
A new Symbiodinium clade (Dinophyceae) from soritid foraminifera in Hawai’i
6
Short Communication A new Symbiodinium clade (Dinophyceae) from soritid foraminifera in Hawai’i Xavier Pochon * , Ruth D. Gates Hawai’i Institute of Marine Biology, University of Hawai’i, Ka ¯ ne’ohe, HI, USA article info Article history: Received 13 November 2009 Revised 28 March 2010 Accepted 31 March 2010 Available online 4 April 2010 Keywords: Ribosomal genes Dinoflagellate Symbiodinium Symbiosis Cryptic diversity abstract Dinoflagellates in the genus Symbiodinium are crucial components of coral reef ecosystems in their roles as endosymbionts of corals and other marine invertebrates. The genus Symbiodinium encompasses eight lineages (clades A–H), and multiple sub-clade types. Symbiodinium in clades A, B, C, and D are most com- monly associated with metazoan hosts while clades C, D, F, G, and H with large soritid foraminifera. Recent studies have described a diversity of new Symbiodinium types within each clades, but no new clades have been reported since 2001. Here, we describe a new clade of Symbiodinium isolated from sori- tid foraminifera from Hawai’i. Published by Elsevier Inc. 1. Introduction Dinoflagellates in the genus Symbiodinium are crucial compo- nents of coral reef ecosystems in their role as endosymbionts of cor- als, other marine invertebrates, and some protists (Trench, 1993). The genus Symbiodinium encompasses eight lineages, delineated phylogenetically using nuclear and chloroplast ribosomal DNA (rDNA), referred to as clades A through H (Pochon et al., 2004). Each clade is further divided into multiple genetic strains based on the nuclear internal transcribed spacer (ITS) regions (e.g., LaJeunesse, 2001; Rodriguez-Lanetty, 2003; van Oppen et al., 2005). Molecular clock inferences have estimated that clade A, the most ancestral Symbiodinium lineage, originated 65–50 Million years ago (MYA) and the other clades diverged from each other throughout the Eo- cene era, until the mid-Miocene (15 MYA) when a dramatic with- in-clade diversification occurred (Pochon et al., 2006; Tchernov et al., 2004). Symbiodinium in clades A, B, C, and D are commonly associated with metazoan hosts and representatives in clades C, D, F, G, and H with the large benthic foraminifera sub-family Soriti- nae (Pochon and Pawlowski, 2006). In recent years, researchers have focused on Symbiodinium with the goal of understanding their evolution, distribution and role in responding to present and future environmental conditions that are threatening the integrity of reefs worldwide (Baker et al., 2008; van Oppen et al., 2008). For example, recent studies conducted in both the Western Atlantic and Indo-Pacific Oceans have surveyed Symbiodinium across host taxa and a variety of spatial and temporal scales (reviewed in Coffroth and Santos, 2005; Stat et al., 2006). These studies have collectively described a diversity of new Symbiodinium types within each clade, but no new clades have been reported since 2001. Here, we describe a new clade of Symbiodinium isolated from soritid foraminifera from Hawai’i. 2. Materials and methods 2.1. Data collection As part of a more detailed study that will be described else- where, hundreds of soritid foraminiferan samples were collected between May 2007 and June 2008 from sites across the entire Hawaiian Archipelago. Genomic DNA was isolated from approxi- mately 100 of these forams using a guanidinium-based extraction protocol (Pochon et al., 2001) and screened for Symbiodinium sub- clade types by amplifying, cloning and sequencing the ITS-2 rDNA using the primers ‘ITS-DINO’ and ‘its2rev2’ (primer set 1) and con- ditions described in Stat et al. (2009). All recovered sequences were identified to sub-clade type or closest relative using the Basic Local Alignment Search Tool (BLAST) in GenBank. Four samples, from two sites on Oahu, Hawai’i (Fig. 1A), amplified ITS-2 Symbiodinium sequences that did not match any published Symbiodinium type closely. To further examine the diversity of Symbiodinium in these DNAs, the primers ‘ITS-DINO’ and ‘L_O’ (primer set 2) were used to amplify the nuclear partial 5.8S, the ITS-2, and the D1–D3 region of the 28S (Pochon et al., 2001) and the primers ‘23S4F’ and ‘23S7R’ (primer set 3) to amplify the chloroplast large subunit Domain V (cp23S; Pochon et al., 2006). 1055-7903/$ - see front matter Published by Elsevier Inc. doi:10.1016/j.ympev.2010.03.040 * Corresponding author. Address: HIMB, 46-007 Lilipuna Road, Ka ¯ ne’ohe, HI 96744, USA. Fax: +1 808 236 7443. E-mail addresses: [email protected] (X. Pochon), [email protected] (R.D. Gates). Molecular Phylogenetics and Evolution 56 (2010) 492–497 Contents lists available at ScienceDirect Molecular Phylogenetics and Evolution journal homepage: www.elsevier.com/locate/ympev
Transcript of A new Symbiodinium clade (Dinophyceae) from soritid foraminifera in Hawai’i

Molecular Phylogenetics and Evolution 56 (2010) 492–497
Contents lists available at ScienceDirect
Molecular Phylogenetics and Evolution
journal homepage: www.elsevier .com/locate /ympev
Short Communication
A new Symbiodinium clade (Dinophyceae) from soritid foraminifera in Hawai’i
Xavier Pochon *, Ruth D. GatesHawai’i Institute of Marine Biology, University of Hawai’i, Kane’ohe, HI, USA
a r t i c l e i n f o a b s t r a c t
Article history:Received 13 November 2009Revised 28 March 2010Accepted 31 March 2010Available online 4 April 2010
Keywords:Ribosomal genesDinoflagellateSymbiodiniumSymbiosisCryptic diversity
1055-7903/$ - see front matter Published by Elsevierdoi:10.1016/j.ympev.2010.03.040
* Corresponding author. Address: HIMB, 46-00796744, USA. Fax: +1 808 236 7443.
