
Taphonomy and zooarchaeology of the Upper Palaeolithic cave of Dzudzuana, Republic of Georgia

COMPOSITION, DISTRIBUTION, AND TAPHONOMY OF NEARSHORE BENTHICFORAMINIFERA OF THE FARASAN ISLANDS, SOUTHERN RED SEA, SAUDI ARABIA
RAMADAN H. ABU-ZIED1,2,3, RASHAD A. BANTAN
1, ALI S. BASAHAM1, MOHAMED H. EL MAMONEY
1AND
HAMAD A. AL-WASHMI1
ABSTRACT
Nearshore benthic foraminifera of the Farasan Islands(southern Red Sea) were investigated to highlight the faunaldistributions and the controlling environmental factors.Forty-one sediment samples were collected in transects offour areas: Khor As Sailah, Ras Sheidah-Ras Abbrah,Jinabah Bay, and Ras Farasan. Cluster and canonicalcorrespondence analyses were performed to determine if anygroups of samples or species correlated with ambientenvironmental factors. Symbiont-bearing species (e.g., Pe-neroplis planatus, Sorites orbiculus, Neorotalia calcar, andCoscinospira hemprichii) dominate the hypersaline lagoon ofKhor As Sailah, which is characterized by muddy sand,seagrasses, and green filamentous algae. Neorotalia calcardominates the hard substrates with green filamentous algae inthe nearshore area of Ras Sheidah-Ras Abbrah. Both JinabahBay and Ras Farasan have sandy substrates with assemblagesdominated by Ammonia convexa, P. planatus, S. orbiculus,N. calcar, Varidentella neostriatula, Quinqueloculina sp., andElphidium sp., and indications of shoreward transport.
Overall, foraminiferal density was highest (210–3930specimens/g) in the Khor As Sailah lagoon and lowest (8–327 specimens/g) in the nearshore sediments of Jinabah Bayand Ras Farasan. Taphonomic effects were evident asdissolution and bioerosion in the calm waters of Khor AsSailah lagoon, and abrasion and breakage in the nearshoresediments of Jinabah Bay and Ras Farasan.
INTRODUCTION
Distributions and compositions of nearshore benthicforaminifera are mainly controlled by factors such as tides,currents, waves, substrates, salinity, temperature, food,light, dissolved oxygen, bioturbation, and predation (Bol-tovskoy and Wright, 1976; Murray, 1991; Sen Gupta, 1999;Murray, 2000, 2006). Due to interactions of these factors,foraminifers have unpredictable life-cycles and patchydistribution patterns (Ribes and others, 2000; Alve andMurray, 2001; Debenay and others, 2006; Wilson andRamsook, 2007). Where there is seagrass or macroalgae,foraminiferal assemblages and abundances can vary fromone part of the substrate to another (Steinker and Clem,1984; Wilson, 1998; Ribes and others, 2000; Debenay andPayri, 2010). Detailed studies on the distributions ofnearshore and intertidal benthic foraminifera (e.g., Buzas-Stephens and Buzas, 2005; Morvan and others, 2006; Vanceand others, 2006; Wilson and Ramsook, 2007; Abu-Zied
and others, 2007; Berkeley and others, 2008; Debenay andPayri, 2010) have shown how they are determined byparticular environmental parameters.
The nearshore benthic foraminifera of the southern RedSea have received much less attention than those in thenorthern sector, which have been thoroughly investigated(Said, 1949, 1950; Reiss and others, 1977; Bahafzallah,1979; Reiss and Hottinger, 1984; Abou-Ouf and others,1988; Abou-Ouf and El-Shater, 1991; Hottinger and others,1993; Haunold and others, 1997). The purpose of our studywas to determine the composition and distribution ofbenthic foraminiferal assemblages, as well as their control-ling environmental factors, in nearshore sediments of theFarasan Islands. We sought this information in order toenhance the use of foraminifera as environmental proxies inregional paleoenvironment and sea-level reconstructions.Although the reliability of the proxies can be compromisedby taphonomic processes, many studies, particularly thoseon the Holocene, have suggested that the basic compositionand structure of fossil benthic foraminiferal assemblages,are retained (e.g., (Murray, 1986; Hippensteel and others,2002; Buzas-Stephens and Buzas, 2005; Vance and others,2006; Wilson and Ramsook, 2007).
STUDY AREA
The Farasan Islands are located in the southern Red Seaplatform of Saudi Arabia between 16.5–17.2uN and 41.5–42.3uE (Fig. 1). They are located approximately 40 kmoffshore Jizan City and 50 km away from the Red Sea axialtrough. Within the tropical sector of the Red Sea, theFarasan Islands are the largest coral island group (c. 128islands totalling 3000 km2 of land). Its largest islands areFarasan Al-Kabir and Sajid (Bantan, 1999).
The Farasan Islands have low topography (,15 m a.s.l)with maximum elevation of 75 m a.s.l. They are composedof reefal limestone that formed during the early to latePleistocene sea-level rise (Dabbagh and others, 1984;Bantan, 1999). Bantan (1999) reported that the limestoneis older toward the centers of the larger islands, suggestingthat the Farasan Islands emerged by progressive uplift dueto salt diapirism. Ages determined from strontium andcarbon isotopes suggest that this uplift is still occurring(Bantan, 1999).
The Farasan Islands are within a monsoon belt where theprevailing winds are northwesterly in summer and south-easterly in winter (Morcos, 1970; Bantan, 1999). Thisaffects surface-water currents around the Farasan Islands,which flow southward in summer and northward in winter.Air temperatures at the Farasan Islands range 19–32uC inwinter and 26.5–43uC in summer. Average annual rainfall isabout 1 cm (El-Beheiry, 2009). Due to the hot, arid climate,surface-water salinity averages ,38 around the islands3 Correspondence author. E-mail: [email protected]
1 Marine Geology Department, Faculty of Marine Science, KingAbdulaziz University, P.O. Box 80207, Jeddah 21589, Saudi Arabia
2 Geology Department, Faculty of Science, Mansoura University,El-Mansoura: 35516, Egypt
Journal of Foraminiferal Research, v. 41, no. 4, p. 349–362, October 2011
349

(Bantan, 1999) and 41 in the lagoons and bays (this study).Tidal cycles around the islands are ,11h with a mean rangeof 0.5 m (Robinson, 1979; Abdelrahman, 1997).
MATERIALS AND METHODS
FIELDWORK
Forty-one sediment samples were collected in June 2008from the upper 2 cm of substrate in nearshore areas ofKhor As Sailah (KS), Jinabah Bay (JB), Ras Sheidah-RasAbbrah (RS-RA), and Ras Farasan (RF; Farasan Islands).The KS and JB sites represent low-energy environments forcomparison with the high-energy environments of RS-RAand RF. Sediment samples were collected in transectsperpendicular to the shoreline. Samples were collected froma variety of substrates with a spatula from the shoreline to,2 m water depth, utilizing SCUBA where necessary.Table 1 lists the basic sample data, such as location(coordinates determined by Garmin II GPS) and substratetype. Collected sediments were stored in sealed plastic bagsand refridgerated until processed in the laboratory. Todetermine salinity, seawater samples from ten sites werecollected in polyethylene bottles.
GRAIN-SIZE AND BENTHIC FORAMINIFERAL ANALYSES
Sediment samples were oven-dried at 50uC for 24 hours,then weighed. They were then washed over stacked sieves(2-, 0.150-, and 0.063-mm openings) to remove mud. Theresidue in each sieve was dried and weighed to determine itspercentage of the total dry weight. Using a microsplitter,foraminifera were picked from portions of each 0.150–2 mmfraction until at least 200 individuals were obtained.Specimens were sorted, identified, and counted (Appendices1 and 2). Species identifications were based primarily on
Loeblich and Tappan (1987), Haig (1988), Cimerman andLanger (1991), Sgarrella and Moncharmont-Zei (1993),Hottinger and others (1993), Jones (1994), and Yassini andJones (1995). The census analysis was based on all testsretaining a proloculus, which were assumed to be notreworked, in order to maximize the integrity of the data(Bernstein and others, 1978; Gooday and others, 1998).Appendix 1 is a taxonomic reference list of the foraminif-eral species found in this study. Density and relativeabundance were calculated for the key species, most ofwhich were imaged using the scanning electron microscope(SEM) at the Faculty of Science, King Abdulaziz Univer-sity, and displayed in Figures 7–9.
CHEMICAL ANALYSES
Salinity
The ten seawater samples were sent to the laboratory atthe Faculty of Marine Science (KAU), where salinity wasmeasured by titration.
