
The taxonomy and growth of a Crypthecodinium species (Dinophyceae) isolated from a brackish-water...
7
African Journal of Marine Science 2006, 28(2): 185–191 Printed in South Africa — All rights reserved Copyright © NISC Pty Ltd AFRICAN JOURNAL OF MARINE SCIENCE EISSN 1814–2338 The taxonomy and growth of a Crypthecodinium species (Dinophyceae) isolated from a brackish-water fish aquarium MW Parrow 1 *, M Elbrächter 2 , MK Krause 3 , JM Burkholder 1 , NJ Deamer 1 , N Htyte 3 and EH Allen 1 1 Center for Applied Aquatic Ecology, North Carolina State University, 620 Hutton Street, Suite 104, Raleigh, NC 27606, USA; current address: Department of Biology, University of North Carolina at Charlotte, 9201 University City Boulevard, Charlotte, NC 28223, USA 2 Deutsches Zentrum für Marine Biodiversitätsforschung, Forschungsinstitut Senckenberg, Wattenmeerstation Sylt, Hafenstrasse 43, D-25992 List/Sylt, Germany 3 Department of Biology, 114 Hofstra University, Hempstead, NY 11549, USA * Corresponding author, e-mail: [email protected] Introduction Biecheler (1938, 1952) described the heterotrophic dinofla- gellate Crypthecodinium setense from the Mediterranean coast of France using silver staining and light microscopy to determine the thecal plate tabulation. Biecheler (1952) placed C. setense in a new family, the Crypthecodiniaceae, and considered the possibility that it might be identical to the previously described Gyrodinium cohnii (Seligo) Schiller, if the same plate tabulation could be proven in the latter. Chatton (1952) regarded these dinoflagellates as conspe- cific, and introduced the combination Crypthecodinium cohnii (Seligo) Chatton, but did not demonstrate a shared thecal structure. Since then, heterotrophic dinoflagellates roughly similar in size and shape have been called C. cohnii and used extensively in research on dinoflagellate physiology, ultra- structure and genetics (Beam and Himes 1984, Barlow and Triemer 1986, Perret et al. 1993, Bhaud et al. 1994, 2000, Lam et al. 2001). Sexual compatibility experiments and genetic analyses of numerous isolates have suggested that the present morphospecies conception of C. cohnii may encom- pass several biological species (Beam and Himes 1977, 1982, Beam et al. 1993). Ucko et al. (1989, 1997) reported a C. cohnii-like dino- flagellate occurring as a deleterious contaminant in commer- cial pond cultures of Porphyridium sp., a unicellular red alga mass-cultured for production of valuable biochemicals. The Crypthecodinium dinoflagellate caused reduced yield and collapse of the Porphyridium cultures by feeding on the red algal cells (Ucko et al. 1989). Therefore, although not known to be toxic, Crypthecodinium may be considered a poten- tially harmful alga because it can negatively affect some commercial mass-algal culture operations, resulting in finan- cial loss. A relatively small heterotrophic dinoflagellate found in abundance in a brackish-water fish aquarium at North Carolina State University, USA, was suspected of being a Crypthecodinium species, based on observations using light microscopy. During similar culture experiments (Parrow et al. 2005), a clonal isolate of this dinoflagellate was estab- lished and has been cultivated for more than 1.5 years using a Chinook salmon Oncorhynchus tshawytscha cell line as the food source. The objectives of this study were to (1) evaluate the taxonomy of this dinoflagellate by examining its morphology and small sub-unit (18S) rDNA sequence; An unidentified heterotrophic dinoflagellate found growing in abundance in a brackish-water fish aquar- ium was isolated and serially cultivated using a fish cell line as the food source. Prominent characteristics of this dinoflagellate included a cingulum that did not fully encircle the motile cell, cell division in non-motile cysts, and a theca composed of thin but structured plates. Morphological analysis of flagellate cells by scanning electron microscopy revealed a Kofoid thecal plate tabulation of 4’, 4a, 4’’, ‘X’, 5 or 6c, ?s, 5’’’, 1p, 1’’’’, most consistent with the original description of Cryp- thecodinium setense Biecheler. This Crypthecodinium species exhibited a high maximum division rate (3.2 divisions day –1 ) and cell yield (>10 6 cells ml –1 ) when fed cultured fish cells. Small sub-unit rDNA phylogenetic analyses supported relatedness with a previously studied Crypthecodinium-like dinoflagellate, but a significant difference in aligned gene sequences was found. This study provides the first clear demonstra- tion of the plate tabulation of a Crypthecodinium spe- cies since the original description over 60 years ago, allowing the original morphological conception of Crypthecodinium to be linked with molecular phylo- genetic information. Keywords: Crypthecodinium, dinoflagellate, phylogeny, small sub-unit rDNA, taxonomy, theca
Transcript of The taxonomy and growth of a Crypthecodinium species (Dinophyceae) isolated from a brackish-water...

