
Comparison of thiol subproteome of the vent mussel Bathymodiolus azoricus from different...
9
Comparison of thiol subproteome of the vent mussel Bathymodiolus azoricus from different Mid-Atlantic Ridge vent sites Rui Company a, ⁎, Amparo Torreblanca b , Miren Cajaraville c , Maria João Bebianno a , David Sheehan d a CIMA, University of Algarve, Faculty of Marine and Environmental Sciences, Campus de Gambelas, 8005‐139 Faro, Portugal b Department of Functional Biology, University of Valencia, 46100 Burjassot, Valencia, Spain c Laboratory of Cell Biology and Histology, Department of Zoology and Cell Biology, University of the Basque Country, P.O Box 644, E-48080 Bilbao, Basque Country, Spain d Proteomics Research Group, Dept. of Biochemistry, University College Cork, Cork, Ireland HIGHLIGHTS ► Thiol proteins play major roles in stress defence and are susceptible to oxidation. ► Activated thiol sepharose was used to isolate thiol proteins in vent mussels. ► Thiol proteins in mussels from high metal levels sites were significantly less abundant. ► Differently expressed proteins reflect adaptation mechanisms to extreme environments. ► Mussels have different susceptibilities to oxidative stress depending on the vent site. GRAPHICAL ABSTRACT abstract article info Article history: Received 28 May 2012 Received in revised form 18 July 2012 Accepted 27 July 2012 Available online 7 September 2012 Keywords: Bathymodiolus azoricus Hydrothermal vents Thiol sub-proteome Activated thiol sepharose Oxidative stress Deep-sea hydrothermal mussels Bathymodiolus azoricus live in the mixing zone where hydrothermal fluid mixes with bottom seawater, creating large gradients in the environmental conditions and are one of the most studied hydrothermal species as a model of adaptation to extreme conditions. Thiol proteins, i.e. proteins containing a thiol or sulfhydryl group (\SH) play major roles in intracellular stress defense against reactive oxygen species (ROS) and are especially susceptible to oxidation. However, they are not particularly abundant, representing a small percentage of proteins in the total proteome and therefore are difficult to study by proteomic approaches. Activated thiol sepharose (ATS) was used for the rapid and quantitative selection of proteins comprising thiol- or disulfide-containing subproteomes. This study aims to isolate thiol-containing proteins from the gills of B. azoricus collected in distinct hydrothermal vents and to study the thiol-containing subproteome as a function of site-specific susceptibility to ROS. Results show that ATS is a powerful tool to isolate the thiol-containing sub-proteome and differently-expressed protein spots showed significant differences among the three vent sites, supporting previous findings that specific environmental conditions are crucial for ROS formation and that B. azoricus have different susceptibilities to oxidative stress depending on the vent site they inhabit. © 2012 Elsevier B.V. All rights reserved. 1. Introduction Although 35 years have passed since the discovery of deep-sea hy- drothermal vents, scientists are still trying to understand and explain the adaptation mechanisms of vent species to one of the most extreme, toxic and stressful environments recognized on earth. Remarkably, Science of the Total Environment 437 (2012) 413–421 ⁎ Corresponding author. Tel.: +351 289 800 900; fax: +351 289 800 069. E-mail address: [email protected] (R. Company). 0048-9697/$ – see front matter © 2012 Elsevier B.V. All rights reserved. http://dx.doi.org/10.1016/j.scitotenv.2012.07.089 Contents lists available at SciVerse ScienceDirect Science of the Total Environment journal homepage: www.elsevier.com/locate/scitotenv
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Comparison of thiol subproteome of the vent mussel Bathymodiolus azoricus from different...

Science of the Total Environment 437 (2012) 413–421
Contents lists available at SciVerse ScienceDirect
Science of the Total Environment
j ourna l homepage: www.e lsev ie r .com/ locate /sc i totenv
Comparison of thiol subproteome of the vent mussel Bathymodiolus azoricus fromdifferent Mid-Atlantic Ridge vent sites
Rui Company a,⁎, Amparo Torreblanca b, Miren Cajaraville c, Maria João Bebianno a, David Sheehan d
a CIMA, University of Algarve, Faculty of Marine and Environmental Sciences, Campus de Gambelas, 8005‐139 Faro, Portugalb Department of Functional Biology, University of Valencia, 46100 Burjassot, Valencia, Spainc Laboratory of Cell Biology and Histology, Department of Zoology and Cell Biology, University of the Basque Country, P.O Box 644, E-48080 Bilbao, Basque Country, Spaind Proteomics Research Group, Dept. of Biochemistry, University College Cork, Cork, Ireland
H I G H L I G H T S G R A P H I C A L A B S T R A C T
► Thiol proteins play major roles instress defence and are susceptible tooxidation.
► Activated thiol sepharose was used toisolate thiol proteins in vent mussels.
► Thiol proteins in mussels from highmetal levels sites were significantlyless abundant.
► Differently expressed proteins reflectadaptation mechanisms to extremeenvironments.
► Mussels have different susceptibilitiesto oxidative stress depending on thevent site.
⁎ Corresponding author. Tel.: +351 289 800 900; faxE-mail address: [email protected] (R. Company).
