
Contribution of endogenously expressed Trp1 to a Ca2+-selective, store-operated Ca2+ entry pathway
12
Contribution of endogenously expressed Trp1 to a Ca 21 -selective, store-operated Ca 21 entry pathway GEORGE H. BROUGH, SONGWEI WU, DONNA CIOFFI, TIMOTHY M. MOORE, MING LI, NICHOLAS DEAN,* AND TROY STEVENS 1 Department of Pharmacology, University of South Alabama, College of Medicine, Mobile, Alabama 36688, USA; and *ISIS Pharmaceuticals, Carlsbad, California 92008, USA ABSTRACT Heterologous expression of the tran- sient receptor potential-1 gene product (Trp1) encodes for a Ca 21 entry pathway, though it is unclear whether endogenous Trp1 contributes to a selective store-oper- ated Ca 21 entry current. We examined the role of Trp1 in regulating both store-operated Ca 21 entry and a store-operated Ca 21 entry current, I SOC , in A549 and endothelial cells. Twenty different ‘chimeric’ 2*-O-(2- methoxy)ethylphosphothioate antisense oligonucleo- tides were transfected separately using cationic lipids and screened for their ability to inhibit Trp1 mRNA. Two hypersensitive regions were identified, one at the 5* end of the coding region and the second in the 3* untranslated region beginning six nucleotides down- stream of the stop codon. Antisense oligonucleotides stably decreased Trp1 at concentrations ranging from 10 to 300 nM, for up to 72 h. Thapsigargin increased global cytosolic Ca 21 and activated a I SOC , which was small (235 pA @ 280 mV), reversed near 140 mV, inhibited by 50 mM La 31 , and exhibited anomalous mole fraction dependence. Inhibition of Trp1 reduced the global cytosolic Ca 21 response to thapsigargin by 25% and similarly reduced I SOC by 50%. These data collectively support a role for endogenously expressed Trp1 in regulating a Ca 21 -selective current activated upon Ca 21 store depletion.—Brough, G. H., Wu, S., Cioffi, D., Moore, T. M., Li, M., Dean, N., Stevens, T. Contribution of endogenously expressed Trp1 to a Ca 21 -selective, store-operated Ca 21 entry pathway. FASEB J. 15, 1727–1738 (2001) Key Words: capacitative Ca 21 entry z I CRAC z signal transduc- tion z endothelial cells Activation of Ca 21 entry across the cell membrane is an important inflammatory stimulus in nonexcitable cells. This stimulus produces degranulation in mast cells (1–3) and activates cell-mediated immunity in T lymphocytes (4 – 8). In endothelial cells, Ca 21 entry across the cell membrane initiates a retraction needed to deliver plasma-rich exudate to inflamed tissue (9 – 15) and for transmigration of circulating leukocytes (16 –19). These cell types all share a common mode of Ca 21 entry: this Ca 21 entry process is activated after the depletion of Ca 21 stored in the endoplasmic reticulum, a so-called store-operated Ca 21 entry pathway (20 –25). However, several apparent store-operated Ca 21 entry pathways exist, each possessing distinctive electrophys- iological characteristics and each making unique con- tributions to the global rise in cytosolic Ca 21 ([Ca 21 ] i ) after Ca 21 store depletion. Indeed, mast cells and lymphocytes express a highly Ca 21 -selective current, called a Ca 21 release-activated current or I CRAC (1, 4). The endothelial cell current may or may not exhibit less Ca 21 selectively (26 –28) and is generally referred to as a store-operated Ca 21 current or I SOC . The molecular identity of ion channels that mediate store-operated Ca 21 entry is still poorly understood, though electrophysiologic studies suggest the presence of multiple, different channels. Discovery that the transient receptor potential (trp) gene product in Dro- sophila melanogaster encoded a retinal store-operated Ca 21 entry channel provided impetus for cloning a whole family of related mammalian homologues (22, 29). These products belong to a six-transmembrane spanning domain family of cation channels that in- clude the Drosophila Trp and seven mammalian Trp homologues (Trp1-Trp7). Predicted topography sug- gests the functional channel is a tetramer, wherein the pore is formed by cohesion of membrane-delimited domains bridging the S5 and S6 membrane spanning regions from four different monomers (29). Thus, multiple combinations of Trp proteins could result in unique channel characteristics. Trp1, 4, and 5 appear to be activated upon Ca 21 store depletion, though not all mammalian Trp channels are store operated (22). Most available data are derived from the heterologous expression of individual gene products, where ex- pressed proteins may form channels with properties that do not resemble their endogenous phenotype. Indeed, coexpression of Trp1 with Trp3 produced a channel with biophysical and regulatory properties that were different from the expression of either Trp1 or Trp3 alone (30). There is therefore a need to examine the contribution that individual Trp proteins make to endogenous modes of Ca 21 entry. Although Trp1 contributes to formation of a Ca 21 1 Correspondence: Department of Pharmacology, Univer- sity of South Alabama College of Medicine, 307 N. University Blvd., MSB 3360, Mobile, AL 36688, USA. E-mail: tstevens @jaguar1.usouthal.edu 1727 0892-6638/01/0015-1727 © FASEB
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Contribution of endogenously expressed Trp1 to a Ca2+-selective, store-operated Ca2+ entry pathway

Contribution of endogenously expressed Trp1 to aCa21-selective, store-operated Ca21 entry pathway
GEORGE H. BROUGH, SONGWEI WU, DONNA CIOFFI, TIMOTHY M. MOORE,MING LI, NICHOLAS DEAN,* AND TROY STEVENS1
Department of Pharmacology, University of South Alabama, College of Medicine,Mobile, Alabama 36688, USA; and *ISIS Pharmaceuticals, Carlsbad, California 92008, USA
ABSTRACT Heterologous expression of the tran-sient receptor potential-1 gene product (Trp1) encodesfor a Ca21 entry pathway, though it is unclear whetherendogenous Trp1 contributes to a selective store-oper-ated Ca21 entry current. We examined the role of Trp1in regulating both store-operated Ca21 entry and astore-operated Ca21 entry current, ISOC, in A549 andendothelial cells. Twenty different ‘chimeric’ 2*-O-(2-methoxy)ethylphosphothioate antisense oligonucleo-tides were transfected separately using cationic lipidsand screened for their ability to inhibit Trp1 mRNA.Two hypersensitive regions were identified, one at the5* end of the coding region and the second in the 3*untranslated region beginning six nucleotides down-stream of the stop codon. Antisense oligonucleotidesstably decreased Trp1 at concentrations ranging from10 to 300 nM, for up to 72 h. Thapsigargin increasedglobal cytosolic Ca21 and activated a ISOC, which wassmall (235 pA @ 280 mV), reversed near 140 mV,inhibited by 50 mM La31, and exhibited anomalousmole fraction dependence. Inhibition of Trp1 reducedthe global cytosolic Ca21 response to thapsigargin by25% and similarly reduced ISOC by 50%. These datacollectively support a role for endogenously expressedTrp1 in regulating a Ca21-selective current activatedupon Ca21 store depletion.—Brough, G. H., Wu, S.,Cioffi, D., Moore, T. M., Li, M., Dean, N., Stevens, T.Contribution of endogenously expressed Trp1 to aCa21-selective, store-operated Ca21 entry pathway.FASEB J. 15, 1727–1738 (2001)
Key Words: capacitative Ca21 entry z ICRAC z signal transduc-tion z endothelial cells
Activation of Ca21 entry across the cell membrane isan important inflammatory stimulus in nonexcitablecells. This stimulus produces degranulation in mastcells (1–3) and activates cell-mediated immunity in Tlymphocytes (4–8). In endothelial cells, Ca21 entryacross the cell membrane initiates a retraction neededto deliver plasma-rich exudate to inflamed tissue (9–15) and for transmigration of circulating leukocytes(16–19). These cell types all share a common mode ofCa21 entry: this Ca21 entry process is activated after thedepletion of Ca21 stored in the endoplasmic reticulum,a so-called store-operated Ca21 entry pathway (20–25).
However, several apparent store-operated Ca21 entrypathways exist, each possessing distinctive electrophys-iological characteristics and each making unique con-tributions to the global rise in cytosolic Ca21 ([Ca21]i)after Ca21 store depletion. Indeed, mast cells andlymphocytes express a highly Ca21-selective current,called a Ca21 release-activated current or ICRAC (1, 4).The endothelial cell current may or may not exhibitless Ca21 selectively (26–28) and is generally referredto as a store-operated Ca21 current or ISOC.