Organic Carbon
Organic carbon in the sediment samples was determinedby the Loss On Ignition (LOI) technique. Approximatelyone gram of each sediment sample was placed in a cleaned,weighed ceramic crucible, and oven-dried at 105uC. Driedsamples were left to cool in a desiccator in order to preventabsorption of atmospheric moisture. They were thenweighed and transferred to a muffle furnace at 550uC forfour hours to remove organic carbon (Heiri and others,2001; Shuman, 2003). After cooling in the desiccator, eachresidual ash sample was weighed. Then, the percentage ofLOI was calculated as LOI% 5 dry wt 2 ash wt/dry wt 3100.
FIGURE 1. Location map of Farasan Islands (modified after Bantan 1999). Sample sites are arranged in transects (Transects 1–7, and 9).
350 ABU-ZIED AND OTHERS

Calcium Carbonate
Calcium carbonate (CaCO3) content was determined bytreating each weighed sediment sample (,0.5 g dry wt) with1 mol HCl for 2 hours at 60uC to dissolve CaCO3. Thesample was filtered to remove the solution and the residuewas oven-dried at 50uC and weighed. Percent CaCO3 wascalculated as CaCO3% 5 dry sample wt 2 dry residue wt/dry sample wt 3 100.
FAUNAL DIVERSITY AND MULTIVARIATE
STATISTICAL ANALYSES
Diversity indices and cluster analyses were performedusing PRIMER v. 4.0 (Clarke and Warwick, 1994). Simplediversity (S) is the total number of species in an assemblage.Following Buzas and Gibson (1969), the Shannon-Wienerdiversity index (H) was calculated as H 5 2 sum (Pi * logPi), whereas Pi is the proportion of each species. Clusteranalysis of square-root transformed, standardized benthicforaminiferal data grouped samples according to their
faunal similarity, using Euclidean distance as a similaritymeasure and complete linkage method.Canonical correspondence analysis, using Past v. 1.98
(Hammer and others, 2001), was applied to show relation-ships between key benthic foraminiferal species andenvironmental variables such as water depth, substratetexture, LOI, and CaCO3.
RESULTS
SUBSTRATE AND SALINITY
The results of the grain-size analyses, plotted on theternary diagram of Figure 2, show that most of the sampledsubstrates are sandy.
Khor As Sailah (KS)
The nearshore substrate of Khor As Sailah (KS) consistsof reefal limestone, most of which is covered by a thin layerof soft sediments vegetated by seagrasses, green filamentous
TABLE 1. Coordinates, water depths (m), type of substrates, CaCO3, LOI and salinity measurements of nearshore areas of Khor As Sailah, RasSheidah-Ras Abbrah, Jinabah Bay and Ras Farasan (Farasan Islands).
AreaSamplenumber Latitude (N) Longitude (E)
Water depth(m) Substrate
Calciumcarbonate (%) LOI (%)
Salinity(%)
Khor AsSailah
T1A 16u43933.850 42u4957.820 0 Muddy sand-sized sediments — — —T1B 16u43933.650 42u4955.570 0.3 Muddy sand-sized sediments 91.8 3.82 41.1T1C 16u43933.310 42u4953.110 1 Hard with seagrasses and algae 91.2 5.4 —T2A 16u4490.910 42u4955.970 0 Muddy sand-sized sediments 86.6 3.37 —T2B 16u4490.500 42u4953.000 1 Hard with seagrasses and algae 89.8 4.81 —T2C 16u43959.910 42u4950.500 1.5 Hard with seagrasses and algae 88.2 4.72 41.9T3A 16u44931.810 42u4943.610 0 Sand-sized sediments 78.42 8.14 —T3B 16u44931.370 42u4941.560 1 Hard with seagrasses and algae 87.8 7.15 —T3C 16u44930.710 42u4939.290 1.5 Hard with seagrasses and algae 92.4 3.54 —C1 16u4598.200 42u3948.160 0.2 Firm sand-sized, mangroves — — —C2 16u4592.280 42u3949.080 1 Firm sand-sized sediments — — 43.6
RasSheidah-RasAbbrah
T4A 16u36928.490 42u8932.020 0 Sand-sized sediments 88.4 5.62 —T4B 16u36927.920 42u8930.830 0.5 Sand-sized sediments 91 2.11 —T4C 16u36927.280 42u8929.190 0.6 Hard with green algae 97 2.47 —T4D 16u36926.630 42u8927.930 0.7 Hard with green algae 91.4 2.72 —T4E 16u36926.170 42u8926.770 1.1 Hard with green algae 86.8 2.97 38.7T4F 16u36925.710 42u8925.510 1.5 Muddy sand-sized sediments 90.4 —T4G 16u36925.350 42u8924.160 2 Seagrass and green algae 91.4 3.84 —T5A 16u36955.330 42u7958.840 0 Sand-sized sediments 88.4 3.2 —T5B 16u36954.770 42u7958.150 0.1 Muddy sand-sized sediments 88.2 2.44 —T5C 16u36953.940 42u7957.120 0.2 Hard, seagrasses and algae 79.4 4.5 —T5D 16u36953.390 42u7956.460 0.3 Hard with green algae 87.4 3.68 —T5E 16u36952.750 42u7955.620 0.6 Hard, seagrasses and algae 96.2 2.73 —T5F 16u36952.200 42u7955.060 1 Hard with green algae 96.16 3.12 —T5G 16u36951.740 42u7954.400 1.5 Hard with green algae 93.8 5.82 —T5H 16u36951.100 42u7953.650 2 Hard with green algae 95.6 2.74 39T6A 16u37928.630 42u6943.960 0 Sand-sized sediments 86.6 1.9 —T6B 16u37927.960 42u6944.370 0.7 Muddy sand-sized sediments 90.8 3.9 —T6C 16u37927.280 42u6944.860 1.8 Hard, seagrasses and algae 92.6 5.67 38.5
Jinabah Bay T7A 16u38955.720 42u6936.860 0 Sand-sized sediments 93.4 3.71 —T7B 16u38956.650 42u6935.460 0.2 Hard with filamentous algae 92.6 3.93 39T7C 16u38957.550 42u6934.060 0.5 Hard with thin mud layer 92 2.92 —T7D 16u38958.180 42u6932.830 0.7 Hard with thin mud layer 97.2 2.78 —T7E 16u38958.730 42u6931.990 0.9 Muddy sand-sized sediments 94.2 3.39 —T7F 16u38959.270 42u6931.050 1.6 Sand-sized sediments 89.4 3.43 —T7G 16u38959.680 42u6930.240 2 Sand-sized sediments 86.8 2.72 38.7
Ras Farasan T9A 16u52950.810 41u46948.580 0 Sand-sized sediments 94.6 1.78 —T9B 16u52951.870 41u46949.240 1 Muddy sand-sized sediments 89.6 5.04 39T9C 16u52952.920 41u46949.670 1.4 Muddy sand-sized sediments 92.2 3.69 —T9D 16u52954.370 41u46950.290 1.6 Sandy-sized substrate 94.8 3.75 —T9E 16u52955.600 41u46950.910 1.9 Muddy sand-sized substrate 91.4 2.5 —T9F 16u52956.470 41u46951.110 1.7 Sandy-sized substrate 91.8 3.35 38.5
FARASAN ISLANDS NEARSHORE FORAMINIFERA 351

algae, and calcareous green algae (Halimeda sp.). Thesedimentary veneer of biogenic muddy sand and brokenHalimeda thalli accounts for 30–50% of the total sedimentmass. Gravelly biogenic content increases shorewards, andsmall and large gastropod shells (e.g., Cerithidea cingulataand Melanoides tuberculata, respectively) characterize theintertidal zone. The LOI was 5% (s6 1.6) and the amount ofCaCO3 was 88% (s 6 4.0). Shoreline sediments are mainlyangular to subangular, blackish to white, calcareous sand, inpart derived from nearby coral reef terraces 3.5 m a.s.l. Someforaminifera in these sediments show evidence of dissolutionand bioerosion. Water salinity measured 42 (s 6 1.0).
Ras Sheidah-Ras Abbrah (RS-RA)
The nearshore area of Ras Sheidah-Ras Abbrah (RS-RA)has a hard substrate mostly covered by filamentous greenalgae. Calcareous green algae (Halimeda) and red algae(Jania) are occasionally frequent, and seagrass is sparse. TheRS-RA beach is characterized by white bioclastic sand,which forms the back-beach dunes that are often covered bysmall shrubs. The beach sands contain many Chiton, whichin life would have been attached to the reefal limestone. Thenearshore sediments are biogenic muddy sand andHalimedathalli constitute ,50% of the total sediment. LOI was 3.4%(s 6 1.0) and the amount of CaCO3 was 91% (s 6 4.0). Alsofrequent in these sediments are porous, black pumice shardsdisplaced ,120 km by northerly seawater flow from theJabal Al-Tair Volcano, which last erupted in September2007. Salinity was 38.7 (s 6 0.2).