0048-9697/$ – see front matter © 2012 Elsevier B.V. Allhttp://dx.doi.org/10.1016/j.scitotenv.2012.07.089
a b s t r a c t
a r t i c l e i n f oArticle history:Received 28 May 2012Received in revised form 18 July 2012Accepted 27 July 2012Available online 7 September 2012
Keywords:Bathymodiolus azoricusHydrothermal ventsThiol sub-proteomeActivated thiol sepharoseOxidative stress
Deep-sea hydrothermal mussels Bathymodiolus azoricus live in themixing zonewhere hydrothermal fluidmixeswith bottom seawater, creating large gradients in the environmental conditions and are one of the most studiedhydrothermal species as a model of adaptation to extreme conditions. Thiol proteins, i.e. proteins containing athiol or sulfhydryl group (\SH) play major roles in intracellular stress defense against reactive oxygen species(ROS) and are especially susceptible to oxidation. However, they are not particularly abundant, representing asmall percentage of proteins in the total proteome and therefore are difficult to study by proteomic approaches.Activated thiol sepharose (ATS)was used for the rapid and quantitative selection of proteins comprising thiol- ordisulfide-containing subproteomes. This study aims to isolate thiol-containing proteins from the gills ofB. azoricus collected in distinct hydrothermal vents and to study the thiol-containing subproteome as a functionof site-specific susceptibility to ROS. Results show that ATS is a powerful tool to isolate the thiol-containingsub-proteome and differently-expressed protein spots showed significant differences among the three ventsites, supporting previous findings that specific environmental conditions are crucial for ROS formation andthat B. azoricus have different susceptibilities to oxidative stress depending on the vent site they inhabit.
© 2012 Elsevier B.V. All rights reserved.
: +351 289 800 069.
rights reserved.
1. Introduction
Although 35 years have passed since the discovery of deep-sea hy-drothermal vents, scientists are still trying to understand and explainthe adaptation mechanisms of vent species to one of the most extreme,toxic and stressful environments recognized on earth. Remarkably,

414 R. Company et al. / Science of the Total Environment 437 (2012) 413–421
hydrothermal vents support ecosystems with enormous biomass andproductivity compared with those observed elsewhere in the deepocean. Deep-sea hydrothermal communities live in the interfacialzone or mixing zone where hydrothermal fluid mixes turbulentlywith bottom seawater, creating large gradients in the environmentalconditions (Tunnicliffe, 1991; Childress and Fisher, 1992; Le Bris et al.,2001). The ventmussel Bathymodiolus azoricus is an endemic and dom-inant species at North Atlantic vent fields, in theMid-Atlantic Ridge andperhaps one of themost studied hydrothermal species as amodel of ad-aptation to extreme conditions. B. azoricus is themost abundant speciesin Menez Gwen (37°51′N, 31°31′W, 850 m), Lucky Strike (37°17′N,32°16′W, 1700 m) and Rainbow (36°13′N, 33°54′W, 2300 m) ventsites where these mussels experience a different hydrostatic pressure,temperature and composition of hydrothermal fluid including pH,dissolved oxygen, methane and sulfide concentrations (Charlou et al.,2000; Douville et al., 2002). These three vent sites have been extensive-ly characterized and the differences in fluid composition are reported inseveral studies (Charlou et al., 2000; Desbruyeres et al., 2001; Sarradinet al., 1999). Another important feature of hydrothermal vent systemsis the presence of high levels of toxic chemicals. These include high con-centrations of metals like Cd, Cu, Fe, Hg, Mn and Zn originating from thedischarge of hydrogen sulfide-rich water (Cosson et al., 2008), petro-leum hydrocarbons (Simoneit et al., 2004; Serafimet al., 2008) and nat-ural radionuclides such as 238U, 235U, 234U, 210Po and 210Pb(Charmasson et al., 2009; 2011). The mixing zone where vent organ-isms live is therefore a highly reactive area that due to the presenceof reduced chemical species (H2S, CH4) and toxic compounds in thevent fluid can lead to the formation of reactive oxygen species (ROS),that produce oxidative stress in organisms i.e. a disruption of thebalance between the levels of oxidants (ROS) and antioxidants(Lesser, 2006; Winterbourn and Hampton, 2008; Sheehan andMcDonagh, 2008). To avoid ROS-induced damage, organisms possessboth enzymatic and non-enzymatic antioxidant defenses. Althoughthe amount and specific types of ROS produced in the hydrothermalmixing zone capable of affecting vent fauna remain poorly-understood,many studies have highlighted ROS-induced damage and mechanismsto counteract oxidative stress in hydrothermal organisms (Blum andFridovich, 1984; Dixon et al., 2002; Pruski and Dixon, 2003; Geret et al.,2004; Bebianno et al., 2005; Company et al., 2006a,b; Marie et al.,2006; Gonzalez-Rey et al., 2007; Marchand et al., 2009). These studiesare based mostly on the specific activities of antioxidant enzymes(such as superoxide dismutase, catalase and glutathione peroxidase) oroxidative injury to cell components such as DNA damage and lipidperoxidation. However, recently proteomics has proven to be a robusttool to study oxidative stress (Sheehan and McDonagh, 2008) and theterm “redox-proteomics” was coined to describe the multiple applica-tions of proteomics for the discovery of oxidative posttranslationalmodifications of proteins. It is well-recognized that the main targets ofROS-induced damage are proteins, which absorb approximately 70% ofROS. Among those are thiol proteins, i.e. proteins containing a thiol orsulfhydryl group (\SH) that are especially susceptible to oxidation.Thiol-containing proteins include metallothioneins (MT), thioredoxin(Trx), glutaredoxin (Grx) and peroxiredoxin (Prx) which play majorroles in intracellular stress defense. Under oxidative stress conditions,oxidation of cysteine sulfhydryl (\SH) groups can occur formingsulfenic (\SOH), sulfinic (\SO2H), and sulfonic/cysteic (\SO3H) acids,interchain/intrachain disulfide bridges (\S\S\), or thiolsulfinates(Hu et al., 2010a). Although thiol-containing proteins contribute signifi-cantly to antioxidant defenses they are not particularly abundant,representing only a small percentage of proteins present in the totalproteome and therefore are difficult to identify and study by proteomicapproaches. To overcome this limitation, methods have been developedto enrich for thiol-containing proteins in oxidative stress studies (Baty etal., 2002; Lind et al., 2002; Eaton et al., 2003; Liu et al., 2005). Recently amethod using activated thiol sepharose (ATS) beads was described toselect for thiol-containing proteins in the bacterium Escherichia coli,
fungus Trichoderma harzianum, and the bivalve Mytilus edulis (Hu et al.,2010a,b,c). This method was successfully applied to study oxidativestress in M. edulis exposed to gold nanoparticles (Tedesco et al., 2010).ATS is a commercially-available, thiol-specific resin with an activateddisulfide structure that reacts efficiently with \SH groups to form acovalentmixeddisulfide linkage. Thus, ATS facilitates rapid andquantita-tive selection of proteins comprising thiol- or disulfide-containingsubproteomes relevant to key biological processes (e.g., translation,metabolism, oxidative stress). The present study aimed to isolatethiol-containing proteins from the gills of B. azoricus collected in distincthydrothermal vents using ATS and to study the thiol-containingsubproteome as a function of site-specific susceptibility to ROS inB. azoricus.