The molecular identity of ion channels that mediatestore-operated Ca21 entry is still poorly understood,though electrophysiologic studies suggest the presenceof multiple, different channels. Discovery that thetransient receptor potential (trp) gene product in Dro-sophila melanogaster encoded a retinal store-operatedCa21 entry channel provided impetus for cloning awhole family of related mammalian homologues (22,29). These products belong to a six-transmembranespanning domain family of cation channels that in-clude the Drosophila Trp and seven mammalian Trphomologues (Trp1-Trp7). Predicted topography sug-gests the functional channel is a tetramer, wherein thepore is formed by cohesion of membrane-delimiteddomains bridging the S5 and S6 membrane spanningregions from four different monomers (29). Thus,multiple combinations of Trp proteins could result inunique channel characteristics. Trp1, 4, and 5 appearto be activated upon Ca21 store depletion, though notall mammalian Trp channels are store operated (22).Most available data are derived from the heterologousexpression of individual gene products, where ex-pressed proteins may form channels with propertiesthat do not resemble their endogenous phenotype.Indeed, coexpression of Trp1 with Trp3 produced achannel with biophysical and regulatory properties thatwere different from the expression of either Trp1 orTrp3 alone (30). There is therefore a need to examinethe contribution that individual Trp proteins make toendogenous modes of Ca21 entry.
Although Trp1 contributes to formation of a Ca21
1 Correspondence: Department of Pharmacology, Univer-sity of South Alabama College of Medicine, 307 N. UniversityBlvd., MSB 3360, Mobile, AL 36688, USA. E-mail: [email protected]
17270892-6638/01/0015-1727 © FASEB

entry pathway, the nature of this Trp1-containing chan-nel is not clear. Overexpression of Trp1 likely forms anonselective cation channel that enhances store-oper-ated Ca21 entry, particularly in recalcification experi-ments (29, 31, 32). However, there have been limitedattempts to examine the contribution of Trp1 to itsendogenously formed channel. Recent findings indi-cate that inhibition of Trp1 expression in salivary glandcells (33) and of Trp1 and Trp3 in HEK-293 cells (34)reduces the global [Ca21]i response to activation ofstore-operated Ca21 entry, suggesting that Trp1 maycomprise a functional subunit of certain store-operatedCa21 entry pathways. Neither of these studies specifi-cally examined the electrophysiological nature of Trp1-containing channels. Indeed, the role of Trp1 in ISOC iscontroversial; expression of Trp1 augmented thewhole-cell Ca21 current in CHO cells (31), but did notfunction as a store-operated Ca21 entry channel whenexpressed in the baculovirus/Sf9 cell system (35). Ourpresent studies therefore used an antisense oligonucle-otide approach to inhibit Trp1 mRNA to examine thespecific role of Trp1 in an ISOC identified in twolung-derived nonexcitable cell types (A549 and endo-thelial cells) responsive to inflammatory stimuli.
MATERIALS AND METHODS
Cell culture
A549 (lung epithelial carcinoma cell line) cells were kindlyprovided by Dr. Richard Honkanen (University of SouthAlabama). The cells were maintained in Dulbecco’s modifiedEagle’s medium (DMEM) (Gibco BRL, Rockville, MD) con-taining 1 g glucose per liter and 10% fetal bovine serum (FBS;Hyclone Laboratories, Logan, UT) with 100 units/ml penicil-lin and 0.1 mg/ml streptomycin (Sigma, St. Louis, MO). Cellswere routinely passaged at 90–95% confluence.
Human pulmonary artery endothelial cells were purchasedfrom Clonetics, Inc. (San Diego, CA). Cells were maintainedin 1:1 DMEM/Ham’s F-12 with 10% FBS. Cells were verifiedas endothelial by positive factor VIII staining and by theuptake of 1,19-dioctadecyl-3,3,39,39-tetramethylindocarbocya-nine perchlorate-labeled, acetylated low density lipoprotein.Cells were cultured after standard procedures, as describedelsewhere (14, 15, 36).
Molecular biology
Standard techniques for RT-PCR subcloning were generallyfollowed. Total RNA was extracted with RNA Stat-60 (Tel-Test‘B’, Friendswood, TX) from A549 cells and human pulmo-nary artery endothelial cells were grown to 100% confluence(;107 cells) on 75 cm2 tissue culture flasks. First-strandsynthesis was performed with SuperScript II reverse transcrip-tase (200 units) and oligo(dT)12–18 primer (Life Technolo-gies, Rockville, MD) on ; 1 mg of DNase I-treated total RNA.PCR was conducted with the following sets of primers forTrp1/Trpb: set A, 59-TCG CCG AAC GAG GTG ATG G-39(sense) and 59-GTT ATG GTA ACA GCA TTT CTC C-39(antisense); set B, 59-GTG CTT GGG AGA AAT GCT G-39(sense) and 59-GGG GCT TGG GTA GAG ATA C-39 (anti-sense); PCR products were ligated into TA cloning vectorpCR2.1 (Invitrogen, San Diego, CA) and transformed into
chemically competent Escherichia coli. Plasmids were isolatedby the QIAprep® spin prep system (Qiagen, Valencia, CA)and submitted to The Biopolymer Laboratory at The Univer-sity of South Alabama for automated fluorescence sequenceanalysis (Applied Biosystems 373XL DNA sequencer). Se-quence accuracy was confirmed by sequencing both strandswith universal primers. Nucleotide and amino acid align-ments were achieved with BLAST (NCBI) and DNASIS v2.0(Hitachi Software) programs.
Oligonucleotide synthesis
Synthesis and purification of chimeric deoxyphosphorothio-ate/29-O-methoxyethyl base oligonucleotides were performedusing an Applied Biosystems 380B automated DNA synthe-sizer, as described by ISIS Pharmaceuticals (37).
Cell treatment with antisense oligonucleotides
A549 cells were seeded at 1.2–1.5 3 105 cells per 60 mm dishuntil 80% confluence was reached (;3 days). At that time, 1ml of DMEM containing ISIS antisense oligonucleotide (10–300 nM final concentration) and 15 mg/ml Lipofectin®(DOTMA/DOPE) (Gibco BRL, Rockville, MD) complexeswere added to the cells for 4 h in humidified air at 37°C with5% CO2. The complexes were washed off and cells wereallowed to recover in full-growth media (DMEM containing10% FBS with penicillin/streptomycin antibiotics).
Microinjection of antisense oligonucleotides
In specified experiments, antisense oligonucleotides weremicroinjected into endothelial cells as we have described (38,39). For these studies, human endothelial cells were seededonto 25 mm circle glass coverslips etched with identifyingmarks. Cells were grown for 24 to 48 h in DMEM supple-mented with 10% FBS and penicillin/streptomycin in humid-ified air at 37°C with 5% CO2. Microinjection was performedusing an Olympus IX70 inverted microscope with Transjector5246, Micromanipulator 5179, and Sterile Femtotips II (timesetting 0.2 s, injection pressure 7.0 psi, compensation pres-sure 0.6 psi). After injections, cells were incubated for periodsof 24, 48, or 72 h. Injected cells were subsequently identifiedby their location and [Ca21]i measurements were performed.
Measurement of Trp1 mRNA levels
A standard Northern blotting procedure with modificationsto optimize detection of low abundance messages was used.After the specified treatment period (24 to 72 h) withantisense oligonucleotides/Lipofectin® complexes, totalRNA was extracted from A549 cells lysed with RNA Stat-60™reagent (Tel-Test ‘B’ Inc., Friendswood, TX). Twenty micro-grams of each RNA sample was resolved on a 1% agarose-formaldehyde gel. RNA transfer to nylon membranes (Bright-Star-Plus™, Ambion, Austin, TX) was accomplished byupward capillary transfer technique for 1 h and cross-linkedwith 125 mJ UV light (GeneLinker, Bio-Rad, Hercules, CA).An ssDNA probe (Tst-47, 226 bases) to trp1 mRNA wasradiolabeled with [a-32P]dCTP along with a single-strandedprobe for the RNA molecular size marker (MillenniumMarker™, Ambion) by linear PCR with the respective anti-sense primers. The blots were hybridized with 1–3 3 106
cpm/ml of Trp1 probe and 20,000 cpm/ml of size markerprobe in UltraHyb™ (Ambion) solution in roller tubes(Techne, Cambridge, UK). Hybridization was performed at
1728 Vol. 15 August 2001 BROUGH ET AL.The FASEB Journal

43°C and overnight (16–20 h). The next day, blots werewashed twice with 23 SSC/0.1% SDS for 10 min and twicewith 0.23 SSC/0.1% SDS for 15 min. The blots were placedon Kodak X-OMAT film with an intensifying screen for up to5 days before development. Blots were reprobed with 32P-labeled glycerol-3-phosphate dehydrogenase to confirm equalloading of RNA samples.
