen
Genetics. A Molecular Perspective. William Sklug and Michael R. Cummings. 2003
ISBN 0-13-008530-8).
Trais such as tall or darwf are physical expression of the information contained in unit factors. We now call the physical expression of a trait the phenotype of the individual. / By contrast, physical appearance of each strain -the tall or dwarf charecteristic- is said o be its phenotype. Los caracteres como alto o bajo son la expresión física de la información contenida en los factores de unidad. Que hoy conocemos como la expresión física de un rasgo del fenotipo del individuo. / Por el contrario, el aspecto físico de cada carácter - la característica de alto o bajo- se dice que es su fenotipo. Principes of Genetics.4ªEd.D.Peter Snustad and Micheel J. Simmons. 2006 ISBN 13-
978-0-471-69939)/
Another important term is phenotype, which is the manifestation or appearance of a characteristic. A phenotype can refer to any type of characteristic – physiological, biochemical, or behavioral. The allelic constitution of each strain is said to be its genotype. Genetics a Conceptual Approach. 3º edition. Benjamin. A. Pierce. 2008 .ISBN -13:
978-0-7167-7928-5.W.H.Freman and Company.
Gene mutations result from smaller changes in the stored chemical information in DNA, making up a part of the organism´s genotype. ….Toda la información contenida en los cromosomas se conoce como genotipo; sin embargo, dicha información puede o no manifestarse en el individuo.

[65]
Wikipedía.
El genotipo: es el contenido genético (el genoma específico) de un individuo, en
forma de ADN.
Biología Molecular del Gen . James D. Watson. 3ºEd.1978. ISBN: 84-03-20-259-8
A la apariencia (configuración física) de un individuo la denominamos su fenotipo y a su constitución genética su genotipo. Individuos con fenotipos idénticos pueden tener genotipos diferentes, de modo que, para determinar el genotipo de un organismo, es muy a menudo necesario realizar cruzamientos genéticos por varias generaciones. ………………………………………………………………………………………….
La “Apariencia” (configuración física) en todas las definiciones la
conceptualizan y definen como el fenotipo de un individuo. Y no podemos por
menos que afirmar de forma rotunda que es la definición más vulgar y la más
confusa que quizás existe en la Biología, y más concretamente, en la Genética.
Pero apresurémonos, antes de seguir con los conceptos y definiciones de los
significados de fenotipo y genotipo, a decir que, por otro lado, el concepto de
fenotipo “lo tenemos por intuición” pero su expresión rigurosa y explícita no. Y
esto lleva y sigue llevando a muchas confusiones en el lenguaje de la Biología
(la Genética).
Casi todas definiciones, y conceptos, que hemos leído están sujetas a un
lenguaje coloquial y, por ende, faltos de rigor científico. Sin embargo en nuestro
objetivo, no fácil desde luego, vamos a tratar de con el lenguaje coloquial
podamos hacer lo que hacen los matemáticos, desechar las ideas que
podemos tener o explicar del significado de fenotipo y genotipo. Estas la
mayoría de las veces son subjetivas. Para lo cual lo primero que haremos es
utilizar el concepto matemático de clase. Por tanto, a la luz de estas
definiciones podemos comprobar con qué amplia variedad de términos
empleados (apariencia, individuo, clase, miembro, organismo, forma, tamaño,
carácter, factores hereditarios, aleles, secuencias de nucleótidos,…), puede

[66]
darse un/el concepto de fenotipo y genotipo, y, por lo tanto, parecer complejo
aportar/dar una definición única para explicar qué es fenotipo y genotipo para la
Genética. Sin embargo, siendo conscientes de su complejidad, ese es nuestro
propósito. Pues bien, ¡ lancémonos!,…a tal ardua tarea.
He aquí nuestra forma de proceder: utilizar principalmente argumentos
matemáticos, al más puro estilo de Mendel. Entonces podemos proponer un
primer borrador sin temor a equivocarnos:
… el fenotipo es un intervalo de valores de una magnitud relativa de una parte
independiente o no, de un organismo. Cada valor de esta magnitud se da en un
ente orgánico (individuo), o bionte, como así lo designaba J.H Woodger
(Bilogía y lenguaje. ISBN: 84-309-0738-6). Y todos estos, los valores, que son
de una misma magnitud se dan respectivamente en partes homólogas de otros
individuos. Los intervalos que son los fenotipos, deben ser, o mejor dicho,
conviene que sean, sin duda, intervalos abiertos. Así, entonces, un fenotipo
es un intervalo abierto de valores de una magnitud. Esto es, que los valores
que acotan por el máximo y mínimo no entren como valores numéricos de esas
magnitudes, figura 1

[67]
Fig.1. El fenotipo rojo de los ojos es un intervalo abierto de valores de una magnitud; el color rojo de los ojos de la Drosophila. Cada valor de esta magnitud se da en la Drosophila y todos los valores del color rojo (una magnitud) se dan respectivamente en partes homólogas de otras Drosophilas.
Según lo que hemos planteado, un fenotipo es un intervalo abierto de
valores de una magnitud. Esta magnitud es, o extensiva (longitud, área,
volumen, etc.) o intensiva (color, peso, sonido, etc.). Y cada valor de esta
magnitud se da en un individuo como magnitud de una parte. Podríamos definir
entonces el fenotipo como un intervalo abierto de valores de una clase de
magnitud intensiva o extensiva referente a una cualidad o carácter de un
individuo, figura .2

[68]
Fig.2. El fenotipo es un intervalo abierto de valores de una magnitud intensiva o extensiva referente a una cualidad o carácter; el color rojo y el tamaño de las alas
Por tanto, como un fenotipo es una clase de magnitud, todo individuo que
pertenezca o que posea esta clase de magnitud, pertenecerá a ese fenotipo.
Pero esta nueva palabra, “clase”, que ha aparecido en la explicación del
significado (N.E.Christensen.1968) de fenotipo, que hemos propuesto, es un
concepto matemático, y a su vez un significado lógico de suma importancia.
Clase se ha definido a veces como una serie, grupo, colección o conjunto de
entidades que poseen por lo menos una característica común. Ejemplos de
clases pueden ser: la clase de los hombres, la clase de objetos cuya
temperatura en estado sólido es inferior a 10ºC, la clase de palabras que
empiezan con la letra “C” en esta página, et.
Aun cuando “clase” es un significado lógico matemático, las clases son
entidades abstractas, a pesar de que los miembros de que se componen son

[69]
entidades concretas. Una clase o colección puede ser definida de dos
maneras: se pueden enumerar sus miembros “la clase, la colección, a que me
refiero es Juan, Pedro y Diego”. O se puede mencionar una propiedad que la
defina, como al hablar de la “humanidad”, de los “hombres de España” o la
temperatura de un grupo de bolas, los guisantes de color verde. La definición
que enumera se llama definición por extensión, y la que menciona una
propiedad que la define se llama definición por intensión. (Beltrand Russell),
figura 3.
Fig. 3. Una clase que se define mediante la enumeración de los miembros se llama definición por extensión, y la que menciona una propiedad que la define, se llama definición por intensión. (Beltrand Russell)
Por tanto, las definiciones del fenotipo como una “apariencia externa…”,
además de ser un craso error, no mantienen el rigor científico necesario como
para ser consideradas correctas. Para convencerse de ello basta reflexionar en
que la forma que tiene un guisante –que, supongamos, tengo ahora en mi
mano--, la coloración de él, la pesadez (o ligereza) del mismo, etc., ésta

[70]
apariencia externa, son, ni más ni menos, otros tantos caracteres observables
de dicho objeto que es “el guisante que tengo en la mano y al que estoy
observando ahora”; y así, en cada guisante que pueda colocar en mi mano,
reconoceré su forma, su coloración, su pesadez, etc.: esto son diferentes
caracteres observables de él y no de ningún otro, esto es, su apariencia
externa.

[71]
Fig. 4. El guisante que tengo en la mano y así cada guisante que pueda colocar en mi mano, reconoceré su forma, su coloración, su pesadez, etc., caracteres observables, como de él y no de otro ninguno.
Estos caracteres observables, por otra parte, no podré cogerlos con mis
manos uno por uno, pero sí que puedo verlos o sentirlos. No puedo coger la
coloración del guisante, pero puedo verla, la veo, estoy viéndola ahora;
tampoco puedo coger la forma del guisante, pero no solo puedo verla también,
sino, además, “darme cuenta” de ella tocando el guisante, haciéndolo rodar
entre las yemas de mis dedos; tampoco coger la pesantez, pero sí sentirla
teniendo al guisante sobre mi mano, etc. Por tanto, no podemos dudar de la
objetividad, de la realidad física de estos caracteres observables o, más
rigurosamente expresado, caracteres singulares.
Pero, ¡démonos cuenta de una cosa!, y esto sí podría ser competencia del
Análisis Lingüístico, como decía el eminente Prof. Ayala en una conversación

[72]
epistolar con él mantenida acerca del concepto de fenotipo <<…..los capitulos …
son muy interesantes, pero escritos dentro de un contexto intelectual llamado en ingles
linguistic analysis, como usted sabe,…..>>. Y es cuando le pidiéramos al lector que
compare el sentido que hemos hecho anteriormente de la palabra carácter,
llamándolo carácter singular, con estas otras expresiones que haremos ahora:
“todos estos guisantes que veo tienen un mismo carácter, que es “color
amarillo de los cotiledones”; todos estos guisantes que veo tienen un mismo
carácter, que es “su rugosidad de los cotiledones”,..etc. Advertiremos, como
evidente, que el sentido de la palabra carácter es completamente diferente
ahora que antes. Unas veces, la palabra carácter singular designa algo
determinado para ese guisante, que se ve perteneciendo a un guisante nada
más (cuál es la forma del guisante, la pesantez, la coloración de este guisante
y no de ningún otro), y, otras veces, como en estas frases a las que nos
referimos últimamente donde la palabra carácter designa algo que decimos
que vemos de forma general; que está siendo lo mismo en un guisante que en
otro cualquiera. Y es lo que podríamos llamar el carácter común. Aunque
propiamente ver, solo se conseguirá ver el carácter singular, peculiarísimo y
exclusivo de un guisante que es “la coloración de este mismo”, y así de cada
uno que se mire. Pero aun así, tampoco con la imaginación se consigue más
que representar uno o muchos colores, extendidos sobre guisantes imaginarios
o sobre fantásticas figuras cualesquiera, pero todas tan singulares, aunque
mucho más borrosas y confusas, que como los guisantes realmente existentes,
ya que el carácter utilizado ahora sería el carácter común, que ya no es un
ente real objetivo ni tampoco imaginario, sino, en verdad, una abstracción de la
realidad. Pues bien, habiendo hecho este posible alarde de análisis lingüístico,

[73]
como sugiere el Prof. Ayala, el carácter común podríamos decir que viene a
ser el concepto de fenotipo.
Y mucho más, a lo que se designa mediante la expresión “carácter
observable” hay dos aspectos: uno más teórico o ideal, y otro, más práctico.
Este doble aspecto, ideal por un lado, real por el otro, no es privativo de los
caracteres observables. En efecto, lo mismo que sucede con los caracteres
observables, pasa con las medidas. Varios individuos pueden tener un mismo
carácter observable en cada uno de ellos, como pueden tener una misma
medida practicable también en cada uno de ellos; y nada más abstracto o ideal
que la expresión numérica de una medida, ni tampoco nada más
decisivamente práctico y experimental que la obtención o verificación de la
misma medida. La comparación entre caracteres observables y medidas
todavía revela más analogías entre aquellos y estas, y así, permite aclarar
mejor ciertas confusiones aparentes, de origen más bien lingüístico o
gramatical, en relación con aquellos
.

[74]
Fig.5 Nada más abstracto o ideal que la expresión numérica de una medida, ni tampoco nada más decisivamente práctico y experimental que la obtención o verificación de la misma medida. El carácter = el fenotipo; esto es, como la medida no forma parte del objeto de estudio.
Un carácter de un organismo no puede ser una parte de organismo, como
tampoco lo es la medida de una parte de él, figura 5. El carácter como la
medida no forma parte del objeto de estudio. Es una correspondencia univoca
entre organismo o partes del organismo, como entidad real y las palabras o
sustantivos, después, claro es, de haber realizado la observación o la medida.
Por tanto, los caracteres y las medidas son completamente ideales, en cuanto
significado.
Por tanto el fenotipo “verde”, de un guisante verde, no es lo que vemos
coloreado de verde en el guisante, este carácter singular, color verde. El
fenotipo que designa con la palabra “verde” es una clase de guisantes.
Realmente, el fenotipo verde que tiene un guisante verde no se ve, ni se puede
ver (como, en general, no se puede percibir con los sentidos fenotipo alguno
que tenga un individuo-organismo cualquiera), sino que, al ver el carácter
verde, lo verde en el guisante se indica, se prueba que el guisante está
comprendido dentro de una clase determinada, la cual es el fenotipo verde.

[75]
Fig.6 No se puede percibir con los sentidos fenotipo alguno que tenga un individuo-organismo cualquiera. El carácter verde, por ejemplo, es una clase determinada donde se reconoce que el guisante esta comprendido,
Así únicamente se concibe que dos o más organismos (dos guisantes, tres
guisantes, dos moscas, etc., el número que sean) puedan tener el mismo
fenotipo: porque el fenotipo no es cosa alguna, no tiene propiedades físicas,
sino que es algo ideal que comprende idealmente a aquellos individuos.
La característica más importante que distingue a la clase de tipos que son los
fenotipos, es la de estar definidos, precisamente, porque cada individuo-
organismo que comprende tenga algo observable o medible, ¡un carácter
observable!, como propiedad física de él mismo. Así, por ejemplo, la forma
geométrica o un color de todo el organismo o de alguna parte de él se
reconoce viéndolo y sólo viéndolo; el valor de masa, de volumen, de longitud se
reconoce pesándolo o midiéndolo y sólo pesándolo o midiéndolo; etc.

[76]
Para concluir, podemos afirmar, como anteriormente ya explicamos con rigor
científico, que el fenotipo es un concepto ideal, como son las medidas y los
caracteres.
Por tanto los fenotipos y genotipos son clases de individuos comprendidos
dentro de una especie. Un fenotipo es la clase de todos aquellos individuos que
tienen algún carácter determinado observable en cada uno de ellos. Un
fenotipo es una clase de magnitud, todo individuo que pertenezca o que posea
esta clase de magnitud pertenecerá a ese fenotipo. El fenotipo es un intervalo
de valores de una magnitud relativa de una parte independiente o no, de un
organismo. Cada valor de esta magnitud se da en un ente orgánico (individuo),
bionte, y todos los valores de una misma magnitud se dan respectivamente en
partes homólogas de otros individuos. Los intervalos que son los fenotipos
deben ser, o mejor dicho, conviene que sean, sin duda, intervalos abiertos. Así,
entonces, un fenotipo es un intervalo abierto de valores de una magnitud (este
carácter puede ser anatómico en unos casos, fisiológico en otros, y hasta
psicológico; pero siempre observable en cada individuo perteneciente al
fenotipo de que se trate). Y es a lo que Mendel describe como formas;
<<.. Las diferentes formas de guisantes elegidas para fecundación mostraron
diferencias en la longitud y coloración del tallo, en el tamaño y forma de las hojas, en
la posición…>> hoy fenotipos.
Y más aun en la famosa relación fenotípica de 3:1 que obtiene en el
experimento que realiza para comprobar la descendencia de los hibrido por
autopolinización. Según Mendel,
<<… Si se reúnen los resultados de todos los experimentos, entre el número de
formas con el carácter dominante y el de formas con el carácter recesivo resulta
como proporción media 2´98:1, o sea, 3:1…>>

[77]
Pero, -¡eh hay!- que el significado de genotipo, va a tener un significado
parecido al del fenotipo, eso ya no es aceptable. Ya no es como el caso del
fenotipo, nos contestaría un erudito de la genética molecular, sino que es
aquello que se hereda. Es, desde un punto químico (para la genética
molecular), toda la información contenida en los cromosomas en la estructura
química inestable del ácido orgánico ADN, suma de numerosísimos
nucleótidos. Son conceptos acríticos hoy día. Pero permítanos el lector que
volvamos a la genética propiamente dicha, que con cierto desdén científico
suelen llamar genética clásica.
Un genotipo, para Mendel y para la Genética, es también hoy día, una clase de
individuos, pero una clase de individuos fundada no en algún carácter
observable en cada individuo perteneciente a la clase (a la forma de planta,
según Mendel) o del fenotipo de que se trate, sino fundada en caracteres
observables en los descendientes o en los ascendientes de cada uno de los
individuos pertenecientes al fenotipo, esto es, fundada en el fenotipo o
fenotipos de los ascendientes o descendientes de cada individuo del cual
definiremos el genotipo (fenotipo de sus descendientes).

[78]
Fig.7. Un genotipo para Mendel es una clase de individuos, pero una clase de individuos fundada no en algún carácter observable en cada individuo perteneciente a la clase (a la forma de planta, según Mendel) o del fenotipo de que se trate; sino fundada en caracteres observables en los descendientes por autopolinización.
Así pues, ni el fenotipo ni el genotipo de un organismo, según la Genética, no
son ningún órgano ni ninguna función ni nada que se asimile a órgano o
función de organismo; son más bien caracteres, aunque tampoco propiamente
esto; son clases de individuos fundadas o definidas en caracteres
observables en los organismos. Y es a lo que Mendel describe como formas;

[79]
<<…..de formas con el carácter dominante y el de formas con el carácter recesivo
resulta como proporción media 2´98:1, o sea, 3:1.El carácter dominante puede tener
aquí un doble significado, a saber; la de carácter paternal o la de carácter híbrido.
Acerca de cual de ambas significaciones tenga el carácter en cada caso particular,
sólo puede permitir decidir la generación siguiente……>>.
Y es cuando Mendel en “Die zweite Generation der Hybriden”( La segunda
generación de los híbridos) sigue escribiendo
<<…La razón 3:1, según la cual ocurre la distribución del carácter dominante y del
recesivo en la primera generación, se descompone, por tanto, para todos los
experimentos en la relación 2:1:1……. de las cuales una mitad desarrolla nuevamente
la forma híbrida, mientras que la otra da plantas que permanecen constantes y que en
igual número reciben respectivamente el carácter dominante y el recesivo..>>
Relación esta 2:1:1, que es sin duda alguna, hoy día, una relación genotípica;
ya que 2 caracteres híbridos y dos caracteres constantes(recesivo y
dominante) son caracteres que no se observan directamente en la planta sino
como dice Mendel
<<... darüber kann nur die nächste Generation entscheiden. Als...>>
<<….solo puede permitir decidir la generación siguiente. >>.
Pero no nos cansaremos de decir que todas estas afirmaciones tienen rigor
científico sin duda alguna siempre que se refieran a esta clase de plantas
autógamas.
¡Y es aquí!, donde reside la monumental claridad de los significados de
Mendel, que supo distinguir de forma clara lo que hoy llamamos fenotipo y
genotipo. Significado este último que con los avances en la genética molecular
se ha vuelto mucho más complejo y también confuso.

[80]
El MÉTODO CIENTÍFICO
Distinción entre método y técnica. El método científico.
No cabe duda de que para saber a qué llamamos hoy día método científico
bastaría con leer los experimentos de Mendel en el Versuche über Planzen-
Hybriden, donde la forma de proceder experimentalmente y teóricamente es
un verdadero paradigma del Método Científico.
Pues bien, quizás el hecho más significativo del desarrollo de la enseñanza de
la Ciencia en los últimos años haya sido el alejamiento del concepto de Ciencia
como un cuerpo de conocimiento acumulado pero ordenado y con objetivos
claros, por un acercarse a estos mediante los puros datos experimentales de
experimentos sin ningún objetivo como método de generación de tal
conocimiento.
Hoy se insiste en la necesidad de enseñar "científicamente" y de empujar a los
alumnos a ser "científicos" frente a los problemas. Tal exigencia implica que
hay algo –en el método científico- que puede ser caracterizado, practicado y
enseñado. Así Bunge escribía; "Una ciencia es una disciplina que utiliza el
método científico con la finalidad de hallar estructuras generales." M. Bunge,
“La Investigación Científica.” Col. Convivium. Ed. Ariel. Barcelona, 1986.
Pues bien, al tener que tratar sobre el método científico, entiendo que importa
sobremanera diferenciar entre método y técnica, antes de precisar el método

[81]
científico, que se puede practicar y enseñar, como decíamos anteriormente,
recordando al hierático Fray Luis de León.
Según el Diccionario de la Real Academia de la Lengua Española, la palabra
“método” posee, además de los significados más generales y corrientes, como
son “modo de decir o hacer con orden una cosa”, “modo de obrar o proceder”,
“hábito o costumbre que cada uno tiene y observa”, el significado más filosófico
de “procedimiento que se sigue en las ciencias para hallar la verdad
y enseñarla”. En cambio, según el mismo diccionario, el vocablo “técnica”
significa “conjunto de procedimientos o recursos de que se sirve una ciencia o
arte”; y, en otra acepción, “pericia o habilidad para usar de esos
procedimientos o recursos”.
Tales significados de “método” y “técnica'’ están conformes con la etimología
de estas palabras. 'Método', a través del latín methodus, que, en cuanto verbo,
significa “caminar hacia”, y, en cuanto sustantivo, “búsqueda” -se
sobreentiende, por lo que atañe al significado de nuestras palabras, que se
trata de “caminar hacia la verdad” o de “búsqueda de la verdad”-. A su vez,
'técnica', mediante el latín technicus, significa “relativo o propio de un arte”, y
también “industrioso, hábil, diestro”.
Si acudimos a un diccionario especialmente filosófico, encontraremos
corroborados con más precisión estos significados. Así, en el de Ferrater Mora,
el amplio artículo dedicado al término 'Método' se puede considerar como “un
programa, pero un programa que se dirige a la obtención del saber, por lo
cual el método debe distinguirse de cualquier otro procedimiento cuyo fin
no sea el saber mismo". En tanto que este mismo autor, Ferrater Mora,
comienza el artículo 'Técnica' diciendo que "Técnica significa

[82]
originariamente arte, es decir, modo de hacer una cosa, procedimiento".
Pero nuestras ansias de discernimiento entre método y técnica las hallamos
colmadas en el "Vocabulario Filosófico" de Goblot. En él leemos, primero, "Un
método es una manera razonada de conducir el pensamiento para llegar a
un resultado determinado, y especialmente para descubrir la verdad”;
segundo, "Un conocimiento técnico es un conocimiento que no está
considerado desde el punto de vista de su valor lógico, pues la prueba se
da ya como hecha, sino desde el punto de vista de su aplicación a algún
fin práctico"; y, en fin, como remate de todo ello, "La técnica de una ciencia
es el arte de ejecutar las operaciones manuales que sus métodos exigen;
el método, al contrario, es un conjunto de operaciones lógicas".
Podriamos aventurarnos a dar un ejemplo con cierta claridad aunque
sacrificando el rigor; como decía el Prof. Russell en sus clases, el rigor por la
claridad es una cantidad constante cuando tratamos de explicar un proceso, un
suceso o un concento de algún significado. Pues he ahí, nuestro objetivo es
evaporar un compuesto sólido. Este compuesto es sólido, sabemos que a 60
ºC se sublima; pasa del estado sólido a vapor a una cierta temperatura, sin
licuarse previamente. Entonces aprovechando esta propiedad química
podemos realizar tal evaporación ya mediante colocarla en un soporte Fig 23 y
calentarla o con un mechero o con una resistencia.
Entonces; el método, es el proceso químico-físico de la sublimación. La
técnica es la distinta forma de aportar calor al compuesto; mediante un
mechero de gas o mediante una resistencia eléctrica.