2. Materials and methods
2.1. Animal collection
Deep sea hydrothermal mussels B. azoricus (6.08±0.58 cm shelllength) were collected from three hydrothermal vent sites locatedin the Mid-Atlantic Ridge: Menez-Gwen (37°51′N and 31°31.2′W,840 m), Lucky Strike (37°17′30″N and 32°165.3′W, 1700 m) andRainbow (36°13′N and 33°54.1′W, 2400 m) in July 2001 using aremote operated vehicle during the EU-funded VENTOX Project(Sarradin et al., 2001). Gills were dissected immediately aftercollection, frozen in liquid nitrogen and stored at −80 °C untilfurther analysis.
2.2. Isolation of thiol proteins
For isolation of thiol-containing proteins, 3 pools of gills (of 3 organ-isms each) of mussels from each vent site were homogenized in 10 mMTris/HCl, pH 7.2, 500 mM sucrose, 1 mM ethylenediaminetetraaceticacid (EDTA), 1 mM phenylmethylsulfonyl fluoride, centrifuged at20,000×g and stored at −70 °C. Protein content was calculated by themethod of Bradford (1976) with bovine serum albumin (BSA) as astandard. Protein estimation was performed in triplicate and thepercentage of proteins retained on ATS was calculated from a knownamount of total protein applied. Isolation of thiol-containing proteinswas performed as described by Hu et al. (2010b). Briefly, protein extractsamples (2 mg total protein) were diluted with an 8000 mM urea stockto a final urea concentration of 5000 mM and incubated for 10 min atroom temperature to unfold the proteins. ATS (20 mg unswollenresin) and 200 μl of binding buffer solution (100 mM Tris–HCl[pH 7.5], 500 mM NaCl, and 1 mM EDTA) were then added, followedby incubation on ice for 1.5 h with gentle shaking every 15 min toensure that thiol-containing proteins completely bound to the beads.During this time, ATS beads swelled, efficiently absorbing the proteinsolution. Samples were centrifuged (11,000×g, 3 min) and thenwashed with 500 μl of binding buffer eight times, centrifuged(11,000×g, 3 min) after each wash, and then supernatants werediscarded. Elution buffer (200 μl, binding buffer containing 25 mMDTT) was added to the samples after the final washing step, andbeads were incubated for 1 h with gentle shaking every 15 min torelease thiol-containing proteins. The beads were centrifuged briefly(11,000×g, 3 min), and supernatantwas collected. All buffers used dur-ing this procedure were previously filtered using 0.2-μm nitrocellulosemembranes (Protran, Schleicher & Schuell, Dassel, Germany) anddegassed to minimize air oxidation.
2.3. Sample preparation and 2-DE separation
Thiol-containing proteins of 9 technical replicates for each ventsite were further purified to remove non-protein contaminants byprecipitation and centrifugation, first with 10% trichloroacetic acidand then with acetone. The resulting pellet was resuspended in

Fig. 2. Quantification of thiol-containing protein spots from 2DE in the gills of B. azoricusfrom Menez-Gwen, Lucky Strike and Rainbow hydrothermal vent sites. Data arepresented as mean±SD and were performed in triplicate. Different letters representstatistical differences (pb0.05).
Table 1Thiol-containing proteins retained on activated thiol sepharose in B. azoricus fromthree replicate samples from Menez-Gwen, Lucky Strike and Rainbow vent sites.