Intracellular free calcium measurements
A549 cells were seeded onto 25 mm circle microscope glasscoverslips (Fisher Scientific, Pittsburgh, PA) and grown toconfluence. Cytosolic Ca21 ([Ca21]i) was estimated with theCa21-sensitive fluorophore Fura-2/acetoxymethylester (Mo-lecular Probes, Eugene, OR). Coverslip-attached A549 cellswere transferred into HEPES-buffered physiological salt solu-tion (in mM: 107 NaCl, 6 KCl, 1.2 MgSO4, 1.2 KH2PO4 2CaCl2, 11.5 D-glucose, 25 HEPES, pH 7.35 adjusted withNaOH; HPSS) and loaded with 2 mM Fura-2/acetoxymethyl-ester for 20 min at room temperature in the dark. Afterde-esterification in fresh-loading HPSS for an additional 20min at room temperature, coverslip-attached cells weremounted in an Attofluor® cell chamber. The fluorescence ofFura-2-loaded cells was then monitored with an OlympusIX70 inverted microscope at 3 400 with a xenon arc lampphotomultiplier system (Photon Technology InternationalLtd., Monmouth Junction, NJ), and data were acquired andanalyzed with PTI Felix software. Epifluorescence (signalaveraged) was measured from three to five A549 cells in aconfluent monolayer and the changes in [Ca21]i were ex-pressed as the fluorescence ratio of the Ca21-bound (340 nm)to Ca21-unbound (380 nm) excitation wavelengths emitted at510 nm.
Patch-clamp electrophysiology
Conventional whole-cell voltage-clamp configuration was per-formed to measure transmembrane currents in single A549cells by the standard giga seal patch-clamp technique. Con-fluent A549 cells were trypsin dispersed, seeded onto 35 mmplastic culture dishes, and allowed to reattach 4 to 6 h beforepatch-clamp experiments were performed. Patch-clamp re-cordings were obtained using single (electrically isolated)A549 cells. Recording pipettes were heat polished to producea tip resistance in the range of 3 to 5 MV in our internalsolution. The pipette solution contained (in mM) 130 N-methyl-D-glucamine, 10 HEPES, 1.15 EGTA, 1 Ca(OH)2, 2 Mg21-ATP, 1 NPA, 0.1 NPPB ([Ca21] was estimated as 100 nM) (pH7.2 adjusted with methane sulfonic acid). The external (bath)solution contained (in mM) 120 aspartic acid, 5 Ca(OH)2, 5CaCl2, 10 HEPES, 0.5 3,4-diaminopyridine (pH 7.4 adjustedwith tetraethylammonium hydroxide). Both solutions wereadjusted to 290–300 mosM with sucrose. Currents wererecorded with a computer-controlled EPC9 patch-clamp am-plifier (HEKA; Lambrecht, Germany). Cell capacitance andseries resistance were calculated with the software supportedinternal routines of the EPC9 and compensated before eachexperiment. Voltage pulses were applied from 2100 to 160mV in 20 mV increments after the whole-cell configurationwas achieved, with 200 ms duration during each voltage stepand 2 s interval between steps. The holding potential betweeneach step was 0 mV. Data acquisition and analysis wereperformed with Pulse/PulseFit software (HEKA) and filteredat 2.9 kHz.
RESULTS
Trp1 in A549 cells
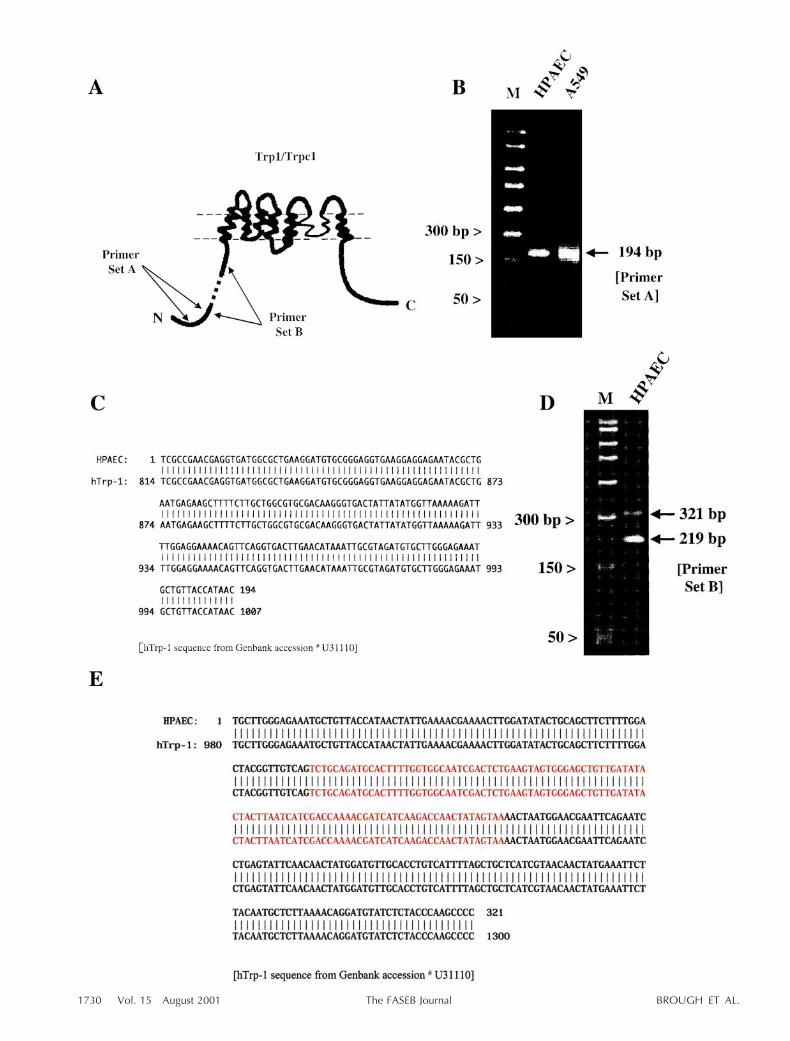
Expression of Trp proteins has not been established inA549 cells. Therefore, initial experiments used RT-PCRcloning to examine whether Trp1 and its splice variant,Trp1b, are expressed in both A549 and human pulmo-nary artery endothelial cells. Sequence specific primerswere generated against discrete regions encoding forportions of the amino-terminal cytosolic loop of Trp1.As indicated in Fig. 1A, primer set A targeted anupstream nucleotide region whereas primer set Bspanned the 102 nucleotide deletion present in Trp1b.Primer set A generated a RT-PCR product of expectedsize in both A549 and endothelial cells which sequenceanalysis verified was Trp1 (100% nucleotide identity)(Fig. 1B, C). Primer set B generated two products ofpredicted size for Trp1 and Trp1b (Fig. 1D, E) that,when sequenced, verified expression of both the full-length and splice variant products. Additional primersets were used to test for homology to Trp1 in trans-membrane spanning and carboxy-terminal regions(data not shown). In each case, single products ofexpected sizes were obtained and shown to be Trp1 bysequence analysis. Using this RT-PCR approach, we alsoidentified a product that encoded an apparent pseudo-gene with high homology to the Trp1d reported inmouse (data not shown) (40). These data collectivelydemonstrate that A549 and endothelial cells expressTrp1, its splice variant Trp1b, and the Trp1d pseudo-gene.
Inhibition of Trp1 translation
To examine the role of endogenous Trp1 in store-operated Ca21 entry, we used a gene walking approachwith 20 different chimeric 29-O-(2-methoxy)ethylphos-phothioate antisense oligonucleotides (Table 1) thatcontained 10 central phosphorothioate oligodeoxy res-idues (‘oligodeoxy gap’) flanked by five 29-O-(2-me-thoxy) residues on the 39 and 59 ends, similar to earlierdescriptions for other mRNA targets (Fig. 2) (41, 42).These methoxy, ethoxy modifications are used to im-part greater hybridizing affinity of oligonucleotides fortheir target and to improve nuclease resistance. Testoligonucleotides were designed to span the start codonand extend through the coding region and into the39-untranslated region of human Trp1 mRNA.