[83]
Fig.1 Objetivo evaporar el compuesto solido de la capsula de porcelana. Sublimación.
Calor por gas o resistencia eléctrica
Es por tanto, que en los procesos humanos para la realización de una
investigación, a poca experiencia que se tenga de todos los distintos momentos
que juntos completan el proceso entero de la investigación científica de
cualquier cuestión experimental definida hasta el esclarecimiento de ésta, cabe
advertir que existen tres principales ingredientes que intervienen en ella:
1º La actividad humana, esto es, el trabajo de los hombres y de las mujeres
dedicados cabalmente a la investigación científica, cualquiera que sea.
2º El objeto material observable al que esta actividad humana se dirige
inmediatamente por medio del conocimiento sensual, es decir, aunque sea
valiéndose de instrumentos de observación.

[84]
3º El lenguaje natural o vernáculo, así como el artificial o simbólico, con los que
aquellos hombres y mujeres expresan y comunican los resultados obtenidos
(consideraciones, reflexiones, especulaciones) relativos a aquel objeto.
Nada más lejos, esto que acabamos de afirmar, de lo que quizás pueda
parecer un desatino: el hecho de dar exagerada importancia al lenguaje
considerándolo nada menos que como uno de los tres principales ingredientes
de la investigación científica, al lado del factor humano y del objeto real del
conocimiento mismo. Pero debemos no olvidar que la ciencia experimental,
analizándola como fruto ya logrado y desprendido de estos quehaceres, viene
a consistir esencialmente en dos conjuntos de enunciados, esto es, en puro
lenguaje: uno describe cierto estado real de cosas observables muy bien
conocido pero que, (sin embargo, no entendemos, por lo que) al no entenderlo,
creemos que requiere explicación; y otro conjunto de enunciados del que, a
pesar de ser lingüística y gramaticalmente bien inteligible (si acaso inventado
ad hoc por nosotros mismos), desconocemos totalmente su veracidad o
falsedad en cuanto a su significado real. No obstante, suponiendo o admitiendo
desde luego la veracidad de este segundo conjunto de enunciados, resulta
posible dar razón o explicar aquel otro primero de ellos muy bien conocido
experimentalmente, con lo cual el segundo conjunto de enunciados viene
entonces a resultar mediata, pero cabalmente probado o comprobado -nunca
demostrado- como verdadero, acrecentando así nuestro conocimiento de la
realidad.
Pues bien, en tanto en cuanto el quehacer de los hombres y de las mujeres
dedicados cabalmente a la investigación científica se dirige inmediatamente al

[85]
objeto material observable, tenemos ante nuestra consideración la fase
propiamente técnica de la investigación científica experimental (en ella entran
en juego principalmente las técnicas).
El registro de los resultados obtenidos en esta fase técnica, ya sea como
abreviadas anotaciones en un cuaderno, ya como una o varias tablas de
valores numéricos, ya como registros continuos de complejísimos aparatos, ya
como explícitas expresiones verbales, formadas mediante una oración
gramatical o por varias, en cuyos términos están comprendidas e incorporadas
precisamente aquellas breves anotaciones, constituye lo que, desde el punto
de vista del método científico, se llama proposiciones protocolarias o
enunciados de observación (cuya característica esencial es la de ser
propiamente verdaderos o falsos, en virtud de su significado, perfectamente
susceptible de ser contrastado inmediatamente en el orden práctico manual).
Pero cuando la actividad humana recae, no ya sobre el objeto material
observable, sino sobre estas mismas proposiciones protocolarias o enunciados
de observación, tratando de compendiarlos en forma de expresiones de lo que
impropiamente se ha dado en llamar "hechos generales" -los hechos, en rigor,
son siempre particulares- y algo más propiamente generalizaciones
inductivas, es cuando comienza el trabajo teórico, presidido por el método
científico propiamente dicho, que entonces entra plenamente en juego.
Encerrándonos en un laboratorio con instrumentos e ingredientes adecuados,
podemos comprobar exacta o muy aproximadamente que 1g de plomo se

[86]
combina con 0,0772g (= 3 x 0,0257) de oxígeno formando
Fig.2 Litargirio Pb O3
Litargirio; que también 1g de plomo se combina con 0,103g (= 4 x 0,0257) de
Fig.3 Plomo rojo Pb O4
oxígeno formando plomo rojo; que análogamente sucede con otros muchos
pares de elementos químicos respecto a determinadas y diferentes cantidades
de ellos siendo fija la de uno en cada par. Pero es prácticamente imposible
comprobar, para todos los pares de elementos químicos que se combinen, que
las diferentes cantidades de uno de ellos que se combinan con una cantidad
fija del otro, están siempre entre sí -es decir lo han estado en el pasado, lo
están actualmente en el presente y lo estarán en el futuro, donde quiera que
sea (en el laboratorio o fuera de él)- en determinadas relaciones numéricas
muy sencillas. Aquellas particulares afirmaciones perfectamente comprobables
son de la clase de las proposiciones protocolarias o enunciados de
observación; esta otra, en cambio, a pesar de su enfático nombre tradicional de
ley de las proporciones múltiples de Dalton,

[87]
Fig. 4 John Dalton (1766-1844) ..hipótesis le conduce a la formulación de la Ley de las proporciones múltiples: Cuando dos elementos forman de manera conjunta más de un compuesto, al unirse distintas masas de uno de ellos con una misma masa del otro, establecen entre sí relaciones múltiples y sencillas.
no es, metodológicamente hablando, más que una hipótesis inductiva o una
generalización inductiva, como nosotros las hemos llamado anteriormente, y
sin embargo es más corriente llamarlas hipótesis inductivas.
El último acto o paso en la ruta teórica inspirada por el método científico es la
formulación de ciertos enunciados inventados para poder deducir de ellos,
como consecuencias lógicas de ellos mismos, entre otros enunciados posibles,
los de las susodichas hipótesis inductivas. Estos últimos, inventados o
formulados completamente “ad hoc”, son las hipótesis explicativas, que ni
sintáctica ni terminológicamente se asemejan a las hipótesis inductivas. Por
tanto, en Genética, como en cualquier otra ciencia experimental, es de los
enunciados de las hipótesis teóricas o explicativas de donde se deducen
aquellos otros que corrientemente se consideran generalizaciones inductivas
de los hechos observados, pero que, en rigor, son generalizaciones inductivas
de las proposiciones protocolarias que expresan los hechos observados.

[88]
Proposiciones protocolarias
(Los datos obtenidos de nuestro experimento)
Generalizaciones inductivas
(Expresión de lo que se suele llamar un "hecho general")
Hipótesis explicativas
(Inventadas de manera que sean relativamente pocas y
se deduzcan de ellas todos los demás
enunciados de la teoría que no sean definiciones)
Las hipótesis explicativas para la hipótesis inductiva, que es la ley de las
proporciones múltiples de Dalton, componen la teoría atómico molecular
formulada por este mismo autor en 1805, la cual sigue siendo esencialmente
correcta, aunque ahora no admitamos, como él afirmaba (desde luego) que los
átomos son individualidades indivisibles e indestructibles.
Dicho esto, afirmábamos antes que el método científico representa la
estructura misma de la ciencia experimental formulada en términos
completamente generales. A este respecto, viene a ser muy ilustrativo recordar
sumariamente la historia que ha seguido durante estos cuatro últimos siglos la
ciencia más venerable y, a la vez, hoy de más actualidad, la cosmología.
El enorme acopio de precisas observaciones astronómicas realizado por
Tycho-Brahe en "Uranienborg" (1576-1597)

[89]
Fig.5. Tycho-Brahe. Astrónomo danés. (Knudstrup, Dinamarca, 1546 - Benatky, actual
Chequia, 1601)
es, ciertamente, un tesoro, pero, en rigor, un tesoro formado exclusivamente de
proposiciones protocolarias o enunciados de observación (la obra entera de
Tycho-Brahe).
Johannes Kepler, que entró como asistente de Tycho cuando este hubo de
trasladarse a Praga, vino a heredar de él su tesoro de observaciones
astronómicas y, además, la sugerencia de calcular, a partir de ellas, la órbita de
Marte. Kepler llevó a cabo este cálculo, pero también hizo lo mismo respecto a
los otros cuatro planetas conocidos: calcular sus respectivas órbitas. Como
ulterior resultado de estos cálculos, mezclándolos con relaciones astrológicas,
consideraciones místicas y hasta referencias a las notas musicales, vino al fin a
formular Kepler sus tres famosas leyes, que publicó después de la muerte de
Tycho en las dos obras "Astronomía nova" (1609) y "De harmonice mundi"
(1619). Pues bien, estas leyes de Kepler son propiamente hipótesis inductivas,

[90]
Fig.6. Johannes Kepler. (Würtemburg, actual Alemania, 1571-Ratisbona, id., 1630)
Astrónomo, matemático y físico alemán.
y, así, en ellas se habla todavía concretamente del sol, de los planetas, de sus
órbitas, de sus revoluciones planetarias. Y como tales hipótesis inductivas que
son, esto es, aplicables a cualquier planeta posible y en todo tiempo, sirvieron
dos siglos y medio después (1846) a Le Verrier para descubrir la existencia de
Neptuno, calculando las características de este nuevo planeta a partir de las
perturbaciones observadas en los movimientos de Urano.

[91]
Fig.7. Isaac Newton. Uno de los más grandes científicos de la historia de la
humanidad. Isaac Newton nació en las primeras horas del 25 de diciembre de 1642 (4
de enero de 1643, según el calendario gregoriano), en la pequeña aldea de
Woolsthorpe, en el Lincolnshire.
Isaac Newton, formulando en 1689, dentro de su monumental "Philosophiae
naturalis Principia Mathematica", los tres axiomas o leyes del movimiento de la
mecánica universal, y definiendo para ello cosas tan abstractas como "cantidad
de materia", "fuerza centrípeta", “magnitud aceleratriz", etc., viene a enunciar la
hipótesis teórica explicativa de la cual se deduce la ley de la gravitación
universal y luego las leyes de Kepler. En resumen: el método científico no es,
en manera alguna, un recetario para fabricar tesis doctorales. El método
científico representa la estructura ideal, puramente formal, intrínseca a la propia

[92]
ciencia experimental, que podemos describir de manera más rigurosa, como
inmediatamente haremos.
Proposiciones protocolarias o de observación.
Una proposición protocolaria es, en cualquier ciencia natural, un enunciado que
tiene como objeto de expresión, o referencia objetiva, un hecho particular
observado, un hecho experimental. El tipo de hecho a que se refiere cualquier
proposición protocolaria es un conjunto de datos (el volumen, los genotipos, el
peso, el fenotipo, una intensidad, etc.); pero -y esto es lo más importante- se
refiere a un conjunto de sucesos observable, manejable y numerable, no sólo
en el sentido teórico de la palabra, sino en el práctico, por cuanto tal conjunto
ha sido efectivamente observado, manejado y contado por un observador o por
varios.
Fig.8. Investigador recogiendo los datos protocolarios, esto es, un conjunto de
sucesos observable, manejable y numerable, no sólo en el sentido teórico de la
palabra, sino en el práctico, por cuanto tal conjunto ha sido efectivamente observado,
manejado y contado por un observador o por varios.

[93]
En este sentido diremos, como característica más distintiva de las
proposiciones protocolarias en cualquier ciencia, que estas se refieren
concretamente a conjuntos, realmente apreciables para el observador.
Fig.9. Las proposiciones protocolarias pueden llamarse también enunciados de
observación, esto es, un conjunto de sucesos observable, manejable y numerable.
Las proposiciones protocolarias relatan, de forma más o menos sumaria, pero
concreta, lo que ocurrió en un tiempo o fecha perfectamente determinado y en
Fig.10. Los datos protocolarios, enunciados de observación relatan, de forma más o
menos sumaria, pero concreta, lo que ocurrió en un tiempo o fecha perfectamente
determinado.

[94]
un sitio concretamente determinado, según observación personal de uno o
varios autores también determinados. Las proposiciones protocolarias pueden
llamarse también enunciados de observación.
Generalizaciones o hipótesis inductivas.
El enunciado de una generalización inductiva es la expresión de lo que se
suele llamar un "hecho general", como primera aproximación o toma de
contacto con la cuestión. Una generalización inductiva es siempre una
expresión/descripción frecuentemente escrita que es planteada en función de
varias proposiciones protocolarias, expresas o tácitas, pero verdaderas, a
menos que la generalización inductiva carezca de toda validez. Siguiendo
todavía con la idea del "hecho general", podemos decir que las proposiciones
protocolarias son las expresiones de otros tantos hechos particulares. El
enunciado de una generalización inductiva difiere muy poco del enunciado de
cualquiera de las proposiciones protocolarias correspondientes. En el
enunciado de una proposición protocolaria, en cualquier ciencia, se dice o
afirma que cierta relación, numéricamente expresada, se da dentro de
determinado conjunto asequible de datos de cierta clase, definida
generalmente por el modo de producción u obtención de estos datos. En el
enunciado de la generalización inductiva resultante de esta se afirma aquella
misma relación o -como más bien suele ser- una relación numéricamente
próxima a aquella, como relación propia del conjunto inasequible constituido
no solo por todos los datos actualmente existentes, sino también por los que ya
han existido y por los que habrán de existir pertenecientes a la clase definida
por su modo de producción u obtención. Es por esto por lo que en los

[95]
enunciados de las generalizaciones inductivas, al contrario que en los
enunciados de las proposiciones protocolarias, no figuran números cardinales
de conjuntos finitos de datos, aunque si relaciones numéricas o cantidades
numéricas, pero si frecuentemente, en su lugar, adjetivos indefinidos, como
'todo', 'todos', 'cualquier', 'cada' o el artículo definido precediendo a un
sustantivo y designando así a la clase de datos de que se trata.
En suma: si las proposiciones protocolarias se caracterizan por referirse tan
sólo a conjuntos asequibles (efectivamente observados, manejados y contados
por algún observador), las generalizaciones inductivas se distinguen, en
cambio, por referirse a conjuntos inasequibles (no observados, manejados, ni
contados, en su totalidad, por nadie).
En rigor, las generalizaciones inductivas son hipótesis genuinas: sus
enunciados expresan lo que se supone que tiene lugar en un conjunto de
hechos tan grande que ningún observador, o equipo de observadores, puede
llevar a efecto su comprobación contando uno por uno los distintos
experimentos que lo componen; y las proposiciones protocolarias son los
únicos enunciados que, en las ciencias experimentales, expresan o se refieren
inmediatamente a los hechos auténticos. Muchos autores prefieren, por esto,
llamar a las generalizaciones inductivas hipótesis inductivas.
Hipótesis teóricas o explicativas. Dos son las notas que caracterizan a una
hipótesis teórica o explicativa que forma parte de alguna teoría científico-
natural: una, que de su enunciado se deduce mediante puro razonamiento, es
decir, lógicamente, el enunciado de una hipótesis inductiva; y la otra, que su
enunciado no se deduce lógicamente de ningún otro de los enunciados que

[96]
forman parte de la teoría. Precisamente por ser esto característico, con
bastante rigor, de las hipótesis teóricas o explicativas, es por lo que se
requieren algunas aclaraciones.
En el desarrollo histórico de cualquier ciencia natural pronto se reúnen muchos
enunciados que son otras tantas hipótesis o generalizaciones inductivas;
entonces, las hipótesis teóricas o explicativas son inventadas, de manera
que sean relativamente pocas y se deduzcan de ellas todos los demás
enunciados de la teoría que no sean definiciones, y, entre ellos, los de las
hipótesis inductivas. Así pues, el valor de las hipótesis teóricas o
explicativas es el de proporcionar orden lógico y síntesis en el conjunto de
enunciados de una ciencia, convirtiéndose en la teoría propia de esta ciencia.
Otra urgente aclaración es que decir que del enunciado de toda hipótesis
teórica o explicativa se deduce lógicamente el enunciado de alguna hipótesis
inductiva, no implica que del solo enunciado de cada hipótesis explicativa se
deduzca el de una hipótesis inductiva. Por lo pronto tenemos que un enunciado
no se deduce de otro sin el auxilio de los principios y reglas de consecuencia
lógica que sean (expresa o tácitamente aplicados). Pero además, lo que suele
ocurrir en las teorías es que cada enunciado de hipótesis inductiva sólo se
deduce del conjunto de varios enunciados de hipótesis explicativas. Una teoría
científica en ciencia natural es, como dice Woodger, (Woodger, Joseph Henry
(1894–1981) ) comparable a un iceberg. Solo una parte pequeña de la teoría es
la que está a la vista; fuera de la vista, como sumergida, queda la mayor parte
de la teoría. La parte de la teoría que está a la vista son los enunciados de la
teoría que hablan de cosas inmediatamente observables. La parte sumergida
es el conjunto de enunciados de la teoría, expresos unos, implícitos los otros,

[97]
pero solo expresados o expresables unos y otros en términos de cosas no
directamente observables.

[98]
EL Laboratorio de Biología y el nacimiento de la Genética experimental en España
En 1913, unos años antes de la primera guerra Mundial, se creó una sala en el
Museo Nacional de Ciencias Naturales de Madrid, que tenía un letrero en la
entrada en el que aparecía la leyenda “Laboratorio de Biología”.
Es allí donde nació la Historia de la Genética en España. El profesor Antonio de
Zulueta Escolano (Barcelona, 1885 - Madrid, 1971) creó ese laboratorio, con
una intensa actividad científica importante hasta 1936.
Fig.1 Antonio de Zulueta, en sus años como Universitario
En aquel laboratorio iniciaron también su actividad científica José Fernández
Nónidez (Madrid 1892 – Augusta, Georgia, USA 1947) y Fernando Galán
Gutiérrez (Luarca 1908 – Salamanca 1999). A estos tres nombres propios se
les considera como los padres de la Genética en España, aunque hay que
señalar que toda la actividad científica de Nonídez estuvo más dedicada a la
citología que a la Genética.

[99]
Antonio de Zulueta fue profesor del Museo Nacional de Ciencias Naturales de
Madrid,
Fig.2 El Prof. Antonio de Zulueta en su “Laboratorio de Biología” –sitio en la residencia de Estudiantes, calle del >>Pinar- del Museo nacional de Ciencias Naturales de Madrid (según foto del Marqués de Villalcázar en septiembre de 1934)
en el que desempeñó el cargo de Jefe del Laboratorio de Biología durante 58
años, concretamente desde la creación de este laboratorio hasta su
fallecimiento, en 1971.
Este Laboratorio de Biología, como nueva unidad de investigación dentro de
dicho Museo, y el llamado Curso Práctico de Biología, como función docente
adscrita a aquel Laboratorio, fueron creados en 1913. Y tanto la dirección de
aquel Laboratorio como la impartición de este Curso Práctico fueron
encomendadas desde un principio a don Antonio de Zulueta, con el propósito
de fomentar, entre los naturalistas de entonces, los estudios experimentales de
biología.
Trabajando siempre en su Laboratorio de Biología, el profesor Zulueta tuvo el
gran mérito de introducir en España, con el ejemplo de su propia labor

[100]
personal, la investigación científica experimental en el campo de la ciencia
genética. Pero además, el profesor Zulueta siguió durante toda su vida
promoviendo la investigación y el cultivo de la ciencia genética en nuestro país:
primero, mediante el magisterio inherente a su personal labor de investigación;
luego, mediante el magisterio propiamente didáctico o docente de sus
excelentes lecciones en la Cátedra de Genética de la Fundación “Conde de
Cartagena” de la Real Academia de Ciencias Exactas, Físicas y Naturales de
Madrid.
Cuando el profesor Zulueta, hacia finales del segundo decenio de hace dos
siglos, decidió dedicarse a los estudios experimentales sobre la herencia
biológica -como entonces se decía-, tales estudios eran considerados por
muchos biólogos y naturalistas con no poco escepticismo, y sus cultivadores
mirados en los congresos de zoología o de botánica con piadosa indulgencia,
como colegas sumamente ingenuos que se entretenían en comprobaciones, al
parecer, tan triviales e intrascendentes como ver cuántos guisantes salían
redondos y cuántos rugosos en una generación.
Zulueta fue quien, entre nosotros los españoles y sin abandonar España (en su
Laboratorio de Biología del Museo Nacional de Ciencias Naturales de
Madrid), no solo supo reconocer tempranamente la grandísima importancia que
entrañaban aquellos, al parecer, ingenuos estudios que con el transcurso del
tiempo se convertirían en la genética moderna, sino, además, quien primero se
dedicó a trabajar experimentalmente en ellos y a procurar, así, su implantación
en nuestros propios lares científicos. En efecto, Zulueta introdujo la
investigación genética en España y llevó a cabo un notabilísimo
descubrimiento: el de la existencia de genes en el cromosoma Y (el

[101]
cromosoma sexual exclusivo de los machos de muchas especies animales, la
humana inclusive). Los trabajos se iniciaron en 1920 con la especie Phytodecta
variabilis y, cinco años después, fue publicado el descubrimiento que le hizo
famoso; en el que se comprobaba experimentalmente, por primera vez, la
existencia de genes en el cromosoma sexual Y, hasta entonces considerado
como un cromosoma vacío de genes
.
Fig.3 Dibujo del Phytodecta variabilis realizado por el profesor Galán cuando colaboraba en el Laboratorio de Biología creado por el profesor Zulueta. Phytodecta variabilis, un coleóptero muy abundante en los alrededores de Madrid, que llama inmediatamente la atención por sus muy diferentes y bien definidos tipos de coloración.
Dije notabilísimo descubrimiento, y casi mejor podría decir sorprendente, por
cuanto la opinión científica autorizada imperante entonces, respaldada ya por la
impresionante labor experimental de la famosa escuela de Morgan en
Drosophila Melanogaster, era la de que el cromosoma Y estaba vacío de
genes, por no conocerse ningún hecho que obligase a localizar genes en él;
era considerado como vacío o agotado, como un corpúsculo vestigial,

[102]
desaparecido en los machos de tipo XO. Y su importancia teórica y
experimental, genética y citológica a la que hemos aludido queda atestiguada
por el mismo profesor Morgan y premio nobel cuando, poco después de la
publicación de las investigaciones de Zulueta en 1925 en Phytodecta, señaló
expresamente la “muy especial importancia” de estas investigaciones, en un
artículo que apareció en 1926 en “Quarterly Review of Biology” (artículo en el
que Morgan recopilaba los principales avances de la genética en aquellos
años). Y en seguida, los datos, las figuras, las genealogías y las explicaciones
de Zulueta relativas al caso de Phytodecta aparecieron extensamente
incorporadas a las monografías y a los tratados de genética. Así, en el gran
tratado o “biblia” de la genética de los años treinta, esto es, en el “Handbuch
der vererbungswissenschaft”, editado por BAUR & HARTMANN, dichos datos,
figuras, genealogías y explicaciones de Zulueta para el caso de Phytodecta
variabilis aparecen insertos reiteradamente, como era procedente por su doble
importancia: primero, con ocasión del tema del alelismo múltiple (tema
redactado por Stern); luego, a propósito del de la herencia ligada al sexo (tema
este otro escrito por Foÿn).
Por último, la consideración del caso de Phytodecta pasó a los libros de texto,
paso que es considerado como la prueba más notoria y objetiva de la
importancia de un descubrimiento científico. A este respecto, fue el profesor
Alvarado, en su excelente y conocidísimo libro de texto de Biología, quien
primero expuso, de manera didáctica en el ámbito universitario, el interesante
caso de “herencia ligada al sexo por el cromosoma Y”, estudiado por Zulueta
en Phytodecta variabilis.