Rep. 1 Rep. 2 Rep. 3 Mean (±SD)
Menez-GwenTotal protein loaded (μg) 2000 2000 2000 2000\SH-containing proteins (μg) 35.8 31.0 41.6 36.1±5.3% retained 1.79 1.55 2.08 1.81±0.27
Lucky StrikeTotal protein loaded (μg) 2000 2000 2000 2000\SH-containing proteins (μg) 32.0 28.2 35.0 31.7±3.4% retained 1.60 1.41 1.75 1.59±0.17
RainbowTotal protein loaded (μg) 2000 2000 2000 2000\SH-containing proteins (μg) 32.0 34.8 26.0 30.9±4.5% retained 1.60 1.74 1.30 1.55±0.22
415R. Company et al. / Science of the Total Environment 437 (2012) 413–421
rehydration buffer containing 5 M urea, 2 M thiourea, 2% Chaps, 4%carrier ampholyte (Pharmalyte 3–10, GE Healthcare), 1% DeStreakreagent (GE Healthcare), and a trace amount of bromophenol blue.Final volumes of 125 μl were loaded on 7-cm pH 3 to 10 nonlinearimmobilized pH gradient (IPG) strips (Bio-Rad, Hercules, CA, USA)and rehydrated overnight for at least 15 h. IPG strips were focusedon a Protean isoelectric focusing (IEF) cell (Bio-Rad) with linearvoltage increases. Following IEF, strips were equilibrated for 20 minin equilibration buffer (6 M urea, 0.375 M Tris [pH 8.8], 2% SDS, and20% glycerol) containing 2% DTT and then for 20 min in equilibrationbuffer containing 2.5% iodoacetamide. Equilibrated strips wereembedded in molten agarose (0.5%) containing trace bromophenolblue atop 12% SDS–polyacrylamide gels and were electrophoresedat a constant voltage (150 V) at 4 °C using an Atto AE-6450 miniPAGE system until the dye front reached the end of the gel. Afterseparation, gels were silver stained by the method of Rabilloud(1992) followed by densitometry scanning.
2.4. Gel scanning and image analysis
Stained gels were scanned with a Bio-Rad densitometer scanner.Differences in protein expression levels among the samples wereanalyzed using PDQuest 2-D analysis software version 8.0.1 (Bio-Rad).Each gel was analyzed for spot background subtraction, spot detection,matching and acquisition of spot coordinates, etc. For comparisonbetween-gels, a set of spot-generation conditions (weakest spot,smallest spot, size of the largest spot, and a selected region ofthe background) were used. The comparison of B. azoricus thiol-containing subproteomes between vent sites was performed be-tween Menez-Gwen (control site) and Lucky Strike and Rainbow.Up- and down-regulated spots were selected according to a
Fig. 1. Representative images obtained for the gills of B. azoricus from (A) Menez-Gwen, (B)marker used is with molecular weight (M.W.) which covers from 14.4 kDa (bottom) to 66
two-fold relative increase or decrease of intensity with respectto controls.
2.5. Statistical analysis
Thiol-containing proteins were expressed as mean±standarddeviation (SD). The data were previously tested for normality andhomogeneity and analyzed by analysis of variance (ANOVA) to deter-mine significant statistical differences of thiol-containing proteinsbetween hydrothermal sites. Post hoc comparison tests for meanswere made using the Tukey's test. The level of significance was setat 0.05. Principal component analysis (PCA) was used in the ordina-tion method to compare differently expressed proteins in each site.Statistical analysis was carried out with Statistica 5.1 and SIMCA-P10 for windows.
3. Results
Thiol-containing proteins from the gills of B. azoricuswere isolatedfrom the total protein extract using ATS. The quantity (by weight) ofthiol-containing proteins retained on ATS was estimated in musselsfrom the three hydrothermal vent sites (Table 1). These resultsshow generally similar percentages of proteins frommussels sampledfrom the three sites.
2-DE separations of thiol-containing proteins in the gills ofB. azoricus were analyzed and representative images are shown inFig. 1. From the gel image analysis the number of detected proteins
Lucky Strike and (C) Rainbow. Isoelectric point (pI) with range of pH units from 3 to 10;kDa (top). Technical replicates were performed at least in triplicate.

Fig. 3. Differently expressed thiol-containing protein spots in the gills of B. azoricus between (A) Menez-Gwen and Lucky Strike (proteins exclusive of Lucky Strike are representedin red squares); and (B) Menez-Gwen and Rainbow (proteins exclusive of Rainbow are represented in red squares).
416 R. Company et al. / Science of the Total Environment 437 (2012) 413–421
varied significantly between sites. The average of thiol-containingprotein spots detected in the replicate gels of B. azoricus fromMenez-Gwen (111±4) was significantly higher than those fromLucky Strike (68±1) and Rainbow (65±3) (pb0.05) (Fig. 2).
PDQuest analysis revealed that 41 protein spots were statisticallydifferent between the gills of the mussels from Menez-Gwen andLucky Strike, 37 of which were only present in Menez-Gwen mussels,while only 3 were exclusive to Lucky Strike (spots ID 4607, 7203 and8503) (Fig. 3A). In a similar analysis, 35 protein spots were statisticallydifferent between Menez-Gwen and Rainbow, 3 of which wereexclusive to Rainbow mussels (spots ID 3510, 4212 and 8503)(Fig. 3B). There is one common spot between Lucky Strike andRainbow sites (Spot ID 8503). Fig. 4 shows that the up-regulated
and down-regulated protein spots in Lucky Strike (Fig. 4A) andRainbow (Fig. 4B) are compared to mussels from Menez-Gwen(control). The Venn diagram shows the total thiol-containingproteins in B. azoricus from three hydrothermal vent sites, thenumber of proteins exclusive from each site and the commonspots between sites (Fig. 5).
PCA was applied to statistically-different spots present in at least80% of the proteomes of the three vent sites (Fig. 6). Together, PC1and PC2 explain 72% of the data variability and there is highreproducibility between replicate samples within each group. Thefirst component (PC1), explaining 53% of data variability, shows aclear separation of mussels from the deepest vent site (Rainbow)plotted on the negative side of PC1 and mussels from the other two

Fig. 4. Thiol-containing protein spots in the gills of B. azoricus from Lucky Strike that are up-regulated (green triangles) and down-regulated (red circles) (A) and Rainbow that areup-regulated (green triangles) and down-regulated (red circles) compared to Menez-Gwen (B).