Since phosphorothioate oligonucleotides act princi-pally through RNase H-mediated cleavage of mRNA(43), antisense inhibition of the Trp1 message wasdetermined by Northern blot analysis using a cDNAprobe that hybridized upstream of the stop codon (Fig.2). This probe was designed to detect both Trp1 andTrp1b mRNA. Cationic lipids (DOTMA/DOPE; Lipo-fectin®) were used to facilitate uptake of antisenseoligonucleotides into A549 cells. Figure 3 illustrateshybridization of the cDNA probe for Trp1 with an ;4.5kb pair transcript, similar to previous reports for Trp1
1729TRP1 AND STORE-OPERATED Ca21 ENTRY
1730 Vol. 15 August 2001 BROUGH ET AL.The FASEB Journal

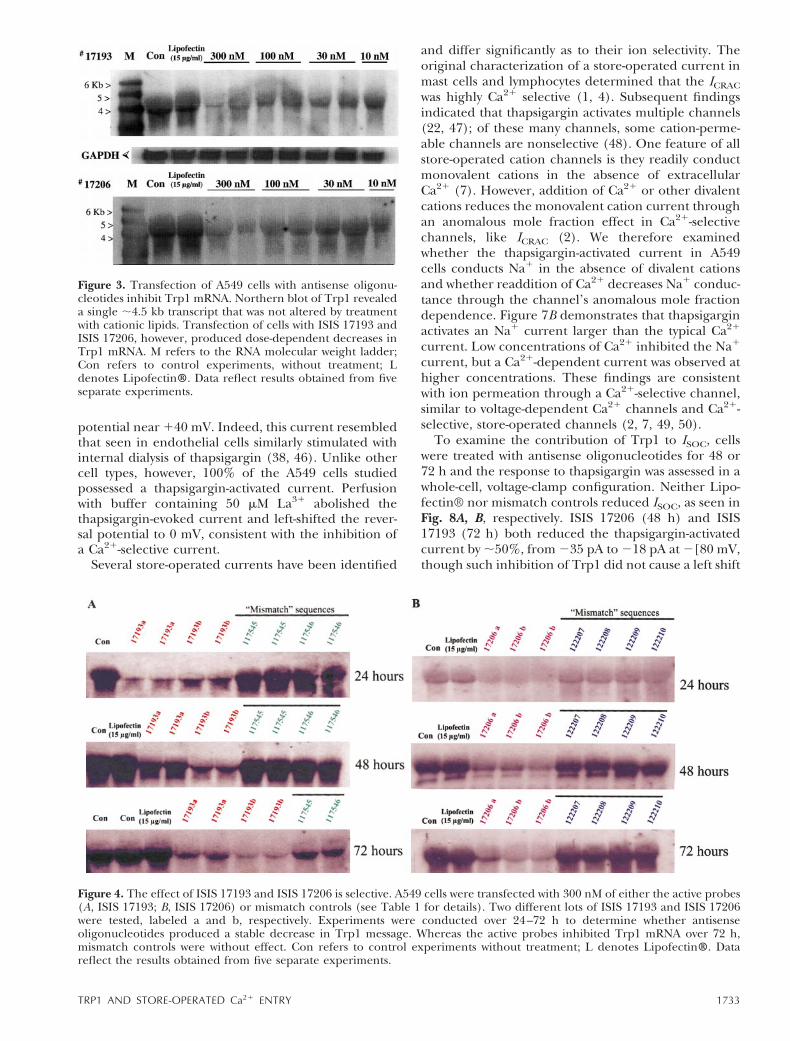
(44, 45). Incubation of A549 cells with cationic lipidsdid not influence mRNA levels. However, two ISISoligonucleotides, 17193 and 17206, produced dose-dependent decreases in Trp1 mRNA. Indeed, of the 20oligonucleotides screened, ISIS 17193 and ISIS 17206reproducibly generated the greatest level of inhibition(data not shown) suggesting these oligonucleotidestarget hypersensitive sites in the Trp1 coding and39-untranslated regions, respectively. Thus, ISIS 17193and ISIS 17206 were selected on the basis of greaterefficacy for further evaluation.
Sequence specificity of antisense oligonucleotides forTrp1 mRNA was examined using a series of ‘mismatch’control analogs, which contain the same base compo-sition as ISIS 17193 and ISIS 17206 but are comprisedof scrambled and noncomplementary sequences toTrp1 (Table 1). These studies were conducted over a72 h time course (Fig. 4). Consistent with the findingsin Fig. 3, ISIS 17193 inhibited Trp1 mRNA for 72 h.Mismatch controls were without effect until at 72 h,when mismatch probes slightly reduced the Trp1 mes-sage. Similarly, ISIS 17206 inhibited Trp1 mRNA for
72 h, but in this case the mismatch controls werewithout effect over the entire time course. These datasupport the specific and stable inhibition of Trp1mRNA by antisense probes.
Trp1 and store-operated Ca21 entry
We next examined the role of Trp1 in store-operatedCa21 entry, using the endoplasmic reticulum Ca21
ATPase inhibitor thapsigargin. Application of thapsi-gargin produced a slowly developing increase in[Ca21]i that peaked near 600 nM, followed by a sus-tained plateau near 300 nM. The thapsigargin responsewas associated with two clearly discernible increases in[Ca21]i, the first due to Ca21 release from intracellularstores and the second due exclusively to Ca21 entryacross the cell membrane (Fig. 5A). In typical [Ca21]iresponses to thapsigargin in other cell types (14, 15,36), Ca21 release and entry phases are not clearlyresolved when physiological concentrations of extracel-lular Ca21 are present. Individual 340 and 380 wave-
TABLE 1. Antisense oligonucleotides targeting Trp1
Oligonucleotide no. Sequence (59339) cDNA target
ISIS 17188 TACAGGGCCGCCATCATCGC AUG codonISIS 17189 GGCGCCCGAGAGGTCCGTGC CodingISIS 17190 GTCACCCTTGTCGCACGCCA CodingISIS 17191 GCATCTGCAGACTGACAACC CodingISIS 17192 CGAAACCTGGAATGCCGGAG CodingISIS 17193 TTCCTGAATTCCACCTCCAC CodingISIS 17194 CCCCGTTTGTCAAGAGGCTC CodingISIS 17195 GTGGGCTTGCGTCGGTAACC CodingISIS 17196 GGCCCCATTGTGTTTTTCTT CodingISIS 17197 TGAGCAACCACTTTGAGGGC CodingISIS 17198 ATGCATCCCAATCCTTCCGA CodingISIS 17199 ATGGACCCAAGATAGAGCTG CodingISIS 17200 TGCCAATGAACGAATGGAAG CodingISIS 17201 CCCACAAAGGACTGCAGTTC CodingISIS 17202 TATGAAGCATTGCCACCAGC CodingISIS 17203 TTTGCCGTTTGACCTTGCCT CodingISIS 17204 ATCTTGGCGCAGTTCGTTTA CodingISIS 17205 TCGCTCCATGATTTAGAAAA 39 UTRISIS 17206 TGAAAATTATTCGCTCCATG 39 UTRISIS 17207 AATATTTTAAAGGAACGGCT 39 UTRISIS 122207 TGAAAATTCTTCGATCCATG Scrambled for 17206ISIS 122208 TAAAGATTATTTGCTCCACG Scrambled for 17206ISIS 122209 GTAAATTTAATCGGTCCATC Scrambled for 17206ISIS 122210 TAAAGATCATGTGATCCCTT Scrambled for 17206ISIS 117545 TCCCCGCATAACTCTTCTAC Scrambled for 17193ISIS 117546 ATTCTAACTTACGCCCCCTC Scrambled for 17193
Š
Figure 1. A549 and pulmonary artery endothelial cells express Trp1 and its splice variant, Trp1b. A) Model of Trp1 (modifiedfrom ref 29) illustrating discrete sites targeted by RT-PCR primer sets. Forward and reverse primers are indicated and weredirected against nucleotides in the amino-terminal coding sequence. Primer set B spanned the region demarcating the Trp1splice variant, Trp1b. B) Primer set A generated a product of expected size in both cell types. C) Cloning and nucleotidealignment revealed 100% identity between the cloned products and human Trp1. Nucleotide sequence of Trp1 is depicted andcompared with the currently cloned partial product. D) Primer set B, which spans the splice site in Trp1b, generated twoproducts. E) Cloning and nucleotide alignment of these two products revealed 100% homology between the cloned productsand Trp1 and Trp1b, respectively. Nucleotide numbers of Trp1 are depicted and compared with the currently cloned partialproduct. The spliced region, deleted in Trp1b, is depicted in red.
1731TRP1 AND STORE-OPERATED Ca21 ENTRY

lengths are shown in Fig. 5A to show that they move inopposition, demonstrating that the lag between Ca21
release and Ca21 entry is not an artifact of fura fluores-cence.