[103]
El languidecer del laboratorio coincidió con la penosa Guerra Civil y con los
intentos de postergación extracientífica de la que fueron objeto el profesor
Zulueta y el profesor Galán. Antonio de Zulueta había sido expulsado del
Museo con lo que su labor científica se hizo cada vez más penosa y casi
impedida por el gobierno golpista, pero, sin embargo, la continuidad de todos
aquellos estudios se trasladaron a la Universidad de Salamanca, donde
Fernando Galán Gutiérrez comenzó a ocupar la Cátedra de Biología. Donde
sus primeros años se le retiró el sueldo, aunque no de su puesto de
funcionario, figura por cierto de suma importancia en los 40 años de la tiranía.
Y siguiendo con estas anécdotas no científicas, diremos, que la dificultad de los
trabajos experimentales que se realizaron durante esos años existen notas en
revistas científicas como Nature, donde J.B.S. Haldane publica el siguiente
comentario en el número del 20 de Febrero de 1937.
<<…."Durante una reciente visita a Madrid para otros propósitos, pude visitar a
algunos de mis colegas de allí, y quedé encantado al comprobar que el trabajo de
investigación en Genética continuaba. El profesor A. de Zulueta, en los intervalos de
guarecer en sótanos los más preciados objetos del museo biológico, continuaba sus
trabajos sobre el polimórfico escarabajo Phytodecta variabilis. El profesor Galán, de
Salamanca, estaba cruzando muy cuidadosamente el "pepino explosivo" Ecballium
elaterium... Nuestra discusión de estas cuestiones se interrumpió por un raid aéreo
considerablemente más severo que cualquiera de los de Londres de 1914-18... Pienso
que la persistencia de Zulueta y Galán bajo condiciones que (lo menos que cabe decir)
no son agradables para investigar, merece ser recordada, y augura favorablemente
para el futuro de la ciencia de la Biología". >>

[104]
Fig.4 . <<….I think, -escribia el profesor Haldane-that the persistence of Zulueta and Galan under conditions which are, to say the least, uncongenial for research, deserves to be recorded, and augurs well for the future of biology in Spain…>>. Pero ni recordados -es verdad- y si postergados con la aquiescencia de los biólogos de su tiempo en el siglo pasado. FERNANDO GALÁN GUTIÉRREZ (1908 – 1999)
Fernando Galán Gutiérrez fue Catedrático de Biología de la Universidad de
Salamanca desde el año 1936 a 1978. En
el año académico de 1927-1928, al estudiar
el 2º curso de la Sección de Naturales,
entra en relación con los naturalistas del
Museo Nacional de Ciencias Naturales de
Madrid. Por entonces, asiste al “Curso

[105]
práctico de Biología Animal” que impartía en el Museo el profesor Antonio de
Zulueta.
De esta manera, Fernando Galán, traba una muy especial relación con el
profesor Zulueta y pasa enseguida, en el año 1928, a ser ayudante de este
profesor en aquel Laboratorio de Biología.
Cautivado por la valía científica del profesor Zulueta, decidió deliberadamente
continuar trabajando sin interrupción a su lado hasta 1939, año en el que se
incorpora de manera definitiva a la Universidad de Salamanca.
Deliberadamente, pues prefirió ser becario (de la “Fundación Premio Rivera”,
Patronato de Museo Nacional de Ciencias Naturales de Madrid en 1930; de la
“Junta para Ampliación de Estudios e Investigaciones Científicas” en 1931-
1933 y 1936-1939; de la “Fundación Conde de Cartagena”, Patronato de la
Academia de Ciencias Exactas, Físicas y Naturales en 1933-1935) al lado del
profesor Zulueta, antes que ejercer como Catedrático de Universidad (en mayo
de 1933 fue nombrado Catedrático numerario de Biología, en virtud de
oposición y con el número uno, de la Facultad de Ciencias de Sevilla en la
sección de Cádiz; y en abril de 1936, Catedrático numerario de Biología de la
Facultad de Ciencias de la Universidad de Salamanca). Estos años que van de
1928 a1939, pasados al lado del Prof. Zulueta, fueron los más fundamentales
para la formación científica superior de Fernando Galán y constituyeron,
principalmente, una etapa dedicada a la investigación científica sobre temas
diversos.
Después de la guerra civil las dependencias del Museo habían quedado
abandonadas y un tanto deterioradas. Deterioro y abandono no solo físico, ya
que se preparaba el espolio científico que se iba a realizar durante la dictadura

[106]
del franquismo. El profesor Galán había recogido de entre la basura del Museo
el letrero del en el que ponía “Laboratorio de Biología”, y lo supo colocar en la
entrada de su nuevo laboratorio en la Universidad de Salamanca. Pero
además, se llevó consigo el espíritu de trabajo existente en el Museo, previo al
languidecer científico de este, como ya decíamos, al ser incorporado al al mal
llamado Consejo de Investigaciones Científicas.
Quizá resulte un tanto sorprendente que el profesor Galán no hubiera ido a
trabajar fuera de España, como lo hicieron Zulueta y Nonídez. Pero no fue
debido a la falta de oportunidades, pues hemos de recordar que en 1936
recibió una beca de la Rockefeller Foundation para trasladarse a Caltech y
trabajar con Thomas Hunt Morgan, como lo había hecho Nonídez, sino a que,
al estallar la guerra civil en España, Galán pensó, en palabras casi literales,
que “su deber patriótico era quedarse por si su país le necesitaba”.
A partir del trasladado de la actividad científica a Salamanca, es cuando se
desarrolla la fase experimental de los trabajos en Ecballium elaterium, especie
a la que Galán dedicó su vida científica.
Fig.5 Imagen de una planta andrógina de Ecballium elaterium.L.Richard.

[107]
Remontándonos a la década de 1925 a 1935 de nuevo, uno de los problemas
científicos más importantes de la genética, que intrigaba sobremanera
entonces a los autores, era el denominado de la “determinación del sexo” en
los animales y en las plantas. Pues bien, atraído por este problema, el profesor
Galán decidió en 1935 emprender una detenida y cuidadosa investigación
experimental que le permitiera dilucidar el problema genético de los
realizadores o determinantes del sexo en los metazoos y antofitas: un problema
este acometido, a partir de Correns en 1908, una decena de veces, por otros
tantos autores, con una veintena de diversas especies, y de solución siempre
frustrada por la esterilidad entre los genitores o por la esterilidad del híbrido.
Para ello, el profesor Galán comenzó en 1935 por llevar a cabo una exploración
de la vegetación natural silvestre de la Península Ibérica, con el singular
propósito de ver si en alguna especie de la flora de esta Península se diese la
particularidad, coincidencia de existir en ella dos variedades sexuales tales que
una tuviese plantas macho, a la vez que plantas hembra (lo que se dice una
variedad dioica), mientras que la otra solo tuviera, en cambio, plantas
hermafroditas o andróginas (lo que se dice una variedad monoica), y con la
esperanza de que, además, ambas variedades se dejasen cultivar y cruzar
entre sí con facilidad. Tal exploración encontró feliz éxito en una conocidísima
especie de cucurbitácea silvestre mediterránea llamada Ecballium elaterium. L.
Richard comprobando que en el norte de España todas las poblaciones de E.
elaterium están compuestas exclusivamente de plantas andróginas y
constituyen, así, una subespecie monoica, mientras que en el sur de España
todas las poblaciones están compuestas exclusivamente de plantas machos y

[108]
plantas hembras, y forman, por tanto, una subespecie dioica. Dicha
comprobación dio como resultado un estudio que se realizó de forma rigurosa,
y que aún no se ha publicado, en el que se llegaba a la conclusión de que las
dos subespecies por especiación alopátrida en la península Ibérica, Ecballium
elaterium L. Rich. subsp. dioicum y subsp. Monoicum, quedaban bien definidas
según las isobaras de la península. Y que en 1981 la profesora Coschi publicó,
eso sí, bajo la tutela e información que el profesor Galán le proporcionó, el
trabajo “Genetic variation in Ecballium elatedurn (Cucurbitaceae): Breeding
system and geographic distribution J.evol.Biol. 5: 589-601 (1992), que no es
más que una pequeña parte de las investigaciones que treinta años antes
habían sido realizadas por el equipo científico del profesor Galán en la
Universidad de Salamanca. Resultados protocolarios que, por cierto, acababan
apilados entre otros muchos que se iban realizando por dicho equipo del
profesor Galán -una forma de proceder de la que nunca se ha sabido el
porqué-. Pero estos hallazgos, resultados experimentales, eran relegados, ¿o
retrasada, intencionadamente su publicación en las revistas científicas? Tal
vez, como se rumoreaba entre sus colaboradores, su admiración por Mendel
era lo que le hacía ser olvidado por la sociedad científica de aquel tiempo, una
sociedad científica Española que estaba dominada e inmersa en el
envilecimiento del espíritu franquista que él despreciaba de manera profunda.
Pero he aquí, el gran hallazgo verdaderamente olvidado por los científicos de
su tiempo y de hoy día: la propuesta y el modelo de la primera hipótesis de la
existencia de lo que hoy ha venido a llamarse morfo genes, cuya concentración
espacio temporal es la que determina el desarrollo de los organismos. Y que
fue comprobada macroscópicamente mediante el estudio estadístico de

[109]
caracteres asociados directamente e indirectamente con el sexo de la
cucurbitácea Ecballium elaterium.L.Richard. En los campos experimentales de
la Universidad de Salamanca.
Fig.6. Campos experimentales con más de 5000 plantas con las que entre otros hallazgos, fue la comprobación estadística anatómica-morfológica de la primera hipótesis sobre la existencia de los gradientes morfo genético. Aquellos campos experimentales donde cultivaba y hacía los cruzamientos con las plantas de Ecballium elaterium estaban situados en lo que habían sido los campos de deportes de la Residencia de Estudiantes, ahora propiedad del centro de investigaciones agrícolas IOATO. Fue el profesor Lucena-Conde, rector de la Universidad de Salamanca, el que apoyó incondicionalmente la cesión de aquellos terrenos.
Así, de los análisis de los datos experimentales obtenidos entre 1935 y 1939
nace la primera propuesta conocida de lo que hoy conocemos por gradientes
morfo genéticos, o, más concretamente, la determinación genética del sexo, en
la planta Ecballium elateriun .L. Richard, mediante un gradiente morfo genético
determinado por de tres genes alelomorfos.
Este determinismo genético fue propuesto por Galán, por primera vez, en 1946,
con posteriores publicaciones en 1951, 1953, 1954, 1957 y 1964.

[110]
Y la divulgación de esta hipótesis del Gradiente Morfo genético comenzó en el
Congreso de Genética de Estocolmo, a mediados de la década de los 40, del
que existe alguna nota de prensa recogida incluso por la agencia EFE.
Fig.7. En la nota se indica que el profesor D. Fernando Galán no pudo acudir al Congreso. La realidad era que la vinculación ideológica de Galán hacía que las autoridades de la época le impidiesen atravesar las fronteras del país. Es en el citado congreso donde el científico holandés Linskens conoce estas
investigaciones y cursa distintas invitaciones al profesor Galán para que acuda
a su laboratorio de Ámsterdam. Ante la insistencia de Linskens, Galán publica
su trabajo, de manera más detallada, en la revista científica alemana

[111]
Physiologie der Fortpflanzung und Sexualität in Fortschritte der Botanik, XIX, pp
387-398 en 1957.

[112]
Fig 8 En esta publicación se recoge por primera vez en la historia de la Genética la relación entre 3 genes que determinan el sexo de la planta a través de un gradiente morfo genético.

[113]
Podría parecer una hipótesis más entre las muchas que existen en la ciencia
de la Biología, pero, como se verá más adelante, este no es el caso.
Como hemos dicho, este trabajo se dio a conocer en 1957, pocos años más
tarde de que Watson y Crick descubrieran la estructura de la doble hélice
(1953) y se iniciase lo que hoy conocemos como Biología Molecular.
Curiosamente, el trabajo de Galán, en el que se relacionaba un sistema de
genes alelomorfos con uno o varios caracteres determinados por las
respectivamente correspondientes concentraciones de una fitohormona, estaría
encuadrado dentro de lo que hoy conocemos como Genética Molecular.
Aquella trascendental e inoportuna hipótesis de mediados del siglo pasado, se
hizo basándose en las pruebas preliminares, no suficientemente rigurosas, que
comenzaron hacia 1935. En realidad, esta hipótesis se había adelantado a las
posibles técnicas científicas experimentales de comprobación existentes, más
aún en una España en la que apenas existían recursos de ningún tipo, y mucho
menos para una persona contraria al régimen.
Sabida la falta de medios para continuar con las investigaciones, sólo cabía
realizar comprobaciones anatómico-fisiológicas y que, además, tuviesen una
correlación clara y cuantificable con la existencia de ese gradiente morfo
genético determinado por tres genes mayores alelomorfos, y así, pudiesen ser
validadas matemáticamente al más puro estilo de Mendel.
Hemos de recordar que, entre los colaboradores del profesor Galán, estuvo,
durante la década de los 60, el joven Francisco J. Ayala (hoy uno de los
científicos de más renombre mundial), que trabajó durante un par de años en
aquel laboratorio de la Universidad Salamanca.

[114]
Fig.9 Heredero intelectual de Theodosius Dobzhansky (1900-1975), el profesor Ayala, nacido en Madrid en 1934, es considerado actualmente una de las mayores autoridades mundiales en genética de poblaciones. Los trabajos experimentales del profesor Galán tenían como organismo
bilógico el Ecballium elateriumL. Richard, vulgarmente llamado el “pepinillo del
diablo”; que eran cultivados en los campos experimentales de, donde se llegó
a tener un registro de 5.000 plantas, fig. 6. Donde cultivaron plantas con los
distintos genotipos: monoicas unas (las flores, tanto masculinas como
femeninas, se encuentran en el mismo individuo) y dioicas otras (los gametos
masculinos y femeninos son portados por individuos distintos de la misma
especie) así como todos los posibles híbridos entre las dos subespecies.
Fig.10 Parte del grupo de colaboradores dirigido por el profesor Galán
Pues como decíamos, de aquella población de más de 5000 plantas de
Ecballium elateriumL. Richard, diocas, monoicas e híbridos inter-específicos se
realizó un registro minucioso por parte del grupo de colaboradores dirigido por

[115]
el profesor Galán. En dicho registro no solo se anotaban todos los detalles de
cada planta, sino que se realizaba un esquema de la distribución de las flores
macho, hembra y hermafroditas. Aquel registro de cada una de las plantas se
repitió durante tres años, algo poco usual en los trabajos experimentales que
son publicados hoy día en revistas científicas.
Fig.11 Ejemplo de uno de los dibujos registrados de las plantas de Woodger cultivadas en el campo experimental del IOATO.
Lógicamente, aquella extensa y compleja etapa de colección de datos
experimentales fue el germen de una fecunda etapa posterior de
publicaciones.
De este registro, y del análisis estadístico del mismo, se pudo comprobar la
existencia de dos polaridades sexuales: una para el tallo y otra para la
inflorescencia. Esta polaridad se ajustaba perfectamente a la hipótesis
publicada 20 años antes. El estudio de Galán sobre los gradientes morfo
genéticos, comunicado en 1953 al IX Congreso Internacional de Genética,
reunido en Bellagio, tuvo continuidad en los trabajos experimentales que llevó a

[116]
cabo Mª Ángeles Sánchez Anta propuestos por Galán. En ellos se destaca la
extraordinaria variabilidad del grado de desarrollo de ambos verticilos sexuales
en flores hermafroditas, y se formula la norma que regula el desarrollo y
configuración de todas las flores de E.elaterium: cuanto mayor es el desarrollo
del gineceo, menor es el del androceo. Y que solo puede entenderse por la
trascendental hipótesis explicativa propuesta por Galán en 1957.
Fig12. En el eje y, el grado de desarrollo del gineceo versus androceo. En el eje x, las concentraciones umbrales y espacio-temporales de una posible fitohormona.
Lo mismo cabe decir sobre la teoría genética de la determinación del fenotipo
sexual de las flores en Ecbalium elaterium propuesta por Galán y comprobada
experimentalmente en 1946. Teoría esta que fue básica para posteriormente
proponer aquel gradiente morfo genético determinado por tres genes
alelomorfos. Sin embargo, sobre la teoría genética, la opinión que el profesor
Kenneth Mather fue en el primer momento (artículo publicado en Nature en
1949, en relación a los trabajos de Galán), que la modificabilidad fenotípica que
se comprobaba en las poblaciones de Ecbalium elaterium era causa de la

[117]
posible existencia de un conjunto de poli genes influyentes sobre el sexo de la
planta.
Fig.13 DR. KENNETH MATHER, 22 June 1911 – 20 March 1990, who has been appointed to the chair of genetics now established in the University of Birmingham, is known for his, in some respects, revolutionary outlook on this subject. Dr. Mather derives this outlook from a fusion of several of the main streams of thought in biology.
Galán propuso a Juan González Julián la investigación experimental de la
existencia de poligenes en las plantas de E. elaterium, y su posible relación con
la modificabilidad fenotípica sexual. El resultado, dado a conocer en 1983, es la
comprobación experimental de la existencia en E. elaterium de un sistema de
poligenes pleiótropos que condiciona directamente la duración de la vida de la
planta y el tamaño de ella, e indirectamente la modificabilidad fenotípica sexual
de E. elaterium.
Del mismo modo, Galán propone a Francisca Gallego Martín la realización de
un trabajo sobre la “micros esporo génesis” de E. elaterium, en el cual se
describe, caracteriza y formula el cariotipo de E elaterium, y se comprueba que
no existen en esta especie cromosomas sexuales. La importancia añadida de
esta investigación radica en que la comprobación de la no existencia de
cromosomas sexuales es incompatible con la hipótesis citogenética de dos
pares distintos de genes propuesta por Westergaad (Advances in Genetics,

[118]
1958), para explicar los resultados de Galán en sus experimentos de
hibridación. Sin embargo, es 100% compatible con la hipótesis genética de
Galán. Finalmente, a partir de una sugerencia que le había hecho el profesor
Linskens, de la Universidad de Nimega, Galán Estella realizó, bajo su
dirección, la investigación titulada; "verificación química experimental de la
hipótesis fisiológica explicativa de la sexualidad de la planta Ecbalium
elaterium L. Richard", que fue publicada en 1988 y en la que se relaciona el
efecto de los niveles del ácido índole-3 acético en los ápices caulinares de los
tallos con el fenotipo sexual de la planta. De esta manera parece que se
comprueba la veracidad y su comprobación experimental de la hipótesis del
gradiente morfo genético determinado por tres genes alelomorfos publicada en
Physiologie der Fortpflanzung und Sexualität in Fortschritte der Botanik, XIX, pp
387-398 en 1957.

[119]
Fig 14. La primera hipótesis de un gradiente morfo genético en el Reino Vegetal
determinado genéticamente por tres genes alelomorfos en elaterium.L.Richard
Ahondando en la importancia de la hipótesis propuesta por el profesor Galán
Gutiérrez, cabe indicar que, en 1995, se le concede el premio Nobel de
Medicina y Fisiología a la profesora Christiane Nüsslein-Volhard, por su modelo
de un gradiente morfo genético en el desarrollo de los embriones de Drosophila
melanogaster, “la organización del huevo se pondría en marcha por la
colocación, controlada por el genoma materno, de varios gradientes morfo
genéticos antero-posteriores“
Fig 15. Foto de Christiane Nüsslein-Volhard
En la autobiografía que remitió a la Academia Sueca indica que, en 1977,
trabajando en la larva de Drosóphila, “interpretó el fenotipo postulando la
existencia de un gradiente. Trabajo que presento en 1978 en el Simposium
Anual de la Sociedad Americana de Biología del Desarrollo. ” Una simple

[120]
reseña escrita por la profesora Christiane Nüsslein-Volhard nos facilitará la
importancia de la hipótesis propuesta por el profesor Galán aproximadamente
30 años antes.
“It also led to a very detailed examination and description of the segmental
pattern of the Drosophila larva which we later used in our screens. I continued
the work on dorsal, discovered the recessive phenotype and interpreted the
phenotype postulating a gradient determining the dorsoventral axis. At that time
(1977), gradients were not widely accepted as mechanisms, in particular
biochemists were highly sceptical, however the Tübingen influence made such
models attractive to me. I presented this and the bicaudal work at the annual
symposium of the American Society of Developmental Biology in Madison in
1978, my first trip to the US” [. A first model describing the three independent
systems involved in establishing the anteroposterior axis was presented in an
article in SCIENCE, with Frohnhöfer and Lehmann, in 1987..]
Con esta reseña –como comentábamos antes- queda evidenciada la
importancia de la hipótesis postulada por Galán y publicada en 1957. Por lo
demás, diremos que el modelo genético propuesto por Galán para explicar la
monoecia y la diecia en Ecballium elateriumL. Richard, ha sido incorporado en
numerosos libros de texto de Genética, tanto extranjeros (Modern Genetics
Ayala y Kieger, 1984) como nacionales (Lacadena, 1988, 1999; Sánchez
Monge y ]ouve, 1989; Puertas, 1999).
El profesor Galán recibió la Medalla de Oro de la Ciudad de Salamanca en
1985, y en el acto de entrega dijo que “después de medio siglo dedicado
exclusivamente al puro negocio del estudio, soy un auténtico proscrito. Trabajo

[121]
con unos pocos de mis más adictos colaboradores, arrinconado en un
vergonzante hacinamiento de libros, aparatos, utensilios y protocolos".
Estos comentarios del profesor Galán ahondaban en la línea de olvido científico
con la que había vivido durante tantos años. Ciertos científicos se salen de las
normas que la sociedad científica -quizás de cualquier sociedad- mantiene y
entonces no se sabe el porqué de tal olvido, que frecuente, se cierne sobre
ellos.
-------------------------------------

[122]
Versuche über Pflanzen-Hybriden
Experimentos de hibridación en plantas.
Por
Gregor Mendel
(Leído en las reuniones del 8 de febrero y del 8 de marzo de 1865)
Traducción del Alemán al Español
Por
F.Galan-Gutierrez y Sancho Blasco, Sonia
Galán-Estella, Fernando1; González Julian, Juan2; Sancho Blasco, Sonia4; Marañón Maroto, Jose Antonio3; Caballero, Ernesto5;. in memorian F.Galan-Gutierrez
1)Universidad Autónoma Madrid, Science Faculty, Analytical Chemistry Dept. Spain 2)
Medicine Faculty. Electronic Microscopy, University of Salamanca 3) 4) Tradichem.
S.L Actives & Ingredients Santiago Grisolía 2- 102. Parque Científico de Madrid.
28760. Tres Cantos. Madrid. SPAIN 4 ) University of Leipzig Institute of Analytical
Chemistry. Linnéstrasse.3. 04103 / Leipzig. Germany; 5) Universidad Miguel
Hernández de Elche. Alicante Spain
______________

[123]
Einleitende Bemerkungen
Consideraciones preliminares
Fecundaciones artificiales, emprendidas en plantas de adorno para obtener
nuevas variedades de color, dieron ocasión a los experimentos de los que se
va a tratar aquí. La sorprendente regularidad con que reaparecían las mismas
formas híbridas, siempre que la fecundación se hacía entre las mismas
especies, vino a ser el estímulo para ulteriores experimentos, cuyo objeto fue
perseguir el comportamiento de los híbridos en sus descendientes.
A este asunto han consagrado parte de su vida, con infatigable perseverancia,
cuidadosos observadores como Kölreuter, Gärtner, Herbert, Lecocq, Wichura y
otros. Particularmente Gärtner, en su obra "Die Bastarderzeugung Im
pflanzenreiche", ha consignado observaciones muy estimables; y muy
recientemente fueron publicadas por Wichura fundamentales investigaciones
acerca de los híbridos de los sauces. Que todavía no se haya conseguido
establecer una ley suficientemente general acerca de la formación y el
comportamiento de los híbridos no puede extrañar a nadie que conozca la
extensión del tema y sepa darse cuenta de las dificultades con que han de
luchar los experimentos de esta índole. Una solución definitiva no se puede
obtener en tanto no se disponga de experimentos detallados acerca de las más
diferentes familias de plantas. Quien considere los trabajos en este campo
llegará a la convicción de que, entre los numerosos experimentos llevados a
cabo, ninguno lo ha sido en extensión y manera que permita determinar el
número de diferentes formas bajo las cuales aparecen los descendientes del
híbrido, ordenar con seguridad estas formas en las distintas generaciones y
establecer entre estas mismas formas sus mutuas relaciones numéricas.
Ciertamente requiere no poco ánimo someterse a tan dilatado trabajo, el cual,
sin embargo, parece ser el único camino seguro por el que pueda ser al fin
alcanzada la solución de un problema de importancia nada despreciable para la
evolución de las formas orgánicas.