E-mail addresses: [email protected] (X. PochoGates).
Dinoflagellates in the genus Symbiodinium are crucial components of coral reef ecosystems in their rolesas endosymbionts of corals and other marine invertebrates. The genus Symbiodinium encompasses eightlineages (clades A–H), and multiple sub-clade types. Symbiodinium in clades A, B, C, and D are most com-monly associated with metazoan hosts while clades C, D, F, G, and H with large soritid foraminifera.Recent studies have described a diversity of new Symbiodinium types within each clades, but no newclades have been reported since 2001. Here, we describe a new clade of Symbiodinium isolated from sori-tid foraminifera from Hawai’i.
Published by Elsevier Inc.
1. Introduction
Dinoflagellates in the genus Symbiodinium are crucial compo-nents of coral reef ecosystems in their role as endosymbionts of cor-als, other marine invertebrates, and some protists (Trench, 1993).The genus Symbiodinium encompasses eight lineages, delineatedphylogenetically using nuclear and chloroplast ribosomal DNA(rDNA), referred to as clades A through H (Pochon et al., 2004). Eachclade is further divided into multiple genetic strains based on thenuclear internal transcribed spacer (ITS) regions (e.g., LaJeunesse,2001; Rodriguez-Lanetty, 2003; van Oppen et al., 2005). Molecularclock inferences have estimated that clade A, the most ancestralSymbiodinium lineage, originated 65–50 Million years ago (MYA)and the other clades diverged from each other throughout the Eo-cene era, until the mid-Miocene (�15 MYA) when a dramatic with-in-clade diversification occurred (Pochon et al., 2006; Tchernovet al., 2004). Symbiodinium in clades A, B, C, and D are commonlyassociated with metazoan hosts and representatives in clades C,D, F, G, and H with the large benthic foraminifera sub-family Soriti-nae (Pochon and Pawlowski, 2006).
In recent years, researchers have focused on Symbiodinium withthe goal of understanding their evolution, distribution and role inresponding to present and future environmental conditions thatare threatening the integrity of reefs worldwide (Baker et al.,2008; van Oppen et al., 2008). For example, recent studiesconducted in both the Western Atlantic and Indo-Pacific Oceans
Inc.
Lilipuna Road, Kane’ohe, HI
n), [email protected] (R.D.
have surveyed Symbiodinium across host taxa and a variety of spatialand temporal scales (reviewed in Coffroth and Santos, 2005; Statet al., 2006). These studies have collectively described a diversityof new Symbiodinium types within each clade, but no new cladeshave been reported since 2001. Here, we describe a new clade ofSymbiodinium isolated from soritid foraminifera from Hawai’i.
2. Materials and methods
2.1. Data collection
As part of a more detailed study that will be described else-where, hundreds of soritid foraminiferan samples were collectedbetween May 2007 and June 2008 from sites across the entireHawaiian Archipelago. Genomic DNA was isolated from approxi-mately 100 of these forams using a guanidinium-based extractionprotocol (Pochon et al., 2001) and screened for Symbiodinium sub-clade types by amplifying, cloning and sequencing the ITS-2 rDNAusing the primers ‘ITS-DINO’ and ‘its2rev2’ (primer set 1) and con-ditions described in Stat et al. (2009). All recovered sequences wereidentified to sub-clade type or closest relative using the Basic LocalAlignment Search Tool (BLAST) in GenBank. Four samples, fromtwo sites on Oahu, Hawai’i (Fig. 1A), amplified ITS-2 Symbiodiniumsequences that did not match any published Symbiodinium typeclosely. To further examine the diversity of Symbiodinium in theseDNAs, the primers ‘ITS-DINO’ and ‘L_O’ (primer set 2) were used toamplify the nuclear partial 5.8S, the ITS-2, and the D1–D3 region ofthe 28S (Pochon et al., 2001) and the primers ‘23S4F’ and ‘23S7R’(primer set 3) to amplify the chloroplast large subunit Domain V(cp23S; Pochon et al., 2006).