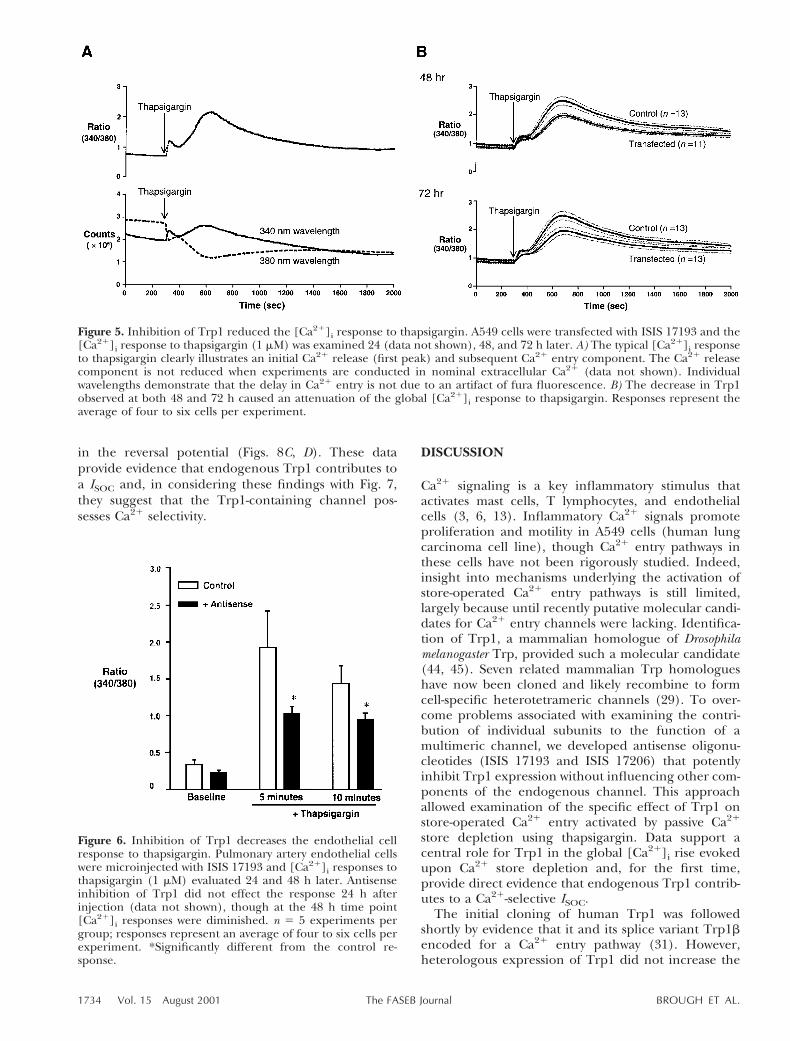
We examined the role of Trp1 on the thapsigargin-induced rise in global [Ca21]i. Twenty-four hours afterA549 cells were treated with ISIS 17193 to inhibit Trp1mRNA, the [Ca21]i response was not different from thecontrol response (data not shown), suggesting Trp1protein turnover may be slow. However, both 48 and72 h treatment with ISIS 17193 resulted in a 25%decrease in the [Ca21]i response to thapsigargin (Fig.5B). We confirmed the role of Trp1 in store-operatedCa21 entry in endothelial cells, first microinjecting theantisense oligonucleotides into the cytosol and measur-ing the [Ca21]i response 24, 48, and 72 h later. Again,the [Ca21]i response to thapsigargin was not differentfrom control responses at 24 h (data not shown), butwas decreased after 48 and 72 h exposure to antisenseoligonucleotide inhibition of Trp1 mRNA (Fig. 6).These data therefore support a central role for Trp1 as
an endogenous component of store-operated Ca21
entry channels.
Trp1 and ISOC
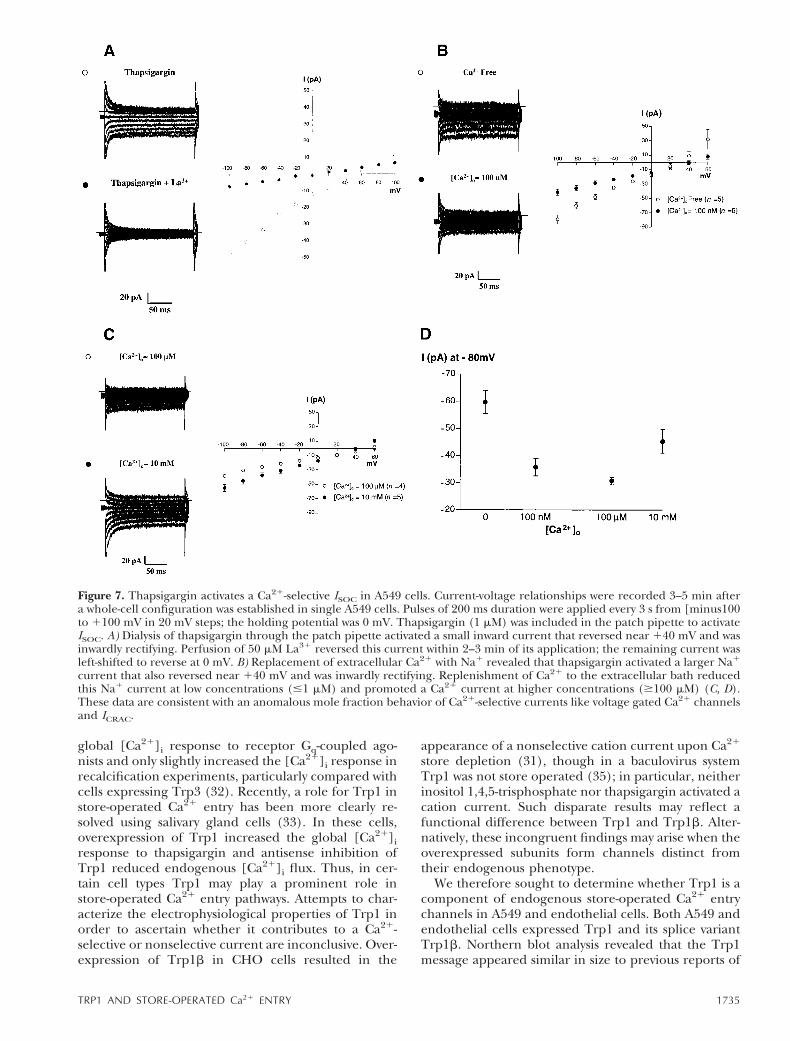
Heterologous expression of Trp1b encoded for a non-selective channel in CHO cells (31), although thischannel was not store operated when expressed in thebaculovirus/Sf9 system (35). Similarly, expression ofTrp1 encoded for a nonselective channel that in-creased whole-cell currents or increased global [Ca21]iresponses (32). The contribution of endogenous Trp1to ISOC, however, is poorly resolved, especially consid-ering that cells express multiple Trp proteins likely toform heteromultimers of various combinations (29).Our next studies therefore examined the contributionof Trp1 to an endogenous ISOC in A549 cells. Figure 7illustrates a typical whole-cell Ca21 current activated oninternal dialysis with thapsigargin through the patchpipette. The Ca21 current was small (; 235 pA at 280mV), inwardly rectifying, and had a positive reversal
Figure 2. Target regions in Trp1 mRNA. Trp1 gene start (green) and stop (red) codons are shown. Sites targeted by ISISantisense probes that stretch across the start codon, through the coding region, and in the 39-untranslated region are underlinedand numbered. Two effective antisense probes, ISIS 17193 and ISIS 17206, are shown in red. The probe generated for Northernblot analyses is illustrated in blue. Note this probe detects both Trp1 and Trp1b mRNA.
1732 Vol. 15 August 2001 BROUGH ET AL.The FASEB Journal
potential near 140 mV. Indeed, this current resembledthat seen in endothelial cells similarly stimulated withinternal dialysis of thapsigargin (38, 46). Unlike othercell types, however, 100% of the A549 cells studiedpossessed a thapsigargin-activated current. Perfusionwith buffer containing 50 mM La31 abolished thethapsigargin-evoked current and left-shifted the rever-sal potential to 0 mV, consistent with the inhibition ofa Ca21-selective current.
Several store-operated currents have been identified
and differ significantly as to their ion selectivity. Theoriginal characterization of a store-operated current inmast cells and lymphocytes determined that the ICRACwas highly Ca21 selective (1, 4). Subsequent findingsindicated that thapsigargin activates multiple channels(22, 47); of these many channels, some cation-perme-able channels are nonselective (48). One feature of allstore-operated cation channels is they readily conductmonovalent cations in the absence of extracellularCa21 (7). However, addition of Ca21 or other divalentcations reduces the monovalent cation current throughan anomalous mole fraction effect in Ca21-selectivechannels, like ICRAC (2). We therefore examinedwhether the thapsigargin-activated current in A549cells conducts Na1 in the absence of divalent cationsand whether readdition of Ca21 decreases Na1 conduc-tance through the channel’s anomalous mole fractiondependence. Figure 7B demonstrates that thapsigarginactivates an Na1 current larger than the typical Ca21
current. Low concentrations of Ca21 inhibited the Na1
current, but a Ca21-dependent current was observed athigher concentrations. These findings are consistentwith ion permeation through a Ca21-selective channel,similar to voltage-dependent Ca21 channels and Ca21-selective, store-operated channels (2, 7, 49, 50).