[124]
La presente memoria trata de un ensayo detallado de tal experimento. Por
conveniencia, este experimento fue limitado a un pequeño grupo de plantas y
en la actualidad, después de una duración de ocho años, está esencialmente
terminado. Si el plan según el cual fueron dispuestos y efectuados los distintos
experimentos corresponde al problema planteado, quede ello a la decisión de
una crítica benevolente.
_____________________________
Auswahl der Versuchsoflanzen
Elección de las plantas de experimento
La validez e importancia de cualquier experimento están condicionadas por la
idoneidad de los medios empleados en él, así como por la adecuada aplicación
de estos mismos. Así, en el presente caso, no puede ser indiferente qué
especies de plantas se eligieron para realizar el experimento y de qué modo
éste se llevó a cabo.
La elección del grupo de plantas que debe servir para experimentos de esta
índole tiene que ser hecha con el mayor cuidado posible, si desde un principio
no se quiere arriesgar todo el éxito.
Las plantas del experimento deben cumplir:
1. Que posean caracteres diferentes constantes.
2. Que sus híbridos estén [naturalmente] protegidos de la intrusión de
todo polen extraño, o puedan serlo con facilidad artificialmente.
3. Que los híbridos y sus descendientes en las sucesivas generaciones no
experimenten en su fertilidad ninguna alteración sensible.
Falseamientos a causa de polen extraño, si ocurrieran en el curso del
experimento y no fueran advertidos, llevarían a conclusiones completamente
erróneas. Escasa fertilidad o completa esterilidad de ciertas formas, como la
que aparece en los descendientes de muchos híbridos, harían muy difícil los
experimentos o los frustrarían totalmente. Para conocer las relaciones en que
las formas híbridas están entre sí y con las formas originarias, parece

[125]
necesario el que puedan ser observados por completo todos los miembros de
la serie de desarrollo de cada distinta generación.
Desde un principio se reparó especialmente en las leguminosas, por su
peculiar constitución floral. Experimentos efectuados con diversos
representantes de esta familia llevaron a la conclusión de que el género Pisum
responde suficientemente a las exigencias requeridas.
Ciertas formas completamente autógamas de este género poseen caracteres
constantes, fáciles y seguros de distinguir, y dan en sus híbridos, en los
recíprocos cruzamientos, descendientes completamente fértiles. Una alteración
a causa de polen extraño tampoco es fácil que ocurra, pues los órganos de la
fecundación están estrechamente envueltos por la quilla, y las anteras estallan
cuando todavía la flor se encuentra en estado de yema, de modo que el
estigma queda recubierto de polen ya antes de abrirse la flor. Esta
circunstancia es de particular importancia. Como otras ventajas, merecen
citarse también el fácil cultivo de estas plantas en el terreno y en macetas, así
como la duración, relativamente breve, de su periodo vegetativo. La
fecundación artificial es ciertamente operación algo minuciosa, pero casi
siempre resulta bien. Para este fin se abre la yema floral aún no
completamente desarrollada, se aparta la quilla y, mediante unas pinzas, se
arranca con cuidado cada estambre, después de lo cual el estigma se puede
recubrir enseguida con el polen ajeno.
En varios comercios de semillas fueron adquiridas en total 34 variedades de
guisantes, más o menos diferentes, y se las sometió a una prueba de dos años.
En una variedad, entre un gran número de plantas iguales, se encontraron
algunas formas considerablemente dispares. Estas formas no variaron, sin
embargo, al año siguiente y concordaron completamente con otra variedad
adquirida en la misma tienda: sin duda, por puro accidente, las semillas habían
sido mezcladas. Todas las otras variedades dieron descendientes iguales y
constantes; al menos en los dos años de prueba no se advirtió ninguna
alteración esencial. Para la fecundación se eligieron 22 de ellas, que fueron
cultivadas cada año, durante toda la duración de los experimentos. Sin
excepción, permanecieron constantes.

[126]
La clasificación sistemática de estas mismas variedades es difícil e
incierta. Si se quisiera aplicar la definición más rigurosa del concepto de
especie, según la cual pertenecen a una especie solo aquellos individuos que
en circunstancias completamente iguales muestran caracteres también
completamente iguales, no se podrían contar dos de ellas como pertenecientes
a una misma especie. No obstante, según la opinión de los especialistas, la
mayoría pertenece a la especie Pisum sativum, mientras que las demás son
consideradas y descritas, ya como subespecies de Pisum sativum, ya como
genuinas especies, como P. quadratum, P. saccharatum, P. umbellatum.
Por lo demás, es completamente indiferente, para los experimentos de que se
trata, la jerarquía que se les asigne en el sistema taxonómico. Así como no se
puede trazar una línea precisa de separación entre especies y variedades,
tampoco se ha conseguido hasta ahora establecer una diferencia fundamental
entre los híbridos de especies y los de variedades.
______________
Eintheilung und Ordnung der Versuche
Clasificación y orden de los experimentos
Si se cruzan dos plantas que difieren de modo constante en uno o en
varios caracteres, entonces, como prueban numerosos experimentos, los
caracteres comunes se transmiten inalterados a los híbridos y a sus
descendientes; por el contrario, cada dos caracteres diferenciales se fusionan
en el híbrido en un nuevo carácter, el cual regularmente experimenta en los
descendientes siempre las mismas variaciones. Observar estas variaciones
para cada dos caracteres diferenciales y descubrir la ley según la cual estas
mismas variaciones ocurren en las sucesivas generaciones, fue el objeto de la
investigación. Esta se subdivide, en consecuencia, en tantos experimentos
distintos como caracteres diferenciales constantes hay en las plantas del
experimento.
Las diferentes formas de guisantes elegidas para fecundación mostraron
diferencias en la longitud y coloración del tallo, en el tamaño y forma de las
hojas, en la posición, color y tamaño de las flores, en la longitud del pedúnculo

[127]
floral, en el color, forma y tamaño de las legumbres, y en la coloración de los
tegumentos de las semillas y del albumen. Una parte de los caracteres citados
no permite, sin embargo, una separación segura y precisa, puesto que la
diferencia consiste en un "más o menos", frecuentemente difícil de determinar.
Tales caracteres no eran utilizables en los experimentos, éstos pudieron ser
limitados a caracteres que resultasen claros y decisivos en las plantas. El
resultado tenía al fin que mostrar si en reunión híbrida los caracteres siguen un
comportamiento concordante todos ellos y si, por lo tanto, cabe formar algún
juicio acerca de aquellos caracteres, los cuales tienen en el tipo de la especie
una importancia subordinada.
Los caracteres que se tomaron para los experimentos se refieren:
1. A la diferencia en la forma de las semillas maduras. Estas son: o bien
esféricas o redondeadas, de modo que las depresiones -si las hay- de su
superficie son sólo poco marcadas; o bien, irregularmente angulosas,
marcadamente rugosas (P. quadratum).
2. A la diferencia en la coloración del albumen de la semilla ('"endospermo"). El
albumen de las semillas maduras, o bien está coloreado de amarillo pálido,
amarillo, vivo o naranja; o bien tiene un color verde más o menos intenso. Esta
diferencia de color se reconoce con claridad en las semillas, ya que sus
tegumentos son transparentes.
3. A la diferencia en la coloración del tegumento de la semilla. Este tegumento,
o bien es de color blanco, con lo que va unido también constantemente el color
blanco de las flores, o bien es gris, pardo-grisáceo, pardo-cuero con
puntuaciones violeta o sin ellas, y entonces aparece el color del estandarte
violeta; el de las alas, púrpura; y el pedúnculo, en las axilas foliares, rojizo. Los
grises tegumentos de las semillas se vuelven pardo-negruzcos en agua
hirviendo.
4. A la diferencia en la forma de la legumbre madura. O bien está simplemente
abultada, sin estrangulaciones de trecho en trecho; o bien está profundamente
estrangulada entre las semillas, y es más o menos rugosa P. saccharatum.

[128]
5. A la diferencia en el color de la legumbre inmadura. Esta aparece coloreada:
o bien de verde brillante a verde oscuro, o bien de amarillo vivo, color del que
también participan el pedúnculo, las nerviaciones foliares y el cáliz1.
6. A la diferencia en la posición de las flores. Estas son, o bien axiales, esto es,
distribuidas a lo largo del tallo; o bien son terminales, agrupadas en el extremo
del tallo en pequeña pseudo-umbela; al mismo tiempo, la parte superior del
pedúnculo en su sección transversal está más o menos ensanchada P.
umbellatus.
7. A la diferencia en la longitud del tallo. La longitud del tallo es muy diferente
en las distintas formas, sin embargo, para cada una es un carácter constante,
diferente en las distintas formas, si bien es un carácter constante en cuanto
que, en plantas sanas que hayan sido cultivadas en el mismo suelo, solo
experimenta variaciones insignificantes. Para una diferenciación segura, en los
experimentos con este carácter, fueron siempre combinados el tallo largo de 6-
7` con el corto de 3-4` hasta 11/2.
Cada dos de los citados caracteres diferenciales, fueron [reunidos] combinados
por fecundación.
Para él
1. Experimento: se hicieron 60 fecundaciones en 15 plantas
2. “ “ 58 “ 10 “
3. “ “ 35 “ 10 “
4. “ “ 40 “ 10 “
5. “ “ 23 “ 05 “
6. “ “ 34 “ 10 “
7. “ “ 37 “ 10 “
De un gran número de plantas de la misma especie se escogieron para
fecundación solo las más vigorosas. Los ejemplares débiles dan siempre
resultados inseguros, pues ya en la primera generación de los híbridos, y
1 Una especie posee vainas de un hermoso color castaño rojizo, que, en la época de la madurez, pasa a violeta y azul. El ensayo sobre este carácter sólo se comenzó el año pasado.

[129]
todavía más en las siguientes, muchos descendientes apenas llegan a florecer,
o bien producen pocas y malas semillas.
Por último, en todos los experimentos se llevó a cabo el cruzamiento recíproco,
esto es, que aquella de las dos variedades que en unas fecundaciones sirvió
como planta productora de semillas fue empleada en las otras como planta
donante de polen.
Las plantas fueron cultivadas en arriates, si bien una pequeña parte de ellas,
en macetas, y mediante barrotes, varas y cuerdas tirantes se las mantuvo en
posición natural erguida. Por cada experimento, unas cuantas plantas en
maceta se pusieron durante la floración en un invernadero; estas plantas
deberían servir como testigos para los experimentos principales en el jardín
respecto a posibles alteraciones a causa de los insectos. Entre aquellos
[insectos] que visitan las plantas de guisantes, los escarabajos de la especie
Bruchus pisi podrían llegar a ser un peligro para el experimento, en el caso de
que apareciesen en grandes cantidades. Como es bien sabido, la hembra de
esta especie deposita sus huevos en la flor y, para ello, abre la quilla; en los
tarsos de un ejemplar capturado en una flor pudieron, con auxilio de una lupa,
verse claramente algunas células-polen. Se debe hacer aquí mención de una
circunstancia que pudiera ocasionar la intrusión de polen extraño, a saber:
ocurre, en algunos raros casos, que ciertas partes de flores, por lo demás,
normalmente desarrolladas, se atrofian; a causa de lo cual se produce una
denudación parcial de los órganos de la fecundación. Así, se observa un
desarrollo escaso de la quilla, por el que el estilo y las anteras quedaban en
parte al descubierto.
También ocurre a veces que el polen no llega a su completa formación. En este
caso, durante la floración ocurre un progresivo alargamiento del estilo, hasta
llegar el estigma a sobrepasar el extremo de la quilla. Este notable fenómeno
fue observado también en los híbridos de Phaseolus y Lathylus.
El peligro de falseamiento a causa de polen extraño es, sin embargo, muy
pequeño en Pisum y de ninguna manera puede alterar el resultado en su
totalidad. En más de 10.000 plantas que fueron investigadas
escrupulosamente, solo algunas veces se dio el caso de que fuera indudable la
ocurrencia de una intrusión. El hecho de que en el invernadero nunca fuera

[130]
observada tal perturbación deja suponer que Bruchus pisi y quizá también las
mencionadas anomalías en la constitución de la flor son la causa de ello.
______________
Die Gestalt der Hybriden.
La forma de los híbridos.
Ya los experimentos que fueron hechos en años anteriores con plantas
ornamentales proporcionaron la prueba de que los híbridos, por lo regular, no
representan la forma exactamente media entre las especies genitoras. En
particulares caracteres que saltan más a la vista, tales como los que se refieren
a la forma y tamaño de las hojas, a la pubescencia de algunas partes, etc., es,
en efecto, la forma media casi siempre evidente; en otros casos, por el
contrario, uno de ambos caracteres paternos posee una preponderancia tan
grande que es difícil, o completamente imposible, descubrir el otro en el
híbrido.
Exactamente así es como pasa con los híbridos en Pisum. Cada uno de los 7
caracteres-híbridos, o bien uno de ambos caracteres paternos es tan igual, que
el otro desaparece a la observación, o bien es a él tan semejante que es
imposible efectuar una distinción segura. Esta circunstancia es de gran
importancia para la determinación y clasificación de las formas bajo las cuales
aparecen los descendientes de los híbridos. En lo sucesivo, aquellos
caracteres que se transmiten completamente, o casi completamente invariables
a la combinación híbrida, puesto que propiamente representan el carácter
híbrido, serán designados como dominantes, y aquellos que en dicha
combinación vienen a ser latentes, como recesivos. El término “recesivo” ha
sido elegido porque los caracteres así designados retroceden o desaparecen
completamente en los híbridos, pero, como luego se dirá, vuelven a aparecer
invariables en los descendientes de estos mismos híbridos.
Finalmente, resulta probado por todos los experimentos que es completamente
indiferente que el carácter dominante pertenezca a la planta que forme las
semillas o a la que proporcione el polen; en ambos casos, la forma híbrida es
exactamente la misma. Este interesante fenómeno también fue señalado por
Gärtner, con la advertencia de que aun el más experimentado conocedor no es
capaz de distinguir en un híbrido cuál de las dos variedades combinadas fue la

[131]
planta productora de semillas y cuál la donante de polen.
De los diferentes caracteres que fueron empleados en los experimentos,
son dominantes los siguientes:
1. La forma redonda redondeada de las semillas con o sin leves
hundimientos.
2. La coloración amarilla del albumen de la semilla.
3. El color gris, pardo-grisáceo o pardo-cuero del tegumento de la
semilla, junto con flores rojo-violetas y manchas rojizas en las axilas
foliares.
4. La forma simplemente abultada de la legumbre.
5. La coloración verde de la legumbre no madura, junto, con el mismo
color en el tallo, en las nerviaciones de las hojas y en el cáliz.
6. La distribución de las flores a lo largo del tallo.
7. La longitud del tallo más grande.
Respecto al último carácter, hay que observar que el más largo de los tallos
genitores generalmente es sobrepasado por el híbrido, lo que quizá solo sea
atribuible a la gran exuberancia que aparece en todas las partes de la planta
cuando se combinan tallos de muy diferente longitud. Así, por ejemplo, en
repetidos experimentos, tallos de 1' y 6' de longitud dieron, sin excepción en
combinación híbrida, tallos cuya longitud osciló entre 6 y 71/2'. Los híbridos
relativos al tegumento de la semilla son frecuentemente más punteados;
asimismo, a veces los puntos confluyen en pequeñas manchas violeta-
azuladas. La puntuación también aparece frecuentemente aun cuando falte en
el carácter genitor.
Las formas híbridas relativas a la forma de la semilla y al albumen se
manifiestan inmediatamente después de la fecundación artificial por la mera
influencia del polen extraño. Pueden ser observadas, por tanto, ya en el primer
año de experimento, mientras que todas las demás, evidentemente, no pueden

[132]
serlo sino hasta el año siguiente en aquellas plantas que hayan sido obtenidas
de las semillas producto del cruzamiento.
______________
Die erste Generation der Hybriden.
La primera generación de los híbridos.
En esta generación reaparecen en toda su peculiaridad, juntamente con los
caracteres dominantes, también los recesivos, y, por cierto, en la decisiva
relación media 3:1, de modo que, de cada cuatro plantas de esta generación,
tres presentan el carácter dominante y una el recesivo. Esto vale, sin
excepción, para todos los caracteres que fueron empleados en los
experimentos. La forma rugosa y angulosa de las semillas, la coloración verde
del albumen, el color blanco del tegumento de la semilla y de la flor, las
estrangulaciones en las legumbres, el color amarillo de la legumbre inmadura,
del pecíolo, del cáliz y de las nerviaciones foliares, la inflorescencia en umbela
y el tallo enano reaparecen en la citada relación numérica, sin ninguna
alteración esencial. En ningún experimento fueron observadas formas de
transición.
Puesto que los híbridos provenientes de cruzamientos recíprocos tenían
forma completamente igual y, en su ulterior desarrollo, tampoco se llegó a
advertir ninguna divergencia apreciable, los resultados de cada experimento
en uno y otro sentido pudieron ser reunidos en una sola cuenta. Las relaciones
numéricas que fueron obtenidos para cada par de caracteres diferentes son los
siguientes:
1. Experimento. Forma de las semillas. De 253 híbridos, se obtuvieron
en el segundo año de experimentos 7324 semillas, de las cuales fueron
redondas o redondeadas 5474 y rugosas angulosas 1850. De aquí resulta la
razón 2,96:1.
2. Experimento. Coloración del albumen. 258 plantas dieron 8023
semillas: 6022 amarillas y 2001 verdes; de donde aquellas son a estas en la
razón de 3,01:1.
En estos dos experimentos se obtienen generalmente de cada legumbre
ambas clases de semillas. En legumbres bien formadas, que por termino medio

[133]
contienen 6-9 semillas, ocurrió a veces que todas las semillas fueran redondas
(experimento 1) o que todas fueran amarillas (experimento 2); por el contrario,
nunca fueron observadas en una legumbre más de 5 angulosas o de 5 verdes.
No parece haber diferencia en que la legumbre se desarrolle más pronto o
más tarde en el híbrido, como tampoco en que pertenezca al eje principal o a
una rama lateral. En algunas pocas plantas, en las legumbres primeramente
formadas, solo se desarrollaron semillas aisladas, y estas poseían
exclusivamente uno de ambos caracteres; pero en las legumbres formadas
después se mantuvo la proporción normal. De la misma manera que varía la
distribución de los caracteres en las distintas legumbres, varía en las distintas
plantas. Como ilustración pueden servir los 10 primeros miembros de ambas
series experimentales
1. Experimento 2. Experimento
Forma de las semillas Coloración del albumen
Planta Redondas Angulosas Amarillas Verdes
1 45 12 25 11
2 27 08 32 07
3 24 07 14 05
4 19 10 70 27
5 32 11 24 13
6 26 06 20 06
7 88 24 32 13
8 22 10 44 09
9 28 06 50 14
10 25 07 44 18
Como extremos en la distribución de ambos caracteres de la semilla en una
planta, se observaron en el experimento 1,43 semillas redondas y solo 2
angulosas; finalmente, 14 redondas y 15 angulosas. En el experimento 2,32
semillas amarillas y solo 1 verde, pero también 20 amarillas y 19 verdes.
Estos experimentos son importantes para el establecimiento de las relaciones
numéricas medias, ya que muestran cómo con un número escaso de plantas
de experimento son posibles de promedio muy considerables. En el recuento
de las diferentes semillas hace falta cierta atención, particularmente en el

[134]
experimento 2, ya que en algunas semillas de muchas plantas la coloración
verde del albumen se desarrolla menos y, al pronto, fácilmente puede ser
inadvertida. La causa de la parcial desaparición de la coloración verde no está
en dependencia alguna con el carácter híbrido de la planta, en cuanto que lo
mismo ocurre en la planta genitora; esta peculiaridad se limita solo al individuo
y no se hereda en los descendientes. En plantas lujuriantes este fenómeno fue
observado frecuentemente. Las semillas que durante su desarrollo son
dañadas por los insectos varían de color y forma; sin embargo, con alguna
práctica en la clasificación, es fácil evitar errores -casi no hace falta decir que
las legumbres han de permanecer en la planta hasta que lleguen a estar
completamente maduras y secas, ya que la forma y el color de las semillas no
alcanzan antes su pleno desarrollo- .
3. Experimento. Color del tegumento de la semilla. De 929 plantas, 705
produjeron flores rojo-violetas y tegumentos seminales pardo-grisáceos; 224,
flores blancas y tegumentos seminales blancos. De lo que resulta la relación
3,l5 : l.
4. Experimento. Forma de las legumbres. De 1181 plantas, 882
tuvieron legumbres simplemente abultadas y 299 con estrangulaciones. De
aquí la relación 2,95 :1.
5. Experimento. Coloración de la legumbre inmatura. El número de
plantas de experimento ascendió a 580, de las cuales 428 tuvieron legumbres
verdes y 152 amarillas. De aquí que aquellas estén a estas en la relación
2,82:1.
6. Experimento. Posición de las flores. De 858 casos, las flores fueron
651 veces asilares y 207 terminales. De donde la relación, 3,l4 : l.
7. Experimento. Longitud del tallo. De 1061 plantas, 787 tuvieron el
tallo largo y 277 corto. De aquí la relación 2,84:1. En este experimento las
plantas enanas fueron escogidas cuidadosamente y trasplantadas a parcelas
especiales. Esta precaución era necesaria, porque, de lo contrario, entre sus
trepadoras hermanas no se hubieran desarrollado correctamente (se hubieran
perdido). Son fáciles de distinguir, ya en sus primeros momentos (estadios) del
desarrollo, por su crecimiento rechoncho y sus gruesas hojas verde oscuras.

[135]
Si se reúnen los resultados de todos los experimentos, entre el número de
formas con el carácter dominante y el de formas con el carácter recesivo
resulta como proporción media 2,98:1, o sea, 3:1.
El carácter dominante puede tener aquí un doble significado, a saber: la de
carácter paternal o la de carácter híbrido. Acerca de cuál de ambas
significaciones tenga el carácter en cada caso particular, solo lo permite decidir
(puede hacerse mediante) la generación siguiente.
________________________________
Die zweite Generation der Hybriden
La segunda generación de los híbridos
Aquellas formas que en la primera generación ostentan el carácter
recesivo ya no varían respecto a este carácter en la segunda generación y
permanecen constantes en sus descendientes.
Otra cosa sucede con aquellas formas que tienen el carácter dominante en la
primera generación. De tales formas, dos partes de ellas dan descendientes
que llevan el carácter dominante y el recesivo en la proporción 3:1, de modo
que muestran exactamente el mismo comportamiento que las formas híbridas;
y solo una parte de tales formas permanece constante con el carácter
dominante.
Los distintos experimentos produjeron los siguientes resultados:
1. Experimento. Entre 565 plantas desarrolladas a partir de semillas
redondas de la primera generación, 193 produjeron solo semillas redondas y
permanecieron, por lo tanto, constantes en este carácter; 372, no obstante,
dieron semillas redondas y angulosas en la proporción de 3 a 1. Por lo tanto, el
número de híbridos, comparado con el de constantes, es de 1,93 a 1.