N
A
Kilometers0 5 10 CUU
UUA
CACCUUCGUGCGGGC
GCGA
UGUUU U C G U G
U C C UG C A C
UCCUGCAAGCCAUCGC
UCAG
AU U U
GC U
UCUGAUGGCUUGUUG A A U G A U U
G GCUGUU U
UGC A A
GC
UCAA
GCG C
U U U G U G A U U C A UA G
C AA A C
C UAU
GGGAUUC
GCUUGGGUCGC
GCUGC
UGAUGCCUA CAGCCUUCA G
CA
UGU
GAAA
CCGC
AUG
CAU
C5’ 3’
20
40
60
Helix I
Helix II
Helix IIIa
Helix IIIb
Helix IV
80
100
120
140160
1801 200
B
MahiWreck
LanaiLookout
OAHU
158°00
21°30
54°75151°851
PACIFIC OCEAN
51°1
254
°12
Fig. 1. (A) Map of the Island of Oahu, Hawai’i, showing two locations where soritid foraminiferans harboring new sequence types were collected. Samples OHU_3, OHU_7, andOHU_15 were collected at ‘Mahi Wreck’ [21�26.019N; 158�12.032W], and sample OHU_66 at ‘Lanai Lookout’ [21�16.316N; 157�41.039W]. (B) ITS-2 secondary structurefolding for the new sequence types. Twelve point mutations were recorded between sequence types, six located in stem loops or in single-stranded regions (green arrows),and six corresponding to hemi-CBC changes within a stem (red arrows). (For interpretation of the references to colour in this figure legend, the reader is referred to the webversion of this article.)
X. Pochon, R.D. Gates / Molecular Phylogenetics and Evolution 56 (2010) 492–497 493
Gene fragments were PCR-amplified using 0.5 U Hotstart Immo-lase Taq polymerase (Bioline) in 50 lL reactions, and following thethermocycling conditions described in Stat et al. (2009) with anannealing temperature of 53 �C. PCR products were purified usingthe QIAquick™ PCR Purification Kit (Qiagen), and ligated into thepGEM-T Easy vector™ (Promega). A minimum of 12 positive in-serts per clone library were amplified using plasmid-specific(M13) primers and sequenced in both directions using the ABIPrism Big Dye™ Terminator Cycle Sequencing Ready Reaction Kitand an ABI 3100 Genetic Analyzer (Perkin-Elmer Applied Biosys-tems) at the University of Hawai’i.
The nuclear and chloroplast clone libraries originating from theforams OHU15 and OHU66 were also screened with two alternativeprimer sets designed to target the low abundance novel sequencetypes in the mixed Symbiodinium communities harbored by theseforams (see Section 3). These primers targeted a 973-bp fragmentof the nuclear rDNA encompassing the ITS-2 and nr28S D1–D3regions (primer set 4: ‘ITS_I_for’ 50-GCTCATTGCACCCTTGG-30 and‘LSU_I_rev’: 50-ACACCTTCAGGCTTCCCA-30), and a 464-bp of thecp23S-DomainV (primer set 5: ‘23S_I_for’ 50-GCTCAAATGA-TATGCTGTAGAAG-30 and ‘23S_I_rev’ 50-GAGCACTCCTATCTGGAC-ACTA-30). Primer sets 4 and 5 were used to PCR screen 96 clonesof the nuclear and chloroplastic clone libraries originally generatedusing primer sets 2 and 3, respectively. Positive amplifications weresequenced in both directions as described above.
2.2. Data analyses
Each sequence was identified via BLAST in GenBank, as well aslocal BLAST to our in house Symbiodinium databases. Threeseparate sequence alignments were created using BioEdit v5.0.9(Hall, 1999): the ITS-2 (from primer set 1), the ITS-2 + nr28S (fromprimer sets 2 and 4), and the cp23S (from primer sets 3 and 5).Each alignment was manually edited and only identical sequencesrecovered from two or more independent samples included in thedownstream analyses. Unconfirmed clone singletons were treatedas artifacts or rare intragenomic repeats and the nucleotide at thesite of polymorphism converted to the consensus (Stat et al., 2009).ITS-2 secondary structure folding was conducted using previouslypublished templates (Hunter et al., 2007; Stat et al., 2009) in theITS-2 database interactive website (Selig et al., 2007) and manuallyedited using the software 4SALE (Seibel et al., 2006).
Edited nr28S (region D1–D3) and cp23S (Domain V) sequenceswere added to alignments previously published by Pochon et al.(2006). The best-fit model of DNA evolution was estimated for each
dataset using the program MrModeltest v2.2 (Posada and Crandall,1998). The maximum likelihood (ML) method was performed witha heuristic search and random addition of sequences using PAUP*
v4.0b10 (Swofford, 2002). Starting tree were obtained via stepwiseaddition of taxa, and then swapped using the tree-bisection–recon-nection (TBR) algorithm. The reliability of internal branches wasassessed using 100 bootstraps. Bayesian tree reconstructions withposterior probabilities and inferred using MrBayes v3.1 (Huelsen-beck and Ronquist, 2001) using the same model of DNA evolutionas for the ML analyses. Four simultaneous Markov chains were runfor 1,000,000 generations with trees sampled every 10 generations,with 50,000 initial trees discarded as ‘‘burn-in”, based on visualinspections.
The position of the novel Symbiodinium sequences in nr28S andcp23S topologies was tested using the Shimodaira–Hasegawatopological congruency test (SH-test; Shimodaira and Hasegawa,1999) implemented in PAUP* v4.0b10 (Swofford, 2002). For bothtopologies, we constrained the novel Symbiodinium sequences toform a monophyletic group with their neighboring clades andtested whether the likelihood values differed significantly fromthose obtained with the best trees. Lastly, uncorrected genetic dis-tances between 13 sequences in the nr28S dataset were calculatedusing the program Mega v4.0 (Tamura et al., 2007). The sequenceshowing the shortest branch length in each Symbiodinium cladeand sub-clade was selected (see Fig. 2A). In cases where several se-quences showed the same short branch length, one sequence washaphazardly chosen among them and included in the analysis.