Jinabah Bay (JB)
The nearshore area of Jinabah Bay (JB) has a narrowinner zone of reefal limestone covered with a thin layer ofmuddy sand retained by filamentous green algae. Beyondthis zone the muddy sand substrate often includes Jania and
broken Halimeda thalli. Small shells of gastropods andbivalves become frequent shoreward. The LOI was 3.3% (s6 0.4) and the amount of CaCO3 was 92% (s 6 3.0). Watersalinity was 38.85 (s 6 0.2).
Ras Farasan (RF)
The nearshore substrate of Ras Farasan (RF) is mainlybiogenic sand with rare seagrasses and green filamentousalgae. Salinity was 39. The beach is white, bioclastic sand thatoften bears aggregates, peloids, and broken Halimeda thalli.LOIwas 3.4% (s6 1.0) and the amount of CaCO3was 92% (s6 1.8). Sediments of both beach and substratum containedsome foraminiferal tests that were abraded or broken.
BENTHIC FORAMINIFERA
Fifty-three benthic foraminiferal species were recordedfromthenearshore substrates sampled in this study.Appendix1is the taxonomic reference list andAppendix 2 presents theircensus. Key species are discussed below and Figures 3 and 4display their density, diversity, and abundance.
Faunal Density and Diversity
Benthic foraminiferal density ranged from 117 specimens/g (s 6 107) at Jinabah Bay and Ras Farasan to 1900specimens/g (s 6 1287) at Khor As Sailah. The nearshoresediments of Ras Sheidah-Ras Abbrah yielded an interme-diate value of 450 specimens/g (s 6 560) (Fig. 3).
Species diversities S and H co-varied among the studiedsample sites, with mean values of S 5 19 (s 6 7) and H 5 2(s 6 0.5; Fig. 3). They increased seaward to highest values(S 5 35. H 5 3) in the deeper sites of KS, RA-RS (transectT4), and the RF transects, and were lowest in transects T5and T6 of RA-RS. In JB, diversity peaked in the middle oftransect T7 (Fig. 3).
General Characteristics of the BenthicForaminiferal Assemblages
Dominant species in the nearshore sediments of Khor AsSailah (KS) were Peneroplis planatus (25%), Soritesorbiculus (14%), Neorotalia calcar (14%), Coscinospirahemprichii (11%), and Varidentella neostriatula (5%). Alsocommon species were Clavulina angularis, Quinqueloculinalamarckiana, Q. poeyana, Q. limbata, and Parrina bradyi. Inthe KS transects, relative abundances of S. orbiculus, C.hemprichii, P. planatus, and C. angularis increased shore-ward, while Neorotalia calcar and Varidentella neostriatulatended to increase seaward. All of these species werecharacteristic of KS, except for Neorotalia calcar and Q.lamarckiana, which occurred in high percentages in the RasSheidah-Ras Abbrah (RS-RA) and Ras Farasan (RF)assemblages. Smaller miliolids account for ,20% of thecomposite KS foraminiferal assemblage.
Neorotalia calcar was dominant at RS-RA, accountingfor ,50% of the composite foraminiferal assemblage.Many species occurred at ,5%, such as P. planatus, S.orbiculus, Q. lamarckiana, Q. seminula, and Trochulinadimidiata. In less abundance were Murrayinella murrayi,Massilina gualtieriana, Glabratella patelliformis, Neoepo-
FIGURE 2. Ternary diagram of grain-size variations in thenearshore sediments of Khor As Sailah (KS), Jinabah Bay (JB), RasSheidah-Ras Abbrah (RS-RA), and Ras Farasan (RF) areas ofFarasan Islands.
352 ABU-ZIED AND OTHERS

nides bradyi, and Discorbinella araucana. All of these specieswere characteristic of RS-RA. In transects T4 and T6, N.calcar increased shoreward, but in transect T5 it increasedseaward. Smaller miliolids represented ,25% of thecomposite RS-RA foraminiferal assemblage.
The nearshore foraminiferal assemblages of Jinabah Bay(JB) were dominated by Ammonia convexa (25%), P.planatus (11%), S. orbiculus (10%), Elphidium striatopunc-tatum (7%), Q. poeyana (7%), and Triloculina fichteliana(5%). Sorites orbiculus generally increased shorewardsediments, while Ammonia convexa, E. striatopunctatum,and T. fichteliana increased seaward. Ammonia convexa, E.striatopunctatum, and T. fichteliana were in their highestabundances in JB, but they were among the dominantspecies elsewhere. Smaller miliolids represent,30% of totalforaminiferal assemblages in the nearshore sediments of JB.
The nearshore assemblages of Ras Farasan (RF) weredominated by Neorotalia calcar (20%), A. convexa (10%), Q.lamarckiana (8%), and E. striatopunctatum (6%). Manyothers species were present in low abundances, such as C.hemprichii, Varidentella neostriatula, Q. limbata, and Elphi-dium carticulatum. Agglutinated forms also occurred in lowabundance (,2%). Neorotalia calcar, Q. lamarckiana, E.striatopunctatum, and agglutinated forms increased sea-wards. Smaller miliolids represented ,35% of total forami-niferal assemblages in the nearshore sediments of RF.
Statistical Analyses
Cluster analysis of the benthic foraminiferal data split thesediment samples/assemblages into bay-lagoonal and openseawater groups (Fig. 5). The bay-lagoon group representsKS and JB, whereas the open-sea group represents RS-RAand RF.
Canonical correspondence analysis showed good positivecorrelation between muddy substrates and S. orbiculus, C.hemprichii, P. planatus, C. angularis, Q. poeyana, and P.bradyi. In contrast, these species had negative correlationwith CaCO3, and showed no relation with sandy or gravellysubstrates. Species such as A. convexa, E. carticulatum, E.striatopunctatum, M. murrayi, V. neostriatula, C. carinatas-triata and D. araucana had positive correlation with sandysubstrates and greater water depth, and N. calcar, Q.limbata, N. bradyi, and T. dimidiata had weak positivecorrelation with gravelly substrates and CaCO3 (Fig. 6).
DISCUSSION
FAUNAL COMPOSITION AND DISTRIBUTION
Cluster analysis divided the nearshore faunas Khor AsSailah (KS), Jinabah Bay (JB), Ras Farasan (RF), and RasSheidah-Ras Abbrah (RS-RA) areas into two majorgroups: the bay-lagoonal group (KS and JB) and theopen-water group (RS-RA and RF). Clusters indicate thatthe benthic foraminiferal assemblages in each study areahave specific characters related to their local environmentsdespite any taphonomic modification of their composition(Hippensteel and others, 2002; Vance and others, 2006).This validates the applicability of the data in localpaleoenvironmental reconstruction, particularly in relationto relative changes in sea level.
Khor As Sailah (KS)
The nearshore KS fauna, characterized symbiont-bearinglarger foraminifera and smaller miliolids and agglutinants,is similar to those that inhabit warm hypersaline lagoons at
FIGURE 3. Benthic foraminiferal density, diversity and water depths (m) of the studied sediment sample sites in Khor As Sailah, Ras Sheidah-RasAbbrah, Jinabah Bay and Ras Farasan areas (Farasan Islands).