African Journal of Marine Science 2006, 28(2): 185–191Printed in South Africa — All rights reserved
Copyright © NISC Pty LtdAFRICAN JOURNAL OF
MARINE SCIENCEEISSN 1814–2338
The taxonomy and growth of a Crypthecodinium species (Dinophyceae)isolated from a brackish-water fish aquarium
MW Parrow1*, M Elbrächter2, MK Krause3, JM Burkholder1, NJ Deamer1, N Htyte3 and EH Allen1
1 Center for Applied Aquatic Ecology, North Carolina State University, 620 Hutton Street, Suite 104, Raleigh, NC 27606, USA;current address: Department of Biology, University of North Carolina at Charlotte, 9201 University City Boulevard, Charlotte,NC 28223, USA2 Deutsches Zentrum für Marine Biodiversitätsforschung, Forschungsinstitut Senckenberg, Wattenmeerstation Sylt,Hafenstrasse 43, D-25992 List/Sylt, Germany3 Department of Biology, 114 Hofstra University, Hempstead, NY 11549, USA* Corresponding author, e-mail: [email protected]
Introduction
Biecheler (1938, 1952) described the heterotrophic dinofla-
gellate Crypthecodinium setense from the Mediterranean
coast of France using silver staining and light microscopy to
determine the thecal plate tabulation. Biecheler (1952)
placed C. setense in a new family, the Crypthecodiniaceae,
and considered the possibility that it might be identical to
the previously described Gyrodinium cohnii (Seligo) Schiller,
if the same plate tabulation could be proven in the latter.
Chatton (1952) regarded these dinoflagellates as conspe-
cific, and introduced the combination Crypthecodiniumcohnii (Seligo) Chatton, but did not demonstrate a shared
thecal structure.
Since then, heterotrophic dinoflagellates roughly similar
in size and shape have been called C. cohnii and used
extensively in research on dinoflagellate physiology, ultra-
structure and genetics (Beam and Himes 1984, Barlow and
Triemer 1986, Perret et al. 1993, Bhaud et al. 1994, 2000, Lam
et al. 2001). Sexual compatibility experiments and genetic
analyses of numerous isolates have suggested that the
present morphospecies conception of C. cohnii may encom-
pass several biological species (Beam and Himes 1977,
1982, Beam et al. 1993).
Ucko et al. (1989, 1997) reported a C. cohnii-like dino-
flagellate occurring as a deleterious contaminant in commer-
cial pond cultures of Porphyridium sp., a unicellular red alga
mass-cultured for production of valuable biochemicals. The
Crypthecodinium dinoflagellate caused reduced yield and
collapse of the Porphyridium cultures by feeding on the red
algal cells (Ucko et al. 1989). Therefore, although not known
to be toxic, Crypthecodinium may be considered a poten-
tially harmful alga because it can negatively affect some
commercial mass-algal culture operations, resulting in finan-
cial loss.