3. Results
3.1. Clone libraries
Consistent with previous studies, Symbiodinium types in cladesC and F were prevalent in most foraminifera examined (data notshown), however, four samples (OHU3, OHU7, OHU15, andOHU66) harbored new ITS-2 sequence types that displayed only78% similarity to the most closely related Symbiodinium sequencein the BLAST analyses. All ITS-2 sequences recovered from samplesOHU3 (14 clones sequenced) and OHU7 (12 clones sequenced) cor-responded to these new types, and they co-occurred with sub-clade F4 Symbiodinium in OHU15 (2 of 12 clones sequenced,respectively) and OHU66 (2 of 9 sequences, respectively).
Twelve additional sequences were then generated for eachclone library corresponding to primer sets 2 (ITS-2 + nr28S) and 3(cp23S), and confirmed that only new types were detectable in

Fig. 2. Maximum likelihood phylograms of the genus Symbiodinium showing the position of clade I based on the previously published (A) nr28S and (B) cp23S datasets(Pochon et al., 2006). Numbers at nodes represent the result of the ML bootstrap analysis (underlined numbers; hundred bootstrap pseudoreplicates performed) and Bayesianposterior probabilities. Black dots represent values of 100% bootstrap support (BP) and Bayesian posterior probabilities (BiPP) of 1.0. Nodes without numbers correspond tosupports weaker than 70% BP and 0.8 BiPP. For each sequence, a sample reference number is indicated, followed by the corresponding GenBank accession number in squaredbrackets. The 13 nr28S sequences selected for the genetic divergence analysis (see Table 2) are indicated by asterisks.
Table 1The diversity and abundance of the new Symbiodinium sequences detected in four foraminifera sampled in Hawaii. Four unique sequence types were identified using both thenuclear (nr-i1–nr-i4) and chloroplastic (cp-i1–cp-i4) markers. Numbers correspond to the number of sequences retrieved from our clone libraries for each sample and sequencetype.
Sample ID Collection site Nuclear (ITS-2 + 28S rDNA) Chloroplastic (23S rDNA)
nr-i1 nr-i2 nr-i3 nr-i4 cp-i1 cp-i2 cp-i3 cp-i4
OHU 3a Mahi Wreck 6 5 0 1 8 2 1 1OHU 7a Mahi Wreck 9 3 0 0 8 4 0 0OHU 15b Mahi Wreck 0 0 1 9 3 1 1 4OHU 66b Lanai Lookout 0 0 5 3 0 0 4 3
a Samples harboring exclusively new sequences.b Samples harboring new sequences co-occurring with clade F sequences.
494 X. Pochon, R.D. Gates / Molecular Phylogenetics and Evolution 56 (2010) 492–497
OHU3 and OHU7. However, the 12 clones sequenced from clone li-braries generated for OHU15 and OHU66 using these primer setswere all sub-clade F4 sequences, reflecting the clear dominanceof F4 in these samples. To address this issue, primers specific tothe new types (primer sets 4 and 5) were used to screen bacterialcolonies in the clone libraries generated for OHU15 and OHU66with primer sets 2 and 3. Of the 384 clones screened using this ap-proach (96 clones per sample and per marker), between 7 and 10 ofthe new sequence types were recovered for OHU66 and OHU15,respectively (Table 1).
3.2. Sequence data