FARASAN ISLANDS NEARSHORE FORAMINIFERA 353
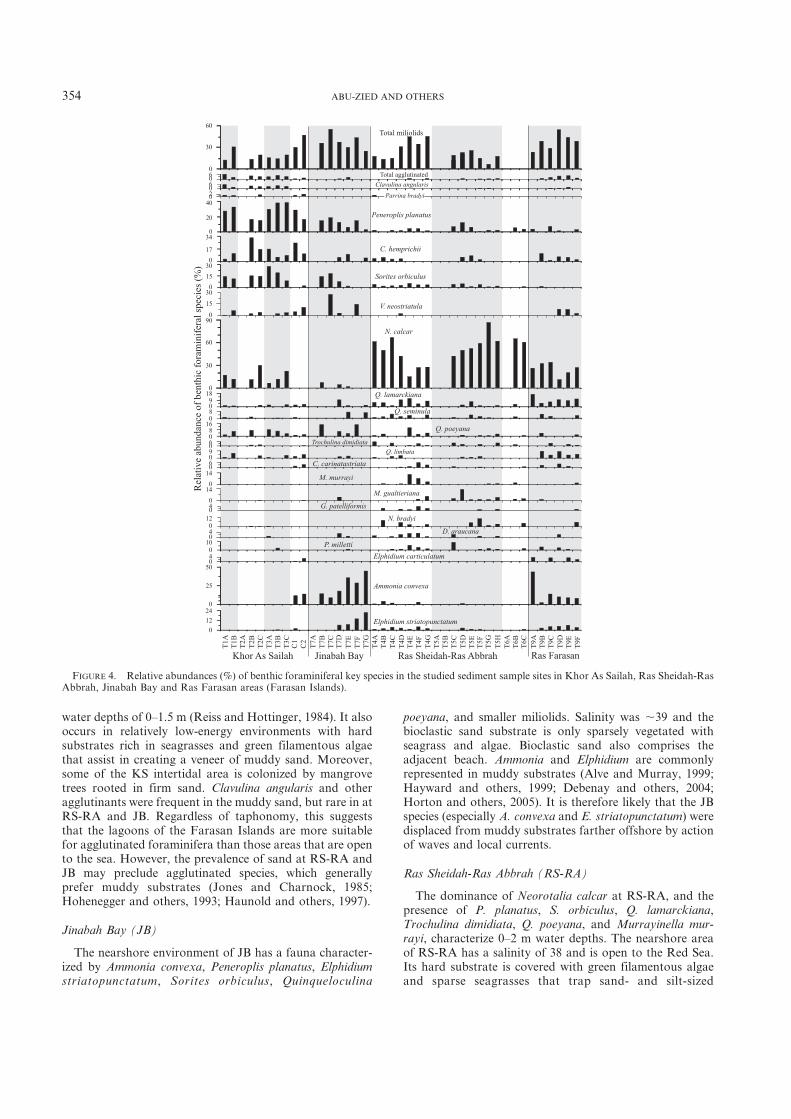
water depths of 0–1.5 m (Reiss and Hottinger, 1984). It alsooccurs in relatively low-energy environments with hardsubstrates rich in seagrasses and green filamentous algaethat assist in creating a veneer of muddy sand. Moreover,some of the KS intertidal area is colonized by mangrovetrees rooted in firm sand. Clavulina angularis and otheragglutinants were frequent in the muddy sand, but rare in atRS-RA and JB. Regardless of taphonomy, this suggeststhat the lagoons of the Farasan Islands are more suitablefor agglutinated foraminifera than those areas that are opento the sea. However, the prevalence of sand at RS-RA andJB may preclude agglutinated species, which generallyprefer muddy substrates (Jones and Charnock, 1985;Hohenegger and others, 1993; Haunold and others, 1997).
Jinabah Bay (JB)
The nearshore environment of JB has a fauna character-ized by Ammonia convexa, Peneroplis planatus, Elphidiumstriatopunctatum, Sorites orbiculus, Quinqueloculina
poeyana, and smaller miliolids. Salinity was ,39 and thebioclastic sand substrate is only sparsely vegetated withseagrass and algae. Bioclastic sand also comprises theadjacent beach. Ammonia and Elphidium are commonlyrepresented in muddy substrates (Alve and Murray, 1999;Hayward and others, 1999; Debenay and others, 2004;Horton and others, 2005). It is therefore likely that the JBspecies (especially A. convexa and E. striatopunctatum) weredisplaced from muddy substrates farther offshore by actionof waves and local currents.
Ras Sheidah-Ras Abbrah (RS-RA)
The dominance of Neorotalia calcar at RS-RA, and thepresence of P. planatus, S. orbiculus, Q. lamarckiana,Trochulina dimidiata, Q. poeyana, and Murrayinella mur-rayi, characterize 0–2 m water depths. The nearshore areaof RS-RA has a salinity of 38 and is open to the Red Sea.Its hard substrate is covered with green filamentous algaeand sparse seagrasses that trap sand- and silt-sized
FIGURE 4. Relative abundances (%) of benthic foraminiferal key species in the studied sediment sample sites in Khor As Sailah, Ras Sheidah-RasAbbrah, Jinabah Bay and Ras Farasan areas (Farasan Islands).
354 ABU-ZIED AND OTHERS

bioclasts. It is conceivable that the spines on N. calcarprovide a means of stabilization that give it an advantageon the algal mat (Hohenegger, 1994; Lobegeier, 2002). Thefilamentous algae at RS-RA allow also the proliferation ofthe epiphytic rosaline Trochulina dimidiata. Most rosalinesare epiphytic, commonly found attached to seagrass orphytal remnants (DeLaca and Lipps, 1972; Dobson andHaynes, 1973; Langer, 1993; Abu-Zied and others, 2007).
Ras Farasan (RF)
The nearshore fauna of RF, characterized by N. calcar,A. convexa, Varidentella neostriatula, Elphidium carticula-tum, E. striatopunctatum, Q. lamarckiana, Q. limbata, C.
hemprichii, and smaller miliolids, is from 0–2 m waterdepths with a salinity of 38.7. The substrata in this areavary from bioclastic muddy sand to sand, in some placeswith isolated seagrass and green filamentous algae. Theseenvironmental conditions and the associated benthicforaminiferal species have some commonality with JB andRS-RA but not with KS.
TAPHONOMY
Although benthic foraminiferal density was much higherat KS than the other areas, faunal diversities were relativelysimilar and often showed a tendency to slightly increaseseaward. The density difference may have resulted frommore favorable conditions in the KS lagoon that promoteda higher reproduction rate, but field observations andcomparison of faunal compositions among the four areasindicates that other factors, such as taphonomy, hadmodified the recovered assemblages. The low-energy ofthe KS environment implies a low sedimentation rate andminimal test destruction that would promote a higherdensity of foraminifera. On the other hand, the energeticenvironments of RS-RA, JB, and RF may destabilizeforaminiferal habitats, winnow tests, or destroy weakertests (Smith, 1987; Berkeley and others, 2007). In addition,these areas could have higher sedimentation rates due toonshore transport by waves and currents.
Many of the foraminiferal tests from nearshore sub-strates of the Farasan Islands show evidence of taphonomicprocesses such as abrasion, breakage, dolomitization,bioerosion, dissolution, and displacement. Abrasion, break-age, and dolomitization were more common at RS-RA andJB, and to a lesser extent at RF, which differ from the KSlagoon by being open to the sea and subject to its waves andcurrents. Abrasion is evident in tests of Peneroplis planatus(Fig. 8.9), Sorites orbiculus (Fig. 8.11), Coscinospira hem-prichii, Neorotalia calcar (Fig. 9.12), and Ammonia convexa(Fig. 9.14) by rounded edges and eroded ornamentation.Erosion of the outer wall of some tests is attributed toprolonged abrasion. Dolomitized specimens, which do notbreak easily with a needle, indicate a long residence timeamong sediment in transport. Abraded foraminiferal testswere highest in the intertidal sediments and decreased withincreasing water depths. Moreover, broken tests werecommon in all samples except those from KS.
At KS and JB, Sorites orbiculus was very abundant in theintertidal assemblages, but decreased seaward in thesubtidal sediments, indicating post-mortem onshore trans-port. Species of this epiphytic genus (e.g., Reiss andHottinger, 1984; Langer, 1993; Hohenegger, 1994; Haunoldand others, 1997) likely detach when blades of seagrassesshed, upon which they are transported by waves andcurrents (Wilson and Ramsook, 2007). Their flat discoidaltests are highly buoyant so they are prone to extendedtransportation in suspension (Hippensteel and others, 2002;Hohenegger, 2004). In Safaga Bay, Sorites was also foundin large numbers in sandy mounds near a seagrass meadow(Haunold and others, 1997). Although species such asPeneroplis planatus and Coscinospira hemprichii wereabundant in the nearshore sediments of KS and JB andalso have large, flat tests, they were not as abundant as S.
FIGURE 5. Cluster analysis dendrogram of the studied sedimentsamples, based on a square-root transformed, standardized benthicforaminiferal data, using Euclidean distance and completelinkage method.
FARASAN ISLANDS NEARSHORE FORAMINIFERA 355

orbiculus in the intertidal sediments. This emphasizes thelink between Sorites and seagrass (Reiss and Hottinger,1984; Langer, 1993; Hohenegger, 1994). Peneroplis planatusand C. hemprichii also live on seagrass as well assedimentary substrates (Langer, 1993; Haunold and others,1997), but they attach weakly to the blades. These findingscould lead to using fossil Sorites orbiculus as a tracer forshoreline development. It has already been suggested thatSorites tests indicate allochthonous shoreline sediments(Hallock and Larsen, 1979; Vance and others, 2006),whereas those of P. planatus and C. hemprichii indicate(par)autochthonous sediments (Haunold and others, 1997).
Sorites orbiculus occurs in low abundance in nearshoresediments of the RF and RS-RA, which have freeconnections with open-marine waters. This is probablyrelated to the lack of seagrass (the primary substrate of S.orbiculus [Reiss and Hottinger, 1984]) in these areas withhard substrates covered with green filamentous algae anddominated by Neorotalia calcar. It is also possible thatjuvenile Sorites cannot attach to seagrass in these high-energy environments.
Agglutinated species (e.g., Clavulina angularis) arerelatively abundant at KS, but absent at RS-RA and JB.This could be related to the low-energy conditions of KSenhancing its preservation potential. In contrast, aggluti-nants in unprotected nearshore areas are subject toabrasion, breakage, and transportation, which disintegratetheir tests immediately after their death (Murray, 2000). AtKS, where mangroves grow in the carbonate-rich sand,agglutinants are absent, probably due to the high oxidationpotential of the substrate. Berkeley and others (2007)reported that muddy substrates with mangroves favoragglutinants because the mangrove roots create a calmenvironment by effectively dissipating waves and currents.