417R. Company et al. / Science of the Total Environment 437 (2012) 413–421
vent sites (Menez-Gwen and Lucky Strike) plotted in the positiveside. The second component, (PC2), explains 19% of variance, with aseparation of vent sites Menez-Gwen from Lucky Strike (Fig. 6). Inorder to better understand the influence of metallic contaminantsand endogenous defenses in B. azoricus on the thiol-containingproteome differences between vent sites, another PCA was performed(Fig. 7). This took into account the metal concentrations (Ag, Cd, Cu,Fe, Mn and Zn) and responses of several antioxidant defensebiomarkers (superoxide dismutase (SOD), catalase, total (GPx-T)and Se-dependent glutathione peroxidase (GPx-Se) and lipid
peroxidation) induced by ROS and previously determined in thegills of comparable organisms (Bebianno et al., 2005). The sampledispersion is very similar to that observed in the previous PCAalthough, in this analysis, samples from Menez-Gwen and LuckyStrike are inversely plotted in the second axis (Fig. 7A). Neverthe-less, separation between sites remains clear and similar. Musselsfrom Rainbow are associated with higher levels of lipid peroxida-tion (LPO), accumulated metals (Ag, Fe and Mg) and catalase(CAT), while mussels from Lucky Strike are associated with highantioxidant enzyme activities (SOD, GPx-T, GPx-Se) and some

Fig. 5. Overlapping Venn diagram of the total thiol-containing proteins detected (numbersoutside the circles) in B. azoricus from Menez-Gwen (MG), Lucky Strike (LS) and Rainbow(Rb) hydrothermal vent sites.
418 R. Company et al. / Science of the Total Environment 437 (2012) 413–421
accumulated metals (Cd and Zn). Samples from the shallowest ventsite Menez-Gwen were only associated with higher Cu levels(Fig. 7B).
4. Discussion
Thiol-containing proteins have important key roles in biologyespecially in antioxidant defense and buffering of transient increasesof ROS (Eaton, 2006; Hansen et al., 2009). The knowledge of antioxi-dant protection in hydrothermal vent organisms is vital to under-standing adaptation of these species to one of the most extreme andpotentially toxic environments. B. azoricus live in the mixing zonewhich is thought to be particularly prone to production of ROS. Inthis area, high abundance of hydrogen sulfide (H2S) and oxygen
Fig. 6. Principal component analysis (PCA) of statistically different thiol-containing proteinshowing the data scores labeled as sites.
leads to oxidation of H2S in the seawater with the production ofboth oxygen radicals (O2•−, H2O2, HO•) and sulfur-centered radicals(HS•, HSO2•, HSO4•) (Tapley et al., 1999). Moreover, the presence ofa high concentration of metals in these areas increases the productionof intracellular hydroxyl radical HO• via the Haber–Weiss reaction.Therefore, it is not surprising that B. azoricus possesses antioxidantenzymatic defenses that are site-specific, i.e. hydrothermal musselscan modulate the specific activity of antioxidant enzymes to betteradapt to their specific habitat conditions (Bebianno et al., 2005).Although several studies have reported on oxidative responses ofB. azoricus to both natural and laboratory-induced stressors, thepresent work constitutes the first analysis of thiol-containing proteinsin oxidative stress responses in this species. The method applied toisolate thiol-containing proteins from gills of B. azoricus is relativelynew and, although still not commonly used, is very promising andhas proven successful in several studies (Hu et al., 2010a,b,c;Tedesco et al., 2010). There is a remarkable consistency in thepercentage of thiol-containing proteins retained by the ATS betweenmussels from different vent sites, ranging from 1.30 to 2.08% of thetotal protein content. This percentage was very similar to thosereported by Hu et al. (2010a) for other species, such as E. coli(2.52±0.41), T. harzianum (1.4±0.05) and M. edulis (1.4±0.69).This strongly suggests that organisms with different biologicalorganization levels and exposed to different environments, containsimilar percentages of thiol proteins and confirms ATS as a robusttool for quantitatively thiol-containing proteins while simultaneouslypreventing their oxidation during experimental manipulation (Hu etal., 2010a).