Four new nuclear ITS-2 + nr28S and four new cp23S sequencetypes were recovered (Table 1). The four nuclear sequence typesdiffered from one another by between 3 and 20 bp and the fourcp23S sequences by between 1 and 13 bp. The secondary structureanalysis of all ITS-2 sequences (N = 71; primer sets 1, 2, and 4) col-lapsed the four ITS-2 sequence types into a single fold (Fig. 1B) cor-responding to the five-stem model previously published by Hunteret al. (2007). The predicted ITS-2 secondary structure for each of
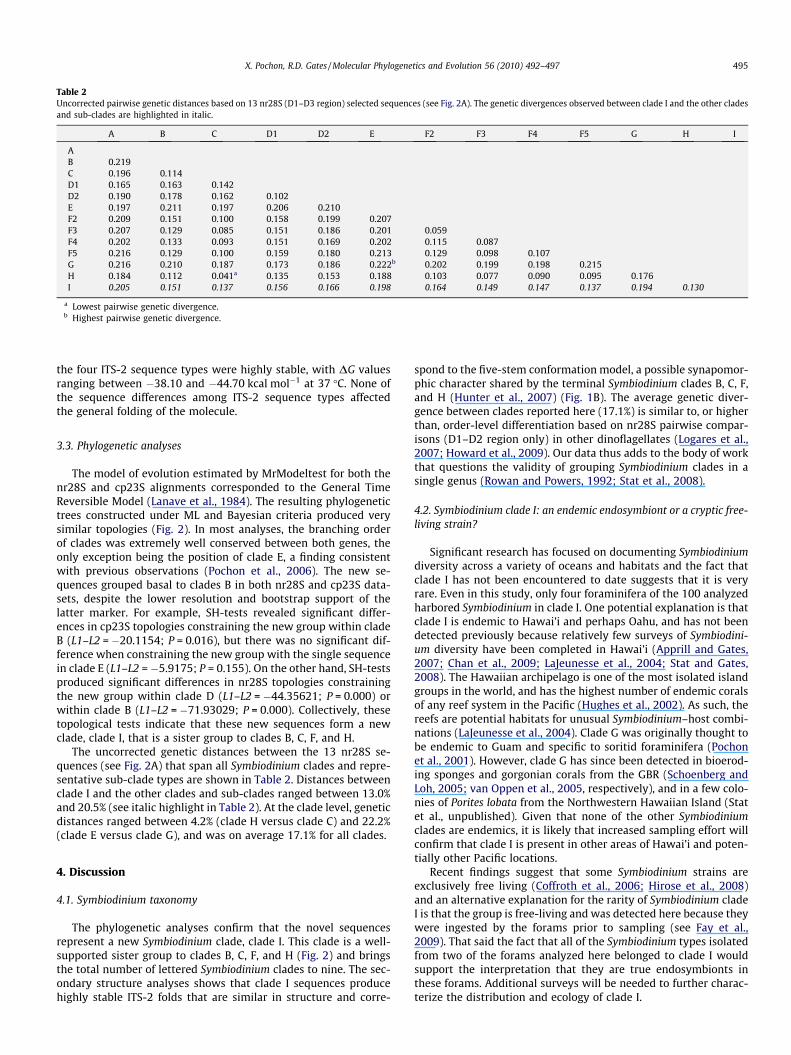
Table 2Uncorrected pairwise genetic distances based on 13 nr28S (D1–D3 region) selected sequences (see Fig. 2A). The genetic divergences observed between clade I and the other cladesand sub-clades are highlighted in italic.
A B C D1 D2 E F2 F3 F4 F5 G H I
AB 0.219C 0.196 0.114D1 0.165 0.163 0.142D2 0.190 0.178 0.162 0.102E 0.197 0.211 0.197 0.206 0.210F2 0.209 0.151 0.100 0.158 0.199 0.207F3 0.207 0.129 0.085 0.151 0.186 0.201 0.059F4 0.202 0.133 0.093 0.151 0.169 0.202 0.115 0.087F5 0.216 0.129 0.100 0.159 0.180 0.213 0.129 0.098 0.107G 0.216 0.210 0.187 0.173 0.186 0.222b 0.202 0.199 0.198 0.215H 0.184 0.112 0.041a 0.135 0.153 0.188 0.103 0.077 0.090 0.095 0.176I 0.205 0.151 0.137 0.156 0.166 0.198 0.164 0.149 0.147 0.137 0.194 0.130
a Lowest pairwise genetic divergence.b Highest pairwise genetic divergence.
X. Pochon, R.D. Gates / Molecular Phylogenetics and Evolution 56 (2010) 492–497 495
the four ITS-2 sequence types were highly stable, with DG valuesranging between �38.10 and �44.70 kcal mol�1 at 37 �C. None ofthe sequence differences among ITS-2 sequence types affectedthe general folding of the molecule.
3.3. Phylogenetic analyses
The model of evolution estimated by MrModeltest for both thenr28S and cp23S alignments corresponded to the General TimeReversible Model (Lanave et al., 1984). The resulting phylogenetictrees constructed under ML and Bayesian criteria produced verysimilar topologies (Fig. 2). In most analyses, the branching orderof clades was extremely well conserved between both genes, theonly exception being the position of clade E, a finding consistentwith previous observations (Pochon et al., 2006). The new se-quences grouped basal to clades B in both nr28S and cp23S data-sets, despite the lower resolution and bootstrap support of thelatter marker. For example, SH-tests revealed significant differ-ences in cp23S topologies constraining the new group within cladeB (L1–L2 = �20.1154; P = 0.016), but there was no significant dif-ference when constraining the new group with the single sequencein clade E (L1–L2 = �5.9175; P = 0.155). On the other hand, SH-testsproduced significant differences in nr28S topologies constrainingthe new group within clade D (L1–L2 = �44.35621; P = 0.000) orwithin clade B (L1–L2 = �71.93029; P = 0.000). Collectively, thesetopological tests indicate that these new sequences form a newclade, clade I, that is a sister group to clades B, C, F, and H.
The uncorrected genetic distances between the 13 nr28S se-quences (see Fig. 2A) that span all Symbiodinium clades and repre-sentative sub-clade types are shown in Table 2. Distances betweenclade I and the other clades and sub-clades ranged between 13.0%and 20.5% (see italic highlight in Table 2). At the clade level, geneticdistances ranged between 4.2% (clade H versus clade C) and 22.2%(clade E versus clade G), and was on average 17.1% for all clades.