A relatively small heterotrophic dinoflagellate found in
abundance in a brackish-water fish aquarium at North
Carolina State University, USA, was suspected of being a
Crypthecodinium species, based on observations using light
microscopy. During similar culture experiments (Parrow etal. 2005), a clonal isolate of this dinoflagellate was estab-
lished and has been cultivated for more than 1.5 years using
a Chinook salmon Oncorhynchus tshawytscha cell line as
the food source. The objectives of this study were to (1)
evaluate the taxonomy of this dinoflagellate by examining
its morphology and small sub-unit (18S) rDNA sequence;
An unidentified heterotrophic dinoflagellate found
growing in abundance in a brackish-water fish aquar-
ium was isolated and serially cultivated using a fish
cell line as the food source. Prominent characteristics
of this dinoflagellate included a cingulum that did not
fully encircle the motile cell, cell division in non-motile
cysts, and a theca composed of thin but structured
plates. Morphological analysis of flagellate cells by
scanning electron microscopy revealed a Kofoid thecal
plate tabulation of 4’, 4a, 4’’, ‘X’, 5 or 6c, ?s, 5’’’, 1p, 1’’’’,
most consistent with the original description of Cryp-thecodinium setense Biecheler. This Crypthecodinium
species exhibited a high maximum division rate (3.2
divisions day–1) and cell yield (>106 cells ml–1) when fed
cultured fish cells. Small sub-unit rDNA phylogenetic
analyses supported relatedness with a previously
studied Crypthecodinium-like dinoflagellate, but a
significant difference in aligned gene sequences was
found. This study provides the first clear demonstra-
tion of the plate tabulation of a Crypthecodinium spe-
cies since the original description over 60 years ago,
allowing the original morphological conception of
Crypthecodinium to be linked with molecular phylo-
genetic information.
Keywords: Crypthecodinium, dinoflagellate, phylogeny, small sub-unit rDNA, taxonomy, theca

Parrow, Elbrächter, Krause, Burkholder, Deamer, Htyte and Allen186
(2) describe its general mode of feeding and reproduction;
and (3) document its high rate of proliferation when culti-
vated on fish cells.
Material and Methods
The fish cell line (ATCC CRL-1681, Manassas, Virginia,
USA) was serially cultivated in Eagle’s minimum essential
medium (MEM) with 10% fetal bovine serum (FBS) in poly-
styrene cell culture flasks following the procedures of
Parrow et al. (2005). Prior to inoculation with dinoflagel-
lates, the MEM + FBS medium overlaying the adherent fish
cell layer was removed and replaced with filter-sterilised
(0.22µm pore size) artificial saltwater medium (ASM)
composed of (in g l–1) NaCl (8.1), MgSO4 (1.6), MgCl2 (1.2),
CaCl2 (0.4), KCl (0.3), NaHCO3 (0.1) and glucose (1.0) in
deionised water. The dinoflagellate culture was initiated by
treating a 20ml sample taken from the aquarium with peni-
cillin (50IU ml–1) and streptomycin (0.1mg ml–1) for 24h,
before isolating a single motile dinoflagellate cell from this
population using a sterile glass micropipette. The cell was
washed by micropipette through eight drops of sterile ASM
under aseptic conditions, and deposited into a culture flask
containing adherent fish cells overlaid with ASM. After the
dinoflagellate had proliferated into a cultured population, a
subsample (0.1ml) was serially transferred every seven
days into a newly prepared flask of fish cells. Observations
and photographs of living cells were made using light
microscopy in culture flask or Petri dish subcultures.
For scanning electron microscopy (SEM), culture aliquots
were treated with a 40% reduction in salinity for 20min and
then fixed in 1% OsO4, 2% glutaraldehyde and 0.1mol
sodium cacodylate (final concentration) at 4°C for 20min.
Fixed cells were filtered onto polycarbonate filters (3µm
pore size), rinsed in 0.1mol sodium cacodylate, dehydrated
through a graded ethanol concentration series (30%, 50%,
70%, 90%, 100%; 15min per step), critical point dried using
CO2, sputter coated with 25nm Au/Pd and viewed at 15kV
on a JEOL 5900LV SEM.
DNA was isolated from dinoflagellate samples using a
Qiagen Plant Mini Kit, and universal dinoflagellate 18S
primers (Saldarriaga et al. 2001) were used for sequence
amplification. Gel-purified 18S rDNA fragments were ampli-
fied by PCR, isolated and sequenced in both directions using
Amersham Thermo SequenaseTM. ClustalW was used for
multiple sequence alignment, and MEGA Version 3.1 was
used for phylogenetic analyses (Kumar et al. 2004). A mini-
mum evolution/maximum likelihood-distance consensus
phylogenetic tree was constructed with 1 500 bootstrap
replicates, based on alignment of sequence data from 41
additional taxa obtained from GenBank covering major
dinoflagellate groups (Table 1). Parameters were estimated
by a Kimura 2-parameter (Gamma) model and calculated with
the Close-Neighbour-Interchange algorithm. Toxoplasmagondii Nicolle et Manceaux was used as an outgroup.