Although mussels from different hydrothermal vents had similarabundance of thiol-containing proteins, 2-DE analysis of the thiol-containing sub-proteome of B. azoricus revealed that the number ofthiol-containing protein spots detected in Menez-Gwen was two-fold higher than that observed in mussels from Lucky Strike andRainbow. This may be consistent with direct oxidation of thiols byROS induced by the extreme environmental conditions at thesesites. B. azoricus was selected from three different vent sites toallow comparison of protein expression signatures in musselsexposed to distinct natural environmental conditions. Based on
spots in the gills of B. azoricus from Menez Gwen, Lucky Strike and Rainbow vent sites
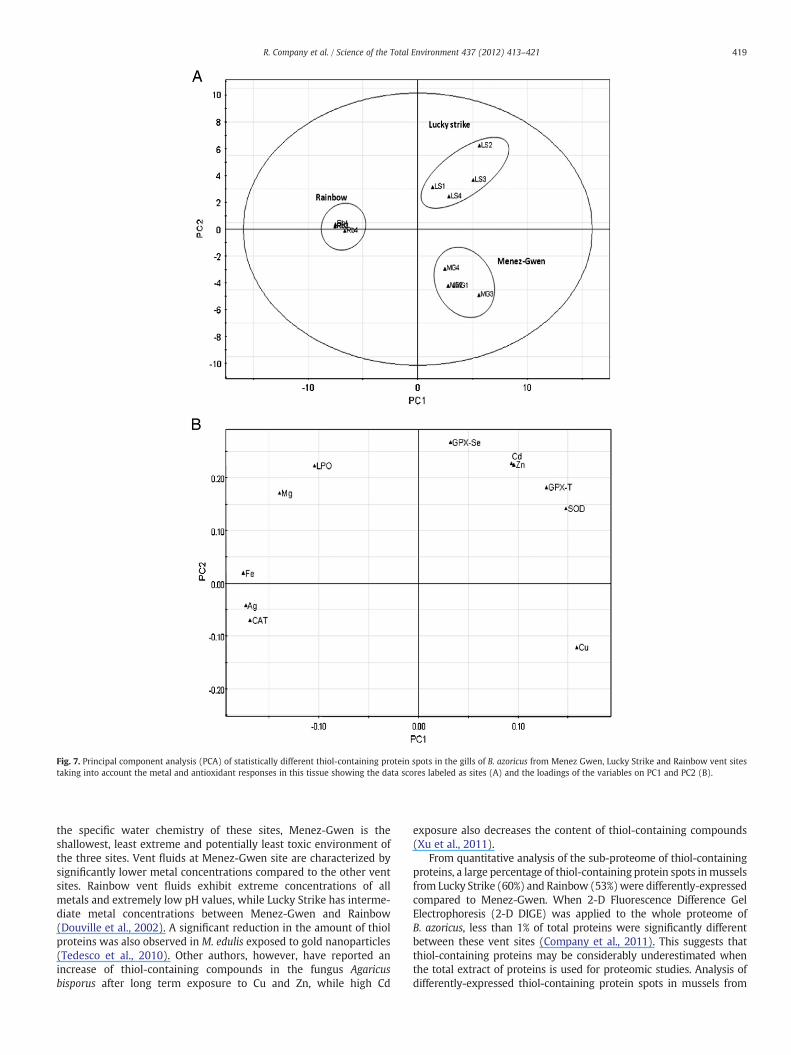
Fig. 7. Principal component analysis (PCA) of statistically different thiol-containing protein spots in the gills of B. azoricus from Menez Gwen, Lucky Strike and Rainbow vent sitestaking into account the metal and antioxidant responses in this tissue showing the data scores labeled as sites (A) and the loadings of the variables on PC1 and PC2 (B).
419R. Company et al. / Science of the Total Environment 437 (2012) 413–421
the specific water chemistry of these sites, Menez-Gwen is theshallowest, least extreme and potentially least toxic environment ofthe three sites. Vent fluids at Menez-Gwen site are characterized bysignificantly lower metal concentrations compared to the other ventsites. Rainbow vent fluids exhibit extreme concentrations of allmetals and extremely low pH values, while Lucky Strike has interme-diate metal concentrations between Menez-Gwen and Rainbow(Douville et al., 2002). A significant reduction in the amount of thiolproteins was also observed in M. edulis exposed to gold nanoparticles(Tedesco et al., 2010). Other authors, however, have reported anincrease of thiol-containing compounds in the fungus Agaricusbisporus after long term exposure to Cu and Zn, while high Cd
exposure also decreases the content of thiol-containing compounds(Xu et al., 2011).
From quantitative analysis of the sub-proteome of thiol-containingproteins, a large percentage of thiol-containing protein spots inmusselsfrom Lucky Strike (60%) and Rainbow (53%)were differently-expressedcompared to Menez-Gwen. When 2-D Fluorescence Difference GelElectrophoresis (2-D DIGE) was applied to the whole proteome ofB. azoricus, less than 1% of total proteins were significantly differentbetween these vent sites (Company et al., 2011). This suggests thatthiol-containing proteins may be considerably underestimated whenthe total extract of proteins is used for proteomic studies. Analysis ofdifferently-expressed thiol-containing protein spots in mussels from

420 R. Company et al. / Science of the Total Environment 437 (2012) 413–421
Lucky Strike and Rainbow shows that very few proteins are identical,i.e., the protein spots that are up-regulated inmussels from Lucky Strikedo not coincide with those from Rainbow. This suggests thatthe thiol-containing sub-proteome of B. azoricus from these ventsites is adapted to different environmental conditions. Based on thedifferently-expressed thiol-containing proteins, PCA analysis clearlygrouped the samples by site. Similar findings were obtained for thetotal proteome of vent mussels (Company et al., 2011). The spotpatterns found in the present study are consistent with the presenceof differing amounts and production pathways of ROS in hydrothermalvents. Oxidation of thiol-containing proteins seems less likely to occurin mussels from Menez-Gwen, the shallowest and least toxic vent site,compared to mussels from Lucky Strike and Rainbow which areespecially prone to ROS formation due to their water chemistry andhigh metal concentrations. SH-containing compounds are capable ofbinding metal ions, and also of scavenging ROS (Romero-Isart andVašák, 2002). PCA analysis of thiol-containing sub-proteomes includingdata for oxidative stress biomarkers in B. azoricus confirmed thatthe oxidative status of vent mussels relies on a balance betweenpro-oxidants, ROS production and antioxidant defenses. This is in agree-mentwith previousfindingswhere total oxyradical scavenging capacity(TOSC) of B. azoricus toward peroxyl (ROO•), hydroxyl radicals (O2•−)and peroxynitrite (HOONO) was significantly higher in mussels fromMenez-Gwen compared to those from Lucky Strike and Rainbow(Bebianno et al., 2005). This suggested an overall impairment inneutralizing ROS in the most toxic MAR vents. The present workcorroborates the usefulness of the thiol-containing sub-proteome fordetection of protein oxidation in response to ROS formation.