4. Discussion
4.1. Symbiodinium taxonomy
The phylogenetic analyses confirm that the novel sequencesrepresent a new Symbiodinium clade, clade I. This clade is a well-supported sister group to clades B, C, F, and H (Fig. 2) and bringsthe total number of lettered Symbiodinium clades to nine. The sec-ondary structure analyses shows that clade I sequences producehighly stable ITS-2 folds that are similar in structure and corre-
spond to the five-stem conformation model, a possible synapomor-phic character shared by the terminal Symbiodinium clades B, C, F,and H (Hunter et al., 2007) (Fig. 1B). The average genetic diver-gence between clades reported here (17.1%) is similar to, or higherthan, order-level differentiation based on nr28S pairwise compar-isons (D1–D2 region only) in other dinoflagellates (Logares et al.,2007; Howard et al., 2009). Our data thus adds to the body of workthat questions the validity of grouping Symbiodinium clades in asingle genus (Rowan and Powers, 1992; Stat et al., 2008).
4.2. Symbiodinium clade I: an endemic endosymbiont or a cryptic free-living strain?
Significant research has focused on documenting Symbiodiniumdiversity across a variety of oceans and habitats and the fact thatclade I has not been encountered to date suggests that it is veryrare. Even in this study, only four foraminifera of the 100 analyzedharbored Symbiodinium in clade I. One potential explanation is thatclade I is endemic to Hawai’i and perhaps Oahu, and has not beendetected previously because relatively few surveys of Symbiodini-um diversity have been completed in Hawai’i (Apprill and Gates,2007; Chan et al., 2009; LaJeunesse et al., 2004; Stat and Gates,2008). The Hawaiian archipelago is one of the most isolated islandgroups in the world, and has the highest number of endemic coralsof any reef system in the Pacific (Hughes et al., 2002). As such, thereefs are potential habitats for unusual Symbiodinium–host combi-nations (LaJeunesse et al., 2004). Clade G was originally thought tobe endemic to Guam and specific to soritid foraminifera (Pochonet al., 2001). However, clade G has since been detected in bioerod-ing sponges and gorgonian corals from the GBR (Schoenberg andLoh, 2005; van Oppen et al., 2005, respectively), and in a few colo-nies of Porites lobata from the Northwestern Hawaiian Island (Statet al., unpublished). Given that none of the other Symbiodiniumclades are endemics, it is likely that increased sampling effort willconfirm that clade I is present in other areas of Hawai’i and poten-tially other Pacific locations.
Recent findings suggest that some Symbiodinium strains areexclusively free living (Coffroth et al., 2006; Hirose et al., 2008)and an alternative explanation for the rarity of Symbiodinium cladeI is that the group is free-living and was detected here because theywere ingested by the forams prior to sampling (see Fay et al.,2009). That said the fact that all of the Symbiodinium types isolatedfrom two of the forams analyzed here belonged to clade I wouldsupport the interpretation that they are true endosymbionts inthese forams. Additional surveys will be needed to further charac-terize the distribution and ecology of clade I.

496 X. Pochon, R.D. Gates / Molecular Phylogenetics and Evolution 56 (2010) 492–497
4.3. Describing Symbiodinium diversity using cloning
Several molecular techniques are routinely used to exploreSymbiodinium diversity and taxonomy. These include the analysisof restriction fragment length polymorphisms (RFLP, e.g., Rowanand Knowlton, 1995), size screening using single-stranded confor-mation polymorphisms (SSCP, e.g., van Oppen et al., 2005), dena-turing gradient gel electrophoresis (DGGE, e.g., LaJeunesse et al.,2004), sequencing of bacterially cloned rDNA (e.g., Apprill andGates, 2007), direct sequencing (e.g., Loh et al., 2001), and real-time PCR (qPCR, e.g., Mieog et al., 2007). Each of these methodsprovides complimentary but different information about the Sym-biodinium samples under investigation. For example, RFLP providesa rapid way of describing taxonomy at the cladal level, DGGE hasbeen used broadly to screen for ecologically dominant types, qPCRmeasures the relative abundance of one or two members in amixed community, and sequencing of bacterially cloned rDNA al-lows the diversity of all Symbiodinium sequences present in amixed sample to be characterized. Each molecular technique hasmethodological pros and cons (see Sampayo et al., 2009), and theyare generally used to address very different questions.
Here, phylogenetic analysis of bacterially cloned nuclear andchloroplastic rDNA gene fragments was used to characterize anew clade of Symbiodinium. Cloning has been criticized for itsinability to distinguish between ecologically relevant types andintragenomic variants and overestimating Symbiodinium diversity(Thornhill et al., 2007). The basis for this criticism lies in high levelsof intragenomic variation in the nuclear rDNA markers used to ex-plore Symbiodinium diversity (see Stat et al., 2009) and reflects thesensitivity of cloning. We exploited this sensitivity while address-ing the problem of intragenomic variation in the nrDNA by analyz-ing a chloroplast gene in parallel with the nuclear rDNA fragments.The chloroplast genes in dinoflagellates are maintained at low copynumbers during division by unique regulatory processes (Kouman-dou and Howe, 2007). As such, we expected that high levels ofintragenomic variation would be reflected in greater number of nu-clear sequence types relative to chloroplast sequence types. Forclade I, we recovered the same number of nuclear and chloroplastrDNA sequence types, suggesting that these sequences representfour independent Symbiodinium types each characterized by onenrDNA and one cpDNA signature.