Results and Discussion
The flagellate cells of this Crypthecodinium species were
8–24µm long, colourless, appeared superficially gymnodinoid
under light microscopy, and were approximately ovoid in
ventral or dorsal view and slightly compressed dorsoven-
trally. The cingulum did not fully encircle the cell, exhibiting
a leftward descending spiral that terminated near the right
lateral side after traversing roughly two-thirds of the cell
circumference (Figure 1a, b). The longitudinal flagellum
was approximately 1.5–2X the cell length (Figure 1a).
Investigation by SEM revealed that the flagellate cells
had thin thecae composed of structured plates. The thecal
plate tabulation was consistent with a Kofoid formula of 4’,
4a, 4”, ‘X’, 5 or 6c, ?s, 5’’’, 1p, 1’’’’ (Figure 2). No distinct apical
pore complex was discerned. The sulcal plates were not
resolved clearly enough to permit description and it was
uncertain whether the number of cingular plates was five or
six. The observed thecal plates were arranged in a pattern
most consistent with the original C. setense description
Table 1: List of species used in the phylogenetic analysis, with
GenBank accession numbers
Species Accession number
Alexandrium catenella AB088284
A. fundyense U09048
A. minutum U27499
Amylax diacantha AY443013
Amyloodinium ocellatum AF080096
Ceratium furca AJ276699
C. fusus AF022153
C. hirundinella AY443014
C. tenue AF022192
Ceratocorys horrida AF022154
Crypthecodinium cohnii M64245
Fragilidium subglobosum AF033869
Gloeodinium viscum L13716
Gonyaulax spinifera AF022155
Gymnodinium beii U37367
G. catenatum AF022193
G. fuscum AF022194
Gyrodinium instriatum AY443015
G. uncatenum AF274263
Hemidinium nasutum AY443016
Karenia brevis AF172714
K. mikimotoi AF022195
Karlodinium micrum AY245692
Lessardia elongata AF521100
Lingulodinium polyedrum AY421788
Noctiluca scintillans AF022200
Peridinium polonicum AY443017
Pfiesteria piscicida AF077055
P. shumwayae AY245694
Polarella glacialis AF099183
Prorocentrum micans AY585526
P. minimum Y16238
Protoceratium reticulatum AF274273
Protoperidinium conicum AY443020
P. pellucidum AY443022
Pyrocystis noctiluca AF022156
Scrippsiella sweeneyae AF274276
S. trochoidea AJ415515
Symbiodinium microadriaticum M88521
Thecadinium dragescoi AY238479
Toxoplasma gondii U00458
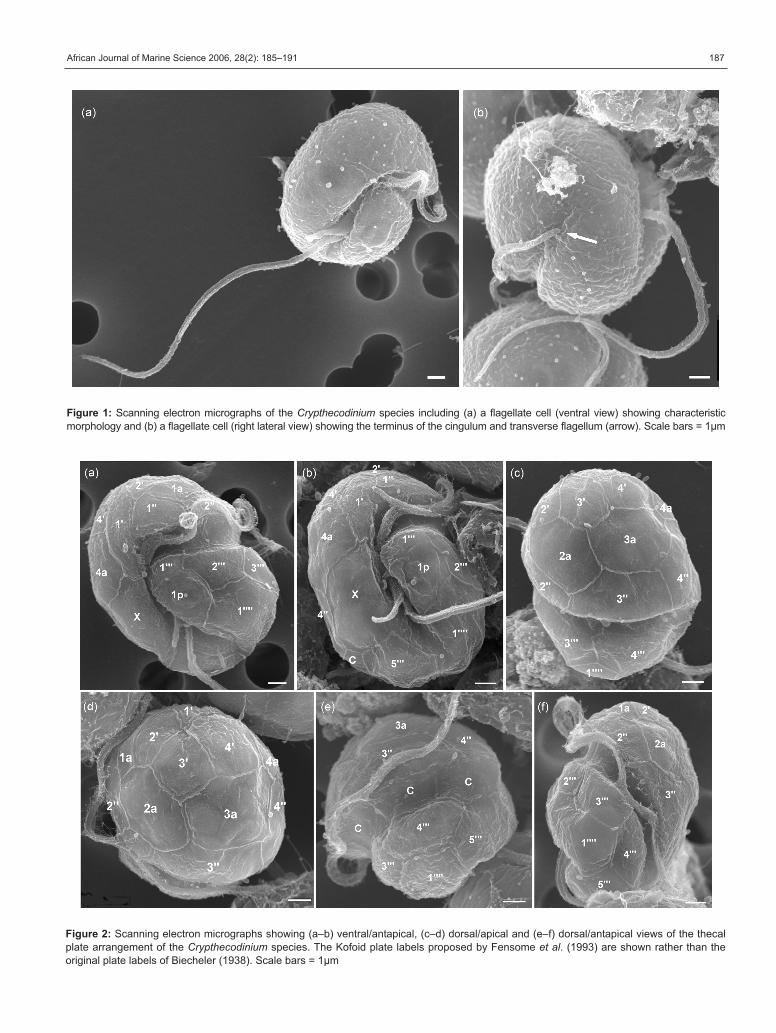
African Journal of Marine Science 2006, 28(2): 185–191 187
Figure 1: Scanning electron micrographs of the Crypthecodinium species including (a) a flagellate cell (ventral view) showing characteristic