5. Conclusion
ATS beads were effective in selecting and isolating thiol-containingproteins of the hydrothermal ventmussel B. azoricus as previously dem-onstrated for coastal mussels. The sub-proteome of thiol-containingproteins in B. azoricus showed considerably less protein spots comparedto the total proteome obtained by DIGE, as expected. However, thepercentage of differently-expressed proteins was considerably higherin the thiol-containing sub-proteome, establishing this selection meth-od as an informative tool in redox proteomics. The quantitative study(number of spots and spot intensity) of differently-expressed proteinspots showed significant differences among the three vent sites,supporting previous findings that specific environmental conditionsare crucial for ROS formation and that B. azoricus have differingsusceptibilities to oxidative stress depending on the vent site theyinhabit.
Acknowledgments
The authors are grateful to the N/O L'Atalante and deep submersibleVICTOR6000 crew and to P.M. Sarradin and D. Dixon, scientific leadersof the ATOS cruise (Ventox Project). This study was partially fundedby the 5th PCRD Ventox project (EVK3CT1999-00003). R. Companywas supported by an FCT Post-doc fellowship (SFRH/BPD/26399).Authors are grateful to Dr. S. Tedesco and Dr.W. Hu from the UniversityCollege Cork for laboratory technical support.
References
Baty JW, Hampton MB, Winterbourn CC. Detection of oxidant sensitive thiol proteins byfluorescence labeling and two-dimensional electrophoresis. Proteomics 2002;2:1261–6.
Bebianno MJ, Company R, Serafim A, Camus L, Cosson RP, Fiala-Medoni A. Antioxidantsystems and lipid peroxidation in Bathymodiolus azoricus from Mid-Atlantic Ridgehydrothermal vent fields. Aquat Toxicol 2005;75:354–73.
Blum J, Fridovich I. Enzymatic defenses against oxygen toxicity in the hydrothermalvents animals Riftia pachyptila and Calyptogena magnifica. Arch Biochem Biophys1984;228:617–20.
BradfordM. A rapid and sensitive method for the quantitation of microgram quantities ofprotein utilizing the principle of protein-dye binding. Anal Biochem 1976;72:248–54.
Charlou JL, Donval JP, Douville E, Jean Baptiste P, Radford‐Knoery J, Fouquet Y, et al.Compared geochemical signatures and the evolution of Menez Gwen (37°50′N)and Lucky Strike (37°17′N) hydrothermal fluids, south of the Azores Triple Junctionon the Mid-Atlantic Ridge. Chem Geol 2000;171:49–75.
Charmasson S, Sarradin P-M, Le Faouder A, Agarande M, Loyen J, Desbruyères D. High levelsof natural radioactivity in biota from deep-sea hydrothermal vents: a preliminarycommunication. J Environ Radioact 2009;100:522–6.
Charmasson S, Le Faouder A, Loyen J, Cosson RP, Sarradin P-M. 210Po and 210Pb in thetissues of the deep-sea hydrothermal vent mussel Bathymodiolus azoricus fromthe Menez Gwen field (Mid-Atlantic Ridge). Sci Total Environ 2011;409(4):771–7.
Childress JJ, Fisher CR. The biology of hydrothermal vent animals: physiology, biochemistry,and autotrophic symbioses. Oceanogr Mar Biol Annu Rev 1992;30:337–441.
Company R, Serafim A, Cosson R, Camus L, Shillito B, Fiala‐Médioni A, et al. Effect ofcadmium on antioxidant responses and susceptibility to oxidative stress in thehydrothermal vent mussel Bathymodiolus azoricus. Mar Biol 2006a;148(4):817–25.
Company R, SerafimA, Cosson R, Fiala-Médioni A, Dixon D, BebiannoMJ. Temporal variationin the antioxidant defence system and lipid peroxidation in the gills and mantle ofhydrothermal vent mussel Bathymodiolus azoricus. Deep Sea Res Part 1 Oceanogr ResPap 2006b;53:1101–16.
Company R, Antúnez O, Bebianno MJ, Cajaraville MP, Torreblanca A. 2-D difference gel elec-trophoresis approach to assess protein expression profiles in Bathymodiolus azoricusfrom Mid-Atlantic Ridge hydrothermal vents. J Proteomics 2011;74:2909–19.
Cosson RP, Thiebaut E, Company R, Castrec-Rouelle M, Colaco A, Martins I, et al. Spatialvariation of metal bioaccumulation in the hydrothermal vent mussel Bathymodiolusazoricus. Mar Environ Res 2008;65:405–15.
Desbruyeres D, Biscoito M, Caprais JC, Colaco A, Comtet T, Crassous P, et al. Variations indeep-sea hydrothermal vent communities on the Mid-Atlantic Ridge near theAzores plateau. Deep Sea Res Part 1 Oceanogr Res Pap 2001;48:1325–46.
Dixon DR, Dixon LRJ, Shillito B, Gwynn JP. Background and induced levels of DNAdamage in pacific deep-sea vent polychaetes: the case for avoidance. Cah BiolMar 2002;43:333–6.
Douville E, Charlou JL, Oelkers EH, Bienvenu P, Jove Colon CF, Donval JP, et al. The rainbowvent fluids (36°14′N, MAR): the influence of ultramafic rocks and phase separationon trace metal content in Mid-Atlantic Ridge hydrothermal fluids. Chem Geol2002;184:37–48.
Eaton P. Protein thiol oxidation in health and disease: techniques for measuringdisulfides and related modifications in complex protein mixtures. Free Radic BiolMed 2006;40:1889–99.
Eaton P, Jones ME, McGregor E, Dunn MM, Leeds N, Byers HL, et al. Reversiblecysteine-targeted oxidation of proteins during renal oxidative stress. J Am SocNephrol 2003;14:S290–6.