Acknowledgments
We thank Michael Stat, Hollie Putnam, and Anderson Mayfieldfor valuable discussions on the manuscript and Daniel Burns fortechnical assistance. Phylogenetic analyses were performed viathe Bioportal computer service (http://www.bioportal.uio.no) atthe University of Oslo, Norway. This work was supported by theNSF (OCE-0752604 to R.D.G.) and the School of Ocean and EarthScience and Technology, University of Hawai’i. Collections werepermitted under DAR Special Activity Permit No. 2006-14. This isHIMB contribution number 1386.
References
Apprill, A.M., Gates, R.D., 2007. Recognizing diversity in coral symbioticdinoflagellate communities. Mol. Ecol. 16, 1127–1134.
Baker, A.C., Glynn, P.W., Riegl, B., 2008. Climate change and coral reef bleaching: anecological assessment of long-term impacts, recovery trends and futureoutlook. Estuar. Coast. Shelf. Sci. 80, 435–471.
Chan, Y.L., Pochon, X., Fisher, M.A., Wagner, D., Conception, G.T., Kahng, S.E., Toonen,R.J., Gates, R.D., 2009. Generalist dinoflagellate endosymbionts and hostgenotype diversity detected from mesophotic (67–100 m depths) coralLeptoseris. BMC Ecol. 9, 21.
Coffroth, M.A., Santos, S.R., 2005. Genetic diversity of symbiotic dinoflagellates inthe genus Symbiodinium. Protist 156, 19–34.
Coffroth, M.A., Lewis, C., Santos, S., Weaver, J., 2006. Environmental populations ofsymbiotic dinoflagellates in the genus Symbiodinium can initiate symbioseswith reef cnidarians. Curr. Biol. 16, R985–R987.
Fay, S.A., Weber, M.X., Lipps, J.H., 2009. The distribution of Symbiodinium diversitywithin individual host foraminifera. Coral Reefs 28, 717–726.
Hall, T.A., 1999. BioEdit: a user-friendly biological sequence alignment editor andanalysis program for Windows 95/98/NT. Nucelic Acid Symp. Ser. 41, 95–98.
Hirose, M., Reimer, J.D., Hidaka, M., Suda, S., 2008. Phylogenetic analyses ofpotentially free-living Symbiodinium spp. isolated from coral reef sand inOkinawa, Japan. Mar. Biol. 155, 105–112.
Howard, M.D.A., Smith, G.J., Kudela, R.M., 2009. Phylogenetic relationships ofyessotoxin-producing dinoflagellates, based on the large subunit and internaltranscribed spacer ribosomal DNA domains. Appl. Environ. Microbiol. 75, 54–63.
Huelsenbeck, J.P., Ronquist, F., 2001. MrBayes: a program for the Bayesian inferenceof phylogeny. Bioinformatics 17, 754–755.
Hughes, T.P., Bellwood, D.R., Connolly, S.R., 2002. Biodiversity hotspots, centers ofendemicity, and the conservation of coral reefs. Ecol. Lett. 5, 775–784.
Hunter, R.L., LaJeunesse, T.C., Santos, S.R., 2007. Structure and evolution of the rDNAinternal transcribed spacer (ITS) region 2 in the symbiotic dinoflagellates(Symbiodinium, Dinophyta). J. Phycol. 43, 120–128.
Koumandou, V.L., Howe, C.J., 2007. The copy number of chloroplast gene minicircleschanges dramatically with growth phase in the dinoflagellate Amphidiniumoperculatum. Protist 158, 89–103.
LaJeunesse, T.C., 2001. Investigating the biodiversity, ecology, and phylogeny ofendosymbiotic dinoflagellates in the genus Symbiodinium using the ITS region:in search of a ‘‘species” level marker. J. Phycol. 37, 866–880.
LaJeunesse, T.C., Thornhill, D.J., Cox, E.F., Stanton, F.G., Fitt, W.K., Schmidt, G.W.,2004. High diversity and host specificity observed among symbioticdinoflagellates in reef coral communities from Hawai’i. Coral Reefs 23, 596–603.
Lanave, C., Preparata, G., Saccone, C., Serio, G., 1984. A new method for calculatingevolutionary substitution rates. J. Mol. Evol. 20, 86–93.
Logares, R., Shalchian-Tabrizi, K., Boltovskoy, A., Rengefors, K., 2007. Extensivedinoflagellate phylogenies indicate infrequent marine–freshwater transitions.Mol. Phylogenet. Evol. 45, 887–903.
Loh, W.K., Loi, T., Carter, D., Hoegh-Guldberg, O., 2001. Genetic variability of thesymbiotic dinoflagellates from the wide ranging coral species Seriatopora hystrixand Acropora longicyathus in the Indo-West Pacic. Mar. Ecol. Prog. Ser. 222, 97–107.
Mieog, J.C., van Oppen, M.J.H., Cantin, N.E., Stam, W.T., Olsen, J.L., 2007. Real-timePCR reveals a high incidence of Symbiodinium clade D at low levels in fourscleractinian corals across the Great Barrier Reef: implications for symbiontshuffling. Coral Reefs 26, 449–457.
Pochon, X., Pawlowski, J., 2006. Evolution of the soritids–Symbiodinium symbiosis.Symbiosis 42, 77–88.
Pochon, X., Pawlowski, J., Zaninetti, L., Rowan, R., 2001. High genetic diversity andrelative specificity among Symbiodinium-like endosymbiotic dinoflagellates insoritid foraminiferans. Mar. Biol. 139, 1069–1078.