morphology and (b) a flagellate cell (right lateral view) showing the terminus of the cingulum and transverse flagellum (arrow). Scale bars = 1µm
Figure 2: Scanning electron micrographs showing (a–b) ventral/antapical, (c–d) dorsal/apical and (e–f) dorsal/antapical views of the thecal
plate arrangement of the Crypthecodinium species. The Kofoid plate labels proposed by Fensome et al. (1993) are shown rather than the
original plate labels of Biecheler (1938). Scale bars = 1µm

Parrow, Elbrächter, Krause, Burkholder, Deamer, Htyte and Allen188
(Biecheler 1938; Figure 3), including the asymmetrical first
apical (1’) and the large ‘X’ plate to the right of the sulcus
extending from the epicone to the hypocone. The 1’ and 3’
plates differed from the original description, however, by
joining at a point rather than sharing a suture and having
one less side each (Figures 2d and 4). Possible taxonomic
implications of this morphological difference are uncertain,
because this type of plate variation can occur within some
dinoflagellate species (Elbrächter and Meyer 2001).
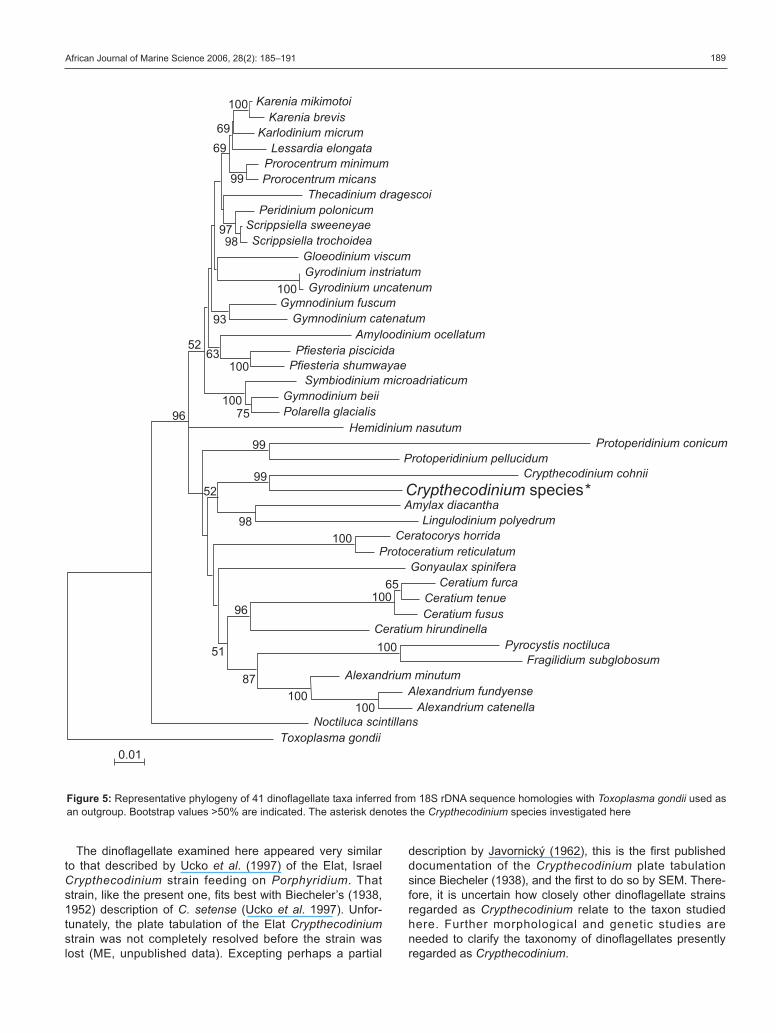
A novel 18S rDNA sequence was obtained from clonal
culture of this dinoflagellate (1 736 nucleotides; GenBank
accession number: DQ322643). Phylogenetic analysis
indicated relatedness between the 18S rDNA sequence of
this dinoflagellate and that of the sole published sequence
for an unspecified strain referred to as C. cohnii by
Gajadhar et al. (1991) (Figure 5). The aligned sequences
shared only 87% of base pairs, however, indicating that
these taxa were not identical. Branching order in the
phylogenetic tree was not well resolved, but there was
support for placement of Crypthecodinium in a major
clade that links dinoflagellate species currently assigned
to the order Gonyaulacales. The position of Cryptheco-dinium in a common gonyaulacoid lineage has been both
supported (Saldarriaga et al. 2001, 2004) and not suppor-
ted (Murray et al. 2005) in previous molecular phyloge-
netic analyses. This phylogenetic analysis did not indicate
close relatedness between Crypthecodinium and a fresh-
water dinoflagellate considered to be Hemidinium nasu-tum Stein, which also has thin thecal plates and a cingu-
lum that does not completely encircle the cell (Stein 1883,
Woloszyñska 1925). Crypthecodinium and Hemidiniumhave reported differences in tabulation pattern, and the
morphological characteristic of a cingulum that does not
fully encircle the cell may have arisen independently in
these taxa.
In this culture system, the dinoflagellates fed on fish cell