Dissolution was frequently observed on tests of P.planatus (Fig. 8.8), S. orbiculus (Fig. 8.12), and N. calcar(Figs. 9.10 and 9.11), especially at KS despite its carbonate-saturated reefal environment. These tests showed signs ofetching, possibly related to the high LOI of the sediment.As organic carbon disintegrates in early diagenesis, itsrelease of CO2 lowers the pH of the water, which results inthe dissolution of CaC03.
Bioerosion was observed as microborings into the tests ofspecies such as Quinqueloculina bosciana (Fig. 7.8), C.hemprichii (Fig. 8.3), P. planatus (Fig. 8.10), S. orbiculusand N. calcar (Fig. 9.11). This was most prevalent at KS.Microborings in reefal foraminiferal have been attributed toendolithic microorganisms and fungi (Lobegeier, 2002) thatproliferate where the eater is calm (Cottey and Hallock,1988; Martin and Liddell, 1991; Lobegeier, 2002). Endolithsmay feed on symbiotic algae as well as foraminiferalprotoplasm, so some of the borings could be post-mortem.Microborings were not observed in the foraminiferal testsfrom RF, RS-RA, and JB, possibly because their higherenergy waters preclude endoparasites from attaching(Kloos, 1982). At RF, RS-RA, and JB, tests are onlyimpacted by abrasion, which polishes N. calcar specimensand erodes their spines.
CONCLUSIONS
Foraminiferal assemblages differed between the fournearshore localities (Khor As Sailah, Ras Sheidah-RasAbbrah, Jinabah Bay, and Ras Farasan) sampled in theFarasan Islands. The uniqueness of each assemblage relatesto its local physico-chemical environment. Symbiont-bearing larger foraminifera (Peneroplis planatus, Soritesorbiculus, Neorotalia calcar, and Coscinospira hemprichii)
FIGURE 6. Canonical correspondence analysis of benthic foraminiferal key species and their environmental variables (e.g., water depth, mud-sized, sand-sized, gravel-sized, LOI and CaCO3). The environmental variables are indicated by arrows, whereas the key species are indicated by solidcircle. Shaded areas indicate the locality of each area.
356 ABU-ZIED AND OTHERS
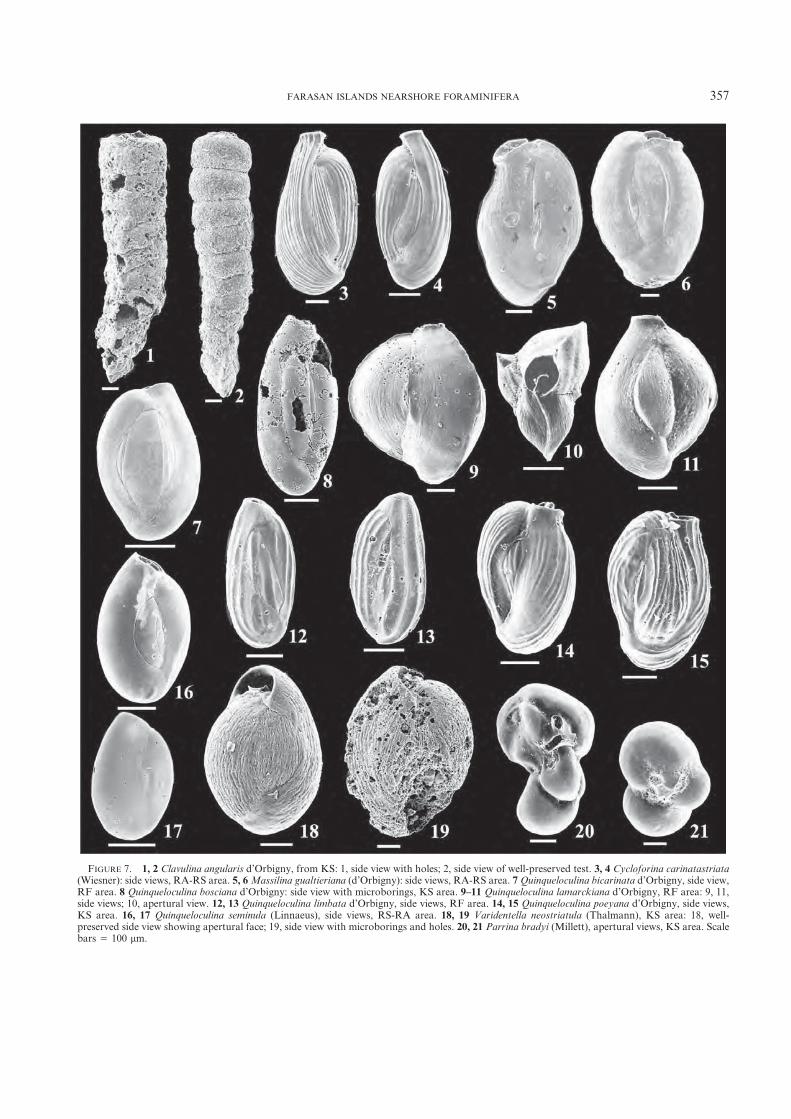
FIGURE 7. 1, 2 Clavulina angularis d’Orbigny, from KS: 1, side view with holes; 2, side view of well-preserved test. 3, 4 Cycloforina carinatastriata(Wiesner): side views, RA-RS area. 5, 6Massilina gualtieriana (d’Orbigny): side views, RA-RS area. 7 Quinqueloculina bicarinata d’Orbigny, side view,RF area. 8 Quinqueloculina bosciana d’Orbigny: side view with microborings, KS area. 9–11 Quinqueloculina lamarckiana d’Orbigny, RF area: 9, 11,side views; 10, apertural view. 12, 13 Quinqueloculina limbata d’Orbigny, side views, RF area. 14, 15 Quinqueloculina poeyana d’Orbigny, side views,KS area. 16, 17 Quinqueloculina seminula (Linnaeus), side views, RS-RA area. 18, 19 Varidentella neostriatula (Thalmann), KS area: 18, well-preserved side view showing apertural face; 19, side view with microborings and holes. 20, 21 Parrina bradyi (Millett), apertural views, KS area. Scalebars 5 100 mm.
FARASAN ISLANDS NEARSHORE FORAMINIFERA 357

dominated the low-energy, hypersaline lagoon of Khor AsSailah that is rich in seagrass and green filamentous algae.In contrast, Neorotalia calcar dominated the high-energynearshore of Ras Sheidah-Ras Abbrah, where greenfilamentous algae cover the hard substrate. The assemblag-
es from the moderate-energy environments of both JinabahBay and Ras Farasan were characterized by Ammoniaconvexa, Peneroplis planatus, Sorites orbiculus, Neorotaliacalcar, Varidentella neostriatula, Quinqueloculina sp., andElphidium sp.
FIGURE 8. 1–4 Coscinospira hemprichii Ehrenberg, KS area: 1, side view of an adult specimen; 2, side view of a juvenile specimen; 3, side view withmicroborings; 4, outer surface at high magnification. 5–10 Peneroplis planatus (Fichtel and Moll), KS area: 5, side view of an adult specimen; 6, outersurface with high magnification; 7, side view of a juvenile specimen; 8, side view of a specimen with rough surface; 9, side view of a broken andabraded specimen with microborings; 10, side view with microborings and holes. 11–14 Sorites orbiculus (Forskal), KS area: 11, side view of abradedspecimen; 12, side view of specimen with rough surface; 13, peripheral view showing apertures; 14, side view of well-preserved specimen. 15–17Neoeponides bradyi (Le Calvez), RA-RS area: 15, umbilical view; 16, peripheral view; 17, spiral view. 18, 19 Trochulina dimidiata (Jones and Parker),RA-RS area: 18, spiral view; 19, umbilical view. 20, 21 Rosalina bradyi (Cushman), KS area: 20, umbilical view; 21, spiral view. Scale bars 5 100 mm.
358 ABU-ZIED AND OTHERS

In the calm waters of Khor As Sailah lagoon, the releaseof CO2 from high amounts of decomposing organic carbonin the soft substratum promotes calcareous test dissolution.This environment is conducive to endolithic microorgan-isms that bore into foraminiferal tests, which promotes theirdestruction and removal from the sediments.
Sorites orbiculus was found living on seagrass blades andits dead were in great abundance in shoreline sediments,especially at Khor As Sailah. This indicates their shorewardtransport by waves and currents, which was probablyfacilitated by attachment to leaf fragments. This suggeststhat the species may be useful in defining ancient shorelines.