[136]
2. Experimento. De 519 plantas, las cuales fueron obtenidas de semillas
cuyo albumen tenía en la primera generación color amarillo, 166 dieron
exclusivamente semillas amarillas, pero 353 las dieron amarillas y verdes en la
proporción 3:1. Ocurre, por tanto, una distribución en formas híbridas y
constantes según la proporción 2,13:1.
Para cada uno de los distintos experimentos siguientes fueron tomadas 100
plantas de las que en la primera generación poseyeron el carácter dominante,
y de cada una de ellas, a fin de comprobar la significación de este mismo
carácter, fueron cultivadas 10 semillas.
3. Experimento. Los descendientes de 36 plantas tuvieron los tegumentos
seminales pardo-grisáceos; de 64 plantas, se obtuvieron en parte
descendientes de tegumentos seminales pardo-grisáceos, en parte
descendientes de tegumentos seminales blancos.
4. Experimento. Los descendientes de 29 plantas solo tuvieron legumbres
simplemente abultadas; de 71, por el contrario, parte de los descendientes las
tuvieron simplemente abultadas, parte, con estrangulaciones.
5. Experimento. Los descendientes de 40 plantas tuvieron solo legumbres
verdes, los de 60, parte de ellos las tuvieron verdes, parte amarillas.
6. Experimento. Los descendientes de 33 plantas tuvieron flores solamente
axiales, en los de 67, por el contrario, las flores fueron axiales en algunos y
terminales en otros.
7. Experimento. Los descendientes de 28 plantas tuvieron eje largo; los de
72 plantas, algunos lo tuvieron largo, otros corto.
En cada uno de estos experimentos cierto número de plantas fueron
constantes con el carácter dominante. Para apreciar la proporción según la cual
ocurre la separación de las formas con el carácter constante, son de particular
importancia los dos primeros experimentos, porque en ellos cabe comparar
gran número de plantas. Las proporciones 1'93:1–2´13:1 dan reunidas casi
exactamente la proporción promedio 2:1. El experimento 6 tuvo un resultado
completamente concordante. En los otros, la proporción oscila más o menos,
como no cabía esperar otra cosa dado el escaso número de 100 plantas de

[137]
experimento. El experimento 5, que es el que muestra la desviación más
grande, fue repetido, y, entonces, en vez de la relación 60:40, se obtuvo la de
65:35. La relación promedio 2:1 parece, pues, como segura. Con esto resulta
probado que, de aquellas formas que en la primera generación poseen el
carácter dominante, dos terceras partes llevan el carácter híbrido, pero una
permanece constante con el carácter dominante.
La razón 3:1, según la cual ocurre la distribución del carácter dominante
y del recesivo en la primera generación, se descompone, por tanto, para todos
los experimentos en la relación 2:1:1 cuando el carácter dominante se
distingue, según su significación, como carácter híbrido y como carácter
parental. Puesto que los miembros de la primera generación se producen
inmediatamente de las semillas de los híbridos, es pues evidente que los
híbridos de cada dos caracteres diferentes producen semillas, de las cuales
una mitad desarrolla nuevamente la forma híbrida, mientras que la otra da
plantas que permanecen constantes y que en igual número reciben
respectivamente el carácter dominante y el recesivo
______________
Die weiteren Generationen der Hybriden
Las siguientes generaciones de los híbridos
Las relaciones según las cuales los descendientes de los híbridos se
desarrollan y reparten en la primera generación y en la segunda, valen
verosímilmente para todas las generaciones ulteriores. El experimento 1 y el 2
actualmente ya han sido proseguidos durante 6 generaciones; el 3 y 7, durante
5; el 4, 5 y 6, durante 4, si bien, a partir de la generación 3, con pequeño
número de plantas, sin que se advirtiera ninguna desviación. Los
descendientes de los híbridos se distribuyen en cada generación, según la
relación 2:1:1, en híbridos y formas constantes.
Designe A uno de los dos caracteres constantes, por ejemplo, el dominante, a,
el recesivo, y Aa, la forma híbrida en la que ambos están reunidos, entonces la
expresión:

[138]
A + 2Aa + a
proporciona la serie de desarrollo para los descendientes de cualesquiera dos
caracteres diferentes [contrapuestos]2.
La observación hecha por Gärtner, Kölreuter y otros de que los híbridos poseen
la tendencia de revertir a las formas
originarias resulta también confirmada por los experimentos descritos. Se deja
demostrar que el número de los híbridos resultantes de una generación,
respecto al número de formas que vienen a ser constantes y de sus des-
cendientes, disminuye considerablemente de generación en generación, sin
que, sin embargo, pueda desaparecer por completo. Si, como promedio para
todas las plantas en todas las generaciones, se admite igual fertilidad, y si,
además, se considera que cada híbrido produce semillas, de las cuales una
mitad origina nuevamente híbridos, mientras que la otra mitad viene a ser
constante para ambas formas a partes iguales, entonces las relaciones
numéricas para los descendientes en cada generación son dadas por el si-
guiente resumen, en el que A y a designan otra vez los dos caracteres
originarios y Aa la forma híbrida. En gracia a la brevedad admitiremos que cada
planta en cada generación produce solo 4 semillas.
Expresado en proporción
Generacion A Aa a A : Aa : a
1 1 2 1 1 : 2 : 1
2 6 4 6 3 : 2 : 3
3 28 8 28 7 : 2 : 7
4 120 16 120 15 : 2 : 15
5 496 32 496 31 : 2 : 31
n 2n -1 : 2 : 2n -1
2 dos caracteres diferentes/ dos caracteres contrapuestos

[139]
En la décima generación, por ejemplo, 2n-1 = 1.023. Resulta, por tanto, que, de
cada 2.048 plantas que se originen en esta generación, 1.023 tienen el carácter
dominante constante, 1.023 el recesivo constante y solo hay 2 híbridos.
_____________
Die Nachkommen der Hybriden, in welchen mehrere differirende Merkmale
verbunden sind
Los descendientes de los híbridos en los cuales están reunidos varios
pares de caracteres diferentes.
Para los experimentos descritos anteriormente se utilizaron plantas que solo
diferían en un carácter esencial. El problema siguiente consistió en averiguar si
la ley de desarrollo encontrada vale también para cada dos caracteres
diferentes [contrapuestos] cuando varios caracteres distintos están reunidos
mediante fecundación en el híbrido
En lo que concierne a la forma de los híbridos en este caso, los experimentos
mostraron concordantemente que esta siempre está más próxima a aquella de
ambas plantas genitoras que contiene el mayor número de caracteres
dominantes. Tiene, por ejemplo, la planta productora de semillas, eje corto,
flores blancas terminales y legumbres simplemente abultadas; [y] por el
contrario, la planta donante de polen tiene eje largo, flores rojo-violeta axilares
y legumbres con estrangulaciones. Entonces el híbrido recuerda a la planta
productora de semillas solo por la forma de las legumbres, y coincide con la
planta donante de polen en los demás caracteres. Si una de las formas
progenitoras posee solo caracteres dominantes, entonces el híbrido es
prácticamente o completamente indistinguible. Contiene una de ambas formas
progenitoras solo caracteres dominantes, pues el híbrido apenas, o casi nada,
es posible diferenciarlo de ella.
Con gran número de plantas se llevaron a cabo dos experimentos. En el
primer experimento las plantas progenitoras fueron diferentes en la forma de
las semillas y en la coloración del albumen; en el segundo, en la forma de las
semillas, en la coloración del albumen y en el color del tegumento de la

[140]
semilla. Los experimentos con caracteres de la semilla llevan al resultado final
más fácil y seguramente al resultado final.
Para una más fácil comprensión, fueron designados en estos
experimentos los diferentes caracteres de la planta productora de las semillas
con A, B, C; los de la planta donante de polen, con a, b, c; y las formas híbridas
de estos caracteres, con Aa, Bb, Cc.
Primer experimento:
AB Planta productora de las semillas ab Planta donante de polen
A Forma redonda (aspecto redondeado) a Forma angulosa(aspecto rugoso)
B Albumen amarillo b Albumen verde
Las semillas fecundadas fueron redondas y amarillas, semejantes a las de la
planta productora de semillas. Las plantas obtenidas de ellas dieron semillas de
cuatro clases, las cuales frecuentemente aparecieron en una misma legumbre.
En total, de 15 plantas se obtuvieron 556 semillas, de las cuales fueron:
315 redondas y amarillas.
101 angulosas y amarillas.
108 redondas y verdes.
32 angulosas y verdes.
Todas fueron cultivadas el año siguiente. De las semillas redondas amarillas 11
no germinaron y 3 plantas no llegaron a fructificar. De las demás plantas:
38 dieron semillas redondas amarillas...........................AB
65 redondas amarillas y [redondas] verdes..................ABb
60 redondas amarillas y angulosas amarillas..............AaB
138 redondas amarillas, redondas verdes angulosas amarillas y angulosas
verdes....................................................................AaBb
De las semillas angulosas amarillas llegaron a fructifican 96 plantas, de las
cuales:
28 dieron solo semillas angulosas amarillas...................aB
68 dieron semillas angulosas amarillas y semillas angulosas
verdes……………………………………………………………….aBb
De 108 semillas redondas verdes fructificaron 102 plantas, de las cuales:
35 dieron solo semillas redondas verdes..........................Ab

[141]
67 semillas redondas verdes y semillas angulosas
verdes………………………………………………….....Aab
Las semillas rugosas verdes dieron 30 plantas absolutamente iguales;
permanecieron constantes:.......................................................ab
Así pues, los descendientes de los híbridos aparecen de 9 formas diferentes y,
en parte, en números muy desiguales. Si se reúnen y ordenan, resulta:
38 Plantas con la descripción AB
35 “ “ “ Ab
28 “ “ “ aB
30 “ “ “ ABb
65 “ “ “ aBb
68 “ “ “ AaB
60 “ “ “ Aab
67 “ “ “ Aab
138 “ “ “ AaBb
Todas las formas se dejan distribuir en 3 grupos esencialmente discutidos. El
primero comprende la fórmula AB, Ab, aB, ab; estas tienen solo caracteres
constantes y ya no varían en las siguientes generaciones. Cada una de estas
formas está por término medio representada 33 veces. El segundo grupo
contiene las formas ABb, aBb, AaB, Aab; estas son constantes en un carácter,
híbridas en el otro solo varían en la generación siguiente respecto al carácter
híbrido. Cada una de estas aparece por término medio 65 veces. La forma
AaBb ocurre 138 veces, es híbrida en ambos caracteres y se produce
exactamente como el híbrido del cual ella procede.
Si se comparan los números en que aparecen las formas de estos grupos, no
se puede dejar de reconocer la relación media 1:2:4. Los números 33, 65, 138
son valores muy favorablemente aproximados a los números relativos 33, 66,
132
La serie de desarrollo consiste, según esto, en 9 miembros. De ellos, 4 ocurren
en ella una vez cada uno y son constantes en ambos caracteres las formas AB,
ab iguales a las variedades genitoras; las otras dos representan las demás
combinaciones constantes todavía posibles de los caracteres unidos A, a, B, b.
Cuatro miembros ocurren 2 veces cada uno y son constantes en un carácter

[142]
híbrido en el otro. Un miembro aparece 4 veces y es híbrido en ambos
caracteres. Por lo tanto, los descendientes de los híbridos, cuando en estos
están unidos dos clases de caracteres diferentes, se desarrollan según la
expresión:
AB + Ab + aB + ab + 2ABb + 2aBb + 2AaB + 2Aab + 4AaBb
Esta serie de desarrollo es, evidentemente, una serie de combinación en la
que están combinadas miembro a miembro las dos series de desarrollo de los
caracteres A y a, B y b. Se obtienen los miembros de la serie completa
mediante la combinación de las expresiones:
A + 2Aa + a
B + 2Bb + b
Segundo experimento:
ABC planta productora de las semillas abc plantas polen
A forma redonda a forma angulosa,
B albumen amarillo b albumen verde
C tegumento [seminal] pardo-grisáceo c tegumento [seminal] blanco
Este experimento se llevó a cabo de modo completamente análogo al
precedente. Entre todos los experimentos fue el que requirió más tiempo y
trabajo. De 24 híbridos se obtuvieron en total 687 semillas, todas las cuales
fueron punteadas pardo-grisáceo o verde-grisáceo, redondas o angulosas.
De ellas llegaron a la fructificación en el año siguiente 639 plantas y, como
mostraron las investigaciones ulteriores, entre ellas:
08 Plantas ABC 22 Plantas ABCc 45Plantas ABCc
14 “ ABc 17 “ AbCc 36 ” ABCc
09 “ AbC 25 “ aBCc 38 ” ABCc
11 “ Abc 20 “ abCc 40 ” ABCc
08 “ aBC 15 “ ABbC 49 ” ABCc
10 “ aBc 18 “ ABbc 48 ” ABCc
10 “ abC 19 “ aBbC
07 “ abc 24 “ aBbc
14 “ AaBC 78 ” ABCc

[143]
18 “ AaBc
20 “ AabC
16 “ Aabc
La serie de desarrollo comprende 27 miembros, de los cuales: 8 son
constantes en todos los caracteres, y cada uno ocurre 10 veces por término
medio; 12 son constantes en dos caracteres, híbridos en el tercer carácter, y
cada uno aparece 19 veces por término medio; 6 son constantes en un
carácter, híbridos en los otros dos, y cada uno de ellos aparece 43 veces por
término medio; una forma acontece 48 veces y es híbrida en todos los
caracteres. La relación 10:19:43:70 está tan próxima a la relación 10:20:40:80
o 1:2:4:8 que, sin duda, estas últimas representan los valores correctos.
El desarrollo de los híbridos, cuando sus variedades genitoras son diferentes
en tres caracteres, resulta, por tanto, según la expresión:
ABC+ABc+AbC+Abc+aBC+aBc+abC+abc+2ABCc+2AbCc+2aBCc+2abCc+2A
BbC+2ABbc+2aBbC+2aBbc+2AaBC+2AaBc+2AabC+2Aabc+4ABbCc+4aBbCc
+4AaBCc+4AabCc+4AaBbC+4AaBbc+8AaBbCc
También aquí se tiene una serie de combinación en la cual están
reunidas las series de desarrollo para los caracteres A y a, B y b, C y c. Las
expresiones:
A + 2Aa + a
B + 2Bb + b
C + 2Cc + c
dan todos los miembros de la serie. Las combinaciones constantes que
aparecieron en ella corresponden a todas las que son posibles entre los
caracteres A, B, C, a, b, c; dos(,) de ellas, ABC y abc, iguales a las dos plantas
genitoras.
Además, se llevaron a cabo todavía varios experimentos con menor número de
plantas, en los cuales los demás caracteres estaban, dos o tres de ellos, en
reunión híbrida; todos estos otros experimentos dieron resultados aproxima-
damente iguales. No hay duda de que para todos los caracteres empleados en
los experimentos tiene validez la proposición: los descendientes de los híbridos

[144]
en los cuales están reunidos varios caracteres esencialmente distintos
representan los miembros de una serie de combinación en la cual están
reunidas las series de desarrollo de cada dos caracteres diferentes
[contrapuestos]. Con esto resulta probado, al mismo tiempo, que el
comportamiento de cada dos caracteres diferentes en combinación híbrida es
independiente de los demás diferencias en ambas plantas genitoras.
Si n designa el número de diferentes características en ambas plantas
genitoras, entonces 3n es el número de miembros de la serie de combinación,
4n el número de individuos que pertenecen a la serie, y 2n el número de
combinaciones que permanecen constantes. Así, por ejemplo, si las plantas
progenitoras son diferentes en 4 caracteres, la serie contiene 34 = 81
miembros, 44 = 256 individuos y 24 = 16 formas constantes; o lo que es lo mis-
mo, entre cada 256 descendientes del híbrido hay 81 combinaciones distintas,
de las cuales 16 son constantes.
Todas las combinaciones constantes que son posibles en Pisum por
combinación de los 7 citados caracteres también fueron obtenidas
efectivamente mediante cruzamiento repetido. Su número viene dado por 27 =
128. Con esto se proporciona al mismo tiempo la prueba experimental de que
los caracteres constantes que ocurren en diferentes variedades de una especie
de plantas pueden aparecer, mediante fecundación artificial repetida, en todas
combinaciones que son posibles según reglas de la combinación.
Respecto al tiempo de floración de los híbridos, los experimentos todavía
no están terminados. En cuanto a lo que cabe ya decir acerca de ello, aquel
tiempo está casi exactamente en el medio entre el de la planta productora de
las semillas y el de la planta donante del polen, y el desarrollo del híbrido
respecto a este carácter sucede probablemente del mismo modo que para los
demás caracteres del caso. Las formas que se elijan para los experimentos de
esta índole tienen que diferir, por lo menos, 20 días respecto al tiempo medio
de floración; también es necesario que las semillas, al cultivarlas, sean todas
hundidas a igual profundidad en la tierra, a fin de obtener una germinación
simultánea; por último, se deben tener en cuenta durante todo el tiempo de
floración las grandes oscilaciones de temperatura y el retardo o la aceleración
en parte ocasionados por ellas en la floración. Como se ve, este experimento
ha de superar toda clase de dificultades y requiere gran atención.

[145]
Si ensayamos resumir brevemente los resultados obtenidos, encontramos que
aquellos caracteres diferentes que permiten una distinción fácil y segura en las
plantas de experimento siguen en combinación híbrida un comportamiento
completamente correspondiente. Los descendientes de los híbridos de cada
dos caracteres diferentes [contrapuestos] son híbridos la mitad de ellos,
mientras que la otra mitad, por partes iguales, es constante con el carácter de
la planta productora de semillas y respectivamente con el de la donante de
polen. Cuando están reunidos (mediante fecundación) en un híbrido varios
caracteres diferentes, entonces los descendientes de este forman los miembros
de una serie de combinación en la cual están reunidas las series de desarrollo
de cada dos caracteres diferentes.
La completa correspondencia que muestran todos los caracteres sometidos a
experimento permite y justifica la admisión de que tengan también igual
comportamiento los demás caracteres, que aparecen en la planta menos
netamente definidos, y por ello no pudieron ser empleados en especiales
experimentos. Un experimento sobre pedúnculos florales de desigual longitud
dio en conjunto un resultado bastante satisfactorio, a pesar de que la distinción
y clasificación de las formas no se pudo efectuar con aquella seguridad que es
indispensable para experimentos correctos.
______________
Die Befruchtungs-Zellen der Hybriden
Las células reproductoras de los híbridos
Los resultados a los que llevaron los precedentes experimentos sugirieron
ulteriores experimentos, cuyo resultado parece idóneo para sacar conclusiones
acerca de la constitución de las células-germen y de las células-polen de los
híbridos. En Pisum ofrece un importante punto de apoyo la circunstancia de
que entre los descendientes de los híbridos aparecen formas constantes -y, por
cierto, en todas las combinaciones [posibles]- de los caracteres reunidos. Hasta
donde la experiencia llega, encontramos confirmado siempre que

[146]
descendientes constantes solo pueden ser formados cuando las células-
germen y el polen fecundante son de igual índole, de manera que ambas están
dotadas de la capacidad de dar individuos completamente iguales, como es el
caso en la fecundación normal de las especies puras. Tenemos, por tanto, que
considerar como necesario que también en la producción de formas constantes
a partir de la planta híbrida cooperan factores completamente iguales. Puesto
que las distintas formas constantes se producen en una [misma] planta, incluso
en una [misma] flor de la misma [planta], parece consecuente la hipótesis de
que en el ovario de los híbridos se forman tantas clases de células-germen
[vesículas germinales], y en las anteras tantas clases de células-polen, como
formas de combinación constantes son posibles, y que estas células-germen y
células-polen corresponden, según su constitución interna, a las distintas
formas.
De hecho, cabe mostrar teóricamente que esta hipótesis bastaría para explicar
completamente el desarrollo de los híbridos en las distintas generaciones si al
mismo tiempo se pudiera suponer que las distintas clases de células-germen y
células-polen se forman en el híbrido en igual número por término medio.
Para someter a prueba por vía experimental estas suposiciones, se eligieron
los siguientes experimentos: dos formas que eran constantemente diferentes
en la forma de la semilla y en la coloración del albumen fueron unidas por
fecundación.
Si otra vez los diferentes caracteres se designan con las letras A, B, a, b
se tiene:
AB Planta producto de las semillas ab planta donante de polen
A forma redonda a forma angulosa
B albumen amarillo b albumen verde
Las semillas fecundadas artificialmente fueron cultivadas junto con
varias semillas de ambas plantas progenitoras y los ejemplares más vigorosos
[obtenidos] de ellas destinados para recíproco cruzamiento. Se fecundó
1. El híbrido con el polen de AB
2. El híbrido “ “ “ “ ab
3. AB “ “ “ del híbrido

[147]
4. ab “ “ “ del híbrido
Para cada uno de estos 4 experimentos fueron fecundadas todas las
flores de 3 plantas. Si la anterior suposición era cierta, tenían que desarrollarse
en el híbrido células-germen y células-polen de las formas AB, Ab, aB, ab, y
unirse:
1. Las células–germen AB, Ab, aB, ab con las células-polen AB.
2. “ AB, Ab, aB, ab “ ab.
3. “ AB “ AB, Ab, aB, ab.
4. “ ab “ AB, Ab, aB, ab
De cada uno de estos experimentos solo podían pues originarse las
siguientes formas:
1. AB, ABb, AaB, AaBb.
2. AaBb, Aab, aBb, ab.
3. AB, ABb, AaB, AaBb.
4. AaBb, Aab, aBb, ab.
Si, además, las distintas formas de células-germen y de células-polen del
híbrido se formaran en igual número por término medio, las ya indicadas 4
combinaciones tendrían que aparecer en igual relación numérica. Una
completa correspondencia de la relación numérica no era, sin embargo, de
esperar, puesto que en toda fecundación, incluso en la normal, alguna célula-
germen permanece sin desarrollarse o se atrofia luego, y aun muchas de las
semillas bien formadas no llegan a germinar después de la siembra. Después
de todo, la precedente suposición se limita a que en la formación de las
distintas clases de células-germen y células-polen se tiende a iguales números,
[pero] sin que esto se tenga que alcanzar con exactitud matemática.
El primer experimento y el segundo tenían, de modo preferente, el propósito de
comprobar la constitución de las células-germen del híbrido, así como el
tercero y el cuarto habían de resolver [análogamente] acerca de las células-
polen. Como resulta del resumen expuesto anteriormente, el primer