cytoplasm using an extensible feeding tube (Figure 6a),
and underwent cell division(s) within non-motile spherical
cysts (Figure 6b) that each produced 2–8 flagellate
offspring cells. Final yields of >106 dinoflagellate cells ml–1
occurred when this Crypthecodinium was cultivated
with fish cells as food (Figure 7), with a corresponding
maximum growth of 3.2 divisions day–1. Most present
Crypthecodinium-like strains are known for their ability to
grow on dissolved compounds in liquid and solid media
(Tuttle and Loeblich 1975, Beam and Himes 1977, 1982,
1984). However, Crypthecodinium has also been described
as a phagotroph that feeds on other protists (Biecheler
1952, Ucko et al. 1997). Feeding by Crypthecodinium on
fish cells in this study may have been a consequence of
the cultivation system rather than an indication of the natu-
ral prey preference of the dinoflagellate. Although this
Crypthecodinium strain might be capable of feeding on
living fish epithelium, no unusual fish deaths occurred in
the aquarium from which the dinoflagellate was isolated.
This Crypthecodinium strain can also feed on some chloro-
phyte and cryptophyte microalgae (MWP, unpublished
data), and is considered therefore to feed naturally on
various organisms.
Figure 3: The reported thecal plate pattern of Crypthecodinium setense: (a) ventral, (b) dorsal, (c) apical and (d) antapical; modified from
Biecheler (1938) and Fensome et al. (1993), using Kofoid plate labels adapted from Fensome et al. (1993)
Figure 4: Illustration of the observed apical plate arrangement of
the Crypthecodinium species
African Journal of Marine Science 2006, 28(2): 185–191 189
The dinoflagellate examined here appeared very similar
to that described by Ucko et al. (1997) of the Elat, Israel
Crypthecodinium strain feeding on Porphyridium. That
strain, like the present one, fits best with Biecheler’s (1938,
1952) description of C. setense (Ucko et al. 1997). Unfor-
tunately, the plate tabulation of the Elat Crypthecodiniumstrain was not completely resolved before the strain was
lost (ME, unpublished data). Excepting perhaps a partial
description by Javornický (1962), this is the first published
documentation of the Crypthecodinium plate tabulation
since Biecheler (1938), and the first to do so by SEM. There-
fore, it is uncertain how closely other dinoflagellate strains
regarded as Crypthecodinium relate to the taxon studied
here. Further morphological and genetic studies are
needed to clarify the taxonomy of dinoflagellates presently
regarded as Crypthecodinium.