FIGURE 9. 1, 2 Glabratella patelliformis (Brady), RA-RS area: 1, umbilical view; 2, peripheral view. 3, 4 Murrayinella murrayi (Heron-Allen andEarland), RA- RS area: 3, spiral view; 4, umbilical view. 5–7 Discorbinella araucana (d’Orbigny), RA- RS area: 5, 7, umbilical views, RA-RS area; 6,spiral view. 8–12 Neorotalia calcar (d’Orbigny): 8, 9, umbilical and spiral views, RA-RS area; 10, 11, umbilical views of specimens with rough surfacesand microborings, KS area; 12, umbilical view of an abraded specimen, RF area. 13–15 Ammonia convexa (Collins), RF area: 13, umbilical viewshowing breakage of the last two chambers; 14, umbilical view of broken and abraded specimen; 15, spiral view. 16–18 Elphidium striatopunctatum(Fichtel and Moll), JB area: 16, peripheral view; 17, umbilical view. 19–20 Elphidium craticulatum (Fichtel and Moll), RF area: 18, umbilical view; 19,peripheral view; 20, umbilical view of broken and abraded specimen. 21, 22 Parellina milletti (Heron-Allen and Earland), RA-RS area: 21, umbilicalview; 22, peripheral view showing apertural face. Scale bars 5 100 mm.
FARASAN ISLANDS NEARSHORE FORAMINIFERA 359

Abrasion, breakage, and transportation are all importanttaphonomic processes in high-energy nearshore environ-ments. The open environments of Jinabah Bay and RasFarasan enable waves and currents to modify foraminiferalassemblages by winnowing smaller specimens and destroy-ing thin-walled tests. As a consequence, the faunal densitywas lowest in these areas.
ACKNOWLEDGMENTS
We would like to thank Mohamed Hariri (former Dean)for approving financial support for our field study. Abdel-Rahman M. A. Hassan, Sultan S. Mahfouz, Aliwy Q. Al-Anzi and many others are also appreciated for their kindhospitality and assistance during our fieldwork at JizanCity and Farasan Islands. Talha Abdulrahman analyzedsamples for their calcium carbonate content. We aregrateful to Johann Hohenegger and Roger Flower forreviewing an early draft of this paper, and Geraint W.Hughes and Robert W. Jones who formally reviewed thesubmitted version. Final editing of the manuscript byKenneth Finger is greatly appreciated.
REFERENCES
ABDELRAHMAN, S. M., 1997, Seasonal fluctuations of mean sea level atGizan, Red Sea: Journal of Coastal Research, v. 13, p. 1166–1172.
ABOU-OUF, M., and EL-SHATER, A., 1991, The relationship betweenthe environmental conditions of the Jeddah coast, Red Sea, andbenthic foraminifera: Journal of King Abdulaziz University,Marine Sciences, v. 2, p. 49–64.
———, RAO, N. V. N. D., and TAJ, R. J., 1988, Benthic foraminiferafrom littoral sediments of Al-Lith-Al Qunfidhah coast, southernRed Sea: Indian Journal of Marine Sciences, v. 17, p. 217–221.
ABU-ZIED, R. H., KEATINGS, K. W., and FLOWER, R. J., 2007,Environmental controls on foraminifera in Lake Qarun, Egypt:Journal of Foraminiferal Research, v. 37, p. 136–149.
ALVE, E., and MURRAY, J. W., 1999, Marginal marine environments ofthe Skagerrak and Kattegat: A baseline study of living (stained)benthic foraminiferal ecology: Palaeogeography, Palaeoclimatol-ogy, Palaeoecology, v. 146, p. 171–193.
———, and ———, 2001, Temporal variability in vertical distributionsof live (stained) intertidal foraminifera, southern England: Journalof Foraminiferal Research, v. 31, p. 12–24.
BAHAFZALLAH, A. A. K., 1979, Recent benthic foraminifera fromJiddah Bay, Red Sea (Saudi Arabia): Neues Jahrbuch furGeologie und Palaeontologie (Monatshefte und Abhandlungen),v. 7, p. 385–398.
BANTAN, R. A., 1999, Geology and sedimentary environments ofFarasan Bank (Saudi Arabia) southern Red Sea: a combinedremote sensing and field study: Unpublished Ph. D. Dissertation,University of London, United Kingdom, 296 p.
BERKELEY, A., PERRY, C. T., SMITHERS, S. G., HORTON, B. P., andTAYLOR, K. G., 2007, A review of the ecological and taphonomiccontrols on foraminiferal assemblage development in intertidalenvironments: Earth-Science Reviews, v. 83, p. 205–230.
———, ———, ———, and HORTON, B. P., 2008, The spatial andvertical distribution of living (stained) benthic foraminifera from atropical, intertidal environment, north Queensland, Australia:Marine Micropaleontology, v. 69, p. 240–261.
BERNSTEIN, B. B., HESSLER, R. R., SMITH, R., and JUMARS, P. A.,1978, Spatial dispersion of benthic foraminifera in the abyssalcentral North Atlantic: Limnology and Oceanography, v. 23,p. 401–416.
BOLTOVSKOY, E., and WRIGHT, R., 1976, Recent Foraminifera: W.Junk, The Hague, 515 p.
BUZAS, M. A., and GIBSON, T. G., 1969, Species diversity: benthonicforaminifera in western North Atlantic: Science, v. 163, p. 72–75.
BUZAS-STEPHENS, P., and BUZAS, M. A., 2005, Population dynamicsand dissolution of foraminifera in Nueces Bay, Texas: Journal ofForaminiferal Research, v. 35, p. 248–258.
CIMERMAN, F., and LANGER, M. R., 1991, Mediterranean Foraminif-era: Slovenian Academy of Science and Arts and Swiss Academyof Natural Sciences, Ljubljana, 118 p.
CLARKE, K. R., and WARWICK, R. M., 1994, Change in marinecommunity: an approach to statistical analysis and interpretation:Plymouth Marine Laboratory, Plymouth, 144 p.
COTTEY, T. L., and HALLOCK, P., 1988, Test surface degradation inArchaias angulatus: Journal of Foraminiferal Research, v. 18,p. 187–202.
DABBAGH, A., HOTZL, H., and SCHNNIER, H., 1984, Farasan Islands, inJado, A. R., and Zotl, J. G. (eds.), Quaternary Period in SaudiArabia, Volume 2: Springer-Verlag, Berlin?, p. 212–218.
DEBENAY, J.-P., and PAYRI, C. E., 2010, Epiphytic foraminiferalassemblages on macroalgae in reefal environments of NewCaledonia: Journal of Foraminiferal Research, v. 40, p. 36–60.
———, GUIRAL, D., and PARRA, M., 2004, Behaviour and taphonomicloss in foraminiferal assemblages of mangrove swamps of FrenchGuiana: Marine Geology, v. 208, p. 295–314.
———, BICCHI, E., GOUBERT, E., and DU CHATELET, E. A., 2006,Spatio-temporal distribution of benthic foraminifera in relation toestuarine dynamics (Vie estuary, Vendee, France): Estuarine,Coastal and Shelf Science, v. 67, p. 181–197.
DELACA, T. E., and LIPPS, J. H., 1972, The mechanism and adaptivesignificance of attachment and substrate pitting in the foraminif-eran Rosalina bradyi d’Orbigny: Journal of ForaminiferalResearch, v. 2, p. 68–72.
DOBSON, M., and HAYNES, J., 1973, Association of foraminifera withhydroides on the deep shelf: Micropaleontology, v. 19, p. 78–90.
GOODAY, A. J., BETT, B. J., SHIRES, R., and LAMBSHEAD, P. J. D.,1998, Deep-sea benthic foraminiferal species diversity in the NEAtlantic and NW Arabian Sea: a synthesis: Deep-Sea Research II,v. 45, p. 165–201.
HAIG, D. W., 1988, Miliolid foraminifera from inner neritic sand andmud facies of the Papuan Lagoon, New Guinea: Journal ofForaminiferal Research, v. 18, p. 203–236.
HALLOCK, P., and LARSEN, A. R., 1979, Coiling direction inAmphistegina: Marine Micropaleontology, v. 4, p. 33–44.
HAMMER, Ø., HARPER, D. A. T., and RYAN, P. D., 2001, Past:Paleontological Statistics Software Package for Education andData Analysis: Palaeontologia Electronica, v. 4, p. 1–9.
HAUNOLD, T. G., BAAL, C., and PILLER, W. E., 1997, Benthicforaminiferal associations in the northern Bay of Safaga, Red Sea,Egypt: Marine Micropaleontology, v. 29, p. 185–210.
HAYWARD, B. W., GRENFELL, H. R., REID, C. M., and HAYWARD, K.A., 1999, Recent New Zealand Shallow-water Foraminifera:Taxonomy, Ecologic Distribution, Biogeography, and Use inPaleoenvironmental Assessments: Institute of Geological andNuclear Sciences, Monograph, v. 21, p. 1–264.