[148]
experimento y el tercero tenían que dar combinaciones completamente iguales;
lo mismo que el segundo y el cuarto; además, el resultado, en la forma y
coloración de las semillas obtenidas por fecundación artificial, debía de ser
parcialmente ya en el segundo año. En el primer experimento y en el tercero
ocurren los caracteres dominantes de forma y de color A y B en cada
combinación, y, por cierto, en parte constantes, en parte en reunión híbrida con
los caracteres recesivos a y b, por lo que tienen que imprimir su peculiaridad a
todas las semillas. En el segundo experimento y en el tercero, por el contrario,
una combinación es híbrida en forma y en color, por lo que las semillas son
redondas y amarillas; otra es híbrida en la forma y constante en el carácter
recesivo del color, por lo que las semillas son redondas y verdes; la tercera es
constante en el carácter recesivo de la forma e híbrida en el del color, de donde
las semillas angulosas y amarillas; la cuarta es constante en ambos caracteres
recesivos, por lo que las semillas son angulosas y verdes. En ambos
experimentos eran de esperar, por lo tanto, cuatro clases de semillas, a saber:
redondas amarillas, redondas verdes, angulosas amarillas, angulosas verdes.
La cosecha correspondió completamente a las exigencias formuladas.
Se obtuvieron:
1 .Experimento: 98 semillas redondas amarilla exclusivamente;
3. “ 94 “ “ “ “ ;
2. Experimento: 31 semillas redondas amarillas, 26 redondas verdes, 27
angulosas amarillas, 26 angulosas verdes.
4. Experimento: 24 semillas redondas amarillas, 25 redondas verdes, 22
angulosas amarillas, 27 angulosas verdes.
Ya apenas cabían dudas de un resultado favorable, la generación siguiente
tenía que deparar la decisión definitiva. De las semillas sembradas llegaron a la
fructificación en el año siguiente 90 plantas en el primer experimento, 87 en el
tercero; de estas, produjeron en él
experimento
1. 3.
20 25 semillas redondas amarillas...........……………………AB
23 19 semillas redondas amarillas y verdes……………….ABb
25 22 semillas redondas amarillas y angulosas.. . . . ….. AaB
22 21 semillas redondas y angulosas, amarilla y verde... AaBb

[149]
En el experimento segundo y en el cuarto las semillas redondas y amarillas
dieron plantas con semillas redondas y angulosas, amarillas y
verdes........................................................................................... AaBb
De las semillas redondas verdes se obtuvieron plantas con semillas redondas y
angulosas verdes............................................................................ .Aab
Las semillas amarillas angulosas dieron plantas con semillas angulosas,
amarrillas y verdes .......................................................................... aBb
De las semillas angulosas verdes salieron plantas que nuevamente solo
produjeron semillas angulosas verdes ............................................ ab
Aunque también en estos experimentos no germinaron algunas
semillas, los números hallados en el año anterior no tuvieron que ser alterados
por ello, puesto que cada clase de semillas dio plantas que, en lo que se refiere
a las semillas, fueron iguales entre sí, y diferentes de las demás. Así pues,
produjeron:
2. el experimento 4.el experimento
31 24 Plantas, semillas de la forma AaBb
26 25 “ “ “ “ Aab
27 22 “ “ “ “ aBb
26 27 “ “ “ “ ab
En todos los experimentos aparecieron, por lo tanto, todas las formas que la
suposición formulada exigió, y, por cierto, en números aproximadamente igual.
En otra prueba fueron tomados en los experimentos los caracteres del
color de las flores y la longitud del eje, y la elección se hizo de modo que en el
tercer año del experimento cada carácter tuviese que aparecer en la mitad de
todas las plantas, en caso de que la anterior hipótesis fuese acertada. A, B, a, b
sirven otra vez para designar los distintos caracteres.
A Flores rojo-violeta a Flores blancas
B Eje largo b Eje corto
La forma Ab fue fecundada con ab, de donde resultó el híbrido Aab. Por
último, fue fecundada aB también con ab, de donde el híbrido aBb. En el
segundo año fueron empleados para las siguientes fecundaciones el híbrido

[150]
Aab como planta productora de las semillas, el otro aBb como planta donante
del polen.
Planta productora de las semillas Aab, Planta donante del polen aBb.
Posibles células-germen Ab, ab, Posibles células-polen aB, ab.
De la fecundación entre las posibles células-germen y células-polen
tenían que resultar cuatro combinaciones, a saber:
AaBb + aBb + Aab + ab
Por lo tanto, es evidente que, según la anterior suposición, en el tercer
año, de todas las plantas la mitad debería tener:
La mitad, Flores rojo violeta (Aa)......................................Términos :1 .3
“ flores blancas (a)..................................................... ” : 2 . 4
“ un eje largo (Bb)..................................................... ” : 1 . 2
“ un eje largo (b)........................................................ ” : 3. 4
De las 45 fecundaciones del segundo año se obtuvieron 187 semillas, de las
cuales llegaron a florecer en el tercer año 166 plantas. Entre ellas aparecieron
los distintos términos en los siguientes números:
Término Color de las flores Eje
1 Rojo-violeta Largo.........47 veces
2 Blanco Corto..........40 “
3 Rojo-violeta Largo.........38 “
4 Blanco Corto..........41 “
Por tanto, apareció: El color rojo-violeta de las flores (Aa) en 85 plantas
“ El color blanco de las flores (a) ” 81 plantas
“ El eje largo (Bb) ” 87 ”
“ El eje corto (b) ” 79 ”
La idea propuesta encuentra también en este experimento una confirmación
satisfactoria.

[151]
Para los caracteres de forma de la legumbre, color de la legumbre y posición
de las flores se dispusieron también experimentos en pequeña escala y se
obtuvieron resultados completamente concordantes. Todas las combinaciones
que eran posibles por la reunión de los diferentes caracteres resultaron
puntualmente y en números aproximadamente Iguales.
Resulta, pues, confirmada por vía experimental la hipótesis de que las plantas
de guisante híbridas forman células-germen y células-polen que, por su
constitución, corresponden en igual número a todas las formas constantes que
resultan de la combinación de los caracteres reunidos por la fecundación.
La diversidad de las formas entre los descendientes de los híbridos, así como
las relaciones numéricas en las cuales estos mismos se observan, encuentran
en la proposición, arriba enunciada, una explicación satisfactoria. El caso más
simple lo ofrece la serie de desarrollo para dos caracteres diferentes
[contrapuestos]. Esta serie viene expresada, como es sabido, por la expresión
A+2Aa+a, en la que A y a representan las formas con los diferentes caracteres
constantes y Aa la forma híbrida de ambos. Tal expresión encierra dentro de
tres términos cuatro individuos. En la formación de estos, intervienen en la
fecundación células-polen y células-germen de la forma A y de la a, por término
medio en igual número de una forma que de la otra, por tanto, cada forma dos
veces, puesto que se forman cuatro individuos. Así pues, participan en la
fecundación:
las células-polen A+A+a+a
las células-germen A+A+a+a.
Queda completamente al azar cuál de las dos clases de polen se combina con
cada distinta célula-germen. Sin embargo, según las leyes de probabilidades,
ocurrirá que, por término medio, cada una de las dos formas de polen, A y a, se
reunirá tantas veces con una como con la otra de las dos formas de células-
germen, A y a; por tanto, en la fecundación una de ambas células-polen A se
encontraría con una célula-germen A; la otra, con una célula-germen a; y,
asimismo, se unirá una célula-polen a con una célula-germen A, y la otra, con
una célula-germen a.

[152]
El resultado de la fecundación se puede representar escribiendo en forma de
quebrado los símbolos de las células-polen y de las células-germen que se
combinan: los de las células-polen encima de la raya, los de las células-germen
debajo. Se obtiene en el presente caso:
a
a
A
a
a
A
A
A
En el primer término y en el cuarto célula-polen y célula-germen son
iguales, por lo tanto, el producto de su combinación tiene que ser constante, a
saber, A y a; por el contrario, en el segundo y en el tercero ocurre una reunión
de los dos caracteres parentales diferentes, de aquí que las formas A
ay
a
A
resultantes de estas fecundaciones sean completamente idénticas al híbrido
del cual ellas proceden. Ocurre, pues, una hibridación repetida. Esto explica el
notable fenómeno de que los híbridos sean capaces de reproducir, juntamente
con las dos formas originarias, también descendientes iguales a ellos mismos;
dan
ambos la misma combinación Aa, puesto que, como ya se ha dicho, es
indiferente para el resultado de la fecundación a cuál de ambos caracteres
corresponde la célula-polen o la célula-germen. Por lo tanto:
Así, de esta forma por término medio, transcurre la autofecundación de
los híbridos cuando en ellos están reunidos dos caracteres diferentes. Sin
embargo, en flores o en plantas por separado, la relación según la cual se
producen las formas de la serie puede experimentar desviaciones no sin
importancia. Aparte de que los números en que se presentan en el ovario
aAaAa
a
A
a
a
A
A
A 2

[153]
ambas clase de células-germen solo en promedio pueden ser consideradas
iguales, queda por completo relegado al azar cuál de ambas clases de células-
polen fecunda efectivamente a cada célula-germen en particular. Por esta
razón, los valores particulares tienen necesariamente que estar sometidos a
oscilaciones y aun ser posibles valores extremos, como antes, a propósito de
los experimentos sobre la forma y la coloración del albumen, que antes
hubieron de ser expuestos. Las relaciones numéricas verdaderas solo pueden
ser dadas por el promedio obtenido de la suma del mayor número posible de
valores particulares: cuanto mayor sea su número, tanto más exactamente será
eliminado el puro azar.
La serie de desarrollo de los híbridos en los cuales están combinados dos
clases de caracteres diferentes encierra [contiene] entre 16 individuos 9 formas
distintas, a saber: AB+Ab+aB+ab+2ABb+2aBb+2AaB+2Aab+4AaBb. Entre los
distintos caracteres de las plantas genitoras A, a y B, b son posibles 4
combinaciones constantes, por lo que el híbrido produce también las
correspondientes 4 formas de células-germen y de células-polen: AB, Ab, aB,
ab, y cada una de ellas intervendrá en la fecundación 4 veces como promedio,
puesto que en la serie están comprendidos 16 individuos. Por lo tanto,
participan en la fecundación:
Células-polen: AB + AB + AB + AB + Ab + Ab + Ab + Ab + aB +
aB + aB + aB + ab + ab + ab + ab
Células-germen: AB + AB + AB + AB + Ab + Ab + Ab + Ab + aB +
aB + aB + aB + ab + ab + ab + ab
En promedio, en el transcurso de la fecundación, cada forma de célula-polen se
combina con cada forma de célula-germen con la misma frecuencia, por lo
tanto, cada una de las 4 células-polen AB se combina una vez con [cada] una
de las clases de células-germen AB, Ab, aB, ab. Exactamente lo mismo ocurre
con la reunión de las demás células-polen de las formas Ab, aB, ab con todas
las otras células-germen. Se obtiene por tanto:
ab
ab
aB
ab
Ab
ab
AB
ab
ab
aB
aB
aB
Ab
aB
AB
aB
ab
Ab
aB
Ab
Ab
Ab
AB
Ab
ab
AB
Ab
AB
Ab
AB
AB
AB

[154]
AB + ABb + AaB + AaBb + ABb + Ab + AaBb + AaB + AaBb + aB + aBb + AaBb + Aab + aBb +
ab + AB + Ab + aB + ab + 2ABb + 2aBb + 2AaB + 2Aab + 4AaBb
De manera completamente análoga se explica la serie de desarrollo de los
híbridos cuando en estos están reunidos caracteres de tres diferentes clases.
El híbrido produce 8 formas distintas de células-germen y de células-polen:
ABC, ABc, AbC, Abc aBC aBc, abC abc, y cada forma de polen se combina,
por término medio, una vez con cada forma de célula-germen.
La ley de la combinación de los caracteres diferentes, según la cual ocurre el
desarrollo de los híbridos, encuentra, por tanto, su fundamento y explicación en
las citadas célula-germen y en la proposición de que los híbridos producen
células-polen, las cuales corresponden en igual número a todas las formas
constantes que resultan de la combinación de los caracteres reunidos mediante
la fecundación.
______________
Versuche ubre die Hybriden anderer Pflnzenarten
Experimentos sobre los híbridos de otras especies de plantas.
Será el tema de ulteriores experimentos averiguar si la ley de desarrollo hallada
en Pisum tiene también valor para los híbridos de otras plantas. Con este fin
fueron emprendidos recientemente varios experimentos. Están terminados dos
pequeños experimentos con especies de Phaseolus, los cuales pueden
encontrar aquí mención.
Un experimento con Phaseolus vulgaris y Phaseolus nanus L. dio un resultado
completamente concordante. Ph. nanus tenía, juntamente con el eje enano,
legumbres verdes simplemente abultadas; Ph. vulgaris, por el contrario, un eje
de 10-12` de altura y legumbres de color amarillo, con estrangulaciones a la
madurez. Las relaciones numéricas en que aparecieron las diferentes formas
en distintas generaciones fueron las mismas que en Pisum. El desarrollo de las
combinaciones constantes se produjo también según la ley de la simple

[155]
combinación de caracteres, exactamente como en el caso de Pisum. Se
obtuvieron:
Combinaciones Coloración de la Forma de la
constantes Eje legumbre inmadura legumbre madura
1 corto verde abultada
2 “ “ con estrangulaciones
3 “ amarillo abultada
4 “ “ con estrangulaciones
5 largo verde abultada
6 “ “ con estrangulaciones
7 “ amarillo abultada
8 “ “ con estrangulaciones
El color verde de las legumbres, la forma abultada de la legumbre y el eje alto
fueron, como en Pisum, caracteres dominantes.
Otro experimento con dos especies muy diferentes de Phaseolus tuvo
éxito solo en parte. Como planta productora de las semillas sirvió Ph. nanus L.,
una especie completamente constante con flores blancas en racimos cortos y
pequeñas semillas blancas en legumbres rectas, abultadas y lisas; como planta
donante del polen, Ph. multiflorus W., con tallo alto voluble, flores rojo-
purpúreas en racimos muy largas, legumbres ásperas, encorvadas en forma de
hoz y semillas gruesas, que están como flameadas y manchadas de negro
sobre fondo rojo flor de albérchigo.
El híbrido tuvo la mayor semejanza con la planta donante del polen, solo las
flores parecían coloreadas menos intensamente. Su fructificabilidad fue muy
restringida: de 17 plantas, que en conjunto produjeron varios cientos de flores,
solo se recogieron 49 semillas. Estas fueron de tamaño mediano y presentaban
un dibujo análogo al de Ph. multiflorus; tampoco el color de fondo fue
esencialmente diferente. Al año siguiente se obtuvieron de ellas 44 plantas, de
las cuales solo 31 llegaron a florecer. Lo caracteres de Ph. nanus, los cuales
permanecieron completamente latentes en el híbrido, volvieron a aparecer en
diferentes combinaciones, sin embargo, la relación numérica de ellos con los

[156]
dominantes, en el escaso número de plantas del experimento, quedó muy
desviado; en ciertos caracteres, como los del eje y de la forma de las
legumbres, fueron, como en Pisum, casi exactamente 1:3.
El resultado de este experimento, si bien escaso para la comprobación de las
relaciones numéricas en que ocurren las distintas formas, ofrece, por otro lado,
el caso de un notable cambio de coloración en las flores y las semillas de los
híbridos. Como es sabido, en Pisum los caracteres de color de las flores y
semillas aparecen inalterados en la primera generación y en las siguientes, y
los descendientes de los híbridos llevan exclusivamente uno u otro de los dos
caracteres originarios. Las cosas son de otra manera en este experimento. El
color blanco de las flores y de las semillas de Ph. nanus apareció ciertamente
igual en la primera generación de un ejemplar bastante fértil, pero las 30
plantas restantes desarrollaron coloraciones florales que representan las
diferentes graduaciones desde el rojo púrpura al violeta pálido. La coloración
del tegumento seminal fue no menos variada que la de la flor. Ninguna planta
resultó completamente fértil; muchas casi no dieron ningún fruto; en otras estos
no se desarrollaron sino en las últimas flores y no llegaron nunca a madurar;
solo de 15 plantas se obtuvieron semillas bien conformadas. La tendencia
mayor a la esterilidad la mostraron las formas con flores preponderantemente
rojas, en las cuales de 16 plantas solo 4 dieron semillas maduras. Tres de ellas
tenían un dibujo en las semillas análogamente como Ph. multiflorus, pero un
color de fondo más o menos pálido; la cuarta planta sólo dio una semilla de
coloración simplemente parda. Las formas con coloración floral
predominantemente violeta tenían semillas pardo oscuras, pardo negruzcas y
completamente negras.
El experimento fue proseguido todavía durante dos generaciones en
circunstancias igualmente desfavorables, pues incluso entre los descendientes
de plantas bastante fértiles nuevamente había una parte menos fértil o
completamente estéril. No aparecieron otros colores de flores y de semillas
más que los ya citados. Las formas que en la primera generación ostentaron
uno o varios caracteres recesivos permanecieron, respecto a ellos, constantes
sin excepción. De aquellas plantas que tenían flores violetas y semillas pardas
o negras, algunas ya no variaron el color de las flores y el de la semillas en las
generaciones siguientes, sin embargo, la mayoría también produjo, al lado de

[157]
descendientes completamente iguales, otros que presentaban flores blancas y
tegumentos seminales coloreados. Las plantas con flores rojas permanecieron
tan poco fértiles que nada cabe decir con certeza acerca de su ulterior
desarrollo.
A pesar de los muchos inconvenientes con los que tuvo que luchar la
observación, todavía resulta de este experimento que, respecto a aquellos
caracteres que atañen a la forma de la planta, el desarrollo de los híbridos
transcurre según la misma ley que en Pisum. Respecto a los caracteres de
coloración parece,
por cierto, difícil hallar una concordancia satisfactoria. Prescindiendo de que de
la unión de una coloración blanca con otra rojo-púrpura resulta toda una serie
de coloraciones, desde el púrpura hasta el violeta pálido y blanco, tiene que
llamar la atención la circunstancia de que entre 31 plantas que florecieron solo
una tuvo el carácter recesivo de la coloración blanca, en tanto que en Pisum
eso ocurre, por término medio, cada cuatro plantas.
Pero también estos enigmáticos fenómenos se dejarían explicar
verosímilmente, según la ley válida para Písum, si se pudiera suponer que el
color de las flores y el de las semillas de Ph. multiflorus está compuesto por
dos o más colores completamente independientes, que se comportan
separadamente, lo mismo que cualquier otro carácter constante de la planta. Si
el color A de la flor estuviese compuesto de los caracteres independientes A1 +
A2+..., los cuales produjesen la impresión total de color rojo púrpura y entonces
por fecundación con el contrapuesto carácter del color blanco a tendrían que
formarse las combinaciones híbridas A1a+ A2a+..., y análogamente ocurriría
con la correspondiente coloración del tegumento de la semilla. Según las
anteriores suposiciones, cada una de estas combinaciones de color híbridas
sería independiente y, por consiguiente, se desarrollaría completamente
independiente de las demás. Fácilmente se ve que de la combinación de
distintas series de desarrollo tendría que resultar una completa serie de
coloraciones.
Si, por ejemplo, fuera A = A1 + A2, corresponderían a los híbridos A1a y A2a las
series de desarrollo:
A1 + 2A1a + a
A2 + 2A2a + a

[158]
Los miembros de estas series pueden aparecer en 9 combinaciones distintas y
cada una de ellas representar la designación para un color diferente.
1 A1 A2 2 A1 a A2 1 A2 a,
2 A1 A2 a 4 A1 a A2 a 2 A2 a a,
1 A1 a 2 A1 a a 1 a a
Los números que preceden a las distintas combinaciones indican cuántas
plantas con la coloración correspondiente pertenecen a la serie. Puesto que la
suma de ellas asciende a 1, todas las coloraciones están distribuidas por
término medio en 16 plantas, aunque como la serie misma muestra, en muy
diferentes proporciones
Si el desarrollo de la coloración ocurriera realmente de esta manera,
podría encontrar también explicación el caso antes citado, a saber, que el color
blanco de las flores y tegumentos apareciera solo una vez entre 31 plantas de
la primera generación. Esta coloración está contenida en la serie solo una vez,
y, por lo tanto, en promedio solo podría manifestarse una vez cada 16 plantas,
y en tres caracteres de color solo una vez entre 64 plantas.
No se debe olvidar que la explicación que es propuesta descansa en una
mera presunción que no tiene más a su favor que el muy incompleto resultado
del experimento citado más arriba. Por lo demás, sería un trabajo provechoso
seguir el desarrollo de la coloración de los híbridos mediante análogos
experimentos, pues es verosímil que por este camino podamos comprender la
extraordinaria variedad de la coloración de nuestras flores de adorno.
Hasta ahora, con certeza, apenas se sabe más que el color de las flores de
nuestras plantas ornamentales es un carácter extremadamente variable.
Frecuentemente se ha manifestado la opinión de que el cultivo conmueve en
alto grado o rompe por completo la estabilidad de las especies, y hay mucha
inclinación a suponer que el desarrollo de las formas de cultivo es un desarrollo
irregular y aleatorio; a este respecto, la coloración de las plantas de adorno es
señalada ordinariamente como ejemplo de la mayor inestabilidad. Sin embargo,
no se comprende por qué el simple trasplante al jardín haya de tener como
consecuencia una revolución tan profunda y persistente en el organismo-
planta. Nadie querrá sostener seriamente que el desarrollo de la planta se rige
en campo libre por leyes distintas a las del jardín. Aquí como allí tienen que

[159]
aparecer típicas modificaciones cuando las condiciones de vida para una
especie son alteradas y esta especie posee la facultad de adaptarse a las
nuevas condiciones. Se concederá de buen grado que el cultivo favorece el
origen de nuevas variedades y que por la mano del hombre se conservan
muchas alteraciones que habrían de sucumbir en estado libre, pero nada nos
autoriza a admitir que la tendencia a la formación de variedades sea exaltada
tan extraordinariamente que las especies pierdan de inmediato toda su
estabilidad y sus descendientes diverjan en una serie infinita de formas
extremadamente cambiantes. Si la alteración de las condiciones de vegetación
fuera la única causa de la variabilidad, cabría esperar que aquellas plantas de
cultivo que han sido cultivadas durante cientos de años en condiciones casi
iguales recuperasen su estabilidad. Como es sabido, no es este el caso, puesto
que precisamente entre estas condiciones se encuentran no solo las formas
más diferentes, sino también las más cambiantes. Solo las leguminosas como
Písum, Phaseolus, Leus, cuyos órganos de reproducción están protegidos por
la quilla, constituyen, a este respecto, una notable excepción. Incluso aquí,
durante un cultivo más que milenario en las más diversas condiciones se han
originado numerosas variedades, las cuales mantienen, sin embargo, bajo
condiciones igualmente constantes, una estabilidad como la de las especies
que crecen silvestres.
Resulta más que verosímil que en la variabilidad de las plantas de cultivo actúe
un factor al que hasta ahora se le ha concedido poca atención. Diferentes
experimentos nos llevan a la opinión de que nuestras plantas de cultivo, con
pocas excepciones, son miembros de diferentes series híbridas cuyo regular
desarrollo ulterior es alterado y retardado por frecuentes ínter cruzamientos.
No se debe pasar por alto la circunstancia de que las plantas cultivadas la
mayoría de las veces están en gran número unas al lado de otras, de modo
que se depara la más favorable ocasión para la fecundación recíproca entre las
variedades presentes e incluso entre las especies. La verosimilitud de este
aserto está apoyada por el hecho de que entre la gran multitud de cambiantes
formas siempre se encuentran algunas que permanecen constantes en uno u
otro carácter solo con impedir cuidadosamente todo influjo extraño. Estas
formas se desarrollan exactamente lo mismo que ciertos miembros de las
complejas series híbridas. Incluso el más sensible de todos los caracteres, el