�������������� �� �
�����������������������������
������������ ������������������������ ��������������� ������� ��
��� ������������ ���������������� �
� �������������������� �������������� ������
������������� ��������������������������� ����
����������� ���������� ������
������������ �������������������� � ����������������������������������� ��������� �
�������������������������� �����
�������������
������������������ ��� ������ ������ ����
������ �������������!�����"���� ����
������������������ ����� ������������
����� ����������� ����������"���������
��������� � ���������� ����������
�������������������� ������� ��� �
#����������������������"����������
���"���������������"������ �������
$� ��� ��� ���������"�������������
������
���
���
��
��
��
��
�����
����
��
�������
�����
��
��
���
����
���
��
��
��
����
Figure 5: Representative phylogeny of 41 dinoflagellate taxa inferred from 18S rDNA sequence homologies with Toxoplasma gondii used as
an outgroup. Bootstrap values >50% are indicated. The asterisk denotes the Crypthecodinium species investigated here

Parrow, Elbrächter, Krause, Burkholder, Deamer, Htyte and Allen190
References
Barlow SB, Triemer RE (1986) Phosphatase localization in the
endomembrane system of the dinoflagellate Crypthecodiniumcohnii. Journal of Histochemistry and Cytochemistry 34: 1021–1027
Beam CA, Himes M (1977) Sexual isolation and genetic diversifica-
tion among some strains of Crypthecodinium cohnii-like dinoflagel-
lates: evidence of speciation. Journal of Protozoology 24: 532–539
Beam CA, Himes M (1982) Distribution of members of the Crypthe-codinium cohnii (Dinophyceae) species complex. Journal ofProtozoology 29: 8–15
Beam CA, Himes M (1984) Dinoflagellate genetics. In: Spector DL
(ed) Dinoflagellates. Academic Press, Orlando, Florida, pp
263–298
Beam CA, Preparata RM, Himes M, Nanney DL (1993) Ribosomal
RNA sequencing of members of the Crypthecodinium cohnii(Dinophyceae) species complex; comparison with soluble enzyme
studies. Journal of Eukaryotic Microbiology 40: 660–667
Biecheler B (1938) Sur un Péridinien cuirasse incolore nouveau
Crypthecodinium n.g. setense n. sp. et la famille nouvelle des
Crypthecodiniacées. Bulletin del la Societé Zoologique de France63: 9–12
Biecheler B (1952) Recherches sur les Péridiniens. Bulletin Biolo-gique de France et de Belgique Supplément 36: 1–149
Bhaud Y, Barbier M, Soyer-Gobillard MO (1994) A detailed study of
the complex cell cycle of the dinoflagellate Crypthecodiniumcohnii Biecheler and evidence for variation in histone H1 kinase
activity. Journal of Eukaryotic Microbiology 41: 519–526
Bhaud Y, Guillebault D, Lennon JF, Defacque H, Soyer-Gobillard
MO, Moreau H (2000) Morphology and behavior of dinoflagel-
late chromosomes during the cell cycle and mitosis. Journal ofCell Science 113: 1231–1239
Chatton E (1952) Classe des dinoflagellés ou péridiniens. In: Grassé
PP (ed) Traité de Zoologie, Vol. 1, Part 1. Masson, Paris, pp 309–
406
Elbrächter M, Meyer B (2001) Plate pattern variability and plate over-
lap in a clonal culture of the freshwater dinoflagellate Peridiniumumbonatum Stein species complex (Dinophyceae). Neues Jahr-buch feur Geologie und Paleaontologie Abhandlungen 219: 221–
227
Fensome RA, Taylor FJR, Norris G, Sarjeant WAS, Wharton DI,
Williams GL (1993) A Classification of Living and Fossil Dinofla-gellates. Micropaleontology, Special Publication Number 7.