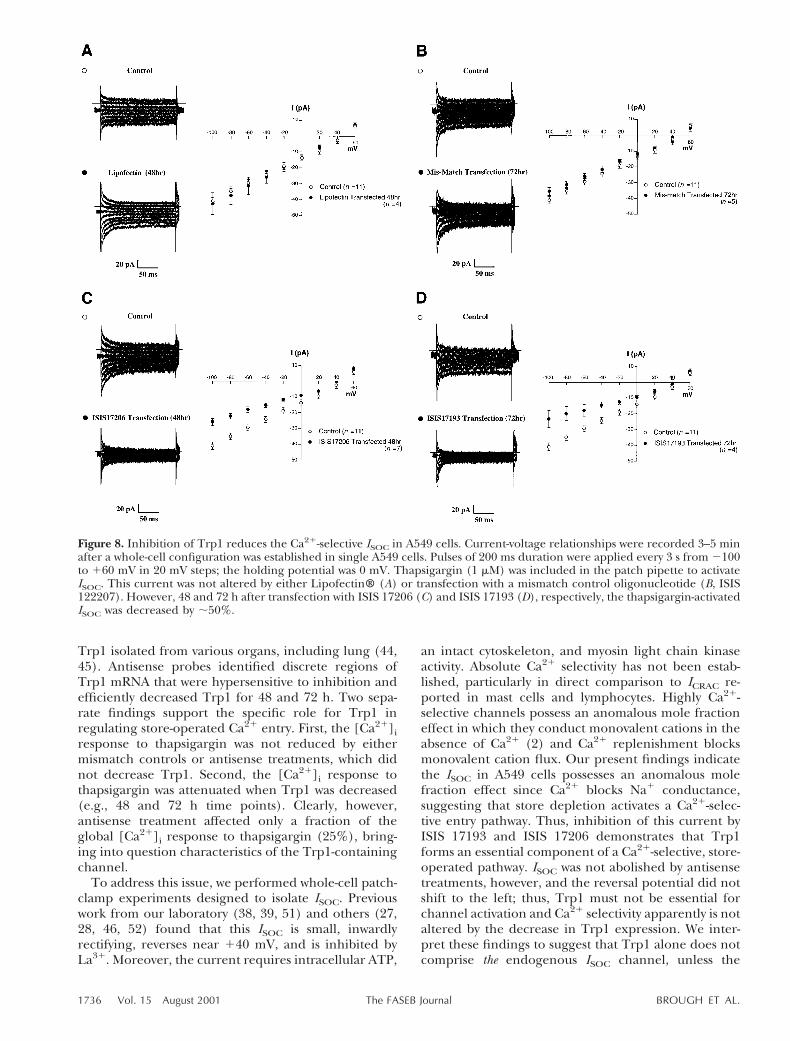
To examine the contribution of Trp1 to ISOC, cellswere treated with antisense oligonucleotides for 48 or72 h and the response to thapsigargin was assessed in awhole-cell, voltage-clamp configuration. Neither Lipo-fectin® nor mismatch controls reduced ISOC, as seen inFig. 8A, B, respectively. ISIS 17206 (48 h) and ISIS17193 (72 h) both reduced the thapsigargin-activatedcurrent by ;50%, from 235 pA to 218 pA at 2[80 mV,though such inhibition of Trp1 did not cause a left shift
Figure 3. Transfection of A549 cells with antisense oligonu-cleotides inhibit Trp1 mRNA. Northern blot of Trp1 revealeda single ;4.5 kb transcript that was not altered by treatmentwith cationic lipids. Transfection of cells with ISIS 17193 andISIS 17206, however, produced dose-dependent decreases inTrp1 mRNA. M refers to the RNA molecular weight ladder;Con refers to control experiments, without treatment; Ldenotes Lipofectin®. Data reflect results obtained from fiveseparate experiments.
Figure 4. The effect of ISIS 17193 and ISIS 17206 is selective. A549 cells were transfected with 300 nM of either the active probes(A, ISIS 17193; B, ISIS 17206) or mismatch controls (see Table 1 for details). Two different lots of ISIS 17193 and ISIS 17206were tested, labeled a and b, respectively. Experiments were conducted over 24–72 h to determine whether antisenseoligonucleotides produced a stable decrease in Trp1 message. Whereas the active probes inhibited Trp1 mRNA over 72 h,mismatch controls were without effect. Con refers to control experiments without treatment; L denotes Lipofectin®. Datareflect the results obtained from five separate experiments.
1733TRP1 AND STORE-OPERATED Ca21 ENTRY
in the reversal potential (Figs. 8C, D). These dataprovide evidence that endogenous Trp1 contributes toa ISOC and, in considering these findings with Fig. 7,they suggest that the Trp1-containing channel pos-sesses Ca21 selectivity.
DISCUSSION
Ca21 signaling is a key inflammatory stimulus thatactivates mast cells, T lymphocytes, and endothelialcells (3, 6, 13). Inflammatory Ca21 signals promoteproliferation and motility in A549 cells (human lungcarcinoma cell line), though Ca21 entry pathways inthese cells have not been rigorously studied. Indeed,insight into mechanisms underlying the activation ofstore-operated Ca21 entry pathways is still limited,largely because until recently putative molecular candi-dates for Ca21 entry channels were lacking. Identifica-tion of Trp1, a mammalian homologue of Drosophilamelanogaster Trp, provided such a molecular candidate(44, 45). Seven related mammalian Trp homologueshave now been cloned and likely recombine to formcell-specific heterotetrameric channels (29). To over-come problems associated with examining the contri-bution of individual subunits to the function of amultimeric channel, we developed antisense oligonu-cleotides (ISIS 17193 and ISIS 17206) that potentlyinhibit Trp1 expression without influencing other com-ponents of the endogenous channel. This approachallowed examination of the specific effect of Trp1 onstore-operated Ca21 entry activated by passive Ca21
store depletion using thapsigargin. Data support acentral role for Trp1 in the global [Ca21]i rise evokedupon Ca21 store depletion and, for the first time,provide direct evidence that endogenous Trp1 contrib-utes to a Ca21-selective ISOC.
The initial cloning of human Trp1 was followedshortly by evidence that it and its splice variant Trp1bencoded for a Ca21 entry pathway (31). However,heterologous expression of Trp1 did not increase the
Figure 5. Inhibition of Trp1 reduced the [Ca21]i response to thapsigargin. A549 cells were transfected with ISIS 17193 and the[Ca21]i response to thapsigargin (1 mM) was examined 24 (data not shown), 48, and 72 h later. A) The typical [Ca21]i responseto thapsigargin clearly illustrates an initial Ca21 release (first peak) and subsequent Ca21 entry component. The Ca21 releasecomponent is not reduced when experiments are conducted in nominal extracellular Ca21 (data not shown). Individualwavelengths demonstrate that the delay in Ca21 entry is not due to an artifact of fura fluorescence. B) The decrease in Trp1observed at both 48 and 72 h caused an attenuation of the global [Ca21]i response to thapsigargin. Responses represent theaverage of four to six cells per experiment.
Figure 6. Inhibition of Trp1 decreases the endothelial cellresponse to thapsigargin. Pulmonary artery endothelial cellswere microinjected with ISIS 17193 and [Ca21]i responses tothapsigargin (1 mM) evaluated 24 and 48 h later. Antisenseinhibition of Trp1 did not effect the response 24 h afterinjection (data not shown), though at the 48 h time point[Ca21]i responses were diminished. n 5 5 experiments pergroup; responses represent an average of four to six cells perexperiment. *Significantly different from the control re-sponse.
1734 Vol. 15 August 2001 BROUGH ET AL.The FASEB Journal
global [Ca21]i response to receptor Gq-coupled ago-nists and only slightly increased the [Ca21]i response inrecalcification experiments, particularly compared withcells expressing Trp3 (32). Recently, a role for Trp1 instore-operated Ca21 entry has been more clearly re-solved using salivary gland cells (33). In these cells,overexpression of Trp1 increased the global [Ca21]iresponse to thapsigargin and antisense inhibition ofTrp1 reduced endogenous [Ca21]i flux. Thus, in cer-tain cell types Trp1 may play a prominent role instore-operated Ca21 entry pathways. Attempts to char-acterize the electrophysiological properties of Trp1 inorder to ascertain whether it contributes to a Ca21-selective or nonselective current are inconclusive. Over-expression of Trp1b in CHO cells resulted in the
appearance of a nonselective cation current upon Ca21
store depletion (31), though in a baculovirus systemTrp1 was not store operated (35); in particular, neitherinositol 1,4,5-trisphosphate nor thapsigargin activated acation current. Such disparate results may reflect afunctional difference between Trp1 and Trp1b. Alter-natively, these incongruent findings may arise when theoverexpressed subunits form channels distinct fromtheir endogenous phenotype.
We therefore sought to determine whether Trp1 is acomponent of endogenous store-operated Ca21 entrychannels in A549 and endothelial cells. Both A549 andendothelial cells expressed Trp1 and its splice variantTrp1b. Northern blot analysis revealed that the Trp1message appeared similar in size to previous reports of
Figure 7. Thapsigargin activates a Ca21-selective ISOC in A549 cells. Current-voltage relationships were recorded 3–5 min aftera whole-cell configuration was established in single A549 cells. Pulses of 200 ms duration were applied every 3 s from [minus100to 1100 mV in 20 mV steps; the holding potential was 0 mV. Thapsigargin (1 mM) was included in the patch pipette to activateISOC. A) Dialysis of thapsigargin through the patch pipette activated a small inward current that reversed near 140 mV and wasinwardly rectifying. Perfusion of 50 mM La31 reversed this current within 2–3 min of its application; the remaining current wasleft-shifted to reverse at 0 mV. B) Replacement of extracellular Ca21 with Na1 revealed that thapsigargin activated a larger Na1
current that also reversed near 140 mV and was inwardly rectifying. Replenishment of Ca21 to the extracellular bath reducedthis Na1 current at low concentrations (#1 mM) and promoted a Ca21 current at higher concentrations ($100 mM) (C, D).These data are consistent with an anomalous mole fraction behavior of Ca21-selective currents like voltage gated Ca21 channelsand ICRAC.