[160]
de color, no puede escapar a la observación cuidadosa que en las distintas
formas la tendencia a la variabilidad se presenta en muy diferente grado. Entre
plantas que proceden de una fecundación espontánea, frecuentemente hay
algunas cuyos descendientes divergen ampliamente en la calidad y disposición
de los colores, mientras otras dan formas menos desviadas, y entre un gran
número se encuentran algunas que transmiten sin cambio el color de sus flores
a sus descendientes. Las especies de Dianthus cultivadas proporcionan a este
respecto un instructivo ejemplo. Un ejemplar de Dianthus Caryophylus de flores
blancas, que procedía de una variedad de flores blancas, fue aislado durante la
floración en un invernadero; las numerosas semillas obtenidas de él dieron
plantas con flores de color blanco exactamente igual. Un resultado análogo se
obtuvo con una subespecie de flores rojas con visos violetas y una de flores
blancas con estrías rojas. Muchas otras, por el contrario, que fueron protegidas
de la misma manera dieron descendientes, más o menos diferentes,
coloreados y marcados con dibujos.
Cualquiera que considere las coloraciones que resultan en las plantas
ornamentales de igual fecundación no podrá sustraerse fácilmente a la
convicción de que también en ese caso el desarrollo ocurra según determinada
ley, la cual posiblemente encuentre su expresión en la combinación de varios
caracteres de coloración independientes.
______________
Schluss-Bemerkungen
Consideraciones finales
No carece de interés comparar las observaciones hechas en Pisum con los
resultados de las investigaciones a los que han llegado las dos autoridades en
este campo, Kölreuter y Gärtner. Según el punto de vista concordante de
ambos, la apariencia externa de los híbridos será, o bien la forma intermedia
entre ambas especies originales, o bien se asemeja más a un tipo u otro, a
veces incluso resultando casi imposible diferenciarlos. De las semillas de los
mismos salen generalmente, cuando la fecundación ha ocurrido con el propio
polen, distintas formas que difieren del tipo normal. Por norma, la mayoría de
los individuos de una misma fecundación mantienen la forma de los híbridos,

[161]
mientras que otros pocos van siendo similares a la planta productora de
semillas y algún que otro individuo se asemeja a la planta productora de polen.
Esto no es válido para todos los híbridos sin excepción. En unos pocos casos,
unos descendientes se han acercado más a una planta originaria y otros a la
otra, o bien se inclinan todos más hacia un lado u otro; en algunos casos, sin
embargo, permanecen completamente idénticos a los híbridos y se reproducen
sin cambios. Los híbridos de las variedades se comportan como los híbridos de
las especies, solo que tienen una todavía mayor variabilidad de la apariencia y
una mayor tendencia a volver a los tipos originales.
En relación a la forma de los híbridos y su desarrollo, por norma general es
inconfundible la concordancia con las observaciones hechas en Pisum. De otra
manera se comportan los casos excepcionales anteriormente nombrados.
Gärtner mismo confiesa que la determinación exacta acerca de si una forma se
parece más a una u otra de ambas plantas originales produce muchas veces
grandes problemas, ya que depende mucho de la percepción del observador.
Pero pudo haber otra circunstancia que hiciera que los resultados, aun
mediante observaciones y diferenciación cuidadosísimas, fuesen fluctuantes e
inciertos. Para los experimentos se emplearon mayoritariamente plantas que se
tienen por buenas especies y que se diferencian en un mayor número de
caracteres. Además de la aparición de los caracteres bien definidos, donde, por
lo general, se trata de una mayor o menor semejanza, aparecen aquellos
caracteres que son difíciles de expresar en palabras, pero que son suficientes,
como todo conocedor de plantas sabe, para dotar a las formas de una
apariencia extraña. Si suponemos que el desarrollo de los híbridos se sucede
de acuerdo con las leyes válidas para Pisum, entonces cada serie de cada
experimento tiene que abarcar muchas formas, ya que el número de términos,
como es conocido, aumenta en potencias de 3 con el número de los caracteres
diferentes. Por lo tanto, con un número relativamente pequeño de plantas de
experimentación, el resultado solo puede ser aproximadamente correcto y en
unos pocos casos diferir notablemente. Si, por ejemplo, las dos especies
originarias se diferencian en 7 caracteres, y si de las semillas de sus híbridos,
para esclarecer el grado de parentesco de los descendientes, se cultivan entre
100 y 200 plantas, entonces vemos con facilidad cuán inseguro es el resultado,
ya que para 7 caracteres diferentes la serie de desarrollo contiene 16384

[162]
individuos en 2187 formas distintas. Podría predominar un parentesco u otro
según el azar hiciese llegar a las manos del observador unas u otras formas en
mayor número.
Si entre los caracteres diferenciadores aparecen a su vez caracteres
dominantes, los cuales pasan a los híbridos sin o casi sin cambios, entonces en
los términos de la serie de desarrollo tiene que predominar aquel progenitor
que tiene un mayor número de caracteres dominantes. En los experimentos
anteriores hechos en Pisum con 3 caracteres diferenciadores, los caracteres
dominantes pertenecían por completo a la planta germen. Aunque los términos
de la serie en su composición interior se inclinan de forma regular hacia las dos
especies originarias, la planta germen obtuvo en este experimento una
preponderancia tan alta que, entre 64 plantas de la primera generación, 54
eran idénticas a ella o solo se diferenciaban en un carácter. Se ve cuán
arriesgado puede ser en ciertas circunstancias sacar conclusiones mediante los
parecidos externos de su parentesco interior.
Gärtner comenta que en aquellos casos donde el desarrollo era regular, entre
los descendientes de los híbridos no se mantienen ambas especies originarias,
sino solamente unos pocos individuos de parentesco más cercano. En series
de desarrollo más amplias no podía ser de otra manera. Para siete caracteres
diferenciadores, por ejemplo, por cada 16000 descendientes de los híbridos
aparecen las dos especies originarias solamente una vez. Por tanto, no es fácil
obtener las mismas con un número pequeño de plantas de experimentación;
sin embargo, hay que contar con una probabilidad pequeña: con la aparición de
algunas formas que se encuentran cercanas a las mismas en la serie.
Nos encontramos con una diferencia fundamental en aquellos híbridos que se
mantienen constantes en sus descendientes y que se reproducen como las
especies puras. Según Gärtner, a estos pertenecen los híbridos
excepcionalmente fértiles: Aquilegia atropurpurea-canadensis, Lavatera
pseudolbia-thuringiaca, Geum urbano-rivale y algunos híbridos de Dianthus; y
según Wichura, los híbridos de especies del sauce. Para la evolución de las
plantas, esta circunstancia es de gran importancia, porque los híbridos
constantes obtienen el significado de especies nuevas. La corrección del
hecho está apoyada por observadores extraordinarios y no puede ser puesta

[163]
en duda. Gärtner tuvo la oportunidad de estudiar a Dianthus Armeria-deltoides
hasta la décima generación, ya que se reproducía en el jardín de forma
autónoma.
En Pisum se comprobó, mediante experimentos, que los híbridos forman
células germen y células polen de naturaleza diferente y que aquí reside el
motivo de la variabilidad de sus descendientes. También en otros híbridos
cuyos descendientes se comportan de forma parecida podemos aventurar que
la causa es la misma; para aquellos, sin embargo, que permanecen constantes,
parece aceptable la suposición de que las células fecundadoras son de la
misma clase y concuerdan con las células básicas de los híbridos. Según la
opinión de famosos fisiólogos, en las fanerógamas3 se unen para la
reproducción una célula polinizadora y otra germinadora en una única célula, la
cual, mediante asimilación de materia y la formación de nuevas células, puede
seguir desarrollándose hacia un organismo autónomo.
Este desarrollo se sucede según una ley constante, la cual está razonada en la
composición material y el orden de los elementos, que en la célula llegan a una
fusión viable. Si las células reproductoras son de la misma clase y concuerdan
con la célula básica de la planta madre, entonces el desarrollo del nuevo
individuo se regirá por la misma ley que vale para la planta madre.
Si se consigue unir una célula germinadora con una polinizadora de una
especie distinta, entonces tenemos que suponer que entre aquellos elementos
de ambas células que determinan las diferencias sucede algún tipo de
compensación. La célula mediadora que sale será la base del organismo
híbrido, cuyo desarrollo necesariamente se guiará por otra ley distinta a la de
cada una de las especies originarias.
________________________
3. En Pisum está fuera de dudas que para la formación del nuevo embrión debe tener lugar una unión
perfecta de los elementos de ambas células reproductoras. ¿Cómo podríamos, sino, explicar que en la
descendencia de los híbridos reaparezcan ambos tipos originales en igual número y con todas sus
peculiaridades? Si la influencia de la célula huevo sobre la célula del polen fuese solo externa y
representara solo un papel de nodriza, entonces el resultado de cada fecundación artificial no podría ser
otro que el híbrido se pareciera exactamente al progenitor productor del polen o, por lo menos, que esto
fuese casi así. Esto no lo han confirmado de ninguna manera los experimentos. Una prueba evidente de la
completa unión del contenido de ambas células es la experiencia adquirida, en todos sentidos, de que no
afecta a la forma de los híbridos cuál de las especies originales sea el progenitor productor de las semillas y
cuál el productor del polen.

[164]
Si tomamos la compensación como completa, en aquel sentido en que el
embrión híbrido se forma de células de la misma clase, en el cual las
diferencias se transmiten de forma completa y permanente, entonces se
sucedería como otra conclusión: que los híbridos, como cualquier otra especie
autónoma de planta, se mantendrían constantes en sus descendientes. Las
células reproductoras que se forman en las anteras y en los vasos seminales
son de la misma clase y concuerdan con las células mediadoras.
En Pisum está fuera de toda duda que, para la creación del nuevo embrión,
tiene que tener lugar la unión completa de los elementos de ambas células
fecundadoras. Cómo se podría explicar, sino, que entre los descendientes de
los híbridos aparecen otra vez ambas especies originarias en igual número y
con las mismas peculiaridades. Si la influencia de la célula germen sobre la
célula polen solo fuera exterior, de manera que le correspondiese solo un papel
de ama de cría, entonces el éxito de cualquier fecundación artificial no podría
ser otro que los híbridos desarrollados solo se pareciesen a la planta
polinizadora o que fueran muy parecidos. Esto no lo apoyan de ninguna
manera los experimentos realizados hasta ahora. Una prueba definitiva de la
completa unión del contenido de ambas células se encuentra en la experiencia
aceptada por todos de que para la forma de los híbridos carece de importancia
cuál de las especies originarias fue la planta polinizadora o la planta germen.
Con respecto a aquellos híbridos cuyos descendientes son variables, se podría
llegar a suponer que entre los elementos diferenciadores de la célula polen y la
germen sucede una mediación tal que es posible la creación de una célula
originaria para los híbridos, pero que la compensación de los elementos
opuestos solo sería de corta duración y no perdurarían más allá de la vida de la
planta híbrida. Como en el desarrollo de la misma durante el periodo vegetativo
no se detectan cambios, tenemos que concluir que los elementos
diferenciadores solo logran salir del enlace obligado durante la formación de las
células fecundadoras. En la creación de estas células toman parte, en un orden
libre y regular, todos los elementos existentes, donde solo se excluyen
mutuamente los diferenciadores. De esta manera se podrían suceder la
creación de tantas células germen y polen como combinaciones diferentes
permitiesen los elementos creadores.

[165]
El intento de explicar que las diferencias fundamentales en el desarrollo de los
híbridos se deben a una unión permanente o transitoria de los elementos
celulares diferenciadores, por supuesto solo puede tener el valor de una
hipótesis, para cuya comprobación, teniendo en cuenta la falta de datos
seguros, aún queda un amplio trabajo. Alguna legitimidad para el punto de vista
comentado se encuentra en las pruebas llevadas a cabo en Pisum: según
estas el comportamiento de dos caracteres diferenciadores en unión híbrida es
independiente de las otras diferencias entre las dos especies originarias; y más
todavía, los híbridos forman tantas células germen y polen como formas de
combinación constantes son posibles. Los caracteres diferenciadores de dos
plantas, finalmente, solo pueden basarse en las diferencias en la composición y
agrupamiento de los elementos que se encuentran en interacción vital en las
células originarias de las mismas.
La validez de lo dicho para Pisum, sin embargo, necesita una comprobación, y,
por tanto, sería deseable una repetición de los experimentos más importantes,
por ejemplo, de aquellos sobre la composición de las células fecundadoras
híbridas.
A un único observador se le puede escapar fácilmente una diferencia, la cual,
aunque inicialmente parezca poco importante, puede crecer de tal forma que
no pueda ser despreciada con respecto al resultado completo. Si los híbridos
variables de otras especies de plantas presentan un comportamiento
totalmente similar, también hay que decidirlo primero mediante experimentos;
mientras, se puede creer que en los puntos importantes no debería aparecer
una diferencia básica, ya que la unidad en el plano de desarrollo de la vida
orgánica se encuentra fuera de toda duda.
Finalmente, también merecen una mención especial los experimentos
realizados por Kölreuter, y Gärtner y otros sobre la “transformación de una
especie en otra por fecundación artificial”. A estos experimentos se les atribuye
una importancia especial; Gärtner los cuenta entre los “más difíciles en la
producción de híbridos”.
Si había que transformar una especie A en otra B, ambas se unían por
fecundación, y los híbridos obtenidos se fecundaban de nuevo con el polen de
B; de entre los descendientes se escogía aquella forma que estaba más

[166]
próxima a la especie B y se fecundaba repetidamente con esta, hasta obtener,
al fin, una forma que era idéntica a B y cuyos descendientes permanecían
constantes. De esta manera, la especie A se había transformado en B. Gärtner
solo había realizado 30 experimentos semejantes con plantas del género
Aquilegia, Dianthus, Geum, Lavatera, Lychnis, Malva, Nicotiana y Oenothera.
El período de la transformación no era el mismo para todas las especies.
Mientras que para algunas valía una fecundación repetida tres veces, en otras
esta se tenía que repetir entre 5 y 6 veces; también para las mismas especies
nombradas se observaron fluctuaciones en distintos experimentos. Gärtner
atribuye estas discrepancias a la circunstancia de que “la fuerza típica con la
que una especie actúa en la procreación que lleva al cambio y transformación
del tipo materno es muy distinta en las diversas plantas, y, por consiguiente, el
número de generaciones por las cuales una especie se transforma en otra
también tienen que ser diferente, y la transformación de algunas especies se
sucede con más y otras con menos generaciones”. Más tarde, el mismo
observador comenta que “también los procesos de transformación dependen
de qué tipo y qué individuo se escogen para las posteriores transformaciones.” Si se
Los híbridos producen tantos tipos de células germen como combinaciones
constantes permiten los caracteres que en ellas se unen, y una de ellas
siempre es de la misma clase que la célula polen. Por tanto, en todos los
experimentos del mismo tipo existe la posibilidad de que, después de la
segunda fecundación, se consiga una forma constante, la cual es idéntica a la
célula polen. Pero que la misma se consiga depende en cada caso tanto del
número de las plantas experimentales como del número de caracteres
diferenciadores que se unieron por la fecundación. Supongamos, por ejemplo,
que las plantas escogidas para el experimento se diferencian en 3 caracteres y
que la especie ABC se transforma en la especie abc por fecundaciones
repetidas con el polen de esta. El híbrido resultante de la primera fecundación
forma 8 clases de células germen distintas:
ABC, ABc, AbC, aBC, Abc, aBc, abC, abc
Estas se unen en el segundo año experimental con la célula polen abc y se
obtiene la serie:
AaBbCc + AaBbc + AabCc + aBbCc + Aabc + aBbc + abCc + abc Com

[167]
dos fecundaciones. Si por casualidad no se hubiera obtenido, habría que
repetir la fecundación con una de las formas próximas: Aabc, aBbc, abCc.
Resulta esclarecedor que un experimento de este tipo se hace más largo
cuanto menor es el número de plantas experimentales y cuanto mayor es el
número de caracteres diferenciadores en las dos especies originarias; que,
además, en las especies nombradas puede suceder un retraso de una, e
incluso de dos generaciones, como ha observado Gärtner. La transformación
de especies muy distintas solo se puede terminar en el quinto o sexto año
experimental, ya que el número de células germen diferentes que se forman en
el híbrido aumenta según la potencia de dos sobre el número de caracteres
diferenciadores.
Gärtner encontró, por experimentos repetidos, que el periodo de transformación
recíproco es distinto en algunas especies, por lo que frecuentemente se puede
convertir una especie A en una B una generación antes que la especie B en la
A. Deduce al mismo tiempo que el punto de vista de Kölreuter, según el cual “la
naturaleza de ambos híbridos guarda el equilibrio más perfecto”, no es tan
infalible. Sin embargo, parece que Kölreuter no merece esta crítica, sobre todo
cuando Gärtner ha pasado por alto un asunto importante sobre el cual él mismo
llamó la atención en otro sitio, al considerar que “depende de qué individuo se
elige para la siguiente transformación”. Experimentos que se realizaron en este
sentido con dos especies de Pisum orientan hacia la elección de los individuos
más apropiados para la fecundación, porque podría existir una gran diferencia,
según cuál de las dos especies es transformada en la otra. Las dos plantas
experimentales se diferenciaban en 5 caracteres; al mismo tiempo, la especie A
poseía todos los dominantes y la otra B todos los caracteres recesivos. Para la
transformación recíproca, se fecundó A con el polen de B, y al revés, B con el
de A, y así se repite lo mismo en los dos híbridos el año siguiente. En el primer
experimento, B/A, después del tercer año experimental había, para la elección
del individuo que interviniese en la siguiente fecundación, 87 plantas de las 32
formas posibles; para el segundo experimento, A/B, se obtuvieron 73 plantas,
que en su aspecto coincidían completamente con la planta polen, pero que en
su composición interior tenían que ser tan distintas como las formas del otro
experimento. Una selección calculada, por lo tanto, solo era posible en el
primer experimento; en el segundo, por puro azar, tenían que ser eliminadas

[168]
algunas plantas. De las últimas, solo una parte de las flores se polinizó con el
polen de A, a las otras se las dejó a la autofecundación. De cada cinco plantas
que se usaron en los dos experimentos para la fecundación, como muestra el
cultivo del año siguiente, coincidían con la planta polen
Primer experimento Segundo experimento
2 plantas --- en todos los caracteres 3 ” --- “ 4 “ --- 2 plantas “ 3 “ --- 2 ” 2 “ --- 1 planta “ 1 carácter
Para el primer experimento la transformación estaba terminada; en el segundo,
que no se siguió realizando, se tendría que haber realizado seguramente dos
fecundaciones más.
Aunque no debiese aparecer con frecuencia el caso de que los caracteres
dominantes pertenezcan solamente a una o a otra planta originaria, siempre
habrá una diferencia de cuál de ambas plantas originarias posee un mayor
número. Si la mayoría de los caracteres dominantes pertenecen a la planta
polen, entonces la elección de la planta para la fecundación posterior tiene que
garantizar un cierto grado de seguridad, más que en el caso inverso, en el que
conlleva un retardo en el período de transformación, suponiendo que el
experimento solo se dé por terminado cuando se obtiene una planta, que no
solo en su apariencia es idéntica a la planta polen, sino que también, como en
esta, los descendientes permanecen constantes.
Por el éxito de los experimentos de transformación, Gärtner se vio movido a
volverse contra la opinión de aquellos investigadores que discutían la

[169]
estabilidad de las especies de plantas y suponen un desarrollo continuo de las
especies. Él ve, en la completa transformación de una especie en otra, la
prueba indiscutible de que la especie está sometida a límites por encima de los
cuales no es capaz de cambiar. Aunque a esta opinión no se le puede
reconocer una validez indiscutible, sí se encuentra, por otro lado, en los
experimentos realizados por Gärtner, una confirmación digna de observación
acerca de la sospecha anteriormente expresada sobre la variabilidad de las
plantas de cultivo.
Entre las especies experimentales había plantas cultivadas, como Aquilegia
atropurpurea y canadensis, Dianthus caryophyllus, chinensis y japonicus,
Nicotiana rustica y paniculada, y tampoco estas habían perdido nada de su
estabilidad tras 4-5 enlaces híbridos.

[170]
BIBLIOGRAFÍA
1869. "Ueber einige aus künstlicher Befruchtung gewonnenen Hieracium-
Bastarde."Verhandlungen des naturforschenden Vereines in Brünn 8
Abhandlungen 26-31. Translated as: "On Hieracium-Hybrids Obtained by
Artificial Fertilisation." in S&S, pp. 49-55.
1906. "The Progress of Genetic Research." Report of the Third International
Conference on Genetics; Hybridisation (The Cross-breeding of Genera or
Species), The Cross-breeding of Varieties, and General Plant-breeding.
London: Royal Horticultural Society, pp.90-97. [published 1907]
1924. Gregor Johann Mendel. Leben, Werk und Wirkung. Berlin: Julius
Springer.
1932. Life of Mendel. Translated by Eden & Cedar Paul. Reprinted, New York:
Hafner Publishing Co., 1966.
1943. "Johann Gregor Mendel wie er wirklich war." Verhandlungen des
naturforschenden Vereines in Brünn 74: 1-263. [This was Teil 2 of the volume
for the year 1942]

[171]
1984. En el centenario de Mendel: la Genética ayer y hoy. Coord. J.R.
Lacadena. Madrid. Ed. Alhambra.
1987. Beyond the Gene: Cytoplasmic Inheritance and the Struggle for Authority
in Genetics. New York: Oxford University Press.
1990. "The Nine Lives of Gregor Mendel." In Experimental Enquiries, ed. H.E.
Le Grand, pp. 137-166. Netherlands: Kluwer Academic Publishers.
2003. Los orígenes de la Genética en España. Editora Milagros Candela.
Madrid. Ed. AM3. Sociedad estatal de conmemoraciones culturales.
Ames, B. N. , Durston, W. E., Yamasaki, E. and Lee, F. D. (1973) Carcinogens
are mutagens: a simple test system combining liver homogenates for activation
and bacteria for detection. Proc. Natl. Acad. Sci. USA 70, 2281-2285.
Ames, B. N., Kammen, H. O. and Yamasaki, E. (1975) Hair dyes are
mutagenic: identification of a variety of mutagenic ingredients. Proc. Natl. Acad.
Sci. USA 72, 2423-2427.
Ames, B. N., Lee, F. D. and Durston, W. E. (1973) An improved bacterial test
system for the detection and classification of mutagens and carcinogens. Proc.
Natl. Acad. Sci. USA 70, 782-786.
Avery, O. T., MacLeod, C. M: and McCarty, M. (1944) Studies on the chemical
nature of the substance inducing transformation of pneumococcal types.
Induction of transformation by a desoxyribonucleic acid fraction isolated from
Pneumococcus type III. J. Exp. Med. 79, 137-158.

[172]
Baltimore, D. (1970) An RNA-dependent DNA polymerase in virions of RNA
tumor viruses. Nature 226, 1209-1211.
Bateson, William & R.C. Punnett. 1911. "On Gametic Series Involving
Reduplication of Certain Terms." Verhandlungen des naturforschenden
Vereines in Brünn 49: 324-344.
Bateson, William. 1902. "The Facts of Heredity in the Light of Mendel's
Discovery." Report of the Evolution Committee of the Royal Society 1:125-160.
Beadle, G. W. and Tatum, E. L. (1941) Genetic control of biochemical reactions
in Neurospora . Proc. Natl. Acad. Sci. USA 27, 499-506
Bertrand Rusell. 1988. El panorama de la ciencia. Ed. Ercilla S.A.
Bishop, B. E. (1996). "Mendel's Opposition to Evolution and to Darwin," Journal
of Heredity 87: 205-213.
Bolívar, F., Rodriguez, R. L., Green, P. J., Betlach M. C., Heyneker, H. L.,
Boyer, H. W., Grosa, J. H. and Folkow, S.(1977) Construction and
characterization of new cloning vehicles. II. A multipurpose cloning system.
Gene 2, 95-113
Brannigan, Augustin. 1981. The Social Basis of Scientific Discovery.
Cambridge: Cambridge University Press.