HEIRI, O., LOTTER, A. F., and LEMCKE, G., 2001, Loss on ignition as amethod for estimating organic and carbonate content in sedi-ments: reproducibility and comparability of results: Journal ofPaleolimnology, v. 25, p. 101–110.
HIPPENSTEEL, S. P., MARTIN, R. E., NIKITINA, D., and PIZZUTO, J. E.,2002, Interannual variation of marsh foraminiferal assemblages(Bombay Hook National Wildlife Refuge, Smyrna, DE): Doforaminiferal assemblages have a memory?: Journal of Forami-niferal Research, v. 32, p. 97–109.
HOHENEGGER, J., 1994, Distribution of living larger foraminifera NWof Sesoko-Jima, Okinawa Japan: P.S.Z.N. I: Marine Ecology,v. 15, p. 291–334.
———, 2004, Depth coenoclines and environmental considerations ofwestern Pacific larger foraminifera: Journal of ForaminiferalResearch, v. 34, p. 9–33.
———, PILLER, W. E., and BAAL, C., 1993, Horizontal and verticalspatial microdistribution of foraminifers in the shallow subtidalGulf of Trieste, northern Adriatic Sea: Journal of ForaminiferalResearch, v. 23, p. 79–101.
HORTON, B. P., WHITTAKER, J. E., THOMSON, K. H., HARDBATTLE, M.I. J., WOODROFFE, S. A., and WRIGHT, M. R., 2005, Thedevelopment of a modern foraminiferal data set for sea-levelreconstructions, Wakatobi Marine National Park, southeast
360 ABU-ZIED AND OTHERS

Sulawesi, Indonesia: Journal of Foraminiferal Research, v. 35,p. 1–14.
HOTTINGER, L., HALICZ, E., and REISS, Z., 1993, Recent foraminifer-ida, Gulf of Aqaba, Red Sea: Opera Academia Scientiarum etArtium Slovenica, Classis IV: Historia Naturalis, v. 33; Paleon-tological Institute ‘‘Ivan Rakovec’’, v. 3, 179 p., 230 pl p..
JONES, R. W., 1994, The Challenger Foraminifera: Oxford UniversityPress, Oxford, 149 p., 117 pl p.
———, and CHARNOCK, M. A., 1985, Morphogroups of agglutinatingforaminifera. Their life positions and feeding habits and potentialapplicability in (paleo)ecological studies: Revue de Paleobiologie,v. 4, p. 311–320.
KLOOS, D. P., 1982, Destruction of tests of the foraminifer Soritesorbiculus by endolithic microorganisms in a lagoon on Curacao(Netherlands Antilles): Geologie en Mijnbouw, v. 61, p. 201–205.
LANGER, M. R., 1993, Epiphytic foraminifera in Langer, M. R. (ed.),Foraminiferal Microhabitats:: Marine Micropaleontology, v. 20,p. 235–265.
LOBEGEIER, M. K., 2002, Benthic foraminifera of the familyCalcarinidae from Green Island reef, Great Barrier Reef Province:Journal of Foraminiferal Research, v. 32, p. 201–216.
LOEBLICH, A. R., JR., and TAPPAN, H., 1987, Foraminiferal Generaand Their Classification, 2 v: van Nostrand Reinhold, NewYork,970 p.
MARTIN, R. E., and LIDDELL, W. D., 1991, The taphonomy offoraminifera in modern carbonate environments, in Donovan, S.K. (ed.), The Processes of Fossilization: Belhaven Press, London,p. 170–193.
MORCOS, S. A., 1970, Physical and chemical oceanography of the RedSea, in Barnes, M. (ed.), Oceanography and Marine Biology,vol. 8: George Allen and Unwin Ltd., London, p. 73–202.
MORVAN, J., DEBENAY, J.-P., JORISSEN, F., REDOIS, F., BENETEAU, E.,DELPLANCKE, M., and AMATO, A.-S., 2006, Patchiness and lifecycle of intertidal foraminifera: implication for environmental andpaleoenvironmental interpretation: Marine Micropaleontology,v. 61, p. 131–154.
MURRAY, J. W., 1986, Living and dead Holocene foraminifera of LimeBay, southern England; Journal of foraminiferal Research, v. 16,p. 347–352.
———, 1991, Ecology and Paleoecology of Benthic Foraminifera:Longman Scientific and Technical, Harlow, Essex, UK, 397 p.
———, 2000, The enigma of the continued use of total assemblages inecological studies of benthic foraminifera: Journal of Foraminif-eral Research, v. 30, p. 244–245.
———, 2006, Ecology and Applications of Benthic Foraminifera:Cambridge University Press, Cambridge, UK, 426 p.
REISS, Z., and HOTTINGER, L., 1984, The Gulf of Aqaba, EcologicalMicropaleontology: Ecological Studies, v. 50, Springer Verlag,Berlin, p. 1–354.
———, LEUTENEGGER, S., HOTTINGER, L., FERMONT, W. J. J.,MEULENKAMP, J. E., THOMAS, E., HANSEN, H. J., BUCHARDT,
B., LARSEN, A. R., and DROOGER, C. W., 1977, Depth-relations ofRecent larger foraminifera in the Gulf of Aqaba–Elat: UtrechtMicropaleontological Bulletin, v. 15, p. 1–244.
RIBES, T., SALVADO, H. J. R., and DEL PILAR GRACIA, A., 2000,Foraminiferal colonization on artificial seagrass leaves: Journal ofForaminiferal Research, v. 30, p. 192–201.
ROBINSON, M. K., 1979, Atlas of N. Atlantic Ocean–Indian Oceanmonthly mean temperatures and mean salinities of the surfacelayer: United States Naval Oceanographic Office, ReferencePublication 18.
SAID, R., 1949, Foraminifera of the Northern Red Sea: CushmanLaboratory for Foraminiferal Research, Special Publication,No. 26, p. l–44.
———, 1950, The distribution of foraminifera in the northern Red Sea:Contributions from the Cushman Foundation for ForaminiferalResearch, v. 1, p. 9–31.
SEN GUPTA, B. K., 1999, Introduction to modern foraminifera, in SenGupta, B. K. (ed.), Modern Foraminifera: Kluwer AcademicPublishers, Dordrecht, p. 3–6.
SGARRELLA, F., and MONCHARMONT-ZEI, M., 1993, Benthic forami-nifera of the Gulf of Naples (Italy): systematic and autoecology:Bollettino della Societa Paleontologica Italiana, v. 32, no. 2,p. 145–264.
SHUMAN, B., 2003, Controls on loss-on-ignition variation in cores fromtwo shallow lakes in the northeastern United States: Journal ofPaleolimnology, v. 30, p. 371–385.
SMITH, R. K., 1987, Fossilization potential in modern shallow-waterbenthic foraminiferal assemblages: Journal of ForaminiferalResearch, v. 17, p. 117–122.
STEINKER, D. C., and CLEM, K. V., 1984, Some near-shoreforaminiferal assemblages from phytal and bottom sediments,Bermuda: The Compass, v. 61, p. 98–115.
VANCE, D. J., CULVER, S. J., CORBETT, D. R., and BUZAS, M. A., 2006,Foraminifera in the Albemarle estuarine system, North Carolina:distribution and recent environmental changes: Journal ofForaminiferal Research, v. 36, p. 15–33.
WILSON, B., 1998, Epiphytal foraminiferal assemblages on the leaves ofthe seagrasses Thalassia testinudum and Syringodium filiforme:Caribbean Journal of Science, v. 34, p. 131–132.
———, and RAMSOOK, A., 2007, Population densities and diversities ofepiphytal foraminifera on nearshore substrates, Nevis, WestIndies: Journal of Foraminiferal Research, v. 37, p. 213–222.
YASSINI, I., and JONES, B. G., 1995, Recent Foraminifera andOstracoda from estuarine and shelf environments on thesoutheastern coast of Australia: The University of WollongongPress, Wollongong, Australia, 484 p.
Received 6 October 2010Accepted 22 April 2011
APPENDIX 1
Taxonomic reference list of benthic foraminifera recorded from nearshore substrates of the Farasan Islands. Listed in alphabetical order are 53species with their original designation and type reference. Bold parenthesised figure numbers refer to the illustrations of key species in this article.