1735TRP1 AND STORE-OPERATED Ca21 ENTRY
Trp1 isolated from various organs, including lung (44,45). Antisense probes identified discrete regions ofTrp1 mRNA that were hypersensitive to inhibition andefficiently decreased Trp1 for 48 and 72 h. Two sepa-rate findings support the specific role for Trp1 inregulating store-operated Ca21 entry. First, the [Ca21]iresponse to thapsigargin was not reduced by eithermismatch controls or antisense treatments, which didnot decrease Trp1. Second, the [Ca21]i response tothapsigargin was attenuated when Trp1 was decreased(e.g., 48 and 72 h time points). Clearly, however,antisense treatment affected only a fraction of theglobal [Ca21]i response to thapsigargin (25%), bring-ing into question characteristics of the Trp1-containingchannel.
To address this issue, we performed whole-cell patch-clamp experiments designed to isolate ISOC. Previouswork from our laboratory (38, 39, 51) and others (27,28, 46, 52) found that this ISOC is small, inwardlyrectifying, reverses near 140 mV, and is inhibited byLa31. Moreover, the current requires intracellular ATP,
an intact cytoskeleton, and myosin light chain kinaseactivity. Absolute Ca21 selectivity has not been estab-lished, particularly in direct comparison to ICRAC re-ported in mast cells and lymphocytes. Highly Ca21-selective channels possess an anomalous mole fractioneffect in which they conduct monovalent cations in theabsence of Ca21 (2) and Ca21 replenishment blocksmonovalent cation flux. Our present findings indicatethe ISOC in A549 cells possesses an anomalous molefraction effect since Ca21 blocks Na1 conductance,suggesting that store depletion activates a Ca21-selec-tive entry pathway. Thus, inhibition of this current byISIS 17193 and ISIS 17206 demonstrates that Trp1forms an essential component of a Ca21-selective, store-operated pathway. ISOC was not abolished by antisensetreatments, however, and the reversal potential did notshift to the left; thus, Trp1 must not be essential forchannel activation and Ca21 selectivity apparently is notaltered by the decrease in Trp1 expression. We inter-pret these findings to suggest that Trp1 alone does notcomprise the endogenous ISOC channel, unless the
Figure 8. Inhibition of Trp1 reduces the Ca21-selective ISOC in A549 cells. Current-voltage relationships were recorded 3–5 minafter a whole-cell configuration was established in single A549 cells. Pulses of 200 ms duration were applied every 3 s from 2100to 160 mV in 20 mV steps; the holding potential was 0 mV. Thapsigargin (1 mM) was included in the patch pipette to activateISOC. This current was not altered by either Lipofectin® (A) or transfection with a mismatch control oligonucleotide (B, ISIS122207). However, 48 and 72 h after transfection with ISIS 17206 (C) and ISIS 17193 (D), respectively, the thapsigargin-activatedISOC was decreased by ;50%.
1736 Vol. 15 August 2001 BROUGH ET AL.The FASEB Journal

residual Trp1 protein (e.g., that not inhibited by anti-sense treatments) is sufficiently high to account for theremaining current. Both A549 and endothelial cellsexpress other Trp proteins, particularly Trp4; indeed,Trp4 is more abundantly expressed than is Trp1(T. Stevens, unpublished observations). Work by Flock-erzi and colleagues has illustrated that Trp4 encodesfor a Ca21-selective channel (53–55). Though specula-tive, Trp1 and Trp4 may form an endogenous channelin which gating and Ca21 selectivity are determined bythe presence of Trp4. Future studies will be required toassess whether Trp1/Trp4 heteromultimers form theendogenous A549 and endothelial cell ISOC channeland whether the current remaining after Trp1 inhibi-tion is due to the presence of Trp4.
In summary, our findings support a role for Trp1 inan endogenous store-operated and selective Ca21 entrypathway. This conclusion was made possible throughantisense inhibition of Trp1 mRNA translation at twodiscrete hypersensitive sites, including one in the cod-ing region and one in the 39-untranslated region, thatallowed for the selective inhibition of endogenousTrp1. The physiological significance of these observa-tions remain speculative. However, earlier studies haveestablished a central role for store-operated Ca21 entryin the response to inflammation. Thus, it is likely thatTrp1 makes an important contribution to the cell’sresponse to inflammation.
The authors wish to thank Ms. Judy Creighton and TrayWeathington for their excellent assistance in cell cultureexperiments. This work was supported by HL56056 andHL60024 (to T.S.), and DK50151 (to M.L.). S.W. is anAmerican Heart Association, Southeastern Consortium Fel-low.
REFERENCES
1. Hoth, M., and Penner, R. (1992) Depletion of intracellularcalcium stores activates a calcium current in mast cells [seecomments]. Nature (London) 355, 353–356
2. Hoth, M. (1995) Calcium and barium permeation throughcalcium release-activated calcium (CRAC) channels. Pf luegersArch. 430, 315–322
3. Huber, M., Hughes, M. R., and Krystal, G. (2000) Thapsigargin-induced degranulation of mast cells is dependent on transientactivation of phosphatidylinositol-3 kinase. J. Immunol. 165,124–133
4. Zweifach, A., and Lewis, R. S. (1993) Mitogen-regulated Ca21
current of T lymphocytes is activated by depletion of intracellu-lar Ca21 stores. Proc. Natl. Acad. Sci. USA 90, 6295–6299
5. Zweifach, A., and Lewis, R. S. (1996) Calcium-dependent poten-tiation of store-operated calcium channels in T lymphocytes.J. Gen. Physiol. 107, 597–610
6. Zweifach, A. (2000) Target-cell contact activates a highly selec-tive capacitative calcium entry pathway in cytotoxic T lympho-cytes. J. Cell Biol. 148, 603–614
7. Lepple-Wienhues, A., and Cahalan, M. D. (1996) Conductanceand permeation of monovalent cations through depletion-activated Ca21 channels (ICRAC) in Jurkat T cells. Biophys. J. 71,787–794
8. Fierro, L., Lund, P. E., and Parekh, A. B. (2000) Comparison ofthe activation of the Ca21 release-activated Ca21 current ICRACto InsP3 in Jurkat T-lymphocytes, pulmonary artery endotheliaand RBL-1 cells. Pfluegers Arch. 440, 580–587
9. Chetham, P. M., Guldemeester, H. A., Mons, N., Brough, G. H.,Bridges, J. P., Thompson, W. J., and Stevens, T. (1997) Ca21-inhibitable adenylyl cyclase and pulmonary microvascular per-meability. Am. J. Physiol. 273, L22–L30
10. Chetham, P. M., Babal, P., Bridges, J. P., Moore, T. M., andStevens, T. (1999) Segmental regulation of pulmonary vascularpermeability by store-operated Ca21 entry. Am. J. Physiol. 276,L41–L50
11. Kelly, J. J., Moore, T. M., Babal, P., Diwan, A. H., Stevens, T., andThompson, W. J. (1998) Pulmonary microvascular and macro-vascular endothelial cells: differential regulation of Ca21 andpermeability. Am. J. Physiol. 274, L810–L819
12. Moore, T. M., Brough, G. H., Babal, P., Kelly, J. J., Li, M., andStevens, T. (1998) Store-operated calcium entry promotes shapechange in pulmonary endothelial cells expressing Trp1. Am. J.Physiol. 275, L574–L582
13. Moore, T. M., Chetham, P. M., Kelly, J. J., and Stevens, T. (1998)Signal transduction and regulation of lung endothelial cellpermeability. Interaction between calcium and cAMP. Am. J.Physiol. 275, L203–L222
14. Stevens, T., Nakahashi, Y., Cornfield, D. N., McMurtry, I. F.,Cooper, D. M., and Rodman, D. M. (1995) Ca21-inhibitableadenylyl cyclase modulates pulmonary artery endothelial cellcAMP content and barrier function. Proc. Natl. Acad. Sci. USA 92,2696–2700
15. Stevens, T., Fouty, B., Hepler, L., Richardson, D., Brough, G.,McMurtry, I. F., and Rodman, D. M. (1997) Cytosolic Ca21 andadenylyl cyclase responses in phenotypically distinct pulmonaryendothelial cells. Am. J. Physiol. 272, L51–L59
16. Huang, A. J., Manning, J. E., Bandak, T. M., Ratau, M. C.,Hanser, K. R., and Silverstein, S. C. (1993) Endothelial cellcytosolic free calcium regulates neutrophil migration acrossmonolayers of endothelial cells. J. Cell Biol. 120, 1371–1380