American Museum of Natural History. Sheridan Press, Hanover,
Pennsylvania, 351pp
Figure 6: Light photomicrographs of the Crypthecodinium species: (a) flagellate cell with extended feeding tube (arrow) attached to a
cultured fish cell (fc); (b) cysts. Scale bars = 10µm
���
���
���
���
���
���
���
���
� � � � � ��� !�"#�$�%
&' !(�"��
� ���))���)*�%
Figure 7: Population growth curve of the Crypthecodinium species
when fed cultured fish cells. Cells were enumerated from preserved
samples (1% acidic Lugol’s solution, final concentration) of batch
cultures sampled daily for six days (means ± 1SD, n = 3)

African Journal of Marine Science 2006, 28(2): 185–191 191
Gajadhar AA, Marquardt WC, Hall R, Gunderson J, Ariztia-Carmona
EV, Sogin ML (1991) Ribosomal RNA sequences of Sacrocystismuris, Theileria annulata and Crypthecodinium cohnii reveal
evolutionary relationships among apicomplexans, dinoflagellates,
and ciliates. Molecular and Biochemical Parasitology 45: 147–
154
Javornický P (1962) Two scarcely known genera of the class Dino-
phyceae: Bernardinium Chodat and Crypthecodinium Biecheler.
Preslia 34: 98–113
Kumar S, Tamura K, Nei M (2004) MEGA3: Integrated software for
Molecular Evolutionary Genetics Analysis and sequence align-
ment. Briefings in Bioinformatics 5: 150–163
Lam CMC, New DC, Wong JTY (2001) cAMP in the cell cycle of
the dinoflagellate Crypthecodinium cohnii (Dinophyta). Journal ofPhycology 37: 79–85
Murray S, Jørgensen MF, Ho SYW, Patterson DJ, Jermiin LS
(2005) Improving the analysis of dinoflagellate phylogeny based
on rDNA. Protist 156: 269–286
Parrow MW, Burkholder JM, Deamer NJ, Ramsdell JS (2005)
Contaminant-free cultivation of Pfiesteria shumwayae (Dino-
phyceae) on a fish cell line. Aquatic Microbial Ecology 39: 97–
105
Perret E, Davoust J, Albert M, Besseau L, Soyer-Gobillard MO
(1993) Microtubule organization during the cell cycle of the prim-
itive eukaryote dinoflagellate Crypthecodinium cohnii. Journal ofCell Science 104: 639–651
Saldarriaga JF, Taylor FJR, Cavalier-Smith T, Menden-Deuer S,
Keeling PJ (2004) Molecular data and the evolutionary history of
dinoflagellates. European Journal of Protistology 40: 85–111
Saldarriaga J[F], Taylor FJR, Keeling PJ, Cavalier-Smith T (2001)
Dinoflagellate nuclear SSU rRNA phylogeny suggests multiple
plastid losses and replacements. Journal of Molecular Evolution53: 204–213
Stein F (1883) Der Organismus der Infusionsthiere. 3. Abt. Der
Organismus der Arthrodelen Flagellaten nach eigenen Forschun-
gen in systematischer Reihenfolge bearbeitet. 2. Hälfte. Einleitung
und Erklärung der Abbildungen. W Engelmann, Leipzig, 30pp + 25
Plates
Tuttle RC, Loeblich AR (1975) An optimal growth medium for the
dinoflagellate Crypthecodinium cohnii. Phycologia 14: 1–8
Ucko M, Cohen E, Gordin H, Arad SM (1989) Relationship between
the unicellular red alga Porphyridium sp. and its predator, the
dinoflagellate Gymnodinium sp. Applied and EnvironmentalMicrobiology 55: 2990–2994
Ucko M, Elbrächter M, Schnepf E (1997) A Crypthecodiniumcohnii-like dinoflagellate feeding myzocytotically on the unicellu-
lar red alga Porphyridium sp. European Journal of Phycology 32:
133–140
Woloszyñska J (1925) Przyczynki do znajomści polskich brózdnic
słodkowoddnych. Beiträge zur Kenntnis der Süsswasser-
Dinoflagellaten Polens. Acta Societatis Botanicorum Poloniae 3:
49–64
Manuscript received February 2005; accepted February 2006




