Agglutinella robusta El-Nakhal 5 Agglutinella robusta El-Nakhal, 1983, p. 130, pl. 1, figs. 4–6, pl. 2, figs. 12–15.Ammonia convexa (Collins) 5 Streblus convexus Collins, 1958, p. 414, pl. 5, fig. 10a–c. (Figs. 9.13–9.15)Amphistegina lobifera Larsen 5 Amphistegina lobifera Larsen, 1976, p. 4, pl. 3, figs. 1–5, pl. 7, fig. 3, pl. 8, fig. 3.Clavulina angularis d’Orbigny 5 Clavulina angularis d’Orbigny, 1826, p. 268, pl. 12, pl.1, figs. 5, 6. (Figs. 7.1, 7.2)Coscinospira hemprichii Ehrenberg 5 Coscinospira hemprichii Ehrenberg, 1839, p. 131, pl. 2, fig. 2. (Figs. 8.1–8.4)Cycloforina carinatastriata (Wiesner) 5 Adelosina milletti var. carinata-striata Wiesner, 1923, p. 76, pl. 14, figs. 190, 191. (Figs. 7.3, 7.4)Cycloforina contorta (d’Orbigny) 5 Quinqueloculina contorta d’Orbigny, 1846, p. 298, pl. 20, figs. 4–6.Cycloforina juleana (d’Orbigny) 5 Quinqueloculina juleana d’Orbigny, 1846, p. 298, pl. 20, figs. 1–3.Cycloforina quinquecarinata (Collins) 5 Quinqueloculina quinquecarinata Collins, 1958, p. 360, pl. 2, fig. 8.Cycloforina stalkeri (Loeblich and Tappan) 5 Quinqueloculina stalkeri Loeblich and Tappan, 1953, p. 40, pl. 5, figs. 5–9.Cycloforina sulcata (d’Orbigny) 5 Quinqueloculina sulcata d’Orbigny, 1900, in Fornasini, 1900, p. 364, text–fig. 9.Cymbaloporella tabellaeformis (Brady) 5 Cymbalopora tabellaeformis Brady, 1884, p. 637, pl. 102, figs. 15–18.Discorbinella araucana (d’Orbigny) 5 Rosalina araucana d’Orbigny, 1839. (Figs. 9.5–9.7)Dyocibicides biserialis Cushman and Valentine 5 Dyocibicides biserialis Cushman and Valentine, 1930, p. 31, pl. 10, figs. 1, 2.Elphidium advenum (Cushman) 5 Polystomella advena Cushman, 1922, p. 56, pl. 9, figs. 11, 12.Elphidium craticulatum (Fichtel and Moll) 5 Nautilus craticulatum Fichtel and Moll, 1798, p. 51, pl. 5, figs. h–k. (Figs. 9.19, 9.20)
FARASAN ISLANDS NEARSHORE FORAMINIFERA 361

Elphidium striatopunctatum (Fichtel and Moll) 5 Nautilus striato-punctatus Fichtel and Moll, 1798, p. 61, pl. 9, figs. a–c. (Figs. 9.16–9.18)Glabratella patelliformis (Brady) 5 Discorbina patelliformis Brady, 1884, p. 647, pl. 88, fig. 3, pl. 89, fig. 1. (Figs. 9.1, 9.2)Glabratellina erecta (Sidebottom) 5 Discorbina erecta Sidebottom, 1908, p. 16, pl. 5, figs. 6, 7.Haynesina depressulus (Walker and Jacob) 5 Nautilus depressulus Walker and Jacob, 1798, p. 641, fig. 33.Lachlanella corrugata (Collins) 5 Massilina corrugata Collins, 1958, p. 362, pl. 2, figs. 11, 12.Lachlanella subpolygona (Parr) 5 Quinqueloculina subpolygona Parr, 1945, p. 196, pl. 12, fig. 2.Massilina gualtieriana (d’Orbigny) 5 Quinqueloculina gualtieriana d’Orbigny, 1839, p. 186, pl. 11, figs. 1–3. (Figs. 7.5, 7.6)Miliolinella dilatata (d’Orbigny) 5 Quinqueloculina dilatata d’Orbigny, 1839, p. 192, pl. 11, figs. 28–30.Murrayinella murrayi (Heron-Allen and Earland) 5 Rotalia murrayi Heron-Allen and Earland, 1915, p. 721. (Figs. 9.3, 9.4)Neoeponides bradyi (LeCalvez) 5 Eponides bradyi Le Calvez, 1974, p. 63. (Figs. 8.15–8.17)Neorotalia calcar (d’Orbigny) 5 Calcarina calcar d’Orbigny, 1839, p. 81, pl. 5, figs. 22–24. (Figs. 9.8–9.12)Parellina milletti (Heron-Allen and Earland) 5 Polystomella milletti Heron-Allen and Earland, 1915, p. 735, pl. 53, figs. 38–42. (Figs. 9.21, 9.22)Parrina bradyi (Millett) 5 Nubecularia bradyi Millett, 1898, p. 261, pl. 5, figs. 6a, b. (Figs. 7.20, 7.21)Peneroplis planatus (Fichtel and Moll) 5 Nautilus planatus Fichtel and Moll, 1798, p. 91, pl. 16, figs. a–f, i. (Figs. 8.5–8.10)Planorbulina mediterranensis d’Orbigny 5 Planorbulina mediterranensis d’Orbigny, 1826, p. 280, no. 2.Quinqueloculina bicarinata d’Orbigny 5 Quinqueloculina bicarinata d’Orbigny, 1878, p. 68, pl. 7, fig. 10. (Fig. 7.7)Quinqueloculina bosciana d’Orbigny 5 Quinqueloculina bosciana d’Orbigny, 1839, p. 191, pl. 11, figs. 22–24. (Fig. 7.8)Quinqueloculina lamarckiana d’Orbigny 5 Quinqueloculina lamarckiana d’Orbigny, 1839, p. 189, pl. 11, figs. 14, 15. (Figs. 7.9–7.11)Quinqueloculina limbata d’Orbigny 5 Quinqueloculina limbata d’Orbigny, 1826, p. 302, no. 20. (Figs. 7.12, 7.13)Quinqueloculina patagonica d’Orbigny 5 Quinqueloculina patagonica d’Orbigny, 1839, p. 74, pl. 4, figs. 14–16.Quinqueloculina poeyana d’Orbigny 5 Quinqueloculina poeyana d’Orbigny, 1839, p. 191, pl. 11, figs. 25–27. (Figs. 7.14–7.15)Quinqueloculina seminula (Linnaeus) 5 Serpula seminula Linnaeus, 1758, p. 786, pl. 2, figs. 1a–c. (Figs. 7.16, 7.17)Rosalina bradyi (Cushman) 5 Discorbis globularis (d’Orbigny) var. bradyi Cushman, 1915, p. 12, pl. 8, fig. 1. (Figs. 8.20, 8.21)Rupertianella rupertiana (Brady) 5 Miliolina rupertiana Brady, 1881, p. 46.Sorites orbiculus (Forskal) 5 Nautilus orbiculus Forskal, 1775, p. 125. (Figs. 8.11–8.14)Spirillina vivipara Ehrenberg 5 Spirillina vivipara Ehrenberg, 1841, p. 422, pl. 3, fig. 41.Spiroloculina antillarum d’Orbigny 5 Spiroloculina antillarum d’Orbigny, 1839, p. 166, pl. 9, figs. 3, 4.Spiroloculina communis Cushman and Todd 5 Spiroloculina communis Cushman and Todd, 1944, p. 63, pl. 9, figs. 4, 5, 7, 8.Spiroloculina corrugata Cushman and Todd 5 Spiroloculina corrugata Cushman and Todd, 1944, p. 51, 61, pl. 8, figs. 22–25.Spiroloculina tricarinata Le Calvez 5 Spiroloculina tricarinata Le Calvez, 1958, p. 207, pl. 8, figs. 84, 85.Textularia agglutinans d’Orbigny 5 Textularia agglutinans d’Orbigny, 1839, p. 144, pl. 1, figs. 17–18, 32–34.Triloculina fichteliana d’Orbigny 5 Triloculina fichteliana d’Orbigny, 1839, p. 171, pl. 9, figs. 8–10.Triloculina serrulata McCulloch 5 Triloculina serrulata McCulloch, 1977, p. 558, pl. 225, figs. 1, 2, 4.Triloculina trigonula (Lamarck) 5 Miliolites trigonula Lamarck, 1804, p. 351.Trochulina dimidiata (Jones and Parker) 5 Discorbis dimidiata Jones and Parker, 1862, in Carpenter and others, 1862, p. 201. (Figs. 8.18, 8.19)Varidentella neostriatula (Thalmann) 5 Quinqueloculina neostriatula Thalmann, 1950, p. 45. (Figs. 7.18, 7.19)Vertebralina striata d’Orbigny 5 Vertebralina striata d’Orbigny, 1826, p. 283.
APPENDIX 2
Census data of benthic foraminiferal species in nearshore surface bottom sediment samples of Khor As Sailah, Ras Sheidah-Ras Abbrah, JinabahBay and Ras Farasan (Farasan Islands). This table can be found on the Cushman Foundation website in the JFR Article Data Repository (http://www.cushmanfoundation.org/jfr/index.html) as item number JFR-DR20110010.
362 ABU-ZIED AND OTHERS
Copyright © 2022 FDOKUMEN




