17. Saito, H., Minamiya, Y., Kitamura, M., Saito, S., Enomoto, K.,Terada, K., and Ogawa, J. (1998) Endothelial myosin light chainkinase regulates neutrophil migration across human umbilicalvein endothelial cell monolayer. J. Immunol. 161, 1533–1540
18. Pfau, S., Leitenberg, D., Rinder, H., Smith, B. R., Pardi, R., andBender, J. R. (1995) Lymphocyte adhesion-dependent calciumsignaling in human endothelial cells. J. Cell Biol. 128, 969–978
19. Simon, S. I., Cherapanov, V., Nadra, I., Waddell, T. K., Seo,S. M., Wang, Q., Doerschuk, C. M., and Downey, G. P. (1999)Signaling functions of L-selectin in neutrophils: alterations inthe cytoskeleton and colocalization with CD18. J. Immunol. 163,2891–2901
20. Berridge, M. J. (1997) Elementary and global aspects of calciumsignalling. J. Physiol. (London) 499, 291–306
21. Berridge, M. J., Lipp, P., and Bootman, M. D. (2000) Signaltransduction. The calcium entry pas de deux. Science 287,1604–1605
22. Hofmann, T., Schaefer, M., Schultz, G., and Gudermann, T.(2000) Transient receptor potential channels as molecularsubstrates of receptor-mediated cation entry. J. Mol. Med. 78,14–25
23. Lewis, R. S. (1999) Store-operated calcium channels. Adv. SecondMessenger Phosphoprotein Res. 33, 279–307
24. Parekh, A. B., and Penner, R. (1997) Store depletion andcalcium influx. Physiol. Rev. 77, 901–930
25. Putney, J. W., Jr. (1999) “Kissin’ cousins”: intimate plasmamembrane–ER interactions underlie capacitative calcium entry.Cell 99, 5–8
26. Vaca, L., and Kunze, D. L. (1993) Depletion and refilling ofintracellular Ca21 stores induce oscillations of Ca21 current.Am. J. Physiol. 264, H1319–H1322
27. Vaca, L., and Kunze, D. L. (1994) Depletion of intracellularCa21 stores activates a Ca21-selective channel in vascular endo-thelium. Am. J. Physiol. 267, C920–C925
28. Vaca, L., and Kunze, D. L. (1995) IP3-activated Ca21 channels inthe plasma membrane of cultured vascular endothelial cells.Am. J. Physiol. 269, C733–C738
29. Birnbaumer, L., Zhu, X., Jiang, M., Boulay, G., Peyton, M.,Vannier, B., Brown, D., Platano, D., Sadeghi, H., Stefani, E., andBirnbaumer, M. (1996) On the molecular basis and regulationof cellular capacitative calcium entry: roles for Trp proteins.Proc. Natl. Acad. Sci. USA 93, 15195–15202
30. Lintschinger, B., Balzer-Geldsetzer, M., Baskaran, T., Graier,W. F., Romanin, C., Zhu, M. X., and Groschner, K. (2000)
1737TRP1 AND STORE-OPERATED Ca21 ENTRY

Coassembly of Trp1 and Trp3 channels generates diacylglyceroland Ca21-sensitive cation channels. J. Biol. Chem. 275, 27799–27805
31. Zitt, C., Zobel, A., Obukhov, A. G., Harteneck, C., Kalkbrenner,F., Luckhoff, A., and Schultz, G. (1996) Cloning and functionalexpression of a human Ca21-permeable cation channel acti-vated by calcium store depletion. Neuron 16, 1189–1196
32. Zhu, X., Jiang, M., Peyton, M., Boulay, G., Hurst, R., Stefani, E., andBirnbaumer, L. (1996) trp, a novel mammalian gene family essen-tial for agonist-activated capacitative Ca21 entry. Cell 85, 661–671
33. Liu, X., Wang, W., Singh, B. B., Lockwich, T., Jadlowiec, J.,O’Connell, B., Wellner, R., Zhu, M. X., and Ambudkar, I. S.(2000) Trp1, a candidate protein for the store-operated Ca21
influx mechanism in salivary gland cells [published erratumappears in J. Biol. Chem., 2000, vol. 275, pp. 9890–9891]. J. Biol.Chem. 275, 3403–3411
34. Wu, X., Babnigg, G., and Villereal, M. L. (2000) Functionalsignificance of human trp1 and trp3 in store-operated Ca21
entry in HEK-293 cells. Am. J. Physiol. 278, C526–C53635. Sinkins, W. G., Estacion, M., and Schilling, W. P. (1998)
Functional expression of TrpC1: a human homologue of theDrosophila Trp channel. Biochem. J. 331, 331–339
36. Stevens, T., Creighton, J., and Thompson, W. J. (1999) Controlof cAMP in lung endothelial cell phenotypes. Implications forcontrol of barrier function. Am. J. Physiol. 277, L119–L126
37. Dean, N. M., and Griffey, R. H. (1997) Identification andcharacterization of second-generation antisense oligonucleo-tides. Antisense Nucleic Acid Drug Dev. 7, 229–233
38. Wu, S., Goodman, S., and Stevens, T. (2000) Non-erythroidspectrin physically links calcium store depletion to activation ofcalcium entry in endothelial cells. FASEB J. 14, A411 (abstr.)
39. Moore, T., Norwood, N., Creighton, J., Babal, P., Brough, G.,Shasby, D., and Stevens, T. (2000) Receptor dependent activa-tion of store operated calcium entry increases endothelialpermeability. Am. J. Physiol. 279, L691–L699
40. Sakura, H., and Ashcroft, F. M. (1997) Identification of fourtrp1 gene variants murine pancreatic beta-cells. Diabetologia 40,528–532
41. Zuo, Z., Dean, N. M., and Honkanen, R. E. (1998) Serine/threonine protein phosphatase type 5 acts upstream of p53 toregulate the induction of p21(WAF1/Cip1) and mediate growtharrest. J. Biol. Chem. 273, 12250–12258
42. Cheng, A., Dean, N. M., and Honkanen, R. E. (2000) Serine/threonine protein phosphatase type 1gamma1 is required forthe completion of cytokinesis in human A549 lung carcinomacells. J. Biol. Chem. 275, 1846–1854
43. Crooke, S. T. (1992) Therapeutic applications of oligonucleo-tides. Annu. Rev. Pharmacol. Toxicol. 32, 329–376
44. Wes, P. D., Chevesich, J., Jeromin, A., Rosenberg, C., Stetten, G.,and Montell, C. (1995) TRPC1, a human homolog of a Drosoph-ila store-operated channel. Proc. Natl. Acad. Sci. USA 92, 9652–9656
45. Zhu, X., Chu, P. B., Peyton, M., and Birnbaumer, L. (1995)Molecular cloning of a widely expressed human homologue forthe Drosophila trp gene. FEBS Lett. 373, 193–198
46. Fasolato, C., and Nilius, B. (1998) Store depletion triggers thecalcium release-activated calcium current (ICRAC) in macrovas-cular endothelial cells: a comparison with Jurkat and embryonickidney cell lines. Pfluegers Arch. 436, 69–74
47. Nilius, B., Viana, F., and Droogmans, G. (1997) Ion channels invascular endothelium. Annu. Rev. Physiol. 59, 145–170
48. Wu, S., Moore, T. M., Brough, G. H., Whitt, S. R., Chinkers, M.,Li, M., and Stevens, T. (2000) Cyclic nucleotide-gated channelsmediate membrane depolarization after activation of store-operated calcium entry in endothelial cells. J. Biol. Chem. 275,18887–18896
49. Warnat, J., Philipp, S., Zimmer, S., Flockerzi, V., and Cavalie, A.(1999) Phenotype of a recombinant store-operated channel:highly selective permeation of Ca21. J. Physiol. (London) 518,631–638
50. Hess, P., and Tsien, R. W. (1984) Mechanism of ion permeationthrough calcium channels. Nature (London) 309, 453–456
51. Norwood, N., Moore, T., Dean, D., Bhattacharjee, R., Li, M., andStevens, T. (2000) Store operated calcium entry and increasedendothelial cell permeability. Am. J. Physiol. 279, L815–L824
52. Schilling, W. P., Cabello, O. A., and Rajan, L. (1992) Depletionof the inositol 1,4,5-trisphosphate-sensitive intracellular Ca21
store in vascular endothelial cells activates the agonist-sensitiveCa21-influx pathway. Biochem. J. 284, 521–530
53. Philipp, S., Cavalie, A., Freichel, M., Wissenbach, U., Zimmer, S.,Trost, C., Marquart, A., Murakami, M., and Flockerzi, V. (1996)A mammalian capacitative calcium entry channel homologousto Drosophila TRP and TRPL. EMBO J. 15, 6166–6171
54. Philipp, S., Hambrecht, J., Braslavski, L., Schroth, G., Freichel,M., Murakami, M., Cavalie, A., and Flockerzi, V. (1998) A novelcapacitative calcium entry channel expressed in excitable cells.EMBO J. 17, 4274–4282
55. Philipp, S., Trost, C., Warnat, J., Rautmann, J., Himmerkus, N.,Schroth, G., Kretz, O., Nastainczyk, W., Cavalie, A., Hoth, M.,and Flockerzi, V. (2000) TRP4 (CCE1) protein is part of nativecalcium release-activated Ca21-like channels in adrenal cells.J. Biol. Chem. 275, 23965–23972
Received for publication February 14, 2001.Accepted for publication April 17, 2001.
1738 Vol. 15 August 2001 BROUGH ET AL.The FASEB Journal




