Geret F, Manduzio H, Company R, Leboulenger F, Bebianno MJ, Danger JM. Molecularcloning of superoxide dismutase (Cu/Zn-SOD) from aquatic mollusks. Mar EnvironRes 2004;58:619–23.
Gonzalez-Rey M, Serafim A, Company R, Bebianno MJ. Adaptation to metal toxicity:a comparison of hydrothermal vent and coastal shrimps. Mar Ecol 2007;28:100–7.
Hansen RE, Roth D, Winther JR. Quantifying the global cellular thioldisulfide status.Proc Natl Acad Sci U S A 2009;106:422–7.
Hu W, Tedesco S, Faedda R, Petrone G, Cacciola SO, O'Keefe A, et al. Covalent selection ofthe thiol proteome on activated thiol sepharose: a robust tool for redox proteomics.Talanta 2010a;80:1569–75.
Hu W, Tedesco S, McDonagh B, Bárcena JA, Keane C, Sheehan D. Selection of thiol- anddisulfide-containing proteins of Escherichia coli on activated thiol-sepharose. AnalBiochem 2010b;398:245–53.
Hu W, Tedesco S, McDonagh B, Sheehan D. Shotgun redox proteomics in sub-proteomestrapped on functionalised beads: identification of proteins targeted by oxidativestress. Mar Environ Res 2010c;69:S25–7.
Le Bris N, Sarradin PM, Pennec S. A new deep-sea probe for in situ pH measurement inthe environment of hydrothermal vent biological communities. Deep Sea Res Part1 Oceanogr Res Pap 2001;48:1941–51.
Lesser MP. Oxidative stress in marine environments: biochemistry and physiologicalecology. Annu Rev Physiol 2006;68:253–78.
Lind C, Gerdes R, Hamnell Y, Schuppe-Koistinen I, von Lowenhielm HB, Holmgren A,et al. Identification of S-glutathionylated cellular proteins during oxidative stressand constitutive metabolism by affinity purification and proteomic analysis. ArchBiochem Biophys 2002;406:229–40.
Liu T, Qian WJ, Chen WN, Jacobs JM, Moore RJ, Anderson DJ, et al. Improved proteomecoverage by using high efficiency cysteinyl-peptide enrichment: the humanmammaryepithelial cell proteome. Proteomics 2005;5:1263–73.
Marchand J, Leignel V, Moreau B, Chénais B. Characterization and sequence analysis ofmanganese superoxide dismutases from Brachyura (Crustacea: Decapoda): hydro-thermal Bythograeidae versus littoral crabs. Comp Biochem Physiol B 2009;153:191–9.
Marie B, Genard B, Rees JF, Zal F. Effect of ambient oxygen concentration on activities ofenzymatic antioxidant defences and aerobic metabolism in the hydrothermal ventworm, Paralvinella grasslei. Mar Biol 2006;150:273–84.
Pruski AM, Dixon DR. Toxic vents and DNA damage: first evidence from a naturallycontaminated deep-sea environment. Aquat Toxicol 2003;64(1):1-13.
Rabilloud T. A comparison between low background silver diammine and silver-nitrateprotein stains. Electrophoresis 1992;13:429–39.
Romero-Isart N, Vašák M. Advances in the structure and chemistry of metallothioneins.J Inorg Biochem 2002;88:388–96.
Sarradin PM, Caprais JC, Riso R, Kerouel R, Aminot A. Chemical environment of thehydrothermal mussel communities in the Lucky Strike and Menez Gwen ventfields, Mid Atlantic Ridge. Cah Biol Mar 1999;40:93-104.

421R. Company et al. / Science of the Total Environment 437 (2012) 413–421
Sarradin PM, Desbruyères D, Dixon DR, Almeida A, Caprais JC, Colaço A, et al. ATOScruise R/V l'Atalante, ROV Victor, June 22nd–July 21st 2001. InterRidge News2001;10(2):18–20.
Serafim A, Lopes B, Company R, Ferreira AM, Bebianno MJ. Comparative petroleum hy-drocarbons levels and biochemical responses in mussels from hydrothermal vents(Bathymodiolus azoricus) and coastal environments (Mytilus galloprovincialis). MarPollut Bull 2008;57(6–12):529–37.
Sheehan D, McDonagh B. Oxidative stress and bivalves: a proteomic approach. InvertSurviv 2008;5:110–23.
Simoneit BRT, Alla YL, Ptersypkin VI, Osipov GA. Composition and origin of hydrothermalpetroleum and associated lipids in the sulfide deposits of the Rainbow Field(Mid-Atlantic Ridge at 36 N). Geochim Cosmochim Acta 2004;68(10):2275–94.
Tapley D, Buettner G, Shick J. Free radicals and chemiluminescence as products of thespontaneous oxidation of sulfide in seawater, and their biological implications.Biol Bull 1999;196:52–6.
Tedesco S, Doyle H, Blasco J, Redmond G, Sheehan D. Oxidative stress and toxicity ofgold nanoparticles in Mytilus edulis. Aquat Toxicol 2010;100:178–86.
Tunnicliffe V. The biology of hydrothermal vents: ecology and evolution. Oceanogr MarBiol Annu Rev 1991;29:319–407.
Winterbourn CC, Hampton MB. Thiol chemistry and specificity in redox signalling. FreeRadic Biol Med 2008;45:549–61.
Xu H, Song P, Gu W, Yang Z. Effects of heavy metals on production of thiol compoundsand antioxidant enzymes in Agaricus bisporus. Ecotoxicol Environ Saf 2011;74(6):1685–92.




