[173]
Brenner, S. (1957) On the impossibility of all overlapping triplet codes in
information transfer from nucleic acid to proteins. Proc. Natl. Acad. Sci. USA 43,
687-694.
Brenner, S., Jacob, F. and Meselson, M. (1961) An unstable intermediate
carrying information from genes to ribosomes for protein synthesis. Nature 190,
576-580. .
Bridges, C. B. (1916) Non disjunction as a proof of the chromosome theory of
heredity. Genetics 1, 1-52, 107-163.
Brown, D. M. and Todd, A. R. (1952) Nucleotides, part X. Some observations
on structure and chemical behaviour of the nucleic acids. J. Chem. Soc. 1, 52-
58.
Cech, T. R., Zaug, A. J. and Grabowski, P. I. (1981) In vitro splicing of the
ribosomal RNA precursor of Tetrahymena : involvement of a guanosine
nucleotide in the excision of the intervening sequence. Cell 27, 487-496.
Chargaff, E. (1950) Chemical specificity of the nucleic acids and mechanism of
their enzymatic degradation. Experientia 6, 201-209.
Chargaff, E. (1951) Structure and function of nucleic acids as cell constituents.
Fed. Proc. 10, 654-659.
Collins, F. S. and Galas, D. (1993) A new five year game plan for the U.S.
genome project. Science 262, 43-46.

[174]
Collins, F. S. et al. (1998) New goals for the U. S. Human Genome Project:
1998-2003. Science 282, 682-689.
Corcos, Alain & Floyd V. Monaghan. 1992. Gregor Mendel's Experiments on
Plant Hybrids. A Guided Study. New Brunswick, New Jersey: Rutgers University
Press.
Creighton, H. B. and McClintock, B. (1931) A correlation of cytological and
genetical crossing-over in Zea mays . Proc. Natl. Acad. Sci. USA 17, 492-497.
Crick, F. H. C. (1958) On protein synthesis. Biological replication of
macromolecules. Symp. Soc. Exp. Biol. 12, 138-163.
Crick, F. H. C. (1966) Codon-anticodon pairing: the wobble hypothesis. J. Mol.
Biol. 19, 548-555.
Crick, F. H. C., Barnett, L., Brenner, S. and Watts-Tobin, R. J. (1961) The
general nature of the genetic code for proteins. Nature 192, 1227-1232.
Darden, L. 1991. Theory Change in Science; Strategies from Mendelian
Genetics. New York: Oxford University Press.
Darwin, Charles R. 1868. The Variation of Animals and Plants under
Domestication. 2 vols. London, John Murray.
De Beer, Sir Gavin. 1965. "Genetics: The Centre of Science." Proceedings of
the Royal Society of London 164: 154-166
De Vries, Hugo. 1899. "Sur la fécondation hybride de l'albumen." Comptes
rendus hebdomadaire de l'Académie des sciences 129: 973.

[175]
Dobzhansky T (1965) Mendelism, Darwinism, and evolutionism. Proceedings of
the American Philosophical Society 109:205-215.
Driesch, Hans A.E.. 1908. The Science and Philosophy of the Organism. The
Gifford Lectures Delivered Before the University of Aberdeen in the Year 1907.
2 vols. London: A. and C. Black.
Dulbecco, R. (1986) A turning point in cancer research: sequencing the human
genome. Science 231, 1055-1056.
Dunn LC (1965) Mendel, his work and his place in history. Proceedings of the
American Philosophical Society 109:189-198.
Dunn P.M. (2003). Gregor Mendel, OSA (1822-1884), founder of scientific
genetic. Arch Dis Child Fetal Neonatal Ed. 88, F537-F539.
East E.M. 1923. Mendel and his contemporaries. The scientific monthly, 16:
225-237.
Eiseley, Loren. 1959. Darwin's Century. Evolution and the Men who Discovered
It. London: Victor Gollancz.
Ephrussi, B. and Weiss, M. C. (1965) Interspecific hybridization of somatic cells.
Proc Natl. Acad. Sci. USA. 53, 1040-1042.
Ephrussi, B. and Weiss, M. C. (1969) Hybrid somatic cells. Scientific Amer. 220,
26-35.
F.R.S” [psuedonym] (1968) Notes of a Not-Watson. Encounters July 31:60-66.

[176]
Fairbanks DJ, Rytting B (2001) Mendelian controversies: A botanical and
historical review. American Journal of Botany 88:737-752.
Fairbanks D.J., Rytting B. 2001. Mendelian controversies: a botanical and
historical review. Am. J.Botany, 88(5): 737-752.
Fisher RA (1936) Has Mendel’s work been rediscovered? Annals of Science
1:115-137.
Fisher, Ronald A. 1936. "Has Mendel's Work Been Rediscovered?" Annals of
Science 1: 115-137. Reprinted in: Stern & Sherwood. 1966. Referred to in
future as S&S], pp. 139-175.
Fraenkel-Conrat, H. (1956) The role of the nucleic acid in the reconstitution of
active tobacco mosaic virus. J. Am. Chem. Soc. 78, 882.
Fraenkel-Conrat, H. and Williams R. C. (1955) Reconstitution of active tobacco
mosaic virus from its inactive protein and nucleic acid components. Proc. Natl.
Acad. Sci. USA 41, 690-698.
Fraser, C. M. et al. (1998) Complete genome sequence of Treponema pallidum,
the syphilis spirochete. Science 281, 375-388.
Gamow, G. (1954) Possible relation between deoxyribonucleic acid and protein
structures. Nature 173, 318.
García-Bellido, A. (1975) Genetic control of wing disk development in
Drosophila . Ciba Foundation Symp. 29, 161-182.

[177]
García-Bellido, A., Ripoll, P. and Morata, G. (1973) Developmental
compartmentalization of the wing disc of Drosophila . Nature 245, 251-253.
Gliboff S (1999) Gregor Mendel and the laws of evolution. History of Science
37:217-235.
González Astorga, J. 2001. Breve cronología de la Genética. Ciencias. 63: 70-
77.
Hartl, D. L., & Orel, V. (1992). "What did Mendel think he discovered?" Genetics
131: 245-253.
Haruna, I. and Spiegelman, S. (1965) Proc. Natl. Acad. Sci. USA 54, 579-587.
Haruna, I., Nozu, K., Ohtaka, Y. and Spiegelman, S. (1963) Proc. Natl. Acad.
Sci. USA 50, 905-911.
Heimans, J. 1971. "Mendel's Ideas on tne Nature of Hereditary Characters. The
Explanation of Fragmentary Records of Mendel's Hybridizing Experiments,"
Folia Mendeliana 6: 91-98.
Hirschhorn R. 1995. Genetic Mosaicism: what Gregor Mendel didn´t know. 95:
443-444.
Iltis, H. (1924). Life of Mendel. London: George Allen & Unwin Ltd.
Iltis, Hugo. 1911. "Vom Mendel denkmal und von seiner Enthüllung."
Verhandlungen des naturforschenden Vereines in Brünn 49: 335-363

[178]
Jacob, F. and Monod, J. (1961) Genetic regulatory mechanisms in the synthesis
of proteins. J. Mol. Biol. 3, 318-356.
Jacob, F. and Wollman, E. L. (1953) Induction of phage development in
lysogenic bacteria. Cold Spring Harbor Symp. Quant. Biol. 18, 101.
Johannsen, Wilhelm. 1911. "The Genotype Conception of Heredity." American
Naturalist 45: 129-159.
Kart R. Popper. Lógica de la investigación científica. 1991. 1ª edición. Ed. Rel.
México.
Khorana, H. G. (1965) Polynucleotide synthesis and the genetic code.
Federation Proc. 24, 1473-1487.
Kimmelman, Barbara A. 1992. "Organisms and Interests in Scientific Research:
R.A. Emerson's Claims for the Unique Contributions of Agricultural Genetics."
In: Clarke & Fujimura (Eds.). 1992, pp. 198-232.
Kornberg, A. (1960) Biological synthesis of deoxyribonucleic acid. Science 131,
1503-1508.
Kornberg, A. and Baker, T. (1992) DNA replication. W. H. Freeman & Co., New
York.
Kornberg, A., Lehman, I. R., Bessman, M. J. and Simms, E. S. (1956) Enzymic
synthesis of deoxyribonucleic acid. Biochim. Biophys. Acta 21, 197-198.
Kornberg, R. D. (1977) Structure of chromatin. Annu. Rev. Biochem. 46, 931-
954.

[179]
Kornberg, R. D. and Lorch, Y. (1999) Twenty-five years of the nucleosome,
fundamental particle of the eukaryote chromosome. Cell 98, 285-294.
Marañón J.A., Caballero-Elipe E, Blasco Sancho S., Álvarez-Martínez A., Galán
F. Mendel: ese gran desconocido. beSEG. 2005; 20: 1-3.
Marmur, J and Doty, P. (1959) Nature 183, 1427.
Maxam, A. M. and Gilbert, W. (1977) A new method for sequencing DNA. Proc.
Natl. Acad. Sci. USA 74, 560-564.
Mayr, Ernst. 1982. The Growth of Biological Thought. Diversity, Evolution, and
Inheritance. Cambridge, Mass., & London: Harvard University Press.
McClintock, B. (1934) The relation of a particular chromosomal element to the
development of the nucleoli in Zea mays . Z. Zellforsch. Mikrosk. Anat. 21, 294-
328.
McClintock, B. (1948) Mutable loci in maize. Carnegie Inst. Wash. Year Book
47, 155-169.
McClintock, B. (1950) The origin and behavior of mutable loci in maize. Proc.
Natl. Acad. Sci. USA 36, 344-355.
McClintock, B. (1951) Chromosome organization and gene expression. Cold
Spring Harbor Symp. Quant. Biol. 16, 13-57.
McClintock, B. (1956) Controlling elements and the gene. Cold Spring Harbor
Symp. Quant. Biol., 21, 197-216.

[180]
McKusicK VA. Human genetics: the last 35 years, the present and the future.
Am J. Med. Genet. 1992; 50: 663-670-
McKusick, V. A. (1989) Mapping and sequencing the human genome. New
England J. Med. 320, 910-915.
McKusick, V. A. (1989) The Human Genome Organisation: history, purposes
and membership. Genomics 5, 385-387.
McKusick, V. A. and Ruddle, F. H. (1977) The status of the gene map of the
human chromosomes. Science 196, 390-405.
Meijer, O.G. 1985. "Hugo de Vries no Mendelian?" Annals of Science 42: 189-
232.
Mendel JG (1865) Versuche über pflanzen-hybriden. Verhandlungen des
Naturforschenden Vereines in Brünn, Abhandlungen 4:3-47.
Mendel JG (1870) Über einge aus künstlicher befruchtung gewonnene
Hieracium bastarde. Verhandlungen des Naturforschenden Vereines in Brünn,
Abhandlungen 8:26-31.
Mendel JG (1966) Experiments on plant hybrids (translation). In The Origin of
Genetics: A Mendel Source Book, C. Stern and E. Sherwood (eds), WH
Freeman: San Francisco. pp. 1-48.
Mendel, G. (1866). "Experiments in Plant Hybridization." On-line document:
http://www.netspace.org/MendelWeb/Mendel.plain.html

[181]
Mendel, G. (1866). "Experiments on Plant Hybrids." In: The Origin of Genetics:
A Mendel Source Book.
Mendel, G. (1866b). "Letters to Carl Nageli: 1866-1873." Trans. L. K. Piternick &
G. Piternick. In: The Origin of Genetics: A Mendel Source Book.
Mendel, G. (1869). "On Hieracium-Hybrids." In: The Origin of Genetics: A
Mendel Source Book.
Mendel, Johann Gregor. 1866. "Versuche über Pflanzenhybriden."
Verhandlungen des naturforschenden Vereines in Brünn 4 Abhandlungen, 3-47.
Reprinted with corrections from a copy of the manuscript in Fundamenta
Genetica, ed. Jaroslav Krízenecký, pp.57-92. Osterhout, The Netherlands &
Prague: Anthropological Publications & the Czechoslovak Academy of
Sciences. [Referred to as Mendel, 1866K]
Mendel G. 1950. Gregor Mednel´s letter to Carl Nageli. Genetics, 35: 1-29.
Meselson, M. and Stahl, F. W. (1958) The replication of DNA in Escherichia coli
. Proc. Natl. Acad. Sci. USA 44, 671-682.
Meselson, M. and Weigle, J. J. (1961) Chromosome breakage accompanying
genetic recombination in bacteriophage. Proc. Natl. Acad. Sci. USA 47, 857-
868.
Moore, R. (1954). Man, Time and Fossils: The Story of Evolution. London:
Jonathan Cape. (cited in Voipio 1990)

[182]
Morgan, T. H. (1910) Sex limited inheritance in Drosophila . Science 32, 120-
122.
Mullis, K. B., Faloona, F. A., Scharf, S., Saiki, R., Horn, G. and Erlich, H. (1986)
Specific enzymatic amplification of DNA in vitro : The polymerase chain
reaction. Cold Spring Harbor Symp. Quant. Biol. 51, 263-273.
Nijhout F. 2004. Importancia del contexto en la Genética. Investigación y
ciencia. Agosto. 62-69.
Nirenberg, M. W. and Matthaei, J. H. (1961) The dependence of cell-free
protein synthesis in E. coli upon naturally occurring or synthetic
polyribonucleotides. Proc. Natl. Acad. Sci. USA 47, 1588-1602.
Okazaki, R. T., Okazaki, K., Sakabe, K., Sugimoto, K. and Sugino, A. (1968)
Mechanism of DNA chain growth. I. Possible discontinuity and unusual
secondary structure of newly synthesized chains. Proc. Natl. Acad. Sci. USA
59, 598-605.
Olby R (1985) Origins of Mendelism, 2nd edition University of Chicago Press:
Chicago.
Olby, R. C. (1966). Origins of Mendelism. Chicago: University of Chicago Press.
Olby, R. C. (1997). "Mendel, Mendelism and Genetics." On-line document:
http://www.netspace.org/MendelWeb/MWolby.html

[183]
Olby, R.C. 1979. "Mendel no Mendelian?" History of Science 17: 53-72.
Reprinted with minor changes in The Origins of Mendelism, 2nd ed., pp.234-
258. Chicago: Chicago University Press, 1985.
Orel, V. (1996). Gregor Mendel: The First Geneticist. New York: Oxford
University Press.
Paul, Diane B., & Barbara A. Kimmelman. 1988. "Mendel in America: Theory
and Practice, 1900-1919." In The American Development of Biology, ed. Ronald
Rainger, Keith R. Benson, & Jane Maienschein, pp.281-310. Philadelphia:
University of Pennsylvania Press. [Paperback, 1991. New Brunswick & London:
Rutgers University Press], pp. 281-310.
Pauling, L. and Delbrück, M. (1940) The nature of the intermolecular forces
operative in biological processes. Science 92, 77-79.
Pontecorvo, Guido. 1966. "Template and Stepwise Processes in Heredity."
Proceedings of the Royal Society of London 164: 167-169.
Porteous J.W. 2004. We still fail to account for Mendel´s observations.
Theoretical Biology and Medical Modelling, 1-4.
http://www.ibiomed.com/content/1/1/4.
Rheinberger, Hans-Jörg. 1995. "When did Carl Correns Read Gregor Mendel's
Paper?" Isis, 86: 612-618.
Richter, Oswald. 1941. "75 Jahre seit Mendels Grosstat und Mendels
Stellungnahme zu Darwins Werken auf Grund seiner Entdeckungen."
Verhandlungen des naturforschenden Vereines in Brünn 72: 110-173.

[184]
Ruzicka, Vladimir. 1925. "Speech as Representative of the University of
Bratislava." Memorial Volume in Honor of the 100th. Birthday of J.G. Mendel.
Prague: Fr. Borovy, pp. 31-49.
Sanger, F. and Thompson, E. O. P. (1953) The amino acid sequence in the
glycyl chain of insulin. Biochem. J. 53, 353-374.
Sanger, F., Air, G. M., Barrell, B. G., Brown, N. L., Coulson, A. R., Fiddes, J. C.,
Hutchison, C. A. III, Slocombe, P. M. and Smith, M. (1977) Nucleotide
sequence of bacteriophage fi X174 DNA. Nature 265, 687-695.
Sanger, F., Nicklen, S. and Coulson, A. R. (1977) DNA sequencing with chain-
terminating inhibitors. Proc. Natl. Acad. Sci. USA 74, 5463-5467.
Sapp, Jan. 1983. "The Struggle for Authority in the Field of Heredity, 1900-
1932: New Perspectives on the Rise of Genetics." Journal of the History of
Biology 16: 311-342.
Schrödinger, E. (1944) What is life? The physical aspect of the living cell.
Cambridge Univ. Press.
Sharp, P. A., Sugden, B. and Sambrook, J. (1973) Detection of two restriction
endonuclease activities in Hemophilus parainfluenzae using analytical agarose-
ethidium bromide electrophoresis. Biochemistry 12, 3055-3062.
Stamhuis, I., 1996. "The Rediscovery of Mendel's Laws was not important to
Hugo de Vries (1848-1935): Evidence from his letters to Jan Willem Moll (1851-
1933)." Folia Mendeliana.

[185]
Standfuss, Max. 1907. "Hybridations-Experimente, im weitesten Sinne des
Wortes, vom Jahre 1873 zur Gegenwart in Ihren Ausblicken auf die Scheidung
der Arten und den Weg, welchen diese Scheidung durchläuft." Seventh Inter.
Congr. Zoology, pp. 111-127.
Stent, G. S. and Calendar, R. (1978) Molecular Genetics. An introductory
narrative. 2 nd ed., pp.465. W. H. Freeman and Co., San Francisco.
Stern, C. (1931) Zytologisch-genetische Untersuchungen als Beweise für die
Morgansche Theorie des Faktoresaustauch. Biol. Zentralbl. 51, 547-587.
Stern, C., & Sherwood, E. R. (1966). The Origin of Genetics: A Mendel Source
Book. San Francisco: W. H. Freeman and Company.
Tatum, E. L. and Lederberg, J. (1947) Gene recombination in the bacterium
Escherichia coli . J. Bacteriol. 53, 673.
Taylor, J. H., Woods, P. S., and Hughes, W. L. (1957) The organization and
duplication of chromosomes as revealed by autoradiographic studies using
tritium-labeled thymidine. Proc. Natl. Acad. Sci. USA 43, 122-128.
Taylor, K., Hradecna, Z. and Szybalski, W. (1967) Asymmetric distribution of the
transcribing regions on the complementary strands of coliphage lambda DNA.
Proc. Natl. Acad. Sci. USA 57, 1618.
Temin, H. M. (1972) RNA-directed DNA synthesis. Scientific Amer. 226, 24-33.

[186]
Tschermak, Erich. 1906. "Ueber Bildung neuer Formen durch Kreuzung." In
Resultats scientifiques du congrés international botanique, Vienne, 1905, ed.
J.P.Lotsy, pp.323-332. Jena.
Venter, J. C., Smith, H. O. and Hood, L. (1996) A new strategy for genome
sequencing. Nature 381, 364-365.
Watson J. D. and Crick, F. H. C. (1953) Genetical implications of the structure
of deoxyribonucleic acid. Nature 171, 964-967.
Watson J. D. and Crick, F. H. C. (1953) Molecular structure of nucleic acids. A
structure for deoxyribose nucleic acid. Nature 171, 737-738.
Watson, J. D. (1990) The Human Genome Project: Past, present and future.
Science 248, 44-49.
Weiling F. (1991). Historical study: Johann Gregor Mendel. Am. J. Med. Genet.
40: 1-25
Weiss K. 2002. Going on in Mendel´s garden. Evolutionary Anthroppology. 11:
40-44.
Weissmann, C., Simon, L. and Ochoa, S. (1963) Proc. Natl. Acad. Sci. USA 49,
407-414.
Weldon WRF (1902) Mendel’s law of alternative inheritance in peas. Biometrika
1:228-254.
Wolins L (1962) Responsibility for raw data. American Psychologist 17:657-666.

[187]
Woodger.JH.(1978). Bilogía y Lenguaje. Ed. Tecnos .ISBN 84-309-0738-6
Yanofsky, C. (1967) Gene structure and protein structure. Scientific Amer, May.
Zirkle C (1968) Mendel and his era. In Mendel Centenary: Genetics,
Development and Evolution, RM Nardone (ed), Catholic University of America
Press: Washington DC. pp. 122-132.

[188]
Agradecimientos:
In memorian Fernando Galán Gutiérrez
Nuestro primer agradecimiento ha de ser para el eminente Prof. F .J.
Ayala, por proporcionarnos el honor de habernos aconsejado en la realización
de este libro y haber sido su primer lector.
A mis colaboradores directos en la elaboración de este libro:
José Ángel Marañón Maroto. (4)
Ernesto Caballero. (5)
Sonia Blasco Sancho. (2)
Juan González Julián. (3)
1) Universidad Autónoma Madrid / Facultad de Ciencias. Depto. Química Analítica.C-XIII.28049./ Madrid / Spain. 2) University of Leipzig Institute of Analytical Chemistry. Linnéstrasse. 3. 04103 / Leipzig. Germany 3) Facultad de Medicina. Microscopia Electronica , Universidad de Salamanca 4) Tradichem. S.L Actives & Ingredients Santiago Grisolía 2- 102. Parque Científico de Madrid.
28760. Tres Cantos. Madrid. SPAIN
5) Universidad Miguel Hernández de Elche. Alicante Spain.
Este libro no podría haberse concluido sin la ayuda de diversas personas. Pero
la más importante, gracias a su propia gran obra, sin duda alguna ha sido la
del Prof. Galán, a la que dedicamos, in memoriam, este libro. La obra se
beneficia del trabajo previo del Prof. Galán, director de un equipo de
colaboradores desde 1962 a 1991 al cual pertenecí. Admirador de Mendel y,
quizás como él, perteneciente también a esos científicos olvidados y, aún hoy,
mal comprendidos hasta por sus propios colaboradores, el Prof. Galán fue
igualmente un proscrito de los científicos de ayer y de hoy.
He de agradecer su cooperación a los más relevantes colaboradores del Prof.
Galán que, de una manera o de otra, lo acompañaron en dichos trabajos, de
los cuales se ha surtido la cristalización de este libro: En primer lugar, al
colaborador inseparable del Prof. Galán, el Dr. Juan González Julián, que me

[189]
ha apoyado de forma directa en la elaboración de este libro con sus acertadas
críticas lingüísticas.Mas lo que decidió de forma categórica la realización de
este libro fue la creación que realicé en la Universidad Autónoma de Madrid de
una asignatura, cuyo título fue “Mendel; paradigma del método científico”, por la
que han pasado cientos de alumnos de psicología, informática, matemáticos,
químicos, físicos y biólogos. Asignatura que sigo impartiendo con la
colaboración de uno de mis más entrañables colaboradores, el Dr. Marañón.
Y no puedo seguir escribiendo si no expreso mi más profundo agradecimiento a
la Dra. Sonia Blasco por su inestimable ayuda en la traducción del alemán al
español del “Versuche Über Pflanzenhybriden”. Y cómo no, mi mayor
agradecimiento a quien, además de alumno de la asignatura “Mendel;
paradigma del método científico”, es uno de mis mejores colaboradores, el Dr.
Ernesto Caballero, sin cuyo profundo romanticismo por la ciencia, su tesón y
dedicación, ya en mis investigaciones, ya en su empeño en hacer posible la
creación y publicación de este libro, hubiera sido imposible el nacimiento de
“Mendel según Mendel”.
Para terminar, quiero expresar mi gran reconocimiento y agradecimiento a la
gran ayuda que nos ha brindado para la corrección lingüística y gramatical la
realizada por el Prof. A .Díez Jambrina.

[190]
.




