Pochon, X., LaJeunesse, T.C., Pawlowski, J., 2004. Biogeographic partitioning andhost specialization among foraminiferan dinoflagellate symbionts(Symbiodinium; Dinophyta). Mar. Biol. 146, 17–27.
Pochon, X., Montoya-Burgos, J.I., Stadelmann, B., Pawlowski, J., 2006. Molecularphylogeny, evolutionary rates, and divergence timing of the symbioticdinoflagellate genus Symbiodinium. Mol. Phylogenet. Evol. 38, 20–30.
Posada, D., Crandall, K.A., 1998. Modeltest: testing the model of DNA substitution.Bioinformatics 14, 817–818.
Rodriguez-Lanetty, M., 2003. Evolving lineages of Symbiodinium-like dinoflagellatesbased on ITS1 rDNA. Mol. Phylogenet. Evol. 28, 152–168.
Rowan, R., Knowlton, N., 1995. Intraspecific diversity and ecological zonation incoral–algal symbiosis. Proc. Natl. Acad. Sci. USA 92, 2850–2853.
Rowan, R., Powers, D., 1992. Ribosomal RNA sequences and the diversity of symbioticdinoflagellates (zooxanthellae). Proc. Natl. Acad. Sci. USA 89, 3639–3643.
Sampayo, E.M., Dove, S., LaJeunesse, T.C., 2009. Cohesive molecular genetic datadelineate species diversity in the dinoflagellate genus Symbiodinium. Mol. Ecol.18, 500–519.
Schoenberg, C.H.L., Loh, W., 2005. Molecular identity of the unique symbioticdinoflagellates found in the bioeroding demosponge Cliona orientalis Thiele,1900. Mar. Ecol. Prog. Ser. 299, 157–166.
Seibel, P.N., Muller, T., Dandekar, T., Schultz, J., Wolf, M., 2006. 4SALE – a tool forsynchronous RNA sequence and secondary structure alignment and editing.BMC Bioinform. 7, 498.
Selig, C., Wolf, M., Muller, T., Dandekar, T., Schultz, J., 2007. The ITS2 Database II:homology modelling RNA structure for molecular systematics. Nucleic AcidsRes. 36, D377–D380.
Shimodaira, H., Hasegawa, M., 1999. Multiple comparisons of log-likelihoods withapplications to phylogenetic inference. Mol. Biol. Evol. 16, 1114–1116.
Stat, M., Gates, R.D., 2008. Vectored introductions of marine endosymbioticdinoflagellates into Hawai’i. Biol. Invasions 10, 579–583.
Stat, M., Carter, D., Hoegh-Guldberg, O., 2006. The evolutionary history ofSymbiodinium and scleractinian hosts – symbiosis, diversity, and the effect ofclimate change. Persp. Plant Ecol. Evol. Syst. 8, 23–43.
Stat, M., Morris, E., Gates, R.D., 2008. Functional diversity in coral–dinoflagellatesymbiosis. Proc. Natl. Acad. Sci. USA 105, 9256–9261.
Stat, M., Pochon, X., Cowie, R.O.M., Gates, R.D., 2009. Specificity in communities ofSymbiodinium in corals from Johnston Atoll. Mar. Ecol. Prog. Ser. 386, 83–96.
Swofford, D.L., 2002. PAUP*: Phylogenetic Analysis Using Parsimony (* and OtherMethods), Version 4.0b10. Sinauer, Sunderland, MA.

X. Pochon, R.D. Gates / Molecular Phylogenetics and Evolution 56 (2010) 492–497 497
Tamura, K., Dudley, J., Nei, M., Kumar, S., 2007. MEGA4: molecular evolutionarygenetics analysis (MEGA) software version 4.0. Mol. Biol. Evol. 24, 1596–1599.
Tchernov, D., Gorbunov, M.Y., de Vargas, C., Yadav, S.N., Milligan, A.J., Häggblom, M.,Falkowski, P.G., 2004. Membrane lipids of symbiotic algae are diagnostic ofsensitivity to thermal bleaching in corals. Proc. Natl. Acad. Sci. USA 101, 13531–13535.
Thornhill, D.J., LaJeunesse, T.C., Santos, S.R., 2007. Measuring rDNA diversity ineukaryotic microbial systems: how intragenomic variation, pseudogenes, andPCR artifacts confound biodiversity estimates. Mol. Ecol. 16, 5326–5340.
Trench, R.K., 1993. Microalgal–invertebrate symbiosis: a review. Endocytobiol. CellRes. 9, 135–175.
van Oppen, M.J.H., Mahiny, A.J., Done, T.J., 2005. Geographic distribution ofzooxanthella groups in three coral species on the Great Barrier Reef sampledafter the 2002 bleaching event. Coral Reefs 24, 482–487.
van Oppen, M.J.H., Baker, A.C., Coffroth, M.A., Willis, B.L., 2008. Bleaching resistanceand the role of algal endosymbionts. In: Lough, J.M., van Oppen, M.J.H. (Eds.),Coral Bleaching: Patterns, Processes, Causes and Consequences. SpringerEcological Studies, Berlin, pp. 84–102.




















